
The Sexually Dimorphic Medial Preoptic Nucleus of Quail: A Key Brain Area Mediating Steroid Action...
75
The Sexually Dimorphic Medial Preoptic Nucleus of Quail: A Key Brain Area Mediating Steroid Action on Male Sexual Behavior GIAN CARLO PANZICA,* CARLA VIGLIETTI-PANZICA,* AND JACQUES BALTHAZART² *Department of Human Anatomy and Physiology, University of Torino, Corso M. D’Azeglio 52, I-10126 Torino, Italy; and ² Laboratory of Biochemistry, University of Lie `ge, 17 place Delcour, B-4020 Lie `ge, Belgium About 10 years ago, a sexually differentiated nucleus was identified in the preoptic area (POA) of the Japanese quail in the course of studies analyzing the dimorphic mechanisms involved in the activation of sexual behavior. In this species, males exposed to testosterone copulate while females never show this masculine behavior. The present paper reviews anatomical, neurochemical, and functional data that have been collected since that time about the quail dimorphic nucleus. The medial preoptic nucleus (POM) is significantly larger in adult male than in adult female quail. Its volume is also steroid- sensitive in adulthood: it decreases when circulating levels of testosterone are low (castration, exposure to short-days) and it increases when testosterone levels are high (treatment with testosterone, exposure to long-days). The POM is a necessary and sufficient site of steroid action for the activation of male copulatory behavior. The volumetric difference of the POM results from a difference in the adult hormonal milieu of males and females (activational effect) and is not affected by embryonic treatments that permanently modify sexual behavior (no organizational effects on POM). In contrast, the size of neurons in the dorsolateral part of POM appears to be irreversibly affected by embryonic steroids and this feature is therefore a better correlate of the behavioral sex difference. The POM is characterized by the presence of a wide variety of neurotransmit- ters, neuropeptides, and receptors. It can, in addition, be specifically distinguished from the surrounding POA by the presence of aromatase-immunoreactive cells, by a high density of a 2 -adrenergic receptors, and by a dense vasotocinergic innervation. Some of these neurochemical markers of the dimorphic nucleus are themselves modulated by steroids. In particular, the aromatase-immunoreactive cells of the lateral POM appear to be a key target for steroids in the activation of male copulatory behavior. The POM is bidirectionally connected to many brain areas. It receives inputs from a variety of sensory areas and from a number of regulatory areas (e.g., catecholaminergic cell groups). This nucleus also sends outputs to ‘‘neurovegetative’’ centers and to brain regions directly connected to the motor pathways. These connections fully support the role of the POM as an integrative center for the control of male sexual behavior. The available data indicate that there is a high degree of steroid-induced neuronal plasticity in the POM, including changes in neuronal function, in protein synthesis, and in specific inputs. These phenom- ena can easily be studied in the POM because they are of a large magnitude, they are localized in a specific brain site, and they develop rapidly after exposure to steroids. They are also directly related to a clear functional output, the activation of male sexual behavior. The quail POM therefore constitutes an exceptional model for the analysis of steroid-induced brain plasticity in a functionally relevant context. KEY WORDS: SDN- POA; Coturnix japonica; avian brain; testosterone; aromatase; male copulatory behavior; catecholamines. r 1996 Academic Press, Inc. Address reprint requests to G.C. Panzica, Department of Human Anatomy and Physiology, University of Torino, Corso M. D’Azeglio 52, I-10126 Torino, Italy. FRONTIERS IN NEUROENDOCRINOLOGY 17, 51–125 (1996) Article No. 0002 51 0091-3022/96 $12.00 Copyright r 1996 by Academic Press, Inc. All rights of reproduction in any form reserved.
Transcript of The Sexually Dimorphic Medial Preoptic Nucleus of Quail: A Key Brain Area Mediating Steroid Action...

The Sexually Dimorphic Medial Preoptic Nucleus of Quail:A Key Brain Area Mediating Steroid Action on Male Sexual Behavior
GIAN CARLO PANZICA,* CARLA VIGLIETTI-PANZICA,* AND JACQUES BALTHAZART†
*Department of Human Anatomy and Physiology, University of Torino, Corso M. D’Azeglio 52,I-10126 Torino, Italy; and † Laboratory of Biochemistry, University of Liege,
17 place Delcour, B-4020 Liege, Belgium
About 10 years ago, a sexually differentiated nucleus was identified in the preopticarea (POA) of the Japanese quail in the course of studies analyzing the dimorphicmechanisms involved in the activation of sexual behavior. In this species, males exposedto testosterone copulate while females never show this masculine behavior. The presentpaper reviews anatomical, neurochemical, and functional data that have been collectedsince that time about the quail dimorphic nucleus. The medial preoptic nucleus (POM) issignificantly larger in adult male than in adult female quail. Its volume is also steroid-sensitive in adulthood: it decreases when circulating levels of testosterone are low(castration, exposure to short-days) and it increases when testosterone levels are high(treatment with testosterone, exposure to long-days). The POM is a necessary andsufficient site of steroid action for the activation of male copulatory behavior. Thevolumetric difference of the POM results from a difference in the adult hormonal milieuof males and females (activational effect) and is not affected by embryonic treatmentsthat permanentlymodify sexual behavior (no organizational effects on POM). In contrast,the size of neurons in the dorsolateral part of POM appears to be irreversibly affected byembryonic steroids and this feature is therefore a better correlate of the behavioral sexdifference. The POM is characterized by the presence of a wide variety of neurotransmit-ters, neuropeptides, and receptors. It can, in addition, be specifically distinguished fromthe surrounding POA by the presence of aromatase-immunoreactive cells, by a highdensity of a2-adrenergic receptors, and by a dense vasotocinergic innervation. Some ofthese neurochemical markers of the dimorphic nucleus are themselves modulated bysteroids. In particular, the aromatase-immunoreactive cells of the lateral POM appear tobe a key target for steroids in the activation of male copulatory behavior. The POM isbidirectionally connected to many brain areas. It receives inputs from a variety of sensoryareas and from a number of regulatory areas (e.g., catecholaminergic cell groups). Thisnucleus also sends outputs to ‘‘neurovegetative’’ centers and to brain regions directlyconnected to the motor pathways. These connections fully support the role of the POM asan integrative center for the control of male sexual behavior. The available data indicatethat there is a high degree of steroid-induced neuronal plasticity in the POM, includingchanges in neuronal function, in protein synthesis, and in specific inputs. These phenom-ena can easily be studied in the POM because they are of a large magnitude, they arelocalized in a specific brain site, and they develop rapidly after exposure to steroids. Theyare also directly related to a clear functional output, the activation of male sexualbehavior. The quail POM therefore constitutes an exceptional model for the analysis ofsteroid-induced brain plasticity in a functionally relevant context. KEY WORDS: SDN-POA; Coturnix japonica; avian brain; testosterone; aromatase; male copulatory behavior;catecholamines.r 1996 Academic Press, Inc.
Address reprint requests to G.C. Panzica, Department of Human Anatomy and Physiology,University of Torino, Corso M. D’Azeglio 52, I-10126 Torino, Italy.
FRONTIERS IN NEUROENDOCRINOLOGY 17, 51–125 (1996)Article No. 0002
51 0091-3022/96 $12.00Copyright r 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

INTRODUCTION
Behavior represents a major output from the brain and the identification andmapping of neural circuits controlling specific behaviors represents a majorchallenge facing contemporary neuroscience. The circuit mediating femalesexual receptivity (the lordosis response in rodents) has been described over thepast 20 to 30 years by charting with tract-tracing and other experimentalapproaches the flow of neural activity from the stimulus provided by the maleto the muscular response by the female rat (206, 207). This approach identified,in the brain, the areas projecting to the major muscles implicated in thelordosis behavior. By contrast, male courtship and sexual behavior implicatethe use of most, if not all, skeletal muscles and the integration of a diversity ofsensory inputs. Therefore such a strategy of retrograde tracing cannot be usedefficiently in the male. There is ample evidence that testosterone (T), actingmainly through aromatization to estrogens, activates mating behavior in theadult male (35, 164, 175) in most species of vertebrates. There are, however, afew exceptions (98–100). The search of the neural site(s) of steroid action onmale copulatory behavior therefore appears as a promising strategy to identifycircuits controlling this type of behavior.The presence of gonadal steroid receptors in similar locations in the central
nervous system (CNS) throughout the entire vertebrate class (169, 170, 247,248) suggests that steroids have evolutionarly stable roles in the differentiationand postnatal development of target cerebral circuits, as well as in the mainte-nance of their functions in the adult brain. An intense research effort wasinitiated in the early 1970s to localize steroid receptors in the brain of a varietyof vertebrate species and to evaluate the biological significance of steroidbinding at these locations. Steroid receptors were found in the anterior hypo-thalamus (including the preoptic region) and in the tuberal region of virtuallyall vertebrates. Stereotaxic implantations of steroids and electrolytic lesions ofdiscrete brain areas also demonstrated that the preoptic area (POA) and thehypothalamus regulate both endocrine and behavioral components of reproduc-tion, i.e., the hypophysial secretion of gonadotrophins, as well as mounting andlordosis behavior (149, 164, 208).At approximately the same time, a number of structural sex dimorphisms
were identified, at light microscopic level, in the vertebrate CNS. Initially, a sexdimorphism in the ultrastructural organization of rat POA neuropil wasdiscovered (211). Large sexually dimorphic structures were thereafter discov-ered in the telencephalon of songbirds (179) and soon after in the POA of rats(127, 128) and of a variety of other mammals (19, 95, 142, 250, 258, 259).Frequently, the sexually dimorphic structures and the receptors for gonadalsteroids were located in the same or in a closely related area, suggesting thatthese neuroanatomical dimorphisms might be influenced by gonadal steroids(193).The work of Nottebohm and Arnold (179) and of Gorski et al. (127, 128), who
first identified, at the light microscopic level, sexually dimorphic structures inthe vertebrate brain, was important not only in establishing that large morpho-
52 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

logical differences exist between the brains of males and females, but also insuggesting a new approach in the study of neural mechanisms controlling thesex differences in reproductive behavior.Since male sexual behavior is also sexually differentiated in many species (it
is more easily activated by androgens in males than in females), it could beexpected that these sexually dimorphic structures would play a key role in theactivation of this behavior (129, 175). This expectation was not really fulfilledand, in particular in rats, it has been difficult to relate in a causal manner thesexual dimorphism of the POA (the sexually dimorphic nucleus of the preopticarea; SDN-POA, is bigger in males than in females) with the mechanismmediating male copulatory behavior (22), although more recent data mayresolve this problem, at least in part (105). The same conclusion was reached inother mammalian species studied (the dimorphic nuclei of the POA do notappear to be specifically implicated in the control of male behavior), except ingerbils, in which a sexually dimorphic area of the POA appears to play a keyrole in the control of male sexual behavior (281, 282).Approximately 10 years ago, our laboratories initiated a collaborative re-
search project utilizing neuroanatomical, biochemical, and behavioral tech-niques to identify the central mechanisms controllingmale reproductive behav-ior in an avian species commonly used in laboratory studies, the Japanese quail(Coturnix japonica). This led to the discovery of a sexually dimorphic nucleus inthe POA of this species. Subsequent studies identified remarkable features ofthis SDNwhich, contrary to the rat SDN-POA, appears to play a key role in theactivation by steroids of male copulatory behavior. The major results of thisresearch are summarized here.
ACTIVATION OF SEXUAL BEHAVIOR BY STEROIDSAND ITS SEXUAL DIMORPHISM
In quail, contrary to what is observed in rodents, the male-type copulatorybehavior is highly differentiated between males and females, while the female-type receptive behavior can be activated in both sexes by an appropriatetreatment with estrogens (6, 10). The sexual dimorphism affectingmale copula-tory behavior is quite extreme in this species: under laboratory test conditions,sexually mature males almost never fail to exhibit the complete copulatorysequence, including grabbing the female’s neck feathers, mount attempts,mounts, and cloacal contact movements. These behaviors are never seen infemales (6, 65). The dimorphism is an all or none phenomenon; it is qualitativein nature (Fig. 1).All of these behaviors are T-dependent and they disappear after castration in
males but their absence in gonadally intact females is not due to a relativeabsence of circulating T. It has indeed been shown that treatment with highdoses of T is not sufficient to activate cloacal contact movements in gonadecto-mized females, while such a treatment restores the full spectrum of sexualbehaviors in males (6, 65, 226). Furthermore, radioimmunoassays of plasma
53THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
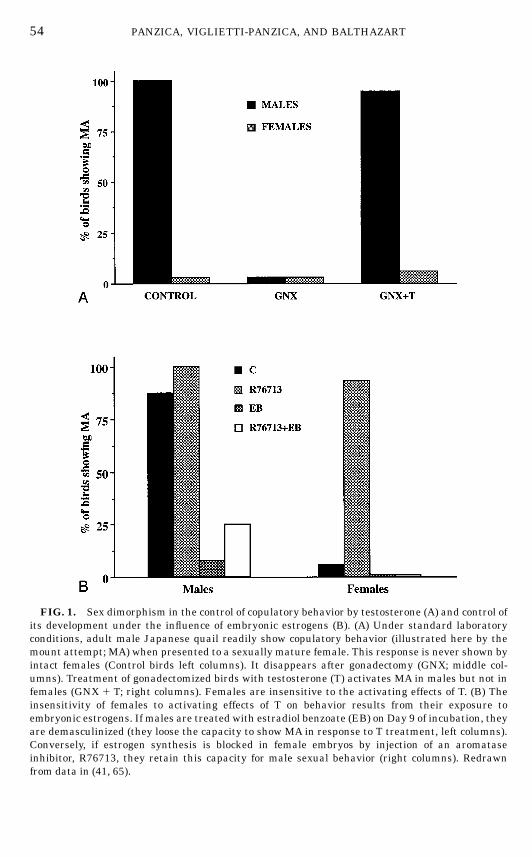
FIG. 1. Sex dimorphism in the control of copulatory behavior by testosterone (A) and control ofits development under the influence of embryonic estrogens (B). (A) Under standard laboratoryconditions, adult male Japanese quail readily show copulatory behavior (illustrated here by themount attempt; MA) when presented to a sexually mature female. This response is never shown byintact females (Control birds left columns). It disappears after gonadectomy (GNX; middle col-umns). Treatment of gonadectomized birds with testosterone (T) activates MA in males but not infemales (GNX 1 T; right columns). Females are insensitive to the activating effects of T. (B) Theinsensitivity of females to activating effects of T on behavior results from their exposure toembryonic estrogens. If males are treated with estradiol benzoate (EB) on Day 9 of incubation, theyare demasculinized (they loose the capacity to show MA in response to T treatment, left columns).Conversely, if estrogen synthesis is blocked in female embryos by injection of an aromataseinhibitor, R76713, they retain this capacity for male sexual behavior (right columns). Redrawnfrom data in (41, 65).
54 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

steroids demonstrate that circulating levels of T overlap to a large extent inmale and female quail even if they are, on average, higher in males (42, 65,112). This implies that the activation by androgens should be sufficient toinduce male-type copulatory behavior in at least some of the females. This isclearly not the case, which suggests that the neuronal circuits supporting malereproductive behavior are sexually differentiated in this species.Experimental studies carried out during the past 20 years unequivocally
demonstrate that the sex difference in sensitivity to the activating effects of Ton copulatory behavior exclusively results from the early exposure of femalebrains to higher levels of circulating estrogens. It was shown by E. Adkins-Regan in the 1970s that a single injection of estradiol benzoate (EB) into malequail eggs produces adults that will never mount or copulate even aftertreatment with high doses of T. These birds are said to be demasculinized andthey have acquired a female phenotype as far as sexual behavior is concerned(6, 8, 9, 14, 15). This effect is robust and can easily be replicated (231). Femaleembryos developing under physiological conditions are similarly exposed tohigh levels of estrogens and these are responsible for the demasculinization oftheir behavior. It has indeed been shown that plasma levels of estradiol-17b aresignificantly higher in female than in male quail embryos during the periodwhen experimental manipulations of steroid levels are able to modify in anirreversible manner the behavioral phenotype of young quail, i.e., betweenDays 9 and 15 of incubation (232). Furthermore, the experimental inhibition ofaromatase (the enzyme catalyzing the synthesis of estrogens) in female em-bryos produces adults that will copulate in a male-like fashion when exposed toT and this effect can be reversed by the simultaneous administration of EB (41).
IMPORTANCE OF T METABOLISM IN THE CONTROL OF SEXUAL BEHAVIOR
In Japanese quail, as in other avian species and in mammals, T is activelymetabolized by the brain into a number of androgenic or estrogenic steroidsthat often participate in the activation of sexual behavior. Other metabolites ofT, such as the 5b-reduced androstanes (a family of steroids that includes5b-dihydrotestosterone), have apparently no behavioral effect on their own andtherefore represent an inactivation shunt that limits T action in the brain (35,145).Two metabolic pathways of T are particularly relevant to the control of male
sexual behavior: 5a-reduction and aromatization. These enzymatic transforma-tions are illustrated in Fig. 2 together with a schematic presentation of theirimplications in the control of this behavior.The 5a-reductase catalyzes the reduction of T into 5a-dihydrotestosterone
(5a-DHT), a steroid that binds with high affinity to the androgen receptor (AR)and, in this way, activates a number of androgen-dependent responses. Theenzymatic complex aromatase catalyzes a complex cascade of reactions (17,151, 244) that leads to the cyclization of the ‘‘A’’ ring of the androgen moleculeand the loss of carbon number 19. The resulting metabolites are aromatic
55THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
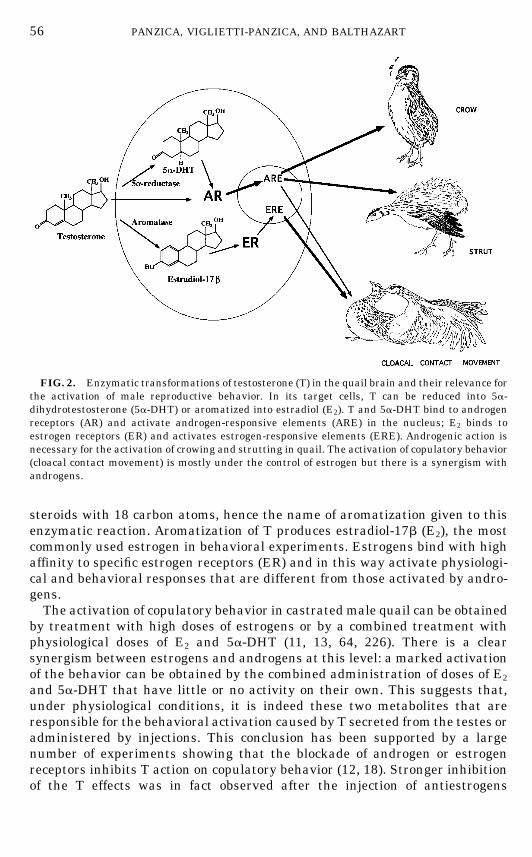
steroids with 18 carbon atoms, hence the name of aromatization given to thisenzymatic reaction. Aromatization of T produces estradiol-17b (E2), the mostcommonly used estrogen in behavioral experiments. Estrogens bind with highaffinity to specific estrogen receptors (ER) and in this way activate physiologi-cal and behavioral responses that are different from those activated by andro-gens.The activation of copulatory behavior in castrated male quail can be obtained
by treatment with high doses of estrogens or by a combined treatment withphysiological doses of E2 and 5a-DHT (11, 13, 64, 226). There is a clearsynergism between estrogens and androgens at this level: a marked activationof the behavior can be obtained by the combined administration of doses of E2
and 5a-DHT that have little or no activity on their own. This suggests that,under physiological conditions, it is indeed these two metabolites that areresponsible for the behavioral activation caused by T secreted from the testes oradministered by injections. This conclusion has been supported by a largenumber of experiments showing that the blockade of androgen or estrogenreceptors inhibits T action on copulatory behavior (12, 18). Stronger inhibitionof the T effects was in fact observed after the injection of antiestrogens
FIG. 2. Enzymatic transformations of testosterone (T) in the quail brain and their relevance forthe activation of male reproductive behavior. In its target cells, T can be reduced into 5a-dihydrotestosterone (5a-DHT) or aromatized into estradiol (E2). T and 5a-DHT bind to androgenreceptors (AR) and activate androgen-responsive elements (ARE) in the nucleus; E2 binds toestrogen receptors (ER) and activates estrogen-responsive elements (ERE). Androgenic action isnecessary for the activation of crowing and strutting in quail. The activation of copulatory behavior(cloacal contact movement) is mostly under the control of estrogen but there is a synergism withandrogens.
56 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

compared to antiandrogens (18, 68), suggesting that estrogenic metabolites of Tare more important for the behavioral activation than T itself or its androgenicmetabolites such as 5a-DHT. This conclusion is also supported by the observa-tion that aromatase inhibitors can completely suppress the effects of T oncopulatory behavior, while 5a-reductase inhibitors have little or no effect at thislevel (11, 18, 45, 49, 68). It should be noted that this comparison may be theresult of a technical artifact due to the higher efficiency of aromatase inhibitorsin general (independent of the response considered). Nevertheless, it is clearthat estrogens alone can activate male copulatory behavior in quail and thatthe inhibition of aromatase completely blocks the effects of T on this behavior.This firmly demonstrates that the aromatization of T is a limiting step for theactivation of sexual behavior by T (46).Interestingly, other reproductive behaviors such as the courtship display
referred to as strut and male-specific vocalizations such as crows are activatedin quail by androgens only (T or 5a-DHT). Estrogens have no effect on thesebehaviors or, even, may inhibit their activation by androgens (7, 13, 35, 62,226). These behaviors can therefore be conveniently used to monitor theendocrine specificity of steroid treatments applied to experimental subjects.
IDENTIFICATION OF A SEXUALLY DIMORPHIC NUCLEUS IN THE QUAIL POA
Adetailed analysis of the central mechanisms controlling male sexual behav-ior required the identification of the brain area(s) where binding of steroids(mostly T and its metabolites) is a prerequisite for the activation of behavior.Converging evidence suggested the POA as a key site in this respect. It wasestablished in birds and in other vertebrates that the POA contains highconcentrations of steroid receptors as revealed by autoradiography or immuno-cytochemistry (34, 57, 169, 170, 247, 248, 276) and that lesions of the POAdrastically impair male sexual behavior, while implants of T or E2 in the POAofcastrated subjects restore a normal or nearly normal level of sexual activity(164, 175, 277, 278). This led us to undertake a detailed morphological analysisof the POA in quail with the hope of identifying a cytoarchitectonically definedgroup(s) of cells that would represent a critical site of steroid action onbehavior. Because male quail reliably mount and copulate when exposed to T,while females never show these behaviors, it was also hoped that this qualita-tive behavioral dimorphism would be reflected in some way at the brain leveland that sexually dimorphic cell groups would be identified in the quail POAashad been done in rats and other mammals.In the quail preoptic region, several magno- and parvocellular masses could
be recognized in Nissl-stained sections (189, 202, 204, 271).Large and intensely stained neurons located in periventricular or more
lateral positions correspond to the neurophysin-immunoreactive system aspreviously described in quail (81, 181, 190, 266, 270). Several groups ofparvocellular neurons are also identified (189). An easily recognizable nucleusis situated ventromedially under the tractus septomesencephalicus and corre-
57THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

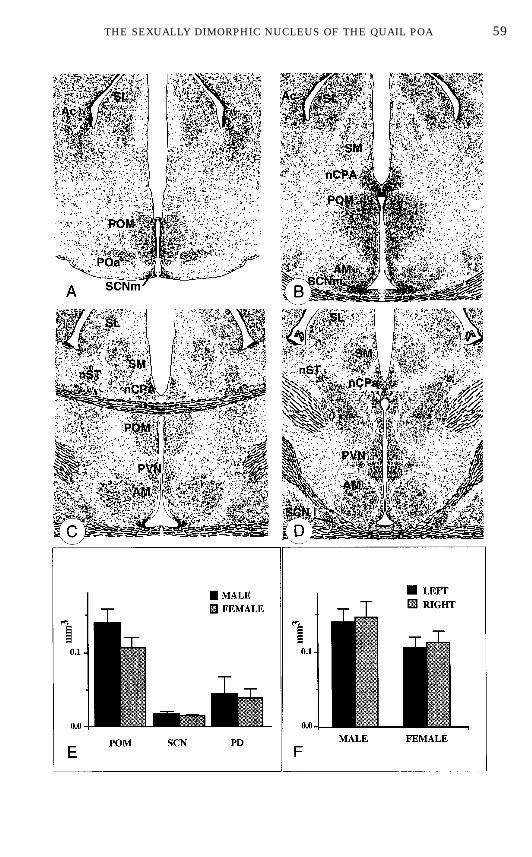
sponds to the nucleus preopticus dorsolateralis (PD) described in the chickenatlas of Kuenzel and van Tienhoven (153). Ventrally, at a slightly more caudallevel, another dense cluster of parvocellular elements that extends laterallylike the wings of a butterfly at the level of the rostral pole of the third ventriclecan be observed (Fig. 3A). This should be identified as the nucleus preopticusanterior (POa) according to Karten and Hodos (148), Stokes et al. (246), andVowles et al. (274). A poorly developed periventricular preoptic nucleus (POP)(102), characterized by a columnar arrangement of a few lines of magnocellularneurons, is located rostrally. Lateral to this small cell group, a large nucleus ispresent that extends throughout most of the preoptic region in the rostrocaudalaxis. At its most rostral level, this nucleus is contiguous with the POP. Morecaudally, it extends dorsolaterally and acquires a characteristic oblong shapejust rostral to the anterior commissure (Figs. 3A–3B). This nucleus extendsdorsally until the level of the anterior commissure and then quickly disappears,although, in some cases, a cluster of cells seems to extend evenmore dorsolater-ally and merges with the bed nucleus striae terminalis (nST). This cell grouphas been called nucleus preopticus medialis (POM) based on several photo-graphic and schematic descriptions of the region in a number of avian species(72, 102, 148, 246, 274, 285). No lateral preoptic nucleus is discernible in thequail preoptic area. It is probably composed of scattered small neurons that arelateral to the POM and interspersed within the lateral forebrain bundle fibers.Dorsal to the anterior commissure close to the median line, a cluster of smalldarkly stained cells can be identified as the bed nucleus of the pallial commis-sure (nucleus commissurae pallii, nCPa; Figs. 3B–3D). They appear to extendcaudally a group of similar neurons located above the organum vasculosumlaminae terminalis. Finally, near the lateral angle of the third ventricle there isan accumulation of cells identified as the nucleus suprachiasmaticus, parsmedialis [SCNm, Figs. 3A–3B; (152)].During our first study of the quail POA, the volume of several preoptic nuclei
was reconstructed in 50-µm-thick paraffin sections that had been stained forNissl material with toluidine blue (271). This demonstrated that the volume ofthe POM is significantly larger (40%) in males than in females. Preoptic nucleithat display such volumetric differences are classically called sexually dimor-phic in the literature and a similar nomenclature has been adopted here. It
FIG. 3. Schematic representation of the cell groups in the quail preoptic area (A–D) andquantitative determination of the volume of selected nuclei illustrating the sex difference affectingthe medial preoptic nucleus (POM). A through D are organized in a rostral to caudal order. E showsthat the volume of POM is significantly larger in males than in females and this difference does notaffect other neighboring nuclei. F demonstrates that the sex difference in POM volume is presenton the left and right sides of the brain and that there is no significant asymmetry in the volume ofthe nucleus. Redrawn from data in (204, 271). Abbreviations: Ac, nucleus accumbens; AM, nucleusanterior (rostralis) medialis hypothalami; nCPA, nucleus commissurae pallii; PD, nucleus preopti-cus dorsolateralis; POa, nucleus preopticus anterior; POM, nucleus preopticus medialis; PVN,nucleus paraventricularis magnocellularis; nST, nucleus striae terminalis; SL, nucleus septalislateralis; SM, nucleus septalis medialis; SCNl, nucleus suprachiasmaticus, pars lateralis; SCNm,nucleus suprachiasmaticus, pars medialis (see text for additional comments on this nomenclature).
58 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
59THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

must be noted, however, that these nuclei are not dimorphic strictly speakingsince they usually have similar shape and differ only in volume.No statistically significant sex difference in volume could be detected for the
other nuclei that were considered (preopticus dorsalis, suprachiasmaticus, andnucleus rotundus thalami; see Fig. 3E). The dimorphism was observed on bothsides of the brain (Fig. 3F) and no lateral asymmetry in the volume of thenucleus could be detected (204). The lack of correlation between the POMvolume and the volume of other surrounding nuclei (e.g., the nucleus rotundus)confirmed that the dimorphism in POM volume is specific and does not resultfrom a gross difference in brain size (194). This sex difference was indepen-dently confirmed in our more recent work (189) and in another laboratory (16).This latter study also failed to identify volumetric sex differences for otherquail brain nuclei (nucleus of the pallial commissure, nucleus taeniae, nucleusof the XIIth nerve) (16).These morphometric studies therefore allow us to add the POM to the list of
sexually dimorphic structures that have been identified at light microscopiclevel in the POA of all classes of tetrapods, including amphibia, reptiles, andmammals, including man (19, 84, 95, 101, 127, 128, 142, 193, 250, 252, 258,259). In view of the fact that the first major sex dimorphism in brain structure(larger brain nuclei controlling song in passerines) was described in birds (179),it is surprising that the Japanese quail is, to our knowledge, the only avianspecies in which a sexually dimorphic nucleus has been identified in the POA.
THE POM IS A STEROID-SENSITIVE STRUCTURE
Because the sexually dimorphic POMwas located in the POA, an area knownto be critical for the control of male reproductive behavior, it seemed likely thatthis nucleus was a key center in the control of behavior. This hypothesis wasoriginally tested by examining the relationships between the sexual dimor-phisms in POM volume and in copulatory behavior. Treatments which wereknown to affect sexual behavior (e.g., castration and testosterone replacement,estrogen injection in the egg; see above) were applied to different groups ofquail and their effects on behavior and on POM volume were quantified.
Effect of T inAdult Castrated Birds
The first experiment tested whether the sex difference in POM volume thathad been described was related to the different hormonal milieu that is presentin adult males and females or rather if it represented a stable differencepresumably organized by neonatal hormones (202). Birds were gonadectomizedor sham-operated just before puberty (about 4 weeks of age) and one-half of thegonadectomized birds received T replacement therapy. As shown previously (6,10, 65), T treatment of gonadectomized birds activated sexual behavior inmales but not in females, despite the fact that the hormone implants produced
60 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

similar levels of T in both sexes (42, 65, 228). This T level was comparable tothat seen in intact mature males (42, 65). A significant decrease in POMvolume, compared with that in sexually mature gonadally intact subjects, wasobserved in castrated males. This decrease was completely reversed whenmales were treated with exogenous T (Fig. 4A). Interestingly, ovariectomizedfemales treated with the same dose of T had POM volumes similar to those ofmales submitted to the same treatment. The POM volume measured in gonad-ectomized T-treated birds was therefore not sexually dimorphic (202). Thissuggested that the sex difference in intact birds is caused by the differentcirculating levels of T in adult males and females. Presumably, this difference isactivational in nature and is not the result of a differential exposure toestrogens during embryonic life (no organizational difference). This conclusionwas directly confirmed in experiments manipulating the steroid milieu ofembryos (see below).
Photoperiodic Control
Under laboratory conditions, the testis size and the testicular production of Tare markedly affected by the photoperiod that the birds are exposed to (120).Males raised on short days (less than 12 h of light per day) have regressedtestes (100–200 mg) and basal levels of circulating T (less than 0.5 ng/ml). Ifsuch birds are transferred to long days (e.g., 18L: 8D), a rapid growth of thetestes follows; the testes reach a combined weight above 5 g in a few weeks andplasma T levels increase to values that range between 2 and 4 ng/ml (59). Inparallel, the rise in plasma T levels activates copulatory behavior (65). Recentexperiments indicate that the volume of the POM also changes as a function ofthe photoperiod under which male quail are maintained (Fig. 4B). Birds livingin short days have smaller POM volumes than birds living in long days. Thisdifference can be observed if the different photoperiods are applied to birdsentering the final phase of sexual maturation, i.e., subjects that are 4–6 weeksold (189), or to fully adult subjects (256). This brings further support to thenotion that the POM volume in adult quail directly correlates with the circulat-ing levels of plasma T.
Effect of Estrogens in Embryos
Behavioral experiments had established that the treatment of male quailembryos with estrogens leads to a complete demasculinization of their sexualbehavior so that they will never show, as adults, the normal copulatorysequence even if exposed to high levels of T (6, 8, 9, 14, 15, 231). This type oftreatment, if performed before Day 12 of incubation, has an irreversibleorganizational effect on the brain: males acquire a female behavioral pheno-type; they are said to be ‘‘demasculinized.’’ In order to further specify therelationships between POM volume and male copulatory behavior, we studied
61THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
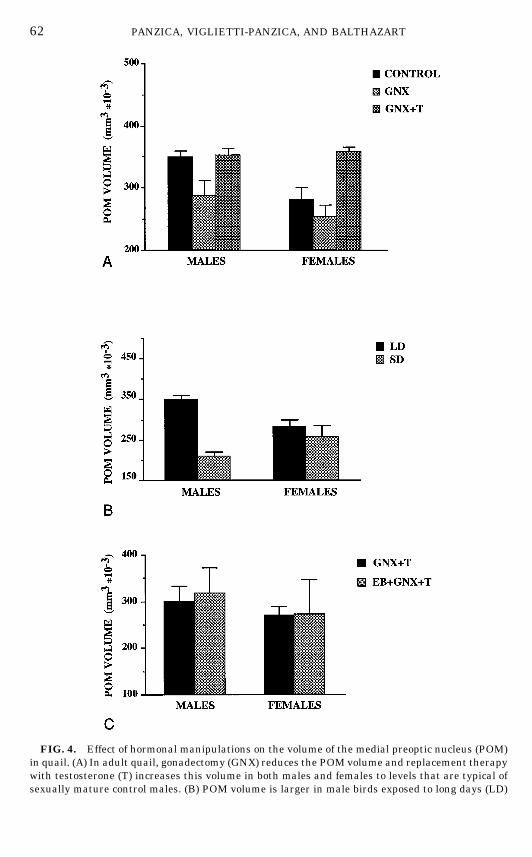
FIG. 4. Effect of hormonal manipulations on the volume of the medial preoptic nucleus (POM)in quail. (A) In adult quail, gonadectomy (GNX) reduces the POM volume and replacement therapywith testosterone (T) increases this volume in both males and females to levels that are typical ofsexually mature control males. (B) POM volume is larger in male birds exposed to long days (LD)
62 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

the effects of an early estrogen treatment on the volume of the dimorphicnucleus in males and females that were as adults either gonadally intact orgonadectomized and submitted to a replacement therapy with exogenous T (26,202). The injection of EB into 9-day-old quail embryos did not affect the volumeof the POM in the adult birds which were grown from the injected eggs (studiedboth as intact or T-treated gonadectomized quail; Fig. 4C). This further sup-ports the notion that the sex difference in POM volume is activational and notorganizational in nature. This is also in agreement with the analysis of thePOM during embryonic stages until hatching that revealed no significant sexdifference in volume (201).Moreover, a recent developmental study of posthatch-ing stages showed that the sexual dimorphism in POM volume is not presentduring the first few weeks of life in quail and becomes apparent only at the ageof 6 weeks (257) when the gonads begin to secrete T in a sexually dimorphicmanner (232).Taken together, these experiments demonstrate that the volume of the quail
POM is directly correlated with adult androgen levels. This nucleus constitutesan excellent marker of T action in the POA. The sexual dimorphism affectingthe overall volume of this structure does not result from an irreversible actionof embryonic steroids but only reflects a differential activation by T in adultmales and females.
Endocrine Specificity in theAction of Steroids on POM Volume
The studies described above showed that, in adult male quail, an increase inPOM volume is always present under physiological conditions that are associ-ated with an activation of male-type copulatory behavior (treatment of cas-trates with exogenous T, photostimulation of gonadally intact birds). It hadbeen previously demonstrated that the effects of T on copulatory behaviorlargely result, at the cellular level, from an interaction of its estrogenic (E2) andandrogenic (5a-DHT) metabolites with their specific receptors (7, 11, 13, 18, 64,226). This provided an independent way of assessing the extent to whichincreases in POM volume are critically associated with the activation of malesexual behavior. Therefore experiments were carried out to analyze the endo-crine specificity of the control of POM volume and compare it with the specific-ity of the behavioral activation. The effect of T and its metabolites E2 and5a-DHT administered alone or in combination were tested in the first experi-ment (24). This confirmed that T increases the volume of the dimorphicnucleus. No treatment with metabolites produced a significant increase in the
that have high circulating levels of testosterone than in birds exposed to short days (SD) with lowlevels of testosterone. This effect of the photoperiod is not observed in females. (C) Injection ofestradiol benzoate (EB) into quail embryos does not affect in a permanentmanner the volume of thePOM.All birds in this experiment were gonadectomized and treated with testosterone in adulthood(GNX 1 T) to avoid confounding organizational and activational effects of steroids. Redrawn fromdata in (189, 202).
63THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

volume of the POM, but the best numerical increment was observed in birdssimultaneously treated with E2 and 5a-DHT, that is, the treatment that alsoprovides the best activation of sexual behavior.Interestingly, another study showed that the blockade of aromatase activity
by a systemic treatment with the aromatase inhibitor R76713 not only sup-presses the activational effects of T on copulatory behavior, but also prevents Tfrom increasing the volume of the sexually dimorphic nucleus, POM (45).Available data are therefore entirely consistent with the idea that T actsmainlythrough the same metabolites on both the copulatory behavior and the POMvolume (major effect of estrogens with a synergistic action of 5a-DHT).
THE POM IS A KEY CENTER FOR STEROID ACTION ON COPULATORY BEHAVIOR
The studies summarized in the previous sections describedmultidimensionalcorrelations between the activation of male sexual behavior by a variety ofsteroid treatments and the increase in volume of the sexually dimorphic POM.They clearly suggested that the two types of responses could be causally linkedbut the demonstration of this idea required direct experimental manipulationof the POM. A series of experiments were therefore undertaken to test thishypothesis.
Electrolytic Lesions
In a first set of studies, bilateral electrolytic lesions aimed at the POM wereplaced in the POA of castrated male quail. Immediately after the lesion, birdsreceived a subcutaneous Silastic implant filled with T and the restoration oftheir copulatory behavior was monitored during the following 3 weeks duringstandard behavioral tests that took place three time a week. Lesions thatdestroyed a significant fraction of the POM (more than 10% of the total volume)markedly delayed or even blocked the restoration by T of copulatory behavior.There was a very significant positive correlation between the volume of thePOM that had been destroyed by the lesion and the behavioral deficit measuredby the latency in days to restore male sexual behavior. The frequencies ofsexual behaviors were similarly affected (Fig. 5A). Lesions of the same sizelocated elsewhere in the POA had no detectable effect on behavior (69). Thebehavior was therefore not related to the absolute size of the lesion but rather to itsposition. These results were independently confirmed in a separate study (70).
T Implants
The lesion studies suggested that the POM plays a key role in the activationof male sexual behavior by T. This conclusion was confirmed in a subsequentexperiment which assessed the behavioral effects of 27-gauge crystalline T
64 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
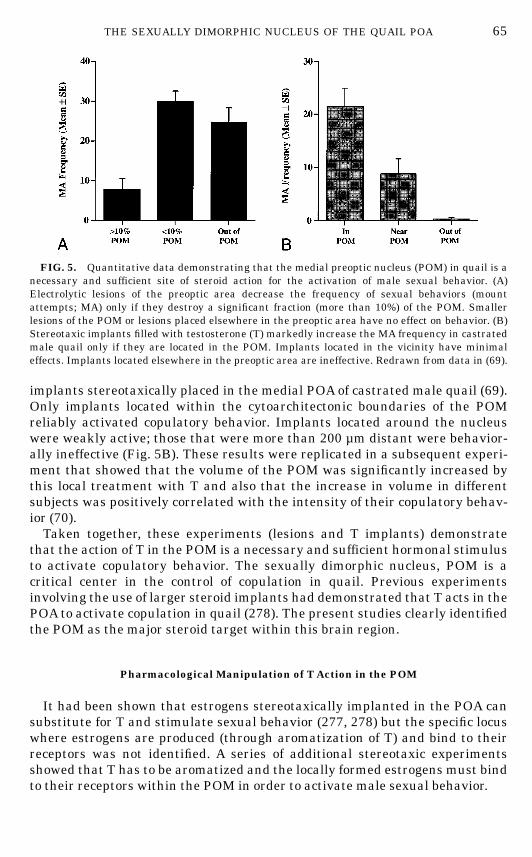
implants stereotaxically placed in the medial POA of castrated male quail (69).Only implants located within the cytoarchitectonic boundaries of the POMreliably activated copulatory behavior. Implants located around the nucleuswere weakly active; those that were more than 200 µm distant were behavior-ally ineffective (Fig. 5B). These results were replicated in a subsequent experi-ment that showed that the volume of the POM was significantly increased bythis local treatment with T and also that the increase in volume in differentsubjects was positively correlated with the intensity of their copulatory behav-ior (70).Taken together, these experiments (lesions and T implants) demonstrate
that the action of T in the POM is a necessary and sufficient hormonal stimulusto activate copulatory behavior. The sexually dimorphic nucleus, POM is acritical center in the control of copulation in quail. Previous experimentsinvolving the use of larger steroid implants had demonstrated that T acts in thePOA to activate copulation in quail (278). The present studies clearly identifiedthe POM as the major steroid target within this brain region.
Pharmacological Manipulation of TAction in the POM
It had been shown that estrogens stereotaxically implanted in the POA cansubstitute for T and stimulate sexual behavior (277, 278) but the specific locuswhere estrogens are produced (through aromatization of T) and bind to theirreceptors was not identified. A series of additional stereotaxic experimentsshowed that T has to be aromatized and the locally formed estrogens must bindto their receptors within the POM in order to activate male sexual behavior.
FIG. 5. Quantitative data demonstrating that the medial preoptic nucleus (POM) in quail is anecessary and sufficient site of steroid action for the activation of male sexual behavior. (A)Electrolytic lesions of the preoptic area decrease the frequency of sexual behaviors (mountattempts; MA) only if they destroy a significant fraction (more than 10%) of the POM. Smallerlesions of the POM or lesions placed elsewhere in the preoptic area have no effect on behavior. (B)Stereotaxic implants filled with testosterone (T) markedly increase the MA frequency in castratedmale quail only if they are located in the POM. Implants located in the vicinity have minimaleffects. Implants located elsewhere in the preoptic area are ineffective. Redrawn from data in (69).
65THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

Implantation of the aromatase inhibitors, R76713 (45), or androstatrienedi-one, ATD (68), into the POM inhibited the induction by T of sexual behavior.Because a larger number of birds had been studied in the ATD experiment, therelationship between the anatomical location of the tip of the implants and thebehavioral inhibition was analyzed in more detail. ATD implants that werepositioned within the POM effectively inhibited the activation of sexual behav-ior by T, while those located in the POA but outside POM had no behavioraleffect and were strictly comparable to control implants filled with cholesterol.Subsequent studies showed that male copulatory behavior can be activated
in castrated quail by stereotaxic implants of the synthetic estrogen diethylstil-bestrol placed in the POM and, conversely, that the behavioral effects of asystemic treatment with T can be blocked by the implantation in the POM ofthe antiestrogen tamoxifen (68).Taken together, all of these studies therefore reveal that the sexually dimor-
phic POM is the area where the behaviorally active estrogenic metabolites of Thave to be produced and bind to estrogen receptors in order to stimulatecopulation.
CELL SIZE IN THE DORSOLATERAL POM IS SEXUALLY DIFFERENTIATED
The studies reviewed so far established that (a) the POM is larger in malesthan in females, (b) its growth in adult birds is controlled by steroids (T and itsestrogenic metabolites), (c) the size of the POM in males correlates with theirreproductive activity, and (d) the POM is a key center for steroid action onbehavior. Moreover, in adult birds, the POM is larger in males than in femalesonly because circulating levels of androgens are higher in the former than inthe latter. When females are treated with T, they have a POM that is as large asthat in males, despite the fact that they still do not show any male-typicalcopulatory behavior. The overall POM volume is therefore not a good correlateof the behavioral dimorphism. Because this nucleus is a key center in thecontrol of behavior, it was reasoned that a more detailed analysis of thisnucleus could reveal features that would be dimorphic in the organizationalsense (irreversibly differentiated by early exposure to steroids) and would leadto themechanismsmediating the dimorphism in behavior. Cytological analysesof the POMwere therefore undertaken.
Effects of Androgens on the Neuronal Size in the Dorsolateral POM
A closer observation of the POM organization in 10-µm-thick paraffin sec-tions suggested that two distinct neuronal populations could be differentiatedwithin this nucleus. The perikarya located in the dorsolateral part of thenucleus appeared much larger than those present in its medial part (Fig. 6A).Quantitative studies confirmed that neurons located in the dorsolateral POM
were significantly larger than those located in the medial part of the nucleus
66 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
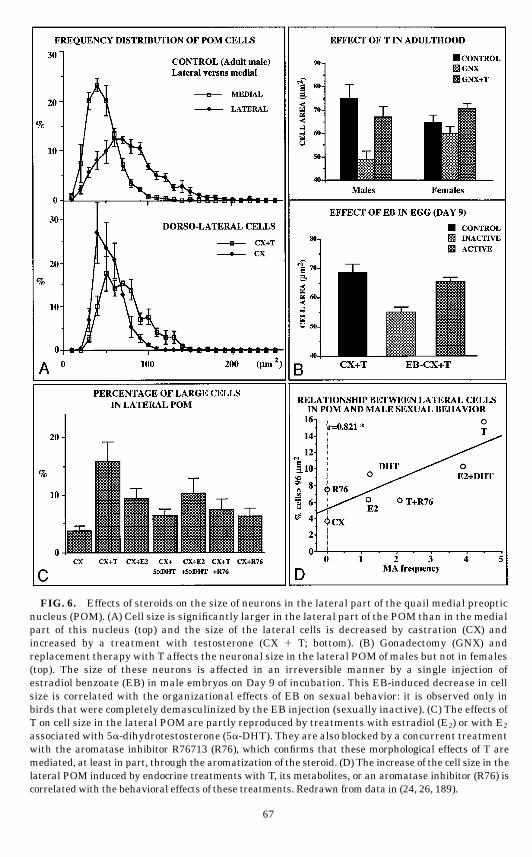
FIG. 6. Effects of steroids on the size of neurons in the lateral part of the quail medial preopticnucleus (POM). (A) Cell size is significantly larger in the lateral part of the POM than in the medialpart of this nucleus (top) and the size of the lateral cells is decreased by castration (CX) andincreased by a treatment with testosterone (CX 1 T; bottom). (B) Gonadectomy (GNX) andreplacement therapy with T affects the neuronal size in the lateral POM ofmales but not in females(top). The size of these neurons is affected in an irreversible manner by a single injection ofestradiol benzoate (EB) in male embryos on Day 9 of incubation. This EB-induced decrease in cellsize is correlated with the organizational effects of EB on sexual behavior: it is observed only inbirds that were completely demasculinized by the EB injection (sexually inactive). (C) The effects ofT on cell size in the lateral POM are partly reproduced by treatments with estradiol (E2) or with E2
associated with 5a-dihydrotestosterone (5a-DHT). They are also blocked by a concurrent treatmentwith the aromatase inhibitor R76713 (R76), which confirms that these morphological effects of T aremediated, at least in part, through the aromatization of the steroid. (D) The increase of the cell size in thelateral POM induced by endocrine treatments with T, its metabolites, or an aromatase inhibitor (R76) iscorrelatedwith the behavioral effects of these treatments. Redrawn fromdata in (24, 26, 189).
67
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

(189). The size of this lateral population of neurons was sensitive to variationsof T levels in males but not in females. The cross-sectional area of theseperikarya was significantly increased in castrated males treated with T (Figs.6A–6B), whereas they were not affected by such treatments in females (189).This differential response to T possibly represents a dimorphic feature in theorganizational sense (a population of neurons is T-sensitive in adult males andnot in females). This dimorphism correlates with the behavioral dimorphism: Tactivates copulatory behavior in males but not in females.Similarly, the comparison of quail exposed to long photoperiods or kept in short
days confirmed that high levels of circulating T associated with long days increasethe cell size within this subpopulation of POM in males, while changes of thephotoperiod have no effect in females (189). This effect of the photoperiod inmales isobserved in developing animals exposed from the age of 3–4 weeks to short or longdays (189), as well as in adult males that were previously fully mature and havebeen photoregressed (256). These experimental manipulations of the circulating Tlevels produce no significant change in cell size in themedial portion of the POM.
Treatment with EB in Egg
Because the previous study suggested that the sex difference in response to Tobserved in the lateral POM could have an organizational nature, we per-formed a morphometrical study of POM cells in quail that had been injectedwith EB during embryonic life. This treatment is known to permanentlydemasculinize copulatory behavior in male quail (see above). To ensure that thedifferences that could be observed would be of an organizational nature, allbirds were gonadectomized after hatching and treated with the same dose of Tto provide an optimal activation of both the copulatory behavior and the POMneurons. EB treatment on Embryonic Day 9 decreased the size of the dorsolat-eral POM neurons in males, but had no effect in females (26). No effect was alsoobserved when the treatment was performed on Day 14 (i.e., when it is notaffecting sexual behavior). The medial POM neurons were not affected. It isinteresting to note that, in this experiment, because we used a dose of EB (5 µg)that was just above the threshold for producing a full demasculinization, a fewmales that had been injected with EB in egg still displayed a low level of sexualactivity. In these birds the POM lateral population was not significantlydifferent from that of controls, i.e., the neuronal size had not been decreased(Fig. 6B, bottom). Therefore, it appears that the size of neurons in the lateralPOM is organized by estrogens in the embryo in parallel with copulatorybehavior. It is important to observe that this effect can be detected even if adultbirds are placed under similar endocrine conditions.
Endocrine Specificity in theAction of Steroids on Neuronal Size in Male POM
We also wondered whether T metabolism is critical for the increase in cellsize of the dorsolateral POM neurons, as it is for the activation of behavior and
68 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

for the increase in the total POM volume. We therefore compared the morpho-logical effects of T, 5a-DHT, and E2 given alone or in combination. To obtain anindependent evaluation of the role played by estrogens, morphological effects ofT associated or not with an aromatase inhibitor (R76713) were also compared(24). As previously observed these experimental treatments significantly af-fected the behavior frequencies. In particular, male sexual behavior was acti-vated by T and by the combined treatment with E2 and 5a-DHT. The effect of Twas partly inhibited by the concurrent administration of the aromatase inhibi-tor. An analysis of the distribution of cell sizes in the lateral part of the POMdemonstrated that the relative frequency of very large perikarya was signifi-cantly affected by the treatments. When we compared between groups thepercentage of large cells (.96 µm2 in cross-sectional area), a significant differ-ence with castrates was found only in birds treated with T or with E2 1 5a-DHT(Fig. 6C). This proportion of large cells in the dorsolateral POM in the differentgroups was strongly correlated with the intensity of the behavioral activation,which further suggested that this morphological parameter is closely related tothe mechanisms controlling male sexual behavior and its possible sex dimor-phism (Fig. 6D).
MECHANISMS UNDERLYING CHANGES IN POM SIZE
The analysis of cell sizes in the lateral POM provided a better correlate to thechanges in behavior induced by endocrine treatments but it also uncovered aproblem relative to the mechanism mediating the changes in POM volume inresponse to steroids. The size of the lateral neurons in POM increases by about36% (cross-sectional area) after treatment of castrated males with T. Thischange in cell size could theoretically be held responsible for the overall changein the volume of the nucleus: a 36% change in area corresponds to a 58% changein cell volume which is the same order of magnitude as the change in the totalvolume of the nucleus (189). In females by contrast, no significant effect of T onthe neuronal size could be detected, whereas previous studies showed that thePOM volume increases to values typical of sexually mature males after expo-sure to T (202). Studies in progress in our laboratory have suggested that theglial compartment of the dimorphic nucleus is also affected by steroid treat-ments and this could explain changes in the POM volume not associated withchanges in neuronal size (272) (N. Aste, C. Castagna, G. C. Panzica and J.Balthazart, unpublished data). There is evidence in other animal models thatthe size of glial cells is steroid-sensitive (123, 124, 253–255, 260). However, thedetail of the mechanism that mediates changes of the POM volume of male andfemale quail in response to T remains unclear at present.It is also important to notice that these changes in overall volume and in cell
size observed in POM in response to steroids were paralleled by variations inthe optical density of the staining obtained with dyes that specifically color theNissl substance. Nissl stains identify at the light microscopic level the highdensities of rough endoplasmic reticulum (RER) and of free polyribosomes.
69THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

Changes in Nissl staining hence reflect differences in the ultrastructuralorganization of the neurons that should presumably be associated with changesin protein synthesis.Quantitative measurement by image analysis of the Nissl stain in the POM
of male and female quail revealed decreases in mean optical density aftercastration and restoration of normal density levels after treatment with T (60).Semiquantitative evaluations confirmed this effect and, in addition, showedthat the effect of T can be mimicked by administration of T metabolites, inparticular E2, alone or in combination with 5a-DHT (24). These data suggestthat a profound reorganization of the POM ultrastructure should take placeafter exposure to T. They also raise the question of the nature of the protein(s)that is potentially synthesized in the POM neurons after treatment with thesteroid.
EFFECTS OF T ON THE ULTRASTRUCTURAL ORGANIZATION OF THE POM
The ultrastructure of the POM was recently described by electron micro-scopic techniques in male quail (200). The neurons of this nucleus generallyshow an extensive development of protein synthesis-related organelles (RER,polysomes) and of secretory structures (Golgi complexes, secretory vesicles,dense bodies). Studies at the light microscopical level demonstrated thatneurons in the lateral POM are larger than in the medial part of the nucleus(189). This was confirmed at the electron microscope level, but no other majordifference was observed in the ultrastructural organization of these neurons. Inboth the medial and the lateral part, the nucleus was characterized by a largevariety of cell bodies, including some that can be considered, on the basis oftheir ultrastructure, as putative peptidergic neurons. Various types of synapticendings were also present, suggesting a rich supply of nerve fibers. The generalmorphological features of these neurons were therefore comparable to thosedescribed for steroid-sensitive neurons in mammals (77, 158, 241). Few glialcells could be detected in the POM by this method. This confirms independentimmunocytochemical data showing that very few elements immunoreactive forthe glialmarkerGFAP (glial fibrillary acid protein) are present in the POM (86).The morphology of the POM neurons was affected by castration and T
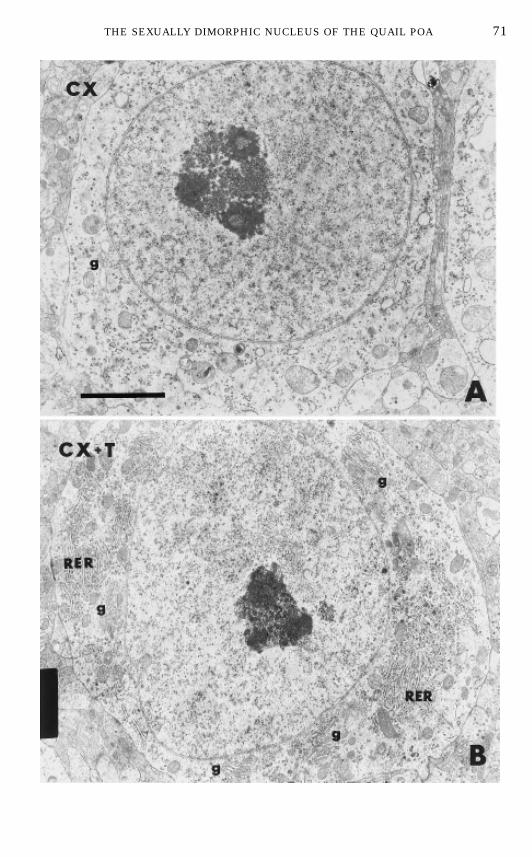
replacement therapy. In castrated birds, a marked decrease of intracytoplas-matic organelles was observed in the large majority of the lateral POM neurons(Fig. 7).This decrease concerned organelles involved in protein synthesis (polyribo-
somes, RER) and secretion (Golgi complexes, vesicles). These changes were
FIG. 7. Photomicrographs at the electronmicroscopic level of typical neurons in the lateral partof the medial preoptic nucleus of a castrated (CX) male quail (A) and of a castrated male that hadbeen treated with testosterone (CX 1 T) for 2 weeks (B). Magnification bar, 2 µm.Abbreviations: g,Golgi complexes; RER, rough endoplasmic reticulum.
70 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
71THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

quantitatively analyzed by stereological methods and were found to be statisti-cally significant in each case (197). Therefore, the changes in cell size and inintensity of Nissl staining that were previously detected at the light micro-scopic level reflect a true reorganization of the neurons. Because few glial cellsare present in the POM of male quail, it can also be concluded that thesteroid-induced plasticity in the POM is largely based on neuronal plasticity.Parallel studies were devoted to the neurochemical characterization of theseneurons and to the identification of the proteins whose synthesis could beactivated by steroids.
NEUROCHEMISTRY OF THE POM
The studies described above identified and characterized a sexually dimor-phic nucleus in the POA based primarily on material stained for the Nisslsubstance. They therefore localized an area that potentially showed a differen-tial rate of protein synthesis in males and females, but it was desirable toidentify independent markers that could single out the dimorphic population ofneurons from the rest of the POA. The morphological heterogeneity observed atthe ultrastructural level also suggested that the neurochemical organization ofthe POM was complex, and a full understanding of the functional significanceof this nucleus required the identification of the neurotransmitters and neuro-peptides present in this area.A broad survey of the neuroactive molecules present in POM was therefore
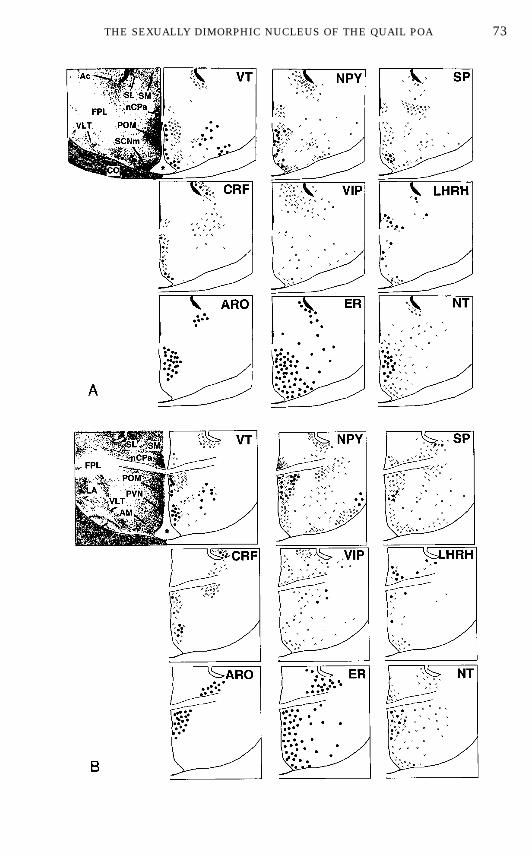
undertaken. At the present time, the presence of at least five different types ofpeptide-containing elements has been confirmed in POMby immunohistochem-istry (Fig. 8).This includes neurons containing neuropeptide Y (NPY) (27), substance P
(SP) (28), molluscan cardioactive tetrapeptide (122), luteinizing hormone-releasing hormone (LHRH) (121, 262), and neurotensin (NT) (2, 283). In
FIG. 8. Schematic drawings of the septal and preoptic regions of the male Japanese quailillustrating the neuroanatomical distribution of vasotocin (VT), neuropeptideY (NPY), substance P(SP), corticotropin-releasing factor (CRF), vasoactive intestinal polypeptide (VIP), luteinizinghormone-releasing hormone (LHRH), aromatase (ARO), estrogen receptors (ER), and neurotensin(NT). The drawings at the top left of each panel represent the arrangement of cell groups in thearea. The top part of the figure (A) represents a level corresponding approximately to the middlepart of the medial preoptic nucleus (POM) in the rostrocaudal axis and it is rostral to the levelshown in (B) that corresponds to the caudal part of the POMat the level of the anterior commissure.Black dots represent immunoreactive perikarya; small crosses represent positive fibers. Thenumber of symbols has been adjusted to represent relative abundance but the number of dots orfibers does not reflect exact numbers of cells of fibers. Abbreviations: Ac, nucleus accumbens; AM,nucleus anterior (rostralis) medialis hypothalami; CO, optic chiasma; FPL, fasciculus prosen-cephali lateralis; GLv, nucleus geniculatus lateralis, pars ventralis; LA, nucleus lateralis anteriorthalami; nCPA, nucleus commissurae pallii; POM, nucleus preopticus medialis; PVN, nucleusparaventricularis magnocellularis; SL, nucleus septalis lateralis; SM, nucleus septalis medialis;SCNm, nucleus suprachiasmaticus, pars medialis, VLT, nucleus ventrolateralis thalami.
72 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
73THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

addition, adjacent to the ependymal wall, a column of perikarya containingLHRH (262), vasotocin (VT) (269), or b-endorphin (263) can be visualized. Adense peptidergic innervation of the POM was also described in these studies,including fibers and punctate structures that were immunoreactive for VT(269), NT (2, 283), NPY (27), SP (28), and corticotrophin-releasing factor (CRF)(203, 284). Each of these peptides (cells and fibers) is heterogeneously distrib-uted in the POM and this distribution varies from one peptide to the other asillustrated schematically in Fig. 8 and described in more detail previously(192).A rich monoaminergic innervation of the POM was originally identified by
histochemistry (235) and later confirmed by immunohistochemistry. Fibersthat are immunoreactive (ir) for serotonin (5HT) (97) and for the catecholamine-synthesizing enzymes tyrosine hydroxylase (TH), the rate-limiting step indopamine (DA) synthesis (29), and dopamine b-hydroxylase (DBH), the limit-ing step in norepinephrine (NE) synthesis (29), were located in the nucleus.This innervation was also confirmed by biochemical assays (high-performanceliquid chromatography coupled with electrochemical detection; HPLC) thatshowed the presence of high concentrations of 5HT, DA, and NE in this nucleus(37, 50).Receptors for these neurotransmitters and hormones have also been identi-
fied in the POM by autoradiography. In particular, very high concentrations ofa2-adrenergic receptors are present in POM (32). By contrast, low levels ofa1-adrenergic (40), muscarinic cholinergic (33), D1 dopaminergic (31), melato-nin (96), GABAa (88), and benzodiazepine (89) receptors are found in themedial preoptic region.Neurochemical studies also confirm that the POM is a primary target for
steroids. The area contains binding sites for T, 5a-DHT, and E2 as demon-strated by autoradiography (276). More recent immunocytochemical studiesreveal the presence of large number of ER-immunoreactive cells (57) and of afew elements that contain immunoreactive AR (56). A high activity of thearomatase enzyme, a critical step in the synthesis of estrogens, has also beenlocalized in the POM (229). Accordingly, large numbers of aromatase-immunoreactive (ARO-ir) neurons are found in this nucleus (48, 55).
NEUROCHEMICAL MARKERS OF POM
Most of these peptides, transmitters, or receptors are present in the POM,but also in adjacent parts of the POA–anterior hypothalamus. In a limitednumber of cases, specific neurochemical markers of the POM were identified.ARO-ir cells (48), NT-ir cells (2), and VT-ir fibers (269) are specifically presentin the POM but absent in the surrounding POA. These neurochemical signalsoutline the boundaries of the dimorphic nucleus like a Nissl stain does. Inaddition, a2-adrenergic receptors are present in much higher densities in thePOM than in the rest of the POA and they also allow one to delineate theboundaries of the nucleus (32, 37, 60). ER-ir cells are present in larger number
74 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

in the POM than in the lateral POA but the difference in immunoreactive celldensity between the POM and its surrounding is not sharp enough to permit anidentification of the boundaries of the nucleus (57).
AromataseActivity andAromatase-Immunoreactive Cells
The critical importance of T aromatization in the brain for the activation ofmale sexual behavior has been established in a large number of experiments(see above). We therefore carried out a number of biochemical studies designedto characterize and localize the enzyme aromatase, which catalyzes this meta-bolic transformation. These experiments showed that a high level of aromataseactivity (AA) is present in the quail POA (228, 230). When a sexually dimorphicnucleus was identified in this region, further studies combining very sensitiveradioenzyme assays with microdissections by the ‘‘Palkovits’’ punch technique(187, 188) were undertaken to better specify the localization of AA in the POA.Very high levels of enzymatic activity were found in the dimorphic nucleus; therest of the POAdisplayed negligible AA (229).A few years ago, an immunocytochemical procedure based on a polyclonal
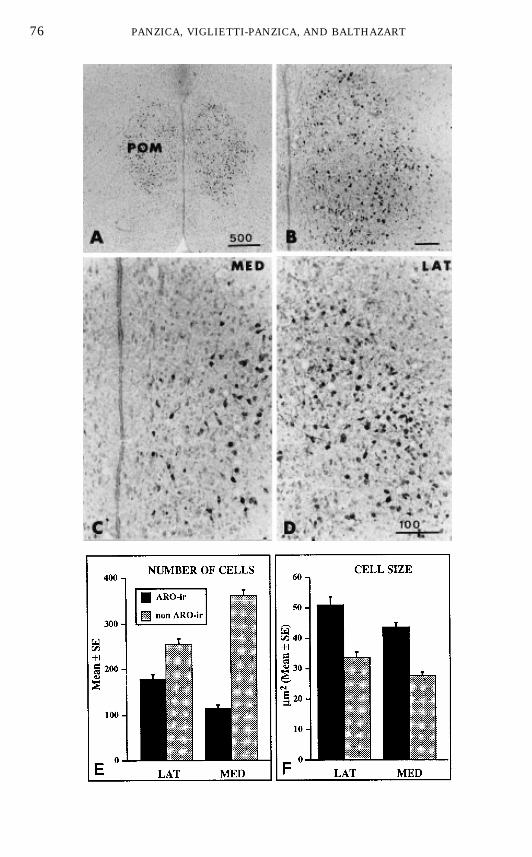
antibody raised against purified human aromatase (134) was set up to visualizeARO-ir cells in the quail brain (48). Four main groups of ARO-ir cells wereobserved by this technique, namely, in the POA, the septal region, at the level ofthe bed nucleus striae terminalis, and in the tuberal hypothalamus (48, 55).However, immunohistochemistry confirmed that all preoptic ARO-ir cells arelocalized within the cytoarchitectonic boundaries of the POM as defined inNissl-stained material. The dense cluster of ARO-ir cells located in the medialPOA actually delineates the boundaries of the dimorphic nucleus: as shown inFigs. 9A–9B, they fill the entire POM throughout its rostral to caudal extent(51).Morphometric analysis of these neurochemically defined cells (25) further
revealed that ARO-ir cells represent a large fraction of the total neuronspresent in the POM (about 40% of the total in the lateral POM; 20% in themedial part of the nucleus; Fig. 9E). The mean size of ARO-ir cells was alsofound to be larger in the lateral than in the medial POM (see Figs. 9C–9D, and9F). Morphological features suggest that all ARO-ir cells in the POM areneurons. This notion was confirmed by immunoelectron microscopic investiga-tions (173) (G. C. Panzica, C. Castagna, and P. J. Sharp, unpublished data).ER-immunoreactive cells had been identified in large numbers in the medial
POA (57). A double label immunocytochemical analysis of the spatial relation-ships between ER-ir and ARO-ir cells in the quail brain was therefore carriedout (52). This showed that, as expected for physiological reasons (52), mostARO-ir cells in the tuberal hypothalamus also contain immunoreactive ER(Fig. 10A). By contrast, this colocalization was unusual in the POM where only17% of ARO-ir neurons contained detectable levels of ER (Fig. 10B). This lowlevel of colocalization in POM has been independently confirmed (186).
75THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
76 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

Neurotensin
NT-ir cells are also specifically present within the boundaries of the POM (2,3, 283). In a way, their association with this nucleus is stronger than foraromatase because NT-ir cells could hardly be detected in other parts of thebrain (just a few weakly positive cells in the paraventricular nucleus and in thetuberal hypothalamus). However, the preoptic NT-ir cells are at least 10 timesless numerous than ARO-ir cells and therefore they do not permit to preciselydelineate the boundaries of the nucleus. Furthermore, they tend in many casesto be observed preferentially in the medial part of the nucleus. They areassociated with high densities of NT-ir fibers that fill the entire POM.It is interesting to note that the POM of females contains a larger number of
NT-ir cells than the POM of males (2). However, this sex difference disappearsafter castration and the treatment of gonadectomized males and females withexogenous T has no significant effect on the number of NT-ir cells in POM,although it increases the number of positive cells located in a position justcaudal to this nucleus (3). Therefore the sex difference observed in the POM ofintact sexually mature birds presumably reflects a larger induction by estro-gens of NT cells in females, which fits in well with the fact that estrogenreceptors have been colocalized with NT in preoptic neurons of mammals (139,140).
a2-Adrenergic Receptors
Autoradiographic studies using the a2-adrenergic ligand para-aminocloni-dine (PAC) demonstrate that the POM is specifically labeled by a high densityof a2-binding sites that allow the distinction of the nucleus from the surround-ing POA. This high density of receptors is visible throughout the rostral tocaudal extent of the nucleus and matches very well its boundaries as defined inNissl-stained material (32).
Vasotocin
VT-ir elements are one of the most studied neuropeptidergic systems of theavian brain. A large majority of VT-ir cells are located laterally or periventricu-
FIG. 9. Photomicrographs of aromatase-immunoreactive (ARO-ir) cells and quantitative analy-sis of these cells in the lateral and medial part of the medial preoptic nucleus (POM). (A–D) Low,medium, and high magnifications of sections through the POM stained by immunocytochemistryfor aromatase illustrating the facts thatARO-ir cells outline the entire POM (A, B) and thatARO-ircells are substantially smaller in the medial (MED; C) than in the lateral (LAT; D) part of thisnucleus. (E–F) Comparison of the number ofARO-ir cells and of other cells in themedial and lateralPOM (E) and comparison of the size of these two types of cells in the two part of the nucleus (F).Redrawn from data in (25).
77THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
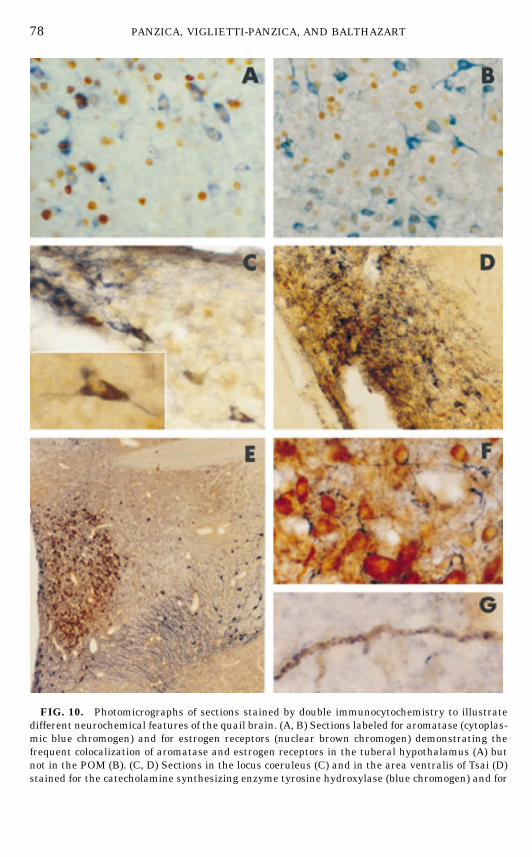
FIG. 10. Photomicrographs of sections stained by double immunocytochemistry to illustratedifferent neurochemical features of the quail brain. (A, B) Sections labeled for aromatase (cytoplas-mic blue chromogen) and for estrogen receptors (nuclear brown chromogen) demonstrating thefrequent colocalization of aromatase and estrogen receptors in the tuberal hypothalamus (A) butnot in the POM (B). (C, D) Sections in the locus coeruleus (C) and in the area ventralis of Tsai (D)stained for the catecholamine synthesizing enzyme tyrosine hydroxylase (blue chromogen) and for
78 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 root

larly in the preoptic and hypothalamic regions (251, 266, 270). VT ir-fibers areobserved in several intra- and extrahypothalamic regions (196). Very recently,we have examined both the lateral septum and the POM of the quail for aspecific innervation by this peptide (268, 269). It was found that VT-ir fibersspecifically label the POM region, with a higher density being present in thedorsolateral region of the nucleus. Close relationships between VT-ir fibers andARO-ir cell bodies or processes (Figs. 10F–10G) were also observed (267).Taken together these data clearly demonstrate that the POM can be distin-
guished from the surrounding POA by a variety of specific neurochemicalmarkers.
FUNCTIONAL STUDIES OF THE PREOPTIC AROMATASE
Because several of the POM neurochemical markers are potentially impli-cated in the control of male sexual behavior, a number of experimental studieswere performed to research whether the activity of these molecules (e.g.,neurotransmitter release, enzyme activity, receptor density) are sexually differ-entiated and/or modulated by steroids in a way that would parallel the changesin behavior. The study of aromatase was extremely enlightening in this respect.
Preoptic AromataseActivity
Radioenzyme assays measuring the in vitro conversion of T to E2 originallyrevealed that AA is sexually dimorphic (higher in males than in females) in thePOA (227). Gonadectomy reduced this enzyme activity to basal levels and,interestingly, a replacement therapy with T differentially restored the enzy-matic levels in males and females (228) so that gonadectomized T-treated birdshad a significantly higher AA than females (Fig. 11A), despite the fact thatbirds of both sexes were exposed to a similar endocrine milieu (42). Thisdifferential response to T is presumably organized by embryonic steroids, andexperimental studies carried out to test this hypothesis provided circumstan-tial evidence supporting this idea (36). However, this difference presumablyconcerns only a small part of the POA neurons and the assay technique thatwas used in these studies did not permit formal identification of an organiza-tional effect at this level.
cholera toxin, B subunit (CTB; brown dotted particles). The CTB stereotaxically injected in thePOM was retrogradely transported in these two areas and accumulated in part in tyrosinehydroxylase-immunoreactive cells, therefore demonstrating that these cells project to the POM.The inset in C shows a highermagnification of a cell containing both tyrosine hydroxylase and CTB.(E, F, G) Sections double labeled for aromatase (brown chromogen) and vasotocin (VT; bluechromogen), illustrating the general distribution of ARO-ir cells and VT-ir cells and fibers in thequail preoptic area (E) and the close association of VT-ir fibers with ARO-ir cells (F) and withARO-ir fibers (G) in the POM.
79THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
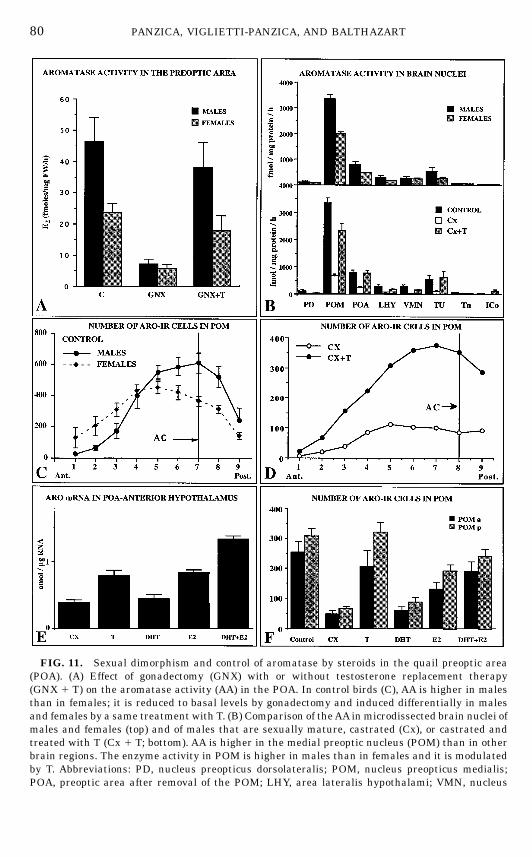
FIG. 11. Sexual dimorphism and control of aromatase by steroids in the quail preoptic area(POA). (A) Effect of gonadectomy (GNX) with or without testosterone replacement therapy(GNX 1 T) on the aromatase activity (AA) in the POA. In control birds (C), AA is higher in malesthan in females; it is reduced to basal levels by gonadectomy and induced differentially in malesand females by a same treatment with T. (B) Comparison of theAA inmicrodissected brain nuclei ofmales and females (top) and of males that are sexually mature, castrated (Cx), or castrated andtreated with T (Cx 1 T; bottom). AA is higher in the medial preoptic nucleus (POM) than in otherbrain regions. The enzyme activity in POM is higher in males than in females and it is modulatedby T. Abbreviations: PD, nucleus preopticus dorsolateralis; POM, nucleus preopticus medialis;POA, preoptic area after removal of the POM; LHY, area lateralis hypothalami; VMN, nucleus
80 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

The sexually differentiated induction of the preoptic AA by T is highlyrelevant to the control of copulatory behavior because gonadectomized T-treated females, which have a lower enzyme activity (228), also fail to copulate(65), and it is known that the aromatization of T into E2 is a limiting step for theactivation of the behavior (46, 47, 49). It must be noted, however, that treat-ment of ovariectomized females with estrogens (which should bypass thelimiting step) still fails to activate male-type sexual behavior (226). The sexdifference in preoptic AA may therefore contribute to the sex difference inbehavior but cannot explain it alone. It is also important to note that theendocrine specificity of the AA induction in castrated male quail is identical tothe endocrine specificity of the behavioral activation: the preoptic AA is in-creased by T or by androgens and estrogens working in synergy (43, 49, 225).This brings further support to the idea that aromatase can be a limiting step forthe activation of behavior.More detailed studies performed on microsamples dissected by the ‘‘Palko-
vits’’ punch technique (187, 188) have shown that the changes in AA describedfor the POAactually concern almost exclusively the enzyme located in the POM(63): AA in the POM is higher in males than in females; it is reduced bycastration and increased by T (Fig. 11B).
Aromatase-Immunoreactive Cells
The changes in AAdescribed above could theoretically result from variationsin the enzymatic activity of a stable number of molecules (regulation byactivators or inhibitors), in the numbers of cells that express the enzyme, or inthe concentration of enzyme in a stable number of aromatase-containing cells,or from any combination of these mechanisms (46, 47). To discriminate amongthese possibilities, ARO-ir cells were analyzed during semiquantitative immu-
ventromedialis hypothalami; TU, tuber; Tn, nucleus taeniae; ICo, nucleus intercollicularis. (C)Distribution of aromatase-immunoreactive (ARO-ir) cells in the medial preoptic nucleus (POM) ofmale and female quail. Nine consecutive sections that were 100 µm apart were stained in the POM.They were realigned for different birds using the organum vasculosum laminae terminalis and theanterior commissure (AC) as landmarks. Section 7 corresponds to the first section in which the fullAC is visible. MoreARO-ir cells are present in males than in females in the sections around theAC.(D) Effects of testosterone on the number of ARO-ir cells in the POM of male quail. Data werecollected as described for C. Birds were either castrated (CX) or castrated and treated with one20-mm-long Silastic implant filled with testosterone (CX 1 T). (E) Effect of testosterone (T) or itsandrogenic (dihydrotestosterone; DHT) and estrogenic (estradiol; E2) metabolites given alone or incombination on the concentration of aromatasemRNAin the preoptic area–anterior hypothalamus.An increase in mRNA concentration was observed after treatment with T, E2, or E2 1 DHT. (F)Effect of testosterone (T) or its metabolites (DHT and E2) given alone or in combination on thenumber of ARO-ir cells in the anterior and posterior parts of the POM (POM a and POM p,respectively). Castration decreased the number ofARO-ir cells by comparison with control sexuallymature birds. A partial or full restoration was observed after treatment with T, E2, or E2 1 DHT.Redrawn from data in (63, 118, 135, 228).
81THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

nocytochemical studies. These experiments established that the changes inenzyme activity mostly reflect differences in the number of cells that expressaromatase under different physiological conditions.The higherAA in the POM of males compared to that in females is paralleled
by the presence of a higher number ofARO-ir cells in the former (Fig. 11C). Thissex difference is actually restricted to the part of the dimorphic nucleus locatedjust rostral to the anterior commissure (118). This may explain why biochemi-cal studies considering the entire POA or even the POM sometimes failed toestablish a sex difference in enzyme activity. Castration reduces the number ofcells that can be visualized by aromatase immunocytochemistry (118), and Treplacement increases this number remarkably (four- to fivefold increase; seeFigs. 11D and 12).This increase in the number of ARO-ir cells after exposure to T has approxi-
mately the same magnitude as the increase in preoptic AA induced by the sametreatment, which suggests that the change in enzyme activity results mostly, ifnot exclusively, from a change in the number of cells that express the enzyme(51). At this level also, the effects of T can be mimicked by its metabolites: acombined treatment with E2 and 5a-DHT (Fig. 11F) or with a combination ofpure androgens (methyltrienolone) and estrogens (diethylstilbestrol) alsomark-edly increases the number of ARO-ir cells (67, 135).Aromatase immunocytochemistry was also used to better localize, at a
cellular level of resolution, the parts of the POA that are implicated in thecontrol of copulatory behavior (70). Electrolytic lesions of the POA disruptedthe activation of male sexual behavior by T only if they destroyed a significantpart of the ARO-ir cells in the POM. Stereotaxic T implants in or close to POMactivatedmale copulatory behavior and increased the number ofARO-ir cells inPOM. These neuroanatomical effects were more prominent on the side of theimplant but they were also detected on the contralateral side. Correlativeanalyses suggested that a part of the POM just rostral to the anterior commis-sure is critical for the activation of copulatory behavior. The best correlationbetween the behavioral deficit induced by electrolytic lesions and the size of thelesion was indeed observed in this area. In addition, high correlations wereobserved between the intensity of the behavior activated by T implants and thenumber of ARO-ir cells that were induced by T.
Aromatase mRNA
The changes in aromatase described by immunocytochemistry strongly sug-gested that the enzyme concentration changes in the quail brain as a function
FIG. 12. Photomicrographs illustrating the effects of castration (CX) with or without replace-ment therapy by testosterone (CX 1 T) on the aromatase-immunoreactive (ARO-ir) cells in thequail POM.A, C, and E present lowmagnifications of the entire POMwhile B, D, and F show highermagnifications of lateral (LAT)ARO-ir cells to demonstrate the changes in cell size produced by theexperimental treatments. Magnification bars are 500 µm (left column) and 100 µm (right column).
82 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
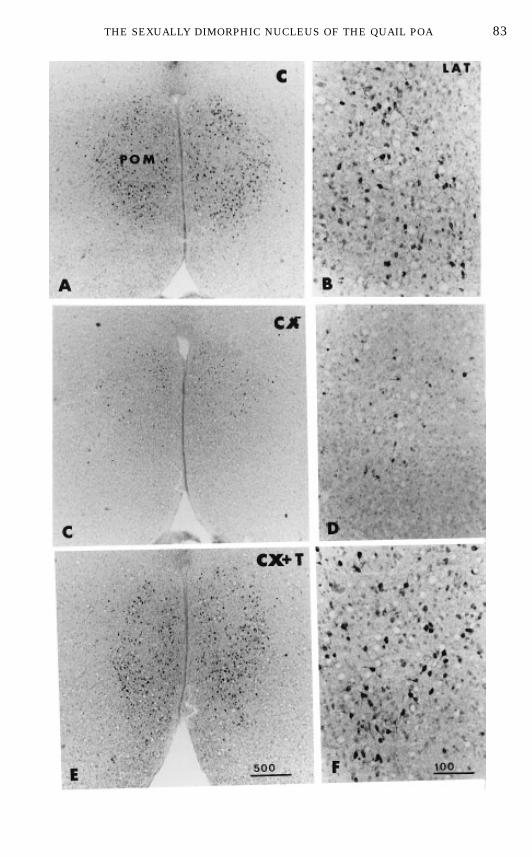
83THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
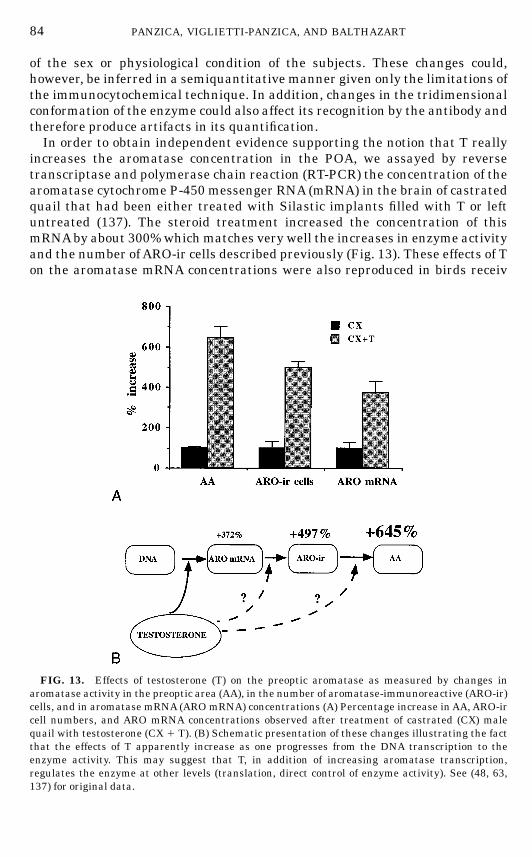
of the sex or physiological condition of the subjects. These changes could,however, be inferred in a semiquantitative manner given only the limitations ofthe immunocytochemical technique. In addition, changes in the tridimensionalconformation of the enzyme could also affect its recognition by the antibody andtherefore produce artifacts in its quantification.In order to obtain independent evidence supporting the notion that T really
increases the aromatase concentration in the POA, we assayed by reversetranscriptase and polymerase chain reaction (RT-PCR) the concentration of thearomatase cytochrome P-450 messenger RNA (mRNA) in the brain of castratedquail that had been either treated with Silastic implants filled with T or leftuntreated (137). The steroid treatment increased the concentration of thismRNAby about 300%whichmatches very well the increases in enzyme activityand the number ofARO-ir cells described previously (Fig. 13). These effects of Ton the aromatase mRNA concentrations were also reproduced in birds receiv
FIG. 13. Effects of testosterone (T) on the preoptic aromatase as measured by changes inaromatase activity in the preoptic area (AA), in the number of aromatase-immunoreactive (ARO-ir)cells, and in aromatase mRNA (AROmRNA) concentrations (A) Percentage increase inAA, ARO-ircell numbers, and ARO mRNA concentrations observed after treatment of castrated (CX) malequail with testosterone (CX 1 T). (B) Schematic presentation of these changes illustrating the factthat the effects of T apparently increase as one progresses from the DNA transcription to theenzyme activity. This may suggest that T, in addition of increasing aromatase transcription,regulates the enzyme at other levels (translation, direct control of enzyme activity). See (48, 63,137) for original data.
84 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

ing a combined treatment with the behaviorally active metabolites E2 and5a-DHT (135) (see Fig. 11E).The effects of T treatments on AA, on the number of ARO-ir cells, and on the
aromatase mRNA concentrations therefore have a very similar magnitude (3.7to 6.4 times). This suggests that an increase in aromatase transcription couldbe the only mechanism implicated in the regulation of the enzyme activity by T.It must be noticed, however, that the magnitude of the T effect increases as oneprogresses from the DNA transcription to the enzymatic activity (Fig. 13B).This might represent only experimental artifacts related to methodologicalproblems arising, for example, from the comparison of quantitative dataobtained through the assay of microdissected samples to neurochemical dataderived by semiquantitative methods from histological sections. Alternatively,this could suggest that additional mechanisms under the control of T modulatethe translation of the mRNA into protein and the activity of the enzyme.
ARO-IR CELLS AS THE MAIN TARGET FOR STEROIDS IN THE POM
As previously mentioned, detailed analyses of the size of ARO-ir and nonim-munoreactive cells in the POMof sexuallymaturemales revealed that immuno-reactive perikarya are, as amean, significantly larger that the nonimmunoreac-tive ones (25). This essentially results from an overrepresentation of theARO-ircells in the large cells classes as illustrated in Fig. 14.More than 75% of the cells that have a cross-sectional area larger than 50
µm2 are immunoreactive for aromatase, while this percentage is negligible insmall cells. This differential representation of ARO-ir cells in the small andlarge cell classes has important consequences for the interpretation of thestudies of Nissl-stained material that have been presented above. We previ-ously indicated that T affects the cell size in the lateral but not in the medialPOM of male quail and that this effect is not seen in females (189). Thissuggests that the sensitivity of the lateral population of POM cells to T issexually differentiated. This idea is also supported by the observation thattreatment of quail eggs with EB permanently decreases the mean cell size inthe lateral POM in male birds (26), essentially because the proportion of largecells decreases and the distribution curve is skewed to the left. Because a largemajority of the big neurons in the lateral POM are in fact aromatase-containingcells (see Figs. 14A–14B), it can be speculated that the effects of steroidmanipulations previously observed in Nissl-stained material primarily affectthese aromatase cells. It is therefore likely thatARO-ir cells in the lateral POMare not sensitive to T in females and that estrogens in the embryo areresponsible for the development of this sexually dimorphic characteristic. Thiscould represent an important part of the sexually differentiated brain mecha-nisms responsible for the activation of male copulatory behavior. It is indeedwell established that females do not show copulatory behavior when exposed toT in adulthood (6, 10, 65, 226) and that the induction of aromatase activity inthe male POA represents a limiting factor for the activation of the behavior in
85THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

males (46, 49). This idea is currently under experimental investigation in astudy of the size of ARO-ir cells in birds exposed to the embryonic endocrinetreatments that are known to permanently affect behavior.Detailedmorphometric studies also revealed that steroids affect in a differen-
tial manner the ARO-ir cells located in the medial or lateral parts of thedimorphic nucleus (25). The mean cell size decreased more after castration inthe medial than in the lateral POM (Fig. 14C). In addition, after castration, thenumber of ARO-ir cells decreased by about 90% in the lateral POM, whereasthis decrease in number was only about 70% in the medial POM (Fig. 14D). Allof these changes were reversed by a treatment with T. It may seem surprisingthat in previous studies T and its metabolites affected cell size in the lateral butnot in the medial POM (24, 189), whereas we demonstrate here that the size of
FIG. 14. Frequency distribution of cells sizes for aromatase-immunoreactive (ARO-ir) and fornonimmunoreactive cells in the lateral (A) andmedial (B) parts of the POMand effects of castration(CX) with or without testosterone replacement (CX 1 T) on the size (C) and number (D) of ARO-ircells in the medial and lateral parts of the nucleus. Redrawn from data in (25).
86 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

ARO-ir cells is affected by these steroids in both parts of the nucleus (25).However,ARO-ir cells represent only about 20% of the cells in the medial POM,while they constitute a larger fraction of the cell population in the lateral POM.The effect onARO-ir cells in themedial part of the nucleus was probably dilutedin a larger population that is presumably not responsive to steroids.The differential effects of the steroid treatments observed in the medial and
lateral parts of the nucleus identify, we believe, the presence of two types ofaromatase cells in the quail POA (25). A small population of ARO-ir cells ispresent in the brain independent of the steroid environment and this repre-sents at least one-third of the cells that are found in the medial part of thePOM. Other larger ARO-ir cells are by contrast exquisitely sensitive to steroidstimulation and they almost completely disappear in castrated birds. Theywould be located mostly in the lateral part of the sexually dimorphic nucleus.The increase in aromatase activity that is observed after exposure to T or itsmetabolite E2 could then result from the combination of two processes: anincrease in the expression of aromatase (suggested by the increase in cell area)in a relatively stable number of medial small cells and a large increase in thenumber of larger cells located mainly in the lateral POM. This differentiallocalization of ‘‘steroid-inducible’’ and ‘‘steroid-insensitive’’ ARO-ir cells in themedial and lateral POM is clearly suggested in Fig. 14: the medial POMcontains more ARO-ir cells than the lateral POM in birds that are not stimu-lated by steroids (CX), while the opposite is true in all other groups (Controland CX 1 T). Most of these effects of T on ARO-ir cells could be reproduced bytreatment with either E2 alone or E2 in combination with 5a-DHT (25). Furtherstudies investigated the ARO-ir system during male quail aging, when thespontaneous decrease in T levels determines a parallel decrease in sexualperformance(s). In aging birds, a significant reduction of the ARO-ir cellnumber was also observed, confirming that the reduction of circulating T bothin the young and in the old animals specifically influence the ARO-ir system(108, 191).
CONNECTIONS OF THE POM
A full understanding of the mechanism underlying male sexual activityrequires the identification of the neural circuitry linked to this behavior and inparticular of the afferents and efferents of the POM. As a first step in thisresearch, in vitro tracing studies with the lipophilic fluorescent tracer dioctadec-yltetramethyl-indocarbocyanine (DiI) demonstrated a number of bidirectionalconnections between the POM and several hypothalamic and thalamic nuclei,including the nucleus dorsolateralis anterior thalami (44, 195).DiI implantation in aldehyde-fixed tissue demonstrated anterograde projec-
tions from the POM to the tuberal hypothalamus, the area ventralis of Tsai,and the substantia grisea centralis (Fig. 15). Dense networks of fluorescentfibers were also seen in several hypothalamic nuclei, such as the anteriormedialis hypothalami, the paraventricularis magnocellularis, and the ventro-
87THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

medialis hypothalami. A major projection in the dorsal direction was alsoobserved from the POM toward the nucleus septalis lateralis and medialis.Fluorescent cells were seen in all these areas, demonstrating that the POM
receives afferent projections from all these regions. Implantation of DiI into thesubstantia grisea centralis also revealed massive bidirectional connectionswith a large number of more caudal mesencephalic and pontine structures. Thesubstantia grisea centralis therefore appears to be an important center connect-ing anterior levels of the brain to brain stem nuclei that may be involved in thecontrol of male copulatory behavior. Most of these bidirectional projectionscould be confirmed by implanting DiI in the identified targets of POM andobserving fluorescent label in POM (cells or fibers). These data have beenpublished in full detail (44). After implantation of DiI in the POM, fluorescentfibers but no fluorescent cells were seen in the nucleus intercollicularis (ICo),suggesting the presence of a unidirectional pathway connecting the POM toICo. However, no experiment so far has confirmed the existence of this connec-tion by applying a retrograde tracer in ICo.In vitro tracing with DiI has technical limitations. In particular, this proce-
dure does not easily identify long distance projections. It is also quite difficult todouble label sections for specific neurochemical markers. Additional studieswere therefore carried out with the tracer cholera toxin B-subunit (CTB) in
FIG. 15. Schematic drawing illustrating, in a parasagittal plane, the afferent and efferentconnections of the medial preoptic nucleus (POM). The POM is represented by the darkly dottedarea. It is bidirectionally connected with all other nuclei indicated in the figure except for thenucleus intercollicularis (efferent projection only). Abbreviations: AM, nucleus anterior (rostralis)medialis hypothalami; AVT, area ventralis of Tsai; GCt, substantia grisea centralis; ICo, nucleusintercollicularis; Loc, locus coeruleus; PVN, nucleus paraventricularismagnocellularis; Scv, nucleussubcoeruleus ventralis; SL, nucleus septalis lateralis; SM, nucleus septalis medialis; Tu, tuber.Redrawn from data in (44).
88 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

order to obtain more information on more distant connections of the POM andto identify the chemical nature of some of the afferent projections (5) (P. Absiland J. Balthazart, unpublished observations). This technique confirmed all theconnections that had been identified with DiI. In addition, a strong input fromthe rostral part of the nucleus taeniae to the POMwas identified. Furthermore,large numbers of retrogradely labeled cells were seen in themajor catecholamin-ergic cell groups, namely, the area ventralis of Tsai (AVT), the nucleus tegmento-pedunculo pontinus, and the locus coeruleus. Double labeling of these cells byimmunocytochemistry for tyrosine hydroxylase, the rate-limiting enzyme incatecholamine synthesis, showed that some of these neurons projecting to POMare in fact catecholaminergic in nature, especially in AVT and in the locuscoeruleus (Figs. 10C–10D).The catecholaminergic projections to the POMare also confirmed by immuno-
cytochemical data demonstrating the presence of TH-ir and DBH-ir fibers inthis nucleus. Double label immunocytochemical studies in which the ARO-ircells and these catecholaminergic fibers were simultaneously visualized (30)demonstrated that both THandDBHfibers apparently innervate the aromatase-containing cells (close association of ARO-ir cells with positive fibers and/orpunctate structures). Interestingly similar contacts have recently been ob-served between VT-ir fibers and ARO-ir cells in the POM (see Figs. 10F–10G).The origin of the vasotocinergic projections has, however, not yet been identi-fied (267).
NEUROCHEMICAL INPUTS TO THE POM: RELATIONSHIPS WITH BEHAVIOR
Catecholamines
In mammals, the involvement of catecholamines in the control of reproduc-tive behavior has been firmly established (103, 104, 161, 164, 166, 167). Theidentification of TH- and DBH-immunoreactive fibers in the POM (see above)suggested that these transmitters could also modulate male sexual behavior inquail. Therefore biochemical and behavioral studies have been undertaken totest this hypothesis. Spectrofluorometric assays first identified the presence ofhigh concentrations of NE and DA in the POA (185). Interestingly, this studyshowed that the NE concentration in the POA was slightly but significantlyhigher in females than in males.This sex dimorphism in NE concentration was confirmed in a more recent
study using HPLC (50). The higher sensitivity of this method also allowed us todissect smaller brain samples by the ‘‘Palkovits’’ punch method (187, 188) andto show that the sexually differentiatedNE concentration is specifically presentin the POM (Fig. 16A). This sex difference is also supported by recent immuno-cytochemical studies showing that the area covered by DBH-ir fibers in themedial POA is wider in females than in males (29).Systemic injections of the noradrenergic neurotoxinN-(2-chlorethyl)-N-ethyl-
2-bromobenzylamine hydrochloride (DSP4) significantly depleted NE concen-
89THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
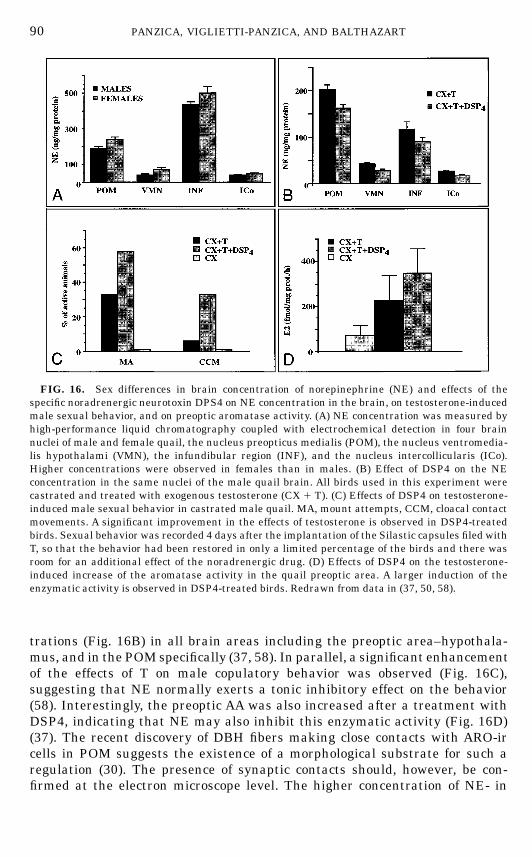
trations (Fig. 16B) in all brain areas including the preoptic area–hypothala-mus, and in the POM specifically (37, 58). In parallel, a significant enhancementof the effects of T on male copulatory behavior was observed (Fig. 16C),suggesting that NE normally exerts a tonic inhibitory effect on the behavior(58). Interestingly, the preoptic AA was also increased after a treatment withDSP4, indicating that NE may also inhibit this enzymatic activity (Fig. 16D)(37). The recent discovery of DBH fibers making close contacts with ARO-ircells in POM suggests the existence of a morphological substrate for such aregulation (30). The presence of synaptic contacts should, however, be con-firmed at the electron microscope level. The higher concentration of NE- in
FIG. 16. Sex differences in brain concentration of norepinephrine (NE) and effects of thespecific noradrenergic neurotoxin DPS4 on NE concentration in the brain, on testosterone-inducedmale sexual behavior, and on preoptic aromatase activity. (A) NE concentration was measured byhigh-performance liquid chromatography coupled with electrochemical detection in four brainnuclei of male and female quail, the nucleus preopticus medialis (POM), the nucleus ventromedia-lis hypothalami (VMN), the infundibular region (INF), and the nucleus intercollicularis (ICo).Higher concentrations were observed in females than in males. (B) Effect of DSP4 on the NEconcentration in the same nuclei of the male quail brain. All birds used in this experiment werecastrated and treated with exogenous testosterone (CX 1 T). (C) Effects of DSP4 on testosterone-induced male sexual behavior in castrated male quail. MA, mount attempts, CCM, cloacal contactmovements. A significant improvement in the effects of testosterone is observed in DSP4-treatedbirds. Sexual behavior was recorded 4 days after the implantation of the Silastic capsules filed withT, so that the behavior had been restored in only a limited percentage of the birds and there wasroom for an additional effect of the noradrenergic drug. (D) Effects of DSP4 on the testosterone-induced increase of the aromatase activity in the quail preoptic area. A larger induction of theenzymatic activity is observed in DSP4-treated birds. Redrawn from data in (37, 50, 58).
90 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

females may therefore contribute to the sex difference in copulatory behaviorand in preoptic aromatase activity (both being inhibited more by NE in femalesthan in males). The POM appears as the most likely site for these regulationsbecause: (a) the preoptic aromatase is exclusively located in this nucleus (48,55, 63, 229), (b) NE concentrations are far higher in the POM than in the rest ofthe POA (50), and (c) the POM is distinguished from the rest of the POA by amuch higher concentration of a2-adrenergic receptors (32). However, no sexdifference in the density of these receptors in POM could be identified (32, 60).DAand its turnover have also been measured in the brain of male and female
quail in several independent studies, using either spectrofluorometric assays(183, 185) or HPLC coupled with electrochemical detection (50). No reliable sexdifference in the baseline level of DA could be detected in the POAbut turnoverstudies showed that the disappearance of DA after one injection of the THinhibitor a-methylparatyrosine (aMPT) was much faster in males than infemales in the POA (183) and also specifically in the POM (50). DA disappear-ance is supposed to reflect the turnover of the amine. This measure wassexually differentiated in sexually mature subjects but also in birds that hadbeen gonadectomized with or without Silastic implants of T (Figs. 17A–17B).Usually, no depletion of DA was found in females after an aMPT injection
except possibly in T-treated ovariectomized subjects, in which a minimaldecrease (still lower than in males submitted to the same treatment) wasobserved. The fact that the sex difference in DA turnover was still observed inmales and females placed under similar endocrine conditions (e.g., in gonadec-tomized subjects) strongly supports the idea that this difference is organiza-tional in nature, but this notion has not yet been experimentally tested.This potentially organized sex difference could have an important physiologi-
cal and behavioral significance. It is indeed well established that DAstimulatesmale copulatory behavior in rats (74, 75, 164). A sexually differentiated controlby DA could therefore be part of the mechanisms that mediate the sex differ-ence in quail behavior. It is already established that experimental manipula-tions of the dopaminergic transmission affect male behavior in quail (4). Inparticular, ip injections of apomorphine, a general dopaminergic agonist, mark-edly reduce the frequency of these behaviors in castrated males treated with T(Fig. 17C). It has, by contrast, consistently been shown in mammals thatapomorphine stimulates male behavior (74, 75, 222).As discussed previously inmore detail (4), several reasons could explain this discrepancy but it can just bementioned here that, since apomorphine acts on both D1- and D2-like receptorsand these are supposed to have opposite effects on sexual behavior (74, 75, 103,164), a differential representation of these two receptor subtypes in birds andmammals could explain why apomorphine has opposite effects in these twovertebrate classes. In support of this interpretation, it can be said that therelative proportion of D2 (inhibiting cAMP production) to D1 (stimulatingcAMP production) receptors may indeed be higher in birds than in mammals(216). Preliminary experiments (4) have also shown that in quail, systemicinjections of the D2 antagonist spiperone enhance the copulatory behavior
91THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
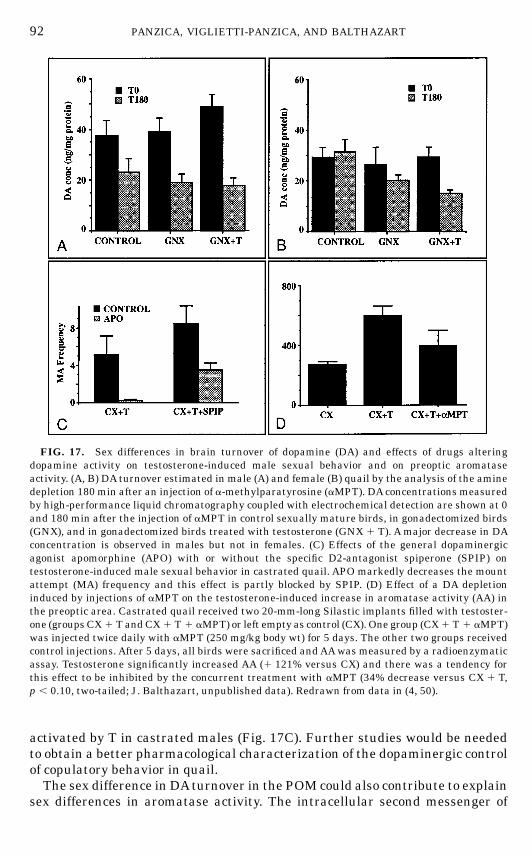
activated by T in castrated males (Fig. 17C). Further studies would be neededto obtain a better pharmacological characterization of the dopaminergic controlof copulatory behavior in quail.The sex difference in DAturnover in the POM could also contribute to explain
sex differences in aromatase activity. The intracellular second messenger of
FIG. 17. Sex differences in brain turnover of dopamine (DA) and effects of drugs alteringdopamine activity on testosterone-induced male sexual behavior and on preoptic aromataseactivity. (A, B) DA turnover estimated in male (A) and female (B) quail by the analysis of the aminedepletion 180 min after an injection of a-methylparatyrosine (aMPT). DAconcentrations measuredby high-performance liquid chromatography coupled with electrochemical detection are shown at 0and 180 min after the injection of aMPT in control sexually mature birds, in gonadectomized birds(GNX), and in gonadectomized birds treated with testosterone (GNX 1 T). Amajor decrease in DAconcentration is observed in males but not in females. (C) Effects of the general dopaminergicagonist apomorphine (APO) with or without the specific D2-antagonist spiperone (SPIP) ontestosterone-induced male sexual behavior in castrated quail. APO markedly decreases the mountattempt (MA) frequency and this effect is partly blocked by SPIP. (D) Effect of a DA depletioninduced by injections of aMPT on the testosterone-induced increase in aromatase activity (AA) inthe preoptic area. Castrated quail received two 20-mm-long Silastic implants filled with testoster-one (groupsCX 1 TandCX 1 T 1 aMPT) or left empty as control (CX). One group (CX1 T 1 aMPT)was injected twice daily with aMPT (250 mg/kg body wt) for 5 days. The other two groups receivedcontrol injections. After 5 days, all birds were sacrificed andAAwas measured by a radioenzymaticassay. Testosterone significantly increased AA (1 121% versus CX) and there was a tendency forthis effect to be inhibited by the concurrent treatment with aMPT (34% decrease versus CX 1 T,p , 0.10, two-tailed; J. Balthazart, unpublished data). Redrawn from data in (4, 50).
92 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

D1-like dopaminergic receptors, cAMP (126), is known to modulate aromataseactivity in several tissues, including the brain (85, 90, 165, 264, 265). It cantherefore be hypothesized that dopaminergic activity significantly affects aro-matase activity in the brain through the modulation of adenylate cyclase by theD1 receptor subtype. This notion is supported bymorphological data demonstrat-ing the presence of catecholaminergic terminals [tyrosine hydroxylase (TH)-immunoreactive] that surround ARO-ir cells in the POM (30). Preliminarypharmacological data also support this hypothesis. Depletion of brain DA levelsby daily injections of aMPT significantly inhibits the increase in AA that isinduced by T in castrated birds (Fig. 17D, J. Balthazart, unpublished data).Since chronic treatment with aMPT decreases both NE (which presumablyinhibits AA in the quail POA based on the DSP4 experiments described above)and DA (50), it can be expected that a selective dopaminergic inhibition wouldlead to an even larger decrease in enzymatic activity. These data are thereforeconsistent with the notion that DAup-regulates brain aromatase, in particularin the POM which contains all the ARO-ir cells of the POA. As discussedelsewhere in more detail (38), this could be one way in which DA modulatesbehavior.In conclusion, available data strongly suggest that both the noradrenergic
and the dopaminergic inputs to the POM have profound effects on the physiol-ogy of the nucleus and in particular modulate the aromatase activity andcopulatory behavior. However, more pharmacological and morphological stud-ies are needed to characterize these effects.
Vasotocin
Among the neuropeptidergic systems that were immunocytochemically de-tected in the POM, VT seems to be of particular interest. The fibers containingthis peptide are distributed along the entire extension of the nucleus, andrecent immunohistochemical data (267) demonstrate a close relationship be-tween them and ARO-ir cell bodies (Figs. 10F–10G). In several species ofmammals, a T-dependent sex dimorphism was observed in regions such as thelateral septum (106, 107). In birds, this was also demonstrated in canaries(273) and quail (268). Additionally, we have recently discovered that thevasotocinergic innervation of the POM is also T-dependent (Fig. 18): a higherdensity of VT-positive fibers is present in intact and castrated males treatedwith T than in untreated castrated males (269).POM neurons could therefore be indirectly regulated by T acting through its
vasotocinergic afferents to the nucleus. These inputs could be very relevant tothe control of male sexual behavior. It is now well established that VT or itshomolog, vasopressin, modulate a wide variety of social behaviors in verte-brates (82, 106, 107, 168). In birds specifically, Kihlstrom and Dannige (150)demonstrated a short-term increase of mating behavior in male chickens andpigeons after systemic administration of VT and oxytocin. More recently, it hasalso been reported that systemic injections of VT or a VT antagonist modulate
93THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
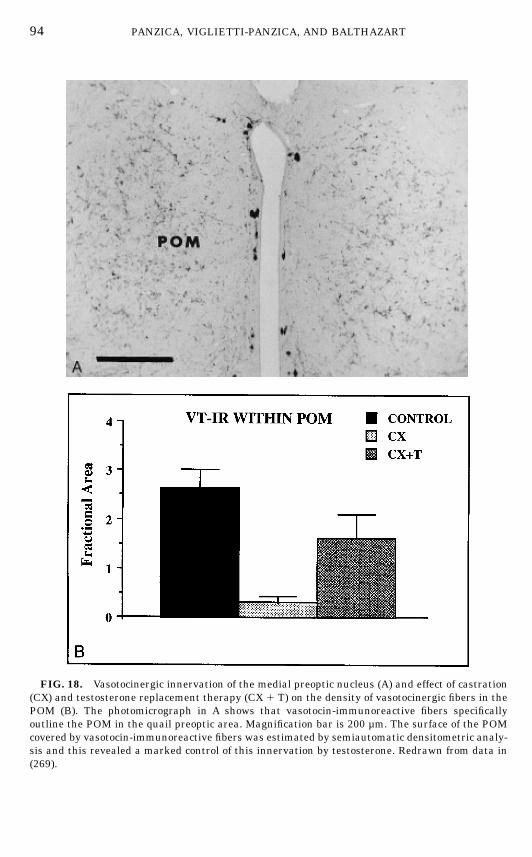
FIG. 18. Vasotocinergic innervation of the medial preoptic nucleus (A) and effect of castration(CX) and testosterone replacement therapy (CX 1 T) on the density of vasotocinergic fibers in thePOM (B). The photomicrograph in A shows that vasotocin-immunoreactive fibers specificallyoutline the POM in the quail preoptic area. Magnification bar is 200 µm. The surface of the POMcovered by vasotocin-immunoreactive fibers was estimated by semiautomatic densitometric analy-sis and this revealed a marked control of this innervation by testosterone. Redrawn from data in(269).
94 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

the appetitive components of male sexual behavior in quail (73). The behavioralpharmacology of VT should therefore be investigated in quail.
DISCUSSION
In summary, the data that have been reviewed above allow us to draw 10main conclusions concerning the dimorphic nucleus of the quail POA and itsfunctional significance.1. The quail POM volume is steroid-sensitive in adults and the sex difference
in the volume of this nucleus reflects the different steroid milieu in adult malesand females, not a difference organized by early steroids.2. Lesion and implantation studies demonstrate that the POM is one of the
main centers involved in the control of copulation. Steroid action in the POM isnecessary and sufficient for the activation of male copulatory behavior.3. The cell population of the POM, as studied by conventional staining
techniques, is heterogeneous and the size of cells situated in the dorsolateralpart of the nucleus is sexually differentiated in the organizational sense.4. The endocrine specificity of the changes observed in the POM volume and
in the size of its lateral cells is the same as the specificity described for theactivation of sexual behavior in males. Both groups of responses are stimulatedby a synergistic action of androgenic (5a-DHT) and estrogenic (E2) metabolitesof T.5. The T-induced changes in the size of lateral cells reflect the ultrastructural
reorganization of these elements, indicating that subcellular compartmentsinvolved in protein synthesis are profoundly influenced by steroids.6. The POM is a heterogeneous region from a neurochemical point of view;
however, specific neurochemical markers can differentiate this region from thesurrounding POA.7. The enzyme aromatase represents one of the most interesting specific
markers of POM because it is directly implicated in the control of sexualbehavior.8. Steroids specifically affect the ARO-ir system and the organizational
differences affecting cell size in the POM lateral population appear to concernmainlyARO-ir cells.9. The POM projects to several brain areas that are relevant to the control of
sexual behavior and receives inputs from a number of regions that playimportant roles in the control of a large array of brain functions, includingreproduction (e.g., catecholaminergic cell groups). This connectivity furthersupports the idea that this nucleus controls male copulatory behavior.10. Some of these transmitter or peptidergic inputs are sexually differenti-
ated (NE, DA) and/or are modulated by the endocrine environment (VT). Thissuggests that these inputs to POM play an important role in the control ofbehavior and its sexual dimorphism. Steroids could therefore affect behavior bya direct action on POM and by a modulation of the inputs to this nucleus.
95THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

Some of these conclusions require additional discussion and also raise anumber of important questions.
1. Is the Entire POM Implicated in the Control of Male Sexual Behavior?
As illustrated above, the sexually dimorphic POM is a necessary and suffi-cient site of steroid action for the activation of male copulatory behavior inquail. This differentiates this nucleus from many other SDN of the POA thathave been identified in mammals. In all mammalian species tested so far, it isestablished that lesions of the POA affect male copulatory behavior (164).However, the SDN that have been identified in the POA of these species do notappear to be directly related to the control of male sexual behavior. In rat orferret, for example, lesions restricted to the SDN have little or no effect on thebehavior (22, 93), although contradictory evidence has been presented for rats(105). The reason for this discrepancy between quail and most mammals is notentirely clear at present. It must be observed, however, that the dimorphicnucleus of the quail (POM) is a very large structure that covers a substantialpart of the POA. It is hence anatomically more comparable to the entire medialpreoptic nucleus of the rat (MPN) as described by Bloch and Gorski (80) than tothe SDN, which is about one order of magnitude smaller (Volume of quail POM:0.3 to 0.6 mm3; volume of rat SDN: 0.01 to 0.03 mm3). The putative homologybetween the quail POM and the rat MPN is also suggested by neurochemicalcomparisons. In both species, these nuclei contain high densities of estrogenreceptors as defined either by autoradiography (170, 205, 247–249, 276) or byimmunocytochemistry of the protein (57, 76, 94, 286) or by in situ hybridizationof the corresponding mRNA (159, 242). These receptor-containing cells do notpermit one to exactly delineate the boundaries of the nuclei because they arestill present but at lower densities in the lateral POA adjacent to thesestructures. By contrast, it appears that the POM of quail and the MPN of ratmay be specifically defined by the presence aromatase-containing cells. Thisneurochemical marker has, however, been observed only by immunocytochemi-cal visualization of the protein in quail (48, 55) and by demonstrating itsmessenger RNAby in situ hybridization in rat (155, 275). Immunocytochemicalstudies have failed so far to identified ARO-ir cells in the medial POA of rats(147, 220, 236, 237) even if these elements are present during development(261). It remains, however, that taken together, these data show that thedimorphic nuclei of the POA in quail and rat are not homologous and there istherefore no reason to expect that they should play similar roles in the controlof behavior.The different parts of the gerbil SDA appear to be homologous to large
fractions of the rat medial POA (117). Recently, clear relationships have beenestablished between specific parts of the SDA and specific components ofreproductive behavior (221, 281, 282). It could therefore be expected, byanalogy, that specific parts of the POM are also preferentially implicated in thecontrol of male copulation.
96 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

Our first studies clearly showed that lesions of the POM abolish malecopulatory behavior in quail, while T implants in this nucleus restore thebehavior in castrated birds (69). Direct experimental evidence derived fromlesion studies and correlative data obtained in anatomical and neurochemicalinvestigations of the nucleus suggest more anatomical specificity. Aromataseimmunocytochemistry was recently used to map, at a cellular level of resolu-tion, the parts of the POM that are destroyed by electrolytic lesions or that arestimulated by stereotaxic implants of T in the POA. These measures of thecellular action of T or of the lesions were then correlated with the behavior ofthe experimental subjects to identify parts of the POM that could be morecritical for the activation of male copulatory behavior (70). As previouslyreported, lesions of the POA disrupted the activation of male sexual behavioronly if they destroyed a significant part of the POM. These lesions also reducedthe volume of the nucleus and the number of ARO-ir cells it contained, and themagnitude of these effects was correlated with the behavioral deficit. Furtheranalyses related the magnitude of the observed behavioral deficit to the surfaceof the lesion and the depletion of ARO-ir cells in successive sections of the POMcollected from its rostral to caudal pole (one section every 100 µm). Interest-ingly, these analyses showed that larger disruptions of the behavior areobtained when lesions are located just rostral to the anterior commissure (70).Similar results were obtained when correlating, in individual birds, the induc-tion of ARO-ir cells in successive section of the POM (one section every 100 µmthroughout the rostrocaudal extent of the nucleus) with the activation of sexualbehavior. Behavioral activation was again maximal when more ARO-ir cellshad been induced in the sections of POM located just rostral to the anteriorcommissure (70).These data therefore suggest that a specific part of the POM, located just
rostral to the anterior commissure, could play a more prominent role inbehavioral activation. This notion is supported by a recent immunocytochemi-cal study showing the presence of a sex difference in the number ofARO-ir cellsrestricted to this part of the POM located before the anterior commissure (118).This is also a part of the dimorphic nucleus (caudal portion) for which earlierstudies had suggested, although not fully demonstrated, a sex difference inaromatase activity (63). Our anatomical and neurochemical work furthersuggests that the lateral part of the POM, at the level of the anterior commis-sure or just rostral, could be more relevant to the control of sexual behaviorthan the medial part of the nucleus (24–26, 189).
2. Is Aromatase Sexually Differentiated in the Organizational Sense?
Two fundamentally different actions of steroids have been described in thecontext of behavioral control even if more recent data indicate that thisdistinction may not be as clear as originally thought (23). Organizationaleffects take place during limited periods of the early ontogeny and are, by and
97THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

large, irreversible; activational effects can be observed throughout the life ofanimals but they are reversible (129, 209).The morphological effects of steroids in the POM take place at both the
organizational and the activational levels, as reflected, for example, by thechange in size of lateral neurons in response to embryonic treatment withestrogen (26) or the increase in overall volume of the nucleus after treatment ofgonadectomized adult subjects with T (189, 202). Both aspects can be related tothe control of male copulatory behavior. Activational effects of T on POM, suchas the increase in overall volume (202), in the size of lateral neurons (189), andin the number of ARO-ir cells (48, 118), may represent part of the cellularmechanisms that translate the action of the steroid into changes in neuralfunction that ultimately produce the behavior. Our initial search for a dimor-phic structure in the quail POAwas, however, prompted by the desire to find acorrelate (and possibly a cause) for the sex difference in quail copulatorybehavior (T activates the behavior in males but not in females). This behavioraldifference is clearly organizational in nature: it cannot be abolished by treat-ment with steroids in adulthood and it is entirely effected by the embryonicexposure to estrogen. A neural explanation of this behavioral dimorphismtherefore requires the identification of a brain sex difference that would also beorganized by embryonic estrogens.Because the enzyme aromatase is a limiting factor in the production of the
behavioral effects of T (46, 49), numerous experiments have analyzed itspossible contribution to the sex difference in behavior. Data currently availablestrongly suggest that this neurochemical system is sexually differentiated inthe organizational sense and therefore plays a significant role in the control ofthe behavioral dimorphism.Our original radioenzymatic studies showed that the preoptic aromatase
activity is higher in sexually mature males than in adult egg laying females(227) and that this difference is also maintained in gonadectomized birdssubmitted to a same replacement therapy with T (228). This suggests that thesex difference in enzyme activity is organized by embryonic steroids. Thisenzymatic difference has since been confirmed in a number of experimentsmeasuring AA in gonadectomized T-treated males and females (36, 43, 49) butexperiments designed to directly test its organizational nature have providedonly limited evidence for a control by embryonic/neonatal estrogens (36). Morerefined studies of intact adult males and females confirmed the existence of asex difference in the AA specifically located in the POM (229) but its organiza-tional nature (presence in adult birds placed under the same endocrine condi-tions) could not be formally established: AA is higher in the caudal part of thePOM of castrated males treated with T than in ovariectomized females treatedwith the same dose of steroid but this difference is not fully significant (63).Immunocytochemical data have shown that, in adult intact birds, the sex
difference in ARO-ir cell number is actually limited to a small part of the POMrostral to the anterior commissure (118). This may explain why biochemicalstudies have often failed to identify sex differences in enzyme activity: theymeasure the enzyme in a large brain sample in which the sex difference is
98 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

diluted in a large amount of nondifferentiated tissue, as discussed by Blausteinand Olster (78). One study of ARO-ir cells in gonadectomized T-treated birdshas, unexpectedly, failed to identify a sex difference in the number of ARO-ircells located in the POM. This failure presumably reflects limitations of theimmunocytochemistry and/or technical problems in this specific experiment(118). A number of indirect data indeed suggest that ARO-ir cells are sexuallydifferentiated in adult birds placed under the same endocrine conditions.Studies of the entire population of neurons in the lateral POM suggest thepresence of an organizational sex dimorphism: these neurons are sensitive to Tin males but not in females (189) and their size is irreversibly affected by anembryonic treatment with EB (26). These effects concern primarily the largestneurons in the area. The recent observation thatARO-ir cells constitute most ofthe largest neurons in the lateral POM (25) strongly suggests that the sexdifference described inNissl-stainedmaterial primarily concerns the aromatase-containing cells and studies specifically devoted to this neurochemically de-fined population are currently in progress to characterize these differencesmore precisely. This dimorphism could represent one major sexually differenti-ated feature of the quail brain directly implicated in the control of the sexuallydimorphic copulatory behavior.
3. Neuronal Plasticity in the Quail POM
The sexually dimorphic nucleus, POM, displays a great deal of plasticity inresponse to steroid action. Major changes in the overall volume of this nucleusare indeed observed when castrated birds are treated with T or its activemetabolites (24, 189, 202). The precise mechanisms that mediate these changesin gross volume have not been fully identified. Increases in neuronal size couldcontribute to the effect (189) but do not seem to explain all its aspects. Nochange in neuronal size has, for example, been identified in ovariectomizedfemales treated with T, while the global volume of their POM increased tomale-typical levels (189). Preliminary data based on themorphometrical analy-sis of semithin sections indicate a possible role for changes in the glialcompartment and in the neuropile of female quail (N. Aste, C. Castagna, G. C.Panzica and J. Balthazart, unpublished data). On the other hand, a castration-induced cell death (e.g., apoptosis) associated with a burst of cell divisions afterexposure to steroids appears to be an unlikely mechanism to explain thesteroid-induced changes in POM volume but this possibility has not beenformally tested. Much experimental work remains to be carried out before anintegrated explanation of these volume changes can be presented. It is alreadyclear, however, that the plasticity induced by steroids in the male POM mostlyconcerns neurons because few glial cells can be identified in the nucleus (86)and, in addition, ultrastructural studies have directly confirmed a drasticreorganization of the intracellular arrangement of neurons in response to T(197).Larger neuronal sizes, as observed in POM after exposure to T, are usually
99THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

thought to reflect increases in the cell metabolism and, in particular, in proteinsynthesis. We have independent ultrastructural evidence showing that there isindeed a marked augmentation of the protein synthesis-related organelles inthe POM of castrated birds treated with T (197). One of the proteins whosesynthesis is increased by steroids should be aromatase as clearly indicated bythe fact that, in the quail POM/POA, T treatment increases the aromataseactivity (63, 228), the number of ARO-ir cells (48, 54, 118, 119), their opticaldensity, which should reflect the enzyme concentration per cell (119), andfinally the concentration of the aromatase mRNA in the quail POA (135, 137).The quail POM is therefore a convenient model for the study of neuronal
plasticity and its control by steroids. Some years ago, studies of steroid action inthe avian brain unexpectedly revealed that an active neurogenesis is main-tained in parts of the adult brain of songbirds (179). Additional work confirmedthat the production of these new neurons is stimulated by T but that it islimited to specific parts of the telencephalon and does not occurs in the preopticarea/hypothalamus (20, 21, 178, 212). Similarly, neurogenesis has been identi-fied in the brain of adult budgerigars (Melopsittacus undulatus) and ring doves(Streptopelia risoria) but in these species also, new neurons were found in theforebrain only (177). These studies have not been replicated in galliforms andthere is therefore no evidence that new neurons could be added in the brain ofthe quail. If an active neurogenesis was identified in quail, it is, however, likelythat it would also be limited to the telencephalon. It is therefore improbablethat new neurons are added to the POM of adult quail under the influence of T.The available data show, however, a different type of plasticity that may also
be very important for the development and maintenance of brain function. Itincludes changes in neuronal function, in protein synthesis, and in specificinputs. These steroid-induced changes observed in quail are therefore similarto the type of plasticity that can be observed in the mammalian and inparticular in the human brain, where no or little neurogenesis takes place.These phenomena can easily be studied in the POM due to their large magni-tude, localization in a specific brain site, and rapid development after exposureto steroids. The fact that steroid action in the POM can be directly related to aclear biological output, the activation of male sexual behavior, also adds to theinterest of this nucleus as a tool for understanding steroid action in the brain.The quail POM therefore constitutes a exceptional model for the analysis ofsteroid-induced brain plasticity in a functionally relevant context.
4. Aromatase and Estrogen Receptors Are Not Colocalized in the POM
The demonstration thatARO-ir cells in POMare generally devoid of immuno-cytochemically detectable ER (108) also raises important physiological ques-tions regarding the mode of action of locally produced estrogens and the controlof aromatase synthesis in this nucleus. This rare colocalization was observed tobe the rule in the POM and in the septal region (17 and 4% of double labeledcells, respectively), while a much larger proportion (about 70%) of the ARO-ir
100 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

cells contained visible ER in the caudal hypothalamus. This contrast suggeststhat the lack of ER in the ARO-ir cells of the POM does not result from atechnical artifact: both markers can be effectively visualized in the samesections and even in the same cells. It must also be stressed that ER-positivecells were observed in all sections that contained ARO-ir cells with no visibleER. This rules out the possibility that the immunocytochemical detection of ERspecifically failed in the sections through the POM. The absence of detectableER in the ARO-ir cells might possibly result from an artifact that would be ofphysiological rather than strictly technical nature. The monoclonal antibodyH222SPg used in these studies recognizes an epitope located in the hormonebinding domain of the ER molecule (154). The free and occupied receptorsmight therefore be differentially recognized by H222SPg. Claims have beenmade that this antibody recognizes equally well both forms of receptors (210) orrecognizes preferentially if not exclusively the free receptor (79, 215). However,biochemical studies have shown that H222SPg binds to both occupied and freeER even if the molar ratios for these interactions are different (125). We believetherefore that the high level of receptor occupancy which might be present inARO-ir cells is not sufficient to explain the absence of visible receptors in thesecells. There is also no reason, as discussed previously (52), to suspect that thelevel of ER occupancy should be higher in the POM (where little colocalizationoccurs) than in the tuberal hypothalamus (where colocalization is the rule).Behavioral and physiological experiments have demonstrated that T aroma-
tization into E2 is required for a full production of the behavioral effects of T (11,45, 46, 49) but also for the stimulation of aromatase synthesis (45, 67, 119).These effects are presumably mediated by the binding of locally producedestrogens to ER, as shown at least in the case of behavior by the fact thattreatment with antiestrogens blocks the activation by T of sexual behavior incastrated males (12, 18, 68). A parsimonious explanation of these multipleeffects would assume that these events take place in a cell which contains boththe aromatase in its cytoplasm and the ER in its nucleus (Fig. 19A).The absence of demonstrable ER in the largest part (about 83%) of theARO-ir
cells in POM clearly challenges this view and asks for a more complex circuitryimplicating at least two neurons (and probably more) to take into account themultiple effects of estrogens produced in the brain (Fig. 19B). The control ofaromatase synthesis and of sexual behavior must here be considered sepa-rately.
Estrogen-Induced Increase in Aromatase Concentration
Although T increases aromatase activity in the POA of all species of highervertebrates examined so far, this effect appears to be largely mediated by theinteraction of the steroid with androgen receptors in rats (218, 219) but by anaction of locally produced estrogens in birds (144, 225). As fully describedabove, this induction of enzyme activity reflects an increase in the concentra-tion of the enzyme and of its mRNA, clearly suggesting an increased transcrip-
101THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
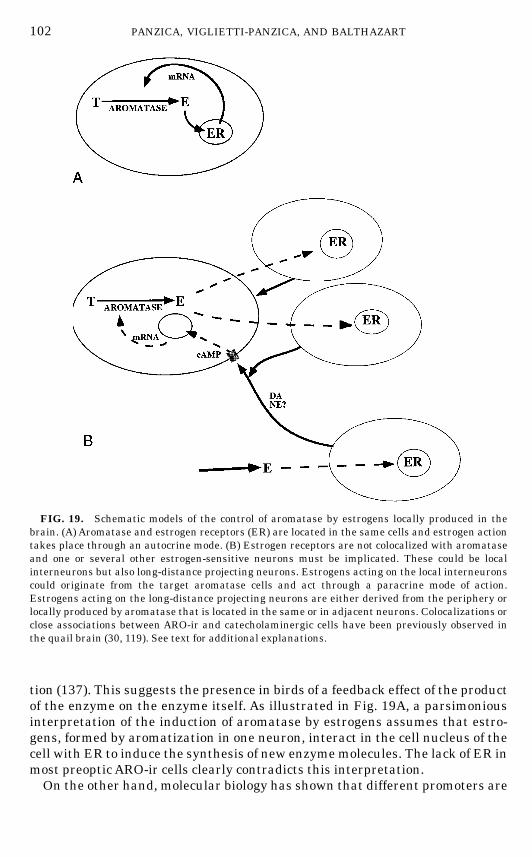
tion (137). This suggests the presence in birds of a feedback effect of the productof the enzyme on the enzyme itself. As illustrated in Fig. 19A, a parsimoniousinterpretation of the induction of aromatase by estrogens assumes that estro-gens, formed by aromatization in one neuron, interact in the cell nucleus of thecell with ER to induce the synthesis of new enzymemolecules. The lack of ER inmost preoptic ARO-ir cells clearly contradicts this interpretation.On the other hand, molecular biology has shown that different promoters are
FIG. 19. Schematic models of the control of aromatase by estrogens locally produced in thebrain. (A) Aromatase and estrogen receptors (ER) are located in the same cells and estrogen actiontakes place through an autocrine mode. (B) Estrogen receptors are not colocalized with aromataseand one or several other estrogen-sensitive neurons must be implicated. These could be localinterneurons but also long-distance projecting neurons. Estrogens acting on the local interneuronscould originate from the target aromatase cells and act through a paracrine mode of action.Estrogens acting on the long-distance projecting neurons are either derived from the periphery orlocally produced by aromatase that is located in the same or in adjacent neurons. Colocalizations orclose associations between ARO-ir and catecholaminergic cells have been previously observed inthe quail brain (30, 119). See text for additional explanations.
102 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

used to regulate the synthesis of aromatase in different human tissues but,unfortunately, the promoter used in the brain has not yet been identified (244).It appears, however that, although several studies have analyzed the aro-matase gene in a number of species, no one has, to this date, identifiedestrogen-responsive elements (nor androgen-responsive elements) that couldmediate a direct effect of steroids on aromatase synthesis (136, 243, 244). Theabsence of direct interaction of steroids with aromatase-containing cells is alsosuggested by the observation that several studies have failed to increasearomatase synthesis (measured by the enzyme activity or the concentration ofits mRNA) in dissociated cell cultures of embryonic POA–hypothalamus (1,157) (J. Balthazart and C. Balthazart-Raze, unpublished observations).The possibility therefore exists that estrogens do not regulate aromatase
activity through binding to ER and association with estrogen-responsive ele-ments in the same cell. A control exerted by a trans-synaptic mechanism needsto be contemplated. The second messenger, cyclic AMP (cAMP), is known tomodulate gene expression through the phosphorylation of transcriptional acti-vator proteins (233) [see also (176) for a review of data pertinent to the controlof behavior]. In particular, it is established that cAMP regulates aromataseactivity in a number of tissues (90, 165, 264, 265), including the brain (85). ThecAMP-induced increase in aromatase activity develops only gradually over a 2-to 4-day period, which suggests that it is due to a progressive increase inenzyme concentration (85, 165). It is therefore tempting to suggest that estro-gens increase aromatase activity and aromatase concentration not through adirect genomic autocrine action in the cells where they were produced butrather by acting on another neuron (local ormore distant) producing a transmit-ter that uses cAMP as second messenger.On the other hand, the observations that aromatase cells are found in close
association with DBH-ir and TH-ir terminals in the preoptic region (30) andthat both NE and DA use cAMP as second messenger (114, 115, 126) suggestthat catecholaminergic neurons could represent long projecting neurons impli-cated in this control. It is already known that noradrenergic agonists regulatearomatase activity in the pineal gland (90) or in the testes (264) and similarcontrols might take place in the hypothalamus (87, 213, 214). Given that thenoradrenergic activity in the brain appears to be under the control of estrogens(163), an estrogen-sensitive noradrenergic transmission might represent theneuroanatomical substrate underlying the regulation of aromatase activity byestrogen. This is consistent with the fact that, in quail, brain areas identified byhigh densities of ARO-ir neurons (medial preoptic nucleus and tuberal hypo-thalamus) also contain high concentrations of norepinephrine (37, 58) and highdensities of a2-adrenergic receptors (32, 37).Anoradrenergic denervation of thequail brain by the specific neurotoxin DSP4 enhances aromatase activity in thepreoptic area (37), which certainly supports the present interpretation (see Fig.16).The same argument can be raised for DA. This neurotransmitter present in
TH-ir positive terminals detected in close proximity to ARO-ir cells (5, 30) ismeasured in significant concentration in the POM (37), and one of its receptor
103THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

subtypes (D1), which has been detected in the POA(31), is also coupled with theactivation of adenylyl cylase (126). In this case also, preliminary pharmacologi-cal evidence suggests a role for DA in the control of the preoptic aromataseactivity (see Fig. 17). Further studies should analyze the precise anatomicalrelationships between noradrenergic and dopaminergic cell groups, their projec-tions, estrogen receptors, and aromatase. It is interesting to note that thismode of control for the aromatase expression would be analogous to what hasbeen described in more detail for the regulation of LHRH-producing neurons.Although steroids exert a strong influence on the synthesis and release of thispeptide, neurons producing LHRH usually contain no estrogen receptors (138,140, 238) and they are controlled by an estrogen-sensitive neuronal networkthat involves several neurotransmitters and neuropeptides (71, 106, 279).
Activation of Male Copulatory Behavior
The lack of ER in preoptic ARO-ir cells also raises the question of how locallyformed estrogens exert their behavioral effects. T acts in the POM to stimulatemale sexual behavior (69). This effect is blocked by the concurrent administra-tion of aromatase inhibitors (11, 45, 49, 68) and of antiestrogens (12, 18, 68),which suggests that locally formed estrogens interact with ER to produce theirbehavioral effects. It was therefore expected that ER would be colocalized witharomatase, at least in the POM, where the biological role of the estradiolderived from T aromatization is so well established. The fact that this colocal-ization was observed in only 17% of the ARO-ir cells (52) leads to the formula-tion of additional hypotheses.It can first be suggested that only a few cells of the POM are implicated in the
activation of behavior and these are the cells that contain both ER andaromatase at the same time. In this case, behavioral activation is mediated bygenomic activation and results from an autocrine action of the steroid (estrogenis produced and exerts its effects in the same cell). This, however, provides noindication for a role of the large majority of ARO-ir neurons. Because ER-positive cells are present in large number in the POM (even if they do notcorrespond to ARO-ir cells), it is also possible that estrogens produced by localaromatization of T exert their biological effects by acting on other ER-containing cells after a passive diffusion through the intercellular space (para-crine rather than autocrine mode of action). Alternatively other nonconven-tional modes of action can be contemplated.It is indeed important to note that, in quail and in rats, biochemical studies
have demonstrated the presence of high levels of aromatase activity in synapto-somes prepared by differential centrifugation (223, 245) even if recent dataseem to contradict this fact in rat (217). In addition, we have shown bypreembedding immunocytochemistry and electron microscopy that numeroussynaptic boutons containing ARO-ir material are present in the quail POA (53,173). Therefore, it appears likely that significant amounts of estrogens areproduced at the presynaptic level. These may be responsible for effects that
104 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

would not be mediated by the nuclear ER but would rather involve a nonge-nomic action of the steroid (162, 224). These might include previously describedsynaptogenic/synaptolytic effects of gonadal steroids (171, 172, 199). Estrogensalso appear to be responsible for the fluctuations of the postsynaptic membraneorganization during the ovarian cycle (172) and they interfere with the bindingof catecholamines to the synaptic membrane (146). Their synthesis at thesynapse level is therefore biologically very relevant. On the other hand,estrogens and their metabolites, the catechol-estrogens, are potent catecholO-methyl transferase inhibitors and in generalmodulate the synthesis, metabo-lism, and binding of catecholamines (160, 163). Their presence in the synapticzone might directly regulate the activity of these neurotransmitters. In connec-tion with this type of action, it is highly relevant to mention here that areas ofthe quail brain that display a high aromatase activity also contain a high levelof 2-hydroxylase activity, i.e., the enzyme that converts estrogens into catechol-estrogens (66). This may not be surprising based on a recent report suggestingthat these two enzymatic activities are in fact due to the same molecule thatshifts its activity as a function of the substrate concentration or pH (182). It iswell established that catecholamines are a part of the neurochemical signalsthat control male sexual behavior (see above). Therefore, these data bringadditional support to the idea that estrogens produced at the presynaptic levelcould affect reproductive behavior by a direct action on the catecholaminesmetabolism at the level of nerve terminals, independent of any genomic action.This hypothesis should now be experimentally tested.In conclusion, the identification of two types ofARO-ir cells (containing ER or
not) in the quail POM raises questions concerning the biological role and themode of action of estrogens produced in the brain.Whether both types ofARO-ircells are implicated in behavior activation remains unclear. The fact thatantiestrogens block the activation of behavior by T suggests that the interac-tion with nuclear ER is critical but does not rule out the possibility that directeffects of estrogen at the synapse level might be important. The specificfunctional significance of these two types of ARO-ir cells should now beinvestigated: do the double labeled (ARO-ER) cells receive specific inputs(transmitters, neuropeptides) that could support a differential regulation, dothey project to areas distinct from the projection areas of non-double-labeledcells? It is interesting to recall that the detailed analysis of the ARO-ir cellreaction to castration associated with or without T replacement therapy alsoidentifies two subpopulations of aromatase cells in the POM: some are nolonger evidenced by the immunocytochemical procedure (70–90% depending onthe part of the nucleus), while others remain clearly labeled (25). The possiblerelationship between these two modes of subdividing ARO-ir cells (containingER or not, steroid-sensitive or not) should also be explored. In particular, oneshould determine whether the small fraction (10–30%) of steroid-insentiveARO-ir cells that are still observed in castrated birds corresponds to theARO-ircells that simultaneously contain ER. They would then represent a key targetpopulation that could play a major role in the activation of the steroid-sensitive
105THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

neurochemical networks when sexually inactive subjects (castrated, photore-gressed) are exposed to steroids.
5. POM Connections
Quail are photoperiodic and they respond to increased daylengths with adramatic growth of the testes that in turn influences the sexual activity. Thiseffect is largely mediated through the stimulation of LHRH release followed bythe secretion of LH and T (59, 120), but it could also be more direct as reflectedby the observation that crowing and copulatory behavior occur with largerfrequencies during some parts of the day–light cycle (110, 130, 184). A malequail exposed to long days can attain the limit of his reproductive developmentin response to photoperiod alone, as no additional effect of females on testiculardevelopment can be detected (111). However, in males exposed to marginallystimulating photoperiods (e.g., 12L:12D), gonadal maturation occurs morerapidly in young males raised in the presence of females (111).The social life of quail, like many other avian species, is organized mainly by
visual and acoustic cues, whereas olfactory and tactile stimuli are usuallyconsidered to play only a minor role (141, 156). In birds, the importance of theauditory and visual sensory pathways for the initiation of reproductive behav-ior has been studied in the most detail in the ring dove. In this species, it wasshown that both auditory information and visual information are necessary toelicit the full behavioral repertoire (91). A male dove when presented with anintact female will immediately display male-typical courtship behaviors andthis response is followed by a significant rise in plasma T levels within 30 min(116, 180). The response is dependent in part on visual and auditory aspects ofthe behavior of the female. Similarly, the female’s endocrine and behavioralresponse to the male is dependent on both visual and acoustic cues emanatingfrom the male (91). Interestingly, the females behavioral response to the malecontributes to her own endocrine development by means of a self-stimulationmechanism (92).In quail, Guyomarc’h has also demonstrated that ovarian development in
females can be stimulated by exposure to male vocalizations (130, 131). In malequail, as is true of many galliform species, visual cues appear to be of primaryimportance in the control of social interactions and seem to be sufficient to elicitcopulatory responses in a male with high levels of T. The presentation of afemale model has been shown to elicit the male copulatory response in turkeys,chicken, and quail (113). In addition, Domjan and Nash have demonstratedthat static visual cues, as obtained from a taxidermically prepared model, aresufficient to induce social proximity behavior and suppress crowing in malequail in the absence of behavioral, auditory, and olfactory cues. The behaviorinduced by the female model was primarily elicited by visual cues from thehead and neck region, which are sexually dimorphic in quail (113). Additionalstimuli, especially vocalizations and behavioral cues such as copulation solicita-
106 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant
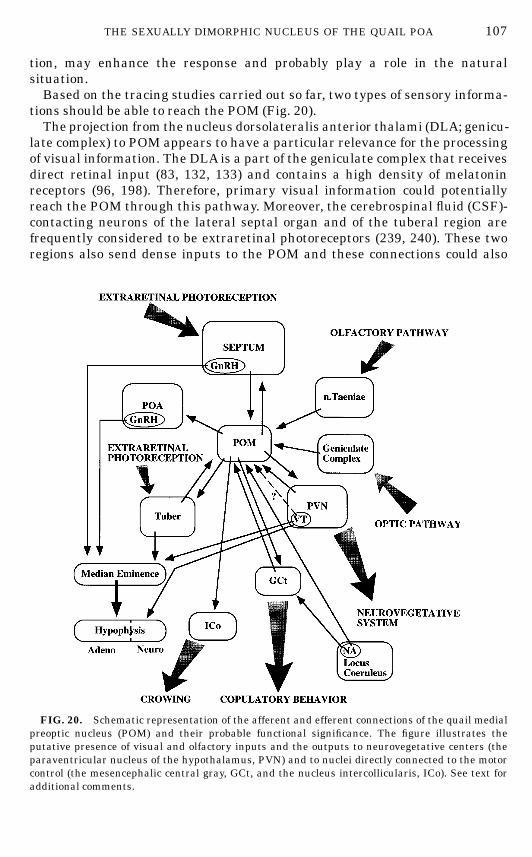
tion, may enhance the response and probably play a role in the naturalsituation.Based on the tracing studies carried out so far, two types of sensory informa-
tions should be able to reach the POM (Fig. 20).The projection from the nucleus dorsolateralis anterior thalami (DLA; genicu-
late complex) to POM appears to have a particular relevance for the processingof visual information. The DLA is a part of the geniculate complex that receivesdirect retinal input (83, 132, 133) and contains a high density of melatoninreceptors (96, 198). Therefore, primary visual information could potentiallyreach the POM through this pathway. Moreover, the cerebrospinal fluid (CSF)-contacting neurons of the lateral septal organ and of the tuberal region arefrequently considered to be extraretinal photoreceptors (239, 240). These tworegions also send dense inputs to the POM and these connections could also
FIG. 20. Schematic representation of the afferent and efferent connections of the quail medialpreoptic nucleus (POM) and their probable functional significance. The figure illustrates theputative presence of visual and olfactory inputs and the outputs to neurovegetative centers (theparaventricular nucleus of the hypothalamus, PVN) and to nuclei directly connected to the motorcontrol (the mesencephalic central gray, GCt, and the nucleus intercollicularis, ICo). See text foradditional comments.
107THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

relay information about the environmental level of light to the dimorphicnucleus. Information about environmental light can therefore reach the POMthrough different routes (retinal input through DLA, deep photoreceptorsthrough septal region and tuberal hypothalamus).We do not know at present whether specific visual information about the
mating partner reaches the dimorphic nucleus. The thalamofugal (132, 133,143) or retinothalamohyperstriatal (83) pathway is connected to POM vianucleus DLA but it is unlikely that the level of integration of the visualinformation in the geniculate complex (DLA) would permit the identification ofthe female as such. Projections to the POM originating from telencephalicregions where the final processing of visual information takes place would berequested for that purpose. We recently identified by retrograde tracing withCTB or with fluorescent microspheres a diffuse projection from the hyperstria-tum accessorium (part of the visual Wulst) to the POM (P. Absil and J.Balthazart, unpublished data). This connection could convey complex visualinformation but more functional studies on the visual Wulst would be needed(132, 133) and a better definition of its connection to the POM should beestablished by specific tract tracing experiments before the significance of thisanatomical relation can be established.No direct auditory input to POM has been identified but it must be kept in
mind that visual cues provided by a taxidermic female model are sufficient tostimulate the appetitive and consummatory aspects of sexual behavior of malequail (113). It is therefore plausible that auditory stimuli are not implicated inthe direct control of male sexual behavior in this species.In mammals, olfactory information derived mainly from the accessory olfac-
tory system reaches the POA and appears to play a key role in the activation ofmale copulatory behavior. The path followed by this chemosensory informationhas been relatively well documented and includes the corticomedial amygdalaand the bed nucleus of the stria terminalis (164, 234). In vivo tracing with CTBhas revealed the presence in quail of an important projection from the archis-triatum and in particular the nucleus taeniae to the POM (5). Nucleus taeniaeis the avian holomog of the mammalian medial amygdala (174, 287), whichsuggests that olfactory inputs could reach the medial preoptic nucleus of thequail. The importance of olfactory information in the control of behavior inbirds is usually considered to be minimal but a limited number of studiessuggest that this widespread idea should perhaps be reconsidered and that thechemical information originating in the female could modulate aspects of thereproductive behavior (61, 280). The projection from nucleus taeniae to POMwould then acquire a particular importance.These studies establish that the POM potentially receives both visual infor-
mation and olfactory information. It must be stressed, however, that theamount of sensory information that reaches the nucleus through these path-ways has not been experimentally determined and we do not know what typesof cells in POM (e.g., steroid-sensitive or not) are connected to these inputs. Inaddition, it is clear that several functional pathways using a variety of neuro-transmitters or neuropeptides reach the dimorphic nucleus. The presence of
108 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

DHB-ir and TH-ir fibers has, for example, been detected in POM (29, 30), andretrograde tracing studies have confirmed that noradrenergic (locus coeruleus)and dopaminergic (area ventralis of Tsai, homolog of the mammalian ventraltegmental area; nucleus tegmentopedunculopontinus, holomog of the mamma-lian substantia nigra) cell groups indeed project to this region (5). Fibers andpunctate structures immunoreactive for VT, NT, NPY, SP, and CRF have alsobeen identified in POM but their origin is unknown at present (192). Takentogether, these anatomical data clearly demonstrate that the POM is a poten-tial site for the integration of various sources of information. Experimentalstudies should now be performed to determine the importance of these differentinputs in the control of behavior.Complex outputs from the POMhave also been identified. They namely reach
the septum, the paraventricular (PVN) and the ventromedial (VMN) nuclei ofthe hypothalamus, the tuberal region, the intercollicular nucleus (ICo), themesencephalic central gray (GCt), and the area ventralis of Tsai. Most of thesestructures have been implicated directly or indirectly in the control of copula-tory behavior in mammals (164). Many of these regions are also steroid-sensitive in quail (56, 57, 276) and in mammals (169, 170, 247, 248). They arepresumably part of the nervous circuitry that controls copulation. The connec-tion with the GCt is potentially of utmost importance here. This area ismassively connected in a bidirectional way with a large number of more caudalmesencephalic and pontine structures (44), and therefore it appears to be animportant center connecting anterior levels of the brain to brainstem nucleiinvolved in the control of the complex motor output represented in malecopulatory behavior. In addition, the projection from the POM to the ICo couldbe implicated in the timing of reproductive behavior in quail. As mentionedbefore, vocalizations of male quail and in particular crows stimulate theovarian development of females (130, 131). Ovarian hormones in turn activatethe receptive behavior of females (10, 109), which facilitates the mounting andcloacal contacts of the male. Several lines of evidence have implicated the ICoin the mediation of crowing and other vocalization in quail (39). It is wellestablished that the quail ICo contains both androgen and estrogen receptors(56, 57, 276) so that steroids could potentially activate in parallel the malecopulatory behavior (by acting in POM) and the vocalizations (by acting in ICo).A finer level of coordination appears to be, in addition, possible thorough theanatomical projection of POM to ICO.
6. Direct and Indirect Effects of Steroids on POM
Potentially, the POM could hence be controlled by T in two separatemanners.On the one hand, the nucleus contains steroid receptors and it could be directlyaffected by steroid hormones. This action would be reflected in the modifica-tions of morphometric characteristics (e.g., cell size, ultrastructural organiza-tion) that are observed in specific subpopulations of cells within this nucleus(dorsolateral neurons) following treatment of castrated birds with Silastic
109THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

implants filled with androgens or estrogens. These morphological changesprobably reflect modifications of the cellular metabolism (e.g., stimulation ordepression of the protein synthesis, aromatase being one of the moleculespotentially regulated in this way). On the other hand, POM neurons could beindirectly regulated by T acting through its afferents (neurotransmitters orneuropeptides) to the nucleus (e.g., the VT, DA, or NE systems). Some of theseafferents (e.g., DA) are sexually differentiated in the organizational sense.Others (e.g., VT) aremodulated by T in adulthood and it is very likely that morerefined studies would also demonstrate that the activity of catecholaminergicinputs to POM are modulated by steroids.These various inputs are likely to affect the physiology of POM neurons and
this has already been proposed in the case of aromatase-containing neurons.Asexplained before, the synthesis of this enzyme and its sexual dimorphismmightbe controlled by the long distance catecholaminergic projections to POM. Thisclaim may appear to contradict the observation that stereotaxic implants of Tin or around POMare able to increase the number ofARO-ir cells in the nucleus(70), despite the fact that most ARO-ir cells are devoid of immunoreactive ER(52). It must be kept in mind, however, that electron microscopic studies havedemonstrated the existence of abundant synaptic contacts in the POM andthese suggest a complex integration of signals (200). In particular, the presenceof numerous axodendritic synapses raises the possibility that ARO-ir cellscould be tonically stimulated by DA or inhibited by NE (long distance control;influenced or not by steroids) but that the catecholaminergic synapses on AROcell in POMcould be, in addition,modulated by local steroid-sensitive aromatase-negative neurons located in or around POM (see Fig. 19B). T and/or its activemetabolite E2 would therefore have a dual control onARO-ir cells.
CONCLUSIONS
In conclusion, the data reviewed in this paper demonstrate that the sexuallydimorphic nucleus of the quail POA represents a key center in the action ofsteroids on male sexual behavior. Various aspects of the POM morphologychange in reaction to T and/or E2, and the structure of this nucleus provides amodel for the action of steroids in the brain. Several forms of neuronalplasticity are observed in the POM and this nucleus constitutes an ideal modelfor the study of sexual behavior in a context that permits a functional interpre-tation of the changes observed at the cellular level.Recent data suggest that the sex differences and the steroid-induced changes
in neuronal size previously described in Nissl-stained material essentiallyconcern aromatase-containing cells. Since aromatization of T plays a limitingrole in the activation of male copulatory behavior, these changes may representthe morphological signature of the mechanisms causally involved in the controlof this behavior and of the mechanism responsible for its sexual dimorphism.Moreover this neuronal system is directly innervated by T-dependent VT fibersand it also receives substantial catecholaminergic inputs. The POM could
110 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

hence be controlled by steroids in direct and indirect ways. The network ofafferent and efferent connections of the nucleus is clearly consistent with thekey role played by the POM in the control of male reproductive behavior.Experimental studies should now analyze the specific contribution of theseinputs and outputs to the dimorphic nucleus of the quail POA.
ACKNOWLEDGMENTS
We thank all of our collaborators who have in the course of the past 10 years helped in thecollection of much of the data presented in this paper. These people include NicolettaAste, PatriziaAimar, Claudia Castagna, and Gian Carlo Anselmetti from the Department of Human Anatomyand Physiology, University of Torino, andMichael Schumacher,Agnes Foidart, Chantal Surlemont,and PhilippeAbsil from the Laboratory of Biochemistry, University of Liege, as well asmore distantcollaborators, in particular, Professor Nobuhiro Harada, Fujita-Gakuen Health University, Toyo-ake, Aichi, Japan, and Professor G. F. Ball, The Johns Hopkins University, Baltimore, Maryland.The research described in this paper has been supported by grants from the MURST (60 and 40%)and CNR (90.02456, 91.00295, 92.01065, 94.02342) to G.C.P. and C.V.P. and by grants from the NIH(MH 50388), the Belgian FRFC (Nbr. 2.9003.91), the French Community of Belgium (Actionconcertee 93/98-171), and the University of Liege (Fonds Speciaux pour la Recherche) to J.B. Thecollaboration between G.C.P. and J.B. is supported by the ESF (RG 14/92) and the EC HumanCapital program (Grant CT94-0472). We also thank Dr. Gregory F. Ball for his critical reading andmany suggestions on an earlier version of this review. The collaboration of J.B. and G. Ball issupported by NATO collaborative Research Grant CRG910526.
REFERENCES
1. Abe-Dohmae S, Tanaka R, Harada N. Cell type- and region-specific expression of aromatasemRNA in cultured brain cells.Mol Brain Res 1994; 24: 153–158.
2. Absil P, Balthazart J. Sex difference in the neurotensin-immunoreactive cell populations ofthe preoptic area in quail (Coturnix japonica). Cell Tissue Res 1994; 276: 99–116.
3. Absil P, Balthazart J. Testosterone effects on neurotensin-immunoreactive cells in the quailpreoptic area.NeuroReport 1994; 5: 1129–1132.
4. Absil P, Das S, Balthazart J. Effects of apomorphine on sexual behavior in male quail.Pharmacol Biochem Behav 1994; 47: 77–88.
5. Absil P, FoidartA, Surlemont C, Balthazart J. Dopaminergic inputs to aromatase-containingareas in the quail brain identified by double label immunocytochemistry and retrogradetracing. Soc Neurosci Abstr 1994; 20: 827.
6. Adkins EK. Hormonal basis of sexual differentiation in the Japanese quail. J Comp PhysiolPsychol 1975; 89: 61–71.
7. Adkins EK. Effects of diverse androgens on the sexual behavior and morphology of castratedmale quail.Horm Behav 1977; 8: 201–207.
8. Adkins EK. Sex steroids and the differentiation of avian reproductive behavior. Am Zool1978; 18: 501–509.
9. Adkins EK. Effect of embryonic treatment with estradiol or testosterone on sexual differen-tiation of the quail brain.Neuroendocrinology 1979; 29: 178–185.
10. Adkins EK, Adler NT. Hormonal control of behavior in the Japanese quail. J Comp PhysiolPsychol 1972; 81: 27–36.
111THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

11. Adkins EK, Boop JJ, Koutnik DL, Morris JB, Pniewski EE. Further evidence that androgenaromatization is essential for the activation of copulation in male quail. Physiol Behav 1980;24: 441–446.
12. Adkins EK, Nock BL. The effects of the antiestrogen CI-628 on sexual behavior activated byandrogen and estrogen in quail.Horm Behav 1976; 7: 417–429.
13. Adkins EK, Pniewski EE. Control of reproductive behavior by sex steroids in male quail. JComp Physiol Psychol 1978; 92: 1169–1178.
14. Adkins-Regan E. Sex steroids and the differentiation and activation of avian reproductivebehaviour. In: Balthazart J, Gilles R, Eds. Hormones and Behaviour in Higher Vertebrates.Berlin: Springer-Verlag, 1983: 219–228.
15. Adkins-Regan E. Nonmammalian psychosexual differentiation. In:Adler NT, Goy R, Pfaff D,Eds.Handbook of Neurobiology, Vol. 7.NewYork: Plenum, 1985: 43–76.
16. Adkins-Regan E, Watson JT. Sexual dimorphism in the avian brain is not limited to the songcontrol system of birds:Amorphometric analysis of the brain of the quail (Coturnix japonica).Brain Res 1990; 514: 320–326.
17. Akhtar M, Njar VCO, Wright JN. Mechanistic studies on aromatase and related C—C bondcleaving P-450 enzymes. J Steroid BiochemMol Biol 1993; 44: 375–387.
18. Alexandre C, Balthazart J. Effects of metabolism inhibitors, antiestrogens and antiandro-gens on the androgen and estrogen induced sexual behavior in Japanese quail. PhysiolBehav 1986; 38: 581–591.
19. Allen LS, Hines M, Shryne JE, Gorski RA. Two sexually dimorphic cell groups in the humanbrain. J Neurosci 1989; 9: 497–506.
20. Alvarez-Buylla A. Mechanism of neurogenesis in adult avian brain. Experientia 1990; 46:948–955.
21. Alvarez-BuyllaA, TheelenM, Nottebohm F. Proliferation ‘‘hot spots’’ in adult avian ventricu-lar zone reveal radial cell division.Neuron 1990; 5: 101–109.
22. Arendash GW, Gorski RA. Effects of discrete lesions of the sexually dimorphic nucleus of thepreoptic area or other medial preoptic regions on the sexual behavior of male rats. Brain ResBull 1983; 10: 147–154.
23. Arnold AP, Breedlove SM. Organizational and activational effects of sex steroids on brainand behavior: A reanalysis.Horm Behav 1985; 19: 469–498.
24. Aste N, Panzica GC, Aimar P, Viglietti-Panzica C, Foidart A, Balthazart J. Implication oftestosteronemetabolism in the control of the sexually dimorphic nucleus of the quail preopticarea. Brain Res Bull 1993; 31: 601–611.
25. Aste N, Panzica GC, Aimar P, Viglietti-Panzica C, Harada N, Foidart A, Balthazart J.Morphometric studies demonstrate that aromatase-immunoreactive cells are the maintarget of androgens and estrogens in the quail medial preoptic nucleus. Exp Brain Res 1994;101: 241–252.
26. Aste N, Panzica GC, Viglietti-Panzica C, Balthazart J. Effects of in ovo estradiol benzoatetreatments on sexual behavior and size of neurons in the sexually dimorphic medial preopticnucleus of Japanese quail. Brain Res Bull 1991; 27: 713–720.
27. Aste N, Viglietti-Panzica C, Fasolo A, Andreone C, Vaudry H, Pelletier G, Panzica GC.Localization of neuropeptide Y-immunoreactive cells and fibres in the brain of the Japanesequail. Cell Tissue Res 1991; 265: 219–230.
28. Aste N, Viglietti-Panzica C, Fasolo A, Panzica GC. Mapping of neurochemical markers inquail central nervous system: VIP- and SP-like immunoreactivity. J Chem Neuroanat 1995;8: 87–102.
29. Bailhache T, Balthazart J. The catecholaminergic system of the quail brain: Immunocyto-chemical studies of dopamine b-hydroxylase and tyrosine hydroxylase. J Comp Neurol 1993;329: 230–256.
112 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

30. Bailhache T, Foidart A, Surlemont C, Harada N, Balthazart J. Catecholaminergic innerva-tion of aromatase and estrogen receptor-immunoreactive cells in the quail brain. SocNeurosci Abstr 1991; 17: 269–269.
31. Ball GF, Casto J, Balthazart J. Autoradiographic localization of D1-like dopamine receptorsin the forebrain of male and female Japanese quail and their relationship with immunoreac-tive tyrosine hydroxylase. J Chem Neuroanat 1995; 9: 121–133.
32. Ball GF, Nock B, McEwen BS, Balthazart J. Distribution of a2-adrenergic receptors in thebrain of the Japanese quail as determined by quantitative autoradiography: Implications forthe control of sexually dimorphic reproductive processes. Brain Res 1989; 491: 68–79.
33. Ball GF, Nock B, Wingfield JC, McEwen BS, Balthazart J. Muscarinic cholinergic receptorsin the songbird and quail brain:Aquantitative autoradiographic study. J CompNeurol 1990;298: 431–442.
34. Balthazart J. Hormonal correlates of behavior. In: Farner DS, King JR, Parkes KC. Eds.Avian Biology.NewYork: Academic Press, 1983: 221–365.
35. Balthazart J. Steroid metabolism and the activation of social behavior. In: Balthazart J, Ed.Advances in Comparative and Environmental Physiology, Vol. 3. Berlin: Springer-Verlag,1989: 105–159.
36. Balthazart J. Correlation between the sexually dimorphic aromatase of the preoptic areaand sexual behavior in quail: Effects of neonatal manipulations of the hormonal milieu. ArchInt Physiol Biochem 1989; 97: 465–481.
37. Balthazart J, Ball GF. Effects of the noradrenergic neurotoxin DSP-4 on luteinizing hormonelevels, catecholamine concentrations, a2-adrenergic receptor binding, and aromatase activ-ity in the brain of the Japanese quail. Brain Res 1989; 492: 163–175.
38. Balthazart J, Ball GF. Is dopamine interacting with aromatase to control of sexual behaviorin male quail? Poultry Sci Rev 1992; 4: 217–233.
39. Balthazart J, Ball GF. Neurochemical differences in two steroid-sensitive areas mediatingreproductive behaviors. Adv Comp Environ Physiol 1993; 15: 133–161.
40. Balthazart J, Ball GF, McEwen BS. An autoradiographic study of a1-adrenergic receptors inthe brain of the Japanese quail (Coturnix coturnix japonica). Cell Tissue Res 1989; 258:563–568.
41. Balthazart J, De Clerck A, Foidart A. Behavioral demasculinization of female quail isinduced by estrogens: Studies with the new aromatase inhibitor, R76713.Horm Behav 1992;26: 179–203.
42. Balthazart J, Delville Y, Sulon Y, Hendrick JC. Plasma levels of luteinizing hormone and offive steroids in photostimulated, castrated and testosterone-treated male and female Japa-nese quail (Coturnix coturnix japonica).Gen Endocrinol (Life Sci Adv) 1987; 5: 31–36.
43. Balthazart J, Devos F, Dohet A, Foidart A, Hugla JL, Radermaker F, Schumacher M. Theinduction of aromatase and sexual behavior by testosterone in male and female Japanesequail: A dose-response study. IRCSMed Sci 1986; 14: 1188–1189.
44. Balthazart J, Dupiereux V,Aste N, Viglietti-Panzica C, Barrese M, Panzica GC.Afferent andefferent connections of the sexually dimorphic medial preoptic nucleus of the male quailrevealed by in vitro transport of DiI. Cell Tissue Res 1994; 276: 455–475.
45. Balthazart J, Evrard L, Surlemont C. Effects of the non-steroidal aromatase inhibitor,R76713 on testosterone-induced sexual behavior in the Japanese quail (Coturnix coturnixjaponica).Horm Behav 1990; 24: 510–531.
46. Balthazart J, Foidart A. Brain aromatase and the control of male sexual behavior. J SteroidBiochemMol Biol 1993; 44: 521–540.
47. Balthazart J, Foidart A. Neural bases of behavioral sex differences in quail. In: Haug M,Whalen RE, Aron C, Olsen KL, Eds. The Development of Sex Differences and Similarities inBehaviour.Dordrecht/Boston/London: KluwerAcademic Publishers, 1993:
48. Balthazart J, Foidart A, Harada N. Immunocytochemical localization of aromatase in thebrain. Brain Res 1990; 514: 327–333.
113THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

49. Balthazart J, Foidart A, Hendrick JC. The induction by testosterone of aromatase activity inthe preoptic area and activation of copulatory behavior. Physiol Behav 1990; 47: 83–94.
50. Balthazart J, Foidart A, Sante P, Hendrick JC. Effects of a-methyl-para-tyrosine on mono-amine levels in the Japanese quail: Sex differences and testosterone effects. Brain Res Bull1992; 28: 275–288.
51. Balthazart J, Foidart A, Surlemont C, Harada N. Preoptic aromatase in quail: Behavioral,biochemical and immunocytochemical studies. In: Balthazart J, Ed. Hormones, Brain andBehavior in Vertebrates. 2. Behavioral Activation in Males and Females—Social Interactionsand Reproductive Physiology. Comp Physiol, Vol. 9. Basel: Karger, 1990: 45–62.
52. Balthazart J, Foidart A, Surlemont C, Harada N. Neuroanatomical specificity in theco-localization of aromatase and estrogen receptors. J Neurobiol 1991; 22: 143–157.
53. Balthazart J, Foidart A, Surlemont C, Harada N, Leranth C, Naftolin F. Immunocytochemi-cal localization of aromatase and estrogen receptors in the brain. Soc Neurosci 1990; 16:1313.
54. Balthazart J, Foidart A, Surlemont C, Harada N, Naftolin F. Neuroanatomical specificity inthe autoregulation of aromatase-immunoreactive neurons by androgens and estrogens: Animmunocytochemical study. Brain Res 1992; 574: 280–290.
55. Balthazart J, Foidart A, Surlemont C, Vockel A, Harada N. Distribution of aromatase in thebrain of the Japanese quail, ring dove, and zebra finch: An immunocytochemical study. JComp Neurol 1990; 301: 276–288.
56. Balthazart J, Foidart A, Wilson EM, Ball GF. Immunocytochemical localization of androgenreceptors in the male songbird and quail brain. J Comp Neurol 1992; 317: 407–420.
57. Balthazart J, Gahr M, Surlemont C. Distribution of estrogen receptors in the brain of theJapanese quail: An immunocytochemical study. Brain Res 1989; 501: 205–214.
58. Balthazart J, Libioulle JM, Sante Ph. Stimulatory effects of the noradrenergic neurotoxinDSP4 on sexual behavior in male quail. Behav Proc 1988; 17: 27–44.
59. Balthazart J, Massa R, Negri-Cesi P. Photoperiodic control of testosterone metabolism,plasma gonadotrophins, cloacal gland growth, and reproductive behavior in the Japanesequail.Gen Comp Endocrinol 1979; 39: 222–235.
60. Balthazart J, Sante Ph, Ball GF. Testosterone effects on the staining density and autoradio-graphic investigations of the a2-adrenergic receptor in the medial preoptic nucleus of theJapanese quail: Relationship to the activation of reproductive behavior. Arch Int PhysiolBioch 1991; 99: 385–392.
61. Balthazart J, Schoffeniels E. Pheromones are involved in the control of sexual behaviour inbirds.Naturwissensch 1979; 66: 55–56.
62. Balthazart J, Schumacher M. Organization and activation of behavior in quail: Role oftestosterone metabolism. J Exp Zool 1984; 232: 595–604.
63. Balthazart J, Schumacher M, Evrard L. Sex differences and steroid control of testosterone-metabolizing enzyme activity in the quail brain. J Neuroendocrinol 1990; 2: 675–683.
64. Balthazart J, Schumacher M, Malacarne G. Interaction of androgens and estrogens in thecontrol of sexual behavior in male Japanese quail. Physiol Behav 1985; 35: 157–166.
65. Balthazart J, Schumacher M, Ottinger MA. Sexual differences in the Japanese quail:Behavior, morphology and intracellular metabolism of testosterone. Gen Comp Endocrinol1983; 51: 191–207.
66. Balthazart J, Stoop R, Foidart A, Granneman JCM, Lambert JGD. Distribution andregulation of estrogen-2-hydroxylase in the quail brain. Brain Res Bull 1994; 35: 339–345.
67. Balthazart J, Stoop R, Foidart A, Harada N. Synergistic control by androgens and estrogensof aromatase in the quail brain.NeuroReport 1994; 5: 1729–1732.
68. Balthazart J, Surlemont C.Androgen and estrogen action in the preoptic area and activationof copulatory behavior in quail. Physiol Behav 1990; 48: 599–609.
114 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

69. Balthazart J, Surlemont C. Copulatory behavior is controlled by the sexually dimorphicnucleus of the quail POA. Brain Res Bull 1990; 25: 7–14.
70. Balthazart J, Surlemont C, Harada N.Aromatase as a cellular marker of testosterone actionin the preoptic area. Physiol Behav 1992; 51: 395–409.
71. Barraclough CA, Wise PM. The role of catecholamines in the regulation of pituitaryluteinizing hormone and follicle stimulating-hormone secretion. Endocrine Rev 1982; 3:91–119.
72. Berk ML, Butler AB. Efferent projections of the medial preoptic nucleus and medialhypothalamus in the pigeon. J Comp Neurol 1981; 203: 379–399.
73. Bernroider G, Leutgeb S. V1-receptor mediated effects of vasotocin on motivational, mne-monic and aversive components of sexual learning in quail. Soc Neurosci Abstr 1994; 20:1602.
74. Bitran D, Hull EM. Pharmacological analysis of male rat sexual behavior.Neurosci BiobehavRev 1987; 11: 365–389.
75. Blackburn JR, Pfaus JG, Phillips AG. Dopamine functions in appetitive and defensivebehaviours. Prog Neurobiol 1992; 39: 247–279.
76. Blaustein JD. Estrogen receptor immunoreactivity in rat brain: Rapid effects of estradiolinjection. Endocrinology 1993; 132: 1218–1224.
77. Blaustein JD, LehmanMN, Turcotte JC, Greene G. Estrogen receptors in dendrites and axonterminals in the guinea pig hypothalamus. Endocrinology 1992; 131: 281–290.
78. Blaustein JD, Olster DH. Gonadal steroid hormone receptors and social behaviors. In:Balthazart J, Ed. Advances in Comparative and Environmental Physiology, Vol. 3. Berlin:Springer-Verlag, 1989: 31–104.
79. Blaustein JD, Turcotte JC. Estrogen receptor-immunostaining of neuronal cytoplasmicprocesses as well as cell nuclei in guinea pig brain. Brain Res 1989; 495: 75–82.
80. Bloch GJ, Gorski RA. Cytoarchitectonic analysis of the SDN-POA of the intact and gonadec-tomized rat. J Comp Neurol 1988; 275: 604–612.
81. Bons N. The topography of mesotocin and vasotocin systems in the brain of the domesticmallard and Japanese quail: Immunocytochemical identification. Cell Tissue Res 1980; 213:37–51.
82. Boyd SK. Arginine vasotocin facilitation of advertisement calling and call phonotaxis inbullfrogs.Horm Behav 1994; 28: 232–240.
83. Breazile JE, Kuenzel WJ. Systema nervosum centrale. In: Baumel JJ, KingAS, Breazile JE,Evans HE, Vanden Berge JC, Eds. Handbook of Avian Anatomy: Nomina AnatomicumAvium. Cambridge, MA: Nuttall Ornithological Club, 1993: 493–554.
84. Breedlove SM. Sexual dimorphism in the vertebrate nervous system. J Neurosci 1992; 12:4133–4142.
85. Callard GV. Aromatization is cyclic AMP-dependent in cultured brain cells. Brain Res 1981;204: 461–464.
86. Cameron-Curry P, Aste N, Viglietti-Panzica C, Panzica GC. Immunocytochemical distribu-tion of glial fibrillary acidic protein in the central nervous system of the Japanese quail(Coturnix coturnix japonica). Anat Embryol 1991; 184: 571–581.
87. Canick JA, Tobet SA, Baum MJ, Vaccaro DE, Ryan KJ, Leeman SE, Fox TO. Studies on therole of catecholamines in the regulation of the developmental pattern of hypothalamicaromatase. Steroids 1987; 50: 510–521.
88. Canonaco M, Tavolaro R, Cerra MC, Anastasio M, Franzoni MF. Gonadal regulation ofGABAA receptors in the different brain areas of the male Japanese quail. Exp Brain Res1991; 87: 634–640.
89. Canonaco M, Tavolaro R, Cerra MC, Franzoni MF. Distribution of benzodiazepine bindingsites in the brain of the male Japanese quail and its correlation to a hormonal control:Quantitative autoradiography study.Neuroendocrinology 1992; 55: 35–43.
115THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

90. Cardinali DP, Ritta MN, Gejman PV. Norepinephrine stimulates testosterone aromatizationand inhibits 5a-reduction via b-adrenoreceptors in the rat pineal gland.Mol Cell Endocrinol1982; 28: 199–209.
91. Cheng MF. Progress and prospects in ring dove research: A personal view. Adv Study AnimBehav 1979; 9: 97–129.
92. Cheng MF. For whom does the female dove coo? A case for the role of vocal self-stimulation.Anim Behav 1992; 43: 1035–1044.
93. Cherry JA, Baum MJ. Effects of lesions of a sexually dimorphic nucleus in the preoptic/anterior hypothalamic area on the expression of androgen- and estrogen-dependent sexualbehaviors in male ferrets. Brain Res 1990; 522: 191–203.
94. Cintra A, Fuxe K, Harfstrand A, Agnati LF, Miller LS, Greene JL, Gustafsson JA. On thecellular localization and distribution of estrogen receptors in the rat tel- and diencephalonusing monoclonal antibodies to human estrogen receptors.Neurochem Int. 1986; 8: 587–595.
95. Commins D, Yahr P. Adult testosterone levels influence the morphology of a sexuallydimorphic area in the mongolian gerbil brain. J Comp Neurol 1984; 224: 132–140.
96. Cozzi B, Stankov B, Viglietti-Panzica C, Capsoni S, Aste N, Lucini V, Fraschini F, Panzica C.Distribution and characterization of melatonin receptors in the brain of the Japanese quail,Coturnix japonica. Neurosci Lett 1993; 150: 149–152.
97. Cozzi B, Viglietti-Panzica C, Aste N, Panzica GC. The serotoninergic system in the brain ofthe Japanese quail. An immunohistochemical study. Cell Tissue Res 1991; 263: 271–284.
98. Crews D. Diversity of hormone–behavior relations in reproductive behavior. In: Becker JB,Breedlove SM, Crews D, Eds. Behavioral Endocrinology. Cambridge, MA: MIT Press, 1992:143–186.
99. Crews D, Camazine B, Diamond M, Mason R, Tokarz R, Gartska WR. Hormonal indepen-dence of courtship behavior in the male garter snake.Horm Behav 1984; 18: 29–41.
100. Crews D, Gartska WR. The ecological physiology of a garter snake. Sci Am 1982; 159–168.101. Crews D, Wade J, Wilczynski W. Sexually dimorphic areas in the brain of whiptail lizards.
Brain Behav Evol 1990; 36: 262–270.102. Crosby EC, Showers MJ. Comparative anatomy of the preoptic and hypothalamic areas. In:
Haymaker W, Anderson E, Nauta WT, Eds. The Hypothalamus. Springfield: Thomas, 1969:61–135.
103. Crowley WR, O’Connor LH, Feder HH. Neurotransmitter systems and social behavior. In:Balthazart J, Ed. Molecular and Cellular Basis of Social Behavior in Vertebrates. Berlin:Springer-Verlag, 1989: 162–208.
104. Crowley WR, Zemlan FP. The neurochemical control of mating behavior. In: Adler NT, Ed.Neuroendocrinology of Reproduction.NewYork: Plenum, 1981: 451–484.
105. De Jonge FH, Louwerse AL, Ooms MP, Evers P, Endert E, Van De Poll NE. Lesions of theSDN-POA inhibit sexual behavior of male Wistar rats. Brain Res Bull 1989; 23: 483–492.
106. DeVries GJ. Sex differences in neurotransmitter systems. J Neuroendocrinol 1990; 2: 1–13.107. DeVries GJ, Crenshaw BJ, Ali Al-Shamma H. Gonadal steroid modulation of vasopressin
pathways. Ann NY Acad Sci 1992; 652: 387–396.108. Dellovade TL, Rissman EF, Thompson N, Harada N, Ottinger MA. Co-localization of
aromatase enzyme and estrogen receptor immunoreactivity in the preoptic area duringreproductive aging. Brain Res 1995; 674: 181–187.
109. Delville Y, Balthazart J. Hormonal control of female sexual behavior in the Japanese quail.Horm Behav 1987; 21: 288–309.
110. Delville Y, Sulon J, Balthazart J. Diurnal variations of sexual receptivity in the femaleJapanese quail.Horm Behav 1986; 20: 13–33.
111. Delville Y, Sulon J, Hendrick JC, Balthazart J. Effect of the presence of females on thepituitary–testicular activity in male Japanese quail (Coturnix coturnix japonica). Gen CompEndocrinol 1984; 55: 295–305.
116 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

112. Doi O, Takai T, Nakamura T, TanabeY. Changes in the pituitary and plasma LH, plasma andfollicular progesterone, and estradiol, and plasma testosterone and estrone concentrationsduring the ovulatory cycle of the quail (Coturnix coturnix japonica). Gen Comp Endocrinol1980; 41: 156–163.
113. DomjanM, Nash S. Stimulus control of social behaviour inmale Japanese quail.AnimBehav1988; 36: 1006–1015.
114. Etgen AM, Petitti N. Norepinephrine-stimulated cyclic AMP accumulation in rat hypotha-lamic slices: Effects of estrous cycle and ovarian steroids. Brain Res 1986; 375: 385–390.
115. Etgen AM, Ungar S, Petitti N. Estradiol and progesterone modulation of norepinephrineneurotransmission: Implications for the regulation of female reproductive behavior. JNeuro-endocrinol 1992; 4: 255–271.
116. Feder HH, StoreyA, Goodwin D, Reboulleau C, Silver R. Testosterone and 5a-dihydrotestos-terone levels in peripheral plasma ofmale and female ring doves (Streptopelia risoria) duringthe reproductive cycle. Biol Reprod 1977; 16: 666–677.
117. Finn PD, de Vries GJ, Yahr P. Efferent projections of the sexually dimorphic area of the gerbilhypothalamus: Anterograde identification and retrograde verification in males and females.J Comp Neurol 1993; 338: 491–520.
118. Foidart A, De Clerck A, Harada N, Balthazart J. Aromatase-immunoreactive cells in thequail brain: Effects of testosterone and sex dimorphism. Physiol Behav 1994; 55: 453–464.
119. Foidart A, Harada N, Balthazart J. Effects of steroidal and non steroidal aromataseinhibitors on sexual behavior and aromatase-immunoreactive cells and fibers in the quailbrain. Brain Res 1994; 657: 105–123.
120. Follett BK. Birds. In: Lamming GE, Ed.Marshall’s Physiology of Reproduction. Edinburgh:Longman Greene, 1984: 283–350.
121. Foster RG, Panzica GC, Parry DM, Viglietti-Panzica C. Immunocytochemical studies on theLHRH system of the Japanese quail: Influence by photoperiod and aspects of sexualdifferentiation. Cell Tissue Res 1988; 253: 327–335.
122. Fujii K, Kobayashi H. FMRFamide-like immunoreactivity in the brain of the Japanese quail,Coturnix coturnix japonica. Neuroendocrinol Lett 1992; 14: 97–102.
123. Garcia-Segura LM, Chowen JA, Duenas M, Torres-Aleman I, Naftolin F. Gonadal steroids aspromoters of neuro-glial plasticity. Psychoneuroendocrinol 1994; 19: 445–453.
124. Garcia-Segura LM, Luquın S, Parducz A, Naftolin F. Gonadal hormone regulation of glialfibrillary acidic protein immunoreactivity and glial ultrastructure in the rat neuroendocrinehypothalamus.Glia 1994; 10: 59–69.
125. Giambiagi N, Pasqualini JR. Interaction of three monoclonal antibodies with the nonacti-vated and activated forms of the estrogen receptor. Endocrinology 1990; 126: 1403–1409.
126. Gingrich JA, Caron MG. Recent advances in the molecular biology of dopamine receptors.Ann Rev Neurosci 1993; 16: 299–321.
127. Gorski RA, Gordon JH, Shryne JE, SouthamAM. Evidence for amorphological sex differencewithin the medial preoptic area of the rat brain. Brain Res 1978; 148: 333–346.
128. Gorski RA, Harlan RE, Jacobson CD, Shryne JE, SouthamAM. Evidence for the existence ofa sexually dimorphic nucleus in the preoptic area of the rat. J Comp Neurol 1980; 193:529–539.
129. Goy RW, McEwen BS. Sexual Differentiation of the Brain. Cambridge, MA: The MIT Press,1980.
130. Guyomarc’h C, Guyomarc’h JC. La stimulation du developpement sexuel des femelles decaille japonaise, Coturnix coturnix japonica, par des chants de males: mise en evidence deperiode privilegiees dans le nycthemere. C R Acad Sci Paris, Serie III 1982; 295: 37–40.
131. Guyomarc’h C, Guyomarc’h JC, Garnier DH. Influence de la perception de vocalisations demales sur la reproduction chez les femelles de Coturnix coturnix japonica. Biol Behav 1981;6: 167–182.
117THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

132. Gunturkun O. The functional organization of the avian visual system. In: Andrew RJ, Ed.Neural and Behavioural Plasticity.Oxford: Univ. Press, 1991: 92–105.
133. Gunturkun O, Miceli O, Watanabe M. Anatomy of the avian thalamofugal pathway. In:Zeigler HP, Bischof HJ, Eds. Vision, Brain and Behavior in Birds. Cambridge, MA: MITPress, 1993: 115–135.
134. Harada N. Novel properties of human placental aromatase as cytochrome P-450: Purifica-tion and characterization of a unique form of aromatase. J Biochem 1988; 103: 106–113.
135. HaradaN,Abe-Dohmae S, Loeffen R, FoidartA, Balthazart J. Synergism between androgensand estrogens in the induction of aromatase and its messenger RNA in the brain. Brain Res1993; 622: 243–256.
136. Harada N, Utsumi T, Takagi Y. Tissue-specific expression of the human aromatase cyto-chrome P-450 gene by alternative use of multiple exons 1 and promoters, and switching oftissue-specific exons 1 in carcinogenesis. Proc Natl Acad Sci USA 1993; 90: 11312–11316.
137. Harada N, Yamada K, Foidart A, Balthazart J. Regulation of aromatase cytochrome P-450(estrogen synthetase) transcripts in the quail brain by testosterone.Mol Brain Res 1992; 15:19–26.
138. Herbison AE, Robinson JE, Skinner DC. Distribution of estrogen receptor-immunoreactivecells in the preoptic area of the ewe: Co-localization with glutamic acid decarboxylase but notluteinizing hormone-releasing hormone.Neuroendocrinology 1993; 57: 751–759.
139. Herbison AE, Theodosis DT. Neurotensin-immunoreactive neurons in the rat medial preop-tic area are oestrogen-receptive. J Neuroendocrinol 1991; 3: 587–589.
140. HerbisonAE, Theodosis DT. Localization of oestrogen receptors in preoptic neurons contain-ing neurotensin but not tyrosine hydroxylase, cholecystokinin or luteinizing hormone-releasing hormone in the male and female rat.Neuroscience 1992; 50: 283–298.
141. Hinde RA. Interaction of internal and external factors in integration of canary reproduction.In: Beach FA, Ed. Sex and Behavior.NewYork: Wiley, 1965: 381–415.
142. Hines M, Davis FC, Coquelin A, Goy RW, Gorski RA. Sexually dimorphic regions in themedial preoptic area and the bed nucleus of the stria terminalis of the guinea pig brain: Adescription and investigation of their relationship to gonadal steroids in adulthood. JNeurosci 1985; 5: 40–47.
143. Horn G.Memory, Imprinting, and the Brain.Oxford: Clarendon Press, 1985.
144. Hutchison JB, Steimer TH. Formation of behaviorally effective 17b-estradiol in the dovebrain: Steroid control of preoptic aromatase. Endocrinology 1986; 118: 2180–2187.
145. Hutchison JB, Steimer Th. Brain 5b-reductase. A correlate of behavioral sensitivity toandrogen. Science 1981; 213: 244–246.
146. Inaba M, Kamata K. Effect of estradiol-17b and other steroids on noradrenaline anddopamine binding to synaptic membrane fragments of rat brain. J Steroid Biochem 1979; 11:1491–1497.
147. Jakab RL, Horvath TL, Leranth C, Harada N, Naftolin F. Aromatase immunoreactivity inthe rat brain: Gonadectomy-sensitive hypothalamic neurons and an unresponsive ‘‘limbicring’’ of the lateral septum-bed nucleus–amygdala complex. J Steroid Biochem Mol Biol1993; 44: 481–498.
148. Karten HJ, Hodos W. A Stereotaxic Atlas of the Brain of the Pigeon (Columba livia).Baltimore: Johns Hopkins, 1967.
149. Kelley DB, Pfaff DW. Generalizations from comparative studies on neuroanatomical andendocrinemechanisms of sexual behaviour. In: Hutchison JB, Ed.Biological Determinants ofSexual Behaviour. Chichester: Wiley, 1978: 225–254.
150. Kihlstrom JE, Danninge I. Neurohypophysial hormones and sexual behavior in males of thedomestic fowl (Gallus domesticus L.) and the pigeon (Columbia livia Gmel). Gen CompEndocrinol 1972; 18: 115–120.
118 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

151. Korzekwa KR, Trager WF, Mancewicz J, Osawa Y. Studies on the mechanism of aromataseand other cytochrome P450 mediated deformylation reactions. J Steroid Biochem Mol Biol1993; 44: 367–373.
152. Kuenzel WJ, Masson M. A Stereotaxic Atlas of the Brain of the Chick (Gallus domesticus).Baltimore: Johns Hopkins Univ. Press, 1988.
153. Kuenzel WJ, VanTienhovenA. Nomenclature and location of avian hypothalamic nuclei andassociated circumventricular organs. J Comp Neurol 1982; 206: 293–313.
154. Kumar V, Green S, Staub A, Chambon P. Localisation of the oestradiol-binding and putativeDNA-binding domains of the human oestrogen receptor. EMBO J 1986; 5: 2231–2237.
155. Lauber ME, Lichtensteiger W. Pre- and postnatal ontogeny of aromatase cytochrome P450messenger ribonucleic acid expression in the male rat brain studied by in situ hybridization.Endocrinology 1994; 135: 1661–1668.
156. Lehrman DS. Interaction between internal and external environments in the regulation ofthe reproductive cycle of the ring dove. In: Beach FA, Ed. Sex and Behavior.NewYork: Wiley,1965: 355–380.
157. Lephart ED, Simpson ER, Ojeda SR. Effects of cyclic AMP and androgens on in vitro brainaromatase enzyme activity during prenatal development in the rat. J Neuroendocrinol 1992;4: 29–35.
158. Liposits Z, Kallo I, Coen CW, Paull WK, Flerko B. Ultrastructural analysis of estrogenreceptor immunoreactive neurons in the medial preoptic area of the female rat brain.Histochemistry 1990; 93: 233–239.
159. Lisciotto CA, Morrell JI. Circulating gonadal steroid hormones regulate estrogen receptormRNA in the male rat forebrain.Mol Brain Res 1993; 20: 79–90.
160. MacLusky NJ, Philip A, Hurlburt C, Naftolin F. Estrogen metabolism in neuroendocrinestructures. In: Celotti F, Naftolin F, Martini L, Eds.Metabolism of Hormonal Steroids in theNeuroendocrine Structures.NewYork: Raven Press, 1984: 103–116.
161. Mason ST. Catecholamines and Behavior. Cambridge, UK: Cambridge Univ. Press, 1984.
162. McEwen BS. Steroid hormone actions on the brain: when is the genome involved? HormBehav 1994; 28: 396–405.
163. McEwen BS, Krey LC. Properties of estrogen-sensitive neurons: Aromatization, progestinreceptor induction and neuroendocrine effects. In: Celotti F, Naftolin F, Martini L, Eds.Metabolism of Hormonal Steroids in the Neuroendocrine Structures.NewYork: Raven Press,1984: 117–128.
164. Meisel RL, Sachs BD. The physiology of male sexual behavior. In: Knobil E, Neill JD et al.,Eds. The Physiology of Reproduction, Vol. 2.NewYork, Raven Press, 1994: 3–105.
165. Mendelson CR, Smith ME, Cleland WH, Simpson ER. Regulation of aromatase activity ofcultured adipose stromal cells by catecholamines and adrenocorticotropin.Mol Cell Endocri-nol 1984; 37: 61–72.
166. Meyerson BJ, Malmnas CO, Everitt BJ. Neuropharmacology, neurotransmitters, and sexualbehavior in mammals. In:Adler N, Pfaff D, Goy RW, Eds.Handbook of Behavioral Neurobiol-ogy, Vol. 7, Reproduction.NewYork: Plenum, 1985: 495–536.
167. Meyerson BJ, PalisA, SietniksA. Hormone-monoamine interactions and sexual behavior. In:Beyer C, Ed. Endocrine Control of Sexual Behavior. New York: Raven Press, 1979: 389–405.
168. Moore FL, Wood RE, Boyd SK. Sex steroids and vasotocin interact in a female amphibian(Taricha granulosa) to elicit female-like egg-laying behavior or male-like courtship. HormBehav 1992; 26: 156–166.
169. Morrell JI, Kelley DB, Pfaff DW. Sex steroid binding in the brain of vertebrates. In: KniggeKM, Scott DE, Kobayashi H, Miura S, Ishii S, Eds. Brain–Endocrine Interactions II. Basel:Karger, 1975: 230–256.
170. Morrell JI, Pfaff DW. A neuroendocrine approach to brain function: Localization of sexsteroid concentrating cells in vertebrate brains. Am Zool 1978; 18: 447–460.
119THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

171. Naftolin F, Brawer JR. The effect of estrogens on hypothalamic structure and function. Am JObst Gynecol 1978; 132: 758–765.
172. Naftolin F, Garcia-Segura LM, Keefe D, Leranth C, MacLusky NJ, Brawer JR. Estrogeneffects on the synaptology and neural membranes of the rat hypothalamic arcuate nucleus.Biol Reprod 1990; 42: 21–28.
173. Naftolin F, Leranth C, Balthazart J. Ultrastructural localization of aromatase immunoreac-tivity in hypothalamic neurons. Endocrine Soc Abstr 1990; 669: 192–192.
174. Nauta WJH, Karten HJ. A general profile of the vertebrate brain with sidelights on theancestry of the cerebral cortex. In: Quarton, Melnecek, Schmitt Eds. The NeuroscienceSecond Study Program.NewYork: Rockefeller Univ. Press, 1970: 7–26.
175. Nelson RJ. An Introduction to Behavioral Endocrinology. Sunderland, MA: Sinauer Associ-ates, Inc, 1994.
176. Nock B, Feder HH. Neurotransmitter modulation of steroid action in target cells thatmediate reproduction and reproductive behavior.Neurosci Biobehav Rev 1981; 5: 437–447.
177. Nottebohm F. Neuronal replacement in adulthood. Ann NY Acad Sci 1985; 457: 143–161.178. Nottebohm F. From bird song to neurogenesis. Sci Am 1989; 260: 74–79.179. NottebohmF,ArnoldAP. Sexual dimorphism in the vocal control areas in the song bird brain.
Science 1976; 194: 211–213.180. O’Connell ME, Reboulleau C, Feder HH, Silver R. Social interactions and androgen levels in
birds. I. Female characteristics associated with increased plasma androgen levels in themale ring dove (Streptopelia risoria).Gen Comp Endocrinol 1981; 44: 454–463.
181. Oksche A, Wilson OW, Farner DS. The hypothalamic neurosecretory system of Coturnixcoturnix japonica. Zeitschr Zellforsch 1964; 61: 688–709.
182. OsawaY, Higashiyama T, ShimizuY, Yarborough C. Multiple functions of aromatase and theactive site structure: Aromatase is the placental estrogen 2-hydroxylase. J Steroid BiochemMol Biol 1993; 44: 469–480.
183. Ottinger MA, Balthazart J. Brain monoamines in Japanese quail: Effects of castration andsteroid replacement therapy. Behav Proc 1987; 14: 197–216.
184. Ottinger MA, Scheidt WM, Russek E. Daily patterns of courtship andmating behavior in themale Japanese quail. Behav Proc 1982; 7: 223–233.
185. Ottinger MA, Schumacher M, Clarke RN, Duchala CS, Balthazart J. Comparison of mono-amine concentrations in the brains of adult male and female Japanese quail. Poultry Sci1986; 65: 1413–1420.
186. Ottinger MA, Thompson N, Dellovade T, Harada N, Rissman EF. Aging and preoptic areaaromatase immunoreactivity in male Japanese quail. Soc Neurosci Abstr 1991; 17: 1409.
187. Palkovits M. Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Res1973; 59: 449–450.
188. Palkovits M, Brownstein MJ. Microdissection of brain areas by the punch technique. In:Cuello AC, Ed. Brain Microdissection Techniques.NewYork: Wiley, 1983: 1–36.
189. Panzica GC, Viglietti-Panzica C, Sanchez F, Sante P, Balthazart J. Effects of testosterone ona selected neuronal population within the preoptic sexually dimorphic nucleus of theJapanese quail. J Comp Neurol 1991; 303: 443–456.
190. Panzica GC. Vasotocin-immunoreactive elements and neuronal typology in the suprachias-matic nucleus of the chicken and Japanese quail. Cell Tissue Res 1985; 242: 371–376.
191. Panzica GC, Aste N, Dellovade T, Rissman E, Foidart A, Balthazart J, Ottinger MA.Aromatase-containing cells of themedial preoptic nucleus respond differentially to testoster-one in young and aged male quail. Ann Endocrinol 1994; 55: 37.
192. Panzica GC,Aste N, Viglietti-Panzica C, Fasolo A. Neuronal circuits controlling quail sexualbehavior. Chemical neuroanatomy of the septo-preoptic region. Poultry Sci Rev 1992; 4:249–259.
120 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

193. Panzica GC, Aste N, Viglietti-Panzica C, Ottinger MA. Structural sex differences in thebrain: Influence of gonadal steroids and behavioral correlates. J Endocrinol Invest 1995; 18:232–252.
194. Panzica GC, Balthazart J, Viglietti-Panzica C. Anatomical and biochemical studies of thesexually dimorphic medial preoptic nucleus of the quail. In: Balthazart J, Ed. Brain andBehaviour in Vertebrates. 1. Sexual Differentiation, Neuroanatomical Aspects, Neurotransmit-ters and Neuropeptides. Comp Physiol, Vol. 8. Basel: Karger, 1990: 104–120.
195. Panzica GC, Barrese M,Aste N, Viglietti-Panzica C. Direct connections between septum andmedial preoptic nucleus of the Japanese quail. A DiI study on fixed tissue. Neurosci Lett1992; 43(Suppl.): S82.
196. Panzica GC, Calcagni M, Ramieri G, Viglietti-Panzica C. Extrahypothalamic distribution ofvasotocin-immunoreactive fibers and and perikarya in the avian central nervous system.Bas Appl Histochem 1988; 32: 89–94.
197. Panzica GC, Castagna C,Aste N, Balthazart J, Viglietti-Panzica C. Morphometrical analysisof testosterone-induced ultrastructural changes in neurons of male quail medial preopticnucleus.Neuroendocrinology 1994; 60(Suppl. 1): 98.
198. Panzica GC, Fraschini F, Aste N, Lucini V, Viglietti-Panzica C, Cozzi B, Stankov B. Thedensity of melatonin receptors is dependent upon the prevailing photoperiod in the Japanesequail (Coturnix japonica).Neurosci Lett 1994; 173: 111–114.
199. Panzica GC, Malacarne G, De Bernochi A, Viglietti-Panzica GC. Effects of steroid hormoneson the neuropil of the hypothalamic paraventricular nucleus of male chickens. Cell TissueRes 1985; 240: 169–174.
200. Panzica GC, Spigolon S, Castagna C. Ultrastructural characterization of the sexuallydimorphic medial preoptic nucleus of the male Japanese quail. Cell Tissue Res 1995; 279:517–527.
201. Panzica GC, Viglietti-Panzica C, Balthazart J. Sex dimorphism of the avian medial preopticnucleus: Cytoarchitecture, morphometry and hormonal control. Acta Anat 1987; 130: 70.
202. Panzica GC, Viglietti-Panzica C, Calcagni M, Anselmetti GC, Schumacher M, Balthazart J.Sexual differentiation and hormonal control of the sexually dimorphic preoptic medialnucleus in quail. Brain Res 1987; 416: 59–68.
203. Panzica GC, Viglietti-Panzica C, Fasolo A, Vandesande F. CRF like immunoreactive systemin the quail brain. J Hirnforsch 1986; 27: 539–547.
204. Panzica GC, Viglietti-Panzica C, Fiori MG, Calcagni M, Anselmetti GC, Balthazart J.Cytoarchitectural analysis of the quail preoptic area. Evidence for a sex-related dimorphismin the medial preoptic nucleus. Bull Zool 1987; 54: 13–17.
205. Pfaff DW. The neuroanatomy of sex hormone receptors in the vertebrate brain. In: AnandKumar TC, Ed.Neuroendocrine Regulation of Fertility. Basel: Karger, 1976: 30–45.
206. Pfaff DW. Estrogen and Brain Function.NewYork: Springer-Verlag, 1980.207. Pfaff DW. Features of a hormone-driven defined neural circuit for a mammalian behavior.
Principles illustrated, neuroendocrine syllogisms, and multiplicative steroid effects. Ann NYAcad Sci 1989; 563: 131–147.
208. Pfaff DW, Schwartz-Giblin S, McCarthy MM, Kow LM. Cellular and molecular mechanismsof female reproductive behaviors. In: Knowbil E, Neill JD, Eds. The Physiology of Reproduc-tion.NewYork: Raven Press, 1994: 107–220.
209. Phoenix CH, Goy RW, Gerall AA, Young WC. Organizational action of prenatally adminis-tered testosterone propionate on the tissues mediating behavior in the female guinea pig.Endocrinology 1959; 65: 369–382.
210. Press MF, Greene GL. Recent development in the use of anti-receptor antibodies to studysteroid hormone receptors. In: Clark CR, Ed. Steroid Hormone Receptors. Their IntracellularLocalization.Weinheim: Verlag Chemie, 1987: 251–275.
211. Raisman G, Field PM. Sexual dimorphism in the neuropil of the preoptic area of the rat andits dependence on neonatal androgens. Brain Res 1973; 54: 1–29.
121THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

212. Rasika S, Nottebohm F, Alvarez-Buylla A. Testosterone increases the recruitment and/orsurvival of new high vocal center neurons in adult female canaries. Proc Natl Acad Sci USA1994; 91: 7854–7858.
213. Raum WJ, Marcano M, Swerdloff RS. Nuclear accumulation of estradiol derived from thearomatization of testosterone is inhibited by hypothalamic beta-receptor stimulation in theneonatal female rat. Biol Reprod 1984; 30: 388–396.
214. Raum WJ, Swerdloff RS. The role of hypothalamic adrenergic receptors in preventingtestosterone-induced androgenization in the female rat brain. Endocrinology 1981; 109:273–278.
215. Reger RL, Gerall AA, Givon L. Distribution and hormonal alterations of immunoreactiveestrogen receptors in the brain of prairie voles, Microtus ochrogaster. Abstr Conf ReprodBehav, Saratoga Springs, NY 1989; 62.
216. Richfield ER, Young AB, Penney JB. Comparative distribution of dopamine D-1 and D-2receptors in the basal ganglia of turtles, pigeons, rats, cats, and monkeys. J Comp Neurol1987; 262: 446–463.
217. Roselli CE. Subcellular localization and kinetic properties of aromatase in the rat brain. SocNeurosci Abstr 1994; 20: 1770.
218. Roselli CE, Horton LE, Resko JA. Time-course and steroid specificity of aromatase inductionin rat hypothalamus–preoptic area. Biol Reprod 1987; 37: 628–633.
219. Roselli CE, Resko JA. Androgens regulate brain aromatase activity in adult male ratsthrough a receptor mechanism. Endocrinology 1984; 114: 2183–2189.
220. Sanghera MK, Simpson ER, McPhaul MJ, Kozlowski G, ConleyAJ, Lephart ED. Immunocy-tochemical distribution of aromatase cytochrome P450 in the rat brain using peptide-generated polyclonal antibodies. Endocrinology 1991; 129: 2834–2844.
221. Sayag N, Hoffman NW, Yahr P. Telencephalic connections of the sexually dimorphic area ofthe gerbil hypothalamus that influence male sexual behavior. Behav Neurosci 1994; 108:743–757.
222. Scaletta LL, Hull EM. Systemic or intracranial apomorphine increases copulation in long-term castrated male rats. Pharmacol Biochem Behav 1990; 37: 471–475.
223. Schlinger BA, Callard GV. Localization of aromatase in synaptosomal and microsomalsubfractions of quail (Coturnix coturnix japonica) brain. Neuroendocrinology 1989; 49:434–441.
224. Schumacher M. Rapid membrane effects of steroid hormones: an emerging concept inneuroendocrinology. Trends Neurosci 1990; 13: 359–362.
225. Schumacher M, Alexandre C, Balthazart J. Interactions des androgenes et des oestrogenesdans le controle de la reproduction. C R Acad Sci Paris, Serie III 1987; 305: 569–574.
226. Schumacher M, Balthazart J. The effects of testosterone and its metabolites on sexualbehavior and morphology in male and female Japanese quail. Physiol Behav 1983; 30:335–339.
227. Schumacher M, Balthazart J. Sexual dimorphism of the hypothalamic metabolism oftestosterone in the Japanese quail (Coturnix coturnix japonica). Prog Brain Res 1984; 61:53–61.
228. Schumacher M, Balthazart J. Testosterone-induced brain aromatase is sexually dimorphic.Brain Res 1986; 370: 285–293.
229. Schumacher M, Balthazart J. Neuroanatomical distribution of testosterone metabolizingenzymes in the Japanese quail. Brain Res 1987; 422: 137–148.
230. Schumacher M, Contenti E, Balthazart J. Testosterone metabolism in discrete areas of thehypothalamus and adjacent brain regions of male and female Japanese quail. Brain Res1983; 278: 337–340.
231. Schumacher M, Hendrick JC, Balthazart J. Sexual differentiation in quail: critical periodand hormonal specificity.Horm Behav 1989; 23: 130–149.
122 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

232. Schumacher M, Sulon J, Balthazart J. Changes in serum concentrations of steroids duringembryonic and post-hatching development of male and female Japanese quail (Coturnixcoturnix japonica). J Endocrinol 1988; 118: 127–134.
233. Schwartz JH, Kandel ER. Synaptic transmission mediated by second messengers. In:Kandel ER, Schwartz JH, Jessel TM, Eds. Principles of Neural Science. New York: Elsevier,1991: 173–193.
234. Segovia S, GuillamonA. Sexual dimorphism in the vomeronasal pathway and sex differencesin reproductive behaviors. Brain Res Rev 1993; 18: 51–74.
235. Sharp PJ, Follett BK. The distribution of monoamines in the hypothalamus of the Japanesequail, Coturnix coturnix japonica. Zeitschr Zellforsch 1968; 90: 245–262.
236. Shinoda K, Mori S, Ohtsuki T, Osawa Y. An aromatase-associated cytoplasmic inclusion, the‘‘stigmoid body,’’ in the rat brain. I. Distribution in the forebrain. J Comp Neurol 1992; 322:360–376.
237. Shinoda K, Yagi H, Fujita H, Osawa Y, Shiotani Y. Screening of aromatase-containingneurons in rat forebrain: An immunohistochemical study with antibody against humanplacental antigen X-P2 (hPAX-P2). J Comp Neurol 1989; 290: 502–515.
238. Shivers BD, Harlan RE, Morrell JI, Pfaff DW. Absence of oestradiol concentration in cellnuclei of LHRH-immunoreactive neurones.Nature 1983; 304: 345–347.
239. Silver R, Ramos C. Vasoactive intestinal polypeptide in avian reproduction. In: BalthazartJ., Ed. Hormones, Brain and Behaviour in Vertebrates. 1. Sexual Differentiation, Neuroana-tomical Aspects, Neurotransmitters and Neuropeptides. Comp Physiol, Vol. 8. Basel: Karger,1990: 191–204.
240. Silver R, Witkovsky P, Horvath P, Alones V, Barnstable CJ, Lehman MN. Coexpression ofopsin- and VIP-like immunoreactivity in CSF-contacting neurons of the avian brain. CellTissue Res 1988; 253: 189–198.
241. SilvermanA-J, DonCarlos LL,Morrell JI. Ultrastructural characteristics of estrogen receptor-containing neurons of the ventrolateral nucleus of the guinea-pig hypothalamus. J Neuroen-docrinol 1991; 3: 623–634.
242. Simerly RB, Chang C, Muramatsu M, Swanson LW. Distribution of androgen and estrogenreceptor mRNA-containing cells in the rat brain: An in situ hybridization study. J CompNeurol 1990; 294: 76–95.
243. Simpson ER, Mahendroo MS, Means GD, Kilgore MW, Corbin CJ, Mendelson CR. Tissue-specific promoters regulate aromatase cytochrome P450 expression. J Steroid Biochem MolBiol 1993; 44: 321–330.
244. Simpson ER, Mahendroo MS, Means GD, Kilgore MW, Hinshelwood MM, Graham-LorenceS, Amarneh B, Ito Y, Fisher CR, Michael MD, Mendelson CR, Bulun SE. Aromatasecytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocrine Rev 1994; 15:342–355.
245. Steimer Th.Aromatase activity in rat brain synaptosomes. Is an enzyme associated with theneuronal cell membrane involved in mediating non-genomic effects of androgens. Eur JNeurosci Suppl 1988; 9.
246. Stokes TM, Leonard CM, Nottebohm F. The telencephalon, diencephalon, and mesencepha-lon of the canary, Serinus canaria, in stereotaxic coordinates. J Comp Neurol 1974; 156:337–374.
247. StumpfWE, SarM. Steroid hormone target sites in the brain: The differential distribution ofestrogen, progestin, androgen and glucocorticosteroid. J Steroid Biochem 1976; 7: 1163–1170.
248. Stumpf WE, Sar M. Anatomical distribution of estrogen, androgen, progestin, corticoid andthyroid hormone target sites in the brain of mammals: phylogeny and ontogeny. Am Zool1978; 18: 435–445.
249. Stumpf WE, Sar M, Keefer DA. Atlas of estrogen target cells in rat brain. In: Stumpf WE,Grant LD, Eds. Anatomical Neuroendocrinology. Basel: Karger, 1975: 104–119.
123THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

250. Swaab DF, Fliers E. A sexually dimorphic nucleus in the human brain. Science 1985; 228:1112–1115.
251. Sanchez F, Panzica GC, Viglietti-Panzica C, Aste N, Carretero J, Vasquez R. A comparativeanalysis of the vasotocin and vasopressin systems in the chicken and rat hypothalamus. Animmunocytochemical study. J Hirnforsch 1991; 32: 27–37.
252. Takami S, Urano A. The volume of the toad medial amygdala–anterior preoptic complex issexually dimorphic and seasonally variable.Neurosci Lett 1984; 44: 253–258.
253. Theodosis DT, Poulain DA. Evidence for structural plasticity in the supraoptic nucleus of therat hypothalamus in relation to gestation and lactation.Neuroscience 1984; 11: 183–193.
254. Theodosis DT, Poulain DA. Neuronal–glial and synaptic plasticity in the adult rat paraven-tricular nucleus. Brain Res 1989; 484: 361–366.
255. Theodosis DT, Poulain DA. Activity-dependent neuronal-glial and synaptic plasticity in theadult mammalian hypothalamus.Neuroscience 1993; 57: 501–535.
256. Thompson R, Adkins-Regan E. Photoperiod affects the morphology of a sexually dimorphicnucleus within the preoptic area of male Japanese quail. Brain Res 1994; 667: 201–208.
257. Thompson RR, Adkins-Regan E. Ontogeny of a sexually dimorphic nucleus in the preopticarea of the Japanese quail (Coturnix japonica).Dev Brain Res 1992; 70: 231–237.
258. Tobet SA, Zahniser DJ, Baum MJ. Sexual dimorphism in the preotic/anterior hypothalamicarea of ferrets: Effects of adult exposure to sex steroids. Brain Res 1986; 364: 249–257.
259. Tobet SA, Zahniser DJ, Baum MJ. Differentiation in male ferrets of a sexually dimorphicnucleus of the preoptic/anterior hypothalamic area requires prenatal estrogen.Neuroendocri-nology 1986; 44: 299–308.
260. Torres-Aleman I, Rejas MT, Pons S, Garcia-Segura LM. Estradiol promotes cell shapechanges and glial fibrillary acid protein redistribution in hypothalamic astrocytes in vitro: aneuronal-mediated effect.Glia 1992; 6: 180–187.
261. Tsuruo Y, Ishimura K, Fujita H, Osawa Y. Immunocytochemical localization of aromatase-containing neurons in the rat brain during pre- and postnatal development. Cell Tissue Res1994; 278: 29–39.
262. Van Gils J, Absil P, Grauwels L, Moons L, Vandesande F, Balthazart J. Distribution ofluteinizing hormone-releasing hormones I and II (LHRH-I and -II) in the quail and chickenbrain as demonstrated with antibodies directed against synthetic peptides. J Comp Neurol1993; 334: 304–323.
263. Van Gils J, Absil P, Moons L, Grauwels L, Vandesande F, Balthazart J. Distribution ofb-endorphin-like-immunoreactive structures in the chicken and quail brain as demonstratedwith a new homologous antibody directed against a synthetic peptide. J Comp Neurol 1994;350: 382–396.
264. VerhoevenG. Effect of neurotransmitters and follicle-stimulating hormone on the aromatiza-tion of androgens and the production of 38,58-monophosphate by cultured testicular cells. JSteroid Biochem 1980; 12: 315–322.
265. Verhoeven GP, Dierckx P, de Moor P. Stimulation effect of neurotransmitters on thearomatization of testosterone by Sertoli cell-enriched cultures.Mol Cell Endocrinol 1979; 13:241–253.
266. Viglietti-Panzica C. Immunohistochemical study of the distribution of vasotocin reactingneurons in avian diencephalon. J Hirnforsch 1986; 27: 559–566.
267. Viglietti-Panzica C,Absil P, Panzica GC,Aste N, Balthazart J. Immunohistochemical studieson vasotocin innervation of aromatase-containing regions in the male quail forebrain. SocNeurosci Abstr 1994; 20: 1739.
268. Viglietti-Panzica C, Anselmetti GC, Balthazart J, Aste N, Panzica GC. Vasotocinergicinnervation of the septal region in the Japanese quail: Sexual differences and the influence oftestosterone. Cell Tissue Res 1992; 267: 261–265.
124 PANZICA, VIGLIETTI-PANZICA, AND BALTHAZART
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant

269. Viglietti-Panzica C,Aste N, Balthazart J, Panzica GC. Vasotocinergic innervation of sexuallydimorphic medial preoptic nucleus of the male Japanese quail: Influence of testosterone.Brain Res 1994; 657: 171–184.
270. Viglietti-Panzica C, Panzica GC. Peptidergic neurons in the avian brain. Ann Sci Natur, ZoolParis 1991; 12: 137–155.
271. Viglietti-Panzica C, Panzica GC, Fiori MG, Calcagni M, Anselmetti GC, Balthazart J. Asexually dimorphic nucleus in the quail preoptic area.Neurosci Lett 1986; 64: 129–134.
272. Viglietti-Panzica, C, Spigolon S,Aste N, Balthazart J, Panzica GC. Ultrastructural modifica-tions of the male quail medial preoptic nucleus after adult testosterone treatment. Eur JNeurosci Suppl 1991; 4: 225.
273. Voorhuis TAM, Kiss JZ, de Kloet ER, de Wied D. Testosterone-sensitive vasotocin-immunoreactive cells and fibers in the canary brain. Brain Res 1988; 442: 139–146.
274. Vowles DM, Beazley L, Harwood DH. Stereotaxic atlas of the brain of the barbary dove(Streptopelia risoria). In: Wright P, Caryl PG, Vowles D, Eds. Neural and Endocrine Aspectsof Behaviour in Birds. Amsterdam: Elsevier, 1975: 351–394.
275. Wagner CK, Morrell JI. The distribution of aromatase mRNA in the brain of adult male andfemale rats using in situ hybridization. Soc Neurosci Abstr 1994; 20: 1740.
276. Watson JT, Adkins-Regan E. Neuroanatomical localization of sex steroid-concentrating cellsin the Japanese quail (Coturnix japonica): Autoradiography with [3H]-testosterone, [3H]-estradiol, and [3H]-dihydrotestosterone.Neuroendocrinology 1989; 49: 51–64.
277. Watson JT, Adkins-Regan E. Testosterone implanted in the preoptic area of male Japanesequail must be aromatized to activate copulation.Horm Behav 1989; 23: 432–447.
278. Watson JT, Adkins-Regan E. Activation of sexual behavior by implantation of testosteronepropionate and estradiol benzoate into the preoptic area of the male Japanese quail(Coturnix japonica).Horm Behav 1989; 23: 251–268.
279. Weiner RI, Findell PR, Kordon C. Role of classic and peptide neuromediators in theneuroendocrine regulation of LH and prolactin. In: Knobil E, Neill J, et al., Eds. ThePhysiology of Reproduction.NewYork: Raven Press, 1988: 1235–1281.
280. Wenzel BM. Chemoreception. In: Farner DS, King JR, Parkes KC, Eds. Avian Biology, Vol. 3.NewYork: Academic Press, 1973: 389–415.
281. Yahr P, Finn PD, Hoffman NW, Sayag N. Sexually dimorphic cell groups in the medialpreoptic area that are essential for male sex behavior and the neural pathways needed fortheir effects. Psychoneuroendocrinology 1994; 19: 463–470.
282. Yahr P, Gregory JE. The medial and lateral cell groups of the sexually dimorphic area of thegerbil hypothalamus are essential for male sex behavior and act via separate pathways.Brain Res 1993; 631: 287–296.
283. Yamada S, Mikami S. Immunocytochemical localization of neurotensin-containing neuronsin the hypothalamus of the Japanese quail, Coturnix coturnix japonica. Cell Tissue Res 1981;218: 29–39.
284. Yamada S, Mikami SI. Immunohistochemical localization of corticotropin releasing factor(CRF)-containing neurons in the hypothalamus of the Japanese quail. Cell Tissue Res 1985;239: 299–304.
285. Yamauchi K, Yasuda M. Cyto-, dendro- and fibro-architectonic studies on the chickenhypothalamus. J Hirnforsch 1985; 26: 509–519.
286. Yuri K, Kawata M. The effect of estrogen on the estrogen receptor-immunoreactive cells inthe rat medial preoptic nucleus. Brain Res 1991; 548: 50–54.
287. Zeier H, Karten HJ. The archistriatum of the pigeon: Organization of afferent and efferentconnections. Brain Res 1971; 31: 313–326.
125THE SEXUALLY DIMORPHIC NUCLEUS OF THE QUAIL POA
FIN 133@sp1/disk3/CLS_jrnl/GRP_finn/JOB_finnps/DIV_145z02 jant




















