
The Role of Early Fungal Colonizers in Leaf-Litter Decomposition in Portuguese Streams Impacted by...
11
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim 1434-2944/09/408-0399 Internat. Rev. Hydrobiol. 94 2009 4 399–409 DOI: 10.1002/iroh.200811154 K. R. SRIDHAR 1 , SOFIA DUARTE 2 , FERNANDA CÁSSIO 2 and CLÁUDIA PASCOAL* , 2 1 Microbiology and Biotechnology, Department of Biosciences, Mangalore University, Mangalore 574 199, India 2 Centre of Molecular and Environmental Biology (CBMA), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal; e-mail: [email protected] Research Paper The Role of Early Fungal Colonizers in Leaf-Litter Decomposition in Portuguese Streams Impacted by Agricultural Runoff key words: leaf decomposition, aquatic hyphomycetes, nutrient loading, transplant experiment Abstract We conducted a transplant experiment between two streams in NW Portugal impacted by agricultural runoff, mainly differing in phosphate concentration, to determine whether fungi on decomposing leaves would adapt to the new environment or would be replaced by fungi of the recipient stream. The most nutrient enriched stream had lower fungal diversity but faster leaf decomposition. Leaf transplanta- tion did not alter fungal activity or species dominance. Multidimensional scaling ordination of fungal communities, from DNA fingerprint or conidial production, revealed that transplanted communities resembled more those of the original stream than the recipient stream. Results suggest that early fungal colonizers will determine the development and activity of fungal communities on decomposing leaves in streams impacted by agricultural practices. 1. Introduction In low-order temperate streams, the canopy cover can constrain light availability restrict- ing primary production by autotrophs (ALAN and CASTILLO, 2007). In such ecosystems, leaf litter constitutes the main source of nutrients and energy for microbial decomposers and invertebrate detritivores (BÄRLOCHER, 2005). Both fungi and bacteria colonize plant detritus submerged in streams; however, higher biomass of fungi than bacteria has been often found on decomposing leaves pointing to a major role of fungi in leaf decomposition (PASCOAL and CÁSSIO, 2004; BALDY et al., 2007). Fungi, particularly aquatic hyphomycetes, release extracellular enzymes capable to degrade the polysaccharides of leaf cell walls and their hyphae penetrate leaf tissues, causing softening and improving leaf palatability for inverte- brates (BÄRLOCHER, 2005). Aquatic hyphomycete diversity and functions are strongly influenced by nutrient con- centrations in the stream water. Usually, elevated concentrations of inorganic N and P are reported to stimulate the activity of fungi (e.g., biomass accrual, reproduction, respiration or productivity), leading to the enhancement of organic matter turnover (SRIDHAR and BÄR- LOCHER, 2000; GULIS and SUBERKROPP, 2003; STELZER et al., 2003; PASCOAL and CÁSSIO, 2004; PASCOAL et al., 2003, 2005a; FERREIRA et al., 2006). Fungal diversity is also reported * Corresponding author
Transcript of The Role of Early Fungal Colonizers in Leaf-Litter Decomposition in Portuguese Streams Impacted by...

© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim 1434-2944/09/408-0399
Internat. Rev. Hydrobiol. 94 2009 4 399–409
DOI: 10.1002/iroh.200811154
K. R. SRIDHAR1, SOFIA DUARTE2, FERNANDA CÁSSIO2 and CLÁUDIA PASCOAL*, 2
1Microbiology and Biotechnology, Department of Biosciences, Mangalore University, Mangalore 574 199, India
2Centre of Molecular and Environmental Biology (CBMA), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal; e-mail: [email protected]
Research Paper
The Role of Early Fungal Colonizers in Leaf-Litter Decomposition in Portuguese Streams Impacted by Agricultural Runoff
key words: leaf decomposition, aquatic hyphomycetes, nutrient loading, transplant experiment
Abstract
We conducted a transplant experiment between two streams in NW Portugal impacted by agricultural runoff, mainly differing in phosphate concentration, to determine whether fungi on decomposing leaves would adapt to the new environment or would be replaced by fungi of the recipient stream. The most nutrient enriched stream had lower fungal diversity but faster leaf decomposition. Leaf transplanta-tion did not alter fungal activity or species dominance. Multidimensional scaling ordination of fungal communities, from DNA fingerprint or conidial production, revealed that transplanted communities resembled more those of the original stream than the recipient stream. Results suggest that early fungal colonizers will determine the development and activity of fungal communities on decomposing leaves in streams impacted by agricultural practices.
1. Introduction
In low-order temperate streams, the canopy cover can constrain light availability restrict-ing primary production by autotrophs (ALAN and CASTILLO, 2007). In such ecosystems, leaf litter constitutes the main source of nutrients and energy for microbial decomposers and invertebrate detritivores (BÄRLOCHER, 2005). Both fungi and bacteria colonize plant detritus submerged in streams; however, higher biomass of fungi than bacteria has been often found on decomposing leaves pointing to a major role of fungi in leaf decomposition (PASCOAL and CÁSSIO, 2004; BALDY et al., 2007). Fungi, particularly aquatic hyphomycetes, release extracellular enzymes capable to degrade the polysaccharides of leaf cell walls and their hyphae penetrate leaf tissues, causing softening and improving leaf palatability for inverte-brates (BÄRLOCHER, 2005).
Aquatic hyphomycete diversity and functions are strongly influenced by nutrient con-centrations in the stream water. Usually, elevated concentrations of inorganic N and P are reported to stimulate the activity of fungi (e.g., biomass accrual, reproduction, respiration or productivity), leading to the enhancement of organic matter turnover (SRIDHAR and BÄR-LOCHER, 2000; GULIS and SUBERKROPP, 2003; STELZER et al., 2003; PASCOAL and CÁSSIO, 2004; PASCOAL et al., 2003, 2005a; FERREIRA et al., 2006). Fungal diversity is also reported
* Corresponding author

400 K. R. SRIDHAR et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
to be higher in nutrient enriched streams compared to oligotrophic streams (GULIS and SUBERKROPP, 2004; ARTIGAS et al., 2008).
The transfer of colonized leaf litter from one stream to another with contrasting water chemistry has been used as an useful approach to measure how aquatic hyphomycetes react to sudden alterations in their habitat condition (SUBERKROPP and CHAUVET, 1995; SRIDHAR et al., 2005; DUARTE et al., 2008). Nevertheless, these transplantation experiments gave vari-able results that appear to be dependent on water chemistry and the initial fungal community (ROSSET and BÄRLOCHER, 1985; SUBERKROPP and CHAUVET, 1995; SRIDHAR et al., 2005). For example, fungal communities did not change when leaves colonized in a nutrient enriched stream were transplanted to a nutrient poor stream, but in the reverse experiment the initial community was rapidly replaced by the indigenous one (SUBERKROPP and CHAUVET, 1995). The transfer of leaves colonized at a moderately metal polluted stream to a highly metal polluted stream lowered fungal sporulation and almost stopped leaf decomposition (SRIDHAR et al., 2005). However, if the initial fungal community is highly diverse, it appears to mitigate the impact of pollution on leaf decomposition (DUARTE et al., 2008).
Although water chemistry can alter fungal communities at various stages of decay, the initial imprint of aquatic fungal species received by leaf litter in a stream appears to deter-mine the subsequent development of fungal communities and thus leaf-litter decomposition (ROSSET and BÄRLOCHER, 1985; SRIDHAR and BÄRLOCHER, 2000; DUARTE et al., 2008). In order to elucidate this issue, we conducted a transplant experiment in two streams impact-ed by agricultural runoff, but greatly differing in phosphorus concentrations, to determine whether fungal communities on decomposing leaves would adapt to new environmental conditions or would be replaced by fungi of the recipient stream.
2. Material and Methods
2.1. Study Site and Field Experiment
The Souto (N: 41º31′09.15′′; W: 8º17′12.75′′) and the Donim (N: 41º31′34.43′′; W: 8º17′16.03′′) streams are located in the Ave River basin, Northwest of Portugal, in an area with high population density and intensive agricultural activities. At the Souto stream, the selected site has about 20 cm depth and 1 m width with granite rocks and pebbles at the bottom, while the Donim stream has about 30 cm depth and 90 cm width with gravel and muddy substrate. At both the sites, the riparian vegetation is mainly Alnus glutinosa (L.) GAERTN. and Quercus robur L.
Temperature, pH, redox potential, oxygen concentration and conductivity of the stream water were measured in situ with field probes (Multiline F/set 3 no 400327, WTW) and water samples were col-lected at each sampling date to determine nitrate concentration by cadmium reduction (HACH kit, program 355; 0–30.0 mg L–1 N–NO3
–), reactive phosphorus concentration by the ascorbic acid method (HACH kit, program 490; 0–2.5 mgL–1 PO4
3–), ammonia concentration by the salicylate method (HACH kit, program 385; 0–0.5 mg L–1 N–NH3) and nitrite concentration by the diazotization method (HACH kit, program 371; 0–0.3 mg L–1 N–NO2
–) using a HACH DR/2000 photometer (Hach Company, Love-land, CO, USA). Moreover, standard solutions were used for each method calibration.
Alnus glutinosa leaves, collected during October 2005 and dried at room temperature, were weighted in 4 g groups and distributed into 66 fine-mesh bags (0.5 mm mesh size; 15 × 15 cm). Thirty-three leaf bags were immersed at each stream and after 7 days of immersion (to allow leaf colonization by native fungi, see e.g., PASCOAL and CÁSSIO, 2004; PASCOAL et al., 2005a), 12 leaf bags were transplanted from Souto to Donim streams and vice versa. Three leaf bags were randomly sampled from each site after 0, 7, 14, 21, 28 and 35 days of leaf immersion, between 12th May and 16th June 2006. At the labora-tory, leaves from each bag were rinsed with deionized water, to remove sediments and invertebrates, and cut into disks (12 mm diameter). Two subsets of leaf disks were freeze-dried (48 h) for subsequent ergosterol analysis (8 leaf disks) and DNA extraction (4 leaf disks) and a third subset (5 leaf disks) was used for aeration to generate spores. The rest of the leaves were dried at 80ºC for 48 ± 4 h, and cooled to room temperature before weighted to the nearest 0.01 mg. At day 0, three leaf bags were immersed

Early Fungal Colonizers in Portuguese Streams 401
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
in the study streams for 10 min, and returned to the laboratory where dry mass was estimated as above, to allow the calculation of initial air-dry mass to initial oven-dry mass conversion factor.
2.2. Fungal Biomass and Sporulation
Fungal biomass on leaf disks was estimated from ergosterol concentration according to GESSNER (2005). Lipids were extracted from leaves by heating (80 ºC, 30 min.) in KOH/methanol (8 g L–1) and the extract was purified by solid-phase extraction. Ergosterol was quantified by HPLC using a LiChrospher RP18 column (250 × 4 mm, Merck), protected by guard columns, and connected to a Beckmann Gold liquid chromatographic system. The system was run isocratically with HPLC-grade methanol (Riedel de-Haën), at 1.4 mL min–1 at 33 ºC, and ergosterol was detected at 282 nm. Standard series of ergosterol (Fluka) in isopropanol were used to estimate the ergosterol concentration in leaf samples.
Fungal sporulation was induced by placing 5 leaf disks into 100 mL Erlenmeyer flasks containing 50 mL sterilized stream water (121 ºC, 20 min.), under continuous aeration with aquarium pumps at 18 ºC. After 48 h, conidial suspensions were mixed with 200 μL of 0.5% of Tween 80 and filtered (5 μm pore size, Millipore); the trapped conidia were stained with 0.05% cotton blue in lactic acid (Fluka). Conidia were identified under a light microscope at 400× magnification (Leica) and at least 300 conidia were counted per filter.
2.3. Molecular Analyses
DNA was isolated from a total of 4 halves of 4 freeze-dried leaf disks with the soil DNA extrac-tion kit (MoBio Laboratories, Solana Beach, California), according to manufacturer instructions. For determination of fungal diversity, the primer pair ITS3GC and ITS4, which amplifies the ITS2 region of fungal ribosomal DNA (WHITE et al., 1990), was used. The ITS3 primer had an additional 40-bp GC tail on the 5’end, which ensures separation on DGGE gels (MUYZER et al., 1993). For polymerase chain reaction (PCR), 1× of Taq buffer (mixture of KCl: (NH4)2 SO4 buffers), 3 mM of MgCl2, 0.2 mM of dNTPs, 0.4 μM of each primer, 1.5 U of DNA Taq polymerase and 1 μL of DNA in a final volume of 50 μL were used. All PCR reagents were purchased from MBI Fermentas, except for primers, which were supplied by MWG-Biotech AG. PCR was carried out in an iCycler Thermal Cycler (BioRad Laboratories, Hercules, CA, USA).
The amplification program started with an initial denaturation of 2 min. at 95 °C, followed by 36 cycles of denaturation for 30 s at 95 °C, primer annealing for 30 s at 55 °C and extension for 1 min. at 72 °C. Final extension was at 72 °C for 5 min. (NIKOLCHEVA et al., 2005).
Denaturing Gradient Gel Electrophoresis (DGGE) analysis was performed using a DCodeTM Univer-sal Mutation Detection System (BioRad Laboratories, Hercules, CA, USA). Samples of 750 ng from the amplification products of 380–400 bp were loaded on 8% (w/v) polyacrilamide gels in 1× TAE with a denaturing gradient from 30 to 70% (100% denaturant corresponds to 40% formamide and 7 M urea). The gels were run at 55 V, 56 °C for 16 h and stained for 5 min. with 1 μg mL–1 of ethidium bro-mide (BioRad). A mixture of DNA of the fungal species Alatospora acuminata INGOLD (UMB-173.01), Articulospora tetracladia INGOLD (UMB-22.01), Anguillospora filiformis GREATH. (UMB-225.02), Lem-onniera aquatica DE WILD. (UMB-143.01), Tricladium chaetocladium INGOLD (UMB-163.01) and Vari-cosporium elodeae W. KEGEL (UMB-20.01) was included in the DGGE gel as ladder to calibrate and normalize the gel for further analyses. The gel images were captured under UV light in a transiluminator Eagle eye II (Stratagene, La Jolla, CA, USA).
2.4. Statistical Analyses
The remaining dry mass of alder leaves was fit to the exponential model mt = m0 · e–kt, where mt is the leaf dry mass remaining at time t, m0 is the initial leaf dry mass, and k is the rate of leaf decomposi-tion. Regression lines of ln-transformed values of leaf dry mass were compared by ANCOVA followed by Tukey’s tests (ZAR, 1996).
Two-way analyses of variance (Two-way ANOVA) were used to test whether stream and time signifi-cantly influenced the fungal biomass and sporulation (ZAR, 1996). To achieve normal distribution, data

402 K. R. SRIDHAR et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
from fungal biomass and sporulation were ln transformed (ZAR, 1996). Differences in water parameters between streams were compared by a paired t-test (ZAR, 1996).
Shannon’s diversity (H′) and Pielou’s equitability (J′) indices were used to assess the diversity of
sporulating aquatic fungi as: H′ = – ∑ i = 1
s Pi (ln Pi) and J′ = H′/ln S, where Pi is the relative abundance of
conidia of taxon i, and S is the total number of sporulating taxa (LEGENDRE and LEGENDRE, 1998).The DGGE gel was aligned and normalized using Gelcompar II (Applied Maths, Belgium) and fungal
diversity was determined by counting the number of bands in each gel. Differences between fungal communities were assessed by nonmetric multidimensional scaling
(MDS) of sporulating species or DNA fingerprints (KRUSKAL and WISH, 1978). In MDS ordination, samples are represented as points in a low-dimension space, and the relative distances between points are in the same rank order as the relative dissimilarities of the samples as measured by an appropri-ate resemblance matrix calculated from the data matrix. Pearson coefficient was used to quantify the correlation between indigenous and transplanted fungal communities, for both sporulation and DNA fingerprinting data.
The ANCOVA, ANOVA and MDS analyses were done with Statistica 6.0 for Windows (Statsoft, Inc.), while Shannon’s and Pielou’s indices were calculated by Primer v6 software package.
3. Results
3.1. Physical and Chemical Characterization of the Stream Water
The analysis of stream water parameters indicated that pH (6.0 vs. 6.6), conductivity (50.5 vs. 98.0 μS cm–1) and concentrations of nitrite (0.005 vs. 0.01 mg L–1 N–NO2
–), ammonium (0.01 vs. 0.07 mg L–1 N–NH4
+) and phosphate (0.06 vs. 0.6 mg L–1 P–PO42–) were signifi-
cantly lower in the Souto stream than in the Donim stream (t-test, P < 0.05), while redox potential (60.7 vs. 18.0 mV) was significantly higher in the Souto stream than in the Donim stream (t-test, P = 0.0007; Table 1).
3.2. Leaf Decomposition and Fungal Activity
Mass loss of alder leaves followed an exponential decay model, with approximately 60% of the initial leaf mass being lost in the Donim stream and 40% in the Souto stream after 35 days (Fig. 1). Leaf decomposition rate was significantly higher in the Donim stream than
Table 1. Physical and chemical characteristics of the stream water in the Souto stream (S) and the Donim stream (D).
Parameter S D n
pH 6.0 ± 0.1 6.6 ± 0.07 6Temperature (ºC) 13.2 ± 0.1 13.9 ± 0.3 6Oxygen (mg L–1) 9.7 ± 0.3 9.1 ± 0.2 6Redox potential (mV) 60.7 ± 4.3 18.0 ± 5.2 4Conductivity (μS cm–1) 50.5 ± 2.6 98.0 ± 8.7 6N–NH4
+ (mg L–1) 0.01 ± 0.004 0.07 ± 0.04 5N–NO3
– (mg L–1) 2.4 ± 0.3 2.4 ± 0.3 5N–NO2
– (mg L–1) 0.01 ± 0.002 0.02 ± 0.002 5P–PO4
3– (mg L–1) 0.07 ± 0.04 0.6 ± 0.2 5
Mean ± SE; n = number of samples.

Early Fungal Colonizers in Portuguese Streams 403
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
in the Souto stream (Tukey test, P = 0.003). The transfer of alder leaves from the Souto stream to the Donim stream or vice versa did not significantly alter leaf decomposition rates (Table 2; Tukey test, P > 0.05).
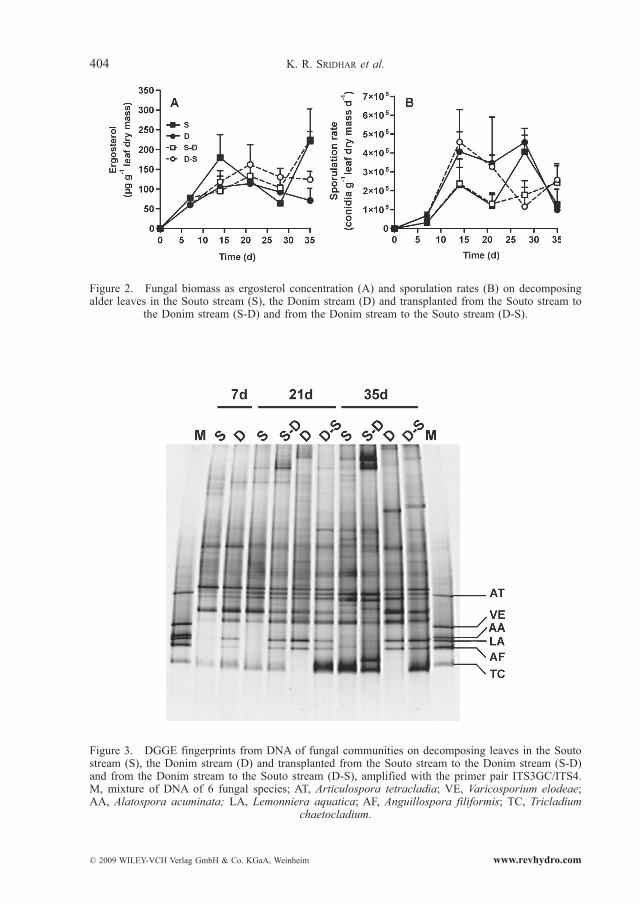
Fungal biomass on alder leaves attained maximum values after 21 days of leaf immersion in the Donim stream (114 μg ergosterol g–1 leaf dry mass) and after 35 days in the Souto stream (221 μg ergosterol g–1 leaf dry mass) (Fig. 2A). Fungal biomass was affected by time of leaf immersion, but did not differ between streams (Two-way ANOVA, P < 0.05 and P > 0.05 for time and streams, respectively). The transfer of leaves from the Souto stream to the Donim stream and vice versa did not affect ergosterol content (Two-way ANOVA, P > 0.05).
The rates of sporulation of aquatic fungi on decomposing leaves did not differ between streams (Two-way ANOVA, P > 0.05), and peak values of 4.1 × 105 and 4.6 × 105 conidia g–1 leaf dry mass d–1 were found in the Souto stream and in the Donim stream, respectively, after 28 days (Fig. 2B). However, an earlier peak (14 days) with a similar magnitude was observed in the Donim stream. The transfer of leaves between streams did not alter fungal sporulation rates (Two-way ANOVA, P > 0.05).
3.3. Fungal Diversity
During the study, a total of 19 species of aquatic hyphomycetes were found sporulating on leaves immersed in the Souto stream, while only 14 species were found in the Donim
Table 2. Decomposition rates (k) of alder leaves immersed in the Souto stream (S), the Donim stream (D) and transplanted from the Souto stream to the Donim stream (S-D) and from the Donim stream to the Souto stream (D-S). SE, standard error; W0, intercept; r2, coefficient of determination; n = 18. Identical superscript letters indicate no significant dif-
ferences (ANCOVA, Tukey test P > 0.05).
Site k (d–1) ± SE W0 (%) r2
S 0.014 ± 0.002a 88.0 0.77D 0.022 ± 0.003b 93.7 0.79S-D 0.016 ± 0.002a 90.3 0.79D-S 0.018 ± 0.003a, b 92.5 0.75
Figure 1. Leaf dry mass remaining of decomposing alder leaves in the Souto stream (S), the Donim stream (D) and transplanted from the Souto stream to the Donim stream (S-D) and from the Donim
stream to the Souto stream (D-S).
404 K. R. SRIDHAR et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Figure 2. Fungal biomass as ergosterol concentration (A) and sporulation rates (B) on decomposing alder leaves in the Souto stream (S), the Donim stream (D) and transplanted from the Souto stream to
the Donim stream (S-D) and from the Donim stream to the Souto stream (D-S).
Figure 3. DGGE fingerprints from DNA of fungal communities on decomposing leaves in the Souto stream (S), the Donim stream (D) and transplanted from the Souto stream to the Donim stream (S-D) and from the Donim stream to the Souto stream (D-S), amplified with the primer pair ITS3GC/ITS4. M, mixture of DNA of 6 fungal species; AT, Articulospora tetracladia; VE, Varicosporium elodeae; AA, Alatospora acuminata; LA, Lemonniera aquatica; AF, Anguillospora filiformis; TC, Tricladium
chaetocladium.

Early Fungal Colonizers in Portuguese Streams 405
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
stream (Table 3). Shannon’s and Pielou’s indices were higher in the Souto stream than in the Donim stream. Dominant sporulating species were Articulospora tetracladia (56–79%) in the Souto stream and Anguillospora filiformis (52–99%) in the Donim stream. Transfer of leaves between streams did not alter the dominant conidial producers or fungal diversity on decomposing leaves.
Fungal DNA fingerprinting from DGGE showed the presence of 21 different bands on leaves decomposing in the Souto stream and 19 bands in the Donim stream (Fig. 3). Some DGGE bands appeared in almost all fingerprints and matched with bands from DNA of pure cultures of A. tetracladia, A. filiformis and Lemonniera aquatica.
Table 3. Percentage contribution of aquatic hyphomycete species to conidial production on decomposing leaves along time in the Souto stream (S), the Donim stream (D) and trans-planted from the Souto stream to the Donim stream (S-D) and from the Donim stream to
the Souto stream (D-S).
Species 7 d 21 d 35 d
S D S S-D D D-S S S-D D D-S
Alatospora acuminata INGOLD 0.8 <0.1 0.1Anguillospora crassa INGOLD 0.7Anguillospora filiformis GREATH. 1.0 96 9.1 21.7 99 86 24.3 25.9 51.8 94.5Anguillospora longissima (SACC. and SYD.) INGOLD
0.2 <0.1 1.3 0.2
Articulospora antipodea ROLDAN and HONRUBIA ined. <0.1Articulospora tetracladia INGOLD
79 1.6 72 77 0.6 9.9 55.8 33 1.5 4.8
Clavatospora longibrachiata (INGOLD) SV. NILSSON ex MARVANOVÁ and SV. NILSSON
0.2
Flagellospora curta J. WEBSTER 7.8 1.3 2.0 0.7 0.6 12.8 5.6 4.0 <0.1Flagellospora curvula INGOLD 2.0 0.2Geniculospora grandis (GREATH.) SV. NILSSON and NOLAN
0.2 0.2 0.9
Heliscus lugdunensis SACC. and THÉRRY
0.3 0.4 <0.1 3.0
Lemonniera alabamensis R.C. SINCLAIR and MORGAN-JONES
4.6 0.4 0.2 4.0 3.4 0.1
Lemonniera aquatica DE WILD. 2.1 <0.1 29.0Tetrachaetum elegans INGOLD 0.2Tetracladium marchalianum DE WILD.
0.5 0.2
Tricladium chaetocladium INGOLD
5.3 <0.1 10.7 0.1 3.1 2.3 25.2 0.2
Tricladium splendens INGOLD 0.2 0.9 0.4 4.0Triscelophorus monosporus INGOLD
0.1
Triscelophorus sp. 0.5 0.9 0.1 9.1
Nº of species 13 7 13 5 3 7 5 10 8 6
Shannon’s index 0.50 0.19 0.74 0.52 0.043 0.55 0.90 1.48 1.24 0.20
Pielou’s index 0.19 0.10 0.30 0.32 0.039 0.28 0.56 0.64 0.60 0.11

406 K. R. SRIDHAR et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Multidimensional scaling (MDS) ordination based on spore identification (Fig. 4A) and DNA fingerprints (Fig. 4B) described well the overall differences in aquatic fungal assem-blages since stress values were low. The MDS ordination showed that fungal communities decomposing in the Souto stream or transplanted to the Donim stream were separated from those decomposing in the Donim stream or transplanted to the Souto stream (Fig. 4), except for communities assessed as DNA fingerprints in the Donim stream after 7 days. These findings suggest that transplanted fungal communities resembled more those of the original stream than the recipient stream.
4. Discussion
Environmental pollution usually results in a decrease of diversity, biomass and ecosys-tem functions of microbial communities. The diversity and ecological functions of aquatic hyphomycetes in polluted waters depends on the type and extent of pollution. For instance, in heavy metal polluted surface and ground waters, an impoverishment of aquatic hyphomyc-ete communities occurred, but leaf decomposition became compromised only at high lev-els of metal contamination (KRAUSS et al., 2005; SRIDHAR et al., 2000, 2001). Although
Figure 4. MDS ordination from Pearson coefficients of fungal diversity on decomposing leaves in the Souto stream (S), the Donim stream (D) and transplanted from the Souto stream to the Donim stream (S-D) and from the Donim stream to the Souto stream (D-S). Fungal diversity was assessed from spore
identification (A) and DNA fingerprinting (B).

Early Fungal Colonizers in Portuguese Streams 407
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
80% of aquatic hyphomycetes were lost due to organic pollutants in an Indian stream, leaf decomposition was not severely hampered (RAVIRAJA et al., 1998). Similarly, in a low-order Portuguese stream impacted by high nutrient load and heavy metals, the decline of aquatic hyphomycete diversity was not accompanied by decreased fungal biomass or leaf decompo-sition (PASCOAL et al., 2005b).
In our study, the streams had high nutrient concentrations due to agricultural runoff, but had contrasting phosphate concentrations. As expected, faster leaf decomposition was found in the most nutrient-enriched stream (Donim stream), in which fungal sporulation peaked early. On the other hand, species richness and diversity indices were higher in the Souto stream than in the Donim stream. The extent of impacts due to agricultural runoff might not be severe enough to compromise fungal performances, as seen in other polluted streams (e.g., SRIDHAR et al., 2000; PASCOAL et al., 2005b). Based on the algal response to mine drainage (acidic pH, high Zn concentration), NIYOGI et al. (2002) hypothesized that even though the diversity is very sensitive to low or moderate stress, the biomass of communi-ties and ecosystem functions may elevate or remain stable, and only decline when the stress is high. In our study, the response of aquatic hyphomycetes to nutrient enrichment in both streams supports this hypothesis and suggests that redundancy among these fungi may occur. However, the identity of dominant fungal species (Articulospora tetracladia in the Souto stream; Anguillospora filiformis in the Donim stream) seems to be important in driving leaf litter decomposition under the stress imposed by agricultural runoff in these Portuguese streams, highlighting that traits of certain species may have a greater influence on ecosystem functioning than species richness per se.
Leaf breakdown rates in streams can differ according to different nutrient regimes. Addi-tions of N and P in streams accelerate leaf breakdown and microbial activity on leaves (GULIS and SUBERKROPP, 2003). At a moderately heavy metal polluted site, in spite of depressed fungal diversity and reproduction, the exponential decay rate of A. glutinosa leaves was high (k = 0.065 d–1), which was predicted from the high levels of N and P in the stream water (SRIDHAR et al., 2001). In our study, the rate of leaf decomposition was higher in the Donim stream than in the Souto stream, probably due to elevated P concentration in the stream water. However, changes in nutrient concentrations seemed to have a marginal effect, because the functions of the early colonizers on leaves were not altered after transplantation. Indeed, leaf decomposition, fungal biomass and sporulation did not change by switching decomposing leaves between streams. These results are in agreement with others showing that the transfer of pre-colonized leaves to polluted streams appears to buffer the impacts of pollution on microbial decomposing activity (SRIDHAR et al. 2005; DUARTE et al., 2008). Some aquatic hyphomycetes may be sensitive to pollution, and if such species are important to leaf litter decomposition, the extent of stream pollution may selectively affect at least their reproductive potential and thus their ecological functions at longer times.
The dominance of single fungal species on decomposing leaves was evident throughout our study, with A. tetracladia and A. filiformis contributing to more than 50% to the total conidial production in the Souto stream and the Donim stream, respectively. When the leaves were switched between streams, the pattern of fungal dominance did not change much. This was also supported by MDS analysis, from microscopic conidial examination and DNA fin-gerprinting, in which transplanted fungal communities resembled more those of the original stream than the recipient stream. These findings suggest that the early fungal colonizers rapidly adjust their performances to the available resources and are not easily replaced by new sets of species. This probably contributes to explain why ergosterol content and leaf mass loss were not significantly altered after leaf transfer.
Overall, our study suggests that the conditions of the selected streams were conducive to fungal colonization and leaf decomposition. The early fungal colonizers in these streams took-over and utilized the available resources of leaves, and the subsequent switching to less favourable or more favourable situations did not alter biomass and reproductive potential of

408 K. R. SRIDHAR et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
fungi and did not lead to replacement of species. Thus, early fungal colonizers of leaf litter in streams affected by agricultural runoff appear to be crucial in organic matter turnover and addressing this issue in additional streams with different agricultural runoff history will be rewarding. It would be pertinent to investigate further whether agricultural runoff favours single species dominance on submerged leaf litter or selectively inhibit fungal species in terms of biomass build-up or sporulation. If agricultural runoff will continue for several years, will sensitive fungal species adapt to the new existing environment? Responses to these questions will help to understand up to what extent nutrient load interfere with decom-poser communities and their functions, and possibly contribute to develop management strategies to minimize the impacts of agricultural runoff in streams.
5. Acknowledgements
K. R. SRIDHAR is grateful to Mangalore University (India) for granting leave to visit the Centre of Molecular and Environmental Biology (CBMA) of the University of Minho (Portugal), and acknowl-edges the support of the CBMA during the tenure of this research. S. DUARTE was supported by the Portuguese Foundation for the Science and Technology (SFRH/BD/13482/2003).
6. References
ALAN, J. D. and M. M. CASTILLO, 2007: Stream Ecology. 2nd ed, Springer, Dordrecht, The Nether-lands.
ARTIGAS, J., A. M. ROMANI and S. SABATER, 2008: Effect of nutrients on the sporulation and diversity of aquatic hyphomycetes on submerged substrata in a Mediterranean stream. – Aquat. Bot. 88: 32–38.
BALDY, V., V. GOBERT, F. GUÉROLD, E. CHAUVET, D. LAMBRIGOT and J.-Y. CHARCOSSET, 2007: Leaf litter breakdown budgets in streams of various trophic status: effects of dissolved inorganic nutrients on microorganisms and invertebrates. – Freshw. Biol. 52: 1322–1335.
BÄRLOCHER, F., 2005: Freshwater fungal communities. – In: DEIGHTON, J., P. OUDEMANS and J. WHITE (eds.), The fungal community: its organization and role in the ecosystem, 3rd ed., Taylor and Francis, CRC Press, Boca Raton, Florida, pp. 39–59.
DUARTE, S., C. PASCOAL and F. CÁSSIO, 2008: High diversity of fungi may mitigate the impact of pol-lution on plant litter decomposition in streams. – Microb. Ecol. 56: 688–695.
FERREIRA, V., V. GULIS and M. A. S. GRAÇA, 2006: Whole-stream nitrate addition affects litter decom-position and associated fungi but not invertebrates. – Oecologia 149: 718–729.
GESSNER, M. O., 2005: Ergosterol as a measure of fungal biomass. – In: Graça, M. A. S., F. BÄRLOCHER and M. O. GESSNER (eds.), Methods to study litter decomposition: a practical guide, Springer, Berlin, pp. 171–176.
GULIS, V. and K. SUBERKROPP, 2003: Leaf litter decomposition and microbial activity in nutrient-enriched and unaltered reaches of a headwater stream. – Freshw. Biol. 48: 123–134.
GULIS, V. and K. SUBERKROPP, 2004: Effects of whole-stream nutrient enrichment on the concentration and abundance of aquatic hyphomycete conidia in transport. – Mycologia 96: 57–65.
KRAUSS, G., K. R. SRIDHAR and F. BÄRLOCHER, 2005: Aquatic hyphomycetes and leaf decomposition in contaminated groundwater wells in Central Germany. – Arch. Hydrobiol. 162: 416–428.
KRUSKAL, J. B. and M. WISH, 1978: Multidimensional scaling. Sage Publications, Beverly Hills.LEGENDRE, P. and L. LEGENDRE, 1998: Numerical Ecology. Developments in environmental modeling,
20, 2nd ed., Elsevier, Amsterdam.MUYZER, G., E. C. DE WAAL and A. G. UITTERLINDEN, 1993: Profiling of complex microbial populations
by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes cod-ing for 16S rRNA. – Appl. Environ. Microbiol. 59: 695–700.
NIKOLCHEVA, L. G., T. BOURQUE and F. BÄRLOCHER, 2005: Fungal diversity during initial stages of leaf decomposition in a stream. – Mycol. Res. 109: 246–253.

Early Fungal Colonizers in Portuguese Streams 409
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
NIYOGI, D. K., W. M. JR. LEWIS and D. M. MCKNIGHT, 2002: Effects of stress from mine drainage on diversity, biomass, and function of primary producers in mountain streams. – Ecosystems 5: 554–567.
PASCOAL, C., M. PINHO, F. CÁSSIO and P. GOMES, 2003: Assessing structural and functional ecosystem condition using leaf breakdown: studies on a polluted river. – Freshw. Biol. 48: 2033–2044.
PASCOAL, C. and F. CÁSSIO, 2004: Contribution of fungi and bacteria to leaf litter decomposition in a polluted river.– Appl. Environ. Microbiol. 70: 5266–5273.
PASCOAL, C. and F. CÁSSIO, A. MARCOTEGUI, B. SANZ and P. GOMES, 2005a: The role of fungi, bac-teria, and invertebrates in leaf litter breakdown in a polluted river. – J. N. Am. Benthol. Soc. 24: 784–797.
PASCOAL, C., F. CÁSSIO and L. MARVANOVÁ, 2005b: Anthropogenic stress may affect aquatic hyphomyc-ete diversity more than leaf decomposition in a low order stream. – Arch. Hydrobiol. 162: 481–496.
RAVIRAJA, N. S., K. R. SRIDHAR and F. BÄRLOCHER, 1998: Breakdown of Ficus and Eucalyptus leaves in an organically polluted river in India: fungal diversity and ecological functions. – Freshw. Biol. 39: 537–545.
ROSSET, J. and F. BÄRLOCHER, 1985: Transplant experiments with aquatic hyphomycetes. – Verh. Inter-nat. Verein. Limnol. 22: 2786–2790.
SRIDHAR, K. R. and F. BÄRLOCHER, 2000: Initial colonization, nutrient supply and fungal activity on leaves decaying in streams. – Appl. Environ. Microbiol. 66: 1114–1119.
SRIDHAR, K. R., G. KRAUSS, F. BÄRLOCHER, R. WENNRICH and G. J. KRAUSS, 2000: Fungal diversity in heavy metal polluted waters in central Germany. – Fungal Divers. 5: 119–29.
SRIDHAR, K. R., G. KRAUSS, F. BÄRLOCHER, N. S. RAVIRAJA, R. WENNRICH, R. BAUMBACH and G.-J. KRAUSS, 2001: Decomposition of alder leaves in two heavy metal polluted streams in Central Ger-many. – Aquat. Microb. Ecol. 26: 73–80.
SRIDHAR, K. R., F. BÄRLOCHER, G.-J. KRAUSS and G. KRAUSS, 2005: Response of aquatic hyphomycete communities to changes in heavy metal exposure.– Internat. Rev. Hydrobiol. 90: 21–32.
STELZER, R. S., J. B. HEFFERNAN, and G. E. LIKENS, 2003: The Influence of dissolved nutrients and par-ticulate organic matter quality on microbial respiration and biomass in a forest stream. – Freshw. Biol. 48: 1925–1937.
SUBERKROPP, K. and E. CHAUVET, 1995: Regulation of leaf breakdown by fungi in streams: influences of water chemistry. – Ecology 76: 1433–1445.
WHITE, T. J., T. BRUNS, S. LEE and J. W. TAYLOR, 1990: Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. – In: INNIS, M. A., D. H. GELFAND, J. J. SNINSKY and T. J. WHITE (eds.), PCR protocols: a guide to methods and applications, Academic Press, Inc., New York, USA, pp. 315–322.
ZAR, J. H., 1996: Biostatistical Analysis, 3rd ed., Prentice-Hall, Englewood Cliffs, NJ, USA.
Manuscript received September 28th, 2008; accepted March 1st, 2009




















