
The recent decline of a New Zealand endemic: how and why did populations of Archey's frog Leiopelma...
11
The recent decline of a New Zealand endemic: how and why did populations of Archey’s frog Leiopelma archeyi crash over 1996–2001? Ben D. Bell a, * , Scott Carver a , Nicola J. Mitchell a , Shirley Pledger b a School of Biological Sciences, Victoria University of Wellington, P.O. Box 600, Wellington, New Zealand b School of Mathematical and Computing Sciences, Victoria University of Wellington, P.O. Box 600, Wellington, New Zealand Received 6 January 2004; received in revised form 9 February 2004; accepted 10 February 2004 Abstract Dramatic changes have been documented in New Zealand’s vertebrate faunas since human settlement, involving major declines and extinctions, but over recent years few species have declined in numbers so rapidly as the terrestrial Archey’s frog Leiopelma archeyi (Anura: Leiopelmatidae). Long-term monitoring over more than 20 years revealed a major population reduction of the species over 1996–2001 and L. archeyi is now classified as Nationally Critical under the New Zealand threat classification system. The decline progressed northwards in the Coromandel ranges, and mostly larger (female) frogs survived. On a 100 m 2 study plot at Tapu Ridge, annual population estimates averaged 433 frogs (SE 32) over 1984–1994, declining by 88% to average 53 frogs (SE 8) over 1996–2002. A mean annual survival rate of 82% for most years declined to 33% over 1994–1997. There is mounting ev- idence to suggest that disease is the major agent of decline, supported by (1) the rapidity and severity of decline, (2) the progressive (south to north) nature of decline, and (3) finding frogs with chytriodiomycosis from Batrachochytrium dendrobatidis at the time of decline. Surprisingly, sympatric populations of the semi-aquatic Leiopelma hochstetteri have not declined dramatically, nor has a western population of L. archeyi at Whareorino, despite chytridiomycosis occurring in some frogs there. Sustaining and restoring populations of L. archeyi in New Zealand raises major challenges for conservation management. Ó 2004 Elsevier Ltd. All rights reserved. Keywords: Amphibian declines; Batrachochytrium dendrobatidis; Biodiversity conservation; Demography; Leiopelma archeyi; Leiopelma hochstetteri 1. Introduction Impacts of human settlement have had major effects on New Zealand’s terrestrial vertebrate populations, involving major declines and extinctions (King, 1984). Over recent years, however, few endemic vertebrates have declined in numbers so rapidly as Archey’s frog Leiopelma archeyi. This species is one of three extant terrestrial Leiopelma (L. archeyi, Leiopelma hamiltoni, Leiopelma pakeka) that inhabit forests and open ridge tops, while one semi-aquatic species (Leiopelma hoch- stetteri) occurs in wetter habitats alongside creeks and damp watercourses, and three other species are extinct (Worthy, 1987a; Bell et al., 1998a). Historical declines are thought to be due mainly to the impact of intro- duced mammalian predators and/or competitors, and to loss of habitat through forest removal (Bell, 1985a; Worthy, 1987a; Newman, 1996). The genus Leiopelma is one of the two least-derived (most ‘primitive’) living anuran genera, and as such, extant species are of high conservation value. Until recently L. archeyi was widely distributed, generally at higher altitude (400–830 m), through much of the Coromandel Peninsula on New Zealand’s North Island, with a new population being discovered in the northern King Country, 150 km SW of the Coromandel Peninsula in 1991 (Bell, 1994; Thurley and Bell, 1994; Bell et al., 1998b). In both the Coromandel Peninsula and the King Country, L. archeyi broadly occurs in * Corresponding author. Fax: +64-4-463-5331. E-mail address: [email protected] (B.D. Bell). 0006-3207/$ - see front matter Ó 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocon.2004.02.011 Biological Conservation 120 (2004) 189–199 www.elsevier.com/locate/biocon BIOLOGICAL CONSERVATION
Transcript of The recent decline of a New Zealand endemic: how and why did populations of Archey's frog Leiopelma...

BIOLOGICAL
CONSERVATION
Biological Conservation 120 (2004) 189–199
www.elsevier.com/locate/biocon
The recent decline of a New Zealand endemic: how and whydid populations of Archey’s frog Leiopelma archeyi crash
over 1996–2001?
Ben D. Bell a,*, Scott Carver a, Nicola J. Mitchell a, Shirley Pledger b
a School of Biological Sciences, Victoria University of Wellington, P.O. Box 600, Wellington, New Zealandb School of Mathematical and Computing Sciences, Victoria University of Wellington, P.O. Box 600, Wellington, New Zealand
Received 6 January 2004; received in revised form 9 February 2004; accepted 10 February 2004
Abstract
Dramatic changes have been documented in New Zealand’s vertebrate faunas since human settlement, involving major declines
and extinctions, but over recent years few species have declined in numbers so rapidly as the terrestrial Archey’s frog Leiopelma
archeyi (Anura: Leiopelmatidae). Long-term monitoring over more than 20 years revealed a major population reduction of the
species over 1996–2001 and L. archeyi is now classified as Nationally Critical under the New Zealand threat classification system.
The decline progressed northwards in the Coromandel ranges, and mostly larger (female) frogs survived. On a 100 m2 study plot at
Tapu Ridge, annual population estimates averaged 433 frogs (SE �32) over 1984–1994, declining by 88% to average 53 frogs (SE
�8) over 1996–2002. A mean annual survival rate of 82% for most years declined to 33% over 1994–1997. There is mounting ev-
idence to suggest that disease is the major agent of decline, supported by (1) the rapidity and severity of decline, (2) the progressive
(south to north) nature of decline, and (3) finding frogs with chytriodiomycosis from Batrachochytrium dendrobatidis at the time of
decline. Surprisingly, sympatric populations of the semi-aquatic Leiopelma hochstetteri have not declined dramatically, nor has a
western population of L. archeyi at Whareorino, despite chytridiomycosis occurring in some frogs there. Sustaining and restoring
populations of L. archeyi in New Zealand raises major challenges for conservation management.
� 2004 Elsevier Ltd. All rights reserved.
Keywords: Amphibian declines; Batrachochytrium dendrobatidis; Biodiversity conservation; Demography; Leiopelma archeyi; Leiopelma hochstetteri
1. Introduction
Impacts of human settlement have had major effectson New Zealand’s terrestrial vertebrate populations,
involving major declines and extinctions (King, 1984).
Over recent years, however, few endemic vertebrates
have declined in numbers so rapidly as Archey’s frog
Leiopelma archeyi. This species is one of three extant
terrestrial Leiopelma (L. archeyi, Leiopelma hamiltoni,
Leiopelma pakeka) that inhabit forests and open ridge
tops, while one semi-aquatic species (Leiopelma hoch-
stetteri) occurs in wetter habitats alongside creeks and
damp watercourses, and three other species are extinct
* Corresponding author. Fax: +64-4-463-5331.
E-mail address: [email protected] (B.D. Bell).
0006-3207/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.biocon.2004.02.011
(Worthy, 1987a; Bell et al., 1998a). Historical declines
are thought to be due mainly to the impact of intro-
duced mammalian predators and/or competitors, and toloss of habitat through forest removal (Bell, 1985a;
Worthy, 1987a; Newman, 1996). The genus Leiopelma is
one of the two least-derived (most ‘primitive’) living
anuran genera, and as such, extant species are of high
conservation value.
Until recently L. archeyi was widely distributed,
generally at higher altitude (400–830 m), through much
of the Coromandel Peninsula on New Zealand’s NorthIsland, with a new population being discovered in the
northern King Country, 150 km SW of the Coromandel
Peninsula in 1991 (Bell, 1994; Thurley and Bell, 1994;
Bell et al., 1998b). In both the Coromandel Peninsula
and the King Country, L. archeyi broadly occurs in

Fig. 1. Location of study sites in the Coromandel Peninsula and at
Whareorino, New Zealand.
190 B.D. Bell et al. / Biological Conservation 120 (2004) 189–199
sympatry with L. hochstetteri. L. archeyi lives and
breeds in cool, secluded terrestrial sites under the cover
of rocks, logs or vegetation under forest or on open
mist-prone ridges. It is endotrophic and exoviviparous
(following the terminology of Altig and Johnston, 1989)laying 1–10 eggs, with the male undertaking parental
care of eggs and the hatchlings (Archey, 1922; Bell,
1985b; Bell and Wassersug, 2003).
Global declines of amphibians have been of interna-
tional concern for more than a decade (Wake, 1991;
Waldman and Tocher, 1998; Alford et al., 2001; Gard-
ner, 2001; Halliday, 2001), and declines of New Zealand
frogs were feared before they eventuated (Bell, 1996;Newman, 1996). In January 1995, 29 dead L. archeyi
were collected by the Department of Conservation on
Tokatea Ridge after a prolonged drought (R. Chappell,
pers. commun.). By late 1996 a major population re-
duction on Tapu Ridge was detected. Concerns in-
creased when a further population reduction of L.
archeyi was recorded in 1998 on Tokatea Ridge. In 1999
Batrachochytrium dendrobatidis, a fungal pathogen, wasrecognised for the first time in New Zealand in Litoria
raniformis (Waldman et al., 2001) and pathological in-
vestigations were initiated. In July 2001 we found a dead
L. archeyi from relatively pristine forest on Te Moehau,
Coromandel, infected with chytrid fungus. L. archeyi is
now classified as Nationally Critical under the New
Zealand threat classification system (Hitchmough,
2002). In this paper we detail evidence of the dramaticpopulation reductions of L. archeyi in New Zealand,
and we review possible reasons for these declines.
2. Materials and methods
2.1. Study areas
Since the early 1970s, we have surveyed populations
of L. archeyi in three study areas on the Coromandel
Peninsula on the North Island of New Zealand–Tapu
Ridge, Tokatea Ridge and Te Moehau Mountain
(Fig. 1). Periodic observations have also been made in
Whareorino Forest, in the northern King Country, since
the discovery of L. archeyi there in 1991.
2.2. Population monitoring
Unlike many anuran amphibians, Leiopelma species
have only limited vocalisations, hence population
monitoring based on acoustic surveys is not possible.
Leiopelma are instead located by daytime searches of
retreat sites (under rocks or logs or within dense vege-
tation), or by counting numbers of emerged individualsat night. Our most rigorous population estimates for L.
archeyi were obtained from an annual survey of a 10�10
m study plot on Tapu Ridge that has operated since
1983 (a 5�5 m sector of the same plot was first used in
1982). Frogs were located by day in progressive searchesacross the study plot. Captured frogs were weighed,
measured (snout-vent and tibiofibula), and, if new, in-
dividually toe-clipped before release at the site of cap-
ture at the end of the search. Toe samples taken over
1993–2003 were preserved in 70% ethanol and are
available for further study, including retrospective as-
sessment of chytrid fungus infection. Younger frogs
were aged on body size, with reference to known lengthsat metamorphosis determined from captive breeding
studies, and from growth curves of known-age individ-
uals successively recaptured on the study plot (Bell,
1978, 1994). L. archeyi cannot be sexed on external
morphology, except for females attaining a greater body
size than males (Bell, 1978). The sex of some females was
confirmed through observation of yolky eggs through
the abdominal wall (Bell, 1978, 1985b). Those adultsfound guarding eggs or hatchlings at nest sites are
deemed to be males, as the male undertakes parental
care in terrestrial Leiopelma species (Bell, 1985b). Many
individuals have multiple recaptures over many years, so
their ultimate body-size can be established, increasing
the likelihood that their sex can be estimated from size.
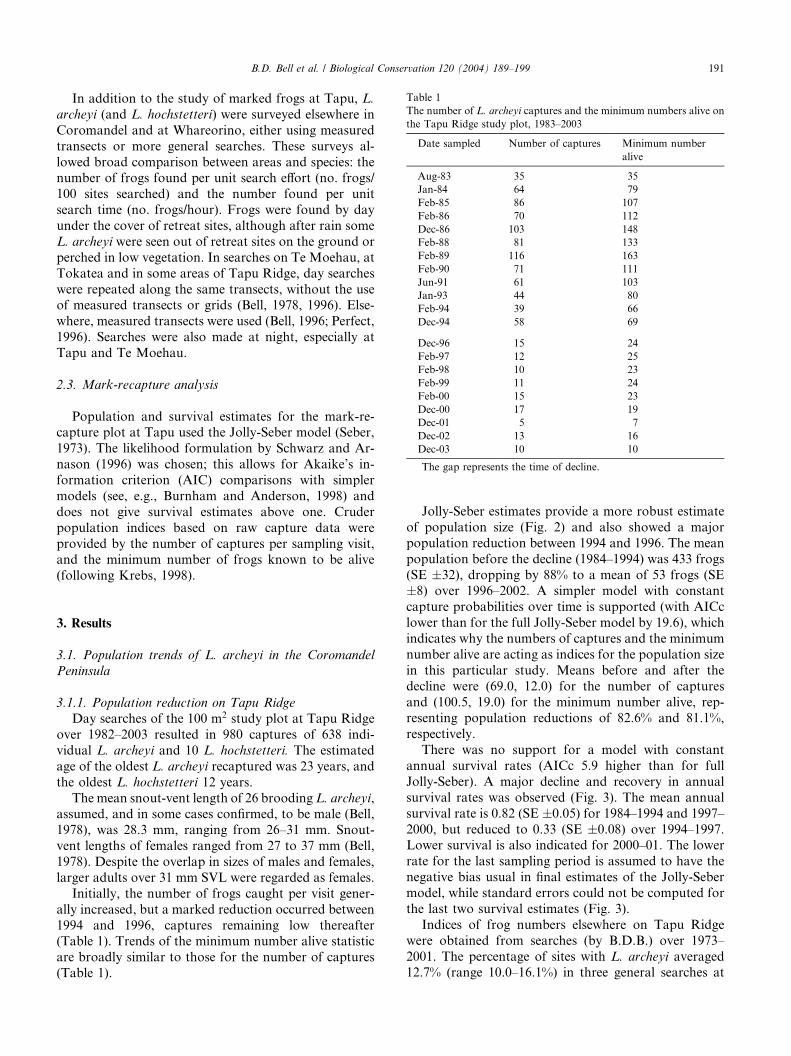
Table 1
The number of L. archeyi captures and the minimum numbers alive on
the Tapu Ridge study plot, 1983–2003
Date sampled Number of captures Minimum number
alive
Aug-83 35 35
Jan-84 64 79
Feb-85 86 107
Feb-86 70 112
Dec-86 103 148
Feb-88 81 133
Feb-89 116 163
Feb-90 71 111
Jun-91 61 103
Jan-93 44 80
Feb-94 39 66
Dec-94 58 69
Dec-96 15 24
Feb-97 12 25
Feb-98 10 23
Feb-99 11 24
Feb-00 15 23
Dec-00 17 19
Dec-01 5 7
Dec-02 13 16
Dec-03 10 10
The gap represents the time of decline.
B.D. Bell et al. / Biological Conservation 120 (2004) 189–199 191
In addition to the study of marked frogs at Tapu, L.
archeyi (and L. hochstetteri) were surveyed elsewhere in
Coromandel and at Whareorino, either using measured
transects or more general searches. These surveys al-
lowed broad comparison between areas and species: thenumber of frogs found per unit search effort (no. frogs/
100 sites searched) and the number found per unit
search time (no. frogs/hour). Frogs were found by day
under the cover of retreat sites, although after rain some
L. archeyi were seen out of retreat sites on the ground or
perched in low vegetation. In searches on Te Moehau, at
Tokatea and in some areas of Tapu Ridge, day searches
were repeated along the same transects, without the useof measured transects or grids (Bell, 1978, 1996). Else-
where, measured transects were used (Bell, 1996; Perfect,
1996). Searches were also made at night, especially at
Tapu and Te Moehau.
2.3. Mark-recapture analysis
Population and survival estimates for the mark-re-capture plot at Tapu used the Jolly-Seber model (Seber,
1973). The likelihood formulation by Schwarz and Ar-
nason (1996) was chosen; this allows for Akaike’s in-
formation criterion (AIC) comparisons with simpler
models (see, e.g., Burnham and Anderson, 1998) and
does not give survival estimates above one. Cruder
population indices based on raw capture data were
provided by the number of captures per sampling visit,and the minimum number of frogs known to be alive
(following Krebs, 1998).
3. Results
3.1. Population trends of L. archeyi in the Coromandel
Peninsula
3.1.1. Population reduction on Tapu Ridge
Day searches of the 100 m2 study plot at Tapu Ridge
over 1982–2003 resulted in 980 captures of 638 indi-
vidual L. archeyi and 10 L. hochstetteri. The estimated
age of the oldest L. archeyi recaptured was 23 years, and
the oldest L. hochstetteri 12 years.
The mean snout-vent length of 26 brooding L. archeyi,assumed, and in some cases confirmed, to be male (Bell,
1978), was 28.3 mm, ranging from 26–31 mm. Snout-
vent lengths of females ranged from 27 to 37 mm (Bell,
1978). Despite the overlap in sizes of males and females,
larger adults over 31 mm SVL were regarded as females.
Initially, the number of frogs caught per visit gener-
ally increased, but a marked reduction occurred between
1994 and 1996, captures remaining low thereafter(Table 1). Trends of the minimum number alive statistic
are broadly similar to those for the number of captures
(Table 1).
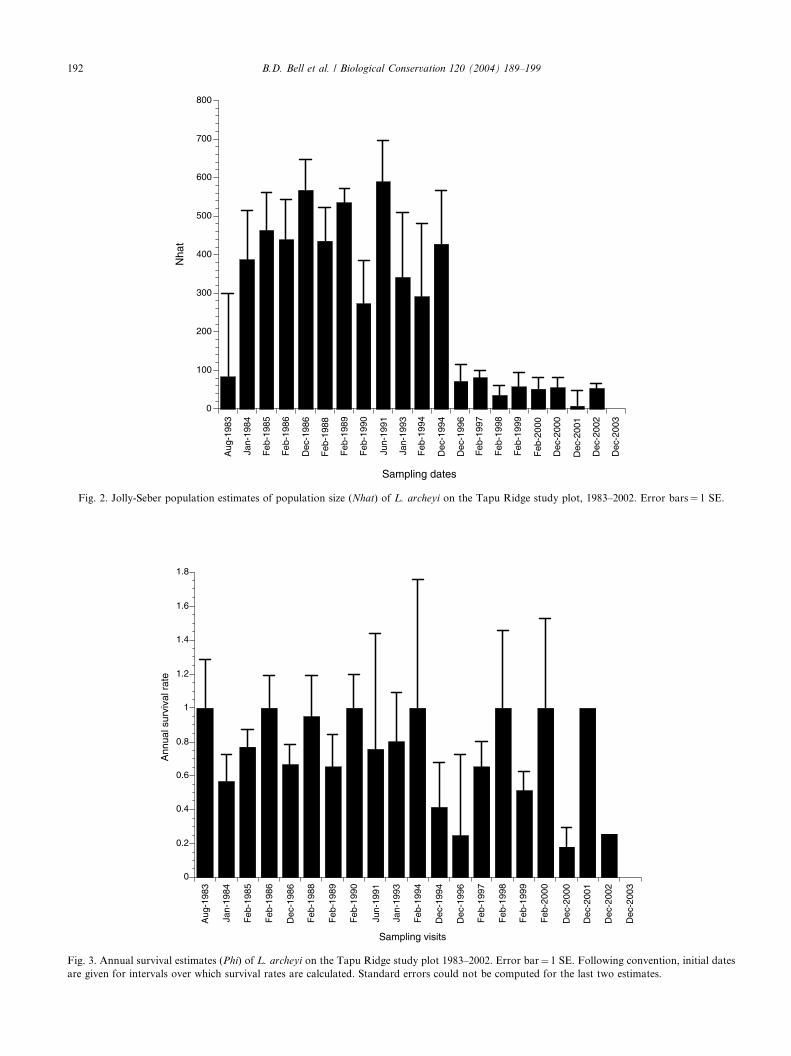
Jolly-Seber estimates provide a more robust estimate
of population size (Fig. 2) and also showed a major
population reduction between 1994 and 1996. The mean
population before the decline (1984–1994) was 433 frogs
(SE �32), dropping by 88% to a mean of 53 frogs (SE�8) over 1996–2002. A simpler model with constant
capture probabilities over time is supported (with AICc
lower than for the full Jolly-Seber model by 19.6), which
indicates why the numbers of captures and the minimum
number alive are acting as indices for the population size
in this particular study. Means before and after the
decline were (69.0, 12.0) for the number of captures
and (100.5, 19.0) for the minimum number alive, rep-resenting population reductions of 82.6% and 81.1%,
respectively.
There was no support for a model with constant
annual survival rates (AICc 5.9 higher than for full
Jolly-Seber). A major decline and recovery in annual
survival rates was observed (Fig. 3). The mean annual
survival rate is 0.82 (SE �0.05) for 1984–1994 and 1997–
2000, but reduced to 0.33 (SE �0.08) over 1994–1997.Lower survival is also indicated for 2000–01. The lower
rate for the last sampling period is assumed to have the
negative bias usual in final estimates of the Jolly-Seber
model, while standard errors could not be computed for
the last two survival estimates (Fig. 3).
Indices of frog numbers elsewhere on Tapu Ridge
were obtained from searches (by B.D.B.) over 1973–
2001. The percentage of sites with L. archeyi averaged12.7% (range 10.0–16.1%) in three general searches at
Aug
-198
3
Jan-
1984
Feb
-198
5
Feb
-198
6
Dec
-198
6
Feb
-198
8
Feb
-198
9
Feb
-199
0
Jun-
1991
Jan-
1993
Feb
-199
4
Dec
-199
4
Dec
-199
6
Feb
-199
7
Feb
-199
8
Feb
-199
9
Feb
-200
0
Dec
-200
0
Dec
-200
1
Dec
-200
2
Dec
-200
3
0
100
200
300
400
500
600
700
800
Nha
t
Sampling dates
Fig. 2. Jolly-Seber population estimates of population size (Nhat) of L. archeyi on the Tapu Ridge study plot, 1983–2002. Error bars¼ 1 SE.
Aug
-198
3
Jan-
1984
Feb
-198
5
Feb
-198
6
Dec
-198
6
Feb
-198
8
Feb
-198
9
Feb
-199
0
Jun-
1991
Jan-
1993
Feb
-199
4
Dec
-199
4
Dec
-199
6
Feb
-199
7
Feb
-199
8
Feb
-199
9
Feb
-200
0
Dec
-200
0
Dec
-200
1
Dec
-200
2
Dec
-200
3
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
Ann
ual s
urvi
val r
ate
Sampling visits
Fig. 3. Annual survival estimates (Phi) of L. archeyi on the Tapu Ridge study plot 1983–2002. Error bar¼ 1 SE. Following convention, initial dates
are given for intervals over which survival rates are calculated. Standard errors could not be computed for the last two estimates.
192 B.D. Bell et al. / Biological Conservation 120 (2004) 189–199

Table 2
Comparison of 1983 and 1998 transect searches for L. archeyi in a
relatively undisturbed area of native forest on Tapu Ridge
Dec-1983 Feb-1998 % Decline
No. L. archeyi found 25 6 76%
No. L. archeyi/h 26.3 6.2 76%
No. L. archeyi/100 sites 9.2 2.0 78%
B.D. Bell et al. / Biological Conservation 120 (2004) 189–199 193
Tapu over 1973–83, but only 1.7% (range 1.5–1.8%) in
two searches over 1998–2001 – a significant population
reduction of 87.0% (generalised linear model with bi-
nomial responses, v2 ¼ 79:6751, p < 0:001, df¼ 1).
Searches along a 25�2 m transect in December 1983 and
February 1998, in an otherwise undisturbed area adja-
cent to the mark-recapture plot, revealed that a signifi-
cant reduction of 76–78% had occurred (v2 ¼ 14:163,p < 0:001, df¼ 1; Table 2).
The mean number of L. archeyi found by Perfect
(1996) on 10 monthly transect counts on Tapu Ridge
over January–October 1995 was 87.4 frogs (SE 8.66,
range 42–123), but a repeat count in February 1997 re-
vealed only 15 frogs – a mean reduction of 83%.
3.1.2. Population reduction on Tokatea Ridge
While L. archeyi had become scarce on Tapu Ridge
by December 1996, an equivalent decline was not evi-
dent on Tokatea Ridge in February 1997, but numbers
had crashed by November 1998 (Table 3). More general
searches for L. archeyi at Tokatea confirm the decline
registered on the main transect.
There is evidence to suggest that L. archeyi in the
central Coromandel ranges between Tapu and Tokateaare also scarcer than formerly. In a two-hour search in
1993, R. Thorpe (pers. commun.) found six frogs at
Manaia, but an eight hour search in the same area in
1999 revealed no frogs.
3.1.3. Population reduction on Te Moehau
In suitable areas on Te Moehau L. archeyi was for-
merly abundant (Stephenson and Stephenson, 1957; Bell,1978). Search indices over the period September 1973 to
October 2001 show that a marked decline occurred there
Table 3
Leiopelmatid frogs located along the transect on Tokatea Ridge 1995–2003.
Nov-1995 Feb-1997
L. archeyi/100 sites 2.77 2.34
L. hochstetteri/100 sites 0.48 0.88
L. archeyi/h 7.11 6.23
L. hochstetteri/h 1.24 2.34
Sites searched 831 684
Search time (min) 194 154
No. L. archeyi 23 16
No. L. hochstetteri 4 6
also, but was not evident until 2001. L. archeyi was
readily located by Ongohi hut in April 2001, but by July
the species was extremely difficult to find, as well as
subsequently (Fig. 4). One of two dead L. archeyi we
found near Ongohi hut in July provided the first case ofchytridiomycosis in Leiopelma. Only four live L. archeyi
were found there in both July and October 2001, while no
L. archeyi were found in a three hour search near Te
Hope hut in October 2001, despite favourably wet
weather.
3.2. Change of size distribution of L. archeyi after the
decline
Comparison of size distributions of L. archeyi before
(1991–1994) and after (1996–2003) the decline on Tapu
Ridge shows that proportionately more large frogs (>30
mm snout-vent length) were caught after the decline
(Mann–Whitney U test, z ¼ �2:7775, p ¼ 0:003, n ¼13). These would be mostly females (Bell, 1978, 1994).
Not only did the frogs’ size distribution change atTapu after the decline (Fig. 5), but their relative condi-
tion (weight for given length) was significantly greater.
The mean condition index (100� (log weight/log snout-
vent length)) of frogs greater than 20 mm snout-vent
length was lower (24.7) in 1993–1994, before the decline,
than afterwards (32.8; Mann–Whitney U test, p < 0:001,n ¼ 237). This difference in part reflects the greater
proportion of larger, gravid females captured after thedecline, but may also indicate a better overall condition
of frogs at lower density, as found in L. pakeka fol-
lowing translocation on Maud Island (Bell et al., 2004).
To check this, weights and lengths of larger frogs with
snout-vent lengths greater than 31.0 mm (females) were
compared. Their condition indices before (33.6) and
after (38.3) the decline were still significantly different
(Mann–Whitney U test, p < 0:001, n ¼ 94), suggestingthat surviving female frogs of similar size were indeed in
better condition. Several juveniles (<20 mm snout-vent
length) were caught after the decline, providing evidence
of continued breeding (also a male was found on an egg
cluster 5 m from the plot in December 2001). Most
surviving L. archeyi found at Tokatea after the decline
There was one observer over 1995–1998, two observers over 2000–03
Nov-1998 Dec-2000 Dec-2003
0 0 0.26
0.75 1.01 0.61
0 0 0.63
1.92 3.73 1.47
665 2087 1155
156 338 285
0 0 3
5 21 7

Fig. 5. L. archeyi size distribution on the Tapu Ridge study plot before (1991–1994) and after (1996–2003) the population decline.
Fig. 4. Capture rate of L. archeyi on Te Moehau (frogs h�1) over 1973–2001. BDB, surveys by B.D. Bell (west slopes and below summit); DOC,
surveys by Department of Conservation on east slopes at 500–600 m; VUW, surveys by Victoria University of Wellington (N.J.M., S.C.) near Ongohi
and Te Hope huts.
194 B.D. Bell et al. / Biological Conservation 120 (2004) 189–199
(n ¼ 6) also were larger frogs in the female size range
(mean snout-vent 33.3 mm, range 31–35 mm), as at
Tapu (Fig. 5). A young L. archeyi found at Tokatea in
December 2003 (snout-vent length 17.1 mm) provides
evidence of continued breeding there.
3.3. Population trends in L. hochstetteri
Unlike L. archeyi, no major declines of L. hochstetteri
are evident. The mean number of L hochstetteri found
by Perfect (1996) on 10 monthly transect counts on

B.D. Bell et al. / Biological Conservation 120 (2004) 189–199 195
Tapu Ridge over January–October 1995 was 39.2 frogs
(SE 2.98, range 30–55), but a repeat count in February
1997 revealed 22 frogs, which was below the minimum
count for 1995. Along the Tokatea Ridge transect L.
hochstetteri remained common (Table 3). Five counts byB.D.B. along the upper reaches of Driving Creek, west
of Tokatea Ridge, revealed varying numbers of L.
hochstetteri over thirty years (1972–2002), successive
search indices (frogs h�1) being: 12.0 (February 1972);
5.6 (December 1972); 12.5 (September 1973); 7.8 (No-
vember 1995); 6.7 (February 2002). L. hochstetteri re-
mained common during the period of marked L. archeyi
decline on Te Moehau in 2001, search indices being 3.6and 4.6 frogs h�1 at Ongohi Hut in July and October
2001, and 1.9 frogs h�1 at Te Hope Hut in October 2001.
In conclusion, from available survey data there is no
evidence of a decline of L. hochstetteri in Coromandel as
seen in L. archeyi, although more monitoring of the
species is needed to provide data equivalent to those
available for L. archeyi.
4. Discussion
Since human settlement, all extant species of Leiop-
elma have suffered range reductions in New Zealand,
with three species going extinct (Worthy, 1987a; Bell,
1994). It is recognised that these past declines or ex-
tinctions are likely to have resulted particularly from theimpact of introduced mammals, and also from habitat
loss (Bell, 1985a, 1994; Newman, 1996). On the other
hand, faunal surveys over recent decades resulted in
extensions of known ranges of both L. archeyi and L.
hochstetteri in the North Island, including the discovery
of both species in Whareorino forest in the northern
King Country (Bell, 1994; Thurley and Bell, 1994;
Newman, 1996). It should be noted, however, that theirranges were far more extensive in the Late Holocene
(Worthy, 1987b). In 1996, at a time of increasing
worldwide concern for declining amphibian populations
(Vial and Saylor, 1993; Carey and Bryant, 1995; Lau-
rance et al., 1996), it was seen as important for trends in
New Zealand’s amphibian populations to be adequately
monitored for early identification of potential problems
(Bell, 1996; Newman, 1996). Previously, there had beenonly anecdotal reports of recent frog declines in New
Zealand, and these were of introduced Litoria species
(Bishop, 1999).
4.1. Value of long-term monitoring
In order to elucidate real declines from stochastic
fluctuations, Gardner (2001) notes that a long time se-ries is highly desirable, although few studies are longer
than five years, and even fewer are more than 10 (Alford
and Richards, 1999; Houlahan et al., 2000; Marsh, 2001;
Young et al., 2001). Populations of terrestrial-breeding
amphibians fluctuate less than their aquatic-breeding
counterparts (Marsh, 2001), thus our Tapu dataset
spanning two decades provides unequivocal evidence of
a population decline. Had this study not been carriedout, the decline may well have been overlooked, or at
least not identified for some time. The decline was so
substantial that less sensitive search indices also revealed
it.
4.2. Progressive northward decline in Coromandel
The decline of L. archeyi at Tapu Ridge is placedbetween December 1994 and December 1996 from
mark-recapture data (Table 1, Fig. 2). Additional evi-
dence, however, suggests that L. archeyi remained
abundant through 1995: substantial numbers (n ¼ 91)
were located on 1080-monitoring transects as late as
October 1995 (Perfect, 1996). Even by mid-1996 the
species was still comparatively common, for members of
the Native Frog Recovery Group had no difficulty infinding frogs then (D.G. Newman, pers. commun.). By
mid-November 1996, however, only one frog was found
after well over 100 sites were searched (K. Corbett, pers.
commun.). That scarcity was confirmed on the study
plot the following month (Table 1), while a more general
search in early January 1997 again revealed very few
frogs (A. Styche, pers. commun.).
Although dead L. archeyi were found at Tokatea in1995, the major decline there occurred between Febru-
ary 1997 and November 1998 (Table 3). On Te Moehau,
the onset of the decline in the Department of Conser-
vation survey area at 500–600 m altitude appears to
have been by January 2001, although by July numbers
had decreased further (Fig. 4). While the years of decline
of L. archeyi are only estimated reliably for Tapu,
Tokatea and Te Moehau, they indicate that the declineprogressed northwards at an average rate of about 12
km per year – or 1 km per month.
4.3. What caused the decline?
Numerous factors may have contributed to declines
of L. archeyi on the Coromandel Peninsula, including
(1) human disturbance, (2) habitat loss, (3) chemicalssuch as biocides, (4) mammalian predation, (5) climatic
factors, and (6) disease. Each of these potential factors is
discussed below in light of the available evidence. We do
not consider effects of potential increases in ambient
ultraviolet-b (UV-b) radiation (cf. Adams et al., 2001),
as L. archeyi eggs are brooded in cryptic locations and
adults spend daylight hours in retreats.
4.3.1. Human disturbance
In December 1996, when a decline had only been
recorded on the 100 m2 study plot on Tapu Ridge, a

196 B.D. Bell et al. / Biological Conservation 120 (2004) 189–199
local disturbance event was a possibility. Conceivably
frogs might have been deliberately and illegally collected
from the site. Indeed, many rocks had been disturbed
there early in 1997, presumably in a search for frogs, but
not before the decline was first noted in December. Ourannual day searches at the site would also have had
some impact, since retreat sites can be drier as a result of
lifting rocks (Bell, 1996). It is very unlikely, however,
that such activity would cause the sudden and major
decline in 1996. Further, we are aware that frogs are
likely to have experienced initial handling stress through
our individual marking regime. However, survival rates
were relatively high over most of the study period (whenmost frogs were marked), showing that marking had
minimal impact on future survival, and was not a factor
in their sudden decline. Moreover, surveys elsewhere in
the Tapu area in 1997 and 1998 confirmed that the de-
cline was widespread, so human disturbance can be
discounted as a major agent of recent decline.
4.3.2. Habitat loss
More general habitat loss within the range of L. ar-
cheyi in Coromandel did not occur to any extent during
the period of decline. Indeed, the species has been re-
silient to major habitat changes in the area since Euro-
pean settlement, such as logging and gold mining
operations, although such activity is likely to deplete
numbers (Bell, 1985a). Habitat loss can therefore also be
discounted as a major agent of recent decline.
4.3.3. Chemicals
There is no evidence that chemicals, such as pollu-
tants or agrochemicals, had an impact on L. archeyi.
However, the decline at Tapu Ridge did occur the year
after a sodium monofluoroacetate (1080) poison drop,
so it is important to assess whether 1080 was a factor in
the decline (the study plot was within the 1080 dropzone). Fortunately, Tapu Ridge was the focus of a
study to specifically monitor the short-term impact of
1080 on native frogs in 1995, so we have information
on its likely effect. Substantial numbers (n ¼ 91) of L.
archeyi were still found on survey transects in October
1995, four months after the 1080 operation (Perfect,
1996). A re-survey of the transect lines in February
1997 showed that the decline was not confined to areaswhere 1080 had been used, expected if the toxin had an
effect, but also occurred in a control area where 1080
had not been used. This provided evidence that 1080
was not a direct agent of recent decline. The decline at
Tokatea is further evidence, as it was again in an area
in which 1080 had not been used. The literature sug-
gests that L. archeyi may not be very susceptible to
1080, as high LD50 doses have been calculated for arange of amphibians, suggesting that they are highly
resistant (Hudson et al., 1984; McIlroy et al., 1985;
Perfect, 1996).
4.3.4. Mammalian predation
Coromandel Leiopelma populations have coexisted
with introduced mammalian predators for many de-
cades, and no novel mammalian predators entered the
study areas during the study period. Unlike L. archeyi atWhareorino forest (Thurley and Bell, 1994; T. Thurley,
pers. commun.), there has been no direct evidence of rat
predation on Tapu Ridge. Larger frogs might be ex-
pected to be at greater risk to rat predation than smaller
frogs, as they may emerge more often and may be less
able to occupy retreat sites inaccessible to predators: the
rat-predated L. archeyi found at Whareorino in 1991–
1993 were all adults (Thurley and Bell, 1994). Survivorsfound at Tapu and Tokatea tended to be larger indi-
viduals, however, suggesting that predation was not a
prime agent of decline there. Side effects of 1080 poi-
soning, if any, are hard to predict. The 1080 operation
was aimed at reducing numbers of introduced brushtail
possums Trichosurus vulpecula, but it would also have
reduced numbers of potential predators (e.g., ship rat
Rattus rattus, stoat Mustela erminea), as well as inver-tebrates (Atkinson et al., 1995). Whether 1080 poisoning
led to prey-switching in surviving predators is not
known. Now that populations of L. archeyi are reduced
in the Coromandel Peninsula, the impact of mammalian
predation on them may become more significant there.
4.3.5. Climate change
While climatic factors may contribute to populationchange and local distribution, weather conditions in the
Coromandel Peninsula over 1996–2001 were not par-
ticularly exceptional (Bishop, 1999). In January 1995, 29
dead L. archeyi were collected by the Department of
Conservation on a 50 m stretch of the summit track at
their type locality on Tokatea Ridge (R. Chappell, pers.
commun.). This followed a thunderstorm, then sun-
shine, after a prolonged drought. The dead frogs weremainly in the adult size range. It is conceivable that
heavy rainfall initiated movement of relatively weakened
and dehydrated individuals to the exposed track, or had
swept them there, and a sudden clearance to sunny
weather might have exposed them to fatally warm con-
ditions. Alternatively, other factors, such as disease,
may have predisposed them to such mortality, but
subsequent surveys confirmed that good numbers of L.archeyi were still to be found in apparently good con-
dition at Tokatea in 1995 and 1997. The co-occurrence
of a disease outbreak and a climatic anomaly remains a
conceivable explanation for this event, but in general
there is little evidence to suggest that climate has been
the main causal factor in declines of L. archeyi in the
Coromandel Peninsula.
4.3.6. Disease
To date the identification of chytrid fungus has pro-
vided the most convincing relationship with the decline

B.D. Bell et al. / Biological Conservation 120 (2004) 189–199 197
of L. archeyi in the Coromandel region. Pathological
chytrid fungus Batrachochytrium dendrobatidis (phylum
Chytridiomycota), described by Longcore et al. (1999),
was first recognised in New Zealand in the introduced
hylid frog Litoria raniformis in Christchurch in 1999–2000 (Waldman et al., 2001). Chytrid fungus was found
in L. archeyi at Te Moehau, Tapu and Whareorino in
2001–02, being first recorded in a dead L. archeyi from
Te Moehau in July 2001. The progressive northward
declines of L. archeyi populations in Coromandel (1996–
2001) would support the notion of a spreading pathogen,
with an average rate of about 12 km per year. If the
causal factor is chytrid fungus, this suggests the pathogenwas in Coromandel by late 1996, when the first decline
occurred at Tapu, even though chytridiomycosis was not
identified in New Zealand until 1999–2000 (Waldman
et al., 2001) and not at Tapu until 2001. Reports across
New Zealand suggest that populations of introduced
frogs (Litoria spp.) crashed over 1993–1995 (Bishop,
1999). In the Coromandel region, declines of Litoria
aurea were reported over 1997–2000 (P. Thomson, inlitt.). While dead L. archeyi infected with chytrid fungus
have been found at Whareorino, no overall decline has
occurred there, and the reasons for this are obscure.
4.3.7. Conclusion – disease the most likely agent of decline
On a global scale, declining amphibian populations
cannot be attributed to a single cause for it appears that
multiple, interacting causes are involved. We have con-sidered various causal agents for the declines of L. ar-
cheyi in New Zealand, and several of these factors may
have interacted to effect those declines. There is
mounting evidence, however, that disease has been the
major agent of decline, either directly, or indirectly
through the frogs becoming susceptible as a result of
other factors (e.g., Pounds, 2001). The disease argument
is supported by (1) the rapidity and severity of declines,(2) the progressive (south to north) nature of the out-
breaks, and (3) evidence of chytriodiomycosis infection
in dead or sick frogs at the time of decline.
In Australia, montane creek-dwelling species tend to
have been most impacted by recent declines (Laurance
et al., 1996; Berger et al., 1998), so we might have ex-
pected the semi-aquatic L. hochstetteri to be more af-
fected in New Zealand. This was not the case - insteadthe terrestrial L. archeyi declined and was found to have
chytridiomycosis. While chytrid fungus is associated
with the frog’s decline, it is not necessarily the main
causal agent. We do not yet know enough of the pa-
thology and epidemiology of this or other possible am-
phibian pathogens, nor of other factors that may have
predisposed frogs to infection. Further, the mechanism
of spread is uncertain, and those studying these frogs arewell aware of the need to follow meticulous hygiene
protocols, to prevent our spreading the pathogen (Bell,
2002).
4.4. Future scenarios for Leiopelma species
Leiopelma archeyi had sufficient resilience to survive
severe habitat disturbance in Coromandel in the past,
including gold mining, kauri logging, deforestation andintroduced mammalian predators. However, it is a K-selected species with low clutch size, slow maturity and
long lifespan. Theoretical population models (South-
wood et al., 1974) indicate that there is a low population
threshold for K-selected species below which extinction
is likely, and a decline of around 80–90% could exceed
such a threshold.
Several scenarios for the future of Leiopelma speciesin New Zealand can be envisaged. The most optimistic
scenario is that L. archeyi will persist and slowly recover
from its decline and that other Leiopelma species will not
be seriously affected, nor the Whareorino L. archeyi
population. A pessimistic scenario is that L. archeyi will
not recover and is edging towards extinction, and that
the closely related L. hamiltoni and L. pakeka may also
be at high risk. An intermediate scenario is that popu-lations of L. archeyi will not recover to their former
state, having become locally extinct in parts of their
range, but will survive in lower numbers elsewhere, with
other Leiopelma species not declining.
As we understand so little of the pathology and epi-
demiology of any disease that might be affecting L. ar-
cheyi, it is hard to make definite recommendations for
management to ensure protection against disease, al-though prudent preventative measures and contingen-
cies need to be further developed. New Zealand has
justifiably earned international respect for its success in
conservation management of threatened endemic fauna
such as endemic birds, where agents of decline can often
be more readily tackled (Bell and Merton, 2002), but the
appearance of a contagious disease in a native frog
brings new and difficult challenges. Even if populationsof L. archeyi persist, increased conservation manage-
ment is now needed, such as habitat protection and
predator control, supplemented with captive manage-
ment. It is to be hoped that this archaic and attractive
frog will survive despite the major population collapse it
has recently experienced in its former Coromandel
stronghold.
Acknowledgements
Kim McConkey kindly offered constructive com-
ments on a draft of this paper. We thank all those who
have assisted in the frog studies reported here, especially
Ann Bell, Paul Bell, Oliver Berry, Roman Biek, Phil
Bishop, Gill Brackenbury, Finn Buchanan, Rob Chap-pell, Mike Cogswell, Keith Corbett, Alison Cree, Kelly
Hare, Kim McConkey, Leigh Marshall, Don Newman,
Richard Norman, Alison Perfect, Chandra Ramarao,

198 B.D. Bell et al. / Biological Conservation 120 (2004) 189–199
Chris Smuts-Kennedy, Andrew Styche, Rick Thorpe,
Tertia Thurley, Bruce Waldman, and Nadia Webster.
As Leiopelma species are protected under the New
Zealand Wildlife Act 1953, the study was undertaken
under New Zealand Government permits, initially fromthe Wildlife Service and more recently from the De-
partment of Conservation, while the Victoria University
Animal Ethics Committee gave additional approval for
the study. We are indebted to the Department of Con-
servation Native Frog Recovery group for its interest
and support, and to Victoria University of Wellington
and the Department of Conservation for financial
support.
References
Adams, M.J., Schindler, D.E., Bury, R.B., 2001. Association of
amphibians with attenuation of ultraviolet-b radiation in montane
ponds. Oecologia 128, 519–525.
Alford, R.A., Dixon, P.M., Pechmann, J.H.K., 2001. Global amphib-
ian population declines. Nature 412, 498–500.
Alford, R.A., Richards, S.J., 1999. Global amphibian declines: a
problem in applied ecology. Annual Review of Ecology and
Systematics 30, 133–165.
Altig, R., Johnston, G.F., 1989. Guilds of anuran larvae: relationships
among developmental modes, morphologies, and habitats. Herpe-
tological Monographs 3, 81–109.
Archey, G., 1922. The habit and life history of Leiopelma hochstetteri.
Records of the Canterbury (N.Z.) Museum 2, 59.
Atkinson, I.A.E., Campbell, D.J., Fitzgerald, B.M., Flux, J.E.C.,
Meads, M.J., 1995. Possums and possum control; effects on
lowland forest ecosystems. Science for Conservation, vol. 1.
Department of Conservation, Wellington.
Bell, B.D., 1978. Observations on the ecology and reproduction of the
New Zealand native frogs. Herpetologica 34, 340–354.
Bell, B.D., 1985a. Conservation status of the endemic New Zealand
frogs. In: Grigg, G., Shine, R., Ehmann, H. (Eds.), The Biology of
Australasian Frogs and Reptiles. Surrey Beatty & Sons, Chipping
Norton, NSW, pp. 449–458.
Bell, B.D., 1985b. Development and parental care in the endemic New
Zealand frogs. In: Grigg, G., Shine, R., Ehmann, H. (Eds.), The
Biology of Australasian Frogs and Reptiles. Surrey Beatty & Sons,
Chipping Norton, NSW, pp. 269–278.
Bell, B.D., 1994. A review of the status of New Zealand Leiopelma
species (Anura: Leiopelmatidae), including a summary of demo-
graphic studies in Coromandel and on Maud Island. New Zealand
Journal of Zoology 21, 341–349.
Bell, B.D., 1996. Aspects of the ecological management of New
Zealand frogs: conservation status, location, identification,
examination and survey techniques. Ecological Management 4,
91–111.
Bell, B.D., 2002. Frog-friendly islands. New Scientist 17 (2342), 57.
Bell, B.D., Daugherty, C.H., Hay, J.M., 1998a. Leiopelma pakeka, n.
sp. (Anura: Leiopelmatidae), a cryptic species of frog from Maud
Island, New Zealand, and a reassessment of the conservation status
of Leiopelma hamiltoni from Stephens Island. Journal of the Royal
Society of New Zealand 28, 39–54.
Bell, B.D., Daugherty, C.H., Hitchmough, R.A., 1998b. The
taxonomic identity of a population of terrestrial Leiopelma
(Anura: Leiopelmatidae) recently discovered in the northern King
Country, New Zealand. New Zealand Journal of Zoology 25, 139–
146.
Bell, B.D., Merton, D.V., 2002. Critically endangered bird populations
and their management. In: Norris, K., Pain, D. (Eds.), Conserving
Bird Biodiversity: General Principles and their Application. Cam-
bridge University Press, Cambridge, pp. 105–138.
Bell, B.D., Pledger, S., Dewhurst, P.L., 2004. The fate of a population
of the endemic frog Leiopelma pakeka (Anura: Leiopelmatidae)
translocated to restored habitat on Maud Island, New Zealand.
New Zealand Journal of Zoology 31 (2), (in press).
Bell, B.D., Wassersug, R.J., 2003. Anatomical features of Leiopelma
embryos and larvae: implications for anuran evolution. Journal of
Morphology 256, 160–170.
Berger, L., Speare, R., Daszak, P., Green, D.E., Cunningham, A.A.,
Goggin, C.L., Slocombe, R., Ragan, M.A., Hyatt, A.D., McDon-
ald, K.R., Hines, H.B., Lips, K.R., Marantelli, G., Parkes, H.,
1998. Chytridiomycosis causes amphibian mortality associated
with population declines in the rain forests of Australia and Central
America. Proceedings of the National Academy of Sciences 95 (15),
9031–9036.
Bishop, P.J., 1999. Declining frog populations in New Zealand – the
New Zealand Frog Survey and possible future directions. New
Zealand Journal of Zoology 26, 255–256.
Burnham, K.P., Anderson, D.R., 1998. Model Selection and Inference:
A Practical Information-Theoretic Approach. Springer, New York.
Carey, C., Bryant, C.J., 1995. Possible interrelationships among
environmental toxicants, amphibian development, and decline of
amphibian populations. Environmental Health Perspectives 103
(Suppl. 4), 13–17.
Gardner, T., 2001. Declining amphibian populations: a global
phenomenon in conservation biology. Animal Biodiversity and
Conservation 24 (2), 25–44.
Halliday, T., 2001. The wider implications of amphibian population
declines. Oryx 35, 181–182.
Hitchmough, R. (comp.), 2002. New Zealand threat classification
system lists – 2002. Threatened species occasional publication, vol.
23, Department of Conservation, Wellington, 210 p.
Houlahan, J.E., Findlay, C.S., Schidt, B.R., Meyer, A.H., Kuzmin,
S.L., 2000. Quantitative evidence for global amphibian population
declines. Nature 404, 752–755.
Hudson, R.H., Tucker, R.K., Haegele, M.A., 1984. Handbook of
toxicity of pesticides to wildlife (second ed). United States Fish and
Wildlife Service Resource Publication No. 153. United States
Department of the Interior, Washington DC.
King, C., 1984. Immigrant Killers: Introduced Predators and the
Conservation of Birds in New Zealand. Oxford University Press,
Auckland.
Krebs, C.J., 1998. Ecological Methodology, second edition. Addison-
Wesley, Longman, Menlo Park, CA.
Laurance, W.F., McDonald, K.R., Speare, R., 1996. Epidemic disease
and the catastrophic decline of Australian rain forest frogs.
Conservation Biology 10, 406–413.
Longcore, J.E., Pessier, A.P., Nichols, D.K., 1999. Batrachochytrium
dendrobatidis gen. Et ap. Nov., a chytrid pathogenic to amphibians.
Mycologia 91 (2), 219–227.
McIlroy, J.C., King, D.R., Oliver, A.J., 1985. The sensitivity of
Australian animals to 1080 poison. VIII. Amphibians and reptiles.
Australian Wildlife Research 12, 113–118.
Marsh, D.M., 2001. Fluctuations in amphibian populations: a meta-
analysis. Biological Conservation 101, 327–335.
Newman, D.G., 1996. Native Frog (Leiopelma spp.) Recovery Plan.
Threatened Species Recovery Plan No. 18, Department of Con-
servation, Wellington, 40 pp.
Perfect, A.J., 1996. Aspects of the ecology of the native frogs
Leiopelma archeyi and L. hochstetteri, and the impact of compound
1080. Unpublished M.Sc. thesis, Victoria University of Wellington,
New Zealand.
Pounds, J.A., 2001. Climate and amphibian declines. Nature 410, 639–
640.

B.D. Bell et al. / Biological Conservation 120 (2004) 189–199 199
Schwarz, C.J., Arnason, A.N., 1996. A general methodology for the
analysis of capture–recapture experiments in open populations.
Biometrics 52, 860–873.
Seber, G.A.F., 1973. The Estimation of Animal Abundance and
Related Parameters. Griffin, London.
Southwood, T.R.E., May, R.M., Hassell, M.P., Conway, G.R., 1974.
Ecological strategies and population parameters. American Natu-
ralist 108, 791–804.
Stephenson, E.M., Stephenson, N.G., 1957. Field observations on the
New Zealand frog Leiopelma Fitzinger. Transactions of the Royal
Society of New Zealand 84, 867–882.
Thurley, T., Bell, B.D., 1994. Habitat distribution and predation on a
western population of terrestrial Leiopelma (Anura: Leiopelmati-
dae) in the northern King Country, New Zealand. New Zealand
Journal of Zoology 21, 431–436.
Vial, J.L., Saylor, L., 1993. The status of amphibian populations: a
compilation and analysis. IUCN/SSC Declining Amphibian Pop-
ulation Task Force Document No. 1, 98 pp.
Wake, D.B., 1991. Declining amphibian populations. Science 253, 860.
Waldman, B., Tocher, M., 1998. Behavioral ecology, genetic diversity
and declining amphibian populations. In: Caro, T.M. (Ed.),
Behavioral Ecology and Conservation Biology. Oxford University
Press, Oxford.
Waldman, B., van de Wolfshaar, K.E., Klena, J.D., Andjic, V.,
Bishop, P.J., Norman, R.J. de B., 2001. Chytridiomycosis in New
Zealand frogs. Surveillance 28 (3), 9–11.
Worthy, T.H., 1987a. Osteology of Leiopelma (Amphibia, Leiopel-
matidae) and descriptions of three new subfossil Leiopelma
species. Journal of the Royal Society of New Zealand 17, 201–
251.
Worthy, T.H., 1987b. Palaeoecological information concerning mem-
bers of the frog genus Leiopelma: Leiopelmatidae in New Zealand.
Journal of the Royal Society of New Zealand 17, 409–420.
Young, B.E., Lips, K.R., Reaser, J.K., Ibanez, R., Salas, A.W.,
Cedeno, J.R., Coloma, L.A., Santiago, R., La Marca, E., Meyer,
J.R., Munoz, A., Bolanos, F., Chaves, G., Romo, D., 2001.
Population declines and priorities for amphibian conservation in
Latin America. Conservation Biology 15, 1213–1223.




















