
The recalcitrant plant species, Castanospermum australe and ...
10
Journal of Experimental Botany, Vol. 48, No. 314, pp. 1717-1726, September 1997 Journal of Experimental Botany The recalcitrant plant species, Castanospermum australe and Trichilia dregeana, differ in their ability to produce dehydrin-related polypeptides during seed maturation and in response to ABA or water-deficit-related stresses Bin Han 1 , Patricia Berjak 2 , Norman Pammenter 2 , Jill Farrant 3 and Allison R. Kermode 1 ' 4 1 Department of Biological Sciences, Simon Fraser University, Burnaby, BC, V5A 1S6 Canada 2 Department of Biology, University of Natal, Durban 4001, South Africa 3 Department of Botany, University of Cape Town, Cape Town 7700, South Africa Received 12 February 1997; Accepted 15 April 1997 Abstract In constrast to seeds of orthodox species, those of recalcitrant species do not acquire desiccation toler- ance during their development and are shed from the parent plant at high water contents. Dehydrin produc- tion in seeds of recalcitrant species was examined during development and germination, in response to abscisic acid (ABA), and following the imposition of various water-deficit-related stresses, including desic- cation, water stress, high salt, high osmolarity, and low temperature. Two tropical species exhibited a dif- ferential capacity to produce dehydrin-related proteins during seed maturation. Dehydrins were present in axes and cotyledons of Castanospermum australe seeds during mid-maturation and at maturity. In Trichilia dregeana, no dehydrin-related polypeptides were detected in the mature seed. During the develop- ment of C. australe seeds, the nature of the dehydrin- related polypeptides accumulated in the cotyledons and axis changed and new polypeptides were detected in the mature seeds that were not present during mid- maturation. The dehydrins present in cotyledons of mature seeds (31, 37 and 40 kDa) were still detectable after germination (i.e. in untreated seedlings). These dehydrins became less abundant in the cotyledons of C. australe seedlings following ABA and all stress treatments except cold, although most of the dehyd- rins were still detectable. An exception was the desic- cation-treated seedlings, in which no dehydrins were detected. In the roots of C. australe seedlings, no dehydrins were found after germination nor were they induced in the root by ABA or any of the stress treat- ments imposed on seedlings. Seedlings of Trichilia dregeana did not produce dehydrins in the roots or cotyledons when exposed to ABA or water-deficit- related stresses. Key words: Dehydrin, ABA, desiccation, recalcitrant, seed. Introduction The terms 'orthodox' and 'recalcitrant' are used to describe the storage behaviour of seeds. Orthodox seeds are shed from the parent plant at low moisture contents, having undergone maturation drying prior to this event, and can generally be further dried to moisture contents in the range of 1-5% without damage. In this dehydrated state, the seed can resist the vicissitudes of the environment, and unless dormant, will resume full metabolic activity, growth and development when conditions conducive to germina- tion are provided. Because of these properties, such seeds can be stored for long periods. Recalcitrant seeds, on the other hand, do not undergo maturation drying, and are shed at relatively high moisture contents. Such seeds are highly susceptible to desiccation injury, and thus are not storable under conditions suitable for orthodox seeds (reviewed in Vertucci and Farrant, 1995; Smith and Berjak, 1995). Furthermore, the seeds of many recalcitrant species are sensitive to chilling injury at lowered temperatures (Farrant et al., 1988). The cause of desiccation sensitivity of recalcitrant seeds, and indeed the mechanism of desiccation tolerance in 'To whom correspondence should be addressed. Fax: +1 604 291 3496. E-mail: kermode©sfu.ca 6 Oxford University Press 1997 Downloaded from https://academic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 March 2022
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of The recalcitrant plant species, Castanospermum australe and ...

Journal of Experimental Botany, Vol. 48, No. 314, pp. 1717-1726, September 1997Journal ofExperimentalBotany
The recalcitrant plant species, Castanospermum australeand Trichilia dregeana, differ in their ability to producedehydrin-related polypeptides during seed maturationand in response to ABA or water-deficit-related stresses
Bin Han1, Patricia Berjak2, Norman Pammenter2, Jill Farrant3 and Allison R. Kermode1'4
1 Department of Biological Sciences, Simon Fraser University, Burnaby, BC, V5A 1S6 Canada2 Department of Biology, University of Natal, Durban 4001, South Africa3 Department of Botany, University of Cape Town, Cape Town 7700, South Africa
Received 12 February 1997; Accepted 15 April 1997
Abstract
In constrast to seeds of orthodox species, those ofrecalcitrant species do not acquire desiccation toler-ance during their development and are shed from theparent plant at high water contents. Dehydrin produc-tion in seeds of recalcitrant species was examinedduring development and germination, in response toabscisic acid (ABA), and following the imposition ofvarious water-deficit-related stresses, including desic-cation, water stress, high salt, high osmolarity, andlow temperature. Two tropical species exhibited a dif-ferential capacity to produce dehydrin-related proteinsduring seed maturation. Dehydrins were present inaxes and cotyledons of Castanospermum australeseeds during mid-maturation and at maturity. InTrichilia dregeana, no dehydrin-related polypeptideswere detected in the mature seed. During the develop-ment of C. australe seeds, the nature of the dehydrin-related polypeptides accumulated in the cotyledonsand axis changed and new polypeptides were detectedin the mature seeds that were not present during mid-maturation. The dehydrins present in cotyledons ofmature seeds (31, 37 and 40 kDa) were still detectableafter germination (i.e. in untreated seedlings). Thesedehydrins became less abundant in the cotyledons ofC. australe seedlings following ABA and all stresstreatments except cold, although most of the dehyd-rins were still detectable. An exception was the desic-cation-treated seedlings, in which no dehydrins weredetected. In the roots of C. australe seedlings, nodehydrins were found after germination nor were they
induced in the root by ABA or any of the stress treat-ments imposed on seedlings. Seedlings of Trichiliadregeana did not produce dehydrins in the roots orcotyledons when exposed to ABA or water-deficit-related stresses.
Key words: Dehydrin, ABA, desiccation, recalcitrant, seed.
Introduction
The terms 'orthodox' and 'recalcitrant' are used to describethe storage behaviour of seeds. Orthodox seeds are shedfrom the parent plant at low moisture contents, havingundergone maturation drying prior to this event, and cangenerally be further dried to moisture contents in the rangeof 1-5% without damage. In this dehydrated state, theseed can resist the vicissitudes of the environment, andunless dormant, will resume full metabolic activity, growthand development when conditions conducive to germina-tion are provided. Because of these properties, such seedscan be stored for long periods. Recalcitrant seeds, on theother hand, do not undergo maturation drying, and areshed at relatively high moisture contents. Such seeds arehighly susceptible to desiccation injury, and thus are notstorable under conditions suitable for orthodox seeds(reviewed in Vertucci and Farrant, 1995; Smith and Berjak,1995). Furthermore, the seeds of many recalcitrant speciesare sensitive to chilling injury at lowered temperatures(Farrant et al., 1988).
The cause of desiccation sensitivity of recalcitrant seeds,and indeed the mechanism of desiccation tolerance in
'To whom correspondence should be addressed. Fax: +1 604 291 3496. E-mail: kermode©sfu.ca
6 Oxford University Press 1997
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022

1718 Han et al.
seeds of orthodox species, is not well understood. Inorthodox seeds, a group of proteins is synthesized duringmid- to late-maturation that are presumed to play aprotective role during desiccation. This group of proteins,termed dehydrins (a subset of the proteins termed LEAs;Late Embryogenesis Abundant or RABs; Responsive toABA), have some common features in their structure thatmay be important for their putative protective function.Precocious appearance of the proteins and their mRNAscan be induced in cultured immature embryos by ABAtreatment. Specifically, it has been hypothesized that,during development, high concentrations of ABA inducethe accumulation of these polypeptides and hence preparethe embryo for desiccation or possible cellular disruptionupon subsequent rehydration (reviewed in Bray, 1991,1993; Chandler and Robertson, 1994; Kermode, 1990,1995, 1997; Bewley and Oliver, 1992). The proteins arecharacterized by a conserved 15-amino acid, lysine-richsequence near the carboxyl terminus. The consensus poly-peptide forms an amphiphilic a-helix which may serveamong other functions as an ion trap in dehydrating cells,sequestering ions as they become concentrated (Closeet al., 1993a; Dure, 1993).
Dehydrin genes exhibit a flexible expression repertoire,being responsive to both developmental and environ-mental cues (reviewed in Thomas et al., 1991). In additionto being expressed under abnormal (stress) conditions,they exhibit temporal regulation during seed developmentwhere expression is most intense during mid- to late-development. Hence, the protective role of dehydrins inthe survival of water loss is purported to be dual: duringmaturation drying of the developing seed and followinggermination/growth of the mature seed (i.e. in seedlingsor plant vegetative tissues undergoing mild water stress).
The question arises as to whether the desiccation sensit-ivity of recalcitrant seeds is at least partially the result ofan insufficient accumulation of dehydrins, or whetherother factors (including a lack of protective sugars) aremore important. Other features that may be part of thebasis of desiccation-intolerance include an inability torepair desiccation-induced damage upon subsequent rehy-dration and an inappropriate proportion or distributionof freezable and non-freezable (bound) water within theseed (Berjak et al., 1992; reviewed in Bewley and Oliver,1992; Vertucci and Farrant, 1995). An interesting hypo-thesis is that dehydrins have detergent and chaperone-like properties and may interact with compatible solutesto serve as structural stabilizers of macromolecules underconditions of water deficit (Close, 1996). Dehydrins havebeen reported to be absent during the late stages ofdevelopment of recalcitrant mangrove (Avicennia marina)seeds, a finding suggested to account for the lack ofdesiccation tolerance of this species (Farrant et al., 1992,1996). In grains of Zizania palustris, a seed that exhibitscharacteristics intermediate between recalcitrance and
orthodoxy (Bradford and Chandler, 1992; but see alsoVertucci et al., 1994; Probert and Brierley, 1989), dehyd-rins can be detected by the maize dehydrin antiserum inboth embryos and seedlings (Bradford and Chandler,1992). The Z. palustris embryos and seedlings are alsocapable of ABA accumulation during limited dehydra-tion. The intolerance of Z. palustris seeds to dehydrationat low temperature does not seem to be due to an absenceofdehydrinsoran inability to accumulate ABA (Bradfordand Chandler, 1992). This indicates that the presence ofdehydrins alone is not sufficient to prevent desiccationinjury (Blackman et al., 1991; Bradford and Chandler,1992). Finch-Savage et al. (1994) detected dehydrins inmature seeds of five desiccation sensitive (recalcitrant)trees (all temperate species), and Lea mRNA can also beinduced in stored recalcitrant seeds of Quercus robur L.(English oak) by ABA and limited drying treatments.Since desiccation tolerance is a quantitative feature(Vertucci and Farrant, 1995) the amount of dehydrins/LEAs, or the rate at which the proteins accumulate, maydetermine the level of tolerance. Accumulation of otherprotectants is likely to be required in addition to LEAproteins (Blackman et al., 1992) since interactions incombination may be necessary to stabilize macro-molecules under conditions of water deficit (Close, 1996;Ingram and Bartels, 1996).
Although all recalcitrant seeds are considered to bedesiccation sensitive, the degree of water loss toleratedvaries with the species; hence, dehydrin gene expressionneeds to be investigated in a wider range of recalcitrantseed types. Further, temporal studies to examine changesin dehydrin production during development and followinggermination are necessary. This is especially importantsince there is often no clear-cut event delineating the endof seed development and the start of germination inrecalcitrant species (Vertucci and Farrant, 1995).Moreover, even though seeds of recalcitrant species maybe capable of dehydrin synthesis during their develop-ment, a decline in the synthesis of dehydrins (or aninability to produce specific dehydrins) following thetransition to germination/seedling growth may be animportant factor contributing to losses in viability afterseed shedding.
Using a dehydrin antibody (Close et al., 1993a), dehyd-rin-related proteins were compared in immature andmature seeds of two tropical species that are recalcitrant—Castanospermum australe and Trichilia dregeana. Theresponses of seeds and seedlings to ABA and water-deficit-related stresses were further examined.
Materials and methods
Plant material
Trichilia dregeana seeds were collected at mid-maturation andat maturity from trees growing in Durban (rainfall 1018 mm
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022

per year, mean temperature 20 °C with a mean winter minimumabout 10-12 °C and a mean summer maximum about 30 °C).Seeds of C. australe were collected at mid-maturation and atmaturity from trees growing in Pietermaritzburg, which is hotterin summer and cooler in winter than Durban and has slightlylower rainfall. The average minimum temperature duringfruiting of C. australe (May to June) is approximately 8°C.Axes and cotyledons were obtained from immature seeds atmid-maturation, from mature seeds and from seedlings withradicle lengths of approximately 25 mm (obtained by germinat-ing mature seed as outlined below). Some immature seeds andseedlings of C. australe and T. dregeana were first subjected tothe various treatments outlined below.
Seed germination
Mature seeds were surface-sterilized in a solution of 1%hypochlorite, 0.01% SDS for lOmin, rinsed three times insterilized water, and seed coats removed under aseptic conditionsin a flow hood. Seeds were placed in Petri dishes containingsterile filter paper moistened with 10 ml sterile water andgerminated at 26 °C in the dark until they had acquired radiclelengths of approximately 25 mm. Seedlings were then used forthe various treatments.
ABA treatments
Immature seeds and seedlings were incubated under ambientlaboratory conditions for 7 d in different concentrations ofABA ( 1 0 ~ 3 M - 1 0 ~ 6 M ) i n Petri dishes containing sterile filterpaper and 10 ml ABA (sterilized by passing through a0.22/im filter).
Water stress treatment
Immature seeds and seedlings were subjected to water stress byequilibrating them in a desiccator for 7 d under ambientlaboratory conditions over 25% glycerol (osmotic potential—12.6 MPa), which produced a relative humidity in thechamber of approximately 91%.
Dehydrins in recalcitrant seeds 1719
Water-control treatmentAs a control for the ABA and water-deficit-related stresses,immature seeds and seedlings were placed under ambientlaboratory conditions in Petri dishes containing sterile filterpaper moistened with 10 ml of sterile water for the same lengthof time as the ABA or stress treatments.
Extraction of heat-stable proteins and Western blot analysis
Heat-stable proteins were extracted from cotyledons and axesisolated from immature seeds (at mid-maturation), from matureseeds and from seedlings, as outlined in Close et al. (1993a).Briefly, total soluble proteins were extracted by grindingtissues in 30 mM N-Tris(hydroxymethyl)methyl-2-amino-ethanesulphonic acid (TES) buffer pH 8.0, containing 20 mMNaCl and 1 mM phenylmethylsulphonyl fluoride (PMSF) usinga ground-glass homogenizer. This was followed by centrifu-gation at 14000 rpm in a microcentrifuge for 15 min at 4°C.The supernatants were boiled at 100 °C for 10 min, kept on iceand then centrifuged as before. Protein concentrations weredetermined by the Bio-Rad protein assay (Bradford, 1976)using bovine serum albumin (BioRad, Fraction V) as astandard. Aliquots containing equal amounts of heat stableproteins were precipitated with 4 vols acetone and thenresuspended in SDS-PAGE sample buffer (0.125 M TRIS-HC1,pH6.7, 2% SDS, 10% glycerol, 0.0003% bromophenol blue,and 5% j9-mercaptoethanol). The mixture was heated for 10 minat 95 °C, cooled and then fractionated by SDS-PAGE on 12%mini gels, according to the method of Laemmli (1970). For allWestern blots, the same amount of heat-stable protein (64 ^gin 32 /zl sample buffer) was loaded on to each lane. Afterelectroblotting on to nitrocellulose (using a BioRad Trans-BlotSemi-Dry Transfer Cell), dehydrin proteins were detected withrabbit anti-dehydrin serum (diluted 1/6000 before use), followedby goat anti-rabbit IgG (Promega, Madison, WI) to which wasconjugated alkaline phosphatase. The anti-dehydrin serum wasproduced from a synthetic polypeptide containing a highlyconserved sequence (K1KEKLPG) (kindly provided byTJ Close, UC Riverside) (Close et al., 1993a, b). Thepre-immune serum was used as a control.
Desiccation treatment
To impose a more severe water stress, immature seeds andseedlings were dried slowly by placing them in a desiccator overstirred saturated salt solutions. The drying regime used forimmature seeds and for seedlings was as described in Kermodeand Bewley (1985). This involved placing seeds or seedlings for1 d sequentially in desiccators containing stirred saturated saltsolutions maintaining RH values of 92, 86 (2 d), 74, 65, 44,and 22%.
High salt and high osmolahty treatments
Immature seeds and seedlings were placed for 7 d under ambientlaboratory conditions in Petri dishes containing sterile filterpaper moistened with either 10 ml 0.2 M NaCl (high salttreatment) or 10 ml 0.3 M mannitol (high osmolarity treatment).Both solutions were previously sterilized by passing through a0.22 ^m filter.
Cold treatment
Immature seeds and seedlings were placed in Petri dishescontaining sterile filter paper moistened with 10 ml of sterilewater, and then maintained at 4 °C for 2-7 d.
ResultsDehydrin-related proteins are differentially produced duringseed development in Castanospermum australe andTrichilia dregeana
Western blot analysis shows that dehydrins are present inthe axes and cotyledons of mature and immature seedsof C. australe (Fig. 1, lanes 1-4). Two dehydrin-relatedpolypeptides (40 and 37 kDa) were detected in the matureaxes of C. australe seeds (lane 1); an additional 31 kDapolypeptide was also present in the cotyledons (lane 2).In contrast, no dehydrins were detectable in whole seedsof T. dregeana at maturity (data shown in Fig. 5A, MS).
It is noteworthy that new dehydrins were synthesizedas seeds of C. australe matured. In immature seeds,31 kDa and 85 kDa dehydrins were detected in axes(Fig. 1, lane 3); neither dehydrin was present in matureaxes, in which polypeptides of 37 and 40 kDa accumulated(compare lane 3 with lane 1). A similar change in the
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022

1720
13986
47
3328
20
Han
. 9 -. 8 -
8-
.3 -
6 -
7 -
etal.
•m
1MA
S-40- • • 3 7
— " - 31
2MC
immune
139 9 -86.8 '
47 8 -
33.3 -
28 6-
20 7-
pre-immune
t.
IA IC
^ - "'•— i
85
- 4 0-37-31
i1 2 3
Fig. 1. Western blot analysis of dehydnns present in immature seeds(at mid-maturation) and mature seeds of the tropical recalcitrantspecies, Castanospermum australe Lane 1 (mature axes), lane 2 (maturecotyledons), lane 3 (immature axes), lane 4 (immature cotyledons).Dehydrin-related proteins were detected by a dehydrin antiserum andblots using the pre-immune serum served as controls (pre-immune).Heat-stable protein (64 /xg in 32 fi\ sample buffer) was loaded on toeach lane. Numbers on the right indicate the approximate molecularmasses in kDa of the dehydrin-related polypeptides; only those proteinsnot present on a control Western blot (using pre-immune serum) areindicated. Left-most lanes = molecular mass markers of 139.9, 86.8,47.8, 33.3, 28.6 and 20.7 kDa.
nature of the dehydrins accumulated in cotyledonsoccurred as seeds matured (compare lanes 2 and 4).
Dehydrins associated with late maturation of C. australe areinduced in immature seeds by ABA and moderate stressesbut not in response to a more extreme desiccationtreatment
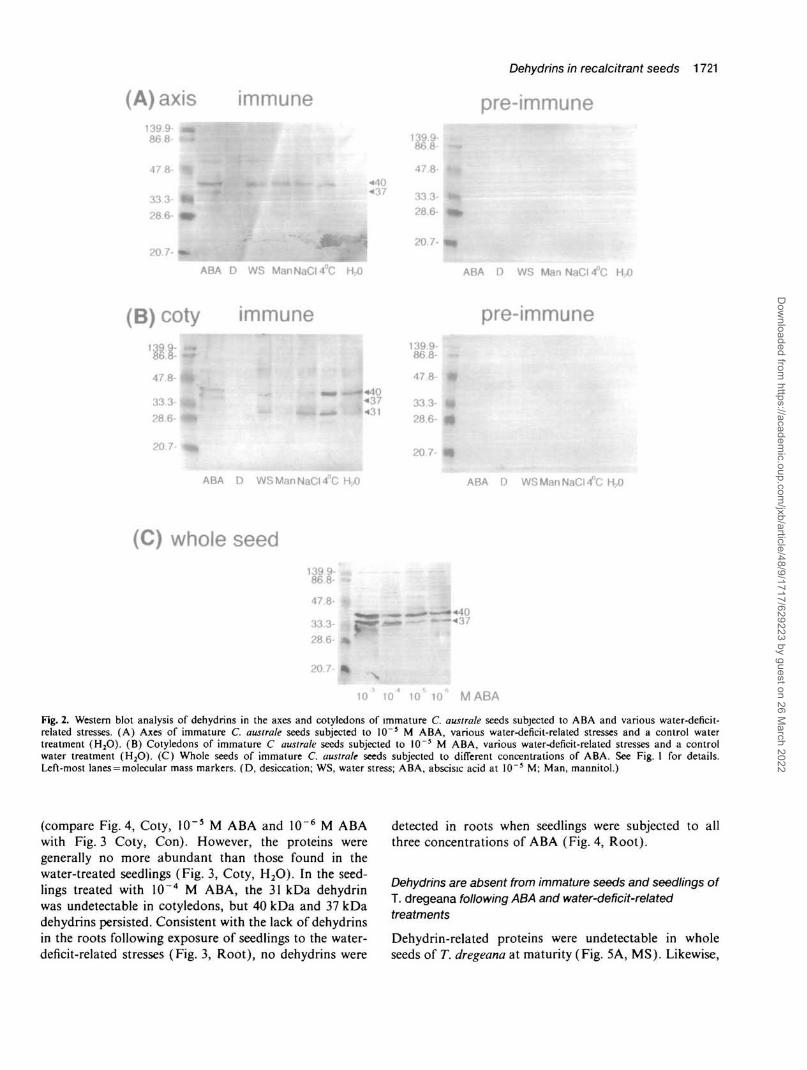
Dehydrin-related proteins were detected by Western blot-ting in axes and cotyledons of immature C. australe seedsfollowing treatment of seeds with ABA or various water-deficit-related stresses (Fig. 2). A 40 kDa dehydrin wasinduced in the immature axis by ABA, water stress, highosmolarity, high salt, and cold (Fig. 2A). The 31 and85 kDa proteins, that were abundant in the immature(untreated) axes (Fig. 1, lane 3), declined (Fig. 2A). Adesiccation treatment that resulted in more extreme waterloss, did not elicit dehydrin synthesis. In the cotyledons,40 kDa dehydrins were induced by the mild water-deficit-related treatments and similar to the situation in axes, no
dehydrins were present following the desiccation treat-ment (Fig. 2B). Thus it appears that dehydrins fail toaccumulate under conditions of more extreme water loss.It is likely that the seeds did not survive the treatmentand therefore were incapable of protein synthesis.Dehydrins were present in cotyledons of seeds subjectedto a control water treatment (see Materials and methods)but not in the axes of these control seeds (Fig. 2A, B,H2O). Thus, the detachment of immature seeds from theparent plant can induce dehydrin production. There wasno 31 kDa dehydrin in ABA-treated cotyledons of imma-ture seeds; instead a 37 kDa protein was detected alongwith the 40 kDa protein (Fig. 2B). The 37 kDa proteinwas also detected in ABA-treated axes (Fig. 2A). Allconcentrations of ABA (10~3-10~6 M) induced 40 kDaand 37 kDa dehydrin proteins in immature whole seeds(axis and cotyledons) (Fig. 2C).
Dehydrins decline following ABA and water-deficit-relatedtreatments in the cotyledons of C. australe seedlings andare undetectable in the roots of the seedlings
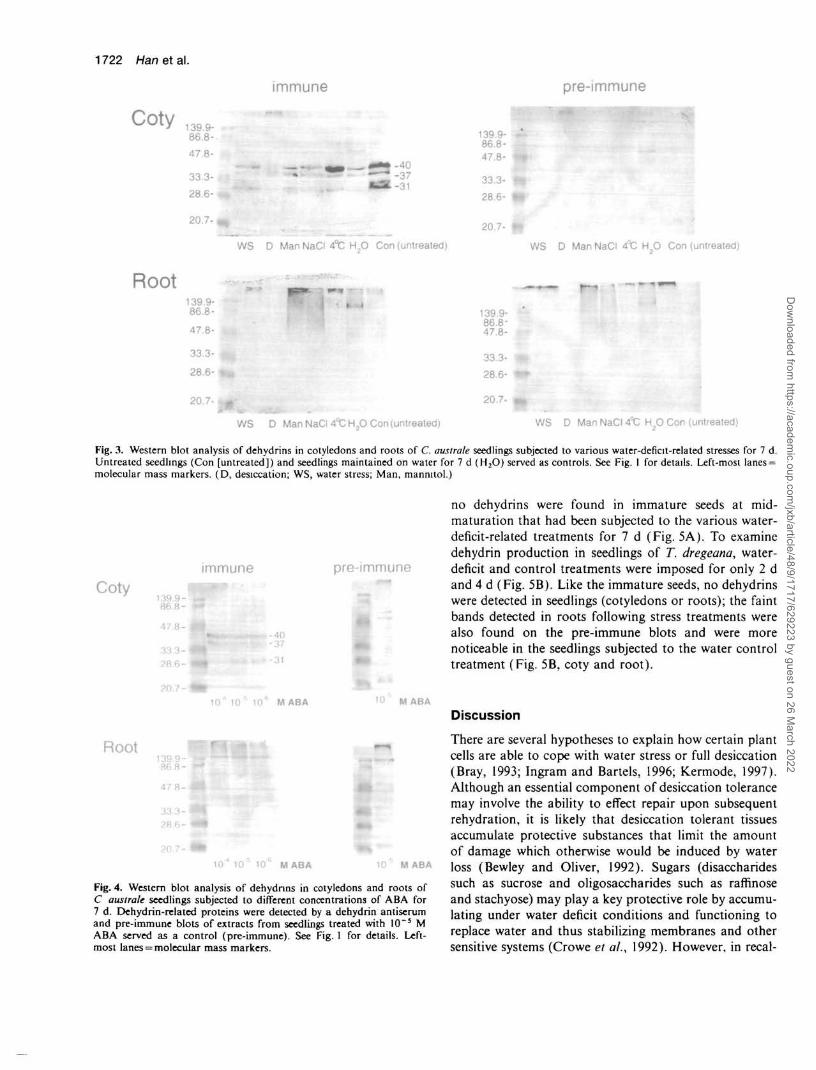
Dehydrin-related proteins in cotyledons and roots of C.australe seedlings exposed to the different water-deficit-related stresses are shown in Fig. 3. In all the experiments,seedlings were subjected to 7 d treatments. Followingimbibition of mature seed (i.e. in untreated seedlings),the dehydrins present in cotyledons of mature seedspersisted (compare Fig. 1 mature cotyledons, lane 2, withFig. 3, Con [untreated]). Following the 7 d treatments,all dehydrins became less abundant in the cotyledons ofthe seedlings, with the exception of the 40 kDa protein incold-treated seedlings (4°C).
Water-treated seedlings served as an additional control(Fig. 3, H2O). These seedlings, which were maintainedon water for the same length of time as that required toimpose the stress treatments (7 d), showed a persistenceof the 40 and 37 kDa dehydrins, although the 31 kDaprotein declined. Dehydrins were not detectable in cotyle-dons of desiccation-treated seedlings (Fig. 3), which wassimilar to the response of the cotyledons of desiccation-treated immature seeds (Fig. 2B).
Interestingly, although dehydrin-related polypeptideswere present in mature axes (Fig. 1, lane 1), no dehydrinswere detectable in the roots of C. australe untreatedseedlings (Fig. 3, Root, Con [untreated]) nor in theseedlings subjected to the different water-deficit-relatedtreatments (Fig. 3). A lack of dehydrins in the roots maycontribute to desiccation sensitivity in C. australe.
The presence of dehydrin-related proteins was exam-ined in cotyledons and axes of C. australe seedlingsfollowing 7 d treatments in different concentrations ofABA (Fig. 4). All three dehydrin proteins (40 kDa,37 kDa and 31 kDa) persisted in the cotyledons of seed-lings, following treatment with 10 ~5 and 10 ~6 M ABA
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022
i) axis139.9- »868
47 8-
33 3- • *
immune
•40••37
m.478
333286
20.7-20 7-
Dehydrins in recalcitrant seeds 1721
pre-immune
ABA D WS Man NaCI 4 C H.0 ABA D WS Man NaCI 4°C HJ)
(B) coty
4 7 . 8 - -•••••
33.3- .„
28.6- « l
20 7 «fe
immune
«37
139.9-86.8-
47.8-
33.3-
286-
#
m20.7-
pre-immune
ABA D WS Man NaCI 4'C H?0 ABA D WS Man NaCI 4°C H.,0
(C) whole seed
40-—•«37
m v -••-47.8-
33.3- r^^* ~r
28.6 *
2 0 7 *
10 10" 10 10 M ABA
Fig. 2. Western blot analysis of dehydrins in the axes and cotyledons of immature C. australe seeds subjected to ABA and various water-deficit-related stresses. (A) Axes of immature C. australe seeds subjected to 10~5 M ABA, various water-deficit-related stresses and a control watertreatment (H2O). (B) Cotyledons of immature C australe seeds subjected to IO~5 M ABA, various water-deficit-related stresses and a controlwater treatment (H2O). (C) Whole seeds of immature C. australe seeds subjected to different concentrations of ABA. See Fig. 1 for details.Left-most lanes = molecular mass markers. (D, desiccation; WS, water stress; ABA, abscisic acid at 10~5 M; Man, mannitol.)
(compare Fig. 4, Coty, 10"5 M ABA and 10"6 M ABAwith Fig. 3 Coty, Con). However, the proteins weregenerally no more abundant than those found in thewater-treated seedlings (Fig. 3, Coty, H2O). In the seed-lings treated with 10 ~A M ABA, the 31 kDa dehydrinwas undetectable in cotyledons, but 40 kDa and 37 kDadehydrins persisted. Consistent with the lack of dehydrinsin the roots following exposure of seedlings to the water-deficit-related stresses (Fig. 3, Root), no dehydrins were
detected in roots when seedlings were subjected to allthree concentrations of ABA (Fig. 4, Root).
Dehydrins are absent from immature seeds and seedlings of
T. dregeana following ABA and water-deficit-related
treatments
Dehydrin-related proteins were undetectable in wholeseeds of T. dregeana at maturity (Fig. 5A, MS). Likewise,
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022
1722 Han et al.
Cotyimmune pre-immune
139.9-86.8-
47.8-
33.3- i28.6- m
20.7- m
-40-37-31
WS D Man NaCI 4°C H2O Con (untreated)
139.9-86.8-47.8- * f
33.3- *a
28.6- m
20.7- #*
WS D Man NaCI 4°C H.,0 Con (untreated)
Root139.9-86.8-
47.8-
33.3-
28.6-
20.7- «*•
139.9-86.8"47.8-
33.3-
28.6-
20.7-
WS D Man NaCI 4 t H , O Con (untreated) WS D Man NaCI 4 t H O Con (untreated)
Fig. 3. Western blot analysis of dehydrins in cotyledons and roots of C. australe seedlings subjected to various water-deficit-related stresses for 7 d.Untreated seedlings (Con [untreated]) and seedlings maintained on water for 7 d (H2O) served as controls. See Fig. 1 for details. Left-most lanes =molecular mass markers. (D, desiccation; WS, water stress; Man, mannitol.)
Coty
Root
immune pre-immune
139.9-86 8-
47 R-
33.3- j28.6-!
20.7 -1
139 086 H -
47 8-
33 3-28 6-
20.7- I
.-40-37
-31
10'' 10" 10* M ABA 10 MABA
10" 10 5 10* MABA 10 MABA
Fig. 4. Western blot analysis of dehydnns in cotyledons and roots ofC australe seedlings subjected to different concentrations of ABA for7 d. Dehydrin-related proteins were detected by a dehydrin antiserumand pre-immune blots of extracts from seedlings treated with 10~s MABA served as a control (pre-immune). See Fig. 1 for details. Left-most lanes = molecular mass markers.
no dehydnns were found in immature seeds at mid-maturation that had been subjected to the various water-deficit-related treatments for 7 d (Fig. 5A). To examinedehydrin production in seedlings of T. dregeana, water-deficit and control treatments were imposed for only 2 dand 4 d (Fig. 5B). Like the immature seeds, no dehydrinswere detected in seedlings (cotyledons or roots); the faintbands detected in roots following stress treatments werealso found on the pre-immune blots and were morenoticeable in the seedlings subjected to the water controltreatment (Fig. 5B, coty and root).
Discussion
There are several hypotheses to explain how certain plantcells are able to cope with water stress or full desiccation(Bray, 1993; Ingram and Bartels, 1996; Kermode, 1997).Although an essential component of desiccation tolerancemay involve the ability to effect repair upon subsequentrehydration, it is likely that desiccation tolerant tissuesaccumulate protective substances that limit the amountof damage which otherwise would be induced by waterloss (Bewley and Oliver, 1992). Sugars (disaccharidessuch as sucrose and oligosaccharides such as raffinoseand stachyose) may play a key protective role by accumu-lating under water deficit conditions and functioning toreplace water and thus stabilizing membranes and othersensitive systems (Crowe et al., 1992). However, in recal-
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022

Dehydrins in recalcitrant seeds 1723
A. Seeds
immune pre-immune
•8*; '»* =47 8 - , 4 7 8 *
28 6- %
2 0 . 7 - ^ 2 0 7 " « »
WS D ManNaCI4uCH,0 MS WS D Man NaCI4(1C H.,0 MS
B. Seedlings
(A) coty immune pre-immune
1399-Q(> Q .' i OQ Q
oc ~o
47 8- ': 47 8-
3 3 3- i 33 32 8 . 6 - •_.-,•) 3 3 ; 3 ;I
— - - 207' ( •D 4"C HO D 4"C HO Con (untreated) D 4 < t H „ D ,o Con (untreated)
( d 3 y S ) 4 4 4 2 2 2 ( d a y s ) 4 4 4 2 2 2
(B) root immune pre-immune
139 9- m
86.8-47 8- K_. ' , 9 47 8
%'£ '" " 33.3-I 28.6-
2 0 7" • • 20.7-
iD 4 " C H 0 D 4 (untreated) D 4"C H 0 D 4(>C HVO Con (untreated)
(days) 4 4 4 2 2 2 (days) 4 4 4 2 2 2
Fig. 5. Western blot analysis to detect dehydrins in mature seeds of T. dregeana and in immature seeds and seedlings of T. dregeana subjected tovarious water-deficit-related stresses. (A) Untreated mature seeds (MS) and immature seeds subjected to water-deficit-related stresses or a controlwater treatment (H2O) for 7 d. (D, desiccation; WS, water stress; Man, mannitol). (B) Cotyledons and roots of seedlings of T. dregeana subjectedto water-deficit-related stresses (desiccation, D and cold, 4°C) or a control water treatment (H2O) for 2 d or 4 d as indicated. Also shown are theuntreated seedlings (Con [untreated]). See Fig.l for details. Left-most lanes = molecular mass markers.
citrant seeds, viability is generally lost at water contents in seeds and plant vegetative tissues undergoing mildfar higher than those at which water replacement (if it water loss or full desiccation—namely dehydrins.occurs in seeds at all) would take place. Another protect- Desiccation tolerance is acquired during developmentive mechanism may involve the group of proteins induced of orthodox seeds; tolerance to full desiccation is generally
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022

1724 Han et al.
lost after germination. Recalcitrant seeds, unlike orthodoxseeds, are sensitive to desiccation when shed from theparent plant, and thus provide a system to study temporaland stress-induced changes in dehydrins.
In this study, dehydrin-related polypeptides were identi-fied in recalcitrant seeds of Castanospermum australe.However, they were absent from mature seeds of Trichiliadregeana, another tropical species and none of the water-deficit-related treatments imposed at any stage wereeffective in eliciting dehydrin production in this species.
Given that dehydrins are produced by mature seeds ofC. australe, but not by those of T. dregeana, a pertinentquestion is whether there are differences in the relativedesiccation sensitivity of the seeds of these tropical species.However, no clear correlations are evident; followingflash drying of mature isolated axes of C. australe, thecritical moisture level at which the electrolyte conductivityof the leachate increases (indicative of membrane damage)is 0.98 g H2O g"1 dry mass. For T. dregeana, the criticalmoisture content for survival of mature axes in tissueculture is 0.2 g H2Og~' dry mass (Pammenter andBerjak, unpublished results). (However, it may not bevalid to directly compare desiccation sensitivity on thebasis of the different indices of survival, i.e. electrolyteleakage and capacity to survive and grow in tissueculture.)
Although more extensive investigation is required, thereis some evidence to suggest that while dehydrin-likeproteins are consistently found in recalcitrant seeds oftemperate species, they are often absent from recalcitrantseeds of tropical wetland species (Farrant et al., 1996).Finch-Savage et al. (1994) detected dehydrins in matureseeds of five desiccation sensitive (recalcitrant) trees (alltemperate species), and Lea mRNA can also be inducedin stored recalcitrant seeds of Quercus robur L. (Englishoak) by ABA and limited drying treatments. Dehydrinshave also been detected in seeds of Zizania palustris, acold temperate aquatic grass, the seeds of which exhibitcharacteristics intermediate between recalcitrance andorthodoxy (Bradford and Chandler, 1992).
In contrast to the consistent presence of dehydrins ina range of recalcitrant seeds of temperate species, dehyd-rins have been reported to be absent during the late stagesof development of recalcitrant mangrove (Avicenniamarina) seeds, a finding suggested to account for the lackof desiccation tolerance of this tropical species (Farrantet al., 1992, 1996). In addition to Avicennia marina, seedsof several other tropical wetland species do not containdehydrins in amounts detectable by Western blotting(Farrant et al., 1996; reviewed in Kermode, 1997). Thusit is important to analyse species that inhabit a range ofclimatic zones and environments. Both seeds studiedherein are of tropical origin, but in constrast toT. dregeana, seeds of C. australe underwent maturationin a temperate climate. C. australe is an Australian species;
the material collected for these studies was from treesgrown in the streets of Pietermaritzburg, South Africa,which is hotter in summer and cooler in winter thanDurban (where seeds of T. dregeana were obtained) andhas slightly lower rainfall.
There were striking changes in the nature of the dehyd-rins present during development of C. australe seeds. Adistinct set of dehydrin-related polypeptides was presentin the axes and cotyledons of immature seeds at mid-maturation; new dehydrins were present in mature seeds.Immature C. australe seeds subjected to ABA or variouswater-deficit-related treatments produced some of thedehydrins associated with late development, but some ofthese were also present in detached hydrated seeds, atleast in the cotyledons. The general characteristics ofdehydrin production in the recalcitrant C. australe seedsduring development (i.e. new polypeptides induced duringlate development and following mild stress or detachmentof immature seeds) are very similiar to those exhibited byseeds of an orthodox tropical species that we have beenstudying, namely castor bean (Ricinus communis L.) (Hanet al., 1995, 1997; Han and Kermode, 1996).
In relation to dehydrin production, the major differ-ences between seeds of C. australe and R. communisappear to be exhibited following the transition to germina-tion and seedling growth. In R. communis, production ofdehydrin-related polypeptides exhibits some tissue-specificity, is dependent upon the physiological stage ofthe seed and exhibits some qualitative and quantitativedifferences in response to different water-deficit-relatedstresses (Han and Kermode, 1996). In other orthodoxspecies such as barley, three lea mRNAs respond differ-ently in seedlings subjected to salt, cold, mannitol andABA (Espelund et al., 1992). These mRNAs also showdifferent expression patterns during barley seed develop-ment. Similar to the endosperm of R. communis seeds,the dehydrin-related polypeptides persist in the cotyledonsof C. australe seeds following imbibition. However, fol-lowing ABA treatments or the imposition of all water-deficit-related stresses (except cold) these dehydrinsbecame less abundant in the cotyledons of C. australeseedlings and no new polypeptides were detected. Incontrast, R. communis seedlings exhibit a strong inductionof dehydrin synthesis in the endosperm, cotyledons andradicle as a consequence of ABA or stress imposition(Han and Kermode, 1996). Furthermore, in the endo-sperm of R. communis seedlings the stress-inducible dehy-drins are distinct from those that are induced during lateseed development and which persist following germina-tion/growth of mature seeds.
Perhaps the most significant finding is the lack ofdehydrins in the axis and roots of C. australe followingseed germination and the inability of high concentrationsof ABA or stress imposition to induce their production.It seems that the axes of C. australe lose their ability to
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022

produce dehydrins after germination, despite having thiscapacity during seed development. In contrast to C.australe, mature seeds of T. dregeana did not containdehydrins and none were induced by stress or ABAtreatments imposed either during seed development orafter germination.
There is limited evidence that seeds of recalcitrantspecies are much less sensitive to applied ABA than aretheir orthodox counterparts; part of the underlying basisfor the apparent insensitivity may be a very active ABAmetabolism demonstrated by a rapid [3H]-ABA turnoverand high levels of endogenous conjugates (e.g. in recalcit-rant Hopea orodata seeds) (Garello and Le Page-Degivry,1995; see also Farrant et al., 1996).
A decline in the ability of the recalcitrant seeds studiedherein to produce dehydrins following the transition toseedling growth may be more important to the viabilityof shed seed than a previous capacity for synthesis duringseed development. It may be significant that the changesthat occur on dehydration of recalcitrant seeds of themangrove, Avicennia marina, are very similar to changesbrought about by desiccation of orthodox seeds duringthe intolerant stage following germination (Farrant et al.,1986). Recalcitrant seeds initiate germination-relatedmetabolism shortly after shedding (reviewed in Vertucciand Farrant, 1995), and in Avicennia marina, 10-15 dbefore shedding (Farrant et al., 1993). As germinationevents progress, the seeds become increasingly sensitiveto drying and attempting to store these seeds is akin toair-dry storage of germinated, orthodox seeds (Farrantet al., 1986, 1988). There is no clear-cut event delineatingthe end of seed development and the start of germination;during both phases, recalcitrant seeds appear to remainmetabolically active, although the axes may undergo avery brief period of relative quiescence. If indeed the shedrecalcitrant seed responds in a manner similar to germin-ated orthodox seeds, it is also important to distinguishbetween dehydrin production in storage tissues (cotyle-dons or endosperm) and that occurring in the axis. Forexample, the axes of orthodox seeds such as soybeanrapidly lose their tolerance to desiccation (brought aboutby air-drying to 10% water content) during the course ofgermination, while the cotyledons remain tolerant for aconsiderably longer period (Senaratna and McKersie,1983).
Finch-Savage et al. (1994) showed an induction of LeamRNA in axes of stored recalcitrant seeds of Quercusrobur L. (English oak, a temperate species) followingtreatment with ABA or limited desiccation. However, inthis temperate species, similar to the tropical species, C.australe, there was no true induction nor even enhancedsynthesis of dehydrin-related polypeptides in response toABA or limited desiccation, which is characteristic oforthodox seeds (reviewed by Bray, 1991, 1993; Thomaset al., 1991; Bewley and Oliver, 1992; Kermode, 1990,
Dehydrins in recalcitrant seeds 1725
1995, 1997; Chandler and Robertson, 1994; Bewley andBlack, 1994). The very small amount of dehydrin-relatedpolypeptides found in the untreated mature or stored Q.robur seeds declined upon application of ABA or imposi-tion of stress (Finch-Savage et al., 1994).
The results of this study support the contention thatdesiccation-sensitivity of recalcitrant seeds is due, in part,to an inability to accumulate sufficient dehydrins and/orto the absence of specific LEA or dehydrin proteins,especially following the transition to a germination/growth programme under stress conditions. Future stud-ies will determine whether there is a direct relationshipbetween a loss of viability during storage (when seeds canbe considered to be under stress) and a decline in theability of the axes of recalcitrant species to synthesizedehydrins. The role of environment during seed matura-tion also needs further investigation. It is likely that theunderlying basis of desiccation tolerance is diverse and isnot simply restricted to the synthesis of specific proteins.
Acknowledgements
We are grateful to TJ Close, UC Riverside, for providing thedehydrin antibody. This research was supported by NaturalSciences and Engineering Research Council of Canada(NSERC) Operating and Equipment Grants to ARK.
References
Berjak P, Pammenter NW, Vertucci C. 1992. Homoiohydrous(recalcitrant) seeds: developmental status, desiccation sensitiv-ity and the state of water in axes of Landolphia kirkii Dyer.Planta 186,249-61.
Bewley J, Black M. 1994. Seeds. Physiology of development andgermination. 2nd edn. New York: Plenum Press.
Bewley JD, Oliver MJ. 1992. Desiccation tolerance in vegetativeplant tissues and seeds: protein synthesis in relation todesiccation and a potential role for protection and repairmechanisms. In: Somero GN, Osmond CB, Bolis CL, eds.Water and life: comparative analysis of water relationships atthe organismic, cellular and molecular levels. Berlin: Spnnger-Verlag, 141-60.
Blackmail SA, Obendorf RL, Leopold AC. 1992. Maturationproteins and sugars in desiccation tolerance of developingsoybean seeds. Plant Physiology 100, 225-30.
Blackman SA, Wettlaufer SH, Obendorf RL, Leopold AC. 1991Maturation proteins associated with desiccation tolerance insoybean. Plant Physiology 96, 868-74.
Bradford M. 1976. A rapid method for the quantitation ofmicrogram quantities of protein utilizing the principle ofprotein-dye binding. Analytical Biochemistry 72, 248-54.
Bradford KJ, Chandler PM. 1992. Expression of'dehydrin-like'proteins in embryos and seedlings of Zizania palustris andOryza saliva during dehydration. Plant Physiology 99, 488-94.
Bray EA. 1991. Regulation of gene expression by endogenousABA during drought stress. In: Davies WJ, Jones HG, eds.Abscisic acid phvsiology and biochemistry. Oxford: BIOS,81-98.
Bray EA. 1993. Molecular responses to water deficit. PlantPhysiology 103, 1035^0.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022

1726 Han et al.
Chandler PM, Robertson M. 1994. Gene expression regulatedby abscisic acid and its relation to stress tolerance. AnnualReview of Plant Physiology and Plant Molecular Biology45, 113-41.
Close TJ. 1996. Dehydrins: emergence of a biochemical role ofa family of plant dehydration proteins. Physiotogia Plantarum97, 795-803.
Close TJ, Fenton RD, Moonan F. 19936. A view of plantdehydrins using antibodies specific to the carboxy terminalpeptide. Plant Molecular Biology 23, 275-86.
Close TJ, Fenton RD, Yang A, Asghar R, DeMason DA, CroneDE, Meyer NC, Moonan F. 1993a. Dehydrin: the protein. In:Close TJ, Bray EA, eds. Responses of plants to cellulardehydration during environmental stress. Rockville, MD:American Society of Plant Physiologists, 104-18.
Crowe JH, Hoekstra FA, Crowe LM. 1992. Anhydrobiosis.Annual Review of Physiology 54, 579-99.
Dure III LS. 1993. A repeating 11-mer amino acid motif andplant desiccation. The Plant Journal 3, 363-9.
Espelund M, Saeboe-Larssen S, Hughes DW, Galau GA, LarsenF, Jakobsen KS. 1992. Late embryogenesis abundant genesencoding proteins with different numbers of hydrophilicrepeats are regulated differentially by abscisic acid andosmotic stress. The Plant Journal 2, 241-52.
Farrant JM, Berjak P, Cutting JGM, Pammenter NW. 1993.The role of plant growth regulators in the development andgermination of the desiccation-sensitive (recalcitrant) seedsof Avicennia marina. Seed Science Research 3, 55-63.
Farrant JM, Berjak P, Pammenter NW. 1992. Protein indevelopment and germination of a desiccation sensitive(recalcitrant) seed species. Plant Growth Regulation 11,257-65.
Farrant JM, Pammenter NW, Berjak P. 1986. The increasingsensitivity of recalcitrant Avicennia marina seeds with storagetime. Physiologia Plantarum 67, 291-8.
Farrant JM, Pammenter NW, Berjak P. 1988. Recalcitrance -a current assessment. Seed Science and Technology 16, 155-66.
Farrant JM, Pammenter NW, Berjak P, Farnsworth EJ, VertucciCW. 1996. Presence of dehydrin-like proteins and levels ofabscisic acid in recalcitrant (desiccation sensitive) seeds maybe related to habitat. Seed Science Research 6, 175-82.
Finch-Savage WE, Pramanik SK, Bewley JD. 1994. Theexpression of dehydrin proteins in desiccation-sensitive (recal-citrant) seeds of temperate trees. Planta 193, 478-85.
Garello G, Le Page-Degivry M-T. 1995. Desiccation-sensitiveHopea odorata seeds: sensitivity to abscisic acid, waterpotential and inhibitors of gibberellin biosynthesis.Physiologia Plantarum 95, 45-50.
Han B, Kermode AR. 1996. Dehydrin-like proteins in castorbean seeds and seedlings are differentially produced inresponse to ABA and water-deficit-related stresses. Journalof Experimental Botany 47, 933-9.
Han B, Hughes DW, Galan GA, Bewley JD, Kermode AR. 1997.
Changes in late embryogenesis abundant (Lea) messengerRNAs and dehydrins during maturation and prematuredrying of Ricinus communis L. seeds. Planta 201, 27-35.
Han B, Pammenter N, Farrant J, Berjak P, Kermode AR. 1995.A comparative analysis of dehydrin synthesis in seeds andseedlings of desiccation tolerant and recalcitrant species.Proceedings of the 22nd Annual Meeting of the Plant GrowthRegulator Society of America. PGRSA Quarterly 23,244-8.
Ingram J, Bartefe D. 1996. The molecular basis of dehydrationtolerance in plants. Annual Reviews of Plant Physiology andPlant Molecular Biology 47, 377^03.
Kermode AR. 1990. Regulatory mechanisms involved in thetransition from seed development to germination. CriticalReviews in Plant Sciences 9, 155-95.
Kennode AR. 1995. Regulatory mechanisms in the transitionfrom seed development to germination: interactions betweenthe embryo and the seed environment. In: Galili G, Kigel J,eds. Seed development and germination. New York: MarcelDekker Inc., 273-332.
Kermode AR. 1997. Approaches to elucidate the basis ofdesiccation-tolerance in seeds. Seed Science Research 7, 75-95.
Kennode AR, Bewley JD. 1985. The role of maturation dryingin the transition from seed development to germination. I.Acquisition of desiccation-tolerance and germinability duringdevelopment of Ricinus communis L. seeds. Journal ofExperimental Botany 36, 1906-15.
Laemmli UK. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680-5.
Probert RJ, Brierley ER. 1989. Desiccation intolerance in seedsof Zizania palustris is not related to developmental age orthe duration of post-harvest storage. Annals of Botany64, 669-74.
Senaratna T, McKersie BD. 1983. Characterization of soluteefflux from dehydration injured soybean (Glycine max L.Merr) seeds. Plant Physiology 72, 911 ^4.
Smith MT, Berjak P. 1995. Deteriorative changes associatedwith the loss of viability of stored desiccation-tolerant anddesiccation-sensitive seeds. In: Galili G, Kjgel J, eds. Seeddevelopment and germination. New York: Marcel DekkerInc., 237-71.
Thomas TL, Vivekananda J, Bogue MA. 1991. ABA regulationof gene expression in embryos and mature plants. In: DaviesWJ, Jones HG, eds. Abscisic acid physiology and biochemistry.Oxford: BIOS, 125-35.
Vertucci CW, Crane J, Porter RA, Oelke EA. 1994. Physicalproperties of water in Zizania embryos in relation to maturitystatus, moisture content and temperature. Seed ScienceResearch 4, 211-24.
Vertucci CW, Fan-ant JM. 1995. Acquisition and loss ofdesiccation tolerance. In: Galili G, Kigel J, eds. Seeddevelopment and germination. New York: Marcel DekkerInc., 701-46.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/9/1717/629223 by guest on 26 M
arch 2022




















