
The promoter regulatory regions of the genes for the cytosolic form of phosphoenolpyruvate...
10
Biochemical and Molecular Roles of Nutrients The Promoter Regulatory Regions of the Genes for the Cytosolic Form of Phosphoenolpyruvate Carboxykinase (GTP) from the Chicken and the Rat Have Different Species-Specific Roles in Gluconeogenesis 1,2 Summer P. Savon, Parvin Hakimi, 3 Deborah R. Crawford, 3 Dwight J. Klemm, Austin L. Gurney and Richard W. Hanson 4 Department of Biochemistry, Case Western Reserve University School of Medicine, Cleveland, OH 44106-4935 ABSTRACT Hepatic expression of the gene for phosphoenolpyruvate carboxykinase (GTP) (PEPCK-C) (EC 4.1.1.32) in birds occurs prior to birth and decreases to negligible levels before hatching, whereas in mammals the gene for PEPCK-C in the liver is expressed at birth and is active throughout the life of the animal. The administration of cyclic AMP to adult chickens results in the induction of transcription of the gene for PEPCK-C and the transient accumulation of PEPCK-C mRNA in the liver. DNase I footprint analysis of 330 bp of the avian PEPCK-C promoter immediately 5* of the start-site of transcription indicated the presence of several protein binding domains, purified CAAT/enhancer binding protein a, cAMP regulatory element binding protein and nuclear factor-1 bound to these regions of the promoter. Sequences corresponding to an hepatic nuclear factor-1 binding domain and to the insulin response sequence, previously identified in the rat PEPCK-C promoter, were also found in the chicken PEPCK-C promoter. Co-transfection of an expression vector for CAAT/enhancer binding protein a or CAAT/enhancer binding protein b markedly stimulated transcription from both the chicken and rat PEPCK-C promoters in human hepatoma cells. Sequences involved in the regulation of gene transcription by cyclic AMP and insulin were found to reside between 0210 and /1 of the avian PEPCK-C promoter. In general, transcription from the avian promoter was more sensitive to inhibition by insulin than was noted for the rat PEPCK-C promoter, which may explain in part the lack of expression of the gene for PEPCK-C in the livers of adult birds. J. Nutr. 127: 276–285, 1997. KEY WORDS: • phosphoenolpyruvate carboxykinase • birds • transcription factors • insulin • cAMP clearly the mechanistic basis for the differential patterns of There are major differences in the regulation of gluconeo- gene expression in birds. This is of special interest in the genesis in birds compared with mammals, which are due in case of PEPCK-C, in view of its very complex arrangement of part to the unique features of P-enolpyruvate carboxykinase hormone-responsive elements (Hanson and Patel 1994). It is (PEPCK) 5 gene expression in birds (Hod et al. 1986, Shen probable that the gene for PEPCK-C in birds represents a and Mistry 1978a and 1978b, Watford et al. 1981). Because functional variant of its counterpart in mammals and that key the gene for the cytosolic form of PEPCK (PEPCK-C) in mam- differential regulatory features may highlight important aspects mals is subjected to a high degree of regulation (Hanson and of promoter architecture involved in gene regulation. An in- Patel 1994, Lamers et al. 1982, Magnunson et al. 1987, sight into the regulation of PEPCK-C gene expression in the McGrane et al. 1988), we have studied the transcription of chicken may also provide a greater understanding of the spe- the gene for PEPCK-C from the chicken to understand more cies-specific variation in the regulation of hepatic gluconeo- genesis. 1 Supported by grants DK 21859 and DK24451 (to R.W.H.) and by the Metabo- Organisms contain varying amounts of the two isozyme lism Training Program grant DK 07319 (to D.J.K. and D.R.C.) from the National forms of PEPCK, the cytosolic PEPCK-C and a mitochondrial Institutes of Health. The authors are indebted to Kulwant Singh Aulak for his PEPCK-M. These two forms of PEPCK are encoded by distinct critical help in the preparation of this manuscript. 2 The costs of publication of this article were defrayed in part by the payment nuclear genes (Hod et al. 1986) which have different patterns of page charges. This article must therefore be hereby marked ‘‘advertisement’’ of regulation. The gene for PEPCK-C in all species studied to in accordance with 18 USC section 1734 solely to indicate this fact. date is acutely regulated by diet and hormones, whereas the 3 Deborah Crawford and Parvin Hakimi made equal contributions to this work. 4 To whom correspondence should be addressed gene for PEPCK-M is largely constitutive in its pattern of 5 Abbreviations used: Bt2cAMP, dibutryl cyclic AMP; CAT, chloramphenicol expression [see (Hanson and Patel 1994) for a review]. Most acetyltransferase; C/EBP, CAAT/enhancer binding protein; CRE, cAMP regulatory mammalian species display almost equal activity of the two element; CREB, cAMP regulatory element-binding protein; GRE, glucorticoid re- sponse element; GRU, glucocorticoid regulatory unit; HNF-1, hepatic nuclear forms of PEPCK in their livers. However, there are metaboli- factor-1; IRS, insulin response sequence; NF-1, nuclear factor-1; PEPCK, phos- cally important variations in this pattern of expression of the phoenolpyruvate carboxykinase; PPARg2, peroxisome proliferator-activated re- ceptor g2. two isozyme forms of PEPCK which occur in several animal 0022-3166/97 $3.00 q 1997 American Society for Nutritional Sciences. Manuscript received 20 November 1995. Initial review completed 14 December, 1995. Revision accepted 28 October 1996. 276 / 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut by guest on January 12, 2016 jn.nutrition.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The promoter regulatory regions of the genes for the cytosolic form of phosphoenolpyruvate...

Biochemical and Molecular Roles of Nutrients
The Promoter Regulatory Regions of the Genes for the Cytosolic Form ofPhosphoenolpyruvate Carboxykinase (GTP) from the Chicken and the RatHave Different Species-Specific Roles in Gluconeogenesis1,2
Summer P. Savon, Parvin Hakimi,3 Deborah R. Crawford,3 Dwight J. Klemm,Austin L. Gurney and Richard W. Hanson4
Department of Biochemistry, Case Western Reserve University School of Medicine, Cleveland,OH 44106-4935
ABSTRACT Hepatic expression of the gene for phosphoenolpyruvate carboxykinase (GTP) (PEPCK-C) (EC4.1.1.32) in birds occurs prior to birth and decreases to negligible levels before hatching, whereas in mammalsthe gene for PEPCK-C in the liver is expressed at birth and is active throughout the life of the animal. Theadministration of cyclic AMP to adult chickens results in the induction of transcription of the gene for PEPCK-Cand the transient accumulation of PEPCK-C mRNA in the liver. DNase I footprint analysis of 330 bp of the avianPEPCK-C promoter immediately 5* of the start-site of transcription indicated the presence of several proteinbinding domains, purified CAAT/enhancer binding protein a, cAMP regulatory element binding protein and nuclearfactor-1 bound to these regions of the promoter. Sequences corresponding to an hepatic nuclear factor-1 bindingdomain and to the insulin response sequence, previously identified in the rat PEPCK-C promoter, were also foundin the chicken PEPCK-C promoter. Co-transfection of an expression vector for CAAT/enhancer binding protein aor CAAT/enhancer binding protein b markedly stimulated transcription from both the chicken and rat PEPCK-Cpromoters in human hepatoma cells. Sequences involved in the regulation of gene transcription by cyclic AMPand insulin were found to reside between 0210 and /1 of the avian PEPCK-C promoter. In general, transcriptionfrom the avian promoter was more sensitive to inhibition by insulin than was noted for the rat PEPCK-C promoter,which may explain in part the lack of expression of the gene for PEPCK-C in the livers of adult birds. J. Nutr.127: 276–285, 1997.
KEY WORDS: • phosphoenolpyruvate carboxykinase • birds • transcription factors • insulin • cAMP
clearly the mechanistic basis for the differential patterns ofThere are major differences in the regulation of gluconeo-gene expression in birds. This is of special interest in thegenesis in birds compared with mammals, which are due incase of PEPCK-C, in view of its very complex arrangement ofpart to the unique features of P-enolpyruvate carboxykinasehormone-responsive elements (Hanson and Patel 1994). It is(PEPCK)5 gene expression in birds (Hod et al. 1986, Shenprobable that the gene for PEPCK-C in birds represents aand Mistry 1978a and 1978b, Watford et al. 1981). Becausefunctional variant of its counterpart in mammals and that keythe gene for the cytosolic form of PEPCK (PEPCK-C) in mam-differential regulatory features may highlight important aspectsmals is subjected to a high degree of regulation (Hanson andof promoter architecture involved in gene regulation. An in-Patel 1994, Lamers et al. 1982, Magnunson et al. 1987,sight into the regulation of PEPCK-C gene expression in theMcGrane et al. 1988), we have studied the transcription ofchicken may also provide a greater understanding of the spe-the gene for PEPCK-C from the chicken to understand morecies-specific variation in the regulation of hepatic gluconeo-genesis.
1 Supported by grants DK 21859 and DK24451 (to R.W.H.) and by the Metabo- Organisms contain varying amounts of the two isozymelism Training Program grant DK 07319 (to D.J.K. and D.R.C.) from the National forms of PEPCK, the cytosolic PEPCK-C and a mitochondrialInstitutes of Health. The authors are indebted to Kulwant Singh Aulak for his
PEPCK-M. These two forms of PEPCK are encoded by distinctcritical help in the preparation of this manuscript.2 The costs of publication of this article were defrayed in part by the payment nuclear genes (Hod et al. 1986) which have different patterns
of page charges. This article must therefore be hereby marked ‘‘advertisement’’ of regulation. The gene for PEPCK-C in all species studied toin accordance with 18 USC section 1734 solely to indicate this fact.date is acutely regulated by diet and hormones, whereas the3 Deborah Crawford and Parvin Hakimi made equal contributions to this work.
4 To whom correspondence should be addressed gene for PEPCK-M is largely constitutive in its pattern of5 Abbreviations used: Bt2cAMP, dibutryl cyclic AMP; CAT, chloramphenicol expression [see (Hanson and Patel 1994) for a review]. Mostacetyltransferase; C/EBP, CAAT/enhancer binding protein; CRE, cAMP regulatory
mammalian species display almost equal activity of the twoelement; CREB, cAMP regulatory element-binding protein; GRE, glucorticoid re-sponse element; GRU, glucocorticoid regulatory unit; HNF-1, hepatic nuclear forms of PEPCK in their livers. However, there are metaboli-factor-1; IRS, insulin response sequence; NF-1, nuclear factor-1; PEPCK, phos- cally important variations in this pattern of expression of thephoenolpyruvate carboxykinase; PPARg2, peroxisome proliferator-activated re-ceptor g2. two isozyme forms of PEPCK which occur in several animal
0022-3166/97 $3.00 q 1997 American Society for Nutritional Sciences.Manuscript received 20 November 1995. Initial review completed 14 December, 1995. Revision accepted 28 October 1996.
276
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from

277TRANSCRIPTIONAL REGULATION OF THE GENE FOR AVIAN PEPCK
species. The rat, mouse and hamster express almost entirely an analysis of the hormonal regulatory elements present in thepromoter of the gene for PEPCK-C from the chicken and(ú90%) the gene for PEPCK-C (Hanson and Garber 1972,
Nordlie and Lardy 1963), whereas animals such as chicken, provide a comparison of the organization and function of theseelements with the better-studied transcriptional control of thepigeon and rabbit predominately express the gene for PEPCK-
M (Soling et al. 1973). The gene for PEPCK-C is hormonally PEPCK-C gene from mammals.responsive, so that starvation or acute diabetes significantlyinduces the activity of PEPCK-C in rabbit liver (Johnson et MATERIALS AND METHODSal. 1970).
Birds are unique among animals studied to date in that the Materials. All chemicals used were of reagent grade or the high-intracellular distribution of PEPCK varies among tissues, with est quality available commercially. Tissue culture media were DMEM
F-12 (GIBCO, Grand Island, NY), supplemented with calf serumthe liver containing only PEPCK-M, whereas both isozymic(GIBCO), and fetal calf serum (Hyclone, Logan, UT), unless notedforms are present in the kidney. This tissue-specific pattern ofotherwise. Radiolabeled nucleotides [a32P]dCTP (111 TBq/mmol),distribution of PEPCK may contribute to the differing abilities[g32P]ATP (222 TBq/mmol) and [35S]dATP (37 TBq/mmol) wereof chicken liver and kidney to support gluconeogenesis fromobtained from New England Nuclear, Wilmington, DE. Restrictiona variety of precursors (Watford et al. 1981). In tissues suchenzymes were purchased from Boehringer Mannheim, Indianapolisas avian liver in which PEPCK-M is the major isozymic form, IN or New England Biolabs, Beverly, MA and used with the buffers
gluconeogenesis occurs predominantly from lactate, whereas recommended by the manufacturer. Other enzymes were obtainedglucose formation from more oxidized substrates such as py- from the sources indicated. DNA ligase, Klenow fragment of DNAruvate and alanine occurs in tissues which contain PEPCK-C, polymerase I, calf intestinal phosphatase, S1 nuclease, DNase I andsuch as the avian kidney (Watford et al. 1981). These differ- polynucleotide kinase were from Boehringer Mannheim; RNase A
was from Sigma, St. Louis, MO. Radiolabeled, double-stranded DNAences in the regulation of gluconeogenesis may be related toprobes were routinely made using the Random Primer DNA labelingthe different ways of generating cytosolic NADH when specifickit from Boehringer Mannheim. The Sequenase Version 2.0 DNAmetabolic precursors are used in gluconeogenesis. If PEPCK-Sequencing kit from U.S. Biochemical, Cleveland, OH, was used toC is present, malate is transported across the mitochondrialobtain promoter sequence. G418 was purchased from GIBCO.membrane, generating NADH in the cytosol where it is oxi- Vectors and oligonucleotides were supplied by commercial anddized to oxaloacetate. Because the requisite cytosolic NADH private sources. pSCAN was obtained from Stratagene, La Jolla, CA
is being generated after the formation of oxaloacetate, a variety and the plasmids pH 4 and pH 6 were constructed as described pre-of gluconeogenic precursors can be used (alanine, pyruvate and viously (Hod et al. 1984). Primers complementary to the T3 andgluconeogenic amino acids), without respect to their initial T7 promoter regions of pSCAN were synthesized or purchased from
Stratagene, respectively. Oligonucleotides were chemically synthe-relative redox states. On the other hand, if only PEPCK-M issized using an Applied Biosystems 380A DNA Synthesizer (Fosterpresent in a tissue, gluconeogenesis occurs from lactate becauseCity, CA). The following proteins (with their source) were used foroxidation to pyruvate produces NADH directly in the cytosolDNase I footprinting analysis of the chicken PEPCK-C promoter:(Hanson and Garber 1972, Soling et al. 1973).CAAT enhancer binding protein a, (C/EBPa) Steven McKnight,This mode of expression of PEPCK in these two majorCarnegie Institute, Baltimore, MD; cAMP regulatory element bindinggluconeogenic organs may be the result of specific adaptations protein (CREB), Mark Montminy, Salk Institute, La Jolla, CA; and
in avian carbohydrate metabolism to meet the novel energy nuclear factor-1 (NF-1) CTF, N. Mermod and R. Tijan, Departmentdemands faced by birds. Avian flight muscle goes through peri- of Biochemistry, University of California, Berkeley, CA. C/EBPa andods of prolonged use in which the cycling of lactate to glucose CREB were purified after over-expression in Esherichia coli. Expressionvia the Cori cycle is essential. Avian liver supports this critical vectors for C/EBPa and C/EBPb were a gift from Steven McKnight.
Animals. Male, Sprague-Dawley rats, weighing 150–200 g, pur-metabolic function by expressing high levels of PEPCK-M. Inchased from Zivic Miller Company (Zelienople, PA), were used incontrast, the net glucose formation from amino acids thatthese studies. The animals were freely fed a nonpurified diet (Formu-occurs during starvation, when muscle protein itself is catabo-lab Chow, Ralston Mills, St. Louis, MO), a high energy, high proteinlized, requires the PEPCK-C activity found in the kidney. Indiet for rodents, and were provided with free access to water. Maleaddition, birds have a less pronounced need for glucose because Leghorn chickens (4–6 weeks old) weighing 250–400 g were hatchedthey have a relatively small brain and nucleated red blood cells in the Case Western Reserve Animal Resource Facility and were
with mitochondria and prefer ketone bodies as a metabolic fuel also fed a nonpurified diet (Lab Chick Chow S-G, Madina Farmers(Sturkie 1965). Birds can also use a diet in which fat replaces Exchange, Madina, OH). The chickens were used as a source ofcarbohydrate as a source of energy (Renner and Elcombe RNA for Northern analysis and for nuclear proteins for the DNase
I footprinting studies. All animals used for research purposes at Case1964).Western Reserve University were housed in a modern, AALAC certi-Despite the negligible level of PEPCK-C in chicken liver,fied animal facility under the supervision of board-certified veterinari-expression of its gene can be induced in the livers of adultans. The animals were handled in accordance with an animal usechickens by the administration of dibutryl cyclic AMPprotocol approved by the Institutional Animal Care and Use Com-(Bt2cAMP) (Hod et al. 1984). There is also a high level of mittee. The chickens were injected intraperitoneally with Bt2cAMPexpression of PEPCK-C in the embryonic liver of the chicken, and theophylline (30 mg/g body weight of each) or with saline (con-
which decreases to negligible levels 2 d before hatching (Savon trols); after 2 h the animals were killed by cervical dislocation, theet al. 1993). This contrasts with the pattern of development livers removed, and nuclear proteins and RNA isolated as describedof mammalian PEPCK-C in which the gene is not expressed below.
Cell culture and stable transfection. LMH cells were deriveduntil immediately after birth (Ballard and Hanson 1968).from a hepatocellular carcinoma which was induced in a male Leg-Thus, differences in the organization of regulatory elements inhorn chicken by long-term treatment with diethynitrosamine (Kawa-the promoter of the avian and mammalian PEPCK-C genesguchi et al. 1987). The LMH cells were kindly provided by Alan G.are likely to account for this variation in transcriptional capac-Goodridge, University of Iowa, Iowa City, IA. The LMH cells main-ity in fully differentiated liver from rats and chickens. In rat tain several differentiated morphological and biochemical features,
liver, a complex interaction between multiple hormones, in- including the expression of the genes for glucose-6-phosphatase andcluding glucagon, glucocorticoids thyroid hormone and insu- ATPase. The cells were cultured in Weymouths MB medium, con-lin, has been shown to control the level of PEPCK-C gene taining 10% fetal calf serum. The H4 and HepG2 cells used in this
study were maintained in Dulbecco’s modified Eagle’s medium, sup-expression (Hanson and Patel 1994). In this report, we present
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from

278 SAVON ET AL.
plemented with 5% calf serum and 5% fetal calf serum. During stable transfection into HepG2 cells (human) or LMH cells (chicken). ThePEPCK-C promoter was linked to the structural gene for chloram-transfection of DNA into the LMH or H4 cells, the cells were grown
to confluence in 10-cm dishes and then transfected with the PEPCK- phenicol acetyltransferase (CAT) and transiently transfected intoLMH or HepG2 cells by asiloglycoprotein receptor–mediated endo-neo gene. The cells were then selected in medium containing 500
mg/L of the antibiotic G418. cytosis (Crawford and Hanson 1993) of 10 mg of the PEPCK-CATvector together with 5 mg of a vector containing the b-galactosidasePreparation of nuclear extracts. Nuclear extracts were prepared
from rat or chicken liver by the method of Gorski et al. (1986). gene driven by the Rous sarcoma virus promoter. The effect of C/EBPa and C/EBPb on transcription from the PEPCK-C promoterThe purification of the cAMP regulatory element (CRE) binding
protein(s) from rat liver nuclear extracts was described previously was determined by co-transfection of 10 mg of the chimeric PEPCK-CAT gene, together with 5 mg of the expression vector containing(Roesler, et al. 1989).
DNase I footprinting. The DNA probes used in the footprinting the cDNA for the transcription factors, into HepG2 cells. Approxi-mately 4 1 106 trypsinized cells were added to the precipitated DNAassays were prepared by end-labeling one strand of DNA using T4
polynucleotide kinase and [g-32P]ATP. The DNase I footprinting and the mixture was plated out. Two days after transfection, thecells were harvested and lysed by freeze-thawing. CAT assays wereconditions were described previously (Roesler et al. 1989).
Construction of recombinant plasmids. The initial chimeric performed on equal amounts of protein from each extract as describedby Gorman et al. (1982). The activity of CAT in the transfectedgene, pSCP/Pf, was engineered by using the Klenow fragment of DNA
polymerase to fill in the ends of the 400 bp PvuII fragment of pH 4, cells is expressed relative to the activity of b-galactosidase to correctfor the efficiency of transfection. Each value is the average of threefollowed by ligation to pSCAN through ends created by digestion
with EcoRI. An additional plasmid, pSC-216del, containing a short- separate transfections experiments or as noted in the figure legends.Statistical analysis. The values derived for both the stable andened promoter sequence, was produced by first isolating the 250 bp
SmaI-ClaI fragment of pSCP/Pf. Compatible ends for ligation were transient transfection of genes containing the PEPCK-C promoterinto chicken and rat hepatocytes were analyzed statistically usingformed in pSCAN by EcoRI digestion, filled in using Klenow enzyme,
and subsequent restriction with ClaI. These two PEPCK-C promoter Student’s t distribution.segments were subsequently subcloned in front of the CAT structuralgene in the plasmid pXSV1CAT (Bokar et al. 1988). The larger RESULTStwo promoter fragments (1.8 kb and 700 bp) were also ligated into
Analysis of the sequence of the PEPCK-C promoter regionpXSV1CAT. Corresponding plasmids containing these two largerfrom the chicken. To analyze the regulation of transcriptionpromoter regions ligated to the neomycin resistance gene were madefrom the promoter for the gene for PEPCK-C from the chicken,by XbaI and ClaI digestion of pH 4 followed by ligation to the filled
in EcoRI site of pSCAN. The block mutation was created from four recombinant plasmids were created: pSCchickPEPC-1.8,pSCPEPC-366 using the approach described previously (Liu and Han- pSCchickPEPC-700, pSCchickPEC-366 and pSCchickPEPC-son 1991). 216. These plasmids contain 1.8 kb, 700 bp, 366 bp and 216
Cell transfection and isolation of RNA. Transfection of DNA bp, respectively, of the 5*-flanking region upstream of the tran-into eukaryotic cells was accomplished using calcium phosphate pre- scriptional start site in the PEPCK-C gene and were madecipitation (Ausubel et al. 1987) on dishes containing 80% confluent using the pSCAN vector which has a convenient multiplecultures. After an overnight exposure to the DNA, the cells were
cloning region immediately 5* to the neo structural gene. Theshocked with glycerol and then incubated with normal completemultiple cloning site is also flanked by oppositely orientedmedia for 2 d. Hormonal treatments were performed as described inT7 and T3 promoters which allow for sequencing of newlydetail previously (Liu et al. 1991). Final amounts of each hormoneintroduced DNA. The nucleotide sequence of the PvuII-PvuIIadded to an individual plate were as follows: Bt2cAMP (2.5 mg in
water), theophylline (1.8 mg in PBS) and insulin (30 mg/plate). The fragment of the PEPCK-C promoter had been previously deter-cells were incubated for 4 h in 10 mL of serum-free media, to which mined (Hod et al. 1984). Sequencing of this region, initiallythe individual hormones were added. performed to confirm the structure of the recombinant plasmid,
Total RNA was extracted from intact tissues and tissue culture indicated several errors in the earlier sequence analysis. Re-cells by homogenization in 4 mol/L guanidinium thiocyanate followed peated sequencing of this 400 bp promoter fragment in bothby centrifugation through a CsCl cushion (Chomczynski and Sacchi directions, combined with complete sequencing of the 1.8 kb1987). Pelleted RNA samples were resuspended, separated by electro-
fragment of pSCchickPEPC-1.8, permitted determination ofphoresis, transferred and hybridized under established conditions (Au-the primary nucleotide structure as indicated in Figure 1.subel et al. 1987).When aligned through the respective transcriptional start sites,S1 nuclease analysis. RNA analysis by S1 nuclease digestion was
performed as described previously (Hod et al. 1984). An end-labeled the promoter regulatory regions of PEPCK-C from the chickenDNA probe was made by digesting pSCP/Pf with NarI, followed by and rat have an overall sequence identity of 25%; this com-treatment with calf alkaline phosphatase. Subsequent restriction with pares with a 95% sequence identity between the rat and mouseEcoRI resulted in the formation of a 646 bp fragment which was for the same region of promoter of the PEPCK-C gene (S.gel isolated and end-labeled with polynucleotide kinase. Precipitated Hakimi, M. Johnson and R. Hanson, personal communication,RNA was resuspended in a formamide-containing hybridization solu- Case Western Reserve University). This large sequence diver-tion to which 1.75 kBq of probe was added. Denaturation of the
gence is evenly distributed throughout the region of the pro-DNA was achieved by a 15-min incubation at 757C, followed immedi-moters being compared, and sizeable local regions of sequenceately by overnight treatment at 527C to optimize DNA/RNA hybrididentity are conspicuously absent. The most extensive correla-formation. The next day, samples were cooled to 377C, digested withtion occurs within the sequence which spans from the pre-S1 nuclease and prepared for electrophoresis in a 6% denaturing
sequencing gel. sumed TATA box to the beginning of the first exon (035 toPrimer extension analysis. A synthetic oligodeoxynucleotide /1) which has 46% identity. A second area of relatedness
(SS-2) was end-labeled and used to prime DNA synthesis, with total (48%) occurs upstream and extends from promoter positionsRNA from chicken kidney serving as a template. The elongated, 01075 to 01050. This region of the PEPCK-C gene fromsingle-stranded DNA species thus formed was subjected to electro- the rat has been shown to contain a peroxisome proliferator-phoresis on a 6% polyacrylamide gel containing urea. The extension activated receptor g2 (PPARg2) binding site, which is re-reaction and electrophoresis were as described previously (Klemm et
quired for the expression of the gene in adipose tissue (Tonto-al. 1990).noz et al. 1995). Additional stretches of sequence identityTranscriptional activity. The transcriptional activity of specificbetween the rat and chicken PEPCK-C promoters are shortsegments of the PEPCK-C promoter was determined by stably(usually 4 base pairs or less) and dispersed throughout thetransfecting chimeric genes into either LMH cells (chicken) or H4
cells (rat) or by determining the level of transcription after transient entire promoter.
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from
279TRANSCRIPTIONAL REGULATION OF THE GENE FOR AVIAN PEPCK
labeled on the 5* end of the coding strand at the NarI siteand extended to the EcoRI site in the pSCAN poly linker.The undigested probe was 646 bases long (see lane 1), whereasthe protected fragment was 272 bases in length. This corre-
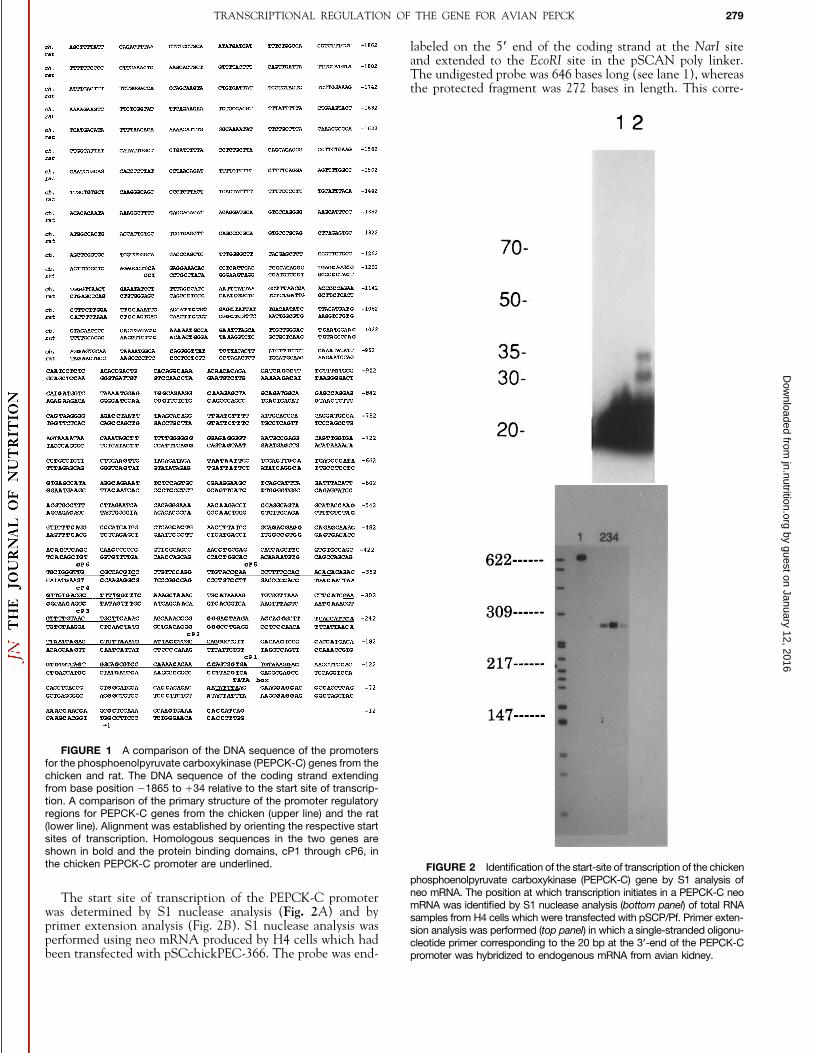
FIGURE 1 A comparison of the DNA sequence of the promotersfor the phosphoenolpyruvate carboxykinase (PEPCK-C) genes from thechicken and rat. The DNA sequence of the coding strand extendingfrom base position 01865 to /34 relative to the start site of transcrip-tion. A comparison of the primary structure of the promoter regulatoryregions for PEPCK-C genes from the chicken (upper line) and the rat(lower line). Alignment was established by orienting the respective startsites of transcription. Homologous sequences in the two genes areshown in bold and the protein binding domains, cP1 through cP6, inthe chicken PEPCK-C promoter are underlined. FIGURE 2 Identification of the start-site of transcription of the chicken
phosphoenolpyruvate carboxykinase (PEPCK-C) gene by S1 analysis ofneo mRNA. The position at which transcription initiates in a PEPCK-C neoThe start site of transcription of the PEPCK-C promotermRNA was identified by S1 nuclease analysis (bottom panel) of total RNAwas determined by S1 nuclease analysis (Fig. 2A) and by samples from H4 cells which were transfected with pSCP/Pf. Primer exten-
primer extension analysis (Fig. 2B). S1 nuclease analysis was sion analysis was performed (top panel) in which a single-stranded oligonu-performed using neo mRNA produced by H4 cells which had cleotide primer corresponding to the 20 bp at the 3*-end of the PEPCK-Cbeen transfected with pSCchickPEC-366. The probe was end- promoter was hybridized to endogenous mRNA from avian kidney.
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from
280 SAVON ET AL.
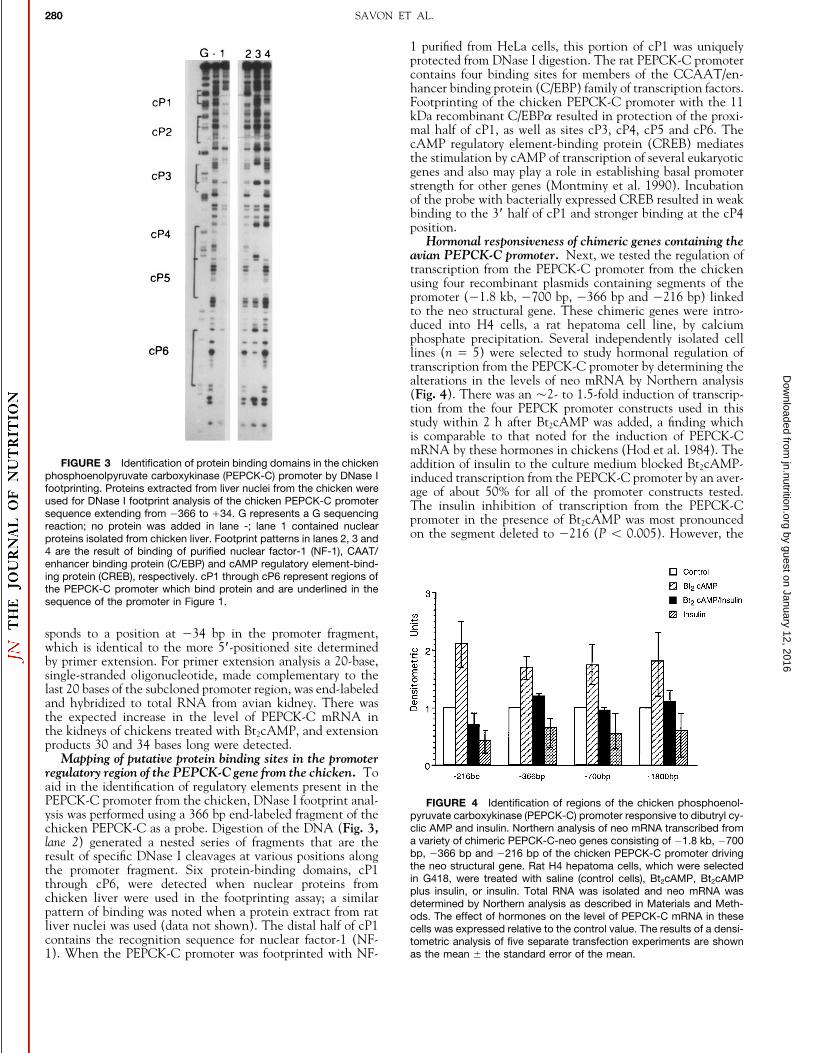
1 purified from HeLa cells, this portion of cP1 was uniquelyprotected from DNase I digestion. The rat PEPCK-C promotercontains four binding sites for members of the CCAAT/en-hancer binding protein (C/EBP) family of transcription factors.Footprinting of the chicken PEPCK-C promoter with the 11kDa recombinant C/EBPa resulted in protection of the proxi-mal half of cP1, as well as sites cP3, cP4, cP5 and cP6. ThecAMP regulatory element-binding protein (CREB) mediatesthe stimulation by cAMP of transcription of several eukaryoticgenes and also may play a role in establishing basal promoterstrength for other genes (Montminy et al. 1990). Incubationof the probe with bacterially expressed CREB resulted in weakbinding to the 3* half of cP1 and stronger binding at the cP4position.
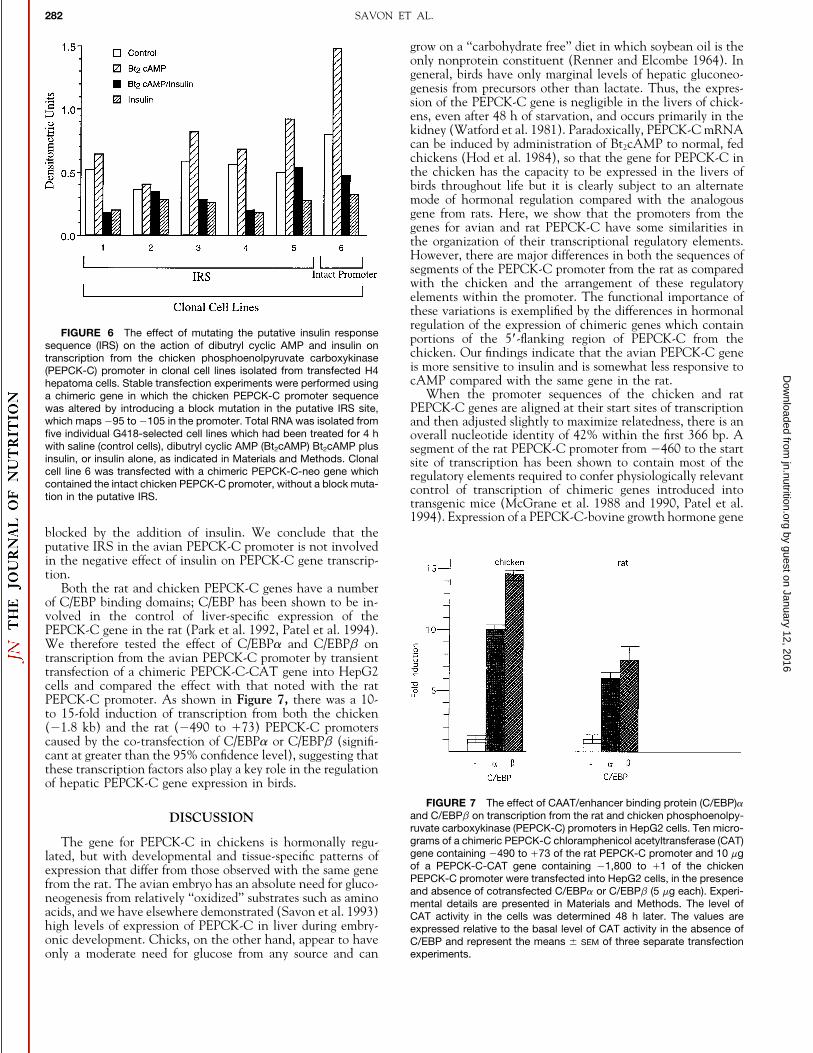
Hormonal responsiveness of chimeric genes containing theavian PEPCK-C promoter. Next, we tested the regulation oftranscription from the PEPCK-C promoter from the chickenusing four recombinant plasmids containing segments of thepromoter (01.8 kb, 0700 bp, 0366 bp and 0216 bp) linkedto the neo structural gene. These chimeric genes were intro-duced into H4 cells, a rat hepatoma cell line, by calciumphosphate precipitation. Several independently isolated celllines (n Å 5) were selected to study hormonal regulation oftranscription from the PEPCK-C promoter by determining thealterations in the levels of neo mRNA by Northern analysis(Fig. 4). There was an Ç2- to 1.5-fold induction of transcrip-tion from the four PEPCK promoter constructs used in thisstudy within 2 h after Bt2cAMP was added, a finding whichis comparable to that noted for the induction of PEPCK-CmRNA by these hormones in chickens (Hod et al. 1984). The
FIGURE 3 Identification of protein binding domains in the chicken addition of insulin to the culture medium blocked Bt2cAMP-phosphoenolpyruvate carboxykinase (PEPCK-C) promoter by DNase I induced transcription from the PEPCK-C promoter by an aver-footprinting. Proteins extracted from liver nuclei from the chicken were age of about 50% for all of the promoter constructs tested.used for DNase I footprint analysis of the chicken PEPCK-C promoter The insulin inhibition of transcription from the PEPCK-Csequence extending from 0366 to /34. G represents a G sequencing promoter in the presence of Bt2cAMP was most pronouncedreaction; no protein was added in lane -; lane 1 contained nuclear
on the segment deleted to 0216 (P õ 0.005). However, theproteins isolated from chicken liver. Footprint patterns in lanes 2, 3 and4 are the result of binding of purified nuclear factor-1 (NF-1), CAAT/enhancer binding protein (C/EBP) and cAMP regulatory element-bind-ing protein (CREB), respectively. cP1 through cP6 represent regions ofthe PEPCK-C promoter which bind protein and are underlined in thesequence of the promoter in Figure 1.
sponds to a position at 034 bp in the promoter fragment,which is identical to the more 5*-positioned site determinedby primer extension. For primer extension analysis a 20-base,single-stranded oligonucleotide, made complementary to thelast 20 bases of the subcloned promoter region, was end-labeledand hybridized to total RNA from avian kidney. There wasthe expected increase in the level of PEPCK-C mRNA inthe kidneys of chickens treated with Bt2cAMP, and extensionproducts 30 and 34 bases long were detected.
Mapping of putative protein binding sites in the promoterregulatory region of the PEPCK-C gene from the chicken. Toaid in the identification of regulatory elements present in thePEPCK-C promoter from the chicken, DNase I footprint anal- FIGURE 4 Identification of regions of the chicken phosphoenol-ysis was performed using a 366 bp end-labeled fragment of the pyruvate carboxykinase (PEPCK-C) promoter responsive to dibutryl cy-
clic AMP and insulin. Northern analysis of neo mRNA transcribed fromchicken PEPCK-C as a probe. Digestion of the DNA (Fig. 3,a variety of chimeric PEPCK-C-neo genes consisting of 01.8 kb, 0700lane 2) generated a nested series of fragments that are thebp, 0366 bp and 0216 bp of the chicken PEPCK-C promoter drivingresult of specific DNase I cleavages at various positions alongthe neo structural gene. Rat H4 hepatoma cells, which were selectedthe promoter fragment. Six protein-binding domains, cP1in G418, were treated with saline (control cells), Bt2cAMP, Bt2cAMPthrough cP6, were detected when nuclear proteins fromplus insulin, or insulin. Total RNA was isolated and neo mRNA waschicken liver were used in the footprinting assay; a similar determined by Northern analysis as described in Materials and Meth-
pattern of binding was noted when a protein extract from rat ods. The effect of hormones on the level of PEPCK-C mRNA in theseliver nuclei was used (data not shown). The distal half of cP1 cells was expressed relative to the control value. The results of a densi-contains the recognition sequence for nuclear factor-1 (NF- tometric analysis of five separate transfection experiments are shown
as the mean { the standard error of the mean.1). When the PEPCK-C promoter was footprinted with NF-
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from

281TRANSCRIPTIONAL REGULATION OF THE GENE FOR AVIAN PEPCK
inhibitory effect of insulin was significant at the 95% confi- from 0140 to 049 bp in the promoter sequence from thechicken was highly homologous to an insulin response se-dence level for all of the segments of the PEPCK-C promoter
with the exception of 01.8 kb, where there was considerable quence (IRS) identified (O’Brien and Granner 1991; O’Brienet al. 1990) in the PEPCK-C gene from the rat. Consequently,variability in the response to insulin. The addition of insulin
also decreased the basal level of transcription of the PEPCK- a pSCAN-based chimeric gene containing a block mutationmade in this location of the chicken PEPCK-C promoter wasneo gene in all four of the PEPCK-C promoter constructs
studied but was significant (P õ 0.05) only for the promoter created and introduced into H4 cells by transfection, followedby selection with G418. An analysis of five individual clonesdeleted to 0216 and 0700. By way of comparison, the expres-
sion of the analogous rat PEPCK-C-neo gene, while retaining of cells expressing this recombinant plasmid (Fig. 6) indicatedno loss of transcriptional responsiveness of the modifiedboth cAMP and dexamethasone responsiveness, does not show
the expected inhibition when treated with insulin (Wynshaw- PEPCK-C promoter to insulin; we noted an average reductionof neo mRNA levels caused by insulin of at least 60% in theBoris et al. 1986). As a control for the effectiveness of the
hormonal treatments, the Northern blot with a cDNA to five clonal cell lines tested. One of the clonal cell lines (line2) did not respond to any treatment, and others (cell lines 1PEPCK-C from the rat was rehybridized with RNA from the
H4 cells, and the stimulatory response of the endogenous rat and 4) demonstrated only a marginal stimulation by Bt2cAMP.In addition, Bt2cAMP-stimulated transcription from thePEPCK-C gene to hormones was as expected (data not
shown). PEPCK-C promoter in all of the clonal cell lines, except line2, was markedly inhibited by insulin addition. Clonal cell lineNext, we compared the effects of Bt2cAMP and insulin on
transcription from the PEPCK-C promoter from the chicken 6 served as a control cell line and was transfected with achimeric gene containing the unmodified segment of the avianand the rat in chicken (LMH) and mammalian (HepG2) hepa-
toma cells by transient transfection. The transcriptional re- PEPCK-C promoter linked to the neo structural gene. In thesecells, we noted the expected increase in transcription from thesponse of the avian PEPCK-C promoter linked to the CAT
structural gene was determined in LMH cells using the same PEPCK-C promoter after the addition of Bt2cAMP, which wasseries of deletions used in the experiments reported in Figure4 (01.8 kb, 0700 bp, 0366 bp and 0216 bp) (Fig. 5A).The 700 bp segment of the PEPCK-C promoter gave the bestresponse to the addition of Bt2cAMP; it was induced threefoldby the cyclic nucleotide from the control value (P õ 0.001).Insulin blocked the induction by Bt2cAMP (P õ 0.05) butdid not lower the basal level of transcription from the avianPEPCK-C promoter in LMH cells. The other segments of thePEPCK-C promoter tested for their transcriptional respon-siveness were less markedly induced by Bt2cAMP. The addi-tion of insulin to the LMH cells did not cause a significantdecrease (at the 95% confidence level) in the ability ofBt2cAMP to induce transcription from segments of the avianPEPCK-C promoter shorter than 0700, an effect which maybe due to the low level of transcriptional induction byBt2cAMP on these segments of the promoter. The reason forthis difference in response between the regions of the chickenPEPCK-C promoter is not clear, but it is possible that thereare critical transcriptional regulatory elements between 0700and 0366 which are required for the full response of thePEPCK-C promoter in the LMH cell. Finally, insulin had nosignificant effect (95% confidence level) on basal transcription FIGURE 5 A comparison of transcriptional regulation from thefrom the PEPCK-C promoter in LMH cells. chicken and rat phosphoenolpyruvate carboxykinase (PEPCK-C) pro-
The rat PEPCK-C promoter (from 0490 to /73), when moter by dibutryl cyclic AMP and insulin in chicken and mammalianintroduced into LMH cells, responded to Bt2cAMP with a hepatoma cells. (A) LMH cells were transfected with chimeric genes
containing the truncated chicken PEPCK-C promoter (01.8 kb to 0216threefold induction in transcription, which was only partiallybp), which was ligated to the structural gene for chloramphenicol acetyl-inhibited by insulin (significant at the 95% confidence level)transferase (CAT) or (insert) the rat PEPCK-C promoter (0490 to /73),(Fig. 5A, insert). As with the avian PEPCK-C promoter, therewhich was also ligated to the CAT structural gene. The activity of CATwas no effect of insulin on the basal level of transcription fromwas determined 4 h after the addition of saline (control cells) dibutrylthe rat PEPCK-C promoter in LMH cells. We next tested thecyclic AMP (Bt2cAMP), Bt2cAMP plus insulin, or insulin alone, as indi-effect of the hormones on a chimeric PEPCK-C-CAT gene cated in Materials and Methods. (B) HepG2 cells were transfected with
containing the avian or rat PEPCK-C promoters, which were a chimeric gene containing 01.8 kb of the promoter from the chickentransfected into HepG2 cells, a mammalian hepatoma cell PEPCK-C gene linked to the structural gene for CAT or (C) a chimericline. Transcription from the avian PEPCK-C promoter was gene containing 0490 to /73 of the rat PEPCK-C gene linked to the
structural gene for CAT. The activity of CAT was determined 4 h afternot stimulated by Bt2cAMP in these cells, and the basal leveltreatment with saline (control cells), Bt2cAMP, Bt2cAMP plus insulin, orof transcription was not inhibited by insulin; in addition, theinsulin alone, as indicated in Materials and Methods. The values areoverall level of transcription from the chicken PEPCK-C pro-expressed as fold induction relative to the control (saline addition) cellsmoter was very low in HepG2 cells (Fig. 5B). This contrastsand were corrected for the relative activity of b-galactosidase. Thewith the chimeric PEPCK-C-CAT gene containing the values presented in this figure are the average of at least three separate
PEPCK-C promoter from the rat which was induced 10-fold transfection experiments and are expressed as the mean { SEM of theby Bt2cAMP but was not responsive to insulin (Fig. 5C). CAT activity in the cells, except for the values for the rat PEPCK-C-
A comparison of the 5*-flanking PEPCK-C sequences from CAT gene which represent the average of two separate transfectionexperiments.the chicken and the rat suggested that the segment extending
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from
282 SAVON ET AL.
grow on a ‘‘carbohydrate free’’ diet in which soybean oil is theonly nonprotein constituent (Renner and Elcombe 1964). Ingeneral, birds have only marginal levels of hepatic gluconeo-genesis from precursors other than lactate. Thus, the expres-sion of the PEPCK-C gene is negligible in the livers of chick-ens, even after 48 h of starvation, and occurs primarily in thekidney (Watford et al. 1981). Paradoxically, PEPCK-C mRNAcan be induced by administration of Bt2cAMP to normal, fedchickens (Hod et al. 1984), so that the gene for PEPCK-C inthe chicken has the capacity to be expressed in the livers ofbirds throughout life but it is clearly subject to an alternatemode of hormonal regulation compared with the analogousgene from rats. Here, we show that the promoters from thegenes for avian and rat PEPCK-C have some similarities inthe organization of their transcriptional regulatory elements.However, there are major differences in both the sequences ofsegments of the PEPCK-C promoter from the rat as comparedwith the chicken and the arrangement of these regulatoryelements within the promoter. The functional importance ofthese variations is exemplified by the differences in hormonalregulation of the expression of chimeric genes which contain
FIGURE 6 The effect of mutating the putative insulin response portions of the 5*-flanking region of PEPCK-C from thesequence (IRS) on the action of dibutryl cyclic AMP and insulin on chicken. Our findings indicate that the avian PEPCK-C genetranscription from the chicken phosphoenolpyruvate carboxykinase
is more sensitive to insulin and is somewhat less responsive to(PEPCK-C) promoter in clonal cell lines isolated from transfected H4cAMP compared with the same gene in the rat.hepatoma cells. Stable transfection experiments were performed using
When the promoter sequences of the chicken and rata chimeric gene in which the chicken PEPCK-C promoter sequencePEPCK-C genes are aligned at their start sites of transcriptionwas altered by introducing a block mutation in the putative IRS site,
which maps095 to0105 in the promoter. Total RNA was isolated from and then adjusted slightly to maximize relatedness, there is anfive individual G418-selected cell lines which had been treated for 4 h overall nucleotide identity of 42% within the first 366 bp. Awith saline (control cells), dibutryl cyclic AMP (Bt2cAMP) Bt2cAMP plus segment of the rat PEPCK-C promoter from 0460 to the startinsulin, or insulin alone, as indicated in Materials and Methods. Clonal site of transcription has been shown to contain most of thecell line 6 was transfected with a chimeric PEPCK-C-neo gene which regulatory elements required to confer physiologically relevantcontained the intact chicken PEPCK-C promoter, without a block muta- control of transcription of chimeric genes introduced intotion in the putative IRS. transgenic mice (McGrane et al. 1988 and 1990, Patel et al.
1994). Expression of a PEPCK-C-bovine growth hormone geneblocked by the addition of insulin. We conclude that theputative IRS in the avian PEPCK-C promoter is not involvedin the negative effect of insulin on PEPCK-C gene transcrip-tion.
Both the rat and chicken PEPCK-C genes have a numberof C/EBP binding domains; C/EBP has been shown to be in-volved in the control of liver-specific expression of thePEPCK-C gene in the rat (Park et al. 1992, Patel et al. 1994).We therefore tested the effect of C/EBPa and C/EBPb ontranscription from the avian PEPCK-C promoter by transienttransfection of a chimeric PEPCK-C-CAT gene into HepG2cells and compared the effect with that noted with the ratPEPCK-C promoter. As shown in Figure 7, there was a 10-to 15-fold induction of transcription from both the chicken(01.8 kb) and the rat (0490 to /73) PEPCK-C promoterscaused by the co-transfection of C/EBPa or C/EBPb (signifi-cant at greater than the 95% confidence level), suggesting thatthese transcription factors also play a key role in the regulationof hepatic PEPCK-C gene expression in birds.
FIGURE 7 The effect of CAAT/enhancer binding protein (C/EBP)aand C/EBPb on transcription from the rat and chicken phosphoenolpy-DISCUSSIONruvate carboxykinase (PEPCK-C) promoters in HepG2 cells. Ten micro-grams of a chimeric PEPCK-C chloramphenicol acetyltransferase (CAT)The gene for PEPCK-C in chickens is hormonally regu-gene containing 0490 to /73 of the rat PEPCK-C promoter and 10 mglated, but with developmental and tissue-specific patterns ofof a PEPCK-C-CAT gene containing 01,800 to /1 of the chickenexpression that differ from those observed with the same genePEPCK-C promoter were transfected into HepG2 cells, in the presencefrom the rat. The avian embryo has an absolute need for gluco-and absence of cotransfected C/EBPa or C/EBPb (5 mg each). Experi-neogenesis from relatively ‘‘oxidized’’ substrates such as amino mental details are presented in Materials and Methods. The level of
acids, and we have elsewhere demonstrated (Savon et al. 1993) CAT activity in the cells was determined 48 h later. The values arehigh levels of expression of PEPCK-C in liver during embry- expressed relative to the basal level of CAT activity in the absence ofonic development. Chicks, on the other hand, appear to have C/EBP and represent the means { SEM of three separate transfection
experiments.only a moderate need for glucose from any source and can
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from

283TRANSCRIPTIONAL REGULATION OF THE GENE FOR AVIAN PEPCK
tions. Both the CREB and C/EBPa can bind within or nearthe cP1 region of the chicken PEPCK-C promoter, althoughCREB binds only weakly to this region of the PEPCK promoter(Fig. 3). Interestingly, the cP1 and cP3 sites have an almostperfect consensus C/EBP binding domain and there is strongbinding of C/EBPa to the cP3 site, suggesting that C/EBP maybe involved in cAMP regulation of PEPCK-C gene transcrip-tion. The role of C/EBPa in mediating the action of cAMPon transcription of the gene for PEPCK from the rat has beensuggested from previous studies (Park et al. 1990) and directlydemonstrated more recently (Roesler et al. 1996). Figure 7demonstrates that both C/EBPa and C/EBPb can stimulatetranscription from the avian PEPCK promoter. In addition,preliminary work in our laboratory has demonstrated that theFIGURE 8 A comparison of transcriptional regulatory elements indeletion of the gene for C/EBPa greatly diminishes the induc-the chicken and the rat phosphoenolpyruvate carboxykinase (PEPCK-tion of PEPCK-C gene expression by cAMP in the livers ofC) genes. Putative transcriptional regulatory elements and transcriptionfetal rats (C. Croniger, V. Poli, G. Darlington and R. Hanson,factor binding sites, identified by protein binding and/or sequence con-personal communication, Case Western Reserve University).servation are compared in the rat (P1 through P4) and chicken PEPCK-
C genes (cP1 through cP4). The abbreviations for the elements and Region 2 of the PEPCK promoter from the rat contains anfactors are: CREB, cAMP regulatory element-binding protein; C/EBP, HNF-1 site which has been shown to be critical for transcrip-CAAT/enhancer binding protein; FOS/JUN, FOS/JUN binding domain; tion of the PEPCK gene in the kidney (Patel et al. 1994). ANF1, nuclear factor-1; HNF-1, hepatic nuclear factor-1; IRE, insulin similar element lies upstream the CRE/NF-1 site in theresponsive element; GRE, glucocorticoid responsive element; T3, thy- PEPCK-C promoter from the chicken. Whether the putativeroid hormone responsive element. hepatic nuclear factor (HNF)-1 site in the avian PEPCK-C
gene is involved in the regulation of PEPCK-C gene transcrip-tion in the kidney remains to be determined. In addition,containing a segment of the rat PEPCK-C promoter from0460Region 2 of the rat PEPCK-C promoter contains a C/EBPto /73, introduced into the germ line of mice, occurs in abinding domain (Park et al. 1990 and 1992) and a thyroidtissue-specific manner, is induced appropriately by diet andhormone regulatory element (Giralt et al. 1991). In the rat,hormones and develops with the same pattern as the endoge-the C/EBP binding sites have been shown to be required fornous PEPCK-C gene (McGrane et al. 1990). The pattern ofthe full transcriptional responsiveness of the promoter totranscriptional regulatory elements in the PEPCK-C promotercAMP (Liu et al. 1991). The PEPCK-C gene from the chickenfrom the rat has been well established (see Hanson and Patelhas a cluster of C/EBP binding sites in a similar region of the1994 for a review); the relative positions of the putative regula-promoter, suggesting a key role for this transcription factor intory elements in the PEPCK-C promoter from the chickenthe control of PEPCK-C in the avian liver. It has recentlydiffer significantly from the locations of these same sequencesbeen shown that C/EBPa is necessary for the expression ofin the PEPCK-C promoter from the rat (Fig. 8). Interestingly,PEPCK in the livers of mice in the perinatal period (Wang etthe promoters for the mouse and the human PEPCK genesal. 1995).have recently been sequenced and shown to be virtually identi-
Region 3 of the PEPCK promoter from the rat is a complexcal to the sequence for the rat, suggesting that the significantdomain with several regulatory elements. Granner and col-divergence in the chicken PEPCK gene relates to the specificleagues (Imai et al. 1990 and 1993) have described a glucocor-metabolic function of PEPCK in birds and to the observedticoid regulatory unit (GRU) in this area of the PEPCK-Cdifferences in hormonal regulation noted in this study.promoter, within which there is a glucocorticoid regulatoryThe rat PEPCK-C promoter contains three regions whichelement (GRE), an insulin regulatory element (IRS) and twohave clusters of regulatory elements. Region 1 contains theaccessory protein binding domains. The analogous region ofCRE and adjacent NF-1 site (P-1). A mutation in the CREthe PEPCK-C promoter in the chicken, which extends frommarkedly reduces the cAMP responsiveness of the PEPCK-C0359 to 0345, contains considerable homology with the GREpromoter in the livers of transgenic mice, whereas a blockconsensus sequence; in addition, its position within the pro-mutation in the NF-1 binding site causes a marked increasemoter is very similar to the comparable position of the GREin the level of basal gene transcription (Patel et al. 1994).in the promoter of the rat PEPCK-C gene. In preliminaryThe same type of response was noted when the PEPCK pro-studies in which the gene was stably transfected into H4 hepa-moter was transfected into hepatoma cells in culture (Liu ettoma cells, we noted that the removal of the putative GREal. 1991). In addition, unpublished work from our laboratoryfrom the chicken PEPCK-C promoter by truncation to 0216has demonstrated that NF-1 has a powerful negative effect onresulted in a loss of the normal twofold induction of transcrip-transcription from the PEPCK-C promoter in the same celltion from the promoter by glucocorticoids (S. Savon, P. Ha-system (D. Crawford, P. Lehey, R. Gronostajski, and R. Han-kimi and R. Hanson, personal communication, Case Westernson, personal communication, Case Western Reserve Univer-Reserve University, Cleveland, OH).sity). The CRE from the PEPCK promoter will bind members
The physiological role of insulin as a negative regulator ofof the leucine zipper family of transcription factors, includingPEPCK-C gene transcription is most likely a critical factor inCREB (Yamamoto et al. 1988), C/EBPa (Roesler et al. 1989),the control of glucose homeostasis in birds. The regulation ofC/EBPb (Park et al. 1992), D-binding protein (Roesler et al.PEPCK gene transcription by insulin is clearly very complex;1992) and fos/jun heterodimers (Gurney et al. 1992).there is no single regulatory element which has been impli-The region of the PEPCK-C promoter from the chickencated in the powerful negative effect of this hormone on thewhich contains the most sequence identity with the PEPCK-transcription of the PEPCK-C gene in the liver. The IRSC promoter from the rat also has a CRE adjacent to an NF-1found within the GRU of the rat PEPCK-C promoter, whichbinding site. The cP1 region in the chicken PEPCK-C pro-maps between 0416 to 0407, accounts for only half of themoter maps between078 and085 bp and contains an octamer
which differs from the consensus CRE sequence in two posi- inhibitory effect of insulin of transcription from the PEPCK-
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from

284 SAVON ET AL.
C promoter in hepatoma cells (O’Brien et al. 1990). Because a high level of basal transcription but there was no regulationthe effect of insulin on PEPCK-C gene transcription most by hormones. However, a similar chimeric gene containinglikely involves an interaction between this hormone and the the promoter for PEPCK-C from the rat was regulated in theCRE of the promoter (which maps between 087 to 074) a expected manner by Bt2cAMP and insulin (Fig. 5). Thus, itmore proximal binding site for transcription factors involved is probable that factors required for the control of avianin this process has been suggested. Most of the work done to PEPCK-C gene transcription are missing from HepG2 cells.date has involved the distal protein binding site; the proximal Segments of the PEPCK-C promoter from01.8 kb to0216site has not yet been identified. The only available data on kb respond almost identically to the administration of hor-the protein(s) which bind to the IRS and the CRE arise from mones when stably transfected into H4 cells (Fig. 4). Thisa recent study by O’Brien et al. (1994) which demonstrated suggests that the regulation of the PEPCK-C gene by cAMPthat C/EBPa, either alone or complexed with an IRS binding and insulin resides within the first 210 bp from the start-siteprotein, termed p20-C/IBP, can gel shift oligonucleotides cor- of gene transcription. This region of the promoter contains aresponding to both the CRE from the PEPCK promoter and CRE adjacent to an NF-1 binding domain, a putative IRS andthe distal IRS. an hepatic nuclear factor-1 (HNF-1) binding site. However,
It is of interest to note that the sequence between0149 and when the same chimeric genes containing the PEPCK-C pro-0140 in the PEPCK-C promoter from the chicken contains a moter deletions were transiently transfected into chickenclose match (9/10) with the consensus IRS found at positions LMH cells, the segment of the promoter from 0700 to /10411 to 0402 in the rat PEPCK-C promoter (O’Brien and demonstrated the most marked activity and was most respon-Granner 1991). This sequence could be a candidate for the sive to hormonal treatment. The reason for this difference inproximal IRS discussed above. Based on the possibility that response is not immediately apparent, but may reflect inherentinsulin sensitivity of the avian PEPCK-C gene is dependent differences between transient and stable DNA transfectionon this sequence, a recombinant chimeric gene containing a techniques.10 bp block mutation corresponding to this area was tested Comparison of hormonal responsiveness of PEPCK-C-CATfor its transcriptional responsiveness to insulin (Fig. 6). It was chimeric genes comprised of promoter sequences from thefound, however, to remain insulin responsive, indicating that chicken or rat, and transiently transfected into a chicken hepa-the relative position of responsive elements both with respect tocyte cell line (LMH cells), further suggests that differencesto each other and with the TATA box has functional signifi- in regulation are at least partially due to differences in tran-cance in regulating gene transcription (Hanson and Patel scription response elements present in the two promoter regu-1994). latory regions (Fig. 5). When introduced into chicken cells,Insulin inhibits transcription from the chicken PEPCK-C transcription of chimeric genes containing promoter sequencespromoter; this contrasts with the absence of such an effect on
from the chicken is inhibited by insulin. Chimeric genes con-transcription from PEPCK-C-neo fusion genes made with rattaining segments of the PEPCK-C promoter from the rat whensequences observed previously in our laboratory (Wynshaw-transfected into chicken cells (Fig. 5), on the other hand,Boris et al. 1986). To demonstrate the effect of insulin onelicit hormonal responses quantitatively more typical of thistranscription from the rat PEPCK promoter in transformedgene when studied in rat cells. In fact, the PEPCK-C promotercells in culture, it has been necessary to carry out more elabo-from the chicken did not respond to hormonal treatment whenrate types of selection procedures. For example, it has not beenintroduced transiently into HepG2 cells, further emphasizingpossible to date to demonstrate a marked negative effect ofthe presence of functionally important differences in transcrip-insulin on transcription when the PEPCK promoter is intro-tional regulatory elements between these two promoters, differ-duced by transient transfection (see Fig. 5C for an example).ences which may result from an altered dependence on chro-The procedure used to identify the IRS in the rat PEPCK-mosomal orientation within the nucleus.promoter (stable transfection in H4 cells) failed to demon-
Patel et al. (1994) used transgenic mice which containedstrate the dramatic inhibitory effects of insulin noted for thechimeric genes in which the promoter for PEPCK-C gene fromendogenous PEPCK-C gene in the same cells (O’Brien et al.the rat contained block mutations in regulatory elements, to1990). In the present study, insulin decreased Bt2cAMP stimu-determine their role in the tissue-specific expression and hor-lated transcription from the avian PEPCK-C promoter andmonal regulation of PEPCK gene expression. When the CREcharacteristically reduced basal gene expression by at leastwas removed by mutation, the level of expression of the50%, suggesting a substantial difference in the transcriptionaltransgene increased four- to fivefold, whereas transcriptionalresponse to insulin by the PEPCK-C gene from the rat andstimulation by cAMP was completely abolished. This findingchicken.suggests that protein binding to the CRE acts to maintain lowA comparison of the hormonal responsiveness of PEPCK-levels of PEPCK gene transcription in the absence of cAMP.C-neo chimeric genes transfected into rat hepatoma cell linesIt is probable that the CRE site of the PEPCK promoter isdemonstrates both the similarities and differences between thealways occupied by proteins in the cell because in vivo foot-promoter regulatory regions of the chicken and rat PEPCK-C
genes. Although cAMP induces transcription from the printing of the promoter using hepatoma cells has indicatedthe presence of a protected region at the CRE, which doesPEPCK-C promoter from the rat four- to sixfold (Liu et al.
1991, Wynshaw-Boris et al. 1984), transcription from the not require added cAMP for protein binding to the promoter(Faber et al. 1993). In addition, NF-1 strongly inhibits tran-chicken PEPCK-C promoter is stimulated only twofold. This
reduced sensitivity to cAMP may be related to the blunted scription from the PEPCK promoter when the genes for NF-1 and PEPCK-CAT are co-transfected into HepG2 hepatomaresponse of the PEPCK-C gene to the high circulating levels
of glucagon in the blood of chickens. Larger concentrations cells (D. Crawford, P. Lehey, R. Gronostajski and R. Hanson,personal communication, Case Western Reserve University).of Bt2cAMP are required to induce the expression of hepatic
PEPCK-C in the chicken, which may explain why there is an It is possible that transcription of the gene for PEPCK-C in thechicken is maintained at a low level due to the arrangement ofobserved lack of induction of PEPCK-C gene expression in
the livers of starved chickens (Hod et al. 1984). Interestingly, the NF-1 site relative to the CREB binding site(s), whichmarkedly differs from the distribution of these elements foundwhen a chimeric gene containing the avian PEPCK-C pro-
moter was transiently transfected into HepG2 cells, there was in the promoter for PEPCK in the rat. Therefore, only an
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from

285TRANSCRIPTIONAL REGULATION OF THE GENE FOR AVIAN PEPCK
Liu, J. & Hanson, R. W. (1991) Regulation of P-enolpyruvate carboxykinaseelevation in the concentration of cAMP above the normalgene transcription. Mol. Cell. Biochem. 104: 89–100.physiological levels will result in the transcription of the Liu, J., Park, E. A., Gurney, A. L., Roesler, W. J. & Hanson, R. W. (1991) Cyclic
PEPCK-C gene in the livers of chickens. Why the gene is AMP induction of P-enolpyruvate carboxykinase gene transcription is medi-ated by multiple promoter elements. J. Biol. Chem. 266: 19095–19102.expressed in the livers of birds before hatching is an interesting
Magnunson, M. A., Quinn, P. A. & Granner, D. K. (1987) Multi hormonal regula-question which remains to be answered. Further investigation tion of P-enolpyruvate carboxykinase-chloramphenicol acetyltransferase fu-of the differences in the hormonal response regions between sion genes. J. Biol. Chem. 262: 14917–14920.
McGrane, M. M., deVente, J., Yun, J., Bloom, J., Park, E. A., Wynshaw-Boris, A.,these species will not only provide insight into the functionalWagner, T., Rottman, F. M. & Hanson, R. W. (1988) Tissue-specific expres-alterations operating in the PEPCK-C gene from the chicken,sion and dietary regulation of a chimeric PEPCK/bGH gene in transgenic
but also increase understanding of how a convergence of influ- mice. J. Biol. Chem. 263: 11443–11451.McGrane, M. M., Yun, J. S., Moorman, A.F.M., Lamers, W. H., Hendrick, G. K.,ences provided by discrete responsive elements results in spe-
Arafah, B. M., Park, E. A., Wagner, T. E. & Hanson, R. W. (1990) Metaboliccifically regulated transcriptional activity.effects of developmental, tissue and cell specific expression of a chimericPEPCK/bGH gene in transgenic mice. J. Biol. Chem. 265: 22571–22379.
Montminy, M. R., Gonzalez, G. A. & Yamamoto, K. K. (1990) Characteristics ofthe cAMP response element. Recent Prog. Horm. Res. 46: 219–230.LITERATURE CITED
Nordlie, R. C. & Lardy, H. A. (1963) Mammalian liver P-enolpyruvate carboxyki-nase. J. Biol. Chem. 238: 2259–2263.Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Seidman,
O’Brien, R. M. & Granner, D. K. (1991) Regulation of gene expression by insu-J. A., Smith, J. A. & Struhl, K. (1987) In: Current Protocols in Molecularlin. Biochem. J. 278: 609–619.Biology, pp. 9.1.1–9.1.4. John Wiley and Sons, New York, NY.
O’Brien, R. M., Lucas, P. C., Forest, C. D., Magnunson, M. A. & Granner, D. A.Ballard, F. J. & Hanson, R. W. (1967) P-Enolpyruvate carboxykinase and py-(1990) Identification of a sequence in the P-enolpyruvate carboxykinaseruvate carboxylase in developing rat liver. Biochem. J. 104: 866–871.gene that mediates the negative effect of insulin on transcription. ScienceBokar, J. A., Roesler, W. J., Vandenbark, G. R., Kaetzel, D. M., Hanson, R. W., &(Washington, DC) 249: 533–537.Nilson, J. (1988) Characterization of the cAMP responsive elements from
O’Brien, R. M., Lucas, P. C., Yamasaki, T., Noisin, E. L. & Granner, D. K. (1994)the genes for the a-subunit of chorionic gonadotropin and P-enolpyruvatePotential convergence of insulin and cAMP signal transduction systems atcarboxykinase. Conserved features of nuclear protein binding between tis-the PEPCK gene promoter through C/EBP. J. Biol. Chem. 269: 30419–30428.sues and species. J. Biol. Chem. 263: 19740–19747.
Park, E. A., Gurney, A. L., Nizielski, S. E., Hakimi, P., Cao, Z., Moorman, A. &Chomczynski, P. & Sacchi, N. (1987) Single step method of RNA isolation byHanson, R. W. (1992) Relative role of members of the C/EBP family ofacid guanididinium thiocyanate-phenol-chloroform extraction. Anal. Biochem.transcription factors in controlling transcription of the gene for phosphoenol-162: 156–159.pyruvate carboxykinase. J. Biol. Chem. 267: 613–619.Crawford, D. R. & Hanson, R. W. (1993) Inhibition of P-enolpyruvate carboxyki-
Park, E. A., Roesler, W. J., Liu, J., Klemm, D. J., Gurney, A. & Hanson, R. W.nase (GTP) gene transcription by protein kinase. C. Life Sci. Advances 12:(1990) The role of CCAAT/enhancer binding protein (C/EBP) in the transcrip-165–170.tional regulation of the gene for P-enolpyruvate carboxykinase (GTP). Mol.Faber, S., O’Brien, R. M., Imai, E., Granner, D. K. & Chalkely, R. (1993) Dy-Cell. Biol. 10: 6264–6272.namic aspects of DNA/protein interactions in the transcriptional initiation com-
Patel, Y. M., Yun, J. S., Liu, J., McGrane, M. M. & Hanson, R. W. (1994) Anplex and the hormone-sensitive domains of the PEPCK promoter in vivo. J.analysis of regulatory elements in the PEPCK gene which are responsible forBiol. Chem. 268: 24976–24985.its tissue-specific expression and metabolic control in transgenic mice. J.Giralt, M., Park, A. E., Liu, J., Gurney, A. & Hanson, R. W. (1991) IdentificationBiol. Chem. 269: 5619–5628.of a thyroid hormone response element in the P-enolpyruvate carboxykinase
Renner, R. & Elcombe, A. M. (1964) Factors affecting the utilization of ‘‘carbo-(GTP) gene: evidence for a synergistic interaction between thyroid hormonehydrate-free’’ diets in the chick. J. Nutr. 84: 327–330.and cAMP cis regulatory elements. J. Biol. Chem. 266: 21991–21996.
Roesler, W. J., Crosson, S. M., Vinson, C. & McFie, P. J. (1996) The a-isoformGorman, C. M., Moffat, L. F. & Howard, B. H. (1982) Recombinant genomesof C/EBP is required for mediating cAMP responsiveness of the P-enolpyru-which express chloramphenicol acetyltransferase in mammalian cells. Mol.vate carboxykinase promoter in hepatoma cells. J. Biol. Chem. 271: 8068–Cell. Biol. 2: 1044–1051. 8074.Gorski, K., Carneino, M. & Schibler, U. (1986) Tissue-specific in vitro transcrip- Roesler, W. J., McFie, P. J. & Dauvin, C. (1992) The liver-enriched transcription
tion from the mouse albumin promoter. Cell 47: 767–776. factor D-site binding protein activates the promoter of the P-enolpyruvateGurney, A. L., Park, E. A., Giralt, M., Liu, J. & Hanson, R. W. (1992) Opposing carboxykinase gene in hepatoma cells. J. Biol. Chem. 267: 21235–21243.
actions of Fos and Jun on transcription of the PEPCK (GTP) gene. Dominant Roesler, W. J., Vandenbark, G. R. & Hanson, R. W. (1989) Identification of mul-negative regulation by Fos. J. Biol. Chem. 267: 18133–18139. tiple protein-binding domains in the promoter-regulatory region of the P-
Hanson, R. W. & Garber, A. J. (1972) P-Enolpyruvate carboxykinase: I. Its role enolpyruvate carboxykinase (GTP) gene. J. Biol. Chem. 264: 9657–9664.in gluconeogenesis. Am. J. Clin. Nutr. 25: 1010–1021. Savon, S., Hakimi, P. & Hanson, R. W. (1993) Expression of the genes for the
Hanson, R. W. & Patel, Y. M. (1994) P-Enolpyruvate carboxykinase: the gene mitochondrial and cytosolic forms of PEPCK in avian liver during develop-and the enzyme. In: Advances in Enzymology (Meister, A., ed.), pp. 203–281. ment. Biol. Neonate 64: 62–68.John Wiley and Sons, New York, NY. Shen, C. S. & Mistry, S. P. (1978a) Development of gluconeogenic, glycolytic
Hod, Y., Cook, J. S., Weldon, S. L., Short, J. M., Wynshaw-Boris, A. & Hanson, and pentose-shunt enzymes in chicken kidney. Poult. Sci. 58: 663–667.R. W. (1986) Differential expression of the genes for the mitochondrial and Shen, C. S. & Mistry, S. P. (1978b) Effect of fasting and fasting and refeedingcytosolic forms of P-enolpyruvate carboxykinase. Ann. N. Y. Acad. Sci. 478: on hepatic and renal gluconeogenesis. Poult. Sci. 58: 890–895.31–45. Soling, H. D., Kleineke, J., Willms, B., Janson, G. & Kuhn, A. (1973) Relation-
Hod, Y., Morris, S. M. & Hanson, R. W. (1984) Induction by cAMP of the mRNA ship between the intracellular distribution of P-enolpyruvate carboxykinase,encoding the cytosolic form of phosphoenolpyruvate carboxykinase (GTP) the regulation of gluconeogenesis and the energy cost of glucose formation.from the chicken. Identification and characterization of a cDNA clone for the Eur. J. Biochem. 37: 233–243.enzyme. J. Biol. Chem. 259: 15603–15608. Sturkie, P. D. (1965) Avian Physiology, 2nd ed. Comstok Pub. Associates, Ith-
Hod, Y., Yoo-Warren, H. & Hanson, R. W. (1984) The gene encoding the cyto- aca, NY.solic form of phosphoenolpyruvate carboxykinase (GTP) from the chicken. J. Tontonoz, P., Hu, E., Devine, J., Beale, E. G. & Spiegelman, B. M. (1995)Biol. Chem. 259: 15609–15614. PPARg2 regulates adipose expression of the PEPCK gene. Mol. Cell. Biol.
Imai, E., Miner, J. N., Mitchel, J. A., Yamamoto, K. R. & Granner, D. A. (1993) 15: 351–357.Glucocorticoid receptor-cAMP response element-binding protein interaction Wang, N., Fingold, M. J., Bradley, A., Ou, C. N., Abdelsayed, S. V., Wilde, M. D.,and the response of the P-enolpyruvate carboxykinase gene to glucocorti- Taylor, L. R., Wilson, D. R. & Darlington, G. J. (1995) Impaired energy ho-coids. J. Biol. Chem. 268: 5353–5356. meostasis in C/EBPa knockout mice. Science (Washington, DC) 269: 1108–
Imai, E., Stromstedt, P., Quinn, P. G., Carlstedt-Duke, J., Gunstaffson, J. & Gran- 1112.ner, D. K. (1990) Characterization of a complex glucocorticoid response Watford, M., Hod, Y., Chiao, Y.-B., Utter, M. F. & Hanson, R. W. (1981) Theunit in the P-enolpyruvate carboxykinase gene. Mol. Cell. Biol. 10: 4712– unique role of the kidney in gluconeogenesis in the chicken: the significance4719. of a cytosolic form of PEPCK. J. Biol. Chem. 256: 10023–10027.
Johnson, D. C., Ebert, K. A. & Ray, P. D. (1970) Hepatic gluconeogenesis of Wynshaw-Boris, A., Lugo, T. G., Short, J. M., Fournier, R.E.K. & Hanson, R. W.the rabbit. Biochem. Biophys. Res. Commun. 39: 750–754. (1984) Identification of a cAMP regulatory region in the gene for rat cytosolic
Kawaguchi, T., Nomura, K., Hirayama, Y. & Kitagawa, T. (1987) Establishment phosphoenolpyruvate carboxykinase (GTP). J. Biol. Chem. 261: 12161–and characterization of a chicken hepatocellular carcinoma cell line, LMH. 12169.Cancer Res. 47: 4460–4464. Wynshaw-Boris,A.,Short,J. M.,Loose,D. S.&Hanson,R. W. (1986) Character-
Klemm, D. J., Roesler, W. J., Liu, J., Park, E. A. & Hanson, R. W. (1990) In vitro ization of the phosphoenolpyruvate carboxykinase (GTP) promoter-regulatoryanalysis of promoter elements regulating transcription of the P-enolpyruvate region. I. Effects of enhancers and multiple hormones regulatory elements.carboxykinase gene. Mol. Cell. Biol. 10: 480–485. J. Biol. Chem. 261: 9714–9720.
Lamers, W. H., Hanson, R. W. & Meisner, H. M. (1982) Cyclic AMP stimulates Yamamoto,K. K.,Gonzalez,G. A.,Briggs,W. H.&Montminy,M. R. (1988) Phos-transcription of the gene for cytosolic phosphoenolpyruvate carboxykinase phorylation-induced binding and transcriptional efficacy of nuclear binding
protein CREB. Nature (Lond.) 334: 494–498.in rat liver nuclei. Proc. Natl. Acad. Sci. U.S.A. 79: 5137–5141.
/ 4w13$$0020 01-17-97 12:36:15 nutra LP: J Nut
by guest on January 12, 2016jn.nutrition.org
Dow
nloaded from




















