
Neutrophils - A key component of ischemia-reperfusion injury

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
The potential role of cobalt ions released from metal prosthesis on the inhibitionof Hv1 proton channels and the decrease in Staphyloccocus epidermidis killingby human neutrophils
Samira Daou a,b, Antoun El Chemaly a, Panayiotis Christofilopoulos c, Louis Bernard d, Pierre Hoffmeyer c,Nicolas Demaurex a,*
aDepartment of Cell Physiology and Metabolism, University of Geneva, 1 rue Michel-Servet CH-1211 Geneva 4, SwitzerlandbVersailles Saint Quentin en Yvelines University, 55 avenue de Paris 78035 Versailles Cedex, FrancecOrthopaedic Surgery Service, Geneva University Hospitals, 4, Rue Gabrielle Perret-Gentil, 1211 Geneva 14, SwitzerlanddBretonneau Hospital, F. Rabelais University, 37000 Tours, France
a r t i c l e i n f o
Article history:Received 13 October 2010Accepted 1 November 2010Available online 8 December 2010
Keywords:Proton channelsPhagocytosisNADPH oxidaseBacterial killingInnate immunity
a b s t r a c t
Infection by Staphylococcus epidermidis is a devastating complication of metal-on-metal (MM) total hiparthroplasty (THA). Neutrophils are the first line of defense against infection, and these innate immunecells are potentially exposed to Co2þ ions released in the peri-prosthetic tissue by the wear of MM THA.The toxicity of Co2þ is still debated, but Co2þ is a potential inhibitor of the Hv1 proton channel thatsustains the production of superoxide by neutrophils. In this study, we show that the Co2þ concentrationin peri-prosthetic tissue from patients with MM THA averages 53 mM and that such high concentrationsof Co2þ alter the antibacterial activity of human neutrophils in vitro by inhibiting Hv1 proton channels.We show that submillimolar concentrations of Co2þ inhibit proton currents, impair the extrusion ofcytosolic acid, and decrease the production of superoxide in human neutrophils. As a result, Co2þ reducesthe ability of human neutrophils to kill two strains of Staphyloccocus epidermidis by up to 7-fold at themaximal concentration tested of 100 mM Co2þ. By inhibiting proton channels, the Co2þ ions released bymetal prostheses might therefore promote bacterial infections in patients with metal-on-metal total hiparthroplasty.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Total hip arthroplasty has greatly improved the quality of life ofpatients suffering from traumatic or degenerative joint diseases.However, the first generation of polyethylene prostheses releasedsignificant amounts of wear particles through friction at the bearinginterface. Release of wear particles has been recognized to bea major concern in the longevity of THA [1] because the ultra-highmolecular weight polyethylene particles can initiate peri-prostheticosteolysis and lead to arthroplasty failure [2, 3]. To circumvent thisproblem, the orthopaedic industry has developed metal-on-metal,metal-on-highly cross-linked polyethylene, ceramic-on-ceramic,and ceramic-on-highly cross-linked polyethylene articulations thatproduce minimal amounts of wear particles [4e7]. Ceramic-on-ceramic bearings offer high wear resistance, low wear and low
friction, but their complications include bearing noise, chippingduring insertion and unanticipated bearing fracture [8]. Highlycross-linked polyethylenes bearings reduce wear of polyethylene[9] but long-term clinical results are not yet available. Among thesenew devices, metal-on-metal (MM) arthroplastics are oftenpreferred because of their low volumetric wear in hip simulators[10, 11] and in clinical practice [12]. Metal-on-metal prostheses,however, still generate metal particles by mechanical wear, andcorrosion of cobalt particles of high surface area subsequentlygenerate cobalt ions (Co2þ) in the peri-prosthetic tissue [13]. Thecobalt ions eventually reach the blood circulation where theirconcentration has been shown to be significantly elevated(>20 nM) in patients with MM THA [14e16]. The cobalt concen-tration in the peri-prosthetic tissues has been measured in onestudy of 6 patients at 20month post-implantation andwas found tobe 1000 fold higher compared to control patients (30 mg/g vs.0.03 mg/g) [17]. Although the toxicity of the cobalt ions released byMMTHA is a cause for concern [18e21], the role of cobalt ions in thephysiopathology of MM THA infection has not been studied so far.
* Corresponding author. Tel.: þ41 22 37 95 212; fax: þ41 22 37 95 260.E-mail address: [email protected] (N. Demaurex).
Contents lists available at ScienceDirect
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
0142-9612/$ e see front matter � 2010 Elsevier Ltd. All rights reserved.doi:10.1016/j.biomaterials.2010.11.016
Biomaterials 32 (2011) 1769e1777

Author's personal copy
Infection after MM THA is a devastating complication that isassociated with a significant increase in hospitalization-relatedmorbidity and mortality [22]. Staphylococcus epidermidis (S. epi-dermidis) is the most frequent etiological agent, recovered in19e37.5% of isolates from infected THA [22]. S. epidermidis is capableof biofilm formation and carries genes that mediate methicillinresistance [23e26]. Upon THA infection, polymorphonuclearleukocytes (PMNs, or neutrophils) are recruited rapidly to peri-prosthetic tissues where they ingest and kill bacteria by releasingenzymes into the phagocytic vacuole [27] together with reactiveoxygen species (ROS) generated by the phagocyte NADPH oxidase[28]. ROS productionplays a key role in host defence against bacteriaas illustrated by the recurrent and often life-threatening microbialinfections occurring in patients with deficient ROS production [29].
The phagocyte NADPH oxidase generates superoxide anions bymoving electrons from the cytosol to extracellular oxygen [28]. Theextrusion of a negative charge is electrogenic and depolarizes theplasma membrane [30e33], whereas the oxidation of NADPH intoNADPþ and Hþ generates substantial amounts of acid in the cytosol[34]. Proton channels prevent both the depolarization and thecytosolic acidification, and are required for the sustained produc-tion of superoxide [35]. Mice lacking the Hv1 proton channel havedefects in superoxide production and in bacterial killing [36,37],and accumulate large quantities of cytosolic acid [36,38]. Hv1proton channels are inhibited by micromolar concentrations ofdivalent cations such as Zn2þ, the prototypical inhibitor of protonchannels [39]. Divalent metal cations such as Cu2þ are equallypotent, whereas physiological cations such as Ca2þ and Mg2þ aremuch less potent inhibitors. Surprisingly, the effects of Co2þ onproton channels are poorly characterized. Co2þ was shown toinhibit proton currents in one pioneering study of snail neurons[40], but has not been tested since as proton channel inhibitor.Neutrophils are most exposed to the potential effects of Co2þ, asthese cells are recruited during infections to metal prostheses thatrelease Co2þ. Given the widespread use of prostheses containingCo2þ, the established inhibitory effect of divalent cations on protonchannels, and the reliance of neutrophils on proton channels forbacterial killing, we aimed to determine whether Co2þ alters thebactericidal capacity of neutrophils. Since Co2þ has been shown toinhibit proton currents in one study [40] we hypothesized that Co2þ
decreases the bactericidal activity of neutrophils by inhibitingproton channels. In this study, wemeasured the Co2þ concentrationin tissue explants from patients with MM THA, and tested whethermicromolar concentrations of Co2þ alter the in vitro antibacterialactivity of human neutrophils against S. epidermidis.
2. Materials and methods
2.1. Patients
Informed consent was obtained from all patients and the study was approved bythe local ethics committee. Peri prosthetic tissues and articular liquid were retrievedfrom twenty six hips of patients with THA (19 MM THA and 7 polyethylene-ceramicTHA) operated in the Orthopaedic Surgery Department of the Geneva UniversityHospitals from August 2009 to September 2010. The causes of revision were asepticloosening (19 cases), hip chronic pain (5 cases), hip chronic bruise (1 case) andinfection (1 cases). Blood samples were collected from the same patients preoper-atively in a standardized manner. All materials used to collect and store sampleswere chosen for their lack of metal investigated in this study. After reception, all thesamples were stored at þ 4 �C until analysis. All the samples (tissues, articular liquidand blood) were analyzed at the same time.
2.2. Analysis of cobalt ions concentration
A Thermo Elemental X7CCT bench top series with PlasmaLab� software andwithout a dynamic reaction cell (Thermo Fisher Scientific, Courtaboeuf, France) wasused for cobalt determination. For sample preparation, briefly, 500 mg tissues weredigested at 70 �C with pure nitric acid and 100 mL of the solution were diluted withthe dilution solution containing indium and rhodium as internal standards [nitric
acid 1% (v/v), butanol 0.5% (v/v), and triton 0.01% (v/v), In and Rh 1 ppb] beforeanalysis. Blood and articular fluid were directly diluted with the same dilutionsolution as tissues. For more details, see Supplemental methods.
2.3. Materials
Cobalt (II) chloride hexahydrate, phorbol 12-myristate 13-acetate (PMA),superoxide dismutase (SOD), zymosan, diphenyl iodonium (DPI) and catalase werepurchased from Sigma Aldrich; dimethyl sulfoxide from Biochemica and zincchloride from Merck.
2.4. Isolation of human neutrophils
According to the quantity of cells needed, neutrophils were obtained either fromcitrated venous blood of healthy consenting volunteers or from buffy coats obtainedfrom the Geneva University Hospitals blood bank. Neutrophils were isolated bysedimentation using Dextran T500 followed by Percoll gradient centrifugation andhypotonic lysis, as previously described [41]. Briefly, citrated blood was layered onthe top of 3 ml of FicollePaque PLUS (final density 1.077 g/ml) and centrifuged at700 g for 20 min at 4 �C. After removal of the Ficoll and supernatant containingmononuclear cells, the enriched granulocyte fraction was mixed with 5 ml of puredistilled water for 30 s to remove remaining erythrocytes. Osmolarity was restoredby addition of the same volume of NaCl 18 g/l and cells centrifuged at 400 g for 5minat 4 �C. The pellet was resuspended in 10ml NaCl 9 g/l and centrifuged again at 400 gfor 5 min at 4 �C. The number (8-15 x106) and purity (>95%) of neutrophils wasassessed by visual counting in a Neubauer chamber, and the cells resuspended inRPMI at a final concentration of 2x106/ml and kept on ice until used.
2.5. Bacterial strains, storage and growth conditions
Two strains of S. epidermidis were used. One commensal strain (V46) isolatedfrom the skin of an healthy donor (Microbiology Department, Versailles University,France) and one clinical strain (JAC) isolated from an infected metalemetal THA(Department of Orthopaedic Surgery, University of Geneva, Switzerland). The JACstrain is icaADBC (ST27 clone) and mecA positive. An overnight bacterial culture ofeach strain was diluted with fresh Luria broth and bacteria were grown underconstant rotation (180 rpm) at 37 �C to stationary growth phase (i.e., an opticaldensity at 600 nm of 0.7). Bacteria were collected by centrifugation (2000 � g,10 min), washed with phosphate-buffered saline (PBS) and stored at �80 �C.Accuracy of concentration of each bacterial aliquot for killing assay was routinelyverified by plating dilution on agar plates and counting colonies to determine CFU(colony- forming unit) per ml.
2.6. Neutrophil phagocytosis assay
Bacteria (107 cells per ml) were opsonized in culture medium (HBSS-HEPES20mM,1% human serum albumin (Sigma Aldrich) and 10% human serum) for 20minat 37 �C under rotation at 220 rpm and kept on ice afterwards. Neutrophils (2 � 106)were allowed to adhere at 37 �C for 30 min in 24-well plates and exposed to theinhibitors for 10min at room temperature. Opsonised bacteria (1�107)were added tocells (MOI: 1/5), phagocytosis allowed to proceed for 30 min at 37 �C, and cells incu-batedwith gentamicin (50 mg/ml) for an additional 30min at 37 �C to kill extracellularbacteria. Intracellular survivalwas thendetermined after lysis of PMNwith 0.1% tritonand bacteriawere plated onMüller Hinton agar. Colonies were enumerated for 1 day.
2.7. Neutrophil bactericidal activity
Killing of S. epidermidis by human PMNs was determined as described [42] withsome modifications. Briefly, after 10 min exposition to the inhibitors, adherent PMNs(2�106)were combinedwithopsonizedbacteria (1�107) in 24-well plates in culturemedium, centrifuged at 400 � g for 5 min (to synchronize phagocytosis) and phago-cytosis was allowed to proceed for 20 min at 37 �C. Cells were then kept for an addi-tional 2hat37 �C in thepresenceof gentamicin (25mg/ml) tokill extracellular bacteria.Intracellular survival was then determined after PMNs lysis with 0.1% triton andbacteria were plated on Müller Hinton agar. Colonies were enumerated for 1e2 days.
2.8. Measurement of ROS production
Superoxide (O2�) production was measured in 96-well microplates by moni-
toring either the superoxide dismutase (SOD) sensitive reduction of ferricytochromec [43], the L-012-enhanced chemiluminescence, or the fluorescence of Amplex red.Neutrophils (100 000 per well) were plated in 0.2 ml of buffer solution (140 mMNaCl, 5 mM KCl, 1 mMMgCl2, 2 mM CaCl210 mM glucose, 20 mM HEPES) containingeither 150 mM cytochrome c, 10 mM L-012 (SigmaeAldrich), or 25 mM Amplex redand 0.005 U/ml horseradish peroxidase. Cobalt (II) chloride hexahydrate was addedat the indicated concentration and the cells stimulated at t ¼ 0 with PMA or withopsonized zymosan (100 mg/ml). Plates were kept at 37 �C and luminescence,fluorescence (590 nm) or absorbance (550 nm) measurements were taken every1 min for 90e120 min on a FLUOstar (BMG Labtech) microplate reader. Each
S. Daou et al. / Biomaterials 32 (2011) 1769e17771770

Author's personal copy
condition was performed in triplicate. Six different concentrations of H2O2 wereused to obtain a calibration curve.
2.9. Intracellular pH measurements
Neutrophilswere loadedwith2mMBCECF/AMfor30min, seededon25-mmglasscoverslips that were inserted in a thermostatic chamber (Harvard Apparatus), andimaged on an Axiovert S100 TV microscope through a 40x 1.3 NA oil-immersionobjective (Carl Zeiss, Inc.) using a cooled 16-bit charge-coupled device back-illumi-nated frame transfer camera (MicroMax; Roper Industries). Cells were alternatelyexcited at 490 and 440 nm with a monochromator (DeltaRam; Photon TechnologyInternational) and images acquired using a 535 � 35-nm emission filter (OmegaOptical). Analysis was performed with the MetaFluor 6.2 software (MDS AnalyticalTechnologies). To exclude any contribution of the Naþ/Hþ antiporters and Cl�/HCO3
�
exchanger and to block vacuolar-type Hþ-ATPase, experiments were performed inHCO3
� and Naþ-free medium containing: 140 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2,10 mM Hepes, 10 mM glucose, 100 nM Bafilomycin A1 (SigmaeAldrich), 1 mM Vali-nomycin (SigmaeAldrich) and 100 nM PMA, pH 7.4. For calibration, 5 mg/ml nigericinand 5 mM monensin (SigmaeAldrich) were added to solutions containing 140 mMKCl, 1 mM MgCl2, 0.2 mM EGTA, and 20 mM Hepes, pH 7.0e7.5, or MES, pH 6.0e6.5.
2.10. Electrophysiology
The whole-cell configuration of the patch-clamp technique was used to recordproton currents via a patch-clamp amplifier (Axoclamp 200B, Axon Instruments)interfaced to a personal computer. Pipettes were made from GC150TF-10 HarvardApparatus borosilicate glass capillaries using a PC-10 Narishige puller. Patch elec-trodes of 3e8 MU were filled with an internal solution containing (in mmol/L):118 N-methyl-D-glucamine, 63 tetramethylammonium methanesulfonate, 1 MgCl2,110 MES, and 1 EGTA. This solution was adjusted to pHi 6.5 with TMAOH. Extra-cellular solution contained (in mmol/L) 130 TMAMeSO3, 1.8 CaCl2, 100 HEPES and40 mM mannitol and adjusted at pHo 7.5. The osmolarities of both internal andexternal solutions were maintained at 290 and 300 mOsm, respectively. Seals wereformed in Ringer solution containing (in mmol/L) 140 NaCl, 5 KCl, 1.8 CaCl2, 1.8MgCl2, 10 Hepes and 10 glucose (pH 7.4). No liquid junction potential correction wasapplied and all experiments were performed at room temperature (20e25 �C).
3. Results
3.1. Co2þ concentrations in tissue samples from patients with metalprostheses
To test whether the wear of metal prostheses could releasesignificant amounts of Co2þ ions near the implant site, we measured
the Co2þ concentrations inperi-prosthetic tissues and articular liquidfrom patients who had undergone surgery for a revision of their MMTHA. As shown in Table 1, the mean Co2þ concentration in tissuesretrieved from 19 patients with a MM THA was 53 mM (range:0.09e410 mM, SD: 94 mM) whereas the Co2þ concentration in tissuesfrom7patientswearing apolyethylene/ceramicTHAaveraged5.8mM(range: 0.037e39mM, SD: 15 mM). Thus, the local Co2þ concentrationswere∼9 foldhigher inperi-prosthetic tissues retrieved fromMMTHApatients than in peri-prosthetic tissues retrieved from polyethylene/ceramic THA patients. These tissue Co2þ concentrations are signifi-cantly higher than the normal plasma Co2þ concentration of<10.9 nM [44]. 15 out of 19 MM THA patients had plasma Co2þ
concentrations higher than this reference value, compared to only 2out of 6 polyethylene/ceramic THA patients whose plasma Co2þ
concentration was on average ∼4 fold lower than patients with MMTHA (78 � 34 nM vs. 19 � 7 nM, Table 1).
3.2. Effects of Co2þ on bacterial killing by PMNs
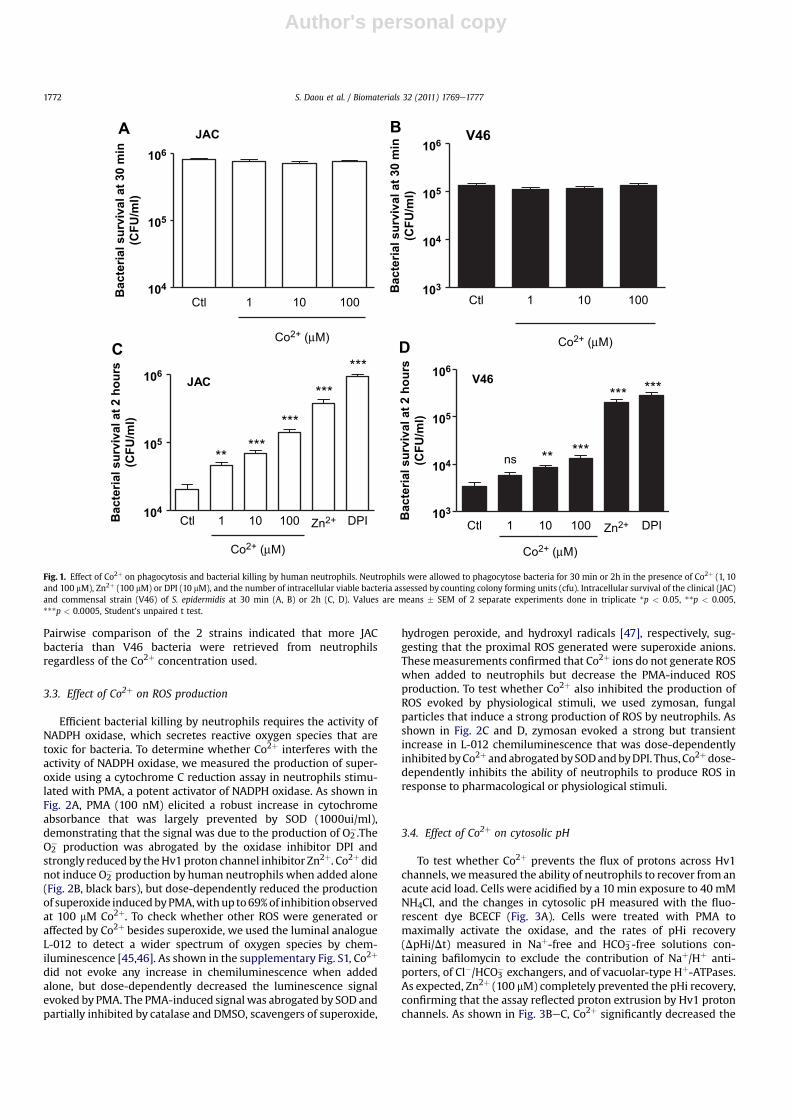
To determine whether Co2þ alters the ability of neutrophils todestroy bacteria, we measured the ability of neutrophils tophagocytose and kill 2 strains of S. epidermidis, a commensal strainisolated from skin (V46) and a clinical strain isolated from aninfected metalemetal THA (JAC). As shown in Fig. 1, the growth ofbacteria and the index of phagocytosis was not altered by thepresence of Co2þ (Fig. 1A and B). We then tested whetherthe intracellular survival of the ingested bacteria was affected bythe presence of Co2þ in the incubationmedium. As shown in Fig.1C,Co2þ significantly improved the intracellular survival of the JACstrain at all concentrations tested. The effect of Co2þ was dose-dependent, with 7 fold more bacteria retrieved from neutrophilsincubated with 100 mM Co2þ during the killing process. Zn2þ
(100 mM) and DPI (10 mM), known inhibitors of Hv1 proton chan-nels and of the NAPDH oxidase, respectively, further increased thesurvival of ingested bacteria. Similar effects were observed with theV46 commensal strain, whose intracellular survival increasedsignificantly at Co2þ concentrations exceeding 100 mM (Fig. 1D).
Table 1Characteristics of patient sample with Co2þ concentrations in peri-prosthetic tissue, blood and articular liquid. na ¼ not available.
Case Sex Patient age (years) Prosthesis age (years) Diagnosis Blood [Co2þ] mM Tissues [Co2þ] mM Articular liquid [Co2þ] mM
Metal/Metal total hip arthroplasty1 M 79 5 Aseptic loosning 0.119 108 na2 F 72 > 15 Aseptic loosning 0.617 410 16.43 M 80 4 Chronic pain 0.142 51 na4 F 85 2 Chronic bruise 0.007 99 2.185 M 65 4 Chronic pain 0.012 8.2 0.1346 F 68 2 Aseptic loosning 0.043 48 na7 M 79 2 infection 0.136 6 0.1758 M 72 15 Aseptic loosning 0.017 19.25 0.0219 M 72 12 Aseptic loosning 0.014 154 na10 M 81 14 Aseptic loosning 0.011 12.5 na11 M 79 2 Aseptic loosning 0.010 37 0.40812 M 81 3 Aseptic loosning 0.144 19 1313 M 66 2 Chronic pain 0.053 15 na14 F 51 4 Chronic pain na 3.14 na15 M 36 4 Aseptic loosning 0.008 0.767 na16 M 61 6 Aseptic loosning 0.024 0.379 2.1317 F 45 18 months Chronic pain 0.020 2 na18 M 36 2 Aseptic loosning 0.011 0.09 na19 M 43 2 Aseptic loosning 0.021 7.15 naPolyethylene/Ceramic total hip arthroplasty20 M 77 8 months infection 0.050 0.253 0.10921 F 69 2 Aseptic loosning na 0.209 0.00922 M 68 10 Aseptic loosning 0.010 39 na23 M 79 4 Aseptic loosning 0.009 0.95 na24 F 62 16 Aseptic loosning 0.010 0.037 na25 F 62 18 Aseptic loosning 0.008 0.059 na26 F 73 3 Aseptic loosning 0.025 0.584 na
S. Daou et al. / Biomaterials 32 (2011) 1769e1777 1771
Author's personal copy
Pairwise comparison of the 2 strains indicated that more JACbacteria than V46 bacteria were retrieved from neutrophilsregardless of the Co2þ concentration used.
3.3. Effect of Co2þ on ROS production
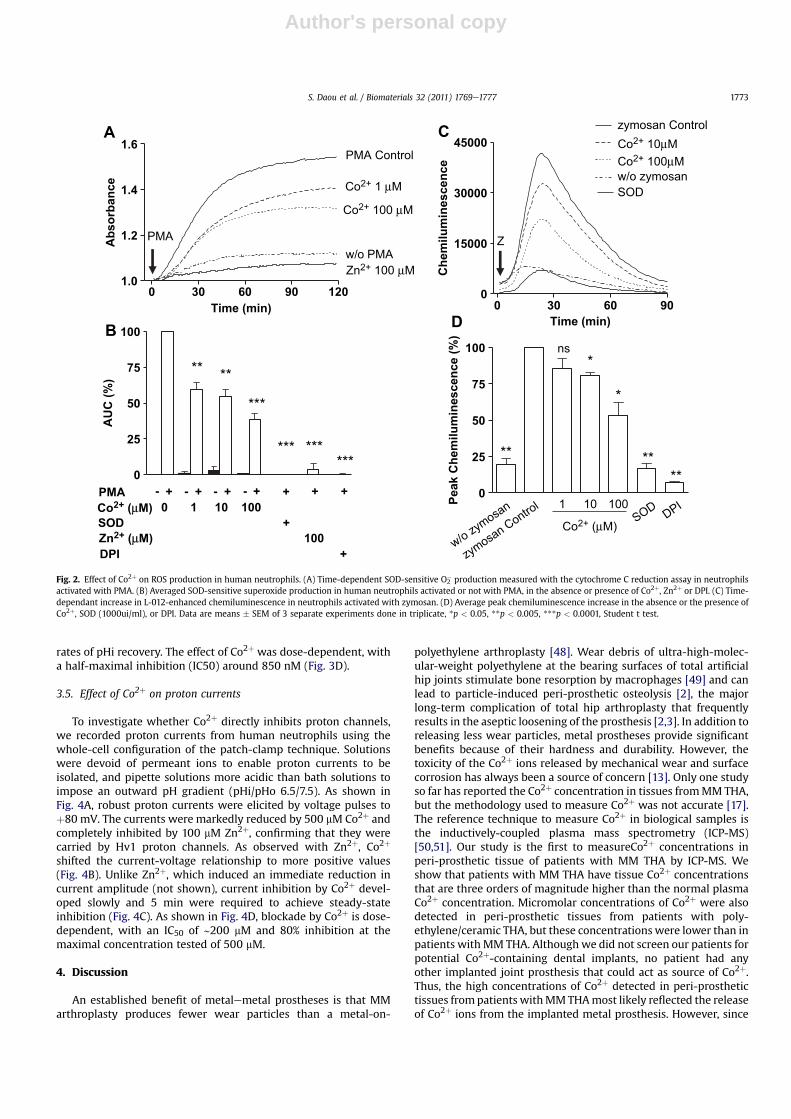
Efficient bacterial killing by neutrophils requires the activity ofNADPH oxidase, which secretes reactive oxygen species that aretoxic for bacteria. To determine whether Co2þ interferes with theactivity of NADPH oxidase, we measured the production of super-oxide using a cytochrome C reduction assay in neutrophils stimu-lated with PMA, a potent activator of NADPH oxidase. As shown inFig. 2A, PMA (100 nM) elicited a robust increase in cytochromeabsorbance that was largely prevented by SOD (1000ui/ml),demonstrating that the signal was due to the production of O2
�.TheO2� production was abrogated by the oxidase inhibitor DPI and
strongly reduced by theHv1proton channel inhibitor Zn2þ. Co2þdidnot induce O2
� production by human neutrophils when added alone(Fig. 2B, black bars), but dose-dependently reduced the productionof superoxide inducedbyPMA,withup to69%of inhibitionobservedat 100 mM Co2þ. To check whether other ROS were generated oraffected by Co2þ besides superoxide, we used the luminal analogueL-012 to detect a wider spectrum of oxygen species by chem-iluminescence [45,46]. As shown in the supplementary Fig. S1, Co2þ
did not evoke any increase in chemiluminescence when addedalone, but dose-dependently decreased the luminescence signalevoked by PMA. The PMA-induced signal was abrogated by SOD andpartially inhibited by catalase and DMSO, scavengers of superoxide,
hydrogen peroxide, and hydroxyl radicals [47], respectively, sug-gesting that the proximal ROS generated were superoxide anions.Thesemeasurements confirmed that Co2þ ions do not generate ROSwhen added to neutrophils but decrease the PMA-induced ROSproduction. To test whether Co2þ also inhibited the production ofROS evoked by physiological stimuli, we used zymosan, fungalparticles that induce a strong production of ROS by neutrophils. Asshown in Fig. 2C and D, zymosan evoked a strong but transientincrease in L-012 chemiluminescence that was dose-dependentlyinhibitedbyCo2þ andabrogatedbySODandbyDPI. Thus, Co2þdose-dependently inhibits the ability of neutrophils to produce ROS inresponse to pharmacological or physiological stimuli.
3.4. Effect of Co2þ on cytosolic pH
To test whether Co2þ prevents the flux of protons across Hv1channels, wemeasured the ability of neutrophils to recover from anacute acid load. Cells were acidified by a 10 min exposure to 40 mMNH4Cl, and the changes in cytosolic pH measured with the fluo-rescent dye BCECF (Fig. 3A). Cells were treated with PMA tomaximally activate the oxidase, and the rates of pHi recovery(DpHi/Dt) measured in Naþ-free and HCO3
�-free solutions con-taining bafilomycin to exclude the contribution of Naþ/Hþ anti-porters, of Cl�/HCO3
� exchangers, and of vacuolar-type Hþ-ATPases.As expected, Zn2þ (100 mM) completely prevented the pHi recovery,confirming that the assay reflected proton extrusion by Hv1 protonchannels. As shown in Fig. 3BeC, Co2þ significantly decreased the
*****
***
***
***JAC
DPI
ns***
*** ***V46
**
V46
104
105
106
104
103
105
106
104
103
105
106
Ctl 1 10 100
Ctl 1 10 100
Ctl 1 10 100
Co2+ (µM)
Co2+ (µM) Co2+ (µM)
Co2+ (µM)
JAC
Ba
cte
ria
l s
urv
iv
al a
t 3
0 m
in
(C
FU
/m
l)
104
105
106
Ba
cte
ria
l s
urv
iv
al a
t 2
h
ou
rs
(C
FU
/m
l)
Ba
cte
ria
l s
urv
iv
al a
t 2
h
ou
rs
(C
FU
/m
l)
Ba
cte
ria
l s
urv
iv
al a
t 3
0 m
in
(C
FU
/m
l)
Zn2+ DPICtl 1 10 100 Zn2+
A
C D
B
Fig. 1. Effect of Co2þ on phagocytosis and bacterial killing by human neutrophils. Neutrophils were allowed to phagocytose bacteria for 30 min or 2h in the presence of Co2þ (1, 10and 100 mM), Zn2þ (100 mM) or DPI (10 mM), and the number of intracellular viable bacteria assessed by counting colony forming units (cfu). Intracellular survival of the clinical (JAC)and commensal strain (V46) of S. epidermidis at 30 min (A, B) or 2h (C, D). Values are means � SEM of 2 separate experiments done in triplicate *p < 0.05, **p < 0.005,***p < 0.0005, Student’s unpaired t test.
S. Daou et al. / Biomaterials 32 (2011) 1769e17771772
Author's personal copy
rates of pHi recovery. The effect of Co2þ was dose-dependent, witha half-maximal inhibition (IC50) around 850 nM (Fig. 3D).
3.5. Effect of Co2þ on proton currents
To investigate whether Co2þ directly inhibits proton channels,we recorded proton currents from human neutrophils using thewhole-cell configuration of the patch-clamp technique. Solutionswere devoid of permeant ions to enable proton currents to beisolated, and pipette solutions more acidic than bath solutions toimpose an outward pH gradient (pHi/pHo 6.5/7.5). As shown inFig. 4A, robust proton currents were elicited by voltage pulses toþ80 mV. The currents were markedly reduced by 500 mM Co2þ andcompletely inhibited by 100 mM Zn2þ, confirming that they werecarried by Hv1 proton channels. As observed with Zn2þ, Co2þ
shifted the current-voltage relationship to more positive values(Fig. 4B). Unlike Zn2þ, which induced an immediate reduction incurrent amplitude (not shown), current inhibition by Co2þ devel-oped slowly and 5 min were required to achieve steady-stateinhibition (Fig. 4C). As shown in Fig. 4D, blockade by Co2þ is dose-dependent, with an IC50 of ∼200 mM and 80% inhibition at themaximal concentration tested of 500 mM.
4. Discussion
An established benefit of metalemetal prostheses is that MMarthroplasty produces fewer wear particles than a metal-on-
polyethylene arthroplasty [48]. Wear debris of ultra-high-molec-ular-weight polyethylene at the bearing surfaces of total artificialhip joints stimulate bone resorption by macrophages [49] and canlead to particle-induced peri-prosthetic osteolysis [2], the majorlong-term complication of total hip arthroplasty that frequentlyresults in the aseptic loosening of the prosthesis [2,3]. In addition toreleasing less wear particles, metal prostheses provide significantbenefits because of their hardness and durability. However, thetoxicity of the Co2þ ions released by mechanical wear and surfacecorrosion has always been a source of concern [13]. Only one studyso far has reported the Co2þ concentration in tissues fromMM THA,but the methodology used to measure Co2þ was not accurate [17].The reference technique to measure Co2þ in biological samples isthe inductively-coupled plasma mass spectrometry (ICP-MS)[50,51]. Our study is the first to measureCo2þ concentrations inperi-prosthetic tissue of patients with MM THA by ICP-MS. Weshow that patients with MM THA have tissue Co2þ concentrationsthat are three orders of magnitude higher than the normal plasmaCo2þ concentration. Micromolar concentrations of Co2þ were alsodetected in peri-prosthetic tissues from patients with poly-ethylene/ceramic THA, but these concentrations were lower than inpatients with MM THA. Although we did not screen our patients forpotential Co2þ-containing dental implants, no patient had anyother implanted joint prosthesis that could act as source of Co2þ.Thus, the high concentrations of Co2þ detected in peri-prosthetictissues frompatients withMMTHAmost likely reflected the releaseof Co2þ ions from the implanted metal prosthesis. However, since
0 30 60 90 120
1.0
1.2
1.4
1.6
Time (min)
Ab
so
rb
an
ce
-- - -
0
25
50
75
100
PMA
0 1 10 100
SOD +
+++ +
100
** **
***
*** ***
DPI +
+ + +
***
AU
C (%
)
A
B
w/o PMAPMA
PMA Control
0 30 60 90
0
15000
30000
45000
w/o zymosan
zymosan Control
SOD
Time (min)
Ch
em
ilu
min
es
ce
nc
e
0
25
50
75
100
w/o zymosan
zymosan Control 1 10 100
SOD DPI
**
*
*
****
ns
Pe
ak
C
he
milu
min
es
ce
nc
e (%
)
C
D
Z
Co2+ 1 µM
Co2+ 100 µM
Co2+ 10µM
Co2+ (µM)
Co2+ 100µM
Zn2+ 100 µM
Zn2+ (µM)
Co2+ (µM)
Fig. 2. Effect of Co2þ on ROS production in human neutrophils. (A) Time-dependent SOD-sensitive O2� production measured with the cytochrome C reduction assay in neutrophils
activated with PMA. (B) Averaged SOD-sensitive superoxide production in human neutrophils activated or not with PMA, in the absence or presence of Co2þ, Zn2þ or DPI. (C) Time-dependant increase in L-012-enhanced chemiluminescence in neutrophils activated with zymosan. (D) Average peak chemiluminescence increase in the absence or the presence ofCo2þ, SOD (1000ui/ml), or DPI. Data are means � SEM of 3 separate experiments done in triplicate, *p < 0.05, **p < 0.005, ***p < 0.0001, Student t test.
S. Daou et al. / Biomaterials 32 (2011) 1769e1777 1773

Author's personal copy
our tissue samples were obtained from patients undergoing revi-sion for THA Co2þ levels might be lower in tissues from patientswith well-tolerated prosthesis.
Upon THA infection, neutrophils are recruited rapidly to peri-prosthetic tissues, where they are found in significant numbers[52,53]. Neutrophils are an essential first-line of defence againstinvading bacteria, and deficiencies in neutrophil function invariablylead to life-threatening bacterial infections. Here, we show thatCo2þ reduces the ability of human neutrophils to secrete super-oxide and to kill bacteria. This effect correlates with the ability ofCo2þ to inhibit Hv1 proton channels. Divalent cations like Zn2þ andCu2þ are well known inhibitors of proton channels, but Co2þ wasonly tested in one pioneering study so far. More than 20 years ago,Byerly and Suen demonstrated that Co2þ partially blocks outwardproton currents in neurones of the snail, Lymnaea stagnalis [40].Here, we show that Co2þ also blocks outward proton currents inhuman neutrophils at concentrations exceeding 10 mM. Asobserved with other divalent cations, Co2þ shifted the current-voltage relationship of the Hv1 channel towards more positivevoltages, effectively reducing the efflux of protons across thechannel. Co2þ also strongly prevented cytosolic acid extrusion fromcells depolarised with valinomycin/Kþ, an assay system that hasbeen used extensively to study the activity of proton channels inintact cells [54]. In intact neutrophils however, efficient inhibitionof proton channels was achieved at much lower concentrations of
Co2þ, with half-maximal inhibition achieved at concentrations inthe 1 mM range. Thus, Co2þ is an effective proton channel blocker inintact activated neutrophils.
Hv1 proton channels are activated by phosphorylation, a prop-erty that couples their activation with the activation of the oxidase[55]. Phosphorylation profoundly enhances the gating of Hv1channels, the phosphorylated channels activating at ∼60 mV lowervoltages, with faster kinetics of activation and slower kinetics ofdeactivation [56]. The enhanced gating of phosphorylated protonchannels facilitates the extrusion of protons from activatedneutrophils, but renders activated cells more sensitive to inhibitionby divalent cations at physiological voltages [56]. Because neutro-phils were activated with PMA during our pH measurements, theirhigher sensitivity to inhibition by Co2þ most likely reflects phos-phorylation of the Hv1 channel. The higher Co2þ sensitivity ofintact, activated neutrophils is more relevant from a physiopatho-logical standpoint, because the bactericidal actions of neutrophilsrequire phosphorylation of the oxidase and of its associated protonchannel.
The main physiological role of proton channels is to sustain theactivity of the phagocyte NADPH oxidase, by providing chargecompensation for the electrons that are translocated from cytosolicNADPH to extracellular oxygen and by extruding the acid releasedin the cytosol during the oxydation of NADPH. This mechanismpostulated long ago by Henderson and colleagues [31] was
15 20 25 30
5.5
5.6
5.7
5.8
5.9
6.0
Control
Co2+ 100 μMZn2+ 100 μM
Co2+ 10 μM
Time (min)
Cy
to
so
lic
p
H
0 5 10 15 20 25 30
5
6
7
8
9 NH4Cl
Time (min)
Cy
to
so
lic
p
H
0.00
0.01
0.02
0.03
**
**
***
*
Ctl 1 10 100 Zn2+
Co2+
(µM)
Reco
very r
ates ( Δ
ΔpH
i/m
in)
0 1 2 3 4 5 6
0.01
0.02
0.03
log [Co2+
] (nM)
Reco
very r
ates (
pH
i/m
in)
A B
C D
Fig. 3. Effect of Co2þ on cytosolic pH recovery from an acute acid load. Human neutrophils were loaded with the pH-sensitive dye BCECF and changes in fluorescence ratio (F490/F440) were measured in bicarbonate-sodium-free solution to minimise the contribution of the Naþ/Hþ antiporters and Cl�/HCO3
� exchanger and in presence of 100 nM Bafilomycinto inhibit vacuolar-type Hþ-ATPase. (A) Experimental protocol. The addition of 40 mM NH4Cl evokes a rapid alkalinisation followed by a gradual decrease in pHi, due to thegeneration of cytosolic acid. The removal of the NH4Cl pulse induced cytosolic acidification followed by cell pHi recovery to pre-stimulatory levels. (B) Effects of Co2þ and Zn2þ oncell pHi recovery. (C) Average pHi recovery rates (dpHi/min) in the absence or presence of Co2þ or Zn2þ. (D) Dose-inhibition curve of Co2þ effects on pHi recovery rates (R2 ¼ 0.95).Data are means � SEM of 3 separate experiments, *p < 0.05, **p < 0.005, ***p < 0.0005, Student’s unpaired t test.
S. Daou et al. / Biomaterials 32 (2011) 1769e17771774

Author's personal copy
confirmed in recent studies using mice genetically ablated for theHv1 channel. Neutrophils from Hv1-deficient mice are more acid,more depolarised, and produce less superoxide anions thanneutrophils from wild-type mice [36]. Consistent with thissequence of events, Co2þ, which inhibits proton channels, impairedthe ability of neutrophils to extrude cytosolic acid and to producesuperoxide. Because of the established causality between protonchannels defects and reduced oxidase activity, the blunted super-oxide production observed in the presence of micromolarconcentrations of Co2þ can therefore be attributed to the inhibitionof Hv1 channels.
The superoxide anion is a precursor of numerous other ROSproduced by phagocytes that contribute to microbial killing [27,57].Predictably, we observed that human PMNs failed to kill correctly 2strains of S. epidermidis in the presence of Co2þ. This ability was notrestricted to the commensal strain, but extended to a clinical strainthat was more resistant to killing by neutrophils. Thus, Co2þ
impairs the ability of neutrophils to kill bacteria. This defect is mostlikely due to the inhibition of proton channels that are required forhigh-levels production of superoxide.
The observation that Co2þ inhibits ROS production by neutro-phils is surprising, because Co2þ was reported to induce ROSproduction in several cell types including neurons, astrocytes, andhepatocytes [58e60]. In these cell types, the production of ROSpersisted in the presence of DPI or in cells depleted ofmitochondrial
DNA [61,62], indicating that Co2þ generates ROS independently ofmitochondria and of NADPHoxidases. In our hands, Co2þ addition toneutrophils did not generate detectable amounts of ROS, as assessedby the cytochrome C reduction assay or by luminol-enhancedchemiluminescence (Figs. 2 and S1). In contrast, Co2þ impaired theability of neutrophils to produce ROS in response to PMA or tozymosan. These findings are consistent with two earlier studiesreporting that Co2þ does not promote ROS production in humanneutrophils [63,64] but instead inhibits the O2
� production evokedby the chemotactic peptide f-MLP [[63,64]. Thus, in neutrophils,Co2þ does not generate ROS but instead inhibits the production ofROS by the phagocytic NADPH oxidase.
Co2þ ions were previously shown to have cytotoxic effects onosteoblasts andmacrophages and to increase the production of TNFby macrophages [18e21]. We now show that Co2þ ions decreasethe ability of neutrophils to kill bacteria by inhibiting protonchannels. Although our results obtained in vitro cannot be readilyextrapolated to a clinical context, these data suggest that Co2þ
might promote bacterial infections in peri-prosthetic tissue frompatients with metal-on-metal total hip arthroplasty. The range ofnoxious effects observed in vitro highlights the potential toxicity ofCo2þ ions released by wear of MM THA. Cobalt alloys are currentlythe preferred materials for MM hip implants due to their hardnessand wear resistance. Engineering efforts should thus aim to furtherreduce wear and Co2þ release. Surface engineering technology of
Fig. 4. Effect of cobalt ions on voltage-activated proton currents. (A) Proton currents recorded from the same cell at þ80 mV in absence (Ctrl) and presence of Co2þ (500 mM) andZn2þ (100 mM). The pH of the solutions used is pHi/o ¼ 6.5/7.5. The protocol used is shown above the traces. (B) Current-voltage relationship showing the effect of Co2þ (10 and100 mM) and Zn2þ (100 mM) on Hv1 currents. The current amplitude is presented as the difference between the end and the beginning of each pulse. (C) Time-course of protoncurrent inhibition by Co2þ. Co2þ (500 mM) was added at the time indicated by the arrow. Data are from the same cell studied in (A). (D) Dose-response relationship of currentinhibition by cobalt ions at þ80 mV. Data were fitted with a sigmoidal equation where IC50 ¼ 186 mM. Results are expressed as mean � SEM with the number of cells tested given inparenthesis.
S. Daou et al. / Biomaterials 32 (2011) 1769e1777 1775

Author's personal copy
MM bearings might reduce wear, as surface-coated componentsprovide a smoother wear surface that should remain undamagedfor longer periods [65e67]. This technology is still in developmenthowever, and further investigations are required to determine itsefficacy.
5. Conclusion
In this study, we show that Co2þ ions reach micromolarconcentrations in peri-prosthetic tissues of patients with metalprostheses, and that similar Co2þ concentrations impair the abilityof human neutrophils to kill two stains of S. epidermidis in vitro. Theimmunosuppressive effect of Co2þ was most likely due to theinhibition of Hv1 proton channels that sustain high-level produc-tion of superoxide by human neutrophils. The high Co2þ concen-trations achieved in peri-prosthetic tissues might therefore impairthe ability of neutrophils to kill bacteria during prosthetic jointinfection in patients with metal-on-metal total hip arthroplasty.
Acknowledgements
The authors would like to thank C. Castelbou for his excellenttechnical assistance and Dr. Vincent Jaquet for expert advice on ROSmeasurements. This research was supported by grant No 3100A0-118393 from the Swiss National Science Foundation (to ND) and bya grant from the ENDO-Stiftung, Hamburg, Germany (to PH).
Appendix. Supplementary material
The supplementary data associated with this article can befound in online version at doi:10.1016/j.biomaterials.2010.11.016.
References
[1] Campbell P, Doorn P, Dorey F, Amstutz HC. Wear and morphology of ultra-high molecular weight polyethylene wear particles from total hip replace-ments. Proc Inst Mech Eng H 1996;210:167e74.
[2] Harris WH. Wear and periprosthetic osteolysis: the problem. Clin Orthop RelatRes; 2001:66e70.
[3] Purdue PE, Koulouvaris P, Nestor BJ, Sculco TP. The central role of wear debrisin periprosthetic osteolysis. HSS J 2006;2:102e13.
[4] Archibeck MJ, Jacobs JJ, Black J. Alternate bearing surfaces in total jointarthroplasty: biologic considerations. Clin Orthop Relat Res 2000:12e21.
[5] Charnley J. Long-term results of low-friction arthroplasty. Hip 1982:42e9.[6] Fisher J, Jin Z, Tipper J, Stone M, Ingham E. Tribology of alternative bearings.
Clin Orthop Relat Res 2006;453:25e34.[7] Naudie D, Roeder CP, Parvizi J, Berry DJ, Eggli S, Busato A. Metal-on-metal
versus metal-on-polyethylene bearings in total hip arthroplasty: a matchedcase-control study. J Arthroplasty 2004;19:35e41.
[8] Bizot P, Nizard R, Hamadouche M, Hannouche D, Sedel L. Prevention of wearand osteolysis: alumina-on-alumina bearing. Clin Orthop Relat Res2001:85e93.
[9] Atienza Jr C, Maloney WJ. Highly cross-linked polyethylene bearing surfaces intotal hip arthroplasty. J Surg Orthop Adv 2008;17:27e33.
[10] Chan FW, Bobyn JD, Medley JB, Krygier JJ, Yue S, Tanzer M. Engineering issuesand wear performance of metal on metal hip implants. Clin Orthop Relat Res1996:96e107.
[11] Medley JB, Chan FW, Krygier JJ, Bobyn JD. Comparison of alloys and designs ina hip simulator study of metal on metal implants. Clin Orthop Relat Res1996:S148e59.
[12] McKellop H, Park SH, Chiesa R, Doorn P, Lu B, Normand P, et al. In vivo wear ofthree types of metal on metal hip prostheses during two decades of use. ClinOrthop Relat Res 1996:S128e40.
[13] Jacobs JJ, Gilbert JL, Urban RM. Corrosion of metal orthopaedic implants.J Bone Jt Surg Am 1998;80:268e82.
[14] Clarke MT, Lee PT, Arora A, Villar RN. Levels of metal ions after small- andlarge-diameter metal-on-metal hip arthroplasty. J Bone Jt Surg Br2003;85:913e7.
[15] Lhotka C, Szekeres T, Steffan I, Zhuber K, Zweymuller K. Four-year study ofcobalt and chromium blood levels in patients managed with two differentmetal-on-metal total hip replacements. J Orthop Res 2003;21:189e95.
[16] Milosev I, Pisot V, Campbell P. Serum levels of cobalt and chromium inpatients with Sikomet metal-metal total hip replacements. J Orthop Res2005;23:526e35.
[17] Bohler M, Kanz F, Schwarz B, Steffan I, Walter A, Plenk Jr H, et al. Adversetissue reactions to wear particles from Co-alloy articulations, increased byalumina-blasting particle contamination from cementless Ti-based total hipimplants. A report of seven revisions with early failure. J Bone Jt Surg Br2002;84:128e36.
[18] Catelas I, Petit A, Vali H, Fragiskatos C, Meilleur R, Zukor DJ, et al. Quantitativeanalysis of macrophage apoptosis vs. necrosis induced by cobalt and chro-mium ions in vitro. Biomaterials 2005;26:2441e53.
[19] Catelas I, Petit A, Zukor DJ, Antoniou J, Huk OL. TNF-alpha secretion andmacrophage mortality induced by cobalt and chromium ions in vitro-quali-tative analysis of apoptosis. Biomaterials 2003;24:383e91.
[20] Catelas I, Petit A, Zukor DJ, Huk OL. Cytotoxic and apoptotic effects of cobaltand chromium ions on J774 macrophages - Implication of caspase-3 in theapoptotic pathway. J Mater Sci Mater Med 2001;12:949e53.
[21] Fleury C, Petit A, Mwale F, Antoniou J, Zukor DJ, Tabrizian M, et al. Effect ofcobalt and chromium ions on human MG-63 osteoblasts in vitro: morphology,cytotoxicity, and oxidative stress. Biomaterials 2006;27:3351e60.
[22] Lentino JR. Prosthetic joint infections: bane of orthopedists, challenge forinfectious disease specialists. Clin Infect Dis 2003;36:1157e61.
[23] Galdbart JO, Allignet J, TungHS, RydenC, El SolhN. Screening for Staphylococcusepidermidis markers discriminating between skin-flora strains and thoseresponsible for infections of joint prostheses. J Infect Dis 2000;182:351e5.
[24] Kozitskaya S, Olson ME, Fey PD, Witte W, Ohlsen K, Ziebuhr W. Clonal analysisof Staphylococcus epidermidis isolates carrying or lacking biofilm-mediatinggenes by multilocus sequence typing. J Clin Microbiol 2005;43:4751e7.
[25] Sivadon V, Rottman M, Quincampoix JC, Prunier E, de Mazancourt P,Bernard L, et al. Polymorphism of the cell wall-anchoring domain of theautolysin-adhesin AtlE and its relationship to sequence type, as revealed bymultilocus sequence typing of invasive and commensal Staphylococcus epi-dermidis strains. J Clin Microbiol 2006;44:1839e43.
[26] Sivadon V, Rottman M, Quincampoix JC, Prunier E, Le Moal M, deMazancourt P, et al. Partial atlE sequencing of Staphylococcus epidermidisstrains from prosthetic joint infections. J Clin Microbiol 2009;47:2321e4.
[27] Nauseef WM. How human neutrophils kill and degrade microbes: an inte-grated view. Immunol Rev 2007;219:88e102.
[28] Babior BM, Lambeth JD, Nauseef W. The neutrophil NADPH oxidase. ArchBiochem Biophys 2002;397:342e4.
[29] Segal AW. Absence of both cytochrome b-245 subunits from neutrophils inX-linked chronic granulomatous disease. Nature 1987;326:88e91.
[30] DeCourseyTE,MorganD,ChernyVV. ThevoltagedependenceofNADPHoxidasereveals why phagocytes need proton channels. Nature 2003;422:531e4.
[31] Henderson LM, Chappell JB, Jones OT. The superoxide-generating NADPHoxidase of human neutrophils is electrogenic and associated with an Hþchannel. Biochem J 1987;246:325e9.
[32] Schrenzel J, Serrander L, Banfi B, Nusse O, Fouyouzi R, Lew DP, et al. Electroncurrents generated by the human phagocyte NADPH oxidase. Nature1998;392:734e7.
[33] Petheo GL, Demaurex N. Voltage- and NADPH-dependence of electron currentsgenerated by the phagocytic NADPH oxidase. Biochem J 2005;388:485e91.
[34] Demaurex N, Downey GP, Waddell TK, Grinstein S. Intracellular pH regulationduring spreading of human neutrophils. J Cell Biol 1996;133:1391e402.
[35] Demaurex N, Petheo GL. Electron and proton transport by NADPH oxidases.Philos Trans R Soc Lond B Biol Sci 2005;360:2315e25.
[36] El Chemaly A, Okochi Y, Sasaki M, Arnaudeau S, Okamura Y, Demaurex N.VSOP/Hv1 proton channels sustain calcium entry, neutrophil migration, andsuperoxide production by limiting cell depolarization and acidification. J ExpMed 2010;207:129e39. S1-2.
[37] Ramsey IS, RuchtiE,Kaczmarek JS, ClaphamDE.Hv1protonchannelsare requiredfor high-level NADPH oxidase-dependent superoxide production during thephagocyte respiratory burst. Proc Natl Acad Sci U S A 2009;106:7642e7.
[38] Murphy R, DeCoursey TE. Charge compensation during the phagocyte respi-ratory burst. Biochim Biophys Acta 2006;1757:996e1011.
[39] DeCoursey TE, Cherny VV. Pharmacology of voltage-gated proton channels.Curr Pharm Des 2007;13:2400e20.
[40] Byerly L, Suen Y. Characterization of proton currents in neurones of the snail,Lymnaea stagnalis. J Physiol 1989;413:75e89.
[41] Miyamasu M, Hirai K, Takahashi Y, Iida M, Yamaguchi M, Koshino T, et al.Chemotactic agonists induce cytokine generation in eosinophils. J Immunol1995;154:1339e49.
[42] Spinner JL, Cundiff JA, Kobayashi SD. Yersinia pestis type III secretion system-dependent inhibition of human polymorphonuclear leukocyte function. InfectImmun 2008;76:3754e60.
[43] Mocsai A, Banfi B, Kapus A, Farkas G, Geiszt M, Buday L, et al. Differentialeffects of tyrosine kinase inhibitors and an inhibitor of the mitogen-activatedprotein kinase cascade on degranulation and superoxide production of humanneutrophil granulocytes. Biochem Pharmacol 1997;54:781e9.
[44] Goulle JP, Mahieu L, Castermant J, Neveu N, Bonneau L, Laine G, et al. Metaland metalloid multi-elementary ICP-MS validation in whole blood, plasma,urine and hair. Reference values. Forensic Sci Int 2005;153:39e44.
[45] Daiber A, Oelze M, August M, Wendt M, Sydow K, Wieboldt H, et al. Detectionof superoxide and peroxynitrite in model systems and mitochondria by theluminol analogue L-012. Free Radic Res 2004;38:259e69.
[46] Yamazaki K, Fukuda K, Matsukawa M, Hara F, Yoshida K, Akagi M, et al.Reactive oxygen species depolymerize hyaluronan: involvement of thehydroxyl radical. Pathophysiology 2003;9:215e20.
S. Daou et al. / Biomaterials 32 (2011) 1769e17771776

Author's personal copy
[47] Klein SM, Cohen G, Cederbaum AI. Production of formaldehyde duringmetabolism of dimethyl sulfoxide by hydroxyl radical generating systems.Biochemistry 1981;20:6006e12.
[48] Doorn PF, Campbell PA, Worrall J, Benya PD, McKellop HA, Amstutz HC. Metalwear particle characterization from metal on metal total hip replacements:transmission electron microscopy study of periprosthetic tissues and isolatedparticles. J Biomed Mater Res 1998;42:103e11.
[49] Green TR, Fisher J, Matthews JB, Stone MH, Ingham E. Effect of size and dose onbone resorption activity ofmacrophages by in vitro clinically relevant ultra highmolecular weight polyethylene particles. J Biomed Mater Res 2000;53:490e7.
[50] Sarmiento-Gonzalez A, Marchante-Gayon JM, Tejerina-Lobo JM, Paz-Jimenez J,Sanz-Medel A. High-resolution ICP-MS determination of Ti, V, Cr, Co, Ni, andMo in human blood and urine of patients implanted with a hip or kneeprosthesis. Anal Bioanal Chem 2008;391:2583e9.
[51] Goulle JP, Saussereau E, Mahieu L, Bouige D, Guerbet M, Lacroix C. [A newmedical concept: the metallic profile]. Rev Med Interne 2010;31:128e34.
[52] Morawietz L, Tiddens O, Mueller M, Tohtz S, Gansukh T, Schroeder JH, et al.Twenty-three neutrophil granulocytes in 10 high-power fields is the besthistopathological threshold to differentiate between aseptic and septicendoprosthesis loosening. Histopathology 2009;54:847e53.
[53] Muller M, Morawietz L, Hasart O, Strube P, Perka C, Tohtz S. [Histopathologicaldiagnosis of periprosthetic joint infection following total hip arthroplasty: useof a standardized classification system of the periprosthetic interfacemembrane]. Orthopade 2009;38:1087e96.
[54] Grinstein S, Romanek R, Rotstein OD. Method for manipulation of cytosolic pHin cells clamped in the whole cell or perforated-patch configurations. AmJ Physiol 1994;267:C1152e9.
[55] Demaurex N, Schrenzel J, Jaconi ME, Lew DP, Krause KH. Proton channels,plasma membrane potential, and respiratory burst in human neutrophils. EurJ Haematol 1993;51:309e12.
[56] Banfi B, Schrenzel J, Nusse O, Lew DP, Ligeti E, Krause KH, et al. A novel H(þ)conductance in eosinophils: unique characteristics and absence in chronicgranulomatous disease. J Exp Med 1999;190:183e94.
[57] Hampton MB, Kettle AJ, Winterbourn CC. Inside the neutrophil phagosome:oxidants, myeloperoxidase, and bacterial killing. Blood 1998;92:3007e17.
[58] Karovic O, Tonazzini I, Rebola N, Edstrom E, Lovdahl C, Fredholm BB, et al.Toxic effects of cobalt in primary cultures of mouse astrocytes. Similaritieswith hypoxia and role of HIF-1alpha. Biochem Pharmacol 2007;73:694e708.
[59] Salnikow K, Costa M, Figg WD, Blagosklonny MV. Hyperinducibility ofhypoxia-responsive genes without p53/p21-dependent checkpoint inaggressive prostate cancer. Cancer Res 2000;60:5630e4.
[60] Zou W, Yan M, Xu W, Huo H, Sun L, Zheng Z, et al. Cobalt chloride inducesPC12 cells apoptosis through reactive oxygen species and accompanied byAP-1 activation. J Neurosci Res 2001;64:646e53.
[61] Chandel NS, Maltepe E, Goldwasser E, Mathieu CE, Simon MC, Schumacker PT.Mitochondrial reactive oxygen species trigger hypoxia-induced transcription.Proc Natl Acad Sci U S A 1998;95:11715e20.
[62] Chandel NS, McClintock DS, Feliciano CE, Wood TM, Melendez JA,Rodriguez AM, et al. Reactive oxygen species generated at mitochondrialcomplex III stabilize hypoxia-inducible factor-1alpha during hypoxia:a mechanism of O2 sensing. J Biol Chem 2000;275:25130e8.
[63] Ciapetti G, Granchi D, Verri E, Savarino L, Cenni E, Savioli F, et al. Fluorescentmicroplate assay for respiratory burst of PMNs challenged in vitro withorthopedic metals. J Biomed Mater Res 1998;41:455e60.
[64] Elferink JG, Deierkauf M. Suppressive action of cobalt on exocytosis andrespiratory burst in neutrophils. Am J Physiol 1989;257:C859e64.
[65] Fisher J, Hu XQ, Stewart TD, Williams S, Tipper JL, Ingham E, et al. Wear ofsurface engineered metal-on-metal hip prostheses. J Mater Sci Mater Med2004;15:225e35.
[66] Williams S, Isaac G, Hatto P, Stone MH, Ingham E, Fisher J. Comparative wearunder different conditions of surface-engineered metal-on-metal bearings fortotal hip arthroplasty. J Arthroplasty 2004;19:112e7.
[67] Leslie IJ, Williams S, Brown C, Anderson J, Isaac G, Hatto P, et al. Surfaceengineering: a low wearing solution for metal-on-metal hip surface replace-ments. J Biomed Mater Res B Appl Biomater 2009;90:558e65.
S. Daou et al. / Biomaterials 32 (2011) 1769e1777 1777
Copyright © 2022 FDOKUMEN




















