
The Physiology of Calcitonin Gene?Related Peptide in the Islet Compared with That of Islet Amyloid...
14
The Physiology of Calcitonin Gene-Related Peptide in the Islet Compared with That of Islet Amyloid Polypeptide (Amylin) D. BRETHERTON-WATT, M. A. GHATEI, H. JAMAL, S. G. GILBEY, P. M. JONES, AND S. R. BLOOM" Department oj Medicine Royal Postgraduate Medical School London W12 ONN, United Kingdom Calcitonin gene-related peptide (a-CGRP or CGRP-1) is a 37-amino acid peptide produced by alternative splicing of the primary transcript of the calcitonin gene.[ CGRP is widely distributed throughout the central nervous system and peripheral tissues, and has been shown to possess a number of biological actions, including potent effects on gastric acid secretion and food intake, pancreatic function, and on the cardiovascular system (for reviews, see Refs. 2 and 3). Subsequently, a second CGRP-encoding gene was discovered and predicted to encode a 37-amino acid ~ e p t i d e . ~ The production of this peptide, known as 0- CGRP or CGRP-2, has been demonstrated to occur in uiuo.s P-CGRP possesses a very similar spectrum of biological actions to CK-CGRP.'.~ However, the peptides exhibit different tissue specificities of distribution, which probably reflects the preferential biological actions of that form,5 such that a-CGRP is the predominant form in central nervous tissue and sensory innervation, while P-CGRP forms the main component of the gut intrinsic innervation.' In 1986, a new peptide of the calcitonin/CGRP family was identified.' Islet amyloid polypeptide (IAPP or amylin) was isolated from the amyloid deposits of an insulinoma' and of a type I1 diabetic pancreas.8 IAPP has a strong familial resemblance to CGRP, with 43% and 46% amino acid sequence homology with human a- and p-CGRPs, respectively (for review, see Ref. 9). Furthermore, both peptides are 37 amino acids long, are C-terminally amidated, and have an N-ter- minal disulfide bridge linking cysteine residues in positions 2 and 7. The similarities between the peptides suggested that IAPP and CGRP are evolutionary homo- logues that arose as a result of a gene duplication event. The location of the IAPP encoding gene to chromosome 12, an evolutionary homologue of chromosome 11 where a- and P-CGRPs are encoded,'(' agrees with this theory. The close structural similarity between IAPP and CGRP suggests they may bind to similar or even identical receptors. Indeed, IAPP has been shown to bind to and activate CGRP receptors in the 1iver.I' This structural homology has also caused problems with antisera and cast doubt upon the validity of earlier reports of CGRP in the pancreas, since it was shown that islet amyloid could react with anti-CGRP antisera." a Address for correspondence: Professor S. R. Bloom, 2"d Floor Francis Fraser Labs., Department of Medicine, Royal Postgraduate Medical School, I50 DuCane Road, London W12 ONN, United Kingdom. 299
Transcript of The Physiology of Calcitonin Gene?Related Peptide in the Islet Compared with That of Islet Amyloid...

The Physiology of Calcitonin Gene-Related Peptide in the Islet
Compared with That of Islet Amyloid Polypeptide (Amylin)
D. BRETHERTON-WATT, M. A. GHATEI, H . JAMAL, S. G. GILBEY, P. M. JONES, AND S. R. BLOOM"
Department o j Medicine Royal Postgraduate Medical School London W12 O N N , United Kingdom
Calcitonin gene-related peptide (a-CGRP or CGRP-1) is a 37-amino acid peptide produced by alternative splicing of the primary transcript of the calcitonin gene.[ CGRP is widely distributed throughout the central nervous system and peripheral tissues, and has been shown to possess a number of biological actions, including potent effects on gastric acid secretion and food intake, pancreatic function, and on the cardiovascular system (for reviews, see Refs. 2 and 3).
Subsequently, a second CGRP-encoding gene was discovered and predicted to encode a 37-amino acid ~ e p t i d e . ~ The production of this peptide, known as 0- CGRP or CGRP-2, has been demonstrated to occur in uiuo.s P-CGRP possesses a very similar spectrum of biological actions to CK-CGRP.'.~ However, the peptides exhibit different tissue specificities of distribution, which probably reflects the preferential biological actions of that form,5 such that a-CGRP is the predominant form in central nervous tissue and sensory innervation, while P-CGRP forms the main component of the gut intrinsic innervation.'
In 1986, a new peptide of the calcitonin/CGRP family was identified.' Islet amyloid polypeptide (IAPP or amylin) was isolated from the amyloid deposits of an insulinoma' and of a type I1 diabetic pancreas.8 IAPP has a strong familial resemblance to CGRP, with 43% and 46% amino acid sequence homology with human a- and p-CGRPs, respectively (for review, see Ref. 9). Furthermore, both peptides are 37 amino acids long, are C-terminally amidated, and have an N-ter- minal disulfide bridge linking cysteine residues in positions 2 and 7. The similarities between the peptides suggested that IAPP and CGRP are evolutionary homo- logues that arose as a result of a gene duplication event. The location of the IAPP encoding gene to chromosome 12, an evolutionary homologue of chromosome 1 1 where a- and P-CGRPs are encoded,'(' agrees with this theory. The close structural similarity between IAPP and CGRP suggests they may bind to similar or even identical receptors. Indeed, IAPP has been shown to bind to and activate CGRP receptors in the 1iver.I' This structural homology has also caused problems with antisera and cast doubt upon the validity of earlier reports of CGRP in the pancreas, since it was shown that islet amyloid could react with anti-CGRP antisera."
a Address for correspondence: Professor S. R. Bloom, 2"d Floor Francis Fraser Labs., Department of Medicine, Royal Postgraduate Medical School, I50 DuCane Road, London W12 ONN, United Kingdom.
299

300 ANNALS NEW YORK ACADEMY OF SCIENCES
IAPP is deposited as amyloid only in certain animals. Comparison of the amino acid sequences of IAPP from different species have pinpointed the amino acids 20-29 as the amyloidogenic region of the m~lecu le . ' ~ This region has the least degree of homology to the CGRP m o l e ~ u l e . ~ It is thought that in certain pathological conditions-for example, non-insulin dependent diabetes (N1DDM)-IAPP mole- cules can polymerize to form the large insoluble P-pleated sheets characteristic of amyloid proteins.I4 In NIDDM, IAPP is depleted from the p-cell, accumulating as fibrils in the islet interstitium.Is
Unlike CGRP, IAPP is not thought to be widespread throughout the body. The primary site of IAPP production is the p-cell of the pancreatic islet, where IAPP is co-localized with insulin to the p-cell secretory granule.16 However, there have been reports of IAPP in the human and rat gastrointestinal tracts.l7-I9 In addition, IAPP mRNA has been identified in rat stomach, lung, and dorsal root ganglia of the spinal cord."' IAPP has been shown to change in parallel with insulin in response to a range of physiological and pharmacological stimuli, both in the c i r~ula t ion~"~ and in isolated pancreatic preparation^.'^-'^
IAPP may be of pathological significance in the development of NIDDM. Deposition of pancreatic amyloid is the most common pathological feature of NIDDM.I4 Furthermore, the disease is characterized by elevated glucose in the presence of normal insulin, suggesting that tissues are not able to respond to insulin and promote lowering of blood glucose-that is, they are insulin re~istant.~' IAPP and CGRP have both been shown to counteract insulin action in peripheral tissues.'8.2'
In this paper, we will review some of the recent work relevant to the glucoregu- latory actions of CGRP and IAPP, both in normal physiology and in the pathology of insulin resistance and NIDDM.
PRODUCTION OF CGRP AND IAPP IN THE PANCREAS
IAPP is synthesized within the p-cell of the pancreatic islet and stored with insulin in the p-cell secretory granule.I6 Levels of IAPP immunoreactivity have been reported to be in the range of 1/3 to 1/100 insulin level^,?^.^^^^" while IAPP mRNA in rat pancreas has been reported as 1\10 insulin levels."' IAPP is released from isolated rat i ~ l e t s , ~ ' p -ce l l~ .~" and perfused whole rat p a n c r e a ~ , ? ~ - ? ~ in parallel with insulin in response to glucose and arginine.
CGRP is also found in the pancreas of several specie^,^'-^^ where it is thought to be present in the innervation around pancreatic blood vessels and islets. Within the islet, CGRP has been localized within cells around the islet periphery, co- localizing with the somatostatin-containing S cells in the rat.33-35 In other species, however-for instance, the mouse-CGRP has been identified in the insulin- secreting p-celk.33
Capsaicin treatment in neonatal rats is known to deplete CGRP-like immunore- activity (-LI) from spinal cord and peripheral sensory innervation, while the intrin- sic innervation of the enteric nervous system is unaffected. Using two radioimmu- noassays, one that was specific for a-CGRP and one that cross-reacted fully with both a- and p-CGRP, Mulderry et d.s established that the non-capsaicin sensitive population was due to P-CGRP innervation, while the capsaicin-sensitive innerva- tion was a-CGRP. In the whole rat pancreas, a- and P-CGRP immunoreactivities are approximately equal. Following capsaicin treatment, there is a specific loss of a-CGRP immunoreactivity. Such a result is consistent with the existence of two separate populations of CGRP within the pancreas.
BRETHERTON-WATT et al.: CGRP IN THE ISLET 301
TABLE 1. Details of the CGRP and IAPP Radioimmunoassays a-CGRP Assav Total CGRP Assav IAPP Assav
Antiserum Synthetic rat a-CGRP (1-37) conjugated to BSA by glutaralde- hyde (CG7)
Specificity 100% cross-reactivity a-CGRP, 2%' cross- reactivity p-CGRP
Sensitivity 1 fmoliassay tube
Synthetic rat p-CGRP (1-37) conjugated to BSA by carbodi- imide (CG3)
100% cross-reactivity with both a- and p- CGRP
2 fmoliassay tube
Synthetic human IAPP (1-37) conjugated to BSA by bis-diazoti- sation
100% cross-reactivity with rat IAPP, <0.01% cross-reac- tion a- or p-CGRPs
3 fmol/assay tube
Recently, we have investigated: (a) whether CGRP is locally produced in the pancreatic islet, and (b) the nature of islet CGRP. Rat pancreatic islets were separated from exocrine tissue by collagenase d i g e ~ t i o n . ~ ~ CGRP immunoreactivity was extracted from pooled islets in acidified ethanol at 4 "C and determined by radioimmunoassay as described in TABLE I . Total RNA was extracted for Northern blotting using a guanidium HCl/phenol/chloroform method of e~traction.~'
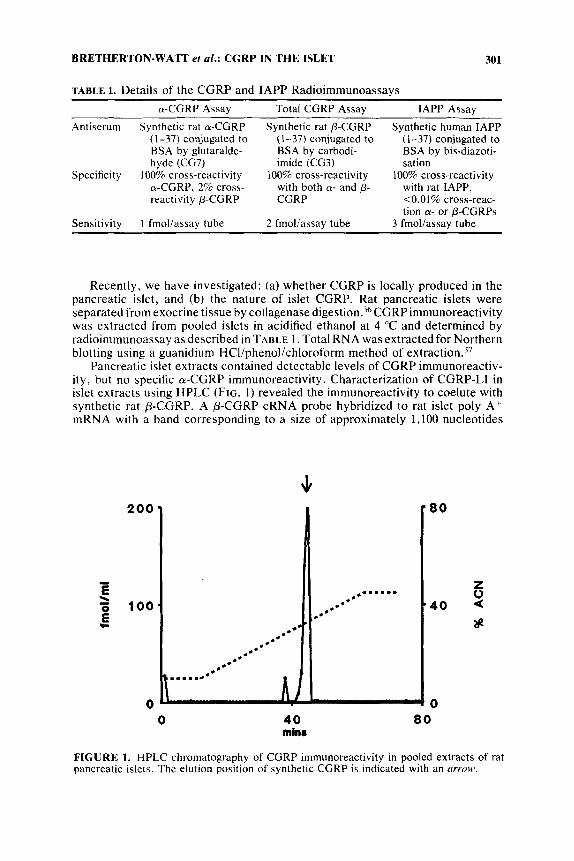
Pancreatic islet extracts contained detectable levels of CGRP immunoreactiv- ity, but no specific a-CGRP immunoreactivity . Characterization of CGRP-LI in islet extracts using HPLC (FIG. I ) revealed the immunoreactivity to coelute with synthetic rat P-CGRP. A P-CGRP cRNA probe hybridized to rat islet poly A + mRNA with a band corresponding to a size of approximately 1,100 nucleotides
200
e = 100 i! r
0 0
80
2 0
40 4 ap
0 40 minr
80
FIGURE 1. HPLC chromatography of CGRP immunoreactivity in pooled extracts of rat pancreatic islets. The elution position of synthetic CGRP is indicated with an arrow.
302 ANNALS NEW YORK ACADEMY OF SCIENCES
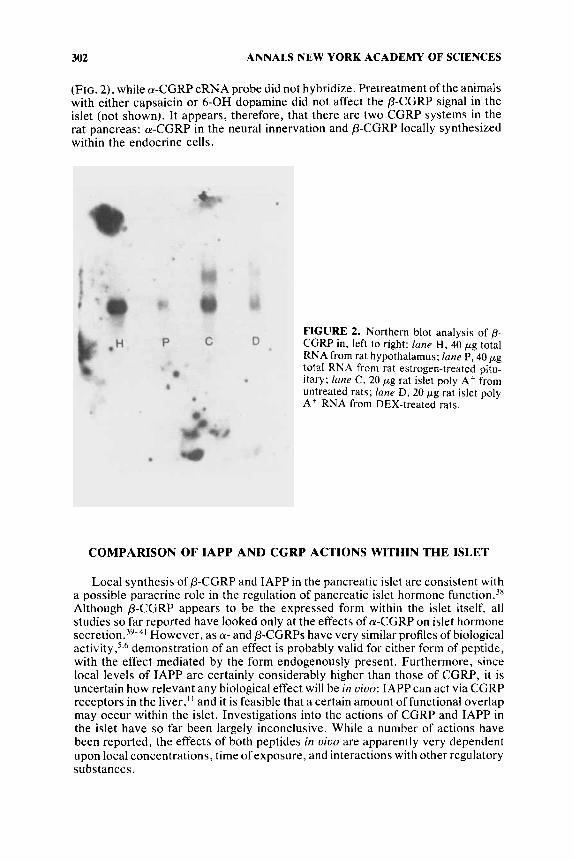
(FIG. Z), while a-CGRP cRNA probe did not hybridize. Pretreatment of the animals with either capsaicin or 6-OH dopamine did not affect the P-CGRP signal in the islet (not shown). It appears, therefore, that there are two CGRP systems in the rat pancreas: LU-CGRP in the neural innervation and P-CGRP locally synthesized within the endocrine cells.
FIGURE 2. Northern blot analysis of p- CGRP in, left to right: lane H, 40 pg total RNA from rat hypothalamus: lane P, 40 pg total RNA from rat estrogen-treated pitu- itary; / m e C, 20 pg rat islet poly A + from untreated rats; lane D, 20 pg rat islet poly A t RNA from DEX-treated rats.
COMPARISON OF IAPP AND CGRP ACTIONS WITHIN THE ISLET
Local synthesis of P-CGRP and IAPP in the pancreatic islet are consistent with a possible paracrine role in the regulation of pancreatic islet hormone function.38 Although P-CGRP appears to be the expressed form within the islet itself, all studies so far reported have looked only at the effects of a-CGRP on islet hormone s e ~ r e t i o n . ~ ~ - ~ ' However, as a- and P-CGRPs have very similar profiles of biological a ~ t i v i t y , ~ . ~ demonstration of an effect is probably valid for either form of peptide, with the effect mediated by the form endogenously present. Furthermore, since local levels of IAPP are certainly considerably higher than those of CGRP, it is uncertain how relevant any biological effect will be in uiuo: IAPP can act via CGRP receptors in the liver," and it is feasible that a certain amount of functional overlap may occur within the islet. Investigations into the actions of CGRP and IAPP in the islet have so far been largely inconclusive. While a number of actions have been reported, the effects of both peptides in uiuo are apparently very dependent upon local concentrations, time of exposure, and interactions with other regulatory substances.

BRETHERTON-WATT er af.: CGRP IN THE ISLET 303
In uitro, CGRP can act directly on isolated rat islets to reduce glucose-stimu- lated insulin release,".40 while the C-terminal fragment, CGRP18-37 can directly inhibit insulin release from isolated rat p -~e l l s .~ ' In whole perfused dog pancreas, CGRP inhibits insulin release at low concentrations, while at higher concentrations insulin is s t im~la ted .~ ' Although CGRP has been identified in somatostatin-con- taining and is a potent stimulator of somatostatin release from the gastroin- testinal tract,43 CGRP does not appear to affect SRIF release from the p a n ~ r e a s . ~ ~ . ~ ~
Similarly, IAPP can suppress insulin release from in uitrn preparations at very high local concentrations (lo-> M)40.45 and, in molar terms, higher than those required to see an effect with CGRP. No effect has been demonstrated at lower concentration^.^^,^^ In whole perfused pancreas, however, while human IAPP had no effect on insulin release,48 a lower concentration of rat IAPP was found to increase insulin and suppress prostaglandin (PG) and somatotropin release inhib- iting factor (SRIF) release.49 Even higher concentrations of rat IAPP suppressed insulin.s0 Recently, both IAPP and CGRP have been shown to be more potent (lo-' M) in suppressing carbachol-stimulated than glucose-stimulated insulin re- lease from isolated perfused rat pancreas.5'
IAPP and CGRP can, therefore, modulate pancreatic hormone secretion under certain conditions. However, because of the variability in responses seen, caution is needed in extrapolating these effects to an in uiuo physiological role.
PATHOPHYSIOLOGY OF IAPP AND CGRP IN INSULIN RESISTANCE AND DIABETES
Peripheral insulin resistance is a predominant feature of NIDDM.27 Both IAPP and CGRP can induce a state of insulin resistance in skeletal m ~ s c l e . ~ * ~ ~ ~ Possible mechanisms for the involvement of IAPP in the pathological process'z include: (a) deposition of amyloid causing disruption and impairment of insulin secretion; (b) increased secretion of IAPP causing peripheral insulin resistance; (c) direct action of IAPP upon insulin secretion within the islet. Since CGRP is also found in the islet and shares biological activity with IAPP, CGRP may also be implicated in NIDDM.
We have looked at changes in both IAPP and CGRP in the pancreas in animal models of insulin resistance, one a spontaneous model of diabetes, the ohloh mouse, and the other induced pharmacologically by dexamethasone.
Dexamethasone Treatment
Dexamethasone (DEX) is a synthetic steroid that leads to the development of peripheral insulin resistance. In order to compensate for the consequent hypergly- cemia, the output of insulin is increased, both by increasing individual p-cell secretory capacity, and, in rodents, by hyperplastic p-cell division.
Adult rats were treated with DEX by 12 daily intraperitoneal injections. For IAPP, either whole pancreatic RNA was extracted for Northern blotting, or pep- tides were solubilized for IAPP determination using a specific IAPP RIA (TABLE 1).53,54 For CGRP, however, islets were isolated by collagenase digestion and pooled prior to extracting either peptides or RNA.
As expected, DEX treatment increased both pancreatic insulin content and
304 ANNALS NEW YORK ACADEMY OF SCIENCES
TABLE t. Changes in IAPP and CGRP after DEX Treatment P-CGRP IAPP
Control DEX Control DEX mRNA (relative to control) 100% 39% I 00% 1600% Peptide content" 2.2 2 0.2 4.8 ? 0.1 657 f 176 1571 -t 137 Ratio peptideiinsulin ( X 1000) 0.61 2 0.3 0.68 i 0.05 85.2 t 8.4 61.7 i 6.0
" CGRP levels are expressed as fmol/islet; lAPP as prnolig wet weight pancreas.
mRNA levels. In the DEX-treated islet, however, CGRP mRNA levels were found to be decreased (TABLE 2, FIG. 2). On the other hand, pancreatic content of CGRP was found to increase; this increase was in parallel with insulin, as seen by an unchanged CGRP: insulin ratio (TABLE 2). Such a result suggests that, along with a decreased synthetic rate of the mRNA for CGRP, there is increased storage of peptide.
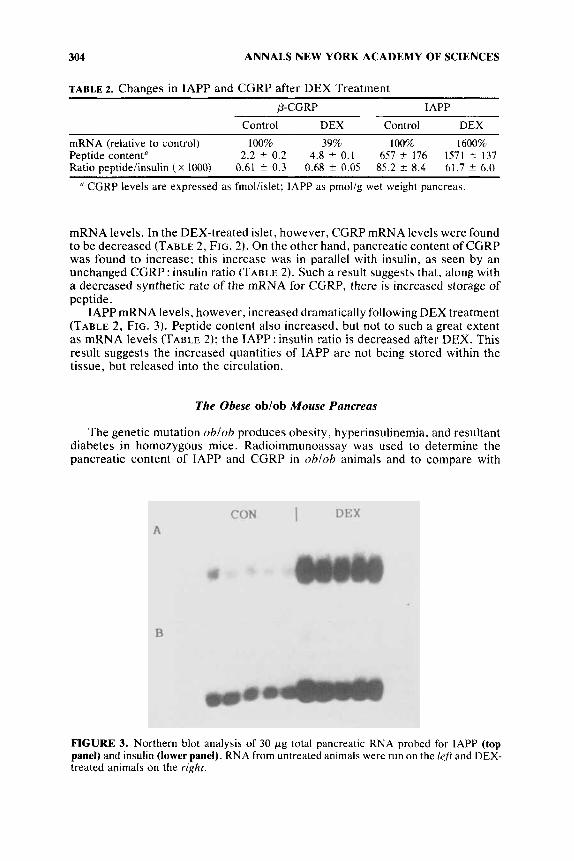
IAPP mRNA levels, however, increased dramatically following DEX treatment (TABLE 2, FIG. 3). Peptide content also increased, but not to such a great extent as mRNA levels (TABLE 2); the IAPP : insulin ratio is decreased after DEX. This result suggests the increased quantities of IAPP are not being stored within the tissue. but released into the circulation.
The Obese oblob Mouse Pancreas
The genetic mutation obloh produces obesity, hyperinsulinemia, and resultant diabetes in homozygous mice. Radioimmunoassay was used to determine the pancreatic content of IAPP and CGRP in ohlob animals and to compare with
FIGURE 3. Northern blot analysis of 30 pg total pancreatic RNA probed for IAPP (top panel) and insulin (lower panel). RNA from untreated animals were run on the left and DEX- treated animals on the right.

BRETHERTON-WATT ef al.: CGRP IN THE ISLET 305
normal littermates either following normal feeding ( n = 5 pairs) or an overnight fast ( n = 8 pairs). Pancreatic insulin content in oblob mice was fivefold higher than control. IAPP was also significantly increased (TABLE 3), while CGRP was unchanged.
In this model, again, IAPP showed a greater relative increase than insulin, as seen by the increased IAPP : insulin ratio (TABLE 3). CGRP content, however, was unchanged, resulting in a decreased CGRP : insulin ratio because of the increased insulin in the oblob mice, although this was significant only in the fasted group (TABLE 3).
While IAPP and CGRP have been shown to have similar effects on peripheral insulin resistance in vivo and in ~ i t r o , ~ ~ . ~ ~ the responses of the endogenous peptides to the development of spontaneous insulin resistance is very different. In particu- lar, the greater relative increase in IAPP compared to insulin is interesting, irre- spective of the initial cause of the insulin resistance. If this change is a direct response to the hyperglycemia, then a similar situation could be postulated to occur in NIDDM. This relative IAPP excess could be contributory to the disruption of both the secretion,ss and the peripheral actions of insulin. This exacerbation of the insulin resistance could lead to eventual arnyloid deposition and p-cell exhaustion.
TABLE 3. IAPP and CGRP in Pancreas of the ohloh Mouse IAPP Total CGRP
Mouse (pmol/panc) Ratio IAPP/ins (pmol/panc) Ratio CGRP/ins ( X 1000)
Fed Control 72 2 18 3.6 ? 0.3 0.7 ? 0.3 0.27 ? 0.13 ob 576 2 165* 6.8 rt 1.2* 0.6 t 0.2 0.08 t 0.04
Fasted Control 106 t 18 3.7 t 0.3 0.7 t 0.2 0.23 2 0.05 ob 1333 5 367* 9.2 ? 1.6* 1 . 1 ? 0.3 0.07 ? 0.03*
* = p < 0.01; n = 5 (fed), n = 8 (fasted).
ARE IAPP AND CGRP HORMONES?
In the circulation of normal, fasting subjects, levels of IAPP have been reported to be in the range of 2-10 pmolll, and to be altered in parallel with insulin in response to a range of physiological and pharmacological stirnuli.2’-23 The source of this circulating IAPP is assumed to be the pancreas, and the extremely low levels of IAPP immunoreactivity in insulin-dependent diabete~,’~ where p-cells are depleted, tends to agree with this.
CGRP immunoreactivity has been identified in plasma of normal rats6 and human.57 Total immunoreactivity is greater than IAPP, with levels of 20-30 pmolll in extracted plasma of both human and rat, with approximately 30% corre- sponding to synthetic CGRP on HPLC. Butler et a/.” measured levels of around 4 pmolil, similar to those of IAPP, and demonstrated that levels were suppressed postprandially. Plasma CGRP is thought to originate both from the thyroid gland

306 ANNALS NEW YORK ACADEMY OF SCIENCES
and as a result of "spillover" from perivascular nerves,56 although other sources cannot be excluded.
In human studies, infusions of CGRP sufficiently high to induce cardiovascular changes had no effect on basals8 or a r g i n i n e - s t i m ~ l a t e d ~ ~ ~ ~ ~ insulin or glucose levels, although CGRP did potentiate arginine-induced hyperglycemia." However, when administered in pigs directly via the pancreatic artery, CGRP suppressed both basal and stimulated-insulin responses and potentiated glucagon release.6' In the rat, an even higher dose decreased insulin response to glucose, while IAPP at the same concentration was i n e f f e ~ t i v e . ~ ~ Yamaguchi ct a/.62 found CGRP-induced hyperglycemia and elevated insulin in the rat that was not mediated via sympathetic mechanisms. In mouse33 or rat" the insulin response to glucose is suppressed with CGRP, resulting in hyperglycemia; although in the rat, basal insulin levels were increased by CGRP. These same concentrations of IAPP administered to both rat and mouse were inef fe~t ive .~ ' ,~ ' Similarly, no effect of IAPP was found on the insulin response to glucose in the rat o r rabbit,M despite the fact that circulating levels of 8 nmolil were achieved. In man, IAPP did not affect the insulin response to an intravenous glucose tolerance test (IVGTT). despite the fact that circulating levels of > 1 nmolil were achieved.6s
Subsequently, we investigated whether even higher doses of IAPP were capable of influencing peripheral insulin sensitivity in man. Six healthy, nonobese volunteers each received, on separate occasions and in random order, an intravenous infusion of either synthetic, human amidated IAPP (nominal doses 25 and 50 pmol k g - ' rnin-') o r normal saline. After a 20-minute infusion, an intravenous glucose bolus was given (0.5 gikg), and infusion continued for a further 30 minutes.
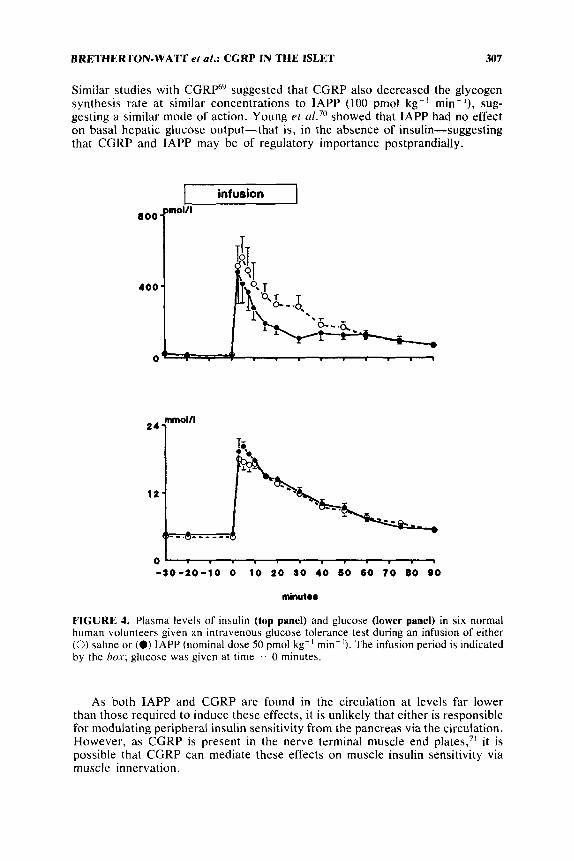
Blood samples were taken throughout the infusion for the measurement of insulin, glucose, and IAPP. Endogenous IAPP-like immunoreactivity did not increase above 30 pmolil after IVGTT, while plateau circulating levels of > 1 nmolil and >2 nmolil were achieved with the low- and high-dose infusions, respectively. IAPP had no effect upon peak plasma glucose, glucose clearance rate, o r overall response, as determined by the area under the curve of the response, during either infusion. However, while the insulin response to glucose was unchanged in the low-dose infusion, it was significantly decreased in the high-dose infusion (FIG. 4; low-dose infusion not shown). Thus, circulating insulin concentrations considerably greater than those found endogenously were necessary to affect the insulin response to glucose.
However, it is acknowledged that tests such as the IVGTT used in these studies are less sensitive than the euglycemic clamp studies: at high-glucose concentrations, glucoregulatory hormones become less important than the pre- vailing glucose concentration.6h In uiuo, hyperglycemic, euglycemic clamp stud- ies in rats demonstrated that CGRP and IAPP can both act via the circulation to cause a decreased glucose disposal rate.?9 Although very high concentrations (5 nmol kgg ' min-I) of both peptides were needed to induce these effects, IAPP and CGRP were found to be equipotent on peripheral muscle, while CGRP was more potent in reversing the insulin-suppressed hepatic glucose output. Two recent studies have looked at the dose-dependancy effects of IAPP in this system. Koopmans el a/.'' recently demonstrated that during hyperglycemic, euglycemic clamps, the liver was sensitive to IAPP at lower concentrations (60 pmol kgg ming I) than peripheral muscle. Conversely, Frontoni et found that IAPP decreased the glucose disposal rate in muscle (as seen by a decreased rate of muscle glycogen synthesis) a t concentrations lower than those required to reverse insulin-suppressed hepatic glucose output (50 pmol kg- ' min- I ) .
BRETHERTON-WATT et al.: CGRP IN THE ISLET 307
Similar studies with CGRP69 suggested that CGRP also decreased the glycogen synthesis rate at similar concentrations to IAPP (100 pmol kg-' min-I), sug- gesting a similar mode of action. Young et ~ 1 1 . ' ~ showed that IAPP had no effect on basal hepatic glucose output-that is, in the absence of insulin-suggesting that CGRP and IAPP may be of regulatory importance postprandially.
infusion
600 fm"'
0 ' . . . I . . . . . . - 1
-30-20-10 0 10 2 0 SO 40 50 60 7 0 80 90
minute.
FIGURE 4. Plasma levels of insulin (top panel) and glucose (lower panel) in six normal human volunteers given an intravenous glucose tolerance test during an infusion of either (0) saline or (0) IAPP (nominal dose 50 pmol kg-' rnin-'). The infusion period is indicated by the box; glucose was given at time = 0 minutes.
As both IAPP and CGRP are found in the circulation at levels far lower than those required to induce these effects, it is unlikely that either is responsible for modulating peripheral insulin sensitivity from the pancreas via the circulation. However, as CGRP is present in the nerve terminal muscle end plates," it is possible that CGRP can mediate these effects on muscle insulin sensitivity via muscle innervation.

308 ANNALS NEW YORK ACADEMY OF SCIENCES
Within the islet, accumulation of IAPP and CGRP by local release into the islet interstitium may reach sufficiently high concentrations to effect an auto- crinelparacrine role on pancreatic secretion. Plasma levels of both peptides may also be considerably higher in the portal vein, and this has been suggested to be the case for CGRP.6’ It is possible, therefore, that levels of CGRP and/or IAPP may reach those necessary to influence liver hepatic glucose output.
SUMMARY
Following the discovery of a second gene containing a CGRP-like sequence, we demonstrated that “0-CGRP” was indeed translated as a 37-amino acid peptide in uiuo and was the predominant form of CGRP produced by the enteric nervous system. The presence of CGRP in the islet has been reported by several groups. We now show that P-CGRP is again the major form.
Another 37-amino acid peptide was recently isolated from islet amyloid depos- its and found to have approximately 50% amino acid sequence homology with CGRP. Islet amyloid polypeptide, or amylin, is co-localized with insulin to the 0- cell secretory granule and is synthesized and released in parallel with insulin in response to a range of physiological and pharmacological stimuli.
IAPP was subsequently shown, like CGRP, to inhibit the release of insulin pharmacologically. Interestingly, it was also shown to decrease the uptake of glucose by striated muscle, though it was considerably less potent than CGRP. This led to the suggestion that IAPP might be a circulating hormone regulating peripheral insulin sensitivity. Infusion of IAPP in human volunteers to produce plasma concentrations more than 100-fold higher than those seen physiologically, however, failed to alter peripheral glucose disposal.
We conclude that P-CGRP and IAPP are likely to play a role in local paracrine control of the islet.
ACKNOWLEDGMENTS
We would like to thank Dr. S. Legon for the IAPP probe used in the DEX study, J . Byrne for help with the Northern blotting, Drs. A. Macrae and J. Beacham for help with the human infusion studies and Dr. R. A. L. Batt (Royal Veterinary College, London) for the oblob mice.
REFERENCES
1. ROSENFELD, M. G., S . G . AMARA, B. A.Roos,E.S.ONG&R.M.EVANS. 1981. Altered expression of the calcitonin gene associated with RNA polymorphism. Nature 290:
2. GOODMAN, E. C. & L. L. IVERSEN. 1986. Calcitonin gene-related peptide: Novel neuropeptide. Life Sci. 38: 2169-2178.
3. ZAIDI, M., H. BREIMER & I. MACINTYRE. 1987. Biology of peptides from the calcitonin genes. Quart. J . Exp. Physiol. 72: 371-408.
4. AMARA, S. G . , J. L. ARRIZA, S. E. LEFF, L. W. SWANSON, R. M. EVANS & M. G. ROSENFELD. 1985. Expression in brain of a messenger RNA encoding a novel neuropeptide homologous to calcitonin gene-related peptide. Science 229: 1094- 1097.
63-65.

BRETHERTON-WATT et al.: CGRP IN THE ISLET 309
5 .
6.
7.
8.
9.
10.
1 1 .
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
MULDERRY, P. K., M. A. GHATEI, R. A. SPOKES, P. M. JONES, A. M. PIERSON, Q. A. HAMID, S. KANSE, J. M. BURRIN, S. LEGON, J. M. POLAK & S. R. BLOOM. 1988. Differential expression of a-CGRP and P-CGRP by primary sensory neurons and enteric autonomic neurons of the rat. Neuroscience 25: 195-205.
WILLIAMS, G., H. CARDOSO, J. A. BALL, P. K. MULDERRY, E. COOKE & S. R. BLOOM. 1988. Potent and comparable vasodilator actions of A and B calcitonin gene-related peptides on the superficial subcutaneous vasculature of man. Clinical Sci. 75:
WESTERMARK, P., C. WERNSTEDT, E . WILANDER & K. SLETTEN. 1986. A novel peptide in the calcitonin gene-related peptide family as a amyloid fibril protein in the endocrine pancreas. Biochem. Biophys. Res. Comrnun. 140: 827-831.
G. J . S. COOPER, A. C. WILLIS, A. CLARK, R. C. TURNER, R. B. SIM & K. B. M. REID. 1987. Purification and characterisation of a peptide from amyloid-rich pancreases of type 2 diabetic patients. Proc. Natl. Acad. Sci. USA 84: 8628-8632.
NISHI, M., T. SANKE, S. NAGAMATSU, G. I. BELL & D. F. STEINER. 1990. Islet amyloid polypeptide: A new P cell secretory product related to islet amyloid deposits. J. Biol. Chem. 265: 4173-4176.
MOSSELMAN, S. , J. W. M. HOPPENER, J . ZANDBERG, A. D. M . VAN MANSFIELD, A. H. M . GEURTS VAN KEUSSEL, C. J. M. LIPS & H. S. JANSZ. 1988. Islet amyloid polypeptide: Identification and chromosomal localisation of the human gene. FEBS Lett. 239: 227-232.
MORISHTA, T., A. YAMAGUCHI, T. FUJITA & T. CHIBA. 1990. Activation of adenylate cyclase by islet amyloid polypeptide with COOH-terminal amide via calcitonin gene-related peptide receptors on rat liver membranes. Diabetes 39: 875-877.
CLARK, A., G. J. S. COOPER, C. E. LEWIS, J. F. MORRIS, A. C. WILLIS, K. B. M. REID & R. C. TURNER. 1987. Islet amyloid formed from diabetes-associated peptide may be pathogenic in type 2 diabetes. Lancet 1: 231-234.
WESTERMARK, P., U. ENGSTROM, K. H. JOHNSON, G. T. WESTERMARK & C. BET- SHOLTDz. 1990. Islet amyloid polypeptide: Pinpointing amino acid residues linked to amyloid fibril formation. Proc. Natl. Acad. Sci. USA 87: 5036-5040.
JOHNSON, K. H., T. D. O’BRIEN, C. BETSHOLTZ & P. WESTERMARK. 1989. Islet amyloid, islet amyloid polypeptide and diabetes mellitus. N. Engl. J . Med. 321:
WESTERMARK, P., E . WILANDER, G. T . WESTERMARK & K. H. JOHNSON. 1987. Islet amyloid polypeptide-like immunoreactivity in the islet B cells of Type 2 (non-insulin- dependent) diabetic and non-diabetic individuals. Diabetologia 30: 887-892.
LUKINIUS, A., E. WILANDER, G. T. WESTERMARK, U . ENGSTROM & P. WESTERMARK. 1989. Co-localisation of islet amyloid polypeptide and insulin in the B cell secretory granules of the human pancreatic islets. Diabetologia 32: 240-244.
NAKAZATO, M., J . ASAI, M. MIYAZATO, S. MATSUKURA, K. KANGAWA, & H. MATSUO. 1990. Isolation and identification of islet amyloid polypeptide in normal human pan- creas. Regul. Pept. 31: 179-186.
ASAI, J . , M. NAKAZATO, K. KANAGAWA, H. MATWO& S. MATSUKURA. 1990. Regional distribution and molecular forms of rat islet arnyloid polypeptide. Biochem. Biophys. Res. Comrnun. 169: 788-795.
TOSHIMORI, H., R. NARITA, M. NAKAZATO. J. ASAI, T. MITSUKAWA, K. KANGAWA, H. MATSUO & S. MATSUKURA. 1990. Islet amyloid polypeptide (IAPP) in the gastroin- testinal tract and pancreas of man and rat. Cell Tissue Res. 262 401-406.
FERRIER, G. J. M., A. M. PIERSON, P. M. JONES, S. R. BLOOM, S. I . GIRGIS & S. LEGON. 1989. Expression of the rat amylin (lAPP/DAP) gene. J . MoI. Endocrinol.
BUTLER, P., J . CHOU, W. B. CARTER, Y-N. WANG, B-H. Bu, D. CHANG & R. A. RIZZA. 1990. Effects of meal ingestion on plasma amylin concentration in NlDDM and nondiabetic humans. Diabetes 39: 752-756.
MITSUKAWA, T., J. TAKEMURA, J. ASAI, M. NAKAZATO, K. KANGAWA, H. MATSUO & S. MATSUKURA. 1990. Islet amyloid polypeptide response to glucose, insulin and somatostatin analogue administration. Diabetes 39: 639-642.
309-3 13.
5 13-5 18.
3: RI-R4.

310 ANNALS NEW YORK ACADEMY OF SCIENCES
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
HARRTER, E., T. SVOBODA, B. L U D V I K , M. SCHULLER, B. LELL, E. KUENBERG, M. BRUNNRAUER, W. WOLOSZCZUK & R. PRAGER. 1991. Basal and stimulated plasma levels of pancreatic amylin indicate its cosecretion with insulin in humans. Diabeto- logia 34: 52-54.
FEHMANN, H-C., V. WEBER, R. GOKE, B. GOKE & R. ARNOLD. 1990. Co-secretion of amylin and insulin from isolated rat pancreas. FEBS Lett. 262 279-28 1.
OGAWA, A,, V. HARRIS, S. K. MCCORKLE, R. H. UNGER & K. L. LUSKEY. 1990. Amylin secretion from the rat pancreas and its selective loss after streptozotocin, treatment. J. Clin. Invest. 85: 973-976.
INOUE, K., A. HISATOMI, F. UMEDA & H. NAWATA. 1990. Amylin release from perfused rat pancreas in response to glucose and arginine. Diabetes Res. Clin. Pract.
DEFRONZO, R. A. 1987. The triumvirate: p-cell, muscle, liver. A collusion responsible for NIDDM. Diabetes 37: 667-687.
LEIGHTON, B. & G. J. S. COOPER. 1988. Pancreatic amylin and calcitonin gene-related peptide cause resistance to insulin in skeletal muscle in vitro. Nature 335: 632-635.
MOLINA, J. M., G. J . S . COOPER, B. LE~GHTON & J. M. OLEFSKY. 1990. Induction of insulin resistance in vivo by amylin and calcitonin gene-related peptide. Diabetes
KAHN, S. E., D. A. D’ALESSIO, M. W. SCHWARTZ, W. Y. FUJIMOTO, J. W. ENSINCK, G. J . TABORSKY, JR. & D. PORTE, JR. 1990. Evidence of cosecretion of islet amyloid polypeptide and insulin by p-cells. Diabetes 39: 634-638.
KANATSUKA, A., H. MAKINO, H. OHSAWA, Y . TOKUYAMA, T. YAMAGUCHI, S. YOSHIDA, & M. ADACHI. 1989. Secretion of islet amyloid polypeptide in response to glucose. FEBS Lett. 259: 199-201.
MULDERRY, P. K., M. A. GHATEI, Y . S. ALLEN, J. M. POLAK & S. R. BLOOM. 1985. Distribution and chromatographic characterisation of CGRP-like inmunoreactivity in the brain and gut of the rat. Regul. Pept. 12: 133-143.
PETTERSSON, M., B. AHREN, G. BOTTCHER & F. SUNDLER. 1986. Calcitonin gene-re- lated peptide: Occurrence in pancreatic islets in the mouse and the rat and inhibition of insulin secretion in the mouse. Endocrinology 119: 865-869.
STERNINI, C., J . R. REEVE, JR. 8z N. BRECHA. 1987. Distribution and characterisation of calcitonin gene-related peptide immunoreactivity in the digestive system of normal and capsaicin-treated rats. Gastroenterology 93: 852-862.
STERNINI, C. & J . P. CARD. 1991. Ultrastructural localisation of calcitonin gene-related peptide-containing fibers and islet cells in the rat pancreas. Pancreas 6: 375-384.
LACY, P. E. & M. KOSTIANOVSKY. 1967. Method for the isolation of intact islets of Langerhans from the rat pancreas. Diabetes 16: 35-39.
CHOMCZYNSKI, P. & N. SACCHI. 1987. Single-step method of RNA isolation by acid- guanidium-thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162: 156-159.
WEIR, G. C. & S. BONNER-WEIR. 1990. Islets of Langerhans: The puzzle of intraislet interactions and their relevance to diabetes. J. Clin. Invest. 85: 983-987.
ISHIZUKA, J. , G. H. GREELEY, JR., G. W. COOPER & J. C. THOMPSON. 1988. Effect of calcitonin gene-related peptide on glucose and gastric inhibitory polypeptide-stimu- lated insulin release from cultured newborn and adult rat islet cells. Regul. Pept. 20:
OHSAWA. H., A. KANATSUKA, T. YAMAGUCHI, H. MAKINO & S. YODHIDA. 1988. Islet amyloid polypeptide inhibits glucose-stimulated insulin secretion from isolated rat pancreatic islets. Biochem. Biophys. Res. Commun. 160: 961-967.
HERMANSEN, K. & B. AHREN. 1990. Dual effects of calcitonin gene-related peptide on insulin secretion in the perfused dog pancreas. Reg. Pept. 27: 149-157.
LEWIS, C. E., A. CLARK, S. J. H. ASHCROFT, G. J . S. COOPER & J . F. MORRIS. 1988. Calcitonin gene-related peptide and somatostatin inhibit insulin release from individual rat B cells. Mol. Cell. Endocrinol. 57: 41-49.
DUNNING, B. E. & G. J . TABORSKY. 1987. Calcitonin gene-related peptide: A potent and selective stimulator of gastrointestinal somatostatin secretion. Endocrinology 120 1774-1781.
10: 189-192.
39: 260-265.
73-82.

BRETHERTON-WATT el al.: CGRP IN THE ISLET 311
44. FUJIMURA, M., G. H. GREELEY, JR., M. B. HANCOCK. A. ALWARK, A. SANTOS, C. W. COOPER, K. J. REUMONT, J. ISHIZUKA & J. C. THOMPSON. 1988. Colocalisation of calcitonin gene-related peptide and somatostatin in pancreatic islet cells and inhibition of insulin secretion by calcitonin gene-related peptide in the rat. Pancreas 3: 49-52.
AR’RAJAB, A. & B. AHREN. 1991. Effects of amidated rat islet amyloid polypeptide on glucose-stimulated insulin secretion in vivo and in vitro in rats. Eur. J . Pharmacol.
TEDSTONE, A. E., T. NEZZER, S. J . HUGHES, A. CLARK & D. R. MATTHEWS. 1990. The effect of islet amyloid polypeptide (amylin) and calcitonin gene-related peptide on glucose removal in the anaesthetised rat and on insulin secretion from rat pancre-
45.
192: 443-445. 46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
atic-islets in vitro. Biosc. Rep. 10: 339-354. PETTERSSON, M. & B. AHREN. 1990. Failure of islet amyloid polypeptide to inhibit
basal and glucose stimulated insulin secretion in model experiments in mice and rats. Acta Physiol. Scand. 138: 389-394.
O’BRIEN, T. D., P. WESTERMARK & K. H. JOHNSON. 1990. Islet amyloid polypeptide (IAPP) does not inhibit glucose-stimulated insulin secretion from isolated perfused rat pancreas. Biochem. Biophys. Res. Commun. 170: 1223-1228.
FEHMANN, H-C., V . WEBER, R. GOKE, B. GOKE, R. EISSELE & R. ARNOLD. 1990. Islet amyloid polypeptide (IAPP; amylin) influences the endocrine but not the exocrine rat pancreas. Biochem. Biophys. Res. Commun. 167: 1102-1 108.
SILVESTRE, R. A., E. PEIRO, P. DEGANO, P. MIRALLES & J. MARCO. 1990. Inhibitory effect of rat amylin on the insulin responses to glucose and arginine in the perfused rat pancreas. Kegul. Pept. 31: 23-31.
KOGIRE, M., J . ISHIZUKA, J . C . THOMPSON & G. H. GREELEY, JR. 1991. Inhibitory action of islet amyloid polypeptide and calcitonin gene-related peptide on release of insulin from the isolated perfused rat pancreas. Pancreas 6 459-463.
JOHNSON, K. H., T. D. O’BRIEN & P. WESTERMARK. 1991. Newly identified pancreatic protein, islet amyloid polypeptide: What is its relationship to diabetes? Diabetes 4 0
BRETHERTON-WATT, D., M. A. GHATEI, S. R. BLOOM, G. J. M. FERRIER, S. I. GIRGIS & S. LEGON. 1989. Altered islet amyloid polypeptide (amylin) gene expression in rat models of diabetes. Diabetologia 32: 88 1-883.
JAMAL, H., D. BRETHERTON-WATT, K. SUDA, M. A. GHATEI & S. R. BLOOM. 1990. Islet amyloid polypeptide-like immunoreactivity (amylin) in rats treated with dexa- methasone and streptozocin. J. Endocrin. 126: 425-429.
POLONSKY, K . S., B. D. GIVEN, L. J. HIRSCH, H. TILLIL, E. T. SHAPIRO, C. BEEBE, B. H. FRANK, J. A. GALLOWAY & E. VAN CAUTER. 1988. Abnormal patterns of insulin secretion in non-insulin dependent diabetes mellitus. N. Engl. J. Med. 318 1231-1239.
ZAIDI, M., P. J . R. BEVIS, G. ABEYASEKERA, S. I . GIRGIS, S. J . WIMALAWANSA, H. R. MORRIS & 1. MACINTYRE. 1986. The origin of circulating calcitonin gene-re- lated peptide in the rat. J . Endocrinol. 110: 185-190.
GIRGIS, S. I., D. W. R. MACDONALD, J. C. STEVENSON, P. J. R. BEVIS, LYNCH, C., S. J. WIMALAWANSA, C. H. SELF, H. R. MORRIS & I . MACINTYRE. 1985. Calcitonin gene-related peptide: Potent vasodilator and major product of calcitoningene. Lancet 2(8445): 14-16.
KRAENZLIN, M. E., J. L. C. CH’NG, P. K. MULDERRY, M. A. GHATEI & S. R. BLOOM. 1985. Infusion of a novel peptide, calcitonin-gene-related peptide (CGRP) in man. Pharmacokinetics and effects on gastric acid secretion and on gastrointestinal hor- mones. Regul. Pept. 18: 189-197.
BEGLINGER, C . , E. KOEHLER, W. BORN, J. A. FISCHER, U. KELLER, L. E. HANSSEN & K. GYR. 1988. Effect of calcitonin gene related peptide on pancreatic functions in man. Gut 29: 243-248.
AHREN. B. 1990. Effects of galanin and calcitonin gene-related peptide on insulin and glucagon secretion in man. Acta Endocrinol. 123: 591-597.
AHREN, B., H. MARTENSSON & A. NOBIN. 1987. Effects of calcitonin gene-related peptide (CGRP) on islet hormone secretion in the pig. Diabetologia 3 0 354-359.
3 10-3 14.
YAMAGUCHI, A, , T. CHIBA, T. MORISHTA, A. NAKAMURA, T. 1NU1, T. YAMATANI, S.

312 ANNALS NEW YORK ACADEMY OF SCIENCES
63.
64.
65.
66.
67.
68.
69.
70.
71.
KADOWAKI, K. CHtHAR.4, M. FUKASE & T. FUJITA. 1990. Calcitonin gene-related peptide and induction of hyperglycemia in conscious rats in vivo. Diabetes 39:
PETTERSSON, M. & B. AHREN. 1988. Insulin and glucagon secretion in rats: Effects of calcitonin gene-related peptide. Regul. Pept. 23: 37-50.
GHATEI, M. A., H. K . DATTA, M. ZAIDI , D. BRETHERTON-WATT, S. J . WIMALA- WANSA, I . MAC~NTYRE & S. R. BLOOM. 1990. Amylin and amylin-amide lack an acute effect on blood glucose and insulin. J . Endocrinol. 124: R9-Rll.
BRETHERTON-WATT, D., S. G. GILBEY, M. A. GHATEI, J. BEACHAM & S. R. BLOOM. 1990. Failure to establish islet amyloid polypeptide (amylin) as a circulating beta cell inhibiting hormone in man. Diabetologia 33: 115-1 17.
BEST, J. D., G. J. TABORSKY, J. B. HALTER & D. PORTE, JR. 1981. Glucose disposal is not proportional to plasma glucose level in man. Diabetes 3 0 847-850.
RADDER, M. FROHLICH, S. F. DE BOER, D. K. KREUTTER, G. C. ANDREWS & J. A. MAASSEN. 1991. Amylin-induced in vivo insulin resistance in conscious rats: The liver is more sensitive to amylin than peripheral tissues. Diabetologia 34: 218-224.
FRONTONI, S., s. B. CHOI, D. BANDUCH &L. ROSSETTI. 1991. Invivoinsulinresistance induced by amylin primarily through inhibition of insulin-stimulated glycogen synthe- sis in skeletal muscle. Diabetes 40: 568-573.
CHOI, S. B., S. FRONTONI & L. ROSSETTI. 1991. Mechanism by which calcitonin gene-related peptide antagonises insulin action in vivo. Am. J . Physiol. 2 3
YOUNG, D. A., R. 0. DEEMS, R. W. DEACON, R. H. MC~NTOSH & J. E . FOLEY. 1990. Effects of amylin on glucose metabolism and glycogenolysis in vivo and in vitro. Am. J. Physiol. 259: E457-E461.
TAKAMI, K.. Y. KAWAI, S. UCHIDA, M. TOYAMA, Y. SHIOTANI, H. YOSHIDA, P. C. EMSON, s. GIRGIS, c. J. HIL.I.YARD & 1. MACINTYRE. 1985. Effect of calcitonin gene-related peptide on contraction of striated muscle in the mouse. Neurosc. Lett. 60: 227-230.
168- 174.
KOOPMANS, S. J., A. D. M. VAN MANSFIELD, H. S. JANSZ, H. M. J. KRANS, J. K.
E231-E235.




















