
Chronology of Events: Conflict in the Chittagong Hill Tracts

SPECIAL SERIES
THE PATHOLOGY OF HEAD AND NECK TUMORS: PAPILLOMAS OF THE UPPER AERODIGESTIVE TRACTS, PART 18
JOHN G. BATSAKIS, MD, A. KEVIN RAYMOND, MD, and DALE H. RICE, MD
Abstract: The human papillomavirus class. of DNA vi- ruses are more than circumstantially related to oral and airway papillomas. Whether they are fully on- cogenic, in the malignant sense, without other agents is questionable. Recent advances in molec- ular virology and the use of genus-specific (com- mon) antigen-antibody reactions have identified papillomavirus in laryngeal and oral papillomas. Laryngeal and oral papillomas could be precancer- ous lesions, but they show a low-risk and long-time interval leading to malignancy unless significant iatrogenic or host variables-such as radiotherapy or immuno-incompetence-are added. Two differ- ent papillomas in the larynx can be recognized; a nonkeratinizing, papillomavirus-related lesion, and a keratinizing, usually solitary papilloma, which may or may not be related to a virus genesis and may be equated with a keratosis or clinical leu- koplakia.
HEAD 81 NECK SURGERY 5:332-344 1983
The literature on papillomas of the upper aerodigestive tracts in humans can be conve- niently divided into three categories: 1) etiology; 2) management of afflicted patients; and 3) the relationship of papillomas to malignancy. All three are in various stages of exposition and ac- ceptance. This report concentrates on the his- togenesis and pathology of papillomas, and thereby presents a summary of our recently ac- quired knowledge of the etiology and neoplastic qualities of these lesions.
Although it has not been conclusively estab- lished, the histogenesis of aerodigestive tract
papillomas is intimately related to specific vi- ruses and, therefore, a brief preamble on the molecular biology of these viruses is necessary.
MOLECULAR BIOLOGY OF DNA VIRUSES
Virus-induced neoplasms have been described among members of nearly all classes of verte- brates. A diverse collection of viruses have been implicated, but there is no obvious correlation be- tween the taxonomic status of the virus and the type of tumor it induces.' Members of all major subdivisions of the viruses containing deoxy- ribonucleic acid (DNA) have been implicated in oncogenesis (Table 1). Historically, only one fam- ily of the viruses containing ribonucleic acid (RNA), the retroviruses, have been found to have oncogenic potential.' We are concerned only with the papovaviruses, and their involvement in pap- illomas of the laryngo-tracheal-bronchial tree.
Papovaviruses. The papovaviruses are small, DNA-containing viruses that belong to the family
From the Department of Pathology The University of Texas M D Ander- son Hospital and Tumor Institute Houston TX (Drs Batsakis and Ray- mond) and the Department of Head and Neck Surgery The Veterans Administration Wadsworth Medical Center Los Angeles CA (or Rice)
Address reprint requests to Dr Batsakis at the Department of Pathology M D Anderson Hospital and Tumor Institute, Houston TX 77030
Accepted for publication September 16 1982
01 48-64031050410332 $01 2510 1983 John Wiley & Sons Inc
332 The Pathology of Head a n d Neck Tumors HEAD & NECK SURGERY MariApr 1983

Table 1. Oncogenic DNA-containing viruses Table 2. Papovaviridae family of DNA-containing viruses.
Family Virus Oncogenic potential Genus papillomavirus Genus polyomavirus
Papovavirus Papilloma virus Papillomas, condy- Rabbit papilloma Mouse polyoma __
lomas; may pro- Rabbit oral papilloma Simian virus 40 (SV40) gress to Human papilloma (wart) Rabbit vacuolating virus
nasopharyngeal carcinoma
Canine papilloma Human BK virus Bovine papilloma Human JC virus
Herpes virus Epstein-Barr virus Burkitt's lymphoma,
Herpes simplex 1 Squamous cell car- Others cinoma (7)
Herpes simplex 2 Cervical carcinoma (7 )
virus ( 7 )
larger (Fig. 1). Virions of each genus member are nonenveloped icosahedral capsids, each contain- ing a circular double-standard DNA genome. The papillomaviruses measure approximately 55 nm in diameter and contain a genome with a molecu- lar weight of approximately 5 x 106.5 The poly-
Hepatitis Hepatitis B Hepatocarcinoma
Poxvi rus Mol I uscu m Nodular epidermal contag iosum hyperplasia (not
neoplastic)
Papovaviridae.2 The name of this virus family is derived from the first two letters of three of its members, papilloma virus, polyoma virus, and uacuolating agent, which was an early name for one of its types. This family is unusual among the DNA viruses in that most of its members are be- lieved to be oncogenic, or potentially so.
The papovavirus family consists of two genera (Tables 2 and 3), the papillomaviruses and the polyoma~iruses .~~~ Of the two, much more is understood about the molecular biology of the polyomaviruses. One of the bases for the classification of these viruses into two genera is their respective sizes; the papillomavirus is
omaviruses measure approximately 40 nm in di- ameter, and contain a 3.3 x lo6 genome.
There are other differences that exist between the two genera. The papillomaviruses have not been successfully propagated in tissue culture and, in general, they produce or promote benign epithelial proliferations in their hosts. The poly- omaviruses can be readily propagated in tissue culture and are not associated with any wide- spread, spontaneous, pathologic alteration in their natural hosts.
The classes within the genera have im- munologically distinct surface antigens. They have few, if any, DNA sequence homology, and exhibit distinguishing restriction endonuclease
Table 3. Papovavirus group (papovaviridae)
Genera Papi I lomavirus Polyomavirus
Family characteristics Nucleic acid: One molecule double-stranded DNA Morphology: Nonenveloped icosahedral particles 45 to 55nm di-
ameter; 72 capsomers in skew arrangement: filamentous forms occur Different species antigenically distinct: common anti- genic determinants shared by some and detectable Cytolytic in cells of host origin: may transform cells of other species; no tissue culture systems for papil- loma viruses Virions transported to nucleus and cellular DNA syn- thesis is stimulated; expression of viral genome in early and late stages: host histones incorporated into virions; virions released by virus Each virus has own host range. Transformation tends to occur in cells unable to support replication of virus
Antigenic properties: Effects on cells:
Replication:
Host range:
Papillomavirus Mol wt of DNA = 5 x lo6; diameter of capsid = 55 nm No tissue cul- ture svstem
The Pathology of Head and Neck Tumors HEAD & NECK SURGERY MariApr 1983 333

Papovaviridae Adenoviridae Herpesviridae Poxviridae (Shope papiilomai (Human Adeno 2 ) (Herpes simplex) ivaccinla)
Figure 1 . Relative sizes of four families of animal viruses
cleavage patterns of their genomes. However, it has been found that each genus of the family pos- sesses a common antigenic determinant that is likely genus-specific, since there is no demonstra- ble immunologic relationship between papil- lomaviruses and polyomavir~ses.~,~ This lack of a relationship is further supported by studies of the homologies of the DNAs of these viruses. The common antigen of the papillomaviruses resides in the internal capsule of the virus. Antibodies are producible from disrupted viral capsids.6
Viral Infection. While the accumulated informa- tion about the papovaviruses equals and, in some instances, surpasses that of any other group of animal DNA viruses, we have only a limited understanding of their in vivo infection. The vi- rus antigen is yet to be demonstrated in clinically normal epithelium.
Most encounters between viruses and human cells do not produce clinical disease-although cellular disease may occur. The type of disease is also dependent on whether the cell itself is killed during intracellular reproduction of the virus.
It is probable that the majority of humans are infected early in life, but primary infection is rarely associated with a major illness. It is also probable that the viral infections persist; if they are to remain for long periods, persistent viruses cannot afford to be pathogenic. The most success- fully persistent viruses are ancient associates of the host species and exist in a state of balanced pathogenicity. The papovaviruses and cytomega- loviruses in humans are successfully persistent parasites and yield few, if any, signs of a. pres- ence.’
Figure 2 illustrates the several phases of cell infection by viruses containing DNA or RNA genomes, regardless of whether they reproduce in the nucleus or the cytoplasm.’ No disease is pro- duced if: 1) the virus cannot attach to the plasma membrane of the cell; 2 ) the virus cannot pene- trate the cell; or 3) after successful penetration, the viral genome is not activated and/or the virus is destroyed or functionally inactivated. If the vi- ral genome is activated, but the virus does not reproduce and the cell is not killed (persistent in- fection), the type of disease is dependent on the type of persistent infection. Usually, if there is no perturbation of the cell, no disease is produced.’
A disease will most often develop after viral infection has altered cell structure or cell func- tion, or both. There are at least five types of in- teractions that lead to clinical disease’: 1) im- mune assault on the changed plasma membranes of cells; 2) immune response directed primarily
Attachment of Virus to Cell
I
I I
Penetration into Cell (Pinocytosis andlor fusion with plasma membrane)
Release of Virus into Cytoplasm and Uncoating of Virus Covering
Activation and Regulation of Virus Genome (New Viral RNA and Protein)
Production of Progeny Blocked Progeny Genomes and Viruses Produced
I I Persistent Infection
New Infective Viruses Released (With or Without Cell Death)
Figure 2. Phases of cell infection by wruses
334 The Pathology of Head and Neck Tumors HEAD & NECK SURGERY MariApr 1983

against extracellular viral proteins and nucleic acids (antigen-antibody, immune complex); 3) vi- ral alteration of the host cell's normal growth- control mechanisms; 4) viral alteration or block- age of normal stem-cell differentiation; and 5) viral-induced changes in cell function without change in the structure of the cell.
The virus-cell interaction between papil- lomaviruses and cells is probably one of viral genome activation, in which the viruses presum- ably interfere with cell differentiation and/or cell growth.
Virus persistence is intimately related to the ability of the virus to be shed to the outside world.8 Virus nucleic acid is present in all types of persistence, often with the virus antigens or in- fectious virus.
One type of persistence is characterized by a persistent infection, eg, subacute sclerosing pan- encephalitis. In this infection, there is no opportu- nity for virus shedding, and only viral nucleic acid sequences are present. A second type of per- sistence is associated with only sporadic shed- ding, which is a result of reactivation of the vi- ruses, such as reactivation of papovaviruses in the kidney. Finally, there are viruses that repli- cate and shed continuously to the outside world, often throughout the host's lifetime via the respi- ratory, urinary, and intestinal tracts. According to Mims,' the second and third types of persis- tence are important, in an evolutionary sense, in that both a maintenance of infection and persis- tence in small groups of individuals allow the vi- ruses to persist in the species.
Virus persistence is favored if as few antigens as possible are produced by the virus. In that re- spect, there is a notable tendency for persistent viruses to infect epithelial sites in glands and body surfaces- which either face, or have a conduit to, the outside world.8 From such loci, the viruses can be discharged and protected from circulating antibodies that fail to reach the luminal surfaces of infected cells where the infectious particles are released. It is not coincidental that viruses often manifest such a striking polarity in epithelia, as viral antigens are confined to the luminal surface of the cells. The infected cell can escape sensitized lymphocytes, which are unable to traverse both the basement membrane and the overlying cell mass unless there is a local inflammation and cell damage.
Antibody-induced antigenic modulation is one way in which cells infected with certain viruses can reduce expression of viral antigens on their
surfaces and also escape immunologic surveil- lance. At least two mechanisms of this type of modulation allow viruses to persist in the cells and escape immune lysis'o: 1) a reduction of the density of antigenic expression on the surface of the cell; and 2) alterations in internal viral pro- teins; thereby favoring future immune selection and the generation of viral mutants, which alone can cause virus persistence.
Certain persistent viruses reactivate periodi- cally. The factors that promote and support reac- tivation are not completely known, but among them are immunosuppression, pregnancy, and in- creasing age.' After (or coinciding with) reactiva- tion, the viruses undergo a period of renewed rep- lication in the host cell. When a reactivated virus is to be shed to the outside world (as discussed above), it will most often replicate in mucous membranes or skin.8
Viral Oncogenesis. Koch's postulate may not be fulfilled by so-called human tumor viruses be- cause most DNA virus infections are cytocidal, and neoplasia is a rare result of a common DNA virus infection. In addition, there is abundant epidemiologic evidence that, in many instances, there are multiple events which lead to the evolu- tion of a neoplasm. Viral infection may be only one of several precipitating factors. Indeed, where there is reasonable evidence for a viral involve- ment, the virus is a common human pathogen, which is neoplastic only in special circumstances or in combination with other factors.
Transformation (alteration in the morphology and behavior of cells) is the in vitro analogue of in vivo tumor formation.' Although transformed cells are often tumorigenic, only certain cell types can be transformed by any particular virus. Thus, cell transformation is not an inevitable outcome of tumor virus infection. This underscores the in- terplay of other factors, such as the environment and genetics.'
Particular subregions of DNA viral genomes are clearly associated with oncogenic potential and, in some instances, subgenomic, noninfec- tious pieces of viral DNA have the capacity to transform cells in vivo or in vitro.'','2 It is prob- able that virus particles carrying damaged or de- fective genomes, in which transforming subre- gions are intact, are the principal transforming particles in a virus population. Table 4l presents possible mechanisms of cell transformation by tumor viruses; a hit-and-run mechanism is nearly
The Pathology of Head and Neck Tumors HEAD & NECK SURGERY MariApr 1983 335

Table 4. Cell transformation by tumor viruses
Mechanism Viral qenome
Hit-and-Run
Hit-and-Remain Transformed cells contain all
Viral genome is absent from transformed cells
or part of viral genome Random or specific integration of viral DNA ablates or abnormally changes expression of host genes Integrated viral DNA contains or provides promoters for viral gene expression whose action affects host gene expression Integrated or free viral DNA encodes a pro- tein whose functioning causes cell transformation
excluded by findings of parts of the viral genome in transformed cells.'
The large-scale replication of DNA tumor vi- ruses is not compatible with survival of the in- fected cells. Therefore, transformed or inappar- ently infected cells do not produce appreciable amounts of virus. However, virus-producing cells usually undergo lysis. In papovaviruses, transfor- mation occurs because either the cell is nonper- missive for virus replication or the virus is both a mutant and defective in its ability to complete a replicative cycle in a permissive host.' In some cases, the virus-cell interaction is semipermissive in that cells transform, but a lytic infectious cycle also occurs, which maintains a low level of virus production.' This phenomenon is especially rele- vant regarding papillomavirus-infected cells in which virus replication is rarely found in the pro- liferating cells of the stratum malpighi, although it does occur mainly in the stratum corneum.
All this data confirms that there is an intri- cate and specific relationship between papovavir- uses and their host cells that has evolved to en- able the viruses to direct their own replication both in permissive cells and, through a complex symbiosis, in a nonpermissive environment.'
In summary, the oncogenic effect of a DNA virus is almost entirely dependent on the pro- longed latency of the viral genome and the sup- pression of most viral functions. As with all DNA, the genome of the DNA tumor virus has been selected for survival in their natural habitat. One or more viral genomes per cell are preserved in certain cells of the host, in a completely latent form, and multiply only in coordination with the host cell genome.
What the cell that harbors the viral genome in vivo undergoes may only be inferred by extrapo-
336 The Pathology of Head and Neck Tumors
lations from in vitro systems; that is, transforma- tion. Transformation is often only the first event in a complex series of events, each modified or even controlled by host factors.
Figure 3 is based on Chan's concept of the na- ture of oncogenic transformation of ce11s.l~ In es- sence, there is a class of genes within the genome of higher cells that function as growth genes. Through intricate programs of gene expression and repression that operate during development, growth genes elicit cell growth to normal phen- otypes. Given proper conditions, a postdevelop- mental expression of any growth gene can lead to oncogenic transformation. In viruses, the trans- formation event may occur within a growth gene that is either randomly or preferentially selected. The transformation gene is, in a sense, a mutant due to alterations of the base sequence of DNA and has its origin in cell genes.
The derepression of growth genes by mutation may be regarded as an initiation process. As pre- viously noted, additional measures beyond muta- tion, specifically promotion, are needed for the consummation of transformation. Promoters, such as other viruses and the environment, basi-
NORMAL STEM CELL - - - - - + CELL LINEAGE - - - - - -* PHENOTYPE
GENES
+
+
I GENOME REPROGRAMMING
f I \ k/ $. \r
STEM CELL ____* CELL LINEAGE - PHENOTYPE
Figure 3. Oncogenic transformation of cells after Chan 73 The combination of host growth genes, virus transforming gene, and promoters yield genomic reprogramming with the production of an abnormal (neoplastic) phenotype.
HEAD & NECK SURGERY MariApr 1983

cally operate by modulating the expression of the transformed phenotype. The combination of virus infection and promoters leads to both a constitu- tive expression of the growth gene from the virus and an unrestrained growth.
Polyoma and virus SV40 appear to mediate transformation via the functioning of viral gene products, some of which are necessary for the vi- rus to complete its cytolytic cell cycle. In contrast, transforming properties of RNA viruses result from their accidental acquisition of normal cel- lular genes that have no role in viral repli- cation.12J4
Papillomavirus warts and/or papillomas re- sult from an abortive infection of one or more epithelial germinative cell(s) which leads to a sta- ble and heritable transformation characterized by an altered response to factors controlling their number andlor mitotic rate. The transformation of germinative cells may be connected with per- sistence and a partial expression of one or several copies of viral genome-either integrated in the cell genome or as an autonomously replicating DNA. l5
Viral-DNA replication, detected by in situ molecular hybridization, is observed only in cells already in the process of keratinization (cells with cytoplasmic granules). Virus production may re- sult from a change in the permissivity of the cells as they undergo specific terminal differentiation on their way to the surface of the epithelium. The histopathologic correlate is that capsid proteins are found only in nuclei of the upper keratinizing cells and in keratinized cells, the very cells in which virus particles have been visualized with an electron r n i c r o s ~ o p e . ~ ~ ~ ~ ' ~ - ~ ~
Viral replication in laryngeal papillomas is exceptionally notable. Infection remains abortive in cytodifferentiating cells, and is revealed by the DNA synthesis that is observed in some of these cells. This contrasts with normal epithelium, in which DNA synthesis is restricted to basal and parabasal cells. It is possible that in laryngeal papillomas, abortive viral infection may prevent the complete inhibition of cell DNA replication which is expected a t the onset of the cytodif- ferentiation process; a demonstration of virus in- teraction in which mechanisms control DNA replication.
PATHOLOGY OF PAPILLOMAS
The lesions attributed to papillomavirus infection of mucosal cells are characteristic, both in gross and in histologic appearance. In the larynx, they
occur mainly on the true vocal cords, the false vocal cords, and in the anterior commissure. The papillomas may extend upward onto the epiglot- tis, pharyngeal walls, and soft palate, and also subglottically in the trachea and bronchi (see be- low). Involvement of the esophagus below the cri- copharyngeus muscle is rare.
Varying in color from whitish-pink to red, the papillomas are typically glistening, elevated, mulberry-like nodular masses (Fig. 4). They range in size from small nodules or sessile plaques, to tumors the size of a cherry.
Microscopically, papillomas contain all com- monly found epithelial cell layers, but in an exag- gerated squamous growth pattern. The sessile, or papillary, configuration of a papilloma is pro- duced by a combination of cell proliferation and an apparent increase in submucosal capillaries, which are arrayed in fibrovascular cores. Second- ary and tertiary branching of the epithelial- covered stalks also contributes to the papilloma- tous appearance.
In the condylomas of the genital tract-but less so in upper airway papillomas-a presumed viral replication within the cells near the epithe- lial surface results in a characteristic degenera- tion of nuclear chromatin." This is manifested as an irregular collapse of the nucleus and a sur-
Figure 4. Laryngeal papillomas slightly superior to the anterior cornrnissure.
The Pathology of Head and Neck Tumors HEAD & NECK SURGERY Mar/Apr 1983 337

rounding zone of cytoplasmic vacuolation. Such Papillornavirus
koilocytes or halo cells are regarded as being I nearly pathognomonic of papillomavirus infec- tions. Florid samples are easily found, but in lesser degrees. The distinction between koilocy- tosis and a cytoplasmic clearing produced by gly- cogen is very subjective.
Atypical Papillomas. In the typical laryngeal papilloma, microscopic evidence of proliferative cellular activity is confined to the deepest part of the epithelium, and is not accompanied by more than a rare mitosis or a cellular pleomorphism. This baseline provides a range of deviation, and Quick et a1.21 have related the magnitude of change to prognosis. They divided laryngeal papillomas of the nonkeratinizing type into two groups: 1) typical (as described above); and 2) atypical. Within the context of their evaluations, atypical referred principally to the width of the epithelium when occupied by cells presenting an appearance similar to that of the basal-parabasal layer of the mucosa, or of typical papilloma. Se- vere atypia was present in cases where almost all of the papilloma was composed of basal-parabasal type cells. Papillomas with moderate atypia man- ifested a similar cell proliferation throughout at least one half of the epithelium. Mild atypia is characterized by a basal-parabasal cell popula- tion in more than one third of the thickness of a papilloma. Mitoses increased with an increase in atypia.
Even though the correlative aspects of the study by Quick, et al.’l lack statistical strength, their findings are important and should be confirmed. Both typical papillomas and those with mild atypia were readily controlled, under- went spontaneous remission in more cases, and did not change their histologic character during the course of the disease. Atypical papillomas with moderate or severe atypia more often re- quired surgical removal. They also changed his- tologic character in relation to the crescendo of the disease, and showed a persistence with active disease for longer periods of time. Extensive le- sions often exhibited high degrees of atypia.
lnvasive Papillomas. There is no typical example that can describe the clinicobiologic behavior of a laryngeal papilloma. Spontaneous regression and relative ease of control are countered by persis- tent and refractory lesions that involve the lower airway (Fig. 5).
A small percentage of patients with laryngeal,
I Mucosa of Larynx, Trachea, Bronchi
Oncogenic (Transformation) Effect
Papillomas
“Remission” Recurrences
I I
-- ---\
Involvement
Post-radiation ”Spontaneous” Carcinoma
Figure 5. Possible consequences of papillomavirus infection of the mucosa of the upper airway.
Carcinoma
tracheal, or bronchial papillomatosis exhibit le- sions that extend beyond the mucosa, eg, into ad- jacent soft tissue, pulmonary or thyroid paren- chyma, and the kin.'','^ Such locally aggressive behavior is not accompanied by acceptable cy- tologic evidence of malignancy o r by a metastatic capability. The extra-mucosal extensions are usu- ally seen in continuity with surface or intralu- minal papilloma. Nearly all invasive papillomas of the larynx have been subjected to prior radio- therapy.” In a few cases, the apparent invasion is due to an outgrowth of the papillomas through surgical defects such as a tracheostomy stoma. In some others, invasion appears to follow normal microanatomic pathways, such as ducts and mucosal folds. This type of growth is similar to that of inverting Schneiderian papillomas of the sinonasal tract, or of verrucous carcinoma.
Two features that together relate almost ex- clusively to papillomas of the larynx are cases where the patient has a history of previous radio- therapy and the tumor has invasive properties. However, radiation therapy is not considered to be as responsible for invasive tracheal or bron- chial p a p i l l o m a t ~ s i s . ~ ~ ~ ~ ~
Malignancy and Papillomas. At present, the malig- nant potential of an unperturbed nonkeratinizing laryngeal papilloma is not definable. This unfor- tunate situation is related not only to the usual problems in defining a histologic malignancy within squamous mucosal lesions, but also to the confusion abundant in the literature. An example of the latter situation is the often-cited paper by
338 The Pathology of Head and Neck Tumors HEAD & NECK SURGERY MariApr 1983
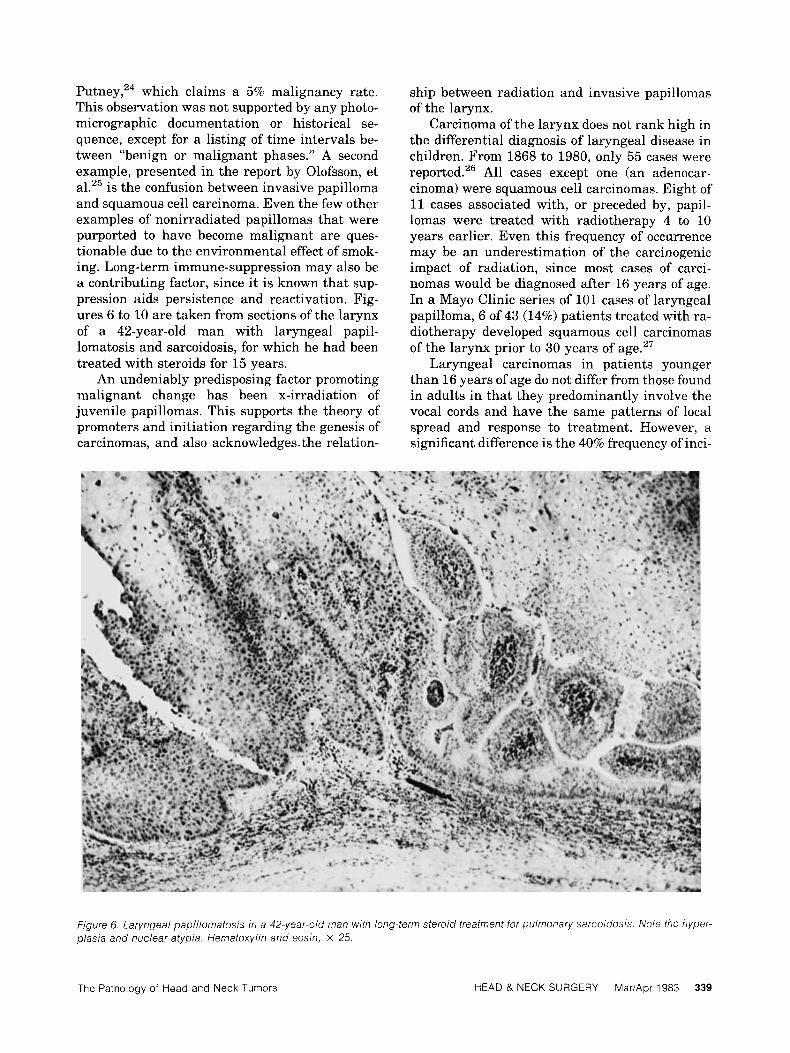
P ~ t n e y , ~ ~ which claims a 5% malignancy rate. This observation was not supported by any photo- micrographic documentation or historical se- quence, except for a listing of time intervals be- tween “benign or malignant phases.” A second example, presented in the report by Olofsson, et al.25 is the confusion between invasive papilloma and squamous cell carcinoma. Even the few other examples of nonirradiated papillomas that were purported to have become malignant are ques- tionable due to the environmental effect of smok- ing. Long-term immune-suppression may also be a contributing factor, since it is known that sup- pression aids persistence and reactivation. Fig- ures 6 to 10 are taken from sections of the larynx of a 42-year-old man with laryngeal papil- lomatosis and sarcoidosis, for which he had been treated with steroids for 15 years.
An undeniably predisposing factor promoting malignant change has been x-irradiation of juvenile papillomas. This supports the theory of promoters and initiation regarding the genesis of carcinomas, and also acknowledges. the relation-
ship between radiation and invasive papillomas of the larynx.
Carcinoma of the larynx does not rank high in the differential diagnosis of laryngeal disease in children. From 1868 to 1980, only 55 cases were reported.26 All cases except one (an adenocar- cinoma) were squamous cell carcinomas. Eight of 11 cases associated with, or preceded by, papil- lomas were treated with radiotherapy 4 t o 10 years earlier. Even this frequency of occurrence may be an underestimation of the carcinogenic impact of radiation, since most cases of carci- nomas would be diagnosed after 16 years of age. In a Mayo Clinic series of 101 cases of laryngeal papilloma, 6 of 43 (14%) patients treated with ra- diotherapy developed squamous cell carcinomas of the larynx prior to 30 years of age.27
Laryngeal carcinomas in patients younger than 16 years of age do not differ from those found in adults in that they predominantly involve the vocal cords and have the same patterns of local spread and response to treatment. However, a significant difference is the 40% frequency of inci-
Figure 6 Laryngeal papillomatosis in a 42-year-old man with long-term steroid treatment for pulmonary sarcoidosis Note the hyper- plas/a and nuclear atypia Hematoxylin and eosin, x 25
The Pathology of Head and Neck Tumors HEAD & NECK SURGERY MariApr 1983 339
340 The Pathology of Head and Neck Tumors
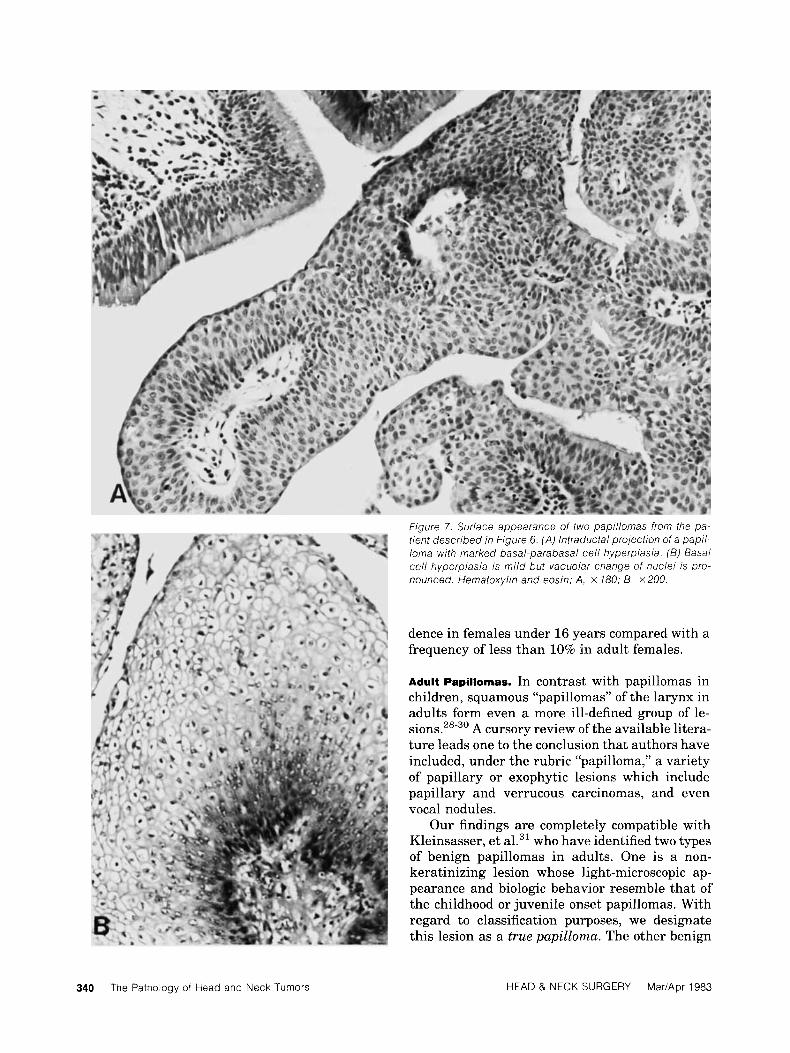
Figure 7. Surface appearance of two papillomas from the pa- tient described in f igure 6. (A) lntraductal projection of a pap/ / - loma with marked basal-parabasal cell hyperplasia. (B) Basal cell hyperplasia is mild but vacuolar change of nuclei is pro- nounced. Hematoxylin and eosin; A, x 180; B. x 200.
dence in females under 16 years compared with a frequency of less than 10% in adult females.
Adult Papillomas. In contrast with papillomas in children, squamous “papillomas” of the larynx in adults form even a more ill-defined group of le- s i o n ~ . ~ ~ - ~ ’ A cursory review of the available litera- ture leads one to the conclusion that authors have included, under the rubric “papilloma,” a variety of papillary or exophytic lesions which include papillary and verrucous carcinomas, and even vocal nodules.
Our findings are completely compatible with Kleinsasser, et al.31 who have identified two types of benign papillomas in adults. One is a non- keratinizing lesion whose light-microscopic ap- pearance and biologic behavior resemble that of the childhood or juvenile onset papillomas. With regard to classification purposes, we designate this lesion as a true papilloma. The other benign
HEAD & NECK SURGERY MariApr 1983
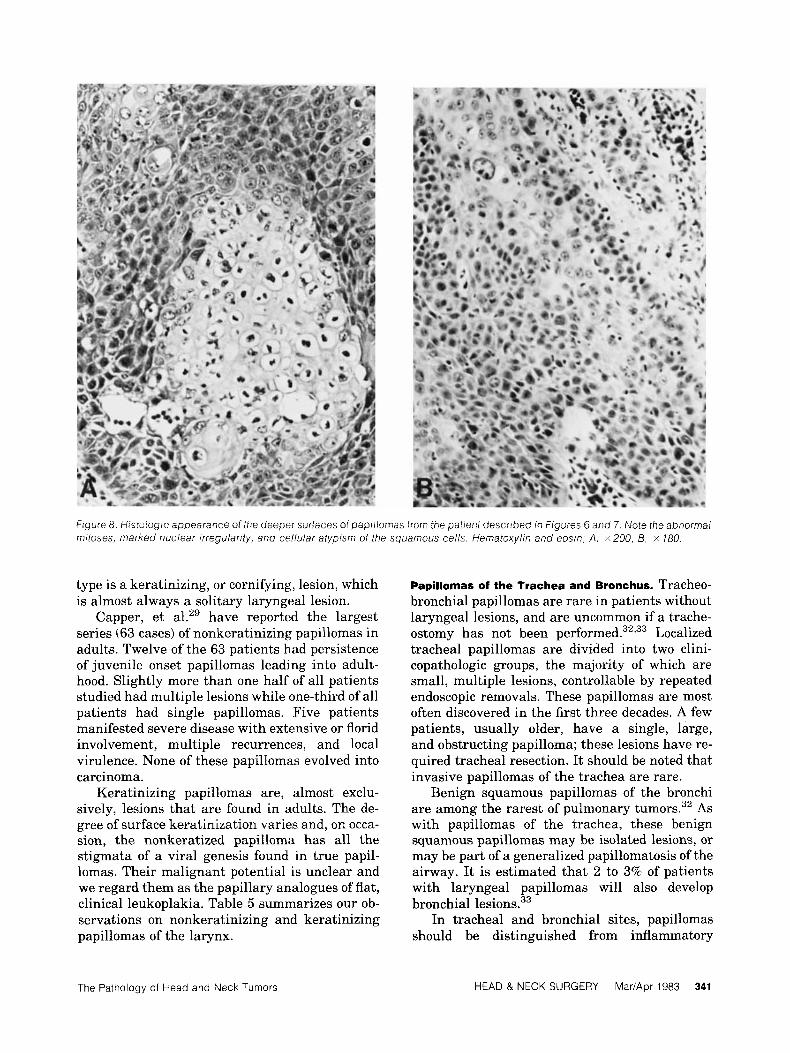
f igure 8. Histoiogic appearance of the deeper surfaces of papiifomas from the patient described in figures 6 and 7. Note fhe abnormal mitoses, marked nuclear irregularity, and ce/lular atypism of the squamous cells. Hematoxylin and eosin; A , x200, B, x 780.
type is a keratinizing, or cornifying, lesion, which is almost always a solitary laryngeal lesion.
Capper, et al.29 have reported the largest series (63 cases) of nonkeratinizing papillomas in adults. Twelve of the 63 patients had persistence of juvenile onset papillomas leading into adult- hood. Slightly more than one half of all patients studied had multiple lesions while one-third of all patients had single papillomas. Five patients manifested severe disease with extensive or florid involvement, multiple recurrences, and local virulence. None of these papillomas evolved into carcinoma.
Keratinizing papillomas are, almost exclu- sively, lesions that are found in adults. The de- gree of surface keratinization varies and, on occa- sion, the nonkeratized papilloma has all the stigmata of a viral genesis found in true papil- lomas. Their malignant potential is unclear and we regard them as the papillary analogues of flat, clinical leukoplakia. Table 5 summarizes our ob- servations on nonkeratinizing and keratinizing papillomas of the larynx.
Papillomas of the Trachea and Bronchus. Tracheo- bronchial papillomas are rare in patients without laryngeal lesions, and are uncommon if a trache- ostomy has not been p e r f ~ r m e d . ~ ~ , ~ ~ Localized tracheal papillomas are divided into two clini- copathologic groups, the majority of which are small, multiple lesions, controllable by repeated endoscopic removals. These papillomas are most often discovered in the first three decades. A few patients, usually older, have a single, large, and obstructing papilloma; these lesions have re- quired tracheal resection. It should be noted that invasive papillomas of the trachea are rare.
Benign squamous papillomas of the bronchi are among the rarest of pulmonary tumors.32 As with papillomas of the trachea, these benign squamous papillomas may be isolated lesions, or may be part of a generalized papillomatosis of the airway. It is estimated that 2 to 3% of patients with laryngeal papillomas will also develop bronchial lesions.33
In tracheal and bronchial sites, papillomas should be distinguished from inflammatory
The Pathology of Head and Neck Tumors HEAD & NECK SURGERY MariApr 1983 341
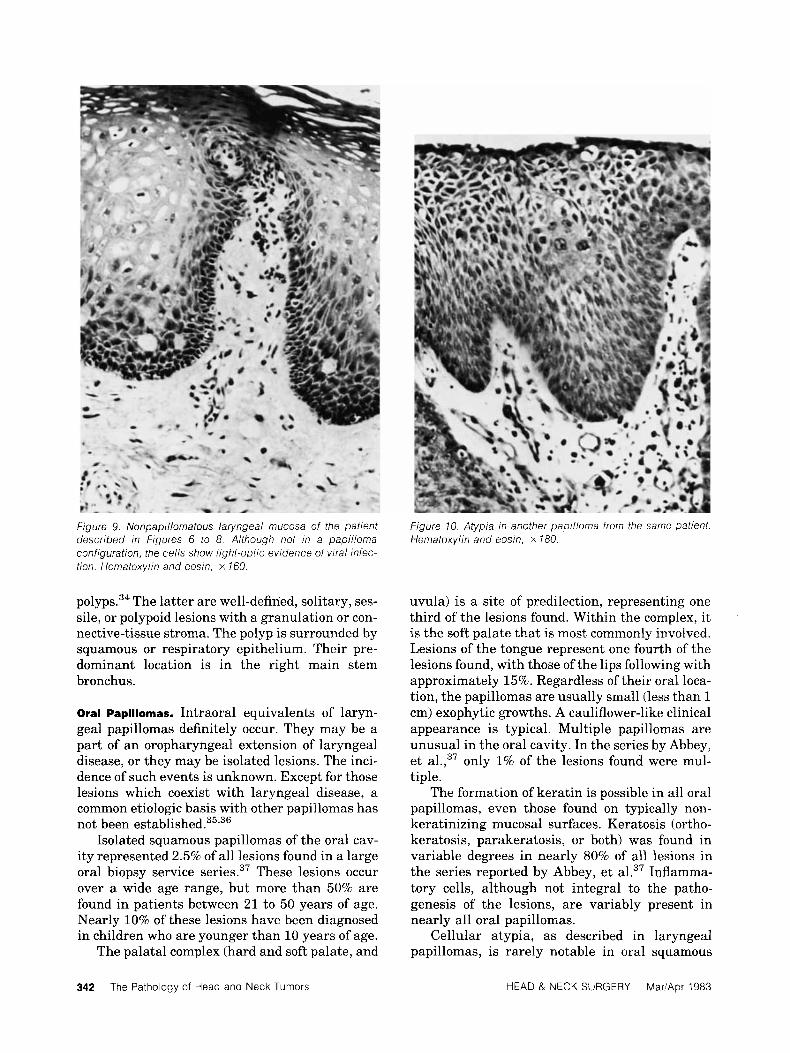
f igure 9. Nonpapillornatous laryngeal mucosa of the patient described in Figures 6 to 8. Although not in a papilloma configuration, the cells show light-optic evidence of viral infec- tion. Hematoxylin and eosin, x 160.
polyps.34 The latter are well-defined, solitary, ses- sile, or polypoid lesions with a granulation or con- nective-tissue stroma. The polyp is surrounded by squamous or respiratory epithelium. Their pre- dominant location is in the right main stem bronchus.
Oral Papillomas. Intraoral equivalents of laryn- geal papillomas definitely occur. They may be a part of an oropharyngeal extension of laryngeal disease, or they may be isolated lesions. The inci- dence of such events is unknown. Except for those lesions which coexist with laryngeal disease, a common etiologic basis with other papillomas has not been e ~ t a b l i s h e d . ~ ~ , ~ ~
Isolated squamous papillomas of the oral cav- ity represented 2.5% of all lesions found in a large oral biopsy service series.37 These lesions occur over a wide age range, but more than 50% are found in patients between 21 to 50 years of age. Nearly 10% of these lesions have been diagnosed in children who are younger than 10 years of age.
The palatal complex (hard and soft palate, and
Figure 10. Atypia In another papilloma from the same patient. Hematoxylin and eosin, x 180.
uvula) is a site of predilection, representing one third of the lesions found. Within the complex, it is the soft palate that is most commonly involved. Lesions of the tongue represent one fourth of the lesions found, with those of the lips following with approximately 15%. Regardless of their oral loca- tion, the papillomas are usually small (less than l cm) exophytic growths. A cauliflower-like clinical appearance is typical. Multiple papillomas are unusual in the oral cavity. In the series by Abbey, et al.,37 only 1% of the lesions found were mul- tiple.
The formation of keratin is possible in all oral papillomas, even those found on typically non- keratinizing mucosal surfaces. Keratosis (ortho- keratosis, parakeratosis, or both) was found in variable degrees in nearly 80% of all lesions in the series reported by Abbey, et al.37 Inflamma- tory cells, although not integral to the patho- genesis of the lesions, are variably present in nearly all oral papillomas.
Cellular atypia, as described in laryngeal papillomas, is rarely notable in oral squamous
342 The Pathology of Head and Neck Tumors HEAD & NECK SURGERY MariApr 1983

Table 5. Papillomas of larynx
TVRe Characteristics
Nonkeratinizing 1. Papillomavirus strongly implicated in etiology
2. Clinical onset most often in childhood, but can present in adult without evi- dence of persistence of disease from childhood
3. Most often multiple: may be solitary
4. Biologic course variable:
a. Controlled by excision b. "Spontaneous" regression c. Persistent, recurrent, and distal
spread d. lnvasive outside of mucosa (rare)
5. "Malignant transformation" mostly lim- ited to those with prior irradiation or other carcinogens; spontaneous malig- nancy is rare
1 , Weak or no association with viruses
2. Almost exclusively an adult lesion
3. Almost always solitary
4. Usually controllable by surgery if stimulating agents removed
5. Biologic implications probably the same as rnucosal keratoses and depen- dent on degree of subkeratotic dvsDlasia
Keratinizing
papillomas; when present, it is of debatable im- portance except as a histologic expression of a rapid turnover of cells. Recurrences of the lesions after surgical removal or their appearance in new sites occur much less often than with laryngeal papillomas. There are also few documented exam- ples of malignant change.
The inverted ductal papilloma of the oral cav- ity, a clinically submucosal lesion with an origin in the excretory duct reserve cells of minor sali- vary glands, should not be confused with the squamous papillomas described previously. In its growth pattern and cytomorphologic appearance, the inverted ductal papilloma closely resembles Schneiderian papillomas of the sinonasal tract. The high recurrence rate and association with carcinoma of the sinonasal tract have not been a feature of these oral lesions.38
Genital Condylomas and Papillomas. Several stud- ies have suggested that there may be a relation- ship between maternal condylomas and laryngeal papilloma^.'^,^^ However, the prospective testing of such a relationship by Cohn, et al.40 indicates that a correlation between laryngeal papillomas
in children and condyloma acuminata in the mother during pregnancy or parturition may not be as strong as retrospective studies have im- plied.
However, the association of genital papil- lomavirus with cervical, vaginal, and vulvar car- cinoma is further advanced than the relationship of laryngeal carcinoma with laryngeal papil- lomavirus infection. Reid, et aL4' have found definite histologic evidence of subclinical papil- lomavirus infection in 91% of a balanced sample of women treated for preinvasive, or invasive, cer- vical cancer. A matched control group manifested only a 12% prevalence of infection. There also were similarities in epidemiologic profiles of cer- vical papillomaviral infection and cervical neo- plasia. It is postulated that cervical neoplasia may arise from a mutagenesis within infected cells at the squamocolumnar junction.
SUMMARY
The papillomaviruses are one of two classes of DNA viruses that are strongly implicated in the production of benign, nonmetastasizing tumors in humans. The mucosal lesions associated with the epitheliotropic papillomaviruses are typified by an exuberant epithelial growth with little or no keratinization; these include, for example, laryngeal and oral papillomas, and condylomata acuminata. Human papillomavirus (HPV) cannot be grown or tested in tissue culture and, as with other members of its genera, it is highly species- specific. In addition, there is a significant plural- ity for HPV. The different types of HPV have lit- tle or no polynucleotide sequence homology, and there is no cross reaction between HPV and virion surface antigens. The type of papillomavirus ap- pears to determine, in part, the clinical appear- ance, location, and natural fate or progression of the lesion it is putatively associated with.
We postulate that an activation of a persistent viral infection, coupled with an unknown promo- ter, is the etiologic basis for laryngeal papillomas, and perhaps also for oral papillomas. The non- keratinizing, or typical, papillomas have the same genesis whether they occur in children or in adults. Keratinizing papillomas in adults may not be pathogenetically related.
The malignant potential of a nonkeratinizing papilloma is unknown. However, on the basis of present knowledge, it is low unless there is a significant environmental enhancement such as x-irradiation exposure or prolonged immunosup- pression.
The Pathology of Head and Neck Tumors HEAD & NECK SURGERY MariApr 1983 343

~
REFERENCES
1. 2.
3.
4.
5 .
6.
7.
8.
9.
Wyke J A Oncogenic viruses. J Pathol 135:39-85, 1981. Takemoto KK: Human papovaviruses. Int Rev Exp Pathol 18:281-301, 1978. Zur Hausen H: Human papillomaviruses and their possi- ble role in squamous cell carcinomas. Curr Top Microbiol Immunol 78:l-30, 1977. Matthews REF: Classification and nomenclature of vi- ruses. Intervirology 12:131-296, 1979. Meeting Report: Workshop on papillomaviruses and can- cer. Cancer Res 39545-546, 1979. Lancaster WD, Jenson AB: Evidence for papillomavirus genus-specific antigens and DNA in laryngeal papilloma. Intervirology 15:204-212, 1981. Lack EE, Jenson AB, Smith HG, Healy GB, Pass F, Vaw- ter GF: Immunoperoxidase localization of human papil- lomavirus in laryngeal papillomas. Intervirology 14:148- 154, 1980. Mims C A Role of persistence in viral pathogenesis, in Mahy BWJ, Mins AC, Darby GK (eds): Virus Persistence, 33rd Symposium of the Society for General Microbiology. March 1982. Cambridge, Cambridge University Press, 1982, pp 1-13. Robb JA: Virus-cell interactions: A classification for vi- rus-caused human disease. Prog Med Virol 2351 -61, 1977.
10. Oldstone MBA, Fujinami RS, Lampert PW: Membrane and cytoplasmic changes in virus-infected cells induced by interactions of antiviral antibody with surface viral anti- gens. Prog Med Virol 26:45-93, 1980.
11. Rowe WP: Genetic interactions between tumor viruses and host cells. Cancer 49:1958-1962, 1982.
12. Watson JD: Induction of cancer by DNA viruses. Ann Otol Laryngol Rhinol 89:489-496, 1980.
13. Chan GL: On the nature of oncogenic transformation of cells. In t Rev Cytol 7O:lOl-137, 1981.
14. Orth G, Jablonska S, Breitburd F, Favre M, Croissant 0: The human papillomaviruses. Bull Cancer 65:151-164, 1978.
15. Quick CA, Faras A, Krzysek R: The etiology of laryngeal papillomatosis. Laryngoscope 88:1789-1795, 1978.
16. Spoendlin H, Kistler G Papovavirus in human laryngeal papillomas. Arch Otol Rhinol Laryngol 218:289-292, 1978.
17. Jenson AB, Rosenthal JD, Olson C, Pass F, Lancaster WD, Shah K: Immunologic relatedness of papillomavi- ruses from different species. J Nut1 Cancer Inst 64:495- 500, 1980.
18. Costa J , Howley PM, Bowling MC, Howard R, Bauer WC: Presence of human papilloma viral antigens in juvenile multiple laryngeal papilloma. Am J Clin Pathol 75194- 197, 1981.
19. Kjeldsberg E: Application of electron microscopy in viral diagnosis. Pathol Res Pract 167:3-21, 1980.
20. Ferenczy A, Braun L, Shah KV: Human papillomavirus (HPV) in condylomatous lesions of cervix. Am J Surg Pathol 5:661-670, 1981.
21. Quick CA, Foucar E, Dehner LP: Frequency and significance of epithelial atypia in laryngeal papil-
lomatosis. Laryngoscope 89550-560, 1979. 22. Fechner RE, Goepfert H, Alford BR: Invasive laryngeal
papillomatosis. Arch Otolaryngol 99:147-151, 1974. 23. Fechner RE, Fitz-Hugh GS: Invasive tracheal papil-
lomatosis. Am J Surg Pathol 4:79-86, 1980. 24. Putney FJ: Borderline malignant lesions of the larynx.
Arch Otolaryngol 61:381-385, 1955. 25. Olofsson J , Bjelkenkrantz K, Grontoft 0: Malignant de-
generation of a juvenile laryngeal papilloma-a follow-up study. J Otolaryngol 9:329-333, 1980.
26. Gindhart TD, Johnston WH, Chism SE, Dedo HH: Carci- noma of the larynx in childhood. Cancer 46:1683-1687. 1980.
27. Majoros M, Devine KD, Parkhill EM: Malignant transfor- mation of benign laryngeal papillomas in children after radiation therapy. Surg Clin N Am 43:1049-1061, 1963.
28. Bjork H, Teir H: Benign and malignant papilloma of the larynx in adults. A comparative clinical and histological study. Acta Otolaryngol 47:95-104, 1957.
29. Capper JWR, Bailey CM, Michaels L: Squamous papil- lomas of the larynx in adults: A review of 63 cases. Clin Otolaryngol 7:138-139, 1982.
30. Johnson JT, Barnes EL, Justice W: Adult onset laryngeal papillomatosis. Otolaryngol Head Neck Surg 89:867-873, 1981.
31. Kleinsasser D, Oliviera E, Cruz G: Juvenile und adulte kehlkopfpapillome. HNO 21:97-106, 1973.
32. Spencer H, Dail DH, Arneud J: Non-invasive bronchial papillary tumors. Cancer 45: 1486-1497, 1980.
33. Glazer G, Webb WR: Laryngeal papillomatosis with pul- monary spread in a 69-year-old man. Am J Roentgen01 1322320-822. 1979.
34. White RP, Cordasco EM, Ahmad M, Sebek B: Multiple endobronchial polyposis; differentiation from squamous cell papillomas. Clew Clin Quart 46:71-76, 1979.
35. Jenson AB, Lancaster WD, Hartmann RP, Shaffer EL: Frequency and distribution of papillomavirus structural antigens in verrucae, multiple papillomas, and condy- lomata of the oral cavity. Am J Pathol107:212-218,1982.
36. Lutzner M, Kuffer R, Blanchet-Bardon C, Croissant 0: Different papillomaviruses as the causes of oral warts. Arch Lfermatol 118:393-397, 1982.
37. Abbey LM, Page DG, Sawyer D R The clinical and his- topathologic features of a series of 464 oral squamous cell papillomas. Oral Surg 49:419-428, 1980.
38. White DK, Miller AS, McDaniel RK, Rothman BN: In- verted ductal papilloma: A distinctive lesion of minor sali- vary gland. Cancer 49519-524, 1982.
39. Quick CA, Watts SL, Krzysek RA, Faras AJ: Relationship between condylomata and laryngeal papillomata. Clinical and molecular virological evidence. Ann Otol Rhinol Laryngol 89:467-471, 1980.
40. Cohn AM, Kos JT, Taber LH, Adams E: Recurring laryngeal papilloma. Am J Otolaryngol 2:129-132, 1981.
41. Reid R, Stanhope CR, Herschman B, Booth E, Phibbs G, Smith J: Genital warts and cervical cancer. I. Evidence of an association between subclinical papillomavirus infec- tion and cervical malignancy. Cancer 50:191-202, 1982.
344 The Pathology of Head and Neck Tumors HEAD & NECK SURGERY Mar/Apr 1983
Copyright © 2022 FDOKUMEN




















