
The lorisiform wrist joint and the evolution of “brachiating” adaptations in the hominoidea
24
The Lorisiform Wrist Joint and the Evolution of "B r ac h i at i ng " Ad aptat ion s i n the H o m i no idea MATT CARTMILL AYD KATHARINE MILTOK Departments ofAnatomj and Anthropology. Duke Uniuersity, Durham, North Carolina 27710und Smithsonran 'Dopica1Research Institute, P 0 Box 2072, Balboa, Canal Zone, U S A KEY WORDS Lorisiformes Hominoidea - Wrist . Brachiation ABSTRACT In lorisines (Loris, Nycticebus, Perodicticus, Arctocebus), the tip of the ulna is reduced to the dimensions of a styloid process, a new and more prox- imal ulnar head is developed, and the pisiform is displaced distally away from its primitive contact with the ulna. In some Nycticebus, intra-articular tissues sepa- rate the ulna from the triquetrum. These traits are not seen in other quadrupe- dal primates, but they are characteristic of extant hominoids. Among hominoids, these features have been interpreted as adaptations to arm-swinging locomotion. Since hominoid-like features of the wrist joint are found in lorisines, but not in New World monkeys that practice arm-swinging locomotion, these features may have been evolved in both lorisines and large hominoids to enhance wrist mobility for cautious arboreal locomotion involving little or no leaping. Most of the other morphological traits characteristic of modern hominoids can be ex- plained as adaptations to cautious quadrupedalism as well as to brachiation, and may have developed for different reasons in different lineages descended from an unspecialized cautious quadruped resembling Alouatta. Although St. George Mivart noted well over a hundred years ago that man, orangutans and chimpanzees differ from monkeys in lack- ing a direct articulation between the ulna and the carpal bones (Mivart, 18671, most of Mivart's more theoretically-oriented succes- sors overlooked this fact when trying to ex- plain the resemblances between men and apes. The specializations of the hominoid wrist joint have come into theoretical promi- nence only in the last ten years, thanks prin- cipally to the work of 0. J. Lewis ('65, '69, '70. '71a,b, '72a,b, '74). Lewis' painstaking ana- tomical studies of the anthropoid wrist have established that extant hominoids (including gorillas and gibbons as well as the species that Mivart looked at) share the following traits, not seen among other higher primates: 1. The hominoid pisiform no longer forms with the triquetrum a compound socket for the head of the ulna-as it does in monkeys and most other mammals, in which the proxi- mally-directed pisiform juts back across the wrist joint to provide a heel-like lever arm for the m. flexor carpi ulnaris. The relatively small pisiform of hominoids projects ventrally and distally (rather than proximally) into the palm, and does not articulate with the ulna. The hypothenar pad, overlying the pisiform, is also displaced distally relative to the distal end of the ulna in hominoids (Midlo, '34). 2. The distal extremity of the hominoid ulna is reduced, forming a more or less narrow styloid process bearing a reduced articular surface at its tip. 3. The styloid process of the ulna is partly (Hylobates, Pan) or entirely (Gorilla, Pongo) excluded from direct contact with the tri- quetrum by an intra-articular meniscus which extends inward from the ulnar side of the joint capsule and attaches dorsally to the ulnar edge of the radius. In most human beings, this meniscus has lost its independent character and is incorporated into the prox- imal articular surface of the radiocarpal joint; but discrete menisci or even chimpan- zee-like ulnotriquetral articulations occur in some individuals (Jones, '42; Lewis et al., '70). 4. The distal radioulnar joint, a simple syn- desmosis in some New World monkeys (Lewis, 249 AM. J. PHYS. ANTHROP., 47: 249-272.
Transcript of The lorisiform wrist joint and the evolution of “brachiating” adaptations in the hominoidea

The Lorisiform Wrist Joint and the Evolution of "B r ac h i at i ng " Ad apt at ion s i n the H o m i no idea
MATT CARTMILL AYD KATHARINE MILTOK Departments o fAnatomj and Anthropology. Duke Uniuersity, Durham, North Carolina 27710und Smithsonran 'Dopica1 Research Institute, P 0 Box 2072, Balboa, Canal Zone, U S A
KEY WORDS Lorisiformes Hominoidea - Wrist . Brachiation
ABSTRACT In lorisines (Loris, Nycticebus, Perodicticus, Arctocebus), the tip of the ulna is reduced to the dimensions of a styloid process, a new and more prox- imal ulnar head is developed, and the pisiform is displaced distally away from its primitive contact with the ulna. In some Nycticebus, intra-articular tissues sepa- rate the ulna from the triquetrum. These traits are not seen in other quadrupe- dal primates, but they are characteristic of extant hominoids. Among hominoids, these features have been interpreted as adaptations to arm-swinging locomotion. Since hominoid-like features of the wrist joint are found in lorisines, but not in New World monkeys that practice arm-swinging locomotion, these features may have been evolved in both lorisines and large hominoids to enhance wrist mobility for cautious arboreal locomotion involving little or no leaping. Most of the other morphological traits characteristic of modern hominoids can be ex- plained as adaptations to cautious quadrupedalism as well as to brachiation, and may have developed for different reasons in different lineages descended from an unspecialized cautious quadruped resembling Alouatta.
Although St. George Mivart noted well over a hundred years ago that man, orangutans and chimpanzees differ from monkeys in lack- ing a direct articulation between the ulna and the carpal bones (Mivart, 18671, most of Mivart's more theoretically-oriented succes- sors overlooked this fact when trying to ex- plain the resemblances between men and apes. The specializations of the hominoid wrist joint have come into theoretical promi- nence only in the last ten years, thanks prin- cipally to the work of 0. J. Lewis ('65, '69, '70. '71a,b, '72a,b, '74). Lewis' painstaking ana- tomical studies of the anthropoid wrist have established that extant hominoids (including gorillas and gibbons as well as the species that Mivart looked a t ) share the following traits, not seen among other higher primates:
1. The hominoid pisiform no longer forms with the triquetrum a compound socket for the head of the ulna-as i t does in monkeys and most other mammals, in which the proxi- mally-directed pisiform juts back across the wrist joint to provide a heel-like lever arm for the m. flexor carpi ulnaris. The relatively
small pisiform of hominoids projects ventrally and distally (rather than proximally) into the palm, and does not articulate with the ulna. The hypothenar pad, overlying the pisiform, is also displaced distally relative to the distal end of the ulna in hominoids (Midlo, '34).
2. The distal extremity of the hominoid ulna is reduced, forming a more or less narrow styloid process bearing a reduced articular surface a t its tip.
3. The styloid process of the ulna is partly (Hylobates, Pan) or entirely (Gorilla, Pongo) excluded from direct contact with the tri- quetrum by an intra-articular meniscus which extends inward from the ulnar side of the joint capsule and attaches dorsally to the ulnar edge of the radius. In most human beings, this meniscus has lost its independent character and is incorporated into the prox- imal articular surface of the radiocarpal joint; but discrete menisci or even chimpan- zee-like ulnotriquetral articulations occur in some individuals (Jones, '42; Lewis et al., '70).
4. The distal radioulnar joint, a simple syn- desmosis in some New World monkeys (Lewis,
249 AM. J. PHYS. ANTHROP., 47: 249-272.

250 MATT CARTMILT, AND KATHARINE MILTON
’651, becomes in hominoids “a fully elaborated diarthrodial . . . joint which incorporates a neomorphical ulnar head” (Lewis, ’74) ; the new ulnar head is produced by the expansion of the ulna’s primitively diminutive articular surface for the radius.
This “drastic reorganization of the wrist joint” (Lewis, ’74) is, in Lewis’ view, “corre- lated with the emergence of a new locomotor pa t te rn . . . characterized by an accomplished ability for rotation of the body from the grasp- ing hand” (Lewis, ’72a). Pronation and supi- nation are obviously facilitated by the expan- sion of the distal radioulnar diarthrosis, and the retreat of the ulna from its carpal articu- lation is also seen by Lewis (’69) as an adapta- tion facilitating the hand’s rotation with the radius around the stationary ulna. Lewis regards the resulting increase in the rotatory mobility of the hominoid hand and radius as an adaptation to brachiation. Lewis defines this term broadly (Lewis, ’72a), but places particular emphasis on slow bimanual suspen- sory locomotion, which involves 180” of rota- tion in the entire forelimb as an animal swings along underneath a branch from one handhold to the next (Avis, ’62; Fleagle, ’74; Carpenter, ’76; Baldwin and Teleki, ’76). Lewis’ analysis thus supports and supple- ments Keith’s (‘23) so-called “brachiationist” model of hominoid differentiation; the distinc- tive features of the wrists of hominoids, like their broad thoraxes, reduced lumbar and cau- dal vertebral regions, mobile shoulders, and reduced olecranon processes, are interpreted as adaptations to swinging by the arms from overhead supports. Since parallel specializa- tions of the wrist are not developed in the ateline monkeys of the New World, which exhibit ape-like suspensory locomotor behav- ior (Mittermeier and Fleagle, ’761, Lewis regards the hominoid wrist specializations as synapomorphies which demonstrate the monophyletic character of the Hominoidea (Lewis, ’69, ’71a). Persistently monkey-like features of the gibbon wrist (substantial ulnotriquetral articulation, distinct and re- stricted meniscus, and shape of the pisiform and triquetrum) suggest t o Lewis t h a t hylobatids diverged from the other hominoids before the hominoid adaptations for brachia- tion had been perfected (Lewis, ’71b). Lewis finds more advanced brachiating adaptations in the Miocene ape Dryopithecus africanus, which he regards as a plausible common ancestor of man and the African apes (Lewis,
’71b, ’72a,b, ’74). Brachiating modifications of the wrist (particularly of the midcarpal joint) reach their greatest degree of perfection, in Lewis’ view, in chimpanzees and gorillas. The “apparent paradox.. . that the least arboreal of extant apes are the possessors of midcarpal joints which seem best suited to forelimb suspension” (Lewis, ’74) is dismissed with the observation that the ricochetal habits of gib- bons and the quadrupedal suspensory pos- tures of orangutans free them from relying exclusively on forelimb rotation in brachia- tion, which was perfected only in the common ancestor of man and the African apes (Lewis, ’73).
Several sorts of objections have been raised to Lewis’ analysis. Many of us still find it odd that “the only primates which consistently brachiate are ill adapted for such activity” (Conroy and Fleagle, ’72). Probably a majority of authors in this area continue to follow Tut- tle (’67, ’69, ’70) in regarding the muscular and bony features which uniquely limit dor- siflexion a t the wrist in chimpanzees and gorillas as adaptations to the knuckle-walk- ing locomotion characteristic of these African apes (Tuttle and Basmajian, ’74a; Tuttle, ’75a,b; OConnor, ’75; Jenkins and Fleagle, ’751, rather than as brachiating adaptions (Lewis, ’74). Inverting Lewis’ argument, Con- roy and Fleagle (’72) suggest that if Dryo- pithecus africanus really resembles chim- panzees and gorillas in features of wrist anat- omy not seen in orangutans and gibbons, that fact should be taken as evidence that D. africanus had a chimpanzee-like locomotor repertoire involving a considerable amount of knuckle-walking. Conroy and Fleagle propose that the retreat of the ulna from the carpals in hominoids may have originated a s a knuckle-walking adaptation, to allow ulnar deviation of the hand when the arm is brought forward during the swing phase of its cycle.
However, knuckle-walking has since been shown to involve very little ulnar deviation a t the wrist (Jenkins and Fleagle, ’75). I t thus seems unlikely that the retreat of the homi- noid ulna had anything to do with knuckle- walking. Furthermore, recent studies have reaffirmed Napier and Davis’ (’59) conclusion that Dryopithecus africanus retained a primi- tive, monkey-like contact between the ulna and pisiform (Schon and Ziemer, ’73; Mor- beck, ’75; Corruccini et al., ’75). In fact, the wrist joint is monkey-like in all the Tertiary hominoids for which evidence exists, and the

LORIS WRIST AND HOMlNOlD EVOLUTION 251
Oligocene and Miocene hominoids known from postcranial remains display other mon- key-like features which suggest that they were all relatively unspecialized quadrupeds (Le Gros Clark and Thomas, ’51; Pilbeam, ’69; Ankel, ’65; Morbeck, ’72; Lewis, ’74; Conroy, ’74; Fleagle et al., ’75; Corruccini et al., ’75, ’76). If the distinctive wrist specializations of modern hominoids represent a current or ancestral adaptation to brachiation where- ever they occur, then one of the following must be true of the extant hominoids’ last common ancestor:
(1) It was a brachiating animal with wrist of modern hominoid type, descended from one of the quadrupedal pongids of the Miocene.
(2) I t was a brachiating animal with wrist of modern hominoid type, but it was a precur- sor or contemporary of the quadrupedal Mio- cene pongids. In other words, the dryopithe- cines are the sister group of all the extant hominoids, gibbons included.
(3) It was a monkey-like quadruped-a “dental ape”-and brachiating adaptations evolved independently in several lineages leading to the hominoids of today.
None of these alternatives is entirely satis- factory. The first one implies a rather short time scale for the divergence of the extant hominoids, perhaps as short as that proposed by Sarich and Wilson (’67). The second alter- native implies that a major evolutionary ra- diation among African and Asian primates is mysteriously unrepresented in the Miocene fossil record. The third alternative is cast into doubt by the failure of the atelines to develop hominoid-like specializations of the wrist joint! in connection with their other adapta- tions for suspensory locomotion.
Another possibility, suggested by Mivart’s (1867) original study, is that the hominoid peculiarities of the wrist may originally have had nothing to do with brachiation, a t least in some lineages. Mivart observed that the tri- quetral (“cuneiforme”) of the Lorisinae artic- ulates only “very slightly” with the ulna, and that the pisiform has no ulnar articulation in these animals. He remarked further that the lorisine pisiform “is very small, . . . and bend- ing distad, is applied to the outer surface of the unciforme, and has its extremity united to the palmar process of that bone” (Mivart, 1867). Among hominoids, such a contact be- tween pisiform and “unciforme” (hamate) is found only in orangutans, and is regarded by Lewis (’74) as an extreme expression of the
hominoid trend toward distal displacement of the pisiform. These putative hominoid-like features of the wrist joint in lorisines are surely unrelated to any history of brachiation in the lorisine ancestry. Convergence in wrist morphology between lorisines and hominoids would accordingly suggest that the specializa- tions shared by these animals, but not seen among atelines, could have originated as an adaptation to some locomotor mode other than bimanual suspension. We accordingly undertook to test and refine Mivart’s observa- tions.
MATERIALS AND METHODS
One preserved specimen each of Galago demidouii, Galago senegalensis, Galago elegan- tulus, Arctocebus calabarensis, and Perodic- ticus potto, two specimens each of Galago crassicaudatus and Loris tardigradus, and four specimens of Nycticebus coucang were ob- tained for study. Specimens of each species were x-rayed in a dorsopalmar direction with the dorsum of the wrist pressed against the plate holder. All the specimens of the three species represented by multiple specimens, together with the single specimens of Perodic- ticus and Galago senegalensis, were dissected by skinning the wrist and cutting through the joint capsule and extensor tendons dorsally to expose the proximal wrist joint. These studies of preserved material were supplemented by examination of skeletal preparations in the collections of the United States National Museum and the Department of Anatomy of Duke University.
RESULTS
Radiographic and osteological findings
The radiographs revealed three principal points of difference between the wrists of Galaginae and Lorisinae (fig. 1):
1. Pisiform. The galagine pisiform has the typical mammalian configuration and posi- tion; it contacts the distal end of the ulna broadly, and is prolonged proximally into the palm, forming a palmar counterpart of the heel of the foot. I t often bears an apparent traction epiphysis (fig. 2). By contrast, the pisiform of all the lorisine specimens exam- ined was relatively small and displaced distal- ly, separated from the ulna by radiolucent tis-
1 Some investigators have found no contact between pisiform and ulna in Ateles (Parsons, 1899; Schwartz, ‘38). while others have (Lewis. ‘71a), the fealure may be variable in this genus. The ulnotri- quetral contact, however, is constant.

252 MATT CARTMILL AND KATHARINE MILTON
Fig. 1 Radiogaphs of hand of Galago crasszcaudutus (left) and Arctocebus cdabarensis (right), illustrating differences between galagines and lorisines in construction of the antehrachiocarpal and distal radioulnar joints. Not to same scale.
Fig. 2 Left ulnocarpal joint of CQZQ~O senegalensis (U. S. Natl. MUB. No. 251625). viewed from the ulnar aspect. Scale on this and succeeding drawings indicates 1 mm. Abbreviations: e, epiphysis of pisiform; P, pisiform; R. radius: T, triquetrum; U. ulna.
sues. The distal displacement of the pisiform in lorisines is accompanied, as in hominoids, by a distal displacement of the hypothenar pad relative to the tip of the ulna (Midlo, '34). The pisiform-hamate contact reported by Mivart (1867) in lorisines seems to be unique to Nycticebus (Etter, '74).
2. Ulnotriquetral joint. In the galagines, the distal end of the ulna articulates with the tri- quetrum, which together with the pisiform
forms a cup-like socket for the reception of the ulna (fig. 2). In the four lorisines exam- ined, the triquetrum and ulna were separated by a substantial radiolucent gap. Extreme forced ulnar deviation of the wrist, attempted on one specimen of Nycticebus, failed to elimi- nate this gap (fig. 3). In the galagines, the dis- tal end of the ulna (homologous with the hominoid styloid process) is comparatively short and stout, approximating the diameter

LORIS WRIST AND HOMINOIU EVOLUTION 253
Fig. 3 Radiograph of right wrist of Nycticebus coucang, preserved specimen. The radiolucent space hetween ulna and carpals persists under forced ulnar deviation.
of the midpoint of the ulnar shaft; in the lorisines, it is a relatively slender projection ending in a smaller articular surface, much like the styloid process of hominoids (figs. 4, 5 ) .
3. Distal radioulnar joint. This joint (fig. 4) is formed in Galago crassicaudatus by a short articular process of the ulna, which projects at an angle from the ulnar shaft and fits against a matching surface on the ulnar edge of the radius, just proximal to the radial artic- ular surface for the lunate. In the smaller galagos, the ulna’s articular surface is not supported on a distinct process (fig. 5). The ra- dioulnar articulation is relatively small and flat in all four galagines, and is oriented roughly perpendicular to an axis passing through the radius and ulna a t right angles to their shafts. In lorisines, a shelflike process of the distal end of the radius extends across toward the ulnar styloid, interposing itself be- tween the ulnar shaft and the carpals (Botez, ’26). This expansion of the radius results in the expansion of two articular surfaces: the surface which contacts the carpals on the dis- tal side of the shelf, and the surface which
contacts the ulna on the proximal side of the shelf. The latter surface of the radius receives a correspondingly expanded distal articular surface of the ulna, constituting a neomorphic secondary ulnar head like that of hominoids. This neomorphic head terminates the shaft of the ulna, and is reoriented to face as much dis- tally as radially (fig. 4). The shelf is thus a functional equivalent of the triangular artic- ular disk in hominoids. The joint surface be- tween radius and ulna is not planar, but ap- proaches a ball-and-socket configuration, with the ulnar “ball” fitting into a radial socket
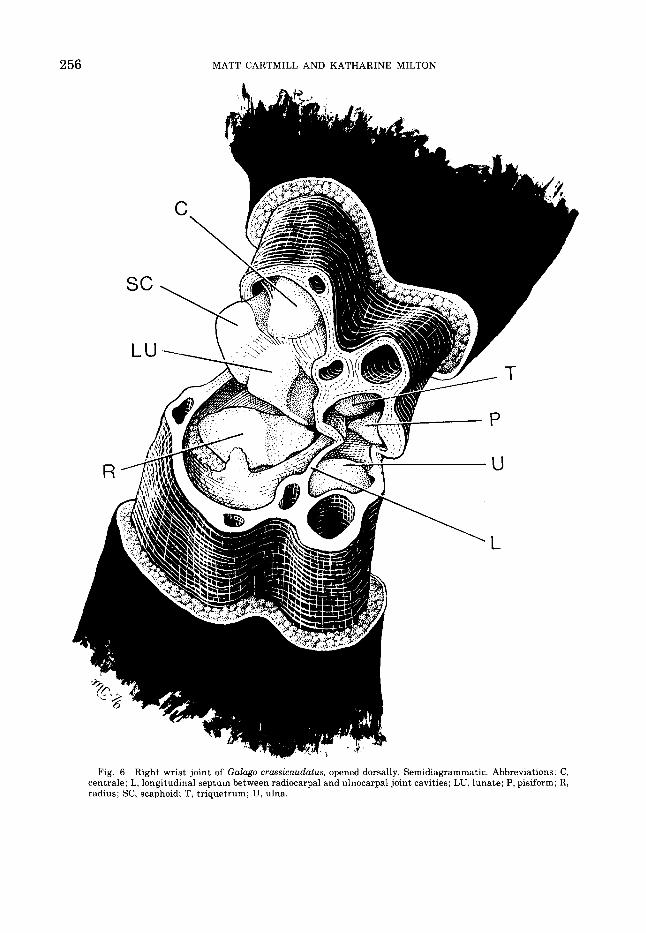
Dissection results 1. Galago crassicaudatus (fig. 6). The prox-
imal wrist joint of the greater bushbaby is divided into separate radial and ulnar com- partments by a longitudinal synovial septum. This septum is continuous dorsally with the capsule of the joint; ventrally, with the palmar ulnocarpal ligament; distally, with the ligaments connecting triquetrum and lunate; and proximally, with the ligamentous bond between distal radius and ulna. A sep-
(fig. 1).

254 MATT CARTMILL AND KATHARINE MILTON
H

LORIS WRIST AND HOMINOID EVOLUTION 255
256 MATT CARTMILL AND KATHARINE MILTON
Fig. 6 Right wrist joint of Galago crassicaudatus, opened dorsally. Semidiagrammatic. Abbreviations: C, centrale; L, longitudinal septum between radiocarpal and ulnocarpal joint cavities; LU, lunate; P, pisiform; R, radius; SC, scaphoid; T, triquetrum; U, ulna.

LORIS WRIST AND HOMINOID EVOLUTION 257
tum in this position was found in all the lorisiforms dissected. Ontogenetically, this septum seems to result from the partial per- sistence of the mesenchymal partition be- tween the primordial radiocarpal and ulnocar- pal joint cavities (Lewis, '70). It may be total- ly resorbed during development, as in Dasyurus, Bradypus, Dasypus, Homo, the pongids, and many monkeys; or be partially resorbed, leaving a perforated septum, as in Daubentonia, Ailuropoda, Lagothrix, Cer- copithecus, a t least some Papio, some Perodic- ticus, and one specimen of Loris examined by us; or persist into adult life as a complete par- tition, as in monotremes, Didelphis, Pha- langer, Tric hosu rus, Macropus, T u p a ia, Microcebus, Lemur fuluus, Tarsius, Callimico, Brachyteles, Ateles, and some Hylobates indi- viduals (Parsons, 1899; Nayak, '33; Hughes, '44; Robertson, '44; Hill, '53, '59, '60, '62, '66; Davis, '64; Altner, '71; Lewis, '71a). We would expect variation in this feature in most spe- cies of mammals, given its ontogenic basis.
The radiocarpal synovial cavity in G. crassicaudatus not only encompasses the ar- ticular surfaces of the radius, lunate, and scaphoid, but extends dorsally between the joint capsule and the dorsal surface of the 0s centrale. The capsule is greatly thickened here to form a fibrous or fibrocartilaginous labrum, bound tightly to a dorsal process pro- jecting from the radius (fig. 4). On dor- siflexion, the centrale comes into close articu- lation with this labrum. On its ulnar edge, the labrum is continuous with the longitudinal septum of the joint, with which it is connected by a fine web of delicate areolar fibers dorsally ("Zotten" of Altner, '71) and more ligamen- tous fibers more ventrally. All these features were also found in the other lorisiforms dis- sected. An intracapsular fat pad intervenes between the radius and the dorsolateral part of the joint capsule, and a synovial fold was found depending from the dorsal labrum onto the dorsal edge of the radial articular surface in one specimen.
The ulnocarpal joint is of the primitive mammalian sort, with the large distal end of the ulna fitting into a cup formed by the tri- quetrum and pisiform.
The distal radioulnar joint of G. crassi- caudatus is a diarthrosis, and the radial and ulnar articular surfaces are closely apposed. In one of the two specimens dissected, the ar- ticular cartilage of the left ulna was sur- rounded by a synovial bursa, but lacked direct
contact with the radius, so that the joint was a t first taken to be a modified syndesmosis. However: the x-ray of this joint presented an abnormal appearance, with a wide radiolu- cent gap between radius and ulna (fig. 5 ) , and we regard the morphology seen here as an artifact of some traumatic dislocation.
2. Galago senegalensis. The specimen of G. senegalensis dissected differed in no signifi- cant respect from G. crassicaudatus.
3. Loris tardigradus (fig. 7). The ulnocar- pal joint of Loris is greatly reduced by com- parison with the more primitive arrangement seen in Galago. The pisiform is displaced dis- tally and ventrally, so that it articulates only with the triquetrum. The distal end of the ulna forms a slender styloid process, bearing a small terminal articular surface that ap- parently comes into direct contact with the triquetrum only during ulnar deviation and dorsiflexion of the hand. The articular sur- faces of the triquetrum and ulnar styloid ar- ticulate principally with meniscus-like in- ward projections of the surrounding joint cap- sule. The longitudinal septum of the wrist joint is excavated on its ulnar side to form a socket for the proximal extremity of the tri- quetrum, and the portion of the capsule which intervenes between the ulna and pisiform bears a clearly demarcated circular articular surface which receives the ventral aspect of the triquetrum's proximal half. During ulnar deviation, the triquetrum evidently swings proximally and radially into its cavity in the longitudinal septum (fig. 71, its ulnar surface sliding along the flat, radiodistally-facing ar- ticular tip of the styloid process. The ulnotri- quetral contact thus does not appear to re- strict ulnar deviation of the hand.
In Loris, as in the other lorisines dissected, the dorsal capsular labrum for the centrale is faired into the distal articular surface of the radius, and a small facet on the dorsal edge of that articular surface receives the proximal extremity of the centrale's dorsal face during dorsiflexion. In one specimen of Loris, the radiocarpal and ulnotriquetral joint cavities communicated through a perforation in the longitudinal septum, ventral to the t r i - quetrum's articular socket in the septum. A similar ventral perforation has been reported in Papio; a dorsal perforation occurs in some other mammals (Hughes, '44; Davis, '64).
The distal radioulnar joint of Loris tar- digadus is a diarthrosis of typical lorisine form (fig. 5).

258 MATT CARTMILL AND KATHARINE MILTON
S
R L
LU
H
-T
I
Fig. 7 Right wrist joint of Loris tardigradus, opened dorsally. Semidiagrammatic. Abbreviations: L, longi- tudinal septum: LU, lunate; R, radius; RL, palmar radiocarpal ligament; S, scaphoid; SU, styloid process of ulna; T, triquetrum. Arrow indicates inferred movement of triquetrum during ulnar deviation. Compare figure 6.
4. Perodicticus potto. In the single speci- quetrum and the styloid process along their men dissected, retreat of the ulna from the postaxial edges. This amounted to little more triquetrum was perhaps slightly more ad- than an exaggeration of a similar capsular vanced than in Loris. A fibrous or fibrocarti- ridge seen in Loris, however, and direct laginous ridge projected radially from the ulnotriquetral contact persists in both. An inner surface of the capsule a t the ulnar edge intracapsular fold of synovial tissue lying of the joint, intervening between the tri- against the dorsal and preaxial edge of the
LOKIS WHIST AND HOMINOID EVOLUTION 259

260 MATT CARTMILL AND KATHARINE MILTON
distal radius was interposed there between radius and scaphoid. In other observed aspects of i ts morphology, Perodicticus resembles Loris. Etter (’74) claims that the potto’s pisiform “participates” in the ulnocarpal joint (“nimmt Anteil am Ulno-Carpal-Ge- lenk”); in the one preserved specimen and four ligamentous skeletons we examined, this was clearly not the case, unless Etter’s phrase implies only that a single synovial cavity enfolds the pisiform, triquetrum and ulnar styloid process.
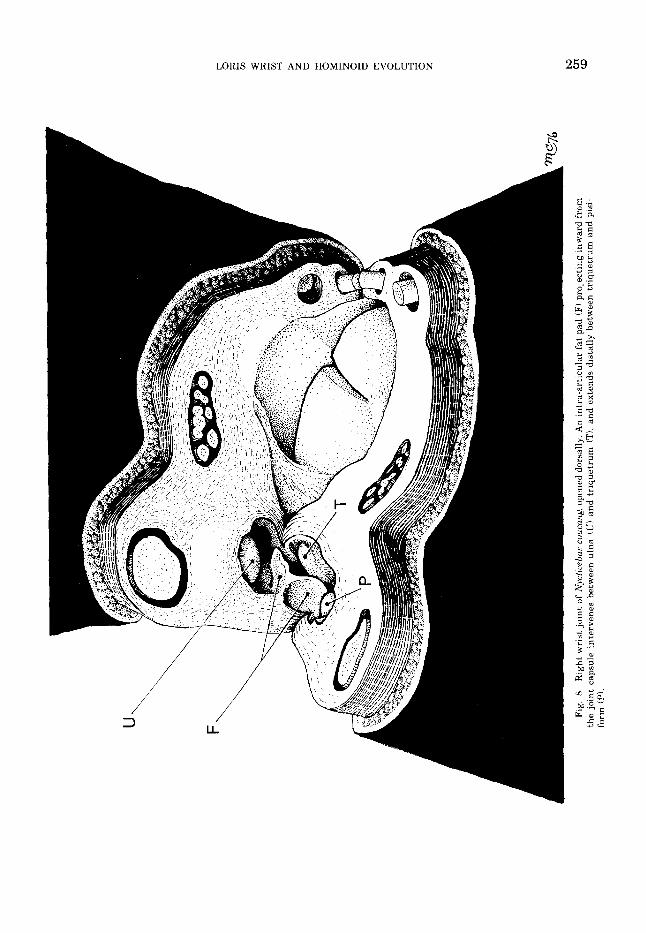
5 . Nycticebus coucang. The proximal wrist joint of N coucangis essentially like that of other lorisines, but the ulnocarpal contacts varied considerably in the four specimens examined. In one (U.S. Natl. Mus. No. 2683111, there was a substantial area of po- tential direct contact between ulna and tri- quetrum, not differing significantly from that seen in Loris and Perodicticus. In a second specimen (U.S. Natl. Mus. No. 105211, the potential area of contact between ulna and triquetrum was much reduced, and the con- formation of the triquetrum’s proximal artic- ular surface appeared to prohibit close-packed contact between the triquetrum and the ulnar styloid. In radial deviation, the ulna and tri- quetrum were apparently widely separated, the intervening space being filled by fatty synovial folds projecting inward from the ulnar side of the joint capsule. In the remain- ing two specimens, procured from the Duke University Primate Facility, a dense intra-ar- ticular fat pad of complex shape intervened
completely between the ulna and triquetrum (fig. 81, eliminating the ulnocarpal contacts altogether. The thin edges, elastic consisten- cy, and white color of this pad led us to misi- dentify it as a true meniscus in a preliminary report (Cartmill and Milton, ’74) ; however, sections stained with methylene blue and an oil-soluble dye conclusively show it to be mature adipose tissue, enclosed in a fold of synovium.
DISCUSSION
Functional significance The observations presented here show that
the wrist of lorisines (and of some Nycticebus in particular) is not typically mammalian, but has undergone a transformation as radical as, and in many respects parallel to, that seen in the extatnt Hominoidea (fig. 9: table 1). I t is no longer possible to maintain that the hominoid wrist joint is characterized by a “morphologi- cal arrangement . . . markedly dissimilar to that of other Primates” (Lewis, ’691, or that hominoid-like specializations of the wrist can be taken as indicators of brachiating locomo- tor habits.
Lorises resemble hominoids and differ from galagos in several other features that have been analyzed as brachiating adaptations in the hominoids. These reportedly include rela- tively high intermembral indices (Napier and Napier, ’67; Walker, ’74); increased length of the scapula’s vertebral border, enlarged infra- spinous fossa, moderate craniolateral redirec- tion of the glenoid cavity of the scapula,
TAB1.E 1
Comparative morphology of the wristjoint in anthropoids and lorzsiforms
Feature Ceboids Cercopithecaids Hnminoids Galagos Lorises
1.
2.
3.
4.
5 .
6 .
Joint divided into radial and ulnar halves by longitudinal septum
i i. r + +
Pisiform articulates with ulna + + Distal extremity of ulna - reduced; expansion of radiocarpal joint surfaces
- - New ulnar head: large distal radioulnar joint
Ulnotnquetral contact lost
Meniscus between ulna and triquetrum incorporated into uroxinial articular surface
The boxed pairs of symbols in the right-hand column indicate features in which lorises differ from galagos in the same way that huminoids differ frnm monkeys.

261

262 MATT CARTMILT, AND KATHARINE MILTON
hominoid-like relative weights and attach- ment patterns of shoulder musculature, and other features of the shoulder girdle (Ashton and Oxnard, ’64a; Oxnard, ’67; Roberts, ’74; Roberts and Davison. ’75); a thoracic cage whose transverse breadth exceeds its dorso- ventral depth (Straus and Wislocki, ’32); caudally-directed spinous processes of all the thoracic and lumbar vertebrae (Mivart, 1865); and a reduced tail. We assume that most of these features, and in particular those of the forelimb girdle and joints, represent locomotor adaptations - rather than, say, specializations for catching insects - and are somehow interrelated. We also assume that the resemblances to hominoids are not coinci- dental. But obviously lorises and pottos are not brachiators, no matter how loosely we de- fine that much-abused category. Do lorisine and hominoid locomotor patterns alike differ from those of other primates in some way that could account for their convergent anatomical specializations?
All the extant species of lorisines share a specialized and peculiar locomotor habit, which has been studied both in the field and on captive animals (Subramoniam, ’57; Bishop, ’64; Walker, ’69; Charles-Dominique, ’71). It has been labelled “slow climbing quad- rupedalism” by Napier and Napier (’67), who characterize i t as “a type of quadrupedal locomotion in which three of the four extremi- ties are applied t o the branch a t any given moment. Movement is always slow and cautious. The limbs may act to suspend the body or to support it.” This is not quite accur- a te - lorisines may stand on or hang by the hind limbs alone, and have a variety of rapid locomotor patterns, including gaits in which three of the four extremities are off the sup- port together a t certain points in the cycle (Cansdale, ’48; Subramoniam, ’57; Hilde- brand, ’67; Walker, ’69; Jewel1 and Oates, ’69; Charles-Dominique, ’7 1; and personal obser- vations) - but it correctly stresses the steal- thy and deliberate character of lorisine move- ment. No one, as far as we know, has ever seen a lorisine jump. When confronted with a gap between one support and the next, lorisines do not release the grip of the hind feet on the support behind until the vise-like hands have secured a fast grip on the support ahead. I t follows that their bodies must be extremely flexible, enabling them to adjust to all possi- ble differences of angulation between one sup- port and the next.
Another habit of lorises of possible impor- tance here is their willingness to walk upside down under horizontal supports as well as on top of them; several observers have reported that lorisines, especially pottos, move about with no great preference for either position, passing from one to the other without chang- ing pace (Harrison, ’49; Loveridge, ’56; Charles-Dominique, ’71). The locomotor classification adopted by Ashton and Oxnard (’64a,b) describes lorisines as “hangers,” and workers from a t least the time of Jones (’16) on have likened them to sloths.2
Either the suspensory habits of lorisines or their need for great post-cranial flexibility might be invoked to explain one or another of their resemblances to hominoids, and i t is pos- sible that both factors have been important in loris evolution. We attempt to assess them separately below.
1. Suspensory postures and tensile stresses. Reduction and retreat of the distal end of the ulna is seen not only in lorisines and hominoids, but also in Choloepus, the two-toed sloth (Yalden, ’72). This suggests the possi- bility that the modifications seen in the wrist joints of all these animals represent a change in the postural stresses borne by the forelimb, from primarily compressive to primarily tensile.
Currey (’68) proposes that, although a limb that acts as a suspensory organ must be loaded under net tension. “it may well be that muscles acting over the whole length of the limb exert a force greater than the dynamic forces that particular segment of the limb has to bear, and so put the bone itself into a state of compression.” This also implies com- pressive loading across intersegmental joints. Some students have argued that bones (and hence diarthroses) are never loaded under net tension, and that “skeletal architecture is invariably designed to reduce, or even elimi- nate, tensile forces” (Tattersall, ’74). How- ever, Tuttle and Basmajian (’74b) find that the muscles of the arm are electrically silent in gorillas hanging from overhead supports. The shoulder and forearm muscles in these animals may maintain net compression across the glenohumeral and elbow joints, but the intervening part of the humeral shaft must be carrying the animals’ weight under tension in this posture, thus proving that bones are
Harrower (’33) reports seeing his captive Nycfrcrbus coucany .after the manner of a bat.“ “sleeping auspended by one hind limb
This extraordinary observation remains unconfirmed.

a b
Fig.
10
Ext
rins
ic f
lexo
rs o
f th
e ha
nd i
n su
spen
sory
pos
ture
s: d
iagr
amm
atic
sec
tion
s th
roug
h cy
lind
rica
l su
ppor
ts.
Whe
n a
hang
ing
anim
al h
as it
s fi
nger
s nv
er t
he to
p of
a s
uppo
rt (
Al.
the
vert
ical
com
pone
nt (M
I of
the
mus
cle
forc
e li
es b
e-
twee
n th
e di
gita
l ske
leto
n an
d th
e su
ppor
t po
int,
and
its
mag
nitu
de th
us e
xcee
ds th
aL o
f th
e pa
rall
el r
eact
ion
forc
e co
mpo
- ne
nt t
hrou
gh t
he s
kele
ton
iR) o
r th
e ve
rtic
al r
eact
ion
forc
e (W) thro
ugh
the
supp
ort
poin
t. S
ince
W is
equ
al t
o th
e p
rtio
n
of t
he a
nim
al’s
wei
ght
born
e by
the
dig
it, m
uscl
e fo
rce
exce
eds w
eigh
t bor
ne f
or a
ach
digi
t an
d th
eref
ore
for
the
hand
as
a w
hole
. If
all
forc
es a
re p
aral
lel,
M/W
is m
inim
al a
nd e
qual
to
(a-
b,/a
. W
hen
mus
cle
forc
e is
col
inea
r w
ith
grav
itat
iona
l re
acti
on f
orce
thr
ough
the
sup
port
tB
j, th
e an
imal
is s
uppo
rted
by
fric
tion
and
the
wei
ght
supp
orte
d (W
l equ
als
or i
s le
ss
than
@ (t
he c
oeff
icie
nt o
f fr
icti
on b
etw
een
body
and
sup
port
sur
face
) ti
mes
A (
the
reac
tion
for
ce n
orm
al t
o th
e co
ntac
t po
int)
. Sin
ce W
@
A an
d A
(M
pjiq
, W =
(M
pp
)iq
; thu
s, W
/’M : (pp
jiq,
and
W>
M o
nly
if p
p>
q.

264 MATT CARTMILL AND KATHARINE MILTON
sometimes loaded under tension even when mechanisms to prevent this are available. Yet even if some of the bones and joints of the loris forelimb arc loaded under net tension, we doubt that this ever occurs a t the wrist joint. The vector of the extrinsic digital flexor force always lies between the support point and a t least one of the joints of the hand (fig. 10A); the muscle force exerted across the wrist must therefore exceed the force exerted by gravity, if the extrinsic flexors are relied upon to maintain the grip. If the hand is fixed into a hook-like configuration by non-muscular structures or by intrinsic hand muscles, so that the extrinsic flexors are not of critical importance in maintaining flexion a t the in- terphalangeal or metacarpophalangeal joints, net tension across the wrist might be possible; but we regard this as unlikely in primates (though it might be true, for example, of sloths). An animal might, in theory, suspend itself from a support without flexion of any of the joints of the hand, relying on frictional forces for support; but forces exerted by the extrinsic flexors will still exceed the animal's weight unless the terminal phalanx is bizarre- ly shaped or the animal's pads have an ex- traordinarily high coefficient of friction (fig. 10B). Net tension across the wrist probably never occurs in primates hanging by their forelimbs.
But even if lorisine wrists sustained net tension in suspensory postures, we still would lack an explanation of the carpal differences between lorisines and galagines, since the forelimbs of some galagos must be loaded under tension at least as frequently as are those of some lorisines. Although the largest and smallest galagos are essentially quad- rupedal in their posture and locomotion, Galago alleni and G. senegalensis are accus- tomed to cling to and climb on vertical sup- ports (Charles-Dominique, '71; Doyle, '74a,b), which necessarily places the upper limb under net tension (Cartmill, '74). Galago senegalen- sis also occasionally shows a potto-like pro- clivity for walking upside down underneath branches (Doyle, '74b). While some published analyses assume that differences in forelimb morphology between galagos and lorises reflect the greater tensile stresses affecting the lorises (Ashton and Oxnard, '64a), others (Walker, '74) invoke tensile stresses to account for certain distinctive features of the forelimbs of gulagos. At present, there seems no adequate reason to think that the two sub-
families a re distinguished by systematic differences in overall tensile loading of the forelimbs.
2. Slow movement and joint mobility. The Galaginae and Lorisinae may not differ in net tensile loading of the forelimb; but they do differ with respect to the maximum levels of compressive stress which their joints must be able to withstand. Galagos jump from one support to another, but lorises do not. The joints and bones of galagos are therefore sub- jected to high transitory stresses on landing, with corresponding risks of incurring disloca- tions like that found in one of the galagos uti- lized in this study (fig. 5 ) . The slow, fluid, unbroken rhythm of lorisine locomotion sub- jects them to no such risks3
Although a loris is not subjected to the strains and sprains involved in leaping from one support to another, it incurs the compen- satory difficulty of having to reach out pre- cisely and grab any support to which it wishes to move. Doing this requires a somewhat brachiator-like reorganization of the shoulder musculature to allow the animal to reach above its head and pull its body weight in a cranial direction (Ashton and Oxnard, '64a; cf. Charles-Dominique, '71: fig. 24). I t also requires enhanced mobility of the limbs. Does the reorganization of the lorisine antebrachio- carpal and distal radioulnar joints serve to enhance their mobility? These joints are involved in rotatory movements of the hand around three roughly perpendicular axes: a longitudinal axis (pronation-supination), a dorsoventral axis (abduction-adduction), and a transverse axis (dorsiflexion-volarflexion) . The last of these is probably not facilitated by the lorisine specializations of the wrist, but the other two may be. Since pronation and supination are the only movements which affect the distal radioulnar joint, the changes here must be related to these movements. I t is easy to see that pronation and supination are limited a t this joint by the relatively flat shape of the articular surfaces in galagos, and facilitated by the joint's conversion into something like a ball-and-socket joint in lorises.
The functional significance of the reduction of ulnocarpal contact in lorisids and homi-
Nyrtkebus will occasionally bridge a large gap between supports by a semiacrobatic "trapeze swing," in which the animal throws its body toward the support ahead and grabs a t it with its hands, while the hind feet retain a firm grip which prevents the animal from fall- ing if i t missea (J. McArdle, personal communication). A8 far as we know. this maneuver represents the extreme in lorisine acrobatics.

LORIS WRIST AND HOMINOID EVOLUTION 265
noids is not so clear. Lewis regards this change as serving essentially to free "the eccentrically situated ulnar styloid process from its restricting articulation with the tri- quetral and pisiform . . . (so that) the radius, carrying with it the whole carpus . . . (can) rotate about the neomorphic ulnar head" (Lewis, '69). In Lewis' view, this change inci- dentally facilities ulnar deviation of the hand. Other investigators (Parsons, 1899; Jones, '42; Conroy and Fleagle, '72) have proposed that reduction of the ulnocarpal joints serves mainly to augment ulnar d e ~ i a t i o n . ~ We see no conflict between these two hypotheses, and both factors may well be involved in the lorisine case. Lorises almost invariably grip branches in such a way that the axis of the branch passes through or to the radial side of the reduced index finger, while the grips of galagos are much more variable in orientation (Bishop, '64). To maintain their stereotyped grasping p a t t e r n while moving among branches of differing angulation, lorisines require a correspondingly enhanced flexibility of the forelimb joints, and in particular of the wrist. I t seems likely that the lorisine special- izations of the wrist serve this requirement by augmenting both ulnar deviation and supina- tion. There is a t present no empirical evidence bearing on this prediction, and it needs to be tested on anesthetized animals.
The fat pad which separates the ulna from the triquetrum in some specimens of Nyc- ticebus is unusual in projecting between the articular surfaces of these bones. Although simple synovial folds may extend between ar- ticulating bones (Grant, '311, intra-articular fat pads are normally restricted to the mar- gins of joints, where they apparently serve to fill up excess space and thus keep the film of synovial fluid in the joint thin enough to per- mit optimally efficient lubrication (Barnett et al., '61). Because they are richly innervat- ed, pain results when they are accidentally nipped between opposing joint surfaces. The variable intrusion of a fat pad into the ulno- triquetral joint of Nycticebus thus indicates that this joint transmits little or no com- pressive force, and never comes into close- packed position. We conclude that in Nyc- ticebus, and perhaps in other lorisines, the retreat of the ulna from the carpals is func- tionally complete, and that the ulna and tri- quetrum do not come into contact even in ex- treme ulnar deviation - a conclusion sup- ported by our radiographic observations.
Evolutionary significance The slow and deliberate locomotion of
lorises, which protects their joints from tran- sitory high stresses but demands great flexibility, has resulted in considerable con- vergence with modern hominoids in the anat- omy of the wrist joint. Admittedly, the extant hominoids share several specialized features of wrist morphology not seen in lorises (where other, functionally equivalent specializations have been developed). These shared hominoid specializations may well have been present, in a rudimentary form in the extant hominoids' last common ancestor, which probably pos- sessed a triangular articular disk and perhaps some sort of incipient ulnocarpal meniscus. But the convergent similarities between lorises and modern hominoids demonstrate that the reduction of the ulnocarpal articula- tions in the lineages leading to man and the extant apes need not in every case represent an adaptation for bimanual suspension. We suggest that reduction or loss of the ulnocar- pal articulations, in the larger apes as well as in lorises (and probably also in Choloepusl, is related to the fact that these animals do not ordinarily leap across gaps between branches, but bridge them by stretching across and grabbing the support ahead before relinquish- ing the one behind.
I t is not clear why lorises have adopted this non-saltatory sort of locomotion in the trees. In the case of the larger hominoids, rather cautious locomotor habits follow inescapably from their large body size. Because of the square-cube law, a large arboreal animal must either utilize disproportionately large sup- ports or distribute its weight over several sup- ports a t once. A large animal is also less likely to survive if it falls from any great height; the kinetic energy which the tissues of i ts body must dissipate on impact increases in propor- tion to the cube of its linear dimensions and to the square of its terminal velocity (which is in turn increased by the low surface-to-mass ratio). Since large arboreal mammals face in- creased risks both of falling and of being killed in a fall, we would expect them to move cautiously in the treetops, grasping several supports at once whenever possible, and cross-
O'Connor ?75: finds that removing the ulnar styloid process from freshly dead cercopithecines results In a slight increase in the range of ulnar deviation, hut not in that of supination. The relevance of this experiment is impossible to assess without determining the relative contributions of muscles, capsular ligaments, and joint surface shape to the observed limitation of motion: evolutionary change would affect all these systems concurrently, which surgery does not.

266 MATT CARTMILL AND KATHARINE MILTON
ing gaps by reaching over to the support ahead and pulling or swinging themselves across instead of jumping - in short, t o move around rather as lorises do, though for dif- ferent reasons. Our model for this kind of be- havior - which Tuttle ('75b) has called "transferring" - is the orangutan, the most arboreal of the large-bodied hominoids.
Mere increase in body size would also be ex- pected to lead to elongation of the forelimbs, for a variety of reasons. In catarrhines, rela- tive forelimb length increases as a function of body size (Biegert and Maurer, '72). This allometric relationship is partly related t o the biomechanics of climbing vertical supports (Cartmill, '74). Large arboreal primates also need relatively longer arms for reaching food growing far out on terminal branches too slender to bear the animals' weight, and for "transferring" across support discontinuities that a big animal cannot afford to cross by leaping.
I t has been proposed that the transversely broad rib cage characteristic of hominoids is also a product of allometry in the larger apes, with gibbons again deviating grossly from the general regression and so displaying a spurious resemblance to the great apes and man (Andrews and Groves, '76). Neither data nor any functional interpretation have yet been advanced in support of this idea. We sug- gest that a transversely broad thorax may reflect not body size per se, nor yet upright posture, as Keith ('23) originally claimed, but a need for increased shoulder mobility; having a broad thoracic cage redirects the glenoid sockets laterally and moves them further apart, and so increases the span of the arms and their range of circumduction. This fact accounts for the moderately accentuated breadth of the rib cage reported in some lorises and sloths (Straus and Wislocki, '321, which are neither large nor orthograde, as well as for that seen in the arm-swinging gib- bons and spider monkeys (Mivart, 1865; Erik- son, '63).
The disproportionately long arms and rela- tively primitive wrist joints of gibbons proba- bly reflect a long history of adaptation to bimanual locomotion not shared by the lineages leading toward the great apes. This hypothesis is compatible with the known fossil evidence. The Miocene hominoids whose post-cranial remains are most completely known appear to have been arboreal quad- rupeds, but they represent two rather
different sorts of arboreal quadrupedalism. Pliopithecus and Dendropithecus, frequently regarded as having special affinities with gib- bons, are distinguished from the dryopith- ecines by their relatively gracile, straight- shafted long bones (Le Gros Clark and Thomas, '51; Zapfe, '58; '60). They resemble gibbons and atelines in these respects, and are often likened to Ateles in overall postcranial morphology (Simons and Fleagle, '73). The other sort of locomotor adaptation seen in the Miocene is that of the smaller dryopithecines, the best-known of which is D. africanus. Its retroflexed humerus and cercopithecid-like joint surfaces show it to have been a largely pronograde quadruped, but it had a more ape- like distal humerus and a more mobile elbow joint than either Pliopithecus or Dendropith- ecus (Napier and Davis, '59; Morbeck, '72, '75, '76; Preuschoft, '73). Dryopithecus africanus resembled Alouatta in size, and has been likened to it in locomotor adaptations by some investigators (Simons, '72; Schon and Ziemer, '73). The postcranial remains of larger dryopithecines are poorly known, although the supposed D. fontani humerus from St. Gaudens indicates that some of them were more ape-like than D. africanus (Pilbeam and Simons, '71; Morbeck, '72; Preuschoft, '73).
Two locomotor types are also distinguisha- ble in the earliest fossil catarrhines, from the Fayum deposits of Egypt. Postcranial ele- ments recovered from these deposits include remains of a t least two primate species, one larger than the other (Conroy, '74). The smaller animal, possibly a species of Apidium, resembles small cebids like Aotus and Saimiri in known parts. The larger animal, probably Aegyptopithecus zeuxis, is known principally from a right ulna. The ulna resembles that of Alouatta in gross size and in certain morpho- logical details - e.g., a small and discrete ar- ticular surface for the proximal radius - which suggests that the forearm of Aegyp- topithecus was capable of a rather large range of pronation and supination (Conroy, '74; Fleagle et al., '75). Preuschoft ('74) also con- cludes, on different grounds, that A e n p - topithecus was "most similar to Alouatta among the recent monkeys."
Published observations of the locomotion of Alouatta in the field, summarized by Stern ('711, as well as our own field studies of this large-bodied ceboid (K. Milton, unpublished data), show that Alouatta is a moderately slow-moving and cautious quadruped, seldom

LORIS WRIST AND HOMINOID EVOLUTION 267
leaping or engaging in other kinds of rapid acrobatic locomotion. As a rule i t moves rather sedately through the trees, retaining a grip with the feet and tail on the terminal branches of one tree as i t stretches its arms and body out to grasp the terminal branches of the next. Although Alouattu practical- ly never engages in bimanual suspension (Richard, ’70; Mendel, ’761, the few specializa- tions which it shares with hominoids (short- ened lumbar region, relatively long forelimbs, and moderate diagonal reorganization of the scapula) have led some researchers to classify i t as a brachiator or “semibrachiator” (Erik- son, ’63; Ashton and Oxnard, ’64b; Ashton e t al., ’65; Oxnard, ’67; Napier and Napier, ’67). The contrary inference seems sounder: name- ly, that the presence of these traits in Alouat- ta shows that some aspects of “brachiating” morphology can appear in connection with cautious arboreal quadrupedalism, and need not have anything to do with bimanual suspension.
The scanty fossil evidence suggests that primitive catarrhines were small animals with an unspecialized quadrupedal locomotor pattern like that of the smaller cebids. The lineage leading toward modern cercopithecids probably preserved this pattern. The largest of the Fayum catarrhines, Aegyptopithecus, shows signs of having been a rather cautious arboreal quadruped like Alouattu. In the New World, quadrupeds of this type gave rise to forms like Ateles, whose straight, elongated forelimbs and reorganized shoulder muscula- ture and scapula reflect their suspensory locomotor habits. By a similar process, the hylobatids evolved out of an ancestor some- thing like Aegyptopithecus. Dendropithecus may represent a transitional stage in this process. In the Old World, the hylobatids’ cautious quadrupedal ancestor lacked Alout- ta’s prehensile tail (itself an adaptation to cautious arboreal locomotion, as in arboreal marsupials, rodents, and edentates). The hylobatids’ defective caudal grasp obliged them to develop a locomotor pattern in which they spend half their time hanging from and rotating around one forelimb while protract- ing the other. The relative insecurity of their one-handed grip has afforded them the oppor- tunity of becoming more acrobatic than the “three-hooked’’ New World arm-swingers, whose third support point improves stability a t the cost of eliminating the phase of free flight in the footfall sequence. As Lewis (’72a)
notes, the phase of free flight in the rapid brachiation of gibbons may have permitted the retention of a more primitive wrist joint than that seen in the larger hominoids.
The great apes, like the atelines, also lack a phase of free flight in their suspensory loco- motion, but for a different reason; they are simply too big to go flying through the treetops like a brachiating hylobatid. The fossil evidence suggests that this has always been the case. Oligocene and early Miocene pongids may have had body weights small enough to have permitted gibbon-like “rico- chetal arm-swinging” (Tuttle, ’691, but their postcranial remains indicate that they re- mained pronograde quadrupeds like Alouatta. The later Miocene and Pliocene dryopith- ecines apparently had body weights roughly covering the range seen in extant great apes (Simons and Pilbeam, ’651, and were probably too large to have been ricochetal arm- swingers. I t thus seems likely that the great apes developed their “brachiating” adapta- tions directly from a cautiously-moving quad- rupedal ancestor without going through a phase characterized by hylobatid-like bi- manual suspensory behavior.
Other recent workers have reached similar conclusions on other grounds (Tuttle, ’75b; Fleagle, ’76). Stern’s (’71) exemplary analysis of primate hip and thigh musculature also indicates that the pongids and hominoids passed through a phase of cautious quad- rupedalism; the great apes and man resemble Alouattu, but differ from other ceboids, cer- copithecoids and gibbons, in a number of myological features mechanically related to slow movement and hindlimb suspension. The most important of these are the extensive and fused origins of the vasti - a feature also characteristic of lorisines (Stern, ’71; J. McArdle, personal communication). Stern concludes that the last common ancestor of the great apes and man
”. . . employed its forelimb much as does the living orangutan and its hindlimbs in a somewhat more pro- nograde quadrupedal manner than does Pongo. Such might be the behavior of a smaller bodied version of the orangutan that had begun to employ its forelimbs extensively in climbing, suspension and other tensile activities, hut had not yet reached the size which was to cause i t to abandon so completely pronoyade quad- rupedality.” (Stern, ’76).
But there are a number of supposed adapta- tions to orthograde posture that are found in hylobatids, pongids, and hominids alike. The most important of these are the following: (1)

268 MATT CARTMILL AND KATHARINE MILTON
ventral displacement of the thoracic vertebral bodies toward the line of gravity of the upright trunk; (2) increase in the number of sacral vertebrae and (3) decrease in the num- ber of lumbar vertebrae, facilitating stabili- zation of the upright trunk; (4) reduction of the tail and refashioning of tail musculature for support of the pelvic viscera in an upright posture; ( 5 ) mesenteric adhesions that pre- vent lapsus of the colon and other abdominal viscera when an upright posture is assumed; and (6) fixation of the fibrous pericardium to the thoracic surface of the diaphragm. In gib- bons, these features have been interpreted by Keith (’23) and his successors as adaptations to the orthograde posture involved in bi- manual suspensory locomotion. How can we account for the presence of these gibbon-like traits in the great apes and man if - as Stern suggests - their last common ancestor had not abandoned pronograde quadrupediality?
Some of these traits are also found in lorisines, demonstrating that they do not necessarily have much to do with upright posture. The ventral displacement of the ver- tebral column is an aspect of the broadening of the thorax, which is at least partly related to the reorientation of the glenoid and associ- ated changes in the shoulder musculature, and is paralleled to a slight extent in lorisines. Lorisines also resemble hominoids in having numerous sacral vertebrae but differ from them in having an unreduced number of lum- bar vertebrae and an increased number of thoracic vertebrae (Mivart, 1865; Schultz, ’61). Alouatta resembles lorisines in the last respect, but has fewer lumbar vertebrae. We suggest that the differences here may be allometric. Since neither lorises nor great apes ordinarily jump across gaps between sup- ports, they must bridge them with arms and trunk. Lorisines are small enough to be able to rely largely on truncal rigidity in bridging such gaps, holding on to the support behind with the hind feet and stretching the rigidly held trunk out toward the support ahead. Elongation of the trunk facilitates this; and i t is logical that it should take place primarily in the more rigid thoracic part of the trunk. The peculiar specialized ribs of Arctocebus have been interpreted as adaptations for this sort of “bridging” behavior by Jenkins (’701, who notes similar specializations in some slow-climbing Xenarthra. The larger hom- inoids also need rigid trunks in reaching out across gaps; but they are so large and top-
heavy (due in part to forelimb elongation) that they cannot afford to retain a supple lum- bar vertebral column, and have accordingly shortened the lumbar region of the spine by reducing both the number and the height of individual vertebrae (Benton, ’67). In both lorisines and hominoids - as in Alouatta and the atelines - the effect is the same: the thorax contributes disproportionately to total trunk length (Schultz, ’61).5 The increased breadth of the thorax in hominoids also improves control over lumbar movements by lengthening the moment arms of the lateral abdominal muscles (Keith, ’23). Ventral dis- placement of the vertebral column may simi- larly improve the leverage of some of the back muscles (e.g., iliocostalis). Sacralization of a number of posterior lumbar or anterior caudal vertebrae, characteristic of lorisines and hominoids, provides what Keith (’23) de- scribed as “a fixed base from which the erector spinae may act.” Keith, with Homo in mind, regarded this as related to upright posture in hominoids; but i t makes sense both in hominoids and in lorises as an additional mechanism for increasing control over move- ment between thorax and pelvis when cross- ing gaps between supports.
The tail of mammals serves as a balancing organ in locomotion, and also as an important site for the radiation of excess heat (Thoring- ton, ’66). Cautious quadrupedal locomotion, which diminishes the importance of both functions, has led to tail reduction in the lorisines. A similar explanation could account for the loss of the external tail in a slow- climbing quadrupedal ancestor of the great apes and man. Catarrhines, including most cercopithecoids, are distinguished from other primates by insertion of the pubococcygeus muscle into the rectum (Elftman, ’321, and even short-tailed cercopithecoids retain an unreduced pubococcygeus for support and retention of the pelvic viscera (Wilson, ’72). Parallel loss of the tail in different hominoid lineages would therefore be expected to result in the conversion of the ventral pelvocaudal musculature into a levator ani, as Elftman (‘32) noted.
The visceral peculiarities of hominoids, however, are still best explained as postural adaptations. Keith (’231 believed that the fibrous pericardium’s attachment to the dia-
This is true even in Loris turdgradus, which has an cxccptianally large number uf lumbar vertebrae and lacks the elongated sacrum seen in the other lurisines (SchulLz, ‘61)

LORIS WRIST AKD HOMINOID EVOLUTION 269
phragm in hominoids converted the medi- astinum into a mechanical link between the diaphragm and the roots of the lungs, forcing the roots of the lungs to descend slightly dur- ing abdominal inspiration and consequently improving the aeration of the apical lobes. In hominoids, these lobes are enlarged a t the ex- pense of the primitively large retrodiaphrag- matic parts of the lungs, and the diaphragm is correspondingly more flat-topped and less obliquely slanted caudally and dorsally. All of this makes sense as an adaptation to upright posture, in which the force of gravity tends to increase intra-abdominal pressure and thus hinder inflation of any part of the lungs which extends down between the diaphragm and the back musculature. The hominoid specializa- tions of the abdominal mesenteries are also presently explicable only as adaptations to upright posture. This may mean that the lineages leading toward the great apes and man went through a gibbon-like “hylobatian” stage. Alternatively, i t may mean that they went through a stage of predominantly or- thograde slow suspensory quadrupedalism resembling that of the orangutan. The latter interpretation is compatible with Stern’s (’76) thesis that the last common ancestor of the great apes and man had not completely aban- doned pronograde quadrupedality. An only partially orthograde quadruped would still be subjected to selection pressure to develop mesocolic and pericardial fixation: and in- deed, Brachyteles and Ateles show some slight convergence with hominoids in these respects (Hill, ‘62; Juraniec and Szostakiewicz- Sawicka, ’68). The importance of suspensory locomotion and postures in an arboreal mam- mal’s behavioral repertoire should vary as a function of its body weight, since a large ani- mal must find it easier to hang below a rela- tively small branch than to balance atop i t (Napier, ’67). A large slow-climbing quad- ruped which had already developed long and mobile forelimbs for the reasons proposed above would develop increasingly suspensory habits if it continued to increase in body size. Its forelimbs would inevitably become the principal organs of suspension, and it would assume a progressively more frequent ortho- grade posture, thus selecting for hominoid- like fixation of the viscera. Again, simple in- crease in body size might be expected to yield fully hominoid morphology from only partly ape-like antecedents.
I t is worth noting in conclusion that the
most ape-like postcranial morphology seen among non-hominoid primates is found in two large-bodied arboreal species: (1) the extinct lemuriform Palaeopropithecus ingens, which shows a number of resemblances both to orangutans and to sloths (Carleton, ’36; Walker, ‘74; Jungers, ’77) and probably had a locomotor pattern of “steady, rather slow, four-limbed climbing” (Walker, ’74) ; and 12) the chimpanzee-sized Miocene catarrhine Oreopithecus barnbolii, which resembles homi- noids sufficiently closely to have been classi- fied with them by many investigators, despite its decidedly unhominoid molars and ankle joint (Simons, ’60, ’72; Straus, ’63; Delson and Andrews, ’75; Szalay, ’75). If Oreopithecus is a hominoid, it demonstrates that some large- bodied Miocene hominoids had progressed beyond the more primitive morphology seen in Dryopithecus africanus; if it is not a hominoid, it shows that a t least some ape-like traits were developed in parallel by more than one catarrhine lineage. In either event: it vitiates some of the arguments for a late divergence of the lines leading to the extant hominoids.
ACKNOWLEDGMENTS
We are grateful to J. Buettner-Janusch, W. C. Hall, F. Vincent, and the Duke University Primate Facility for providing cadavers used in this study, and to H. W. Setzer and R. W. Thorington, Jr . for giving the senior author access to facilities and collections under their care a t the U. S. National Museum. The senior author’s colleague W. Longley kindly allowed him to radiograph cadavers using apparatus designed and built by Doctor Longley for use in x-ray diffraction studies. We thank K. Brown, R. S. Corruccini, J. G. Fleagle, W. L. Hylander, R. F. Kay, A. L. Rosenberger, J. T. Stern, Jr., and T. L. Strickler for their help and advice. The senior author’s research is supported by Grant 5-K04-HD00083-02 from the National Institutes of Health.
LITERATURE CITED
Andrews, P., and C. P. Groves 1976 Gibbons and brachia- tion. Gibbon and Siamang, 4: 167-218.
Al tner , G . 1971 Histologische und vergleichend- anatomische Untersuchungen zur Ontogenie und Phylogenie des Handskeletts von Tupaid glis (Diard 1820) und Microcebus murinus (J. F. Miller 17771. Folia Primatol. (Suppl.). 14: 1-106.
Ankel, F. 1965 Der Canalis Sacralis als Indicator fur die Lange der Caudalregion der Primaten. Folia Primatol., 3: 263-276.
Vertebral morphology of fossil and extant primates. In: The Functional and Evolutionary Biology
1972

270 MATT CARTMILL AND KATHARINE MILTON
of Primates. R. H. Tuttle, ed. Aldine-Atherton. Chicago, pp. 223-240
Functional adapta- lions in the primate shoulder girdle. Proc. Zool. Soc. Lon- don. 242: 49-66.
1964b Locomotor patterns in primates. Proc. Zool. Soc. London. 142: 1-28,
Ashton. E. H., C. E. Oxnard and T. F. Spence 1965 Scapular shape and primate classification. Proc. Zool. Soc. London, 145: 125.142.
Avis, V. 1962 Brachiation: the crucial issue for man’s ancestry. Southwest. J . Anthrop., 18: 119.148.
Baldwin. L.. A., and G. Teleki 1976 Patterns of gibbon be- havior on Hall’s Island, Bermuda. A preliminary ethogram for ffylobates lar. Gibbon and Siamang. 4: 21-105.
1961 Synovial Joints: Their Structure and Mechanics. Charles C Thomas, Springfield. Illinois.
Renton. R S 1967 Morphological evidence for adapta- tions within the epaxial region of the primates. In: The Baboon in Medical Research. Vol. 2. H. Vagthorg, ed. Univ. Texas Press, Austin, pp. 201-216.
Bicgert. J., and R . Maurer 19i2 Rumpfskelettlange, Allometrien und Korperproportionen bei catarrhinen Primaten. Folia Primatol., 17: 142-156.
1964 Use of the hand in lower primates. In: Evolutionary and Genetic Biology of Primates. Vol. 2 . J. Buettner-Janusch, ed. Academic Press, New York, pp. 133 225.
Botez, 1. G. 1926 Etude morphologique et morphogeni- que du squelette du bras et de I’avant-bras chez les Pri- mates. Arch. Murph. Gkn. e t Exp., 24: I 172.
Cansdale, G. S. 1948 The Calabar potto or anpant lbo . Zoo Life (London), 3: 121-122.
Carleton, A. 1936 The limb hones and vertebrae of the extinct lemurs of Madagascar. Proc. Zool. SOC. London.
Carpenter, C. R. 1976 Suspensory behavior of Qbbons. Gibbon and Siamang, 4: 1-20.
Cartmill, M. 1974 Pads and claws in arhoreal locomo- tion. In: Primate Locomotion. F. A. Jenkins, Jr.. ed. Academic Press, New York. pp. 45 83.
Cartmill. M., and K. Milton 1974 The lorisiform wrist joint. Am. J. Phys. Anthrop.. 41: 471.
Charles-Dominique, P. 1971 Eco-ethologie des Prosi- miens du Gabon. Biol. Gabonica. 7: 121-228.
Conroy, G. C. 1974 Primate Postcranial Remains from the Fayum Province, Egypt, U.A.R. Ph.D. thesis. Yale University.
Conroy. G. C.. and J. G. Fleagle 1972 Locomotor behaviour in living and fossil pongids. Nature, 237: 103-104.
Corruccini, R. S.. R. L. Ciochon and H. M. McHenry 1975 Osteometric shape relationships in the wrist joint of some anthropoids. Folia Primatol., 24: 250-274.
The postcranium of Miocene hominoids: were dryopithecines merely “dental apes”? Primates [Japan). 17: 205-223.
Currey. J. D. 1968 The adaptation of hones to stress. J. Theoret. Biol., 20: 91-106.
Davis. D. D. 1964 The giant panda: a morphological study of evolutionary mechanisms. Fieldiana (Zool. Memoirs), 3: 1-339.
Delson, E.. and P. Andrews 1975 Evolution and interrela- tionships of the catarrhine primates. In: Phylogeny of the Primates. W. P. Luckett and F. S. Szalay, eds. Plenum, New York, pp. 405-446.
Doyle, G. A. 1974a The behaviour of the lesser hush- baby. In: Prosimian Biology. R . D. Martin, G. A. Doyle
Ashton. E. H.. and C. E. Oxnard 1964a
Barnett. C. H., D. V. Davies and M. A. MarConaill
Bishop. A.
1936: 281-307.
1976
and A. C. Walker, eds. Duckworth, London, pp. 213-231. 197413 Behaviour of prosimians. In: Behavior of
Nonhuman Primates. Vol. 5. A. M. Schrier and F. Stnllnitz, eds. Academic Press. New York, pp. 155-353.
Elftman. H. 0. 1932 The evolution of the pelvic floor of primates. Am. J. Anat., 51: 307-346.
Erikson, G. E. 1963 Brachiation in New World monkeys and in anthropoid apes. Symp. Zool. SOC. London, 10: 136- 164.
Etter, H.-U. F. 1974 Morphologisch- und metrisch- vergleichende Untersuchung am Handskelett rezenter Primaten. 11. Gegenb. Morph. Jb., 120: 153-171.
Fleagle, J . G. 1974 Dynamics of a brachiating siamang Hylohates iSymphalanyLs/ syndactylus Nature, 248: 259- 260.
Locomotion and posture or the Malayan siamang and implications for hominoid evolution. Folia Primatol., 26: 245-269.
Fleagle, J. G.. E. L. Simons andG. C. Conroy 1975 Ape limb bone from the Oligocene of Egypt. Science, 189: 135-137.
Flower, W. H. 1885 An Introduction to the Osteology of the Manimalia. Macmillan, London.
Grand, T. I. 1972 A mechanical interpretation of termi- nal branch feeding. J . Mamm., 53: 198-201.
Grant. J . C. B. 1931 Interarticular synovial folds. Brit. J. Surgery, 18: 636-640.
Harrison, J. L. 1949 Some mammals of the trees. Malayan Nat. J . , 4: 66-69.
Harrower, G. 1933 Nycticehus malaianus Anderson: a dissection of the abdomino-pelvic viscera. Ceylon J. Sci- ence (BJ, 28: 73-88.
Hildehrand, M. 1967 Symmetrical gaits of primates. Am. <J. Phys. Anthrop., 26: 119-130.
Hill, W. C. 0. 1953 Primates: comparative Anatomy and Taxonomy. I. Strepsirhini. Edinburgh Univ. Press, Edin- burgh.
1959 The anatomy of Callimico goetdii (Tho- mas], a primitive American primate. Trans. Amer. Philos. Soc., 49 (5): 1-116.
1960 Primates: Comparative Anatomy and Tax- onomy. IV. Cebidae, Part A. Edinburgh Univ. Press, Edinburgh.
1962 Primates: Comparatwe Anatomy and Tax- onomy. V. Cehidae, Part 8. Edinburgh Univ. Press, Edin- burgh.
Primates: Comparative Anatomy and Tax- onomy. VI. Catarrhini : Cercopithecoidea; Cercopith- ecinae. Interscience, New York.
Hughes, H. 1944 The evolution and functional sig- nificance of the transverse carpal arch of man. J. Anat. (London). 78: 167-172.
Jenkins, F. A,, Jr. 1970 Anatomy and function of ex- panded ribs in certain edentates and primates. J. Mamm., 51: 288-301.
Jenkins, F. A,, Jr.. and J. G. Fleagle 1975 Knuckle-walk- ing and the functional anatomy of the wrists in living apes. In: Primate Functional Morphology and Evolution. R. H. Tuttle, ed. Mouton, The Hague, pp. 213-227.
Jewel], P. A,, and J. F. Oates 1969 Ecological observations on the lorisoid primates of African lowland coast. Zool. Africana, 4: 231-248.
1976
1966
Jones, F. W. 1916 Arboreal Man. E. Arnold, London. 1942 The Principles of Anatomy as Seen in the
Hand. Williams and Wilkins, Baltimore. Jouffroy, F. K. 1975 Osteology and myologj of the
lernuriform postcranial skeleton. In: Lemur Biology. I. Tattersall and R. W. Sussmann, eds. Plenum, New York, pp. 149-192.
Jungers, W. L. 1977 Subfossil Malagasy lemurs as non-

LORIS WRIST AND HI
primate analogues. Proc. 6th Congr. Internatl. Primat. SOC., in press.
Juraniec, J., and H. Szostakiewicz-Sawicka 1968 The cen- t ra l tendon of the diaphragm in primates. Folia Morphol., 27: 183-194.
Keith, A. 1923 Man’s posture: its evolution and disor- ders. Brit. Med. J., I : 451-454,499-502, 545-548, 587-590, 624-626, 669-672.
Le Gros Clark, W. E., and D. P. Thomas 1951 Associated jaws and limb bones of Lirnnopithecus rnacinnesi. British Museum (Nat. Hist.), London, Fossil Mammals of Africa, 3: 1-27.
Lewis, 0. J. 1965 Evolutionary change in the primate wrist and inferior radio-ulnar joints. Anat. Rec.. 151: 275-286.
1969 The hominoid wrist joint. Am. J. Phys. An- throp., 30: 251-268.
1970 The development of the human wrist joint during the fetal period. Anat. Rec., 166: 499-516.
1971a The contrasting morphology found in the wrist joints of semi-brachiating monkeys and brachiat- ing apes. Folia Primatol., 16: 248-256.
Brachiation and the early evolution of the Hominoidea. Nature, 230: 577-578.
Evolution of the hominoid wrist. In: The Functional and Evolutionary Biology of Primates. R. H. Tuttle, ed. Aldine-Atherton, Chicago, pp. 207-222.
1972b Osteological features characterizing the wrists of monkeys and apes, with a reconsideration of this region in Dryopithecus iProconsu0 africanus. Am. J. Phys. Anthrop., 36: 45-58.
1973 The hominoid os capitatum, with special reference to the fossil bones from Sterkfontein and Olduvai Gorge. J. Hum. Evol., 2: 1-11.
The wrist articulations of the Anthropoi- dea. In: Primate Locomotion. F. A. Jenkins, Jr., ed. Academic Press. New York, pp. 143-169.
Lewis. 0. J., R. J. Hamshere and T. M. Bucknill 1970 The anatomy of the wrist joint. J. Anat. (London), 106: 539- 552.
Loveridge. A. 1956 Forest Safari. Lutterworth Press, London.
Mendel, F. 1976 Postural and locomotor behavior of Alouatta pallinta on various substrates. Folia primatol., 26: 36-53.
Midlo, C. Form of hand and foot in primates. Am. J. Phys. Anthrop., 19: 337-389.
Miller, R. A. 1943 Functional and morphological adapta- tions in the forelimbs of the Slow Lemurs. Am. J. Anat., 73: 153-183.
Mittermeier, R. A., and J. G. Fleagle 1976 The locomotor and postural repertoires of Ateles geofroyi and Colobus guereza, and a reevaluation of the locomotor category semibrachiation. Am. J. Phys. Anthrop., 45: 235-256.
Mivart, St. G. J . 1865 Contributions toward a more com- plete knowledge of the axial skeleton in the Primates. Proc. 2001. Soc. London, 1865: 545-592.
On the appendicular skeleton of the pri- mates. Philos. Trans. Royal Soc. London, 157: 299-429.
Morbeck, M. E. 1972 A Re-examination of the Forelimb of the Miocene Hominoidea. Ph.D. Thesis, Univ. Calif. Berkeley.
1975 Dryopithecus africanus forelimb. J. Hum. Evol.. 4: 39-46.
1976 Problems in reconstruction of fossil anat- omy and locomotor behaviour: the Dryopithecus elbow complex. J. Hum. Evol., 5: 223-233.
Napier, J. R. 1967 Evolutionary aspects of primate locomotion. Am. J. Phys. Anthrop., 27: 333-342.
1971b
1972a
1974
1934
1867
DMINOID EVOLUTION 27 1
Napier, J . R., and P. R. Davis 1959 The fore-limb skeleton and associated remains of Proconsul africanus. British Mus. INat. Hist.), London, Fossil Mammals of Africa, 16: 7-69.
Napier, J. R., and P. H. Napier 1967 A Handbook of Living Primates. Academic Press, London.
Nayak, U. V. 1933 The articulations of the carpus in Chiromys rnudugu.scarensis, with reference to certain other lemurs. J. Anat. (London), 68: 109.115.
O’Connor, B. L. 1975 The functional morphology of the cercopithecoid wrist and inferior radioulnar joints, and their bearing on some problems in the evolution of the Hominoidea. Am. J. Phys. Anthrop., 43: 113.122.
Oxnard, C. E. 1967 The functional morphology of the primate shoulder as revealed by comparative anatomical, osteometric, and discriminant function techniques. Am. J. Phys. Anthrop., 26: 219-240.
Parsons, F. G. 1899 The ,joints of mammals compared with those of man. J. Anat. Physiol., 34: 41-68.
Pilbeam, D. R. 1969 Tertiary Pongidae of East Africa: evolutionary relationships and taxonomy. Bull. Peabody Mus. Nat. Hist. (Yale), 31: 1~185.
Pilbeam, D. R., and E. L. Simons 1971 Humerus of Dryopilhtrcus from Saint Gaudens, France. Nature, 229: 406-407.
Preuschoft. H. 1973 Body posture and locomotion in some East African Miocene Dryopithecinae. In: Human Evolution. M. H. Day, ed. Taylor and Francis, London, pp. 13-46.
1974 Body posture and mode of locomotion in fossil primates - method and example: Aegyptupzthecus zeuxis. Symp. 5th Cong. Int’l. Primatol. Soc., pp. 345-359.
Richard, A. 1970 A comparative study of the activity patterns and behavior of Alouatta uillosa and Ateles geofruyi. Folia Primatol., 12: 241-263.
Roberts, D. 1974 Structure and function of the primate scapula. In: Primate Locomotion. F. A. Jenkins, Jr., ed. Academic Press, New York, pp. 171-200.
Roberts, D., and I. Davidson 1975 The lemur scapula. In: Lemur Biology. I. Tattersall and R. W. Sussman, eds. Plenum, New York, pp. 125-147.
Robertson, D. F. 1944 Anatomy of the South American woolly monkey (Lugothriri. Part 1. The forelimb. Zoologi- ca, 29: 169.192.
Sarich, V. M., and A. C. Wilson 1967 Immunological time scale for hominid evolution. Science, 158: 1200-1203.
Schon, M. A,, and L. K. Ziemer 1973 Wrist mechanism and locomotor behavior of Dryopithecus Proconsuli africanus. Folia Primatol., 20: 1-11,
Schultz, A. H. 1961 Vertebral column and thorax. Pri- matologia, 4 (Lfg. 5): 1-66.
Schwarz, W. 1938 Das 0 s pisiforme. Geg. Morph. Jb., 81: 187-212.
Simons, E. L. 1960 Apzdzum and Oreopithecus. Nature, 186: 824-826.
1972 Primate Evolution: An Introduction to Man’s Place in Nature. Macmillan, New York.
Simons, E. L., and J. G. Fleagle 1973 The history of extinct gibbon-like primates. Gibbon and Siamang, 2: 121-148.
Simons, E. L., and D. R. Pilbeam 1965 Preliminary revision of the Dryopithecinae (Pongidae. Anthropoidea). Folia Primatol., 3: 81.162.
Functional myology of the hip and thigh of cebid monkeys and its implications for the evolution of erect posture. Biblioth. Primatol., 14: 1-318.
1976 Before bipedality. Yearbk. Phys. Anthrop., 19: 59-68.
Straus, W. L., Jr. 1963 The classification of Oreopith-
Stern, J. T., Jr. 1971

272 MATT CARTMILI, AND KATHARINE MILTON
ecus. In: Classification and Human Evolution. S. L. Washburn, ed. Aldine, Chicago, pp. 146~177.
Straus, W. L.. Jr., and G . B. Wislocki 1932 On certain simi- larities between sloths and slow lemurs. Bull. Mus. Comp. Zwl. (Harvard), 74: 45-56.
Subramoniam, S. 1957 Some observations on the habits of the slender loris, Lorls tardigradus L. J. Bombay Nat. Hist. Soc.. 54: 387-398.
Szalay, F. S. 1975 Haplorhine phylogeny and the status of the Anthropnidea. In: Primate Functional Morphology and Evolution. R. H. Tuttle. ed. Mouton, The Hague, pp. 3-22.
Tattersall. I . 1974 Facial structure and mandibular me- chanics in Archaeolemur. In: Prosimian Biology. R. D. Martin, G. A. Doyle and A. C. Walker, eds. Duckworth, London, pp. 563-577.
The biology of rodent tails: a study of form and function. Technical report AAL-TR- 65-8, Arctic Aeromedical Laboratory, Fort Wainwright, Alaska.
Tuttle, R. H. 1967 Knuckle-walking and the evolution of hominoid hands. Am. J. Phys. Anthrop., 26: 171-206.
1969 Quantitative and functional studies on the hands of the Anthropoidea. I. The Hominoidea. J. Morph., 228: 309-364.
1970 Postural, propulsive and prehensile capa- bilities in the cheiridia of chimpanzees and other great apes. In: The Chimpanzee. Vol. 2. G. H. Bourne, ed. Uni- versity Park Press, Baltimore. pp. 167-253.
1975a Knuckle-walking and knuckle-walkers: a commentary on some recent perspectives on hominoid
Thorington, R. W.. Jr . 1966
evolution. In: Primate Functional Morphology and Evolution. H. H. Tuttle, ed. Mouton. The Hague, pp. 203- 212.
1975b Parallelism, brachiation, and hominoid phylogeny. In: Phylogeny of the Primates. W. P. Luckett and F. S. Szalay. eds. Plenum, New York, pp. 447-480.
Tuttle, R. H., and J. V. Basmnjian 1974a Electromyogra- phy of forearm musculature in gorilla and problems re- lated to knuckle-walking. In: Primate Locomotion. F. A. Jenkins, Jr., ed. Academic Press, New York, pp. 293-347.
1974b Electromyography of brachial muscles in Pan gorilla and hominoid evolution. Am. J. Phys. An- throp., 41: 71-90.
Walker, A. 1969 The locomotion of the lurises, with special reference to the potto. E. Afr. Wildlife J., 7: 1-5.
Locomotor adaptations in past and present prosimian primates. In: Primate Locomotion. F. A. Jenkins, Jr.. ed. Academic Press, New York, pp. 349-381.
Wilson, L). R. 1972 Tail reduction In Macaca. In: The Functional and Evolutionary Biology of Primates. R. H. Tuttle, ed. Aldine-Atherton. Chicago, pp. 241-261.
The form and function of the carpal bones in some arboreally adapted mammals. Acta Anat., 82: 383-406.
Zapfe, H. 1958 The skeleton of Pliopithecus @iplio- pilh~cusJ uindobonensis Zapfe and Hbrzeler. Am. J. Phys. Anthrop., 16: 441-458.
1960 Die Prirnatenfunde aus der miozanen Spaltenfullung von Neudorf an der March (Devinska Nova Ves), Tschechoslowakei. Schweiz. Palaontol. Abh., 78: 1~269.
1974
Yalden, D. W. 1972




















