
Functional characterization of gefitinib uptake in non-small cell lung cancer cell lines
Upload
independentCategory
view
3download
0
RESEARCH ARTICLE
The improvement of doxorubicin activity on breast cancercell lines by tangeretin through cell cycle modulation
Edy Meiyanto & Aditya Fitriasari & Adam Hermawan &
Sendy Junedi & Ratna Asmah Susidarti
Received: 24 May 2011 /Accepted: 10 June 2011 /Published online: 5 July 2011# Institute of Oriental Medicine, Kyung Hee University 2011
Abstract Tangeretin, shows cytotoxic effect on COLO 205colon cancer cells. Combination of tangeretin with tamox-ifen showed synergistic effect and increased the cancer cellsensitivity towards tamoxifen on T47D cells. However, thecombination of tangeretin with chemotherapeutic agentdoxorubicin on breast cancer cells have not been exploredyet. Therefore, the aim of this research is to examine theimprovement of cytotoxic effect of doxorubicin by tanger-etin through cell death induction and cell cycle modulationon MCF-7 and T47D cells. The cytotoxic effect oftangeretin, doxorubicin, and their combination on testedcells were carried out by using MTT assay. Cell cycledistribution was determined by flowcytometer FACS-Calibur and the flowcytometry data was analyzed usingModFit LT 3.0 program. Cell death assay were done bydouble staining method using ethydium bromide-acridinorange. Single treatment of tangeretin 5–100 μM did notshow cytotoxic effect on MCF-7 and T47D cells. Thecombination of tangeretin 50 and 100 μM with doxorubicin200 nM (MCF-7) and 7.5 nM (T47D) increased thecytotoxic effect of doxorubicin on both breast cancer celllines. This improvement of cytotoxic effect is due to celldeath induction and cell cycle modulation. Furthermore,single treatment of tangeretin showed cell death only onT47D cell and caused G1-phase arrest on MCF-7 cell and
G2/M-phase arrest on T47D cell. While doxorubicininduced cell accumulation at G2/M phase in both cancercell lines. However, combination of tangeretin and doxoru-bicin increased cell death on both cancer cell lines,compared with doxorubicin by itself. The combination alsoshowed G1-phase arrest on MCF-7 cell and increased cellaccumulation at G2/M phase on T47D cell. Based on thisresult, tangeretin is potential to be developed as co-chemotherapeutic agent for breast cancer by inducingapoptosis and cell cycle arrest. However, the molecularmechanism need to be explored further.
Keywords Tangeretin . Co-chemotherapy . Breast cancer .
Cell cycle arrest . Cell death
Introduction
Breast cancer is one of the death-cause cancer in the world(Jemal et al. 2010). Breast cancer is caused by theuncontrolled proliferation of breast cells which is happenedbecause of their ability to avoid apoptotic mechanism (Yu etal. 2010). One drug for breast cancer therapy is doxorubicin.The using of doxorubicin has a lot of side effects and alsoresistance effects on breast cancer cell (Smith et al. 2006).Therefore, combination of doxorubicin with chemopreven-tive agent (co-chemotherapy) were needed to increase theactivity of doxorubicin by inhibiting proliferation andinducing apoptosis of breast cancer cell.
One of the chemopreventive agents that has beenreported to have antiproliferative effect is tangeretin, apolymethoxy flavone found in citrus species (Citrusaurantifolia). Tangeretin have been proven to inhibit thegrowth of estradiol-stimulated T47D cells (Van Slambroucket al. 2005); inhibit proliferation of MCF-7 cells after
E. Meiyanto (*) :A. Fitriasari :A. Hermawan : S. Junedi :R. A. SusidartiCancer Chemoprevention Research Center, Faculty of Pharmacy,Universitas Gadjah Mada,Sekip Utara,Yogyakarta 55281, Indonesiae-mail: [email protected]
E. Meiyantoe-mail: [email protected]: http://www.ccrc.farmasi.ugm.ac.id
Orient Pharm Exp Med (2011) 11:183–190DOI 10.1007/s13596-011-0016-4

4 days treatment and cause G1-phase arrest on MCF-7 cellsby 24 h, 48 h and 72 h treatment (Morley et al. 2007).Tangeretin has the smallest IC50 value on COLO 205 coloncancer cells, compared with the other flavonoid com-pounds, such as apigenin, kaempferol, myricetin, quercetin,luteolin, nobiletin, and rutin (Pan et al. 2002). Tangeretinalso induce apoptosis on HL-60 leukemia cells (Hirano etal. 1995). The combination of tangeretin and tamoxifen invitro increased the sensitivity of cancer cell synergisticallytowards tamoxifen (Bracke et al. 1999).
Those researches showed the potency of tangeretin aschemopreventive agent and became a basic for thedevelopment of tangeretin as co-chemotherapeutic agentto increase the cytotoxic activity and reduce the side effectsof doxorubicin. Therefore, the purpose of this research is toexamine the effect of tangeretin and its combination withdoxorubicin on cell cycle and apoptosis of breast cancercells.
Materials and methods
Materials
Tangeretin was obtained from Sigma Aldrich ChemieGmBH, Steinheim, Germany (Cat No. T8951) whiledoxorubicin was obtained from P.T. Ferron Par Pharmaceu-tical. A DMSO (Sigma Aldrich Chemie GmBH, Steinheim,Germany) solution of tangeretin was used for in vitroexperiment by diluting desired consentration. The finalDMSO concentration was not more than 1%.
Cell lines
MCF-7 and T47D cells were kindly provided by Prof.Masashi Kawaichi (Nara Institute of Science and Technol-ogy (NAIST)), Nara, Japan.
Cytotoxic assay (MTT assay)
MCF-7 and T47D cells were cultured in Dulbecco’sModified Eagle’s Medium (DMEM) containing Fetal BovineSerum (FBS) 10% (v/v) (FBS qualified, Gibco, Invitro-gen™ USA) and penisillin-streptomisin 1% (v/v) (Gibco,Invitrogen Corporation, Grand Island, NY, 14072, USA).Cells (5×103 cells/well) were transferred to 96-well plate(Iwaki, Japan) and incubated for 24 h (70–80% confluent).Cells were treated by tangeretin, doxorubicin, and theircombination, and incubated for 24 h. At the end of theincubation, 5 mg/ml solution of MTT [3-(4,5-dimethylth-iazole-2-yl)-2,5-diphenyl tetrazolium bromide] (Sigma,Sigma-Aldrich Corp, St. Louis, MO, USA) were added toeach wells and the cells were incubated for 4 h in 37°C.
Viable cells reacts with MTT to form purple formazancrystal. After 4 h, stopper sodium dodesil sulphate 10% in0,1N sulphuric acid solution were added to dissolveformazan crystal. Cells were incubated over night andprotected from light. Cells were shaken for 10 min beforeread by ELISA reader at λ 595 nm. The absorbance of eachwell converted to percentage of viable cells:
%Viable cells ¼ Treated cells abs�Medium control abs
Cells control abs�Medium control abs� 100%
Cell death assay (double staining method)
Cells (5×104 cells/well) were transferred to coverslips(Nunc, Denmark) in 24-well plate (Iwaki, Japan) andincubated for 24 h (50–60% confluent). Cells were treatedby tangeretin, doxorubicin, and their combination, andincubated for 15 h. At the end of the incubation, coverslipscontaining cells were moved to object glass. Mixturesolution of etidium bromide-acridine orange (Sigma,Sigma-Aldrich Corp, St. Louis, MO, USA) were added tothe cells to form fluorescence cells. The fluorescence cellswere examined immediately by fluorescence microscope(Zeiss MC80). Green fluorescence cells showed viablecells, while red fluorescence cells showed dead cells.
Flowcytometry assay
Cells (5×105 cells/well) were transferred into 6-well plate(Iwaki, Japan) and incubated until the cells return to normalcondition. Cells were treated by tangeretin, doxorubicin,and their combination, and incubated for 24 h. At the end ofthe incubation, the media containing free cells suspensionwere taken and transferred into 1.5 ml eppendorfs. Theeppendorfs were centrifugated (2,000 rpm, 3 min) and thesupernatant were removed. The cells in 6-well plate wereadded by phospate buffered saline (PBS), and the PBS weretransferred into previous eppendorfs. The eppendorfs werecentrifugated and the supernatant were removed again. Thissteps were repeated before the cells harvested by trypsin-EDTA 0.25% (Gibco, Invitrogen, Canada). Harvested cellswere transferred into the eppendorfs and centrifugated(2,000 rpm, 30 s). The remaining cells in the 6-well platewere rinsed with PBS and transferred into the eppendorfs.The eppendorfs were centrifugated and the supernatantwere removed. Pellet cells in eppendorfs washed by coldPBS and added by propidium iodide (PI) (Sigma AldrichChemie GmBH, Steinheim, Germany). The eppendorfswere wrapped in aluminum foil and incubated in 37°C for10 min. After 10 min, cell suspension were homogenatedand transferred into the flowcyto-tube to be analyzed byflowcytometer FACS using ModFit LT 3.0 program.
184 E. Meiyanto et al.
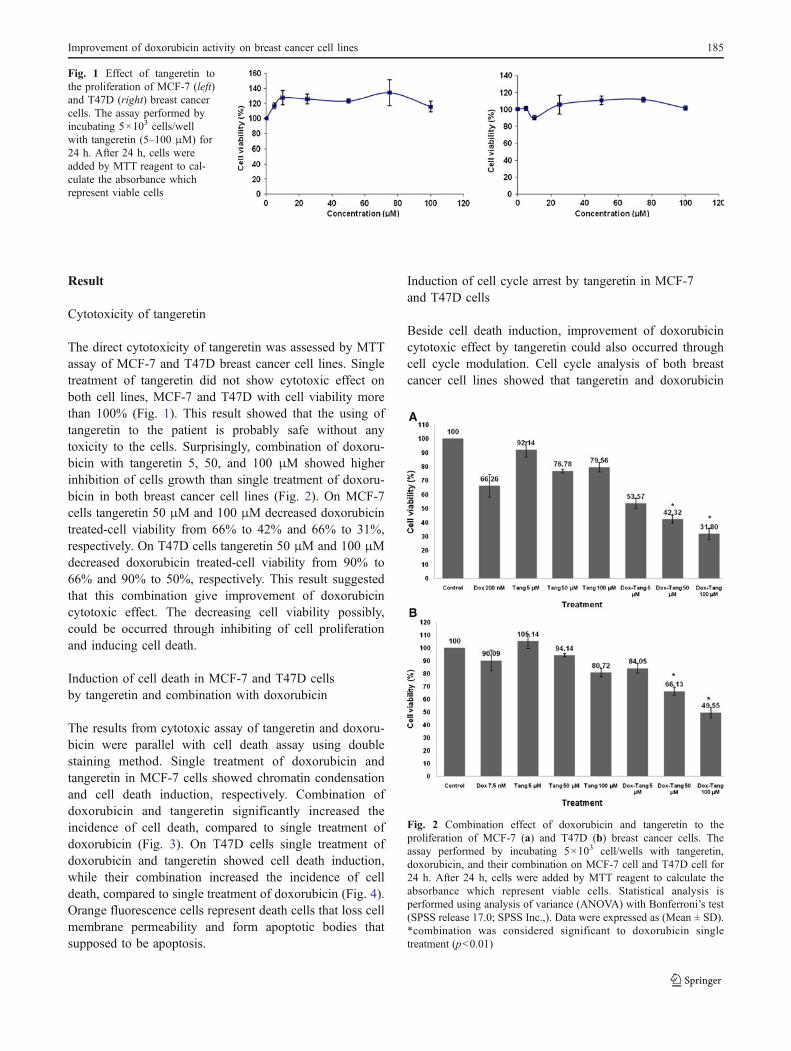
Result
Cytotoxicity of tangeretin
The direct cytotoxicity of tangeretin was assessed by MTTassay of MCF-7 and T47D breast cancer cell lines. Singletreatment of tangeretin did not show cytotoxic effect onboth cell lines, MCF-7 and T47D with cell viability morethan 100% (Fig. 1). This result showed that the using oftangeretin to the patient is probably safe without anytoxicity to the cells. Surprisingly, combination of doxoru-bicin with tangeretin 5, 50, and 100 μM showed higherinhibition of cells growth than single treatment of doxoru-bicin in both breast cancer cell lines (Fig. 2). On MCF-7cells tangeretin 50 μM and 100 μM decreased doxorubicintreated-cell viability from 66% to 42% and 66% to 31%,respectively. On T47D cells tangeretin 50 μM and 100 μMdecreased doxorubicin treated-cell viability from 90% to66% and 90% to 50%, respectively. This result suggestedthat this combination give improvement of doxorubicincytotoxic effect. The decreasing cell viability possibly,could be occurred through inhibiting of cell proliferationand inducing cell death.
Induction of cell death in MCF-7 and T47D cellsby tangeretin and combination with doxorubicin
The results from cytotoxic assay of tangeretin and doxoru-bicin were parallel with cell death assay using doublestaining method. Single treatment of doxorubicin andtangeretin in MCF-7 cells showed chromatin condensationand cell death induction, respectively. Combination ofdoxorubicin and tangeretin significantly increased theincidence of cell death, compared to single treatment ofdoxorubicin (Fig. 3). On T47D cells single treatment ofdoxorubicin and tangeretin showed cell death induction,while their combination increased the incidence of celldeath, compared to single treatment of doxorubicin (Fig. 4).Orange fluorescence cells represent death cells that loss cellmembrane permeability and form apoptotic bodies thatsupposed to be apoptosis.
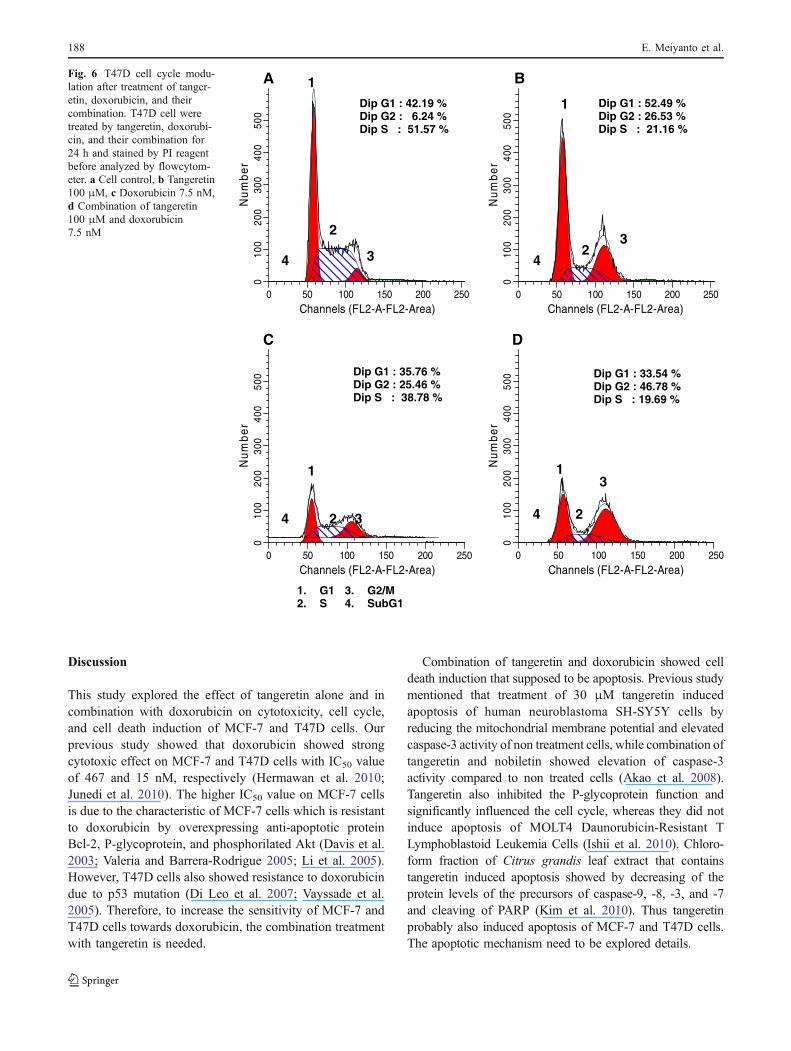
Induction of cell cycle arrest by tangeretin in MCF-7and T47D cells
Beside cell death induction, improvement of doxorubicincytotoxic effect by tangeretin could also occurred throughcell cycle modulation. Cell cycle analysis of both breastcancer cell lines showed that tangeretin and doxorubicin
Fig. 1 Effect of tangeretin tothe proliferation of MCF-7 (left)and T47D (right) breast cancercells. The assay performed byincubating 5×103 cells/wellwith tangeretin (5–100 μM) for24 h. After 24 h, cells wereadded by MTT reagent to cal-culate the absorbance whichrepresent viable cells
Fig. 2 Combination effect of doxorubicin and tangeretin to theproliferation of MCF-7 (a) and T47D (b) breast cancer cells. Theassay performed by incubating 5×103 cell/wells with tangeretin,doxorubicin, and their combination on MCF-7 cell and T47D cell for24 h. After 24 h, cells were added by MTT reagent to calculate theabsorbance which represent viable cells. Statistical analysis isperformed using analysis of variance (ANOVA) with Bonferroni’s test(SPSS release 17.0; SPSS Inc.,). Data were expressed as (Mean ± SD).*combination was considered significant to doxorubicin singletreatment (p<0.01)
Improvement of doxorubicin activity on breast cancer cell lines 185

C D
A BFig. 3 Effect of tangeretin,doxorubicin, and their combina-tion on MCF-7 cell death.MCF-7 cell were treated bytangeretin, doxorubicin, andtheir combination for 15 h andstained by etidium bromide-acridine orange. a Cell control,b Tangeretin 100 μM, c Doxo-rubicin 200 nM, d Combinationof tangeretin 100 μM anddoxorubicin 200 nM. Viablecells give green fluorescence( ), dead cells give orangefluorescence ( )
A B
C D
Fig. 4 Effect of tangeretin,doxorubicin, and their combina-tion on T47D cell death. T47Dcell were treated by tangeretin,doxorubicin, and their combina-tion for 15 h and stained byetidium bromide-acridineorange. a Cell control, b Tan-geretin 100 μM, c Doxorubicin7.5 nM, d Combination of tan-geretin 100 μM and doxorubicin7.5 nM. Viable cells give greenfluorescence ( ), dead cellsgive orange fluorescence ( )
186 E. Meiyanto et al.

have different cell cycle profile. Single treatment oftangeretin induced cell accumulation at G1 phase onMCF-7 cells and showed cell accumulation at G2/M phase(G2/M-phase arrest) on T47D cells. While, doxorubicinarrested both cell lines at G2/M-phase. However, combina-
tion of tangeretin and doxorubicin showed G1-phase arreston MCF-7 cells and increased cell accumulation at G2/Mphase on T47D cells (Table 1; Figs. 5 and 6). These resultssuggested the domination of tangeretin in the improvementof doxorubicin cytotoxic effect.
Cell Treatment Concentration G1 (%) S (%) G2/M (%)
MCF-7 Control – 69.61 22.75 7.64
Tangeretin 100 μM 78.46 13.95 7.59
Doxorubicin 200 nM 67.75 2.84 29.41
Tangeretin-Doxorubicin 100 μM–200 nM 83.42 0.00 16.58
T47D Control – 42.19 51.57 6.24
Tangeretin 100 μM 52.49 21.16 26.35
Doxorubicin 7.5 nM 35.76 38.78 25.46
Tangeretin-Doxorubicin 100 μM–7.5 nM 33.54 19.69 46.78
Table 1 Cell cycle distributionafter treatment of tangeretin,doxorubicin, and their combina-tion for 24 h
Channels (FL2-A-FL2-Area)0 50 100 150 200 250
Num
ber
010
020
030
040
050
060
01
4 2 3
A
Dip G1 : 69.61 % Dip G2 : 7.64 % Dip S : 22.75 %
Channels (FL2-A-FL2-Area)0 50 100 150 200 250
Num
ber
010
020
030
040
050
060
0
B
Dip G1 : 78.46 % Dip G2 : 7.59 % Dip S : 13.95 %
1
4
C
Channels (FL2-A-FL2-Area)0 50 100 150 200 250
Num
ber
010
020
030
040
050
060
0
1
3 4 2
Dip G1 : 67.75 % Dip G2 : 29.41 % Dip S : 2.84 %
1. G12. S
3. G2/M4. SubG1
Channels (FL2-A-FL2-Area)0 50 100 150 200 250
Num
ber
010
020
030
040
050
060
0 Dip G1 : 83.42 % Dip G2 : 16.58 % Dip S : 0.00 %
D
1
2 3
2 3 4
Fig. 5 MCF-7 cell cyclemodulation after treatment oftangeretin, doxorubicin, andtheir combination. MCF-7 cellwere treated by tan3geretin,doxorubicin, and theircombination for 24 h andstained by PI reagent beforeanalyzed by flowcytometer.a Cell control, b Tangeretin100 μM, c Doxorubicin200 nM, d Combination oftangeretin 100 μM anddoxorubicin 200 nM
Improvement of doxorubicin activity on breast cancer cell lines 187
Discussion
This study explored the effect of tangeretin alone and incombination with doxorubicin on cytotoxicity, cell cycle,and cell death induction of MCF-7 and T47D cells. Ourprevious study showed that doxorubicin showed strongcytotoxic effect on MCF-7 and T47D cells with IC50 valueof 467 and 15 nM, respectively (Hermawan et al. 2010;Junedi et al. 2010). The higher IC50 value on MCF-7 cellsis due to the characteristic of MCF-7 cells which is resistantto doxorubicin by overexpressing anti-apoptotic proteinBcl-2, P-glycoprotein, and phosphorilated Akt (Davis et al.2003; Valeria and Barrera-Rodrigue 2005; Li et al. 2005).However, T47D cells also showed resistance to doxorubicindue to p53 mutation (Di Leo et al. 2007; Vayssade et al.2005). Therefore, to increase the sensitivity of MCF-7 andT47D cells towards doxorubicin, the combination treatmentwith tangeretin is needed.
Combination of tangeretin and doxorubicin showed celldeath induction that supposed to be apoptosis. Previous studymentioned that treatment of 30 μM tangeretin inducedapoptosis of human neuroblastoma SH-SY5Y cells byreducing the mitochondrial membrane potential and elevatedcaspase-3 activity of non treatment cells, while combination oftangeretin and nobiletin showed elevation of caspase-3activity compared to non treated cells (Akao et al. 2008).Tangeretin also inhibited the P-glycoprotein function andsignificantly influenced the cell cycle, whereas they did notinduce apoptosis of MOLT4 Daunorubicin-Resistant TLymphoblastoid Leukemia Cells (Ishii et al. 2010). Chloro-form fraction of Citrus grandis leaf extract that containstangeretin induced apoptosis showed by decreasing of theprotein levels of the precursors of caspase-9, -8, -3, and -7and cleaving of PARP (Kim et al. 2010). Thus tangeretinprobably also induced apoptosis of MCF-7 and T47D cells.The apoptotic mechanism need to be explored details.
A
Dip G1 : 42.19 % Dip G2 : 6.24 % Dip S : 51.57 %
1
4
2
3
Channels (FL2-A-FL2-Area)0 50 100 150 200 250
Num
ber
010
020
030
040
050
0
B
1
4
3 2
Dip G1 : 52.49 % Dip G2 : 26.53 % Dip S : 21.16 %
Channels (FL2-A-FL2-Area)0 50 100 150 200 250
Num
ber
010
020
030
040
050
0
Dip G1 : 35.76 % Dip G2 : 25.46 % Dip S : 38.78 %
C
1
4 2 3
Channels (FL2-A-FL2-Area)0 50 100 150 200 250
Num
ber
010
020
030
040
050
0
Channels (FL2-A-FL2-Area)0 50 100 150 200 250
Num
ber
010
020
030
040
050
0
1 3
2 4
Dip G1 : 33.54 % Dip G2 : 46.78 % Dip S : 19.69 %
D
1. G12. S
3. G2/M4. SubG1
Fig. 6 T47D cell cycle modu-lation after treatment of tanger-etin, doxorubicin, and theircombination. T47D cell weretreated by tangeretin, doxorubi-cin, and their combination for24 h and stained by PI reagentbefore analyzed by flowcytom-eter. a Cell control, b Tangeretin100 μM, c Doxorubicin 7.5 nM,d Combination of tangeretin100 μM and doxorubicin7.5 nM
188 E. Meiyanto et al.

There are two main mechanism of apoptosis, intrinsicand extrinsic pathway, through which apoptosis could beoccurred in MCF-7 and T47D cells. These apoptoticmechanisms involve the expression of regulatory protein,such as p53, Bcl-2 family, and activation of caspasefamily (Kumar et al. 2000). MCF-7 cells express p53 andBcl-2. Doxorubicin induces apoptosis through intrinsicmechanism by elevating p53 expression which is neededto induce the expression of pro-apoptosis protein (Bax)(Minotti et al. 2004). In addition, tangeretin increases theexpression of p53 protein in colon cancer cell (Pan et al.2002). Therefore, the increasing of cell death aftercombination treatment of tangeretin and doxorubicin onMCF-7 cells probably by apoptosis in p53-dependentpathway. The proposed mechanism is need to be exploredfurther.
While, T47D cells do not express p53. Therefore,apoptotic mechanism of T47D cells is occurred in p53-independent pathway. Van Slambrouck et al. (2005) showedthat tangeretin inhibit ERK phosphorilation pathway whichis involved in T47D cell proliferation. Perhaps, theincreasing of apoptotic cell after combination treatment oftangeretin and doxorubicin on T47D cells also occurredthrough this mechanism. But the exact mechanism need tobe explored more detail.
The different effect on MCF-7 and T47D cell cycle areoccurred due to different mechanism action of tangeretin inboth cells. Cell cycle arrest by tangeretin involves inhibi-tion of CDK2/CDK4 activity and induction of CDKinhibitor expression, such as p27 and p21, which areproved by Pan et al. (2002) and Morley et al. (2007) oncolorectal cancer (COLO 205), breast cancer (MDA-MB-435 dan MCF-7), and colon cancer (HT-29). Doxorubicininhibits the activity of cdc2 which is involved in G2-Mphase transition (Ling et al. 1996). But the exact mecha-nism need to be explored further.
The significant increasing of cytotoxic effect aftercombination treatment of tangeretin and doxorubicin couldalso occurred through inhibition of multidrugs resistance(MDR) protein. Tanaka et al. (1997) reported that tangeretinincreases vinblastine uptake by inhibiting P-gp efflux pumpin Caco-2 cells. Those three mechanisms, MDR proteininhibition, apoptotic induction, cell cycle modulation arestrengthened each other to form stronger cytotoxic effect onbreast cancer cells.
This result showed the potency of tangeretin to bedeveloped as co-chemotherapeutic agent for doxorubicin byinducing cell death and cell cycle arrest. The use ofdoxorubicin together with tangeretin is expected to increasethe activity and reduce the side effects of doxorubicin.However, the molecular mechanism of cell death inductionand cell cycle arrest by this combination need to beexplored further.
Conclusion
This research shows that combination of tangeretin anddoxorubicin synergically increases the cytotoxic effect ofdoxorubicin through cell death induction and cell cyclearrest. Based on this result, tangeretin is potential to bedeveloped as co-chemotherapeutic agent for doxorubicin.
Acknowledgement This work was supported by Program DIPAUniversitas Gadjah Mada through Research Program Tim HibahPascasarjana Multitahun year 2009 and 2010.
References
Akao Y, Itoh T, Ohguchi K, Iinuma M, Nozawa Y (2008) Interactiveeffects of polymethoxy flavones from citrus on cell growthinhibition in human neuroblastoma SH-SY5Y cells. Bioorg MedChem 16:2803–2810
Bracke ME, Depypere HT, Boterberg T, Van Marck VL, VennekensKM, Vanluchene E, Nuytinck M, Serreyn R, Mareel MM (1999)Influence of tangeretin on tamoxifen’s therapeutic benefit inmammary cancer. J Natl Cancer Inst 91(4):354–359
Davis JM, Navolanic PM, Weinstein-Oppenheimer CR, Steelman LS,Wei H, KonoplevaM, BlagosklonnyMV,McCubrey JA (2003) Raf-1and Bcl-2 induce distinct and common pathways that contribute tobreast cancer drug resistance. Clin Cancer Res 9:1161–1170
Di Leo A, Tanner M, Desmed C, Paesman M, Cardoso F, Durbecq V,Chan S, Parren T, Aapro M, Sotiriou C, Piccart MJ, Larsimont D,Isola J (2007) p-53 gene mutations as a predictive marker in apopulation of advanced breast cancer patients randomly treatedwith doxorubicin or docetaxel in the context of a phase IIIclinical trial. Ann Oncol 18:997–1003
Hermawan A, Meiyanto E, Susidarti A (2010) Hesperidin MeningkatkanAktivitas Sitotoksik Doxorubicin pada SelMCF-7.Majalah FarmasiIndonesia 21(1):8–17
Hirano T, Abe K, Gotoh M, Oka K (1995) Citrus flavone tangeretininhibits leukaemic HL-60 cell growth partially through inductionof apoptosis with less cytotoxicity on normal lymphocytes. Br JCancer 72(6):1380–1388
Ishii K, Tanaka S, Kagami K, Henmi K, Toyoda H, Kaise T (2010)Effects of naturally occurring polymethyoxyflavonoids on cellgrowth, p-glycoprotein function, cell cycle, and apoptosis ofdaunorubicin-resistant T lymphoblastoid leukemia cells. CancerInvest 28:220–229
Jemal A, Siegel R, Xu J, Ward E (2010) Cancer statistics, 2010. CACancer J Clin. doi:10.3322/caac.20073
Junedi S, Susidarti RA, Meiyanto E (2010) Naringenin MeningkatkanEfek Sitotoksik Doxorubicin Pada Sel Kanker Payudara T47DMelalui Induksi Apoptosis. Jurnal Ilmu Kefarmasian Indonesia 8(2):85–90
Kim H, Moon JY, Mosaddik A, Cho SK (2010) Induction of apoptosisin human cervical carcinoma HeLa cells by polymethoxylatedflavone-rich Citrus grandis Osbeck (Dangyuja) leaf extract. FoodChem Toxicol 48:2435–2442
Kumar R, Vadlamudi RK, Adam L (2000) Apoptosis in mammarygland and cancer. Endocr Relat Cancer 7:257–269
Li X, Lu Y, Liang K, Liu B, Fan Z (2005) Differential responsesto doxorubicin-induced phosphorylation and activation of aktin human breast cancer cells. Breast Cancer Res 7(5):R589–R597
Ling YH, el-Naggar AK, Priebe W, Perez-Soler R (1996) Cell cycle-dependent cytotoxicity, G2/M phase arrest, and Disruption of
Improvement of doxorubicin activity on breast cancer cell lines 189

p34cdc2/Cyclin B1 activity induced by doxorubicin in synchro-nized P388 cells. Mol Pharmacol 49(5):832–841
Minotti G, Menna P, Salvatorelli E, Cairo G, Gianni L (2004)Anthracyclins: molecular advances and pharmacologic developmentsin antitumor activity and cardiotoxicity. Pharmacol Rev 56:185–228
Morley K, Ferguson P, Korotpatnick J (2007) Tangeretin and nobiletininduce G1 cell cycle arrest but not apoptosis in human breast andcolon cancer cells. Cancer Lett 251(1):168–178
Pan MH, Chen WJ, Shiau SYL, Ho CT, Lin JK (2002) Tangeretininduces cell-cycle G1 arrest through inhibiting cyclin-dependentkinases 2 and 4 activities as well as elevating Cdk inhibitors p21and p27 in human colorectal carcinoma cells. Carcinogenesis 23(10):1677–1684
Smith L, Watson MB, O’Kane SL, Drew MPJ, Lind MJ, Cawkwell L(2006) The analysis of doxorubicin resistance in human breastcancer cells using antibody microarrays. Mol Cancer Ther 5(8):2115–2120
Tanaka T, Makita H, Ohnishi M, Mori H, Satoh K, Hara A, Sumida T,Fukutani K, Tanaka T, Ogawa H (1997) Chemoprevention of 4-
nitroquinoline 1-oxide-induced oral carcinogenesis in rats byflavonoids diosmin and hesperidin, each alone and in combina-tion. Cancer Res 57:246–252
Valeria P, Barrera-Rodrigue R (2005) Changes in P-glycoproteinactivity are mediated by the growth of a tumour cell line asmulticellular spheroids. Cancer Cell Int 5(20), Summary
Van Slambrouck S, Parmar VS, Sharma SK, De Bondt B, Foré F,Coopman P, Vanhoecke BW, Boterberg T, Depypere HT, LeclercqG, Bracke ME (2005) Tangeretin inhibits extracellular-signal-regulated kinase (ERK) phosphorylation. FEBS Lett 579(7):1665–9
Vayssade M, Haddada H, Faridoni-Laurens L, Tourpin S, Valent A,Bénard J, Ahomadegbe J (2005) p73 functionally replaces p53 inadriamycin-treated, p53-deficient breast cancer cells. Int J Cancer116(6):860–869
Yu B, Shen H, Gao F, Fan Y, Sun Z (2010) Expression of theapoptosis-related genes BCL-2 and BAD in human breastcarcinoma and their associated relationship with chemosensitivity.J Exp Clin Cancer Res 29:107, pp 1–7
190 E. Meiyanto et al.
Copyright © 2022 FDOKUMEN




















