
Horse Embryonic Stem Cell Lines from the Proliferation of Inner Cell Mass Cells
9
STEM CELLS AND DEVELOPMENT 15:523–531 (2006) © Mary Ann Liebert, Inc. Original Research Report Horse Embryonic Stem Cell Lines from the Proliferation of Inner Cell Mass Cells XIHE LI, 1,4 S.G. ZHOU, 2 MARTA P. IMREH, 3 L. ÄHRLUND-RICHTER, 3 and W.R. ALLEN 1 ABSTRACT Inner cell mass (ICM) cells were isolated immunosurgically from day 7–8 horse blastocysts and, af- ter proliferation in vitro for 15–28 passages, three lines of cells were confirmed to be embryonic stem (ES) cells by their continued expression of alkaline phosphatase activity and their ability to bind antisera specific for the recognized stem cell markers, SSEA-1, TRA-1-60, TRA-1-81, and the key embryonic gene Oct-4. When maintained under feeder cell-free conditions in vitro, the three lines of cells differentiated into cells of ectodermal, endodermal, and mesodermal lineages. How- ever, they did not form teratomata when injected into the testes of severe combined immunodefi- ciency (SCID)/beige immunoincompetent mice, thereby indicating a significant difference in phe- notype between ES cells of the horse and those of the mouse and human. 523 INTRODUCTION A T THE BLASTOCYST STAGE OF DEVELOPMENT, the preim- plantation mammalian embryo separates into the in- ner cell mass (ICM) cells of the putative embryo proper and the trophectoderm cells that will eventually form the outermost layer of the fetal placenta (1). Pluripotent ES cell lines were derived originally from the ICM of mouse and human preimplantation embryos with retention of the developmental potential to transform into derivatives of all three embryonic germinal layers, ectoderm, endoderm, and mesoderm, even after prolonged culture in vitro (1–3). Subsequently, it has become apparent that the culture re- quirements for proliferation and maintenance of ES cells in an undifferentiated state, and the nature of the stimuli required to induce ES cells to transform into specialized end-stage cells of specific phenotype, vary greatly between the human and various animal species (1–10). Usefully, it appears that the pig and horse, in contrast to ruminant species, will more readily permit the establishment of gen- uine pluripotent ES cell lines following the isolation and repeated passage of ICM cells in vitro. Li et al. (11) pro- duced a porcine ES cell line in this manner and Saito et al. (12) described the successful production of two horse ES cell lines from the repeated culture of the ICM cells of day-8 frozen-thawed horse blastocysts. In the present study, we also created three lines of horse ES cells by repeated passage of ICM cells recovered im- munosurgically from day-8 blastocysts. We used antis- era generated against a well-recognized panel of human and mouse ES cell markers to study the phenotypic char- acteristics of horse ICM cells and the ES cell lines cre- ated from them. We also evaluated the transformation ca- pabilities of these ES cells when grown in vitro in the 1 Department of Veterinary Medicine, Equine Fertility Unit, Mertoun Paddocks, University of Cambridge, Newmarket, Suffolk CB8 9BH, UK. 2 The Babraham Institute, Babraham, Cambridge CB2 4AT, UK. 3 Clinical Research Centre, Huddinge University Hospital, Karolinska Institute, 141 57 Stockholm, Sweden. 4 Present address: China Agricultural University, College of Animal Science and Technology, Equine Research Centre, No 2 Yangminyang Xilu, Beijing 100049, China.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Horse Embryonic Stem Cell Lines from the Proliferation of Inner Cell Mass Cells

STEM CELLS AND DEVELOPMENT 15:523–531 (2006)© Mary Ann Liebert, Inc.
Original Research Report
Horse Embryonic Stem Cell Lines from the Proliferation of Inner Cell Mass Cells
XIHE LI,1,4 S.G. ZHOU,2 MARTA P. IMREH,3 L. ÄHRLUND-RICHTER,3 and W.R. ALLEN1
ABSTRACT
Inner cell mass (ICM) cells were isolated immunosurgically from day 7–8 horse blastocysts and, af-ter proliferation in vitro for 15–28 passages, three lines of cells were confirmed to be embryonicstem (ES) cells by their continued expression of alkaline phosphatase activity and their ability tobind antisera specific for the recognized stem cell markers, SSEA-1, TRA-1-60, TRA-1-81, and thekey embryonic gene Oct-4. When maintained under feeder cell-free conditions in vitro, the threelines of cells differentiated into cells of ectodermal, endodermal, and mesodermal lineages. How-ever, they did not form teratomata when injected into the testes of severe combined immunodefi-ciency (SCID)/beige immunoincompetent mice, thereby indicating a significant difference in phe-notype between ES cells of the horse and those of the mouse and human.
523
INTRODUCTION
AT THE BLASTOCYST STAGE OF DEVELOPMENT, the preim-plantation mammalian embryo separates into the in-
ner cell mass (ICM) cells of the putative embryo properand the trophectoderm cells that will eventually form theoutermost layer of the fetal placenta (1). Pluripotent EScell lines were derived originally from the ICM of mouseand human preimplantation embryos with retention of thedevelopmental potential to transform into derivatives of allthree embryonic germinal layers, ectoderm, endoderm, andmesoderm, even after prolonged culture in vitro (1–3).Subsequently, it has become apparent that the culture re-quirements for proliferation and maintenance of ES cellsin an undifferentiated state, and the nature of the stimulirequired to induce ES cells to transform into specializedend-stage cells of specific phenotype, vary greatly between
the human and various animal species (1–10). Usefully, itappears that the pig and horse, in contrast to ruminantspecies, will more readily permit the establishment of gen-uine pluripotent ES cell lines following the isolation andrepeated passage of ICM cells in vitro. Li et al. (11) pro-duced a porcine ES cell line in this manner and Saito etal. (12) described the successful production of two horseES cell lines from the repeated culture of the ICM cells ofday-8 frozen-thawed horse blastocysts.
In the present study, we also created three lines of horseES cells by repeated passage of ICM cells recovered im-munosurgically from day-8 blastocysts. We used antis-era generated against a well-recognized panel of humanand mouse ES cell markers to study the phenotypic char-acteristics of horse ICM cells and the ES cell lines cre-ated from them. We also evaluated the transformation ca-pabilities of these ES cells when grown in vitro in the
1Department of Veterinary Medicine, Equine Fertility Unit, Mertoun Paddocks, University of Cambridge, Newmarket, SuffolkCB8 9BH, UK.
2The Babraham Institute, Babraham, Cambridge CB2 4AT, UK.3Clinical Research Centre, Huddinge University Hospital, Karolinska Institute, 141 57 Stockholm, Sweden.4Present address: China Agricultural University, College of Animal Science and Technology, Equine Research Centre, No 2
Yangminyang Xilu, Beijing 100049, China.

absence of a feeder cell layer and/or leukemia inhibitoryfactor (LIF) and in vivo when injected into the testes of severe combined immunodeficiency (SCID)/beigeimmunoincompetent mice.
MATERIALS AND METHODS
Media preparation
STO medium was prepared for the culture of primaryembryonic fibroblasts using basic Dulbecco’s modifiedEagle medium (DMEM; Invitrogen, Groningen, TheNetherlands) containing 10% fetal calf serum (FCS; Hy-Clone, Logan, UT) and 2 mM L-glutamine (Invitrogen).ES medium for the culture of ICM cells consisted of 85%DMEM/nut mix F12 (Invitrogen) containing 15% FCS(HyClone), 2 mM L-glutamine, 1% nonessential aminoacids (Invitrogen), 1 mM sodium pyruvate (Invitrogen),0.1 mM 2-mercaptoethanol (Invitrogen), and 1,000units/ml of human LIF (Invitrogen). This mixture is com-monly used to maintain mouse ES cells in an undiffer-entiated state (13). Conditioned ES medium was recov-ered from culture dishes containing ES cells that had beenco-cultured with a feeder cell monolayer of mouse pri-mary embryonic fibroblasts (mPEF) for 3–4 days.
Preparation of feeder cells for co-culture withICM cells
Two types of fibroblast cells, mPEFs (kindly suppliedby Professor A Surani, The Gurdon Laboratory, Cam-bridge) and horse fetal fibroblasts (hFF; obtained from theculture of limb buds of a 33-day-old horse fetus), wereused as feeder cells for ICM proliferation. Confluentmonolayers of each cell type were treated with mitomycin-C (Sigma, St. Louis, MO) for 2 h, centrifuge-washed threetimes in STO medium, and dispensed into four-well petri-dishes (3,14). ES culture medium was added to these feedercells and the cultures were maintained for at least 1 daybefore they were used for ICM co-culture.
Isolation of ICMs and their proliferation in vitro
Horse blastocysts with a diameter of 500–900 �m andan identifiable ICM containing 100–200 cells were re-covered by nonsurgical uterine lavage on day 8 after ovu-lation, as described previously (15). The ICM of eachblastocyst was isolated by immunosurgery (2,3). Briefly,the trophoblast cells were removed by immersing the em-bryo in a 10% dilution in phosphate-buffered saline(PBS) of a rabbit polyclonal antiserum generated againstthe choriovitelline membrane of a 21-day horse concep-tus, at 38°C for 90 min. The embryos were then trans-ferred to a 5% dilution in PBS of goat anti-rabbit im-
munoglobulin G (IgG; Sigma) for 60 min followed byimmersion in a 20% dilution of guinea pig complement(Invitrogen) in PBS, again for 60 min. The isolated ICMsobtained in this manner were then moved onto a mono-layer of mPEF in ES medium for passage 1.
Following this first passage, the colonies were sepa-rated and then subjected to four different culture condi-tions during passage 2 onwards: (1) continuous culture ona monolayer of mPEF with LIF added to the medium; (2)culture on a monolayer of hFF with LIF in the medium;(3) culture on a monolayer of mPEF without the additionof LIF; (4) culture in ES medium alone with added LIF.
Characterization of proliferated cells for ESmarker expression
Six ES cell markers, alkaline phosphatase (AKP),SSEA-1, SSEA-4, TRA-1-60, TRA-1-81, and the keyembryonic gene Oct-4, were searched for in this im-munocytochemical staining experiment; all have beenidentified previously on human and mouse ES cells (3).The colonies of cultured cells were fixed in 4% parafor-maldehyde for 30 min, washed three times with PBS, andthen incubated for 60 min in a monoclonal antibody gen-erated against one of the following 6 ES cell markers: (1)AKP (Vector Laboratories, Burlingham, CA); (2) SSEA-1 and SSEA-4 (Chemicon International, Temecula, CA);(3) TRA-1-60 and TRA-1-81 (kindly donated by Dr. Pe-ter Andrews, University of Sheffield, UK); and (4) Oct-4 (Santa Cruz Biotechnology, Santa Cruz, CA). After fur-ther wash steps, the cells were incubated for 60 min in abiotinylated anti-mouse gamma globulin solution fol-lowed by a 30-min incubation in an avidin/biotinylatedhorseradish peroxidase (HRP) complex, all supplied inthe ABS Staining System marketed by Vector Laborato-ries). Staining controls were run under the same condi-tions, but in the absence of the primary antibody.
Characterization of in vitro differentiated ES cells
Differentiation of the ES cells in vitro was induced dur-ing passages 15–17 by culturing the cells in feeder cell-free ES medium without LIF in collagen IV (Sigma)-coated four-well Petri dishes. The outgrowths from the EScells were fixed in 4% paraformaldehyde after 20–22 daysof culture and stained for the six ES cell markers by theABC staining technique described previously, and alsoagainst primary monoclonal antibodies specific for �-fe-toprotein (AFP; Sigma), cardiac troponin I (cTnI; SantaCruz Biotechnology), and �-tubulin II (Sigma), which, re-spectively, identify the presence of the three primitivegerm layer cells of endoderm, mesoderm, and ectoderm(16). The nuclei of the differentiated cells were detectedfluorescently by immersing the fixed cells for 10 min inPBS containing 5 �g ml�1 propidium iodide (Sigma).
LI ET AL.
524

TA
BL
E1.
PCR
PR
IME
RS
US
ED
IND
ET
EC
TIN
GG
EN
ET
RA
NS
CR
IPT
SIN
HO
RS
EE
SC
EL
LS
AN
DT
HE
IRD
IFF
ER
EN
TIA
TIO
NPR
OD
UC
TS
Gen
eSe
nse
prim
erA
ntis
ense
pri
mer
Tis
sue
mar
ker
Cat
egor
y
Oct
-4T
CC
CA
GG
AC
AT
CA
AA
GC
TC
TG
CA
GA
TC
TG
GG
CT
CT
CC
CA
TG
CA
TT
CA
AA
CT
GA
Em
bryo
nic
and
ES
germ
cel
lsST
AT
-3T
CT
GG
CT
AG
AC
AA
TA
TC
AT
CG
AC
CT
TT
TA
TT
TC
CA
AA
CT
GC
AT
CA
AT
GA
AT
CT
Em
bryo
nic
and
ES
germ
cel
ls�
-Tub
ulin
III
CA
GA
GC
AA
GA
AC
AG
CA
GC
TA
CT
TG
TG
AA
CT
CC
AT
CT
CG
TC
CA
TG
CC
CT
CN
euro
nsE
ctod
erm
GA
TA
-4C
TC
TG
GA
GG
CG
AG
AT
GG
GA
CG
GG
GA
GC
GG
TC
AT
GT
AG
AG
GC
CG
GC
AG
GC
AT
TH
emat
opoi
etic
Mes
oder
mpr
ecur
sor
cells
�-F
etop
rote
inC
TT
AC
AC
AA
AG
AA
AG
CC
CC
TC
AA
CA
AA
CT
CC
CA
AA
GC
AG
CA
CG
AG
Vis
cera
l ce
llsE
ndod
erm
GA
PDH
GG
GC
TT
GG
CT
TC
GG
TG
AC
AA
CA
CC
AA
GG
CG
GC
CG
AG
CA
AA
GG
CC
TC
TG
CC
AC
CT
TG
CG
GT
TA
ll th
e ce
llsH
ouse
keep
ing

RT-PCR analyses of differentiated ES cells
Gene transcripts for the markers, Oct-4, Stat-3, �-tubu-lin III, GATA-4, �-fetoprotein, and GAPDH were ana-lyzed in undifferentiated and differentiated ES cells us-ing the PCR primers shown in Table 1. ES cells atpassages 6–20 were transferred into suspension cultureand embryoid bodies (EBs) were collected from the cul-tures 4 and 9 days later. mRNA and cDNA samples wereprepared from the undifferentiated ES cells, the EBs andfrom hFF using a Rneasy Midi Kit (Qiagen, 75142) forthe mRNA and an Omniscript RT Kit (Qiagen, 205111)for the cDNA, with Oligo(dt)15 primers, all used ac-cording to the manufacturer’s protocols (Qiagen RT-PCRprotocols). The RT-PCR conditions were 93°C for 2 min,followed by 40 cycles of 93°C for 30 sec, 55°C for 30sec, and 72°C for 1 min. The PCR products were incu-
bated at 72°C for a further 10 min and, finally, the RT-PCR products were run on a 1% agarose gel, stained withEthylene Blue, and captured on a Gel Document.
Teratoma formation in SCID/beige mice
Attempts to generate teratomata in SCID/beige micewere performed as described previously (17). Briefly,approximately 5 � 105 ES cells recovered betweenpassages 14 and 26 from the lines ES/IG P14, ES-PWP16, ES/Balle P20, and ES/OP P26 were each injectedinto one testis of two 8- to 12-week-old male SCID/beige mice. The mice were killed 8–10 weeks later andboth testicles were recovered and sectioned serially.The sections were stained with Hemotoxylin & Eosin(H&E) and examined histologically for signs of tumorformation.
LI ET AL.
526
FIG. 1. Morphology of proliferating ICM cells at passages 1 and 5 when maintained under different culture conditions. Bar,50 �m. (A) A colony of ICM cells at passage 1 after 7 days of co-culture on a feeder cell monolayer of mPEF in the presenceof LIF. (B) A colony of proliferated ES cells at passage 5 being maintained in co-culture on a feeder cell monolayer of mPEFin the presence of LIF. (C) A colony of proliferated ES cells at passage 5 being maintained in co-culture on a feeder cell mono-layer of hFF in the presence of LIF. (D) A colony of proliferated ES cells at passage 5 being maintained in co-culture on a mono-layer of mPEF in the absence of LIF. (E) A colony of proliferated ES cells at passage 5 being maintained in ES medium withadded LIF. (F) A colony of proliferated ES cells at passage 5 being maintained in ES medium without both LIF and feeder cells.
FIG. 2. Phenotypic characteristics of proliferated ES cells at passage 5 following staining in the ABC immunocytochemicalstaining system with monoclonal primary antibodies specific for six ES cell markers, including AKP, SSEA-1, SSEA-4, TRA-1-60, TERA-1-81, and Oct-4. For A–F, the red staining shows the whole colony at low magnification, whereas in A1–F1, theblue staining highlights at higher magnification the positively stained cells at the periphery of each colony. Bar, 50 �m. (A andA1) AKP. (B and B1) TRA-1-60. (C and C1) SSEA-1. (D and D1) Oct-4. (E and E1) TRA-1-81. (F and F1) SSEA-4.
FIG. 3. Differentiation of the ES cells in vitro. The ES cells were maintained for 20–22 days on collagen IV-treated four-wellPetri dishes, in feeder cell-free ES medium without LIF. Blue staining shows a positive reaction for each marker. Bar, 50 �m.(A–C) Morphology of ES cells differentiated into: (A) precursor gland-like cells; (B) fibroblast-like cells; (C) neuron-like cells.(D–F) Positive immunoreactivity for: (D) AFP; (E) cTnI; and (F) �-tubulin III, indicating the presence of precursor endoderm,mesoderm, and ectoderm cells, respectively. (G–I) Nuclear appearance of the three differentiated cell types.
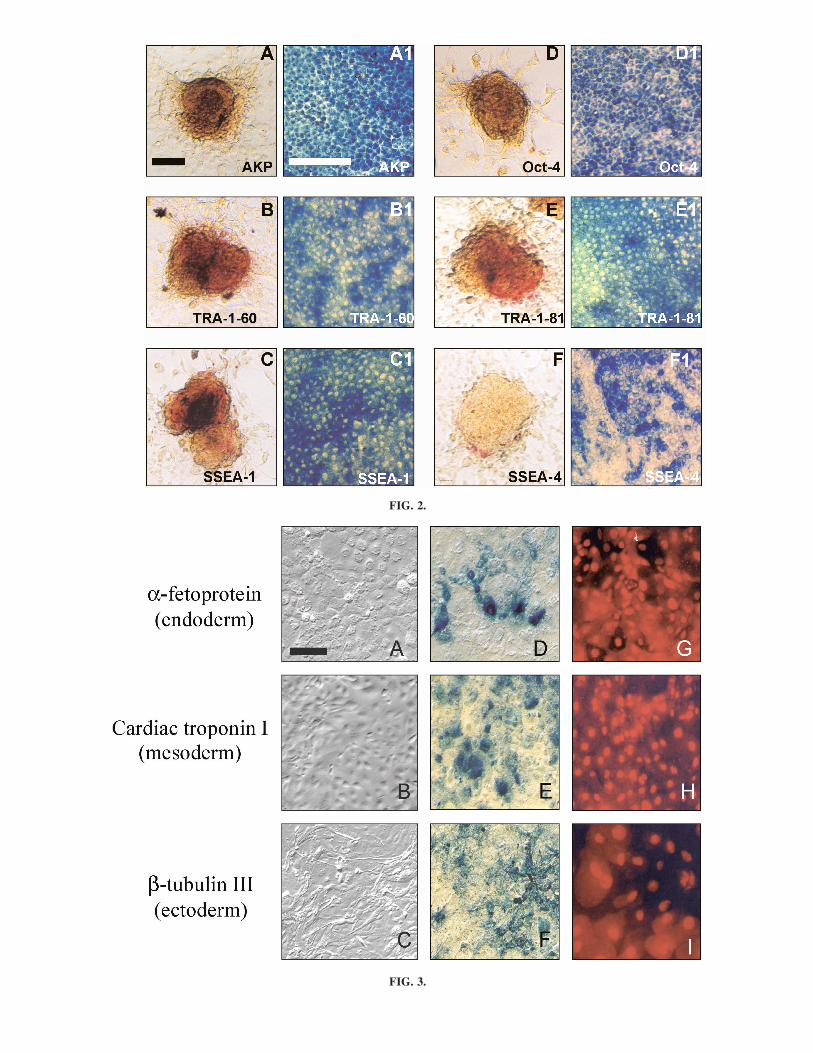
FIG. 2.
FIG. 3.

RESULTS
ICM proliferation in vitro
A total of 18 ICMs were isolated successfully from 24day-7 to -8 blastocysts and, after 7 days of culture on anmPEF or hFF feeder cell monolayer during passage 1,cells from all 18 ICMs had formed expanded colonies(Fig. 1A,B). The morphology of these colonies moreclosely resembled those of mouse ES than human EScells, because they showed underrun cell borders, a smallcytoplasmic/nuclear ratio, and prominent nucleoli fol-lowing cell proliferation during passages 1–5 (1–3; Fig.1C,D). However, the colonies grown in feeder cell-freemedium and those grown without LIF in the medium bothshowed morphological differentiation to the extent thatthe cells had assumed a flattened, epithelioid architectureby the end of passage 5 (Fig. 1E,F).
Cell characteristics of horse ICM andproliferated ES cells
Staining the horse ES cells with five ES markers usedcommonly to identify human and mouse ES cells showedthat they continued to express AKP enzyme activity andthe markers SSEA-1 and -4 and TRA-1-60 and -81 after26 passages (Fig. 2A–E). However, the intensity of stain-ing for SSEA-4 was much lower than the other four mark-ers (Fig. 2C). In a similar manner, the horse ES cells con-tinued to express the POU5f1 gene, encoding thetranscription factor Oct-4 (also called Oct-3 or Oct-3/4)(18–20), over 20 or more passages, thereby illustratingthe undifferentiated pluripotent status of the cells.
Differentiation capacities of the ES cells
The results of growing the ES cells in the absence ofan underlying layer of mEPF of hFF feeder cells areshown in Fig. 3. Under these conditions the cells formedEBs in around 10 days and, by days 20–22, the cellcolonies showed a markedly heterogeneous morphology,including flattened gland-like cells with swollen nuclei(Fig. 3A,C), elongated fibroblast-like cells (Fig. 3B,H),and precursor neuron-like cells with axons and other cellconnections (Fig. 3C,I). The presence of positivelystained cells for AFP, cTnI, and �-tubulin III in thecolonies also heralded the differentiation of the ES cellsinto the three primitive germ layer cells of endoderm (Fig.3D), mesoderm (Fig. 3E), and ectoderm (Fig. 3F).
Molecular marker expression analyzed by RT-PCR
The results of the RT-PCR analyses undertaken tomonitor ES cell differentiation in vitro are shown in Fig.4. ES cells at passages 6–20 cultured in the absence of a
feeder cell layer formed EBs in 4–9 days and they alsoformed blastocyst-like structures when small groups ofthem were injected into cytoplasm-free oocyte zonae pel-lucida (Fig. 4A, b1-2 and c1-3). RT-PCR showed the ex-pression of Oct-4 and Stat-3 genes by both ES cells andEB at day 4 (Fig. 4D). However, although the EBsshowed a weak band for the �-tubulin III gene at day 4,they did not yet express the other markers of the GATA-4 and AFP genes (Fig. 4D). By day 9 of culture, how-ever, the EB and FF cells both showed strong expressionof the three embryonic molecular marker genes, �-tubu-lin III, GATA-4, and AFP (Fig. 4D).
Teratoma formation in SCID/beige mice
Cells from four horse ES cell lines, ES/IG P14, ES/PWP16, ES/Balle P20, and ES/OP P26, when injected intothe testes of a total of 10 immunoincompetentSCID/beige mice (2 mice for each of the ES/IG P14,ES/PW P16, ES/Balle P20 lines, and 4 mice for ES/OPP26), all failed to produce any signs of teratomata. Hu-man ES line H9 cells injected into 6 SCID/beige mice ascontrols all produced, as expected, teratomata that ex-hibited cell types of ectodermal, endodermal, and meso-dermal origin.
DISCUSSION
LIF and mPEF are used commonly to maintain mouseES cells in an undifferentiated state (21). In the presentstudy, we compared the abilities of LIF and two differ-ent types of feeder cells to maintain horse ICM cell pro-liferation and pluripotency in vitro during passages 1–5.The morphology of the resulting colonies more closelyresembled that of mouse ES, than human ES, cells be-cause they showed underrun cell borders, a small cyto-plasmic/nuclear ratio, and prominent nucleoli (1–3) (Fig.1C,D). However, both of the colonies grown in the ab-sence of feeder cells and those grown in the absence ofLIF in the medium showed morphological differentiationto a flattened, epithelioid architecture by the end of pas-sage 5 (Fig. 1E,F). Thus, undifferentiated proliferation ofhorse ICM cells in vitro appears to be dependent uponthe presence of both appropriate feeder cells and LIF inthe culture medium. The cells could not respond ade-quately to LIF alone, as had been shown previously incultures of human ES cells (3).
ES and EC cells, as well as ICM cells of mouse blas-tocysts, express a panel of cell-surface markers that char-acterize their undifferentiated and pluripotent status. Elevated AKP expression is also associated with nondif-ferentiation and pluripotency in mouse and human ES,EG, EC, and ICM cells (1–3,22). In a similar manner, thehorse ES cells in this study still showed raised AKP en-
LI ET AL.
528

HORSE ES CELLS FROM ICM CELLS
529
FIG. 4. The results of EB formation and RT-PCR analysis for in vivo differentiation of the ES cells. (A) ES colony culturedin vitro. (B1 and B2) EBs formed in vitro from differentiated ES masses at: (B1) day 4; (B2) day 9. (C1 and C2) Blastocyst-likestructures formed after 5–6 days culture in vitro following the injection of ES cell masses into empty oocyte zonae pellucida.(C3) A blastocyst-like structure stained with Oct-4 FITC. (D) Results of RT-PCR performed on: (1) ES cells; (2) day 4 EBs; (3)day 9 EBs; and (4) FFs. Oct-4 and Stat-3, ES cell markers; �-Tubulin III, ectoderm cell marker; GATA-4, mesoderm cell marker;AFP, endoderm cell marker; GAPDH, control marker.
FIG. 5. Directed differentiation of ES cells in vitro. The cells were immunostained using the ABC system and applying the an-tibody for the AFP marker for fibroblasts and an antibody specific for collagen type IV for the cartilage cells. (A) ES cells co-cultured with horse fibroblasts for 7 days. (B) ES cells co-cultured with horse cartilage cells for 15 days.
D

zyme activity (Fig. 2A) and expressed the SSEA-1, TRA-1-60, and TRA-1-81 markers at passage 26 (Fig. 2B,D,E),thereby demonstrating their similarity to mouse ES andhuman EG cells. However, in support of previous obser-vations of Saito et al. (12), the horse ES cells showedmuch weaker staining with SSEA-4 than the other EScell marker antibodies. In a similar manner, whereasmouse ES cells express SSEA-1 strongly, human ES cellsdo not express SSEA-1 at all (1–3). Taken together, thesefindings suggest that basic differences exist amongmouse, human, and horse embryos in their expression ofmolecules associated with undifferentiated pluripotency.
Oct-4 is present in the mouse zygote and it is requiredthroughout development to the blastocyst stage to estab-lish and maintain the pluripotency of the ICM and theepiblast (23,24). Oct-4 is also expressed in the primor-dial germ cells of mice and by undifferentiated mouseand human ES cells (18,19,25). The present study dem-onstrated that the Oct-4 gene is also expressed by pro-liferating horse ES cells during more than 20 passages(Fig. 2G,I). The result was amplified by the positive RT-PCR analysis for Oct-4 gene expression by the ES cellsand also by EBs after in vitro differentiation of the EScells (Fig. 4).
The definition of an ES cell line is more than just self-replicating stem cells derived from the embryo that candifferentiate into almost any cell type within the body. Inthe present study, we were able to demonstrate that thehorse ES cells derived from in vitro proliferated ICMs ofday-7 to 8 blastocyst-stage embryos after 15–17 passagesin vitro could still differentiate into cells of the threeprimitive germ cell layers, endoderm, mesoderm, and ec-toderm (Figs. 3 and 5). These results suggest that the threeES cell lines established in the experiment have the equiv-alent transformation capacity of established ES cell linesof other species such as mice and humans. Hence, theycan be employed in a wide range of future in vitro andin vivo transformation studies.
Teratoma formation by ES cells injected intosCID/beige mice is a well-established method to evalu-ate the transformation potential of ES cell lines. Thesemultitussue tumors have been shown to develop in themuscles, joints, or testis tissue of SCID/beige within 8–10weeks of injection of mouse and human ES cells (1–3,22).In the present study, none of the four horse ES cell linesat passages 14–26 that were injected into the testes ofSCID/beige mice showed any cell proliferation or dif-ferentiation in the testis tissue, even after the longergrowth period of 10 weeks. We are at a loss to explainthis clearly negative result and, in the absence of evi-dence to the contrary, must assume that an important dif-ference may exist between horse pluripotent ES cells andthose of at least the mouse and human in their potentialfor multitussue tumor formation in foreign immunologi-cally privileged sites. Perhaps it bodes well for the future
application of horse ES cells to improve the rate and qual-ity of healing of a wide range of exercise-induced injuriesin this essentially athletic species.
In summary, the results of this study demonstrated thatLIF and either mPEF or hFF feeder cells are necessary tomaintain an undifferentiated state in ES cells derived fromthe ICM of horse blastocysts. The cells continue to expressAKP activity, and they also express the cell-surface mark-ers SSEA-1, TRA-1-60, TRA-1-81, and Oct-4, but stainonly weakly for SSEA-4, after prolonged culture in vitro.Furthermore, they have the potential to differentiate in vitrointo the three precursor germ layer cells, endoderm, meso-derm, and ectoderm, even though they appear incapable oftransforming into teratomata in vivo.
ACKNOWLEDGMENTS
We thank Dr. Peter Andrews of the University ofSheffield for kindly providing monoclonal antibodies toTRA-1-60 and TRA-1-81. We are also grateful to MissesAnne-Cécile Lefranc, Sandra Wilsher, and Clare Tipladyfor their assistance in recovering horse embryos. Thestudy was generously supported by the Moller Charita-ble Trust, the Japan Racing Association, and theThroughbred Breeders’ Association of Great Britain.
REFERENCES
1. Martin GR. (1981). Isolation of a pluripotent cell line fromearly mouse embryos cultured in medium conditioned byteratocarcinoma stem cells. Proc Natl Acad Sci USA 78:7634–7638.
2. Evans M and M Kaufman. (1981). Establishment in cul-ture of pluripotental cells from mouse embryos. Nature292:154.
3. Thomson JA, J Itskovitz-Eldor, SS Shapiro, MA Waknitz,JJ Swiergiel, VS Marshall and JM Jones. (1998). Embry-onic stem cell lines derived from human blastocysts. Sci-ence 282:1145–1147.
4. Anderson GB, RH BonDurant, L Goff, J Groff and ALMoyer. (1996). Development of bovine and porcine ter-atomas in athymic mice. Anim Reprod Sci 45:231–240.
5. Lumelsky N, O Blondel, P Laeng, I Velasco, R Ravin andR McKay. (2001). Differentiation of embryonic stem cellsto insulin-secreting structures similar to pancreatic islets.Science 292:1389–1394.
6. Choi D, HJ Oh, UJ Chang, SK Koo, JX Jiang, SY Hwang,JD Lee, GC Yeoh, HS Shin, JS Lee and B Oh. (2002). Invivo differentiation of mouse embryonic stem cells into he-patocytes. Cell Transpl 11:359–368.
7. Wichterle H, I Lieberam, JA Porter and TM Jessell. (2002).Directed differentiation of embryonic stem cells into mo-tor neurons. Cell 110:385–397.
8. Yamamoto H, G Quinn, A Asari, H Yamanokuchi, T Ter-atani, M Terada and T Ochiya. (2003). Differentiation of
LI ET AL.
530

embryonic stem cells into hepatocytes; biological functionsand therapeutic application. Hepatology 37:983–993.
9. Fair JH, BA Cairns, M LaPaglia, J Wang, AA Meyer, HKim, S Hatada, O Smithies and L Pevny. (2003). Induc-tion of hepatic differentiation in embryonic stem cells byco-culture with embryonic cardiac mesoderm. Surgery 134:189–196.
10. Lester L. (2004) Manipulating stem cells to make islets. JInvest Med 52:284–291.
11. Li M, YH Li, Y Hou, XF Sun, Q Sun and WH Wang.(2004). Isolation and culture of pluripotent cells from invitro produced porcine embryos. Zygote 2:43–48.
12. Saito S, H Ugai, K Sawai, Y Yamamoto, A Minamihashi,K Kurosaka, Y Kobayashi, T Murata, Y Obata and KYokoyama. (2002). Isolation of embryonic stem-like cellsfrom equine blastocysts and their differentiation in vitro.FEBS Lett 20:389–396.
13. Williams RL, DJ Hilton, S Pease, TA Willson, CL Stew-art, DP Gearing, EF Wagner, D Metcalf, NA Nicola andNM Gough. (1988). Myeloid leukaemia inhibitory factormaintains the developmental potential of embryonic stemcells. Nature 336:684–687.
14. Li XH, LH-A Morris and WR Allen. (2002). In vitro de-velopment of horse oocytes reconstructed with the nucleiof fetal and adult cells. Biol Reprod 66:1288–1292.
15. Allen WR. (1982). Embryo transfer in the horse. In: Mam-malian Egg Transfer. CE Adams, ed. CRC Press, Boca Ra-ton, pp 135–154.
16. Xu C, MS Inokuma, J Denham, K Golds, P Kundu, JDGold and K Carpenter. (2001). Feeder-free growth of un-differentiated human embryonic stem cells. NatureBiotechnology 19:971–974.
17. Gertow K, S Wolbank, B Rozell, R Sugars, M Andäng, CLParish, MP Imreh, M Wendel and L Ährlund-Richter.(2004). Organised development from human embryonicstem cells after injection into immunodeficient mice. StemCells Dev 13:421–435.
18. Scholer HR, S Ruppert, N Suzuki, K Chowdhury and PGruss. (1990). New type of POU domain in germ line-spe-cific protein Oct-4. Nature 344:435–439.
19. Rosner JL. (1990). Reflections of science as a product. Na-ture 345:108.
20. Nichols J, B Zevnik, K Anastassiadis, H Niwa, D Klewe-Nebenius, I Chambers, H Scholer and AG Smith. (1998).Formation of pluripotent stem cells in the mammalian em-bryo depends on the POU transcription factor, Oct-4. Cell95:379–391.
21. Murray P and D Edgar. (2001). The regulation of embry-onic stem cell differentiation by leukaemia inhibitory fac-tor (LIF). Differentiation 68:227–234.
22. Thomson JA, J Kalishman, TG Golos, M Durning, CP Har-ris, RA Becker and JP Hearn. (1995). Isolation of a pri-mate embryonic stem cell line. Proc Natl Acad Sci USA92:7844–7848.
23. Niwa H, J Miyazaki and AG Smith. (2000). Quantitativeexpression of Oct-3/4 defines differentiation, dedifferenti-ation or self-renewal of ES cells. Nature Genet 24:372–376.
24. Rodda SJ, SJ Kavanagh, J Pathjen and P Rathjen. (2002).Embryonic stem cell differentiation and the analysis ofmammalian development. Int J Devel Biol 46:449–458.
25. Pesce M, X Wang, DJ Wolgemuth and H Scholer. (1998).Differential expression of the Oct-4 transcription factor dur-ing mouse germ cell differentiation. Mech Devel 71:89–98.
Address reprint requests to:Dr. W.R. Allen
The Equine Fertility UnitMertoun Paddocks
Woodditton Road, NewmarketSuffolk CB8 9BH, UK
E-mail: [email protected]
Received April 3, 2006; accepted June 6, 2006.
HORSE ES CELLS FROM ICM CELLS
531




















