
The IGFBP-3 mRNA and protein levels are IGF-I-dependent and GH-independent in MG-63 human...
13
Molecular and Cellular Endocrinology 175 (2001) 15 – 27 The IGFBP-3 mRNA and protein levels are IGF-I-dependent and GH-independent in MG-63 human osteosarcoma cells Roberto Rosato a,1 , Katia Gerland b,2 , He ´le `ne Jammes c,3 , Nelly Bataille-Simoneau b,2 , Berta Segovia a,1 , Louis Mercier b,2 , Andre ´ Groyer a,d,*,1, a Inserm U.142, Ho ˆpital St Antoine, 75571, Paris Cedex 12, France b LHEA, UFR des Sciences Me ´dicales et Pharmaceutiques, 1 rue Haute de Recule ´e, 49045, Angers, France c Laboratoire d’Endocrinologie Mole ´culaire, INRA, 78352, Jouy en Josas, France d Inserm U. 327, Faculte ´ de Me ´decine Zaier Bichat, 75870 - Paris Ce ´dex 18, France Received 17 October 2000; accepted 27 January 2001 Abstract Growth Hormone (GH), Insulin-like Growth Factors (IGFs) and IGF-Binding Proteins which modulate the IGFs’ bioavailabil- ity (e.g. IGFBP-3, -4, -5), are essential regulators of bone remodeling. In this study, MG-63 human osteosarcoma cells were used as a model system to investigate the mechanism(s) whereby IGF-I and GH control IGFBP-3 gene expression. Physiological concentrations of IGF-I (1 – 20 nM) induced a dose-dependent increase in the steady-state amount of IGFBP-3 mRNA (maximal stimulation: 9–10-fold). This increase was detectable 3 h after the onset of IGF-I treatment, was enhanced over a 24 h period, then plateaued until at least 30 h. Consistently, a dose-dependent increase in IGFBP-3 secretion ( 40–50-fold at IGF-I concentrations 16 nM) was observed by western ligand- and immuno-blot analysis of MG-63 cells conditioned medium, and its time course was similar to that observed for IGFBP-3 transcripts. IGFBP-3 mRNA stability (t 1/2 20 h) was identical in the presence or absence of IGF-I treatment. By contrast, human (h) GH treatment (24–72 h) of MG-63 cells did not increase IGFBP-3 secretion in the conditioned medium. Ectopic expression of recombinant rat GH-R resulted in hGH-enhanced expression of GH-responsive reporter gene constructs, but did not increase endogenous IGFBP-3 gene expression, suggesting that the GH unresponsiveness was not only due to the very low level of GH binding sites at the plasma membrane level. Altogether, these results support the conclusions that in MG-63 cells (i) transcriptional rather post-transcriptional mechanisms are involved in the IGF-I-induced increase of IGFBP-3; (ii) the abundance of GH-R is very low at the plasma membrane level; (iii) the dowstream GH-signaling cascade is fully functional in this human osteosarcoma cell line; and (iv) the endogenous IGFBP-3 gene is not responsive to hGH in human MG-63 osteosarcoma cells. © 2001 Elsevier Science Ireland Ltd. All rights reserved. Keywords: IGFBP-3; Osteosarcoma cells; Gene expresssion; Insulin-like growth factor-I; Growth hormone; Growth hormone receptor; Actino- mycin-D www.elsevier.com/locate/mce 1. Introduction Although recent evidence suggests that Growth Hor- mone (GH) participate in bone remodelling, the precise mechanism by which GH affects bone cell functions remains unclear. Consistent with the somatomedin hy- pothesis, Insulin-like Growth Factors (IGFs) have been demonstrated to mediate GH effects in some target organs, but their role in mediating GH effects in os- teoblasts is still controversial (reviewed in Kassem, 1997; Ohlsson et al., 1998). Although there are two types of IGF receptors, most of the experimental studies have shown that IGF-I as well as IGF-II mediate their effects on cell proliferation and differentiation via the type-I receptor (IGF-IR), structurally related to the insulin receptor (Centrella et al., 1990; Oh et al., 1993a; Raile et al., 1994). With regard to skeletal cells, numerous reports show (i) that * Corresponding author. Present address: INSERM U.327, 16, Henri Huchard, BP416, 75870 Paris Cedex 18, France. Fax: +33-1- 44859279. E-mail address: [email protected] (A. Groyer 1 ). 1 Fax: +33-1-43433234. 2 Fax: +33-2-41735888. 3 Fax: +33-1-34652273. 0303-7207/01/$ - see front matter © 2001 Elsevier Science Ireland Ltd. All rights reserved. PII:S0303-7207(01)00434-8
Transcript of The IGFBP-3 mRNA and protein levels are IGF-I-dependent and GH-independent in MG-63 human...

Molecular and Cellular Endocrinology 175 (2001) 15–27
The IGFBP-3 mRNA and protein levels are IGF-I-dependent andGH-independent in MG-63 human osteosarcoma cells
Roberto Rosato a,1, Katia Gerland b,2, Helene Jammes c,3, Nelly Bataille-Simoneaub,2,Berta Segoviaa,1, Louis Mercierb,2, Andre Groyera,d,*,1,
a Inserm U.142, Hopital St Antoine, 75571, Paris Cedex 12, Franceb LHEA, UFR des Sciences Medicales et Pharmaceutiques, 1 rue Haute de Reculee, 49045, Angers, France
c Laboratoire d’Endocrinologie Moleculaire, INRA, 78352, Jouy en Josas, Franced Inserm U. 327, Faculte de Medecine Za�ier Bichat, 75870 - Paris Cedex 18, France
Received 17 October 2000; accepted 27 January 2001
Abstract
Growth Hormone (GH), Insulin-like Growth Factors (IGFs) and IGF-Binding Proteins which modulate the IGFs’ bioavailabil-ity (e.g. IGFBP-3, -4, -5), are essential regulators of bone remodeling. In this study, MG-63 human osteosarcoma cells were usedas a model system to investigate the mechanism(s) whereby IGF-I and GH control IGFBP-3 gene expression. Physiologicalconcentrations of IGF-I (1–20 nM) induced a dose-dependent increase in the steady-state amount of IGFBP-3 mRNA (maximalstimulation: �9–10-fold). This increase was detectable 3 h after the onset of IGF-I treatment, was enhanced over a 24 h period,then plateaued until at least 30 h. Consistently, a dose-dependent increase in IGFBP-3 secretion (�40–50-fold at IGF-Iconcentrations�16 nM) was observed by western ligand- and immuno-blot analysis of MG-63 cells conditioned medium, and itstime course was similar to that observed for IGFBP-3 transcripts. IGFBP-3 mRNA stability (t1/2 �20 h) was identical in thepresence or absence of IGF-I treatment. By contrast, human (h) GH treatment (24–72 h) of MG-63 cells did not increaseIGFBP-3 secretion in the conditioned medium. Ectopic expression of recombinant rat GH-R resulted in hGH-enhancedexpression of GH-responsive reporter gene constructs, but did not increase endogenous IGFBP-3 gene expression, suggesting thatthe GH unresponsiveness was not only due to the very low level of GH binding sites at the plasma membrane level. Altogether,these results support the conclusions that in MG-63 cells (i) transcriptional rather post-transcriptional mechanisms are involvedin the IGF-I-induced increase of IGFBP-3; (ii) the abundance of GH-R is very low at the plasma membrane level; (iii) thedowstream GH-signaling cascade is fully functional in this human osteosarcoma cell line; and (iv) the endogenous IGFBP-3 geneis not responsive to hGH in human MG-63 osteosarcoma cells. © 2001 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: IGFBP-3; Osteosarcoma cells; Gene expresssion; Insulin-like growth factor-I; Growth hormone; Growth hormone receptor; Actino-mycin-D
www.elsevier.com/locate/mce
1. Introduction
Although recent evidence suggests that Growth Hor-mone (GH) participate in bone remodelling, the precisemechanism by which GH affects bone cell functions
remains unclear. Consistent with the somatomedin hy-pothesis, Insulin-like Growth Factors (IGFs) have beendemonstrated to mediate GH effects in some targetorgans, but their role in mediating GH effects in os-teoblasts is still controversial (reviewed in Kassem,1997; Ohlsson et al., 1998).
Although there are two types of IGF receptors, mostof the experimental studies have shown that IGF-I aswell as IGF-II mediate their effects on cell proliferationand differentiation via the type-I receptor (IGF-IR),structurally related to the insulin receptor (Centrella etal., 1990; Oh et al., 1993a; Raile et al., 1994). Withregard to skeletal cells, numerous reports show (i) that
* Corresponding author. Present address: INSERM U.327, 16,Henri Huchard, BP416, 75870 Paris Cedex 18, France. Fax: +33-1-44859279.
E-mail address: [email protected] (A. Groyer1).1 Fax: +33-1-43433234.2 Fax: +33-2-41735888.3 Fax: +33-1-34652273.
0303-7207/01/$ - see front matter © 2001 Elsevier Science Ireland Ltd. All rights reserved.PII: S 0 3 0 3 -7207 (01 )00434 -8

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–2716
IGF-I has prominent anabolic effects on bone in vitro,enhancing proliferation and differentiation of os-teoblasts in rodent and human species (McCarthy et al.,1989; Wergedal et al., 1990; Schmid, 1993; Furlanettoet al., 1994); and (ii) that expression of IGF-I is regu-lated by several hormones and locally produced growthfactors which are known for their prominent effects onbone metabolism in cultured osteoblastic cells (Canalisand Gabbitas, 1994; Kobayashi et al., 1994; Westleyand May, 1994; Delany and Canalis, 1996; Yeh et al.,1996).
In biological fluids, IGFs interact with specific bind-ing proteins (IGFBPs) that modulate their action. Sofar, six IGFBPs referred to as IGFBP-1 to 6 have beenidentified. IGFBPs are also synthesized in cells of theosteoblast lineage, and IGFBPs gene expression as wellas IGFBPs secretion are regulated by hormones andlocally produced growth factors (Mc Carthy et al.,1994; Schmid et al., 1994; Wakao et al., 1994; Canalisand Gabbitas, 1995; Knutsen et al., 1995; Kassem etal., 1996; Zhou et al., 1996). Both processes were shownto be dependent on the proliferative or differentiatedstate of the cells (Birnbaum and Wiren, 1994; Thrailkillet al. 1996).
During the past decade, numerous in vivo (Zapf etal., 1989; Kanety et al., 1993; Jones and Clemmons,1995) and in vitro (reviewed in Jones and Clemmons,1995) studies have been devoted to the regulation ofIGFBP-3 expression. Specially, in human bone cellsIGFBP-3 gene expression and protein secretion hasbeen reported to be enhanced by GH, the IGFs and1,25 (OH)2 vitamin-D3; and inhibited by dexam-ethasone (Bale and Conover, 1992; Mohan et al., 1992;Schmid et al., 1994). With regard to the regulation ofIGFBP-3 by GH, some studies support IGF-I depen-dency, whereas others favor an IGF-I independentregulation.
In contrast to the somatomedin hypothesis, a dualeffector theory of growth hormone action in adipocyteshas been proposed (Zezulak and Green, 1986). Thishypothesis implies that GH stimulates maturation ofpreadipocytes and make them more responsive to IGF-I. Likewise, Langdahl et al. (1998) have reported asynergistic effect of GH and IGF-I on the proliferativeresponse of non-transformed human osteoblasts, andMehlus and Ljunghall (1996) have shown that GH andIGF-I do not activate identical patterns of gene expres-sion in human osteoblasts.
The aim of the present study was to gain moreinsights into the mechanisms by which GH/IGF-I/IGF-BPs exert their action in cells of the osteoblast lineage.The human osteosarcoma cell line MG-63 was chosenas a model system since it exhibits phenotypic featuresrepresentative of the osteoblastic lineage. For example,(i) the expression of phosphatase alkaline, and of os-teocalcin, two characteristic markers of bone derived
cells is induced by 1,25-dihydroxyvitamin D3 in MG-63osteosarcoma cells (Clover and Gowen, 1994; Aubin etal. 1996); (ii) the cell-surface heparan sulfate syndecan-1and -2 are expressed in MG-63 cells as well as inneonatal calvarial osteoblasts cultured in conditionsthat promote osteogenesis (Birch and Skerry, 1999); (iii)�3 and �1 are the major integrin subunits detected onMG-63 as well as bone-derived cells (Clover andGowen, 1994). Moreover, MG-63 cells display addi-tional characteristics that makes them attractive forsuch a study. First, IGF-I-R is expressed in MG-63cells, and is autophosphorylated in response to IGF-I(Lopaczynski et al., 1993). Second, there is no evidencein this cell line neither for IGF-I, nor for IGF-IIexpression under basal culture conditions (Strom et al.,1994; Okasaki et al., 1995). Therefore, any putativeautocrine /paracrine effects of the IGFs can be ruledout in MG-63 cell cultures, and only exogenous IGF-I(i.e. added in the culture medium) would trigger anIGF response. Third, in MG-63 cells, IGFBP-3 geneexpression is correlated with an inhibition of cell prolif-eration and with an induced expression of bone differ-entiation markers (Velez-Yanguas et al., 1996). Inaddition, data from our laboratory support the notionthat when added to the culture medium, IGF-I controlsits own bioavailability via both IGFBP-3 production byMG-63 cells and the regulation of IGFBP-3 proteolysisin the conditioned medium (Lalou and Binoux, 1993).
In this study we investigated the regulation ofIGFBP-3 expression in the MG-63 cell line. The datashow (i) that IGF-I significantly increased IGFBP-3transcript levels and to an even greater extent accumu-lated protein levels in the culture medium; and (ii) thatIGFBP-3 mRNA stability was not affected by IGF-I.In addition, we observed that GH per se was unable toinduce IGFBP-3 gene expression in MG-63 cells, de-spite evidence for an operative GH transduction path-way. Altogether, our results therefore support theconclusion that the endogenous IGFBP-3 gene is notresponsive to hGH in human MG-63 osteosarcomacells.
2. Materials and methods
2.1. Materials
Recombinant human (rh) IGF-I (rhIGF-I) andGrowth Hormone (rhGH) were gifts from Ciba-Geigyand from Sanofi or Pharmacia SA, respectively. IGF-I,IGF-II and GH were iodinated with Na125I (Amer-sham) to a specific activity of 100–200 mCi/mg usingthe chloramine-T method, and purified by gel filtrationat pH 7.0.
The rabbit polyclonal anti-IGFBP-3 antibody wasgenerated in the laboratory against non-glycosylated

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–27 17
rhIGFBP-3, and have been previously described (Lalouand Binoux, 1993). Chemical reagents were purchasedfrom Sigma.
2.2. Oligonucleotides for PCR analyses
GH-R (456 bp amplicon): GH-R1, 5�-CT-CAACTGGACTTTACTGAACG-3� (forward primer,fo), GH-R2, 5�-AATCTTTGGAACTGGAACTGGG-3� (reverse primer, re); GH-R (420 bp — wild-type- or354 bp-GH-Rd3-amplicons) (Urbanek et al., 1992):GH-R3, 5�-CCTACAGGTATGGATCTCTGT-3� (5�-end of exon 2), GH-R4, 5�-CCACCATTGGCTAGT-TAGCTT-3� (exon 5); IGFBP-3 (415 bp amplicon):5�-GCCGCGGGCTCTGCGTCAACGC-3� (fo, exon1), 5�-CTGGGACTCAGCACATTGAGGAAC-3� (re,exon 3); b-actin (227 bp amplicon): 5�-AAGATGACCCAGATCATGTT-3� (fo), 5�-AGGATCTTCAT-GAGGTAGTT-3� (re).
2.3. Plasmids
GH-responsive reporter plasmids (pGHRE-I-CAT,pGHRE-Imut-CAT, pGHRE-II-CAT and pGHRE-IImut-CAT, pLHRE-Luc) and rat GH-R expressionvectors (pECE-GHR and pcDNA3-GHR) were de-scribed elsewhere (Dinerstein et al., 1995; Legraverendet al., 1996; Sotiropoulos et al., 1996), and were kindlyprovided by A. Le Cam (Inserm U.376, France) and byJ. Finidori (Inserm U.344, France).
2.4. Cell culture
MG-63 human osteosarcoma cells were cultured inDulbecco’s Modified Eagle’s Medium (DMEM) supple-mented with 5 or 10% heat-inactivated fetal calf serum(FCS, DMEM-5 or -10), 100 U/ml penicillin-G, 50�g/ml streptomycin and 1 mM glutamine, as previouslydescribed (Lalou et al., 1994).
2.5. Transfection and CAT and Luciferase assays
MG-63 cells seeded in six-well plates and grown to�80% confluency in DMEM-5, were co-transfectedwith 3 �g of the GH-responsive gene reporter con-structs and 1 �g of pCMV-�-Gal (Clontech), an inter-nal monitor for transfection efficiency, �1 �gpECE-GHR, using the calcium phosphate co-precipita-tion procedure (Chen & Okayama, 1987). Twenty hourspost-transfection, cells were rinsed with phosphate-buffered saline (PBS), and the medium was replacedwith fresh DMEM-0.5. Eight hours later (i.e. 28 h aftertransfection), GH treatment (16 nM) was initiated andprotracted for 18 h. Cells were then washed, scrappedin chilled PBS and resuspended in 120 �l of cell lysisbuffer (Promega). CAT activities were measured as
described by Paquereau and Le Cam (1992), and nor-malized relative to �-galactosidase activity.
In some experiments ExGen 5004 (Euromedex) wasused as DNA delivering agent (5 �g plasmid DNA/10�l ExGen 500). The experimental design was as de-scribed above. Luciferase activities were measured ac-cording to Sotiropoulos et al., (1996).
2.6. Western -ligand and -immuno blotting
The conditions used have been extensively describedin Lalou et al. (1994). Briefly, after removal of celldebris by centrifugation, the conditioned media wereconcentrated by ultrafiltration (Centricon, Amicon) andstored at −20°C. IGFBP-3 analysis was performed onaliquots equivalent to 0.5 ml of unconcentrated condi-tioned medium, prior to concentration.
2.7. RNA extraction
Cell monolayers were rinsed with PBS and totalRNA prepared according to the acid-guanidine thio-cyanate/phenol/chloroform method (Sambrook et al.,1989).
2.8. Northern blot analysis
RNA samples (20 �g) were size fractionated on 1.2%formaldehyde-agarose gels and analysed by northern-blotting as previously described (Lalou et al., 1994).Hybridization with the 32P-labeled hIGFBP-3 cDNAwas performed at 42°C for 18–20 h. In some instances,membranes were stripped off (boiling in 0.5% SDS),before re-hybridization with a 32P-labeled GAPDHcDNA probe (used as an internal standard).
2.9. Re�erse transcription
Five micrograms (GH-R) or 1 �g (IGFBP-3, and�-actin) of total cellular RNA were reversed trancribedwith 200 units of MoMLV reverse transcriptase (Gibco-BRL). The reaction mixture (20 �l) contained 50 mMTris–HCl (pH 8.3), 75 mM KCl, 10 mM DTT, 3 mMMgCl2, 1 mM of each dNTP, 2.5 units of RNAsin
4 A dramatic drop in reporter gene activity was always observedwhen co-transfection of two or more plasmids was realized withExgen 500. Since the same observation was made when Luc con-structs were co-transfected with either pUC or pCH110 (�-galactosi-dase expression vector; Pharmacia) DNA, we deduced that thisdecrease was not dependent on the presence of multiple eucaryoticpromoter sequences that could trap component(s) of the basal tran-scriptional machinery. Therefore, Luc assays cannot be normalized,using �-galactosidase or CAT expression vectors. Fortunately, tripli-cate values for reporter gene activity were highly reproducible in allexperiments. All the data obtained with the ExGen500 protocol werethus expressed as Luc activity/�g protein.

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–2718
(Promega) and either random hexanucleotide primers(100 pmoles, GH-R), or specific 3�-reverse primers (20pmoles of each, IGFBP-3, �-actin). Following incuba-tion at 37°C for 1 h, the mixture was heated to 95°C,snap cooled and stored at −20°C.
2.10. Polymerase chain reaction
Replicate aliquots of the reverse transcription mix-tures (equivalent to 5 �g, 250 ng and 50 ng of totalcellular RNA for GH-R, IGFBP-3 or �-actin, respec-tively) were submitted to PCR amplification. The reac-tion mixtures (50 �l) contained 10 mM Tris–HCl, pH9, 50 mM KCl, 1.5–2 mM MgCl2, 200 �M dNTPs,0.5–1 �M of forward and reverse primers, and 1 U TaqDNA-polymerase (Perkin-Elmer, Promega or BioprobeSystems). After an initial denaturation step (94°C for 10min), amplifications were carried out as follows: (i)94°C, 1 min; 60°C, 1 min; 72°C, 1 min (30 cycles,GH-R); and (ii) 94°C, 30 s; 50°C, 20 s; 72°C, 30 s (28cycles-IGFBP-3- or 20 cycles-�-Actin). In all cases, thePCR reaction was completed by a 10 min elongationstep at 72°C. The amplified products were resolved byagarose gel electrophoresis (2% agarose), and stainedwith ethidium bromide.
When semi-quantitative RT-PCR was carried out(IGFBP-3 and �-actin), the PCR reaction mixture con-tained [32P]-labeled primers (�105 cpm/assay). Thebona fide PCR fragments were excised from the agarosegel, and the amount of incorporated radioactivity wasdetermined by liquid scintillation.
2.11. GH binding
Replicate monolayers of MG-63 cells cultured inDMEM supplemented with 5% FCS were grown to�80% confluency in six-wells plates. Cells were thentransferred in serum-free DMEM for 12–24 h prior tothe determination of hGH binding. At the time of theexperiment, the medium was removed, cell monolayerswere rinsed with PBS, and overlaid with 1 ml/well ofPBS containing 1% BSA, 125I-rhGH (105 cpm/ml; AS,�70–80 mCi/mmol) in the absence or presence ofvarious concentrations of unlabeled competitors (hGHor ovine PRL; range: 10−11–10−6 M). After a 2 hincubation at 20°C the incubation mixture was dis-carded, monolayers were rinsed three times with 5 ml ofchilled PBS, and cells were lysed in 0.1% SDS. Cell-as-sociated radioactivity was measured by liquid scintilla-tion counting.
2.12. Quantitati�e analysis
Quantitative analysis of autoradiograms (Northern-,Western ligand- and immuno-blots) was performed bylaser densitometry scanning (Ultroscan XL, Pharmacia-
LKB), using the GSXL (Pharmacia-LKB) or NIH Im-age softwares.
3. Results
3.1. IGFBP-3 protein le�els and transcripts areincreased by IGF-I in MG-63 cells
Under basal conditions the overall secretion ofIGFBP-3 by MG-63 cells was very low. The character-istic 39/42 kDa doublet of IGFBP-3 was barely de-tectable by ligand-blot analysis of the conditionedculture medium, whereas the 24 kDa IGFBP-2 specieswas predominant (Fig. 1A). Immunoblot analysis confi-rmed that the 39/42 kDa bands corresponded toIGFBP-3 (Fig. 1B). Moreover, only the intact forms ofIGFBP-3 were detected, since there was no evidence forany proteolytic fragment in the conditioned medium(i.e. no band at 30 kDa corresponding to the majorproteolytic fragment). This is consistent with the obser-vation reported by Lalou et al. (1994) that the condi-tioned media of MG-63 cells do not contain anysignificant protease activity. Physiological concentra-tions of IGF-I (ranging between 1 and 12 nM) induceda dose-dependent increase in IGFBP-3, as evidenced byligand-blot (Fig. 1A) and immuno-blot analysis (Fig.1B). IGFBP-3 protein levels plateaued at higher con-centrations of IGF-I (�20 nM). Densitometric scan-ning of the ligand- and immuno-blots showed thatIGFBP-3 levels in the culture medium were increased�7–9-fold in presence of 1 nM IGF-I and up to�40–50-fold at 12 nM.
Analysis of total cellular RNAs by northern blottingalso shows that the amount of IGFBP-3 mRNA, fairlylow under basal culture conditions, was increased byIGF-I in a dose dependent manner (Fig. 2A). The�3-fold increase at 1 nM IGF-I, was raised up to�9-fold at IGF-I concentrations �12 nM, as deducedfrom densitometric scanning of the northern blots (Fig.2B).
3.2. Mechanisms in�ol�ed in IGF-I-stimulated IGFBP-3expression
3.2.1. Kinetics of IGFBP-3 RNA expression in MG-63cells and of protein secretion in the conditioned medium
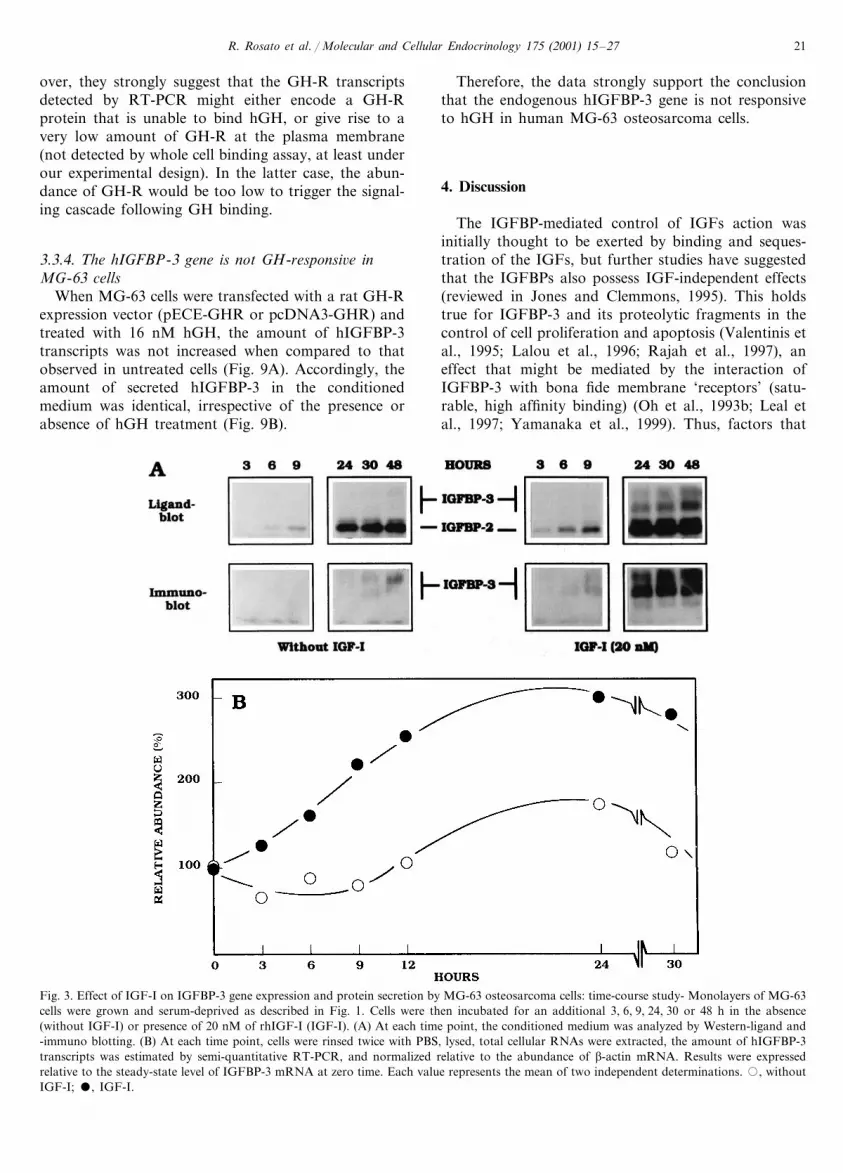
A time course study (3–48 h) was performed foruntreated as well as IGF-I-treated cells. When com-pared to that observed in untreated cells, IGFBP-3accumulation in the conditioned medium was increasedin IGF-I-treated cells. IGFBP-3 was detectable by im-muno-blot as early as 3 h after the onset of IGF-Itreatment (20 nM), and its amount was enhanced by�10–15-fold after 48 h (Fig. 3A, right panels). Bycontrast, the increase with time in secreted IGFBP-3

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–27 19
was fairly low in the medium of untreated cells(IGFBP-3 levels detectable by immunoblotting werenot obtained before 24–30 h) (Fig. 3A, left panels).
Simultaneously, the relative variations in the amountof IGFBP-3 mRNA were assessed by semi-quantitativeRT-PCR (Fig. 3B). In untreated cells, the steady-statelevel of IGFBP-3 mRNA remained almost unchangedover the first 12 h (70–100% of that measured in cells atthe onset of the incubation) and was slightly increasedwith time (e.g. +50% at 24 h; Fig. 3B, open circles). Bycontrast, in treated cells, the steady state amount ofIGFBP-3 transcripts was gradually increased after theonset of IGF-I treatment. The rise in IGFBP-3 mRNAwas detectable as soon as 3 h, (+20%), became moresignificant at 6 h (+53%) and was maximal at 24 h(�2.9-fold-increase) of IGF-I treatment and later (Fig.3B, closed circles).
3.2.2. Lack of effect of IGF-I on IGFBP-3 mRNAhalf-life
MG-63 cells were cultured in the absence or presenceof 20 nM IGF-I for 48 h, and further treated with thepotent transcription inhibitor actinomycin-D (5 �g/ml)for various periods of time (1–24 h). Ethidium bromidestaining of 28S rRNA confirmed that no degradation ofRNA occured, at least during the first 6 h of actino-mycin-D treatment. However, after 24 h, the integrity
of the RNA was affected, as attested by the smearapparent over the 28S rRNA band (Fig. 4).
As expected, the steady-state level of IGFBP-3mRNA was higher (�3.5-fold) in MG-63 cells previ-ously treated for 48 h with 20 nM IGF-I (Fig. 4A, 0 h;i.e. at the onset of actinomycin-D treatment). When thecells were further exposed to actinomycin-D, theamount of IGFBP-3 transcripts decayed slowly overtime, both in untreated and IGF-I-treated cells (Fig.4A). Accordingly, when plotted as percent of controlvs. time (Fig. 4B), the rate of IGFBP-3 mRNA decreasewas similar in untreated (open circles) and in IGF-I-treated cells (closed circles). The estimated IGFBP-3mRNA half-life was not significantly different in theabsence (19.7�5.3 h) or presence of IGF-I (24.8�6.2h) in the culture medium (P�0.2; Student’s t-test).
3.2.3. The synthesis of IGFBP-3 is not stimulated byGH in MG-63 cells
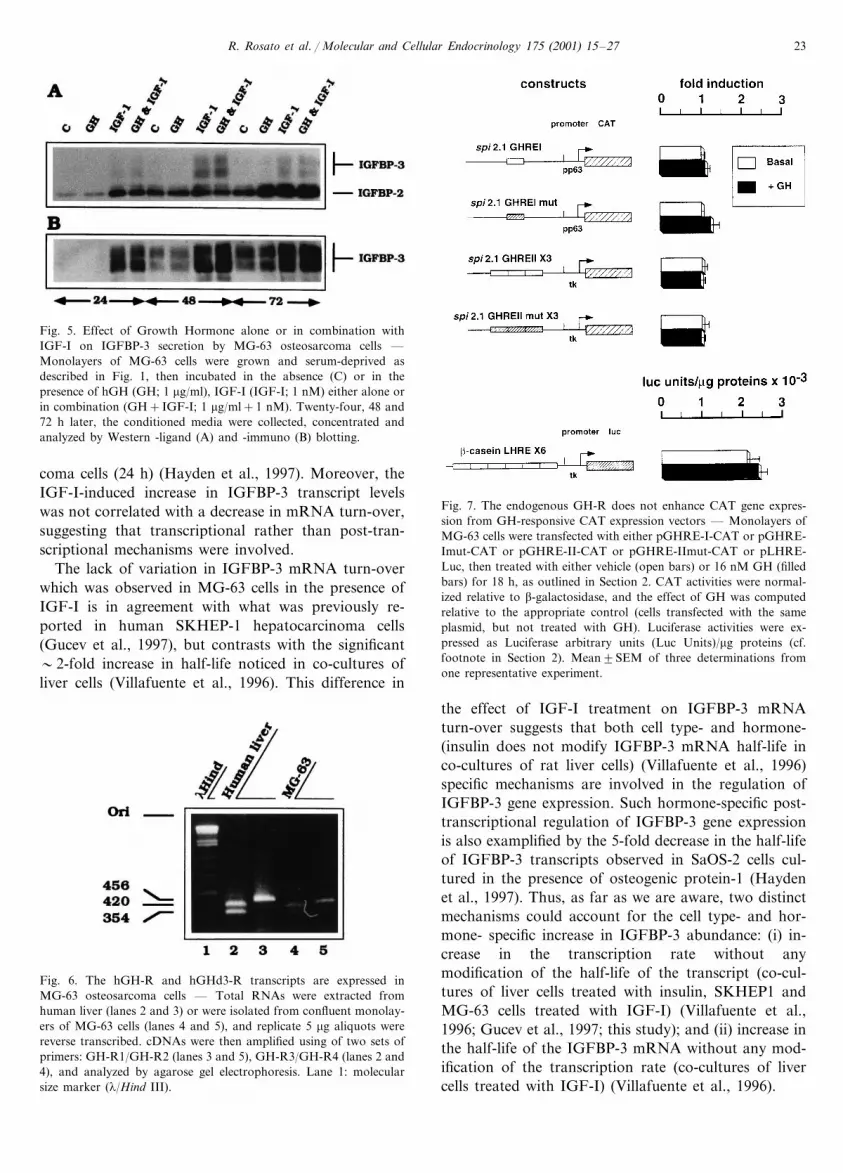
Since GH has been previously shown to mediate theexpression of IGFBP-3 in a IGF-dependent (Langdahlet al., 1998) or -independent manner (Gucev et al.,1997), we addressed the question of whether GH con-trols IGFBP-3 synthesis in MG-63 cells. The amountsof IGFBP-3 that accumulated in the conditionedmedium after 24, 48 or 72 h of treatment with GH wasanalyzed by western ligand- (Fig. 5A) and immuno-
Fig. 1. Effect of IGF-I on IGFBP-3 secretion by MG-63 osteosarcoma cells: dose-response relationship — Monolayers of MG-63 cells were grownto �80% confluency. The culture medium was then replaced by serum-free DMEM, and after an additional 24 h incubation period, theserum-free medium was discarded and replaced by fresh serum-free DMEM containing various concentrations of rhIGF-I. Fourty-eight hourslater, the conditioned medium was collected, concentrated and analyzed by Western -ligand and -immuno blotting. Representative western -ligand(A) and -immuno (B) blots are displayed. Quantitative data were obtained by densitometric scanning of the autoradiograms: (C), Western-ligandblot; (D), Western-immuno blot) (mean of two independent determinations).

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–2720
Fig. 2. Effect of IGF-I on IGFBP-3 gene expression by MG-63osteosarcoma cells: dose-response relationship — Monolayers ofMG-63 cells were grown and treated as described in Fig. 1. Cells werefinally rinsed twice with PBS, lysed, and total cellular RNAs wereextracted. (A) Total RNAs (20 �g/lane) were loaded on 1.2% agarose-formaldehyde gels, blotted onto a nylon membrane, then hybridizedwith a hIGFBP-3 cDNA probe. Ethidium bromide staining of the28S ribosomal RNA was used as a monitor for variations in RNAloading across the lanes. (B) Quantitative data were obtained bydensitometric scanning of the autoradiograms. The abundance ofIGFBP-3 transcripts was normalized relative to 28S rRNA (mean oftwo independent determinations).
456 bp DNA fragment was amplified, identical to thatgenerated from human liver RNA (Fig. 6; comparelanes 3 and 5). Moreover, when RT-PCR was per-formed with a set of primers designed to amplify hGH-R sequences from the beginning of exon 2 down to themiddle of exon 5, two amplification products of 420and 354 bp were generated (Fig. 6; compare lanes 2 and4). They correspond to the amplification of wild-typeand GH-R lacking exon 3 (GH-Rd3) transcripts, re-spectively (Urbanek et al., 1992). However, as attestedby the faint bands resulting from ethidium bromidestaining of the agarose gel, we may consider that hGH-R gene expression is very low in MG-63 cells.
3.3.2. High affinity hGH binding is not detectable bywhole cell binding assay in MG-63 cells
Whether the low amounts of wild-type GH-R orGH-Rd3 transcripts detected in MG-63 cells by RT-PCR yielded detectable amounts of bona fide GH-Rprotein (ie. able to bind GH with high affinity) waschecked in a whole cell binding assay. Incubation ofMG-63 cells with 125I-hGH did not provide any evi-dence for high affinity binding of hGH to MG-63 cells(data not shown). The failure to detect any 125I-hGHbinding was consistently observed over four indepen-dent experiments. Since only low amounts of GH-Rtranscripts were detected in MG-63 cells, we may expectthe GH-R protein to be expressed at a very low level.This was further assessed and confirmed by westernblotting (our unpublished observation).
3.3.3. The GH transduction pathway is functional inMG-63 cells
Transient transfection experiments were performedwith various GH-responsive reporter constructs, tocheck whether or not the GH-R transduction pathwayswere altered in this cell line. Although reporter geneactivity was controlled by GH-response elements in allthe transfected constructs (i.e. pGHRE-I-CAT orpGHRE-II-CAT or pLHRE-Luc), CAT and Luc activ-ities were identical, regardless of the presence of 16 nMGH in the culture medium (GH/vehicle ratios of1.1, 0.93 and 1.14 for GHRE-I, GHRE-II and LHRE,respectively) (Fig. 7). By contrast, when MG-63 cellswere co-transfected with the same reporter plasmidsand with a rat GH-R expression plasmid (pECE-GHRor pcDNA3-GHR), CAT and Luc activities were sig-nificantly enhanced 2.0-, 6.46- and 2.35-fold, respec-tively, in the presence of GH in the culture medium(Fig. 8). This increase was specific since it was abol-ished when the GH-responsive elements were mutated(pGHRE-Imut and pGHRE-IImut in Fig. 8).
Altogether, these data unambiguously demonstratethat the transduction pathways involved in the tran-scriptional activation of GH-responsive promoters byGH are functional in MG-63 osteosarcoma cells. More-
blotting (Fig. 5B). At each time point, the increase inthe level of secreted IGFBP-3 was identical in controland GH-treated cells. By contrast, when hGH and alow concentration of IGF-I (1 nM) were combined, theamount of secreted IGFBP-3 was significantly en-hanced when compared to control and GH treatedcells. However, at each time point, the magnitude of theeffect was identical to that observed in presence of 1nM IGF-I alone. To further investigate the issue of GHinsensitivity, we followed the expresssion of growthhormone receptor (GH-R) and the GH-induced tran-scription of GH-responsive gene constructs in MG-63cells.
3.3. While the le�el of GH-R is low, the GHtransduction pathway is functional in MG-63 cells
3.3.1. Two isoforms of GH-R transcripts are expressedin MG-63 cells
When total cellular RNA extracted from MG-63 cellswas analyzed by RT-PCR with oligonucleotide primersspanning the trans-membrane domain of the hGH-R, a
R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–27 21
over, they strongly suggest that the GH-R transcriptsdetected by RT-PCR might either encode a GH-Rprotein that is unable to bind hGH, or give rise to avery low amount of GH-R at the plasma membrane(not detected by whole cell binding assay, at least underour experimental design). In the latter case, the abun-dance of GH-R would be too low to trigger the signal-ing cascade following GH binding.
3.3.4. The hIGFBP-3 gene is not GH-responsi�e inMG-63 cells
When MG-63 cells were transfected with a rat GH-Rexpression vector (pECE-GHR or pcDNA3-GHR) andtreated with 16 nM hGH, the amount of hIGFBP-3transcripts was not increased when compared to thatobserved in untreated cells (Fig. 9A). Accordingly, theamount of secreted hIGFBP-3 in the conditionedmedium was identical, irrespective of the presence orabsence of hGH treatment (Fig. 9B).
Therefore, the data strongly support the conclusionthat the endogenous hIGFBP-3 gene is not responsiveto hGH in human MG-63 osteosarcoma cells.
4. Discussion
The IGFBP-mediated control of IGFs action wasinitially thought to be exerted by binding and seques-tration of the IGFs, but further studies have suggestedthat the IGFBPs also possess IGF-independent effects(reviewed in Jones and Clemmons, 1995). This holdstrue for IGFBP-3 and its proteolytic fragments in thecontrol of cell proliferation and apoptosis (Valentinis etal., 1995; Lalou et al., 1996; Rajah et al., 1997), aneffect that might be mediated by the interaction ofIGFBP-3 with bona fide membrane ‘receptors’ (satu-rable, high affinity binding) (Oh et al., 1993b; Leal etal., 1997; Yamanaka et al., 1999). Thus, factors that
Fig. 3. Effect of IGF-I on IGFBP-3 gene expression and protein secretion by MG-63 osteosarcoma cells: time-course study- Monolayers of MG-63cells were grown and serum-deprived as described in Fig. 1. Cells were then incubated for an additional 3, 6, 9, 24, 30 or 48 h in the absence(without IGF-I) or presence of 20 nM of rhIGF-I (IGF-I). (A) At each time point, the conditioned medium was analyzed by Western-ligand and-immuno blotting. (B) At each time point, cells were rinsed twice with PBS, lysed, total cellular RNAs were extracted, the amount of hIGFBP-3transcripts was estimated by semi-quantitative RT-PCR, and normalized relative to the abundance of �-actin mRNA. Results were expressedrelative to the steady-state level of IGFBP-3 mRNA at zero time. Each value represents the mean of two independent determinations. �, withoutIGF-I; �, IGF-I.

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–2722
Fig. 4. Effect of IFG-I on IGFBP-3 mRNA stability — Monolayers of MG-63 cells were grown and serum-deprived as described in Fig. 1, thenincubated for 48 h in the absence or presence of 20 nM IGF-I. Fourty-eight hours after the onset of IGF-I treatment, actinomycin-D (5 �g/ml)was added to the serum-free medium, and total cellular RNAs were extracted at 0, 1, 2, 4, 6, and 24 h after the actinomycin-D treatment wasinitiated. (A) Total RNAs (20 �g/lane) were loaded on 1.2% agarose-formaldehyde gels, blotted onto a nylon membrane, then hybridized with ahIGFBP-3 cDNA probe. Ethidium bromide staining of the 28S ribosomal RNA was used as a monitor for variations in RNA loading across thelanes. The data obtained with two independent culture dishes are shown. (− ) and (+ ), culture in the absence and presence of rhIGF-I,respectively; the duration of the incubation period with actinomycin-D is indicated below the lanes; (28S) ethidium bromide staining of 28Sribosomal RNAs. (B) Quantitative data obtained by densitometric scanning of the autoradiograms. IGFBP-3 transcripts were normalized relativeto 28S rRNA, and at each time point their relative abundance was expressed as a percentage of that measured at zero time (mean of twoindependent determinations). �, without IGF-I; �, with IGF-I.
influence both the availability and post-translationalmodification(s) of IGFBP-3 in the extracellular fluids(e.g. gene expression, binding to the extracellular ma-trix, proteolysis) are likely to play key roles in regulat-ing the magnitude of its biological effects. In thisconnection, we may speculate that in vivo, the secretionof IGFBP-3 by osteoblast-like cells contributes to theparacrine control of osteoblast proliferation by bothIGF-I- dependent (paracrine control of IGFsbioavailability) and -independent (the limited proteoly-sis of IGFBP-3 in the extracellular fluid) mechanisms,and thus promotes the induction of bone-specificmarker expression, as already shown ex vivo with MG-63 osteosarcoma cells in culture (Velez-Yanguas et al.,1996). A role for IGFBP-3 in the control of osteoblastproliferation is further supported by our unpublishedobservation that IGFBP-3 transcripts were more abun-dant in nearly confluent monolayers of MG-63 cells(low rate of proliferation), than in low density cultures(exponential growth)(A.G., unpublished observation).
In the present study, we have focused on the mecha-nisms involved in the regulation of IGFBP-3 gene
expression by IGF-I and GH, using MG-63 osteosar-coma cells as a model system. The MG-63 cell lineexpresses a low level of IGFBP-3, reflected in thetranscript and protein levels. IGF-I significantly ele-vated transcript levels and to an even greater extentincreased protein levels in the culture medium, with aprogressive time course. These results are in agreementwith the rapid rise in IGFBP-3 transcripts abundanceobserved in co-cultures of rat hepatocytes and of nor-mal hepatic sinusoidal cells in the presence of 1 �Minsulin (a supra-physiologic concentration at which in-sulin acts, at least partially, through the IGF-IR) (Villa-fuente et al., 1996). By contrast, the IGF-I-inducedincrease in IGFBP-3 mRNA was delayed (12 h) inSKHEP-1 human hepatocarcinoma cells, a cell line thatotherwise exhibited a sooner GH-induced increase inIGFBP-3 transcripts (6 h) (Gucev et al., 1997).
In MG-63 cells, the half-life of IGFBP-3 transcripts(�20 h) was close to that determined under basalculture conditions both in co-cultures of liver cells(17–18 h) (Villafuente et al., 1996), in SKHEP-1 cells(14–18 h)(Gucev et al., 1997) and in SaOS-2 osteosar-
R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–27 23
Fig. 5. Effect of Growth Hormone alone or in combination withIGF-I on IGFBP-3 secretion by MG-63 osteosarcoma cells —Monolayers of MG-63 cells were grown and serum-deprived asdescribed in Fig. 1, then incubated in the absence (C) or in thepresence of hGH (GH; 1 �g/ml), IGF-I (IGF-I; 1 nM) either alone orin combination (GH+IGF-I; 1 �g/ml+1 nM). Twenty-four, 48 and72 h later, the conditioned media were collected, concentrated andanalyzed by Western -ligand (A) and -immuno (B) blotting.
Fig. 7. The endogenous GH-R does not enhance CAT gene expres-sion from GH-responsive CAT expression vectors — Monolayers ofMG-63 cells were transfected with either pGHRE-I-CAT or pGHRE-Imut-CAT or pGHRE-II-CAT or pGHRE-IImut-CAT or pLHRE-Luc, then treated with either vehicle (open bars) or 16 nM GH (filledbars) for 18 h, as outlined in Section 2. CAT activities were normal-ized relative to �-galactosidase, and the effect of GH was computedrelative to the appropriate control (cells transfected with the sameplasmid, but not treated with GH). Luciferase activities were ex-pressed as Luciferase arbitrary units (Luc Units)/�g proteins (cf.footnote in Section 2). Mean�SEM of three determinations fromone representative experiment.
coma cells (24 h) (Hayden et al., 1997). Moreover, theIGF-I-induced increase in IGFBP-3 transcript levelswas not correlated with a decrease in mRNA turn-over,suggesting that transcriptional rather than post-tran-scriptional mechanisms were involved.
The lack of variation in IGFBP-3 mRNA turn-overwhich was observed in MG-63 cells in the presence ofIGF-I is in agreement with what was previously re-ported in human SKHEP-1 hepatocarcinoma cells(Gucev et al., 1997), but contrasts with the significant�2-fold increase in half-life noticed in co-cultures ofliver cells (Villafuente et al., 1996). This difference in
the effect of IGF-I treatment on IGFBP-3 mRNAturn-over suggests that both cell type- and hormone-(insulin does not modify IGFBP-3 mRNA half-life inco-cultures of rat liver cells) (Villafuente et al., 1996)specific mechanisms are involved in the regulation ofIGFBP-3 gene expression. Such hormone-specific post-transcriptional regulation of IGFBP-3 gene expressionis also examplified by the 5-fold decrease in the half-lifeof IGFBP-3 transcripts observed in SaOS-2 cells cul-tured in the presence of osteogenic protein-1 (Haydenet al., 1997). Thus, as far as we are aware, two distinctmechanisms could account for the cell type- and hor-mone- specific increase in IGFBP-3 abundance: (i) in-crease in the transcription rate without anymodification of the half-life of the transcript (co-cul-tures of liver cells treated with insulin, SKHEP1 andMG-63 cells treated with IGF-I) (Villafuente et al.,1996; Gucev et al., 1997; this study); and (ii) increase inthe half-life of the IGFBP-3 mRNA without any mod-ification of the transcription rate (co-cultures of livercells treated with IGF-I) (Villafuente et al., 1996).
Fig. 6. The hGH-R and hGHd3-R transcripts are expressed inMG-63 osteosarcoma cells — Total RNAs were extracted fromhuman liver (lanes 2 and 3) or were isolated from confluent monolay-ers of MG-63 cells (lanes 4 and 5), and replicate 5 �g aliquots werereverse transcribed. cDNAs were then amplified using of two sets ofprimers: GH-R1/GH-R2 (lanes 3 and 5), GH-R3/GH-R4 (lanes 2 and4), and analyzed by agarose gel electrophoresis. Lane 1: molecularsize marker (�/Hind III).

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–2724
Finally, it should be stressed that treatment of MG-63 cells with IGF-I yielded a �3–5-fold smaller in-crease in IGFBP-3 mRNA levels than in the amount ofIGFBP-3 protein released in the conditioned medium,arguing for an IGF-I-induced increase in IGFBP-3stability (e.g. decreased IGFBP-3 proteolysis in theconditioned medium), and/or an IGF-I-induced dissoci-ation of membrane-bound IGFBP-3, and/or a generalincrease in protein synthesis in response to IGF-I treat-ment. According to Lalou et al. (1994), the first hypoth-esis seems very unlikely. Their data showing thatIGFBP-3 was not proteolysed in the conditionedmedium of MG-63 cells whether the cells were culturedor not in the presence of IGF-I support the conclusionthat the effect of IGF-I was not through protection ofIGFBP-3 against intrinsic protease activity. The secondhypothesis also seems unlikely. On the one hand, thetime-course for IGFBP-3 secretion in the conditioned
Fig. 9. The endogenous IGFBP-3 gene is not responsive to hGH inMG-63 cells — Monolayers of MG-63 cells were transfected withpECE-GHR (5 �g per well), then were grown in serum-deprivedmedium. After 16 h, the serum-free medium was renewed and thecells were treated with either vehicle or GH for an additional 48 h.(A) The cell monolayers were rinsed twice with PBS, lysed, and totalcellular RNAs were extracted. Total RNAs (20 �g/lane) were loadedon 1.2% agarose-formaldehyde gels, blotted onto a nylon membrane,then hybridized with a hIGFBP-3 cDNA probe. After visualization ofthe hIGFBP-3 transcripts (upper panel), blots were stripped, thenre-hybridized with a [32P]-labeled GAPDH probe (lower panel). (B)The conditioned serum-free medium was collected, concentrated andanalyzed by Western-immuno blotting.
Fig. 8. The post-GH-R signal transduction pathway is functional inMG-63 cells — Monolayers of MG-63 cells were co-transfected withpECE-GHR or pcDNA3-GHR and with either pGHRE-I-CAT orpGHRE-Imut-CAT or pGHRE-II-CAT or pGHRE-IImut-CAT orpLHRE-Luc, then treated with either vehicle (open bars) or GH(filled bars) for 18 h, as outlined in Section 2. CAT activities werenormalized relative to �-galactosidase, and the effect of GH wascomputed relative to the appropriate control (cells transfected withthe same series of plasmids, but not treated with GH). Luciferaseactivities were expressed as Luciferase arbitrary units (Luc Units)/�gproteins (cf. footnote in Section 2). Mean�SEM of three determina-tions from one representative experiment.
medium, and that observed for IGFBP-3 transcriptswere similar. On the other hand, in numerous studiesand especially in one of the most recent and compre-hensive ones by Maile et al. (1999), it was reported thatthe release of membrane-associated IGFBP-3 onlyyielded proteolytic fragments, and not intact, non-pro-teolysed IGFBP-3 in the conditioned medium. This isdue to the proteolytic cleavage of the membrane-boundIGFBP-3 by membrane-associated protease (s). In thisconnection, since not any trace of IGFBP-3 proteolyticfragments was detected in MG-63 cells conditionedmedium (irrespective of the presence or absence ofIGF-I treatment), our data do not support the possibil-ity that the release of IGFBP-3 (bound to the mem-brane of MG-63 cells at the onset of the treatment)could account for the accumulation of IGFBP-3 in theconditioned medium.
It was previously reported that the amount ofIGFBP-3 that accumulates in the conditioned mediumof INS-1 (De et al., 1995), of SKHEP-1 (Gucev et al.,1997) and of osteoblastic cells (Ernst and Rodan, 1990)was enhanced by GH treatment. This increase (i) wascorrelated with a rise in the abundance of IGFBP-3mRNA (De et al., 1995; Gucev et al., 1997); (ii) wasdependent on GH concentration in the culture medium(De et al., 1995); (iii) was not mediated by IGF-I or -II(De et al., 1995); and (iv) was not associated withstabilization of the transcript (Gucev et al., 1997).Hence, this increase appears to be dependent on thestimulation of IGFBP-3 gene transcription by GH.

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–27 25
Such GH-dependent increase in IGFBP-3 gene tran-scription may rely on the presence of bona fide STAT-binding site(s) in the rat and human IGFBP-3 gene,respectively. Indeed, one such site was localized withinthe 5�-flanking sequences of the human gene (A.G.,unpublished observation).
By contrast, GH did not increase the amount ofsecreted IGFBP-3 by MG-63 cells. This was not due tothe lack of hGH-R gene expression (transcripts forboth wild-type and hGHRd3 isoforms of the hGH-Rwere detected by RT-PCR analysis, albeit their amountwas fairly low), but may arise from the fact that thesetranscripts did not give rise to a sufficient level of bonafide hGH-R protein at the cell surface. Indeed, thenumber of GH binding sites was below the threshold ofthe assay and was not detected by western-blot (K.G.and L.M., unpublished observation). Alternatively, wecannot exclude: (i) the absence of translation of abnor-mal GH-R transcripts; and (ii) that the putative en-dogenous hGH-R expressed in MG-63 cells is notcorrectly routed to the plasma membrane, or corre-spond to a form mutated in critical regions (e.g. thatimpedes hGH binding or GH-R dimerization). ThatMG-63 cells do not respond to GH was further sup-ported by the observation that neither the expression ofan endogenous gene (c-fos)(data not shown), nor theactivity of transfected GH-responsive reporter geneswere enhanced when the cells were treated with GH.This held true regardless of the specificity of the cis-GHResponse Elements.
However, in MG-63 cells, the absence of increase inendogenous IGFBP-3 gene expression following GHtreatment is not exclusively due to an impaired GH-Rfunction (it was also observed when wild-type rat GH-R was over-expressed in the cells by transfection), butprobably results from the fact that the IGFBP-3 gene isnot GH-responsive in MG-63 cells.
Acknowledgements
The authors are indebted to Alphonse Le Cam andJean Djiane for critical reading of the manuscript. Thiswork was supported by the Association pour laRecherche contre le Cancer (ARC) and the LigueFrancaise contre le Cancer and from the EEC- (Humancapital and Mobility, contract N° CHRX-CT-94-00556).
References
Aubin, R., Menard, P., Lajeunesse, D., 1996. Selective effect ofthiazides on the human osteoblast-like cell line MG-63. KidneyInt. 50, 1476–1482.
Bale, L.K., Conover, C., 1992. Regulation of insulin-like growthfactor binding protein-3 messenger ribonucleic acid expression byinsulin-like growth factor I. Endocrinology 131, 608–614.
Birch, M.A., Skerry, T.M., 1999. Differential regulation of syndecanexpression by osteosarcoma cell lines in response to cytokines butnot osteotropic hormones. Bone 24, 571–578.
Birnbaum, R.S., Wiren, K.M., 1994. Changes in insulin-like growthfactor-binding protein expression and secretion during the prolif-eration, differentiation, and mineralization of primary cultures ofrat osteoblasts. Endocrinology 135, 223–230.
Canalis, E., Gabbitas, B., 1994. Bone morphogenetic protein 2 in-creases insulin-like growth factor I and II transcripts and polypep-tide levels in bone cell cultures. J. Bone Miner. Res. 9, 1999–2005.
Canalis, E., Gabbitas, B., 1995. Skeletal growth factors regulate thesynthesis of insulin-like growth factor binding protein-5 in bonecell cultures. J. Biol. Chem. 270, 10 771–10 776.
Centrella, M., Mc Carthy, T.L., Canalis, E., 1990. Receptors forinsulin-like growth factors-I and II in osteoblasts enriched cul-tures from fetal rat bone. Endocrinology 126, 39–44.
Chen, C., Okayama, H., 1987. High-efficiency transformation ofmammalian cells by plasmid DNA. Mol. Cell. Biol. 7, 2745–2752.
Clover, J., Gowen, M., 1994. Are MG-63 and HOS TE85 humanosteosarcoma cell lines representative models of the osteoblasticphenotype? Bone 15, 585–591.
De, W., Breant, B., Czernikow, P., Asfari, M., 1995. Growth hor-mone (GH) and prolactin (PRL) regulate IGFBP-3 gene expres-sion in rat b-cells. Mol. Cell. Endocrinol. 114, 43–50.
Delany, A.M., Canalis, E., 1996. Transcriptional repression of in-sulin-like growth factor I by glucocorticoids in rat bone cells.Endocrinology 136, 4776–4781.
Dinerstein, H., Lago, F., Goujon, L., Ferrag, F., Esposito, N.,Finidori, J., Kelly, P.A., Postel-Vinay, M.C., 1995. The prolinerich region of the growth hormone receptor is essential for Jak2phosphorylation, activation of cell proliferation and gene tran-scription. Mol. Endocrinol. 9, 1701–1707.
Ernst, M., Rodan, G.A., 1990. Increased activity of insulin-likegrowth factor (IGF) in osteoblastic cells in the presence of growthhormone (GH): positive correlation with the presence of GH-in-duced IGF-Binding Protein-3. Endocrinology 127, 807–814.
Furlanetto, R.W., Harwell, S.E., Frick, K.K., 1994. Insulin-likegrowth factor-I induces cyclin-D1 expression in MG-63 humanosteosarcoma cells in vitro. Mol. Endocrinol. 8, 510–517.
Gucev, Z.S., Oh, Y., Kelley, K.M., Labarta, J.I., Worwerk, P.,Rosenfeld, R.G., 1997. Evidence for insulin-like growth factor(IGF)-independent transcriptional regulation of IGF-bindingprotein-3 by growth hormone in SKHEP-1 human hepatocar-cinoma cells. Endocrinology 138, 1464–1470.
Hayden, J.M., Strong, D.D., Baylink, D.J., Powell, D.R., Sampath,T.K., Mohan, S., 1997. Osteogenic-protein-1 stimulates produc-tion of insulin-like growth factor-binding protein-3 nuclear tran-scripts in human osteosarcoma cells. Endocrinology 138,4240–4247.
Jones, J.I., Clemmons, D.R., 1995. Insulin-like growth factors andtheir binding proteins: biological actions. Endocr. Rev. 16, 3–34.
Kanety, H., Karisik, A., Klinger, B., Silbergeld, A., Laron, Z., 1993.Long-term treatment of Laron-type dwarfs with insulin-likegrowth factor-I increases insulin-like growth factor-bindingprotein-3 in the absence of growth hormone. Acta Endocrinol.(Copenh.) 128, 144–149.
Kassem, M., Okazaki, R., De-Leon, D., Harris, S.A., Robinson, J.A.,Spelsberg, T.C., Conover, C.A., Riggs, B.L., 1996. Potentialmechanism of estrogen-mediated decrease in bone formation:estrogen increases production of inhibitory insulin-like growthfactor-binding protein-4. Proc. Assoc. Am. Phys. 108, 155–164.
Kassem, M., 1997. Cellular and molecular effects of growth hormoneand estrogen on human bone cells. APMIS Suppl. 105, 7–30.

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–2726
Knutsen, R., Honda, Y., Strong, D.D., Sampath, T.K., Baylink, D.J.,Mohan, S., 1995. Regulation of insulin-like growth factor systemcomponents by osteogenic protein-1 in human bone cells. En-docrinology 136, 857–865.
Kobayashi, T., Sugimoto, T., Saijoh, K., Fukase, M., Chihara, K.,1994. Calcitonin directly acts on mouse osteoblastic MC3T3-E1cells to stimulate mRNA expression of c-fos, insulin-like growthfactor-1 and osteoblastic phenotypes (type 1 collagen and os-teocalcin). Biochem. Biophys. Res. Commun. 199, 876–880.
Lalou, C., Binoux, M., 1993. Evidence that limited proteolysis ofinsulin-like growth factor binding protein-3 (IGFBP-3) occurs innormal state outside of the bloodstream. Regul. Pept. 48, 179–188.
Lalou, C., Lassarre, C., Binoux, M., 1996. A proteolytic fragment ofIGFBP-3 that fails to bind IGFs inhibits IGF and insulin mito-genic effects. Endocrinology 137, 3206–3212.
Lalou, C., Silve, C., Rosato, R., Segovia, B., Binoux, M., 1994.Interactions between insulin-like growth factor-I (IGF-I) and thesystem of plasminogen activators and their inhibitors in thecontrol of IGF-binding protein-3 production and proteolysis inhuman osteosarcoma cells. Endocrinology 135, 2318–2326.
Langdahl, B.L., Kassem, M., Møller, M.K., Eriksen, E.F., 1998. Theeffects of IGF-I and IGF-II on proliferation and differentiation ofhuman osteoblasts and interaction with growth hormone. Eur. J.Clin. Invest. 28, 176–183.
Leal, S.M., Liu, Q., Huang, H.H., Huang, J.S., 1997. The type Vtransforming growth factor b receptor is the putative insulin-likegrowth factor-binding protein-3 receptor. J. Biol. Chem. 272,20 572–20 576.
Legraverend, C., Simar-Blanchet, A.E., Paul, C., Sotiropoulos, A.,Finidori, J., LeCam, A., 1996. A novel growth hormone responseelement unrelated to STAT (signal transducer and activator oftranscription)-binding site is a bifunctional enhancer. Mol. En-docrinol. 10, 1507–1518.
Lopaczynski, W., Harris, S., Nissley, P., 1993. Insulin-like growthfactor-I (IGF-I) dependent phosphorylation of the IGF-I receptorin MG-63 cells. Regul. Pept. 48, 207–216.
Maile, L.A., Gill, Z.P., Perks, C.M., Holly, J.M., 1999. The role ofcell surface attachment and proteolysis in the insulin-like growthfactor (IGF)-independent effects of IGF-binding protein-3 onapoptosis in breast epithelial cells. Endocrinology 140, 4040–4045.
Mc Carthy, T.L., Casinghino, S., Centrella, M., Canalis, E., 1994.Complex pattern of insulin-like growth factor binding proteinexpression in primary rat osteoblast enriched cultures: regulationby prostaglandin E2, growth hormone, and the insulin-likegrowth factors. J. Cell Physiol. 160, 163–175.
McCarthy, T.L., Centrella, M., Canalis, E., 1989. Regulatory effectsof insulin-like growth factors I and II on bone collagen synthesisin rat calvaria cultures. Endocrinology 124, 301–309.
Mehlus, H., Ljunghall, S., 1996. Growth hormone and insulin-likegrowth factor-I do not activate identical genes in normal humanosteoblast. Biochem. Mol. Biol. Int. 38, 425–428.
Mohan, S., Strong, D.D., Lempert, U.G., Tremollieres, F., Wergedal,J.E., Baylink, D.J., 1992. Studies on the regulation of insulin-likegrowth factor binding protein (IGFBP)-3 and IGFBP-4 produc-tion in human bone cells. Acta Endocrinol. (Copenh.) 127, 555–564.
Oh, Y., Muller, H.L., Pham, H., Rosenfeld, R.G., 1993a. Demonstra-tion of receptors for insulin-like growth factor binding protein-3on Hs578T human breast cancer cells. J. Biol. Chem. 268, 26 045–26 048.
Oh, Y., Muller, H.L., Neely, E.K., Lamson, G., Rosenfeld, R.G.,1993b. New concepts in insulin-like growth factor receptor physi-ology. Growth Regul. 3, 113–123.
Ohlsson, C., Bengtsson, B.-A� ., Isaksson, O.G.P., Andreassen, T.T.,Slootweg, M.C., 1998. Growth hormone and bone. Endocr. Rev.19, 55–79.
Okasaki, R., Conover, C.A., Harris, S.A., Spelsberg, T.C., Riggs,B.L., 1995. Normal human osteoblast-like cells consistently ex-press genes for insulin-like growth factor I and II, but trans-formed human osteoblast cell lines do not. J. Bone Miner. Res.10, 788–794.
Paquereau, L., Le Cam, A., 1992. Electroporation-mediated genetransfer into hepatocytes: preservation of a growth hormoneresponse. Anal. Biochem. 204, 147–151.
Raile, K., Hoflich, A., Kessler, U., et al., 1994. Human osteosarcomacells (U-2 OS) cells express both insulin-like growth factor-I(IGF-I) receptors and insulin-like growth factor-II/mannose-6phosphate (IGF-II/M6P) receptors and synthetise IGF-II. Au-tocrine growth stimulation by IGF-II via IGF-I receptor. J. CellPhysiol. 159, 531–541.
Rajah, R., Valentinis, B., Cohen, P., 1997. Insulin-like growth factor(IGF)-binding protein-3 induces apoptosis and mediates the ef-fects of transforming growth factor-�1 on programmed cell deaththrough a p53 and IGF-independent mechanism. J. Biol. Chem.272, 12 181–12 188.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning:A Laboratory Manual, 2nd. Cold Spring Harbor LaboratoryPress.
Schmid, C., 1993. IGFs: function and clinical importance. 2 Theregulation of osteoblast function by hormones and cytokines withspecial reference to insulin-like growth factors and their bindingproteins. J. Intern. Med. 234, 535–542.
Schmid, C., Schlapfer, I., Peter, M., Boni-Schnetzler, M., Schwander,J., Zapf, J., Froesch, E.R., 1994. Growth hormone and parathy-roid hormone stimulate IGFBP-3 in rat osteoblasts. Am. J. Phys-iol. 267, E226–E233.
Sotiropoulos, A., Moutoussamy, S., Renaudie, F., Clauss, M., Kay-ser, C., Gouilleux, F., Kelly, P.A., Finidori, J., 1996. Differentialactivation of STAT3 and STAT5 by distinct regions of the growthhormone receptor. Mol. Endocrinol. 10, 998–1006.
Strom, P., Ljunghall, S., Melhus, H., 1994. Adult human osteoblast-like cells do not express insulin-like growth factor-I. Biochem.Biophys. Res. Commun. 199, 78–82.
Thrailkill, K.M., Siddhanti, S.R., Fowlkes, J.L., Quarles, L.D., 1996.Differentiation of MC3T3-E1 osteoblasts is associated with tem-poral changes in the expression of IGF-I and IGFBPs. Bone 17,307–313.
Urbanek, M., MacLeod, J.N., Cooke, N.E., Liebhaber, S.A., 1992.Expression of human Growth Hormone (hGH) receptor isoformis predicted dy tissue-specific alternative splicing of exon 3 of theGH receptor gene transcript. Mol. Endocrinol. 6, 279–287.
Valentinis, B., Bhala, A., DeAngelis, T., Baserga, R., Cohen, P.,1995. The human insulin-like growth factor (IGF) bindingprotein-3 inhibits the growth of fibroblasts with a targeted disrup-tion of the IGF-I receptor gene. Mol. Endocrinol. 9, 361–367.
Velez-Yanguas, M.C., Kalebic, T., Maggi, M., Kappel, C.C., Lette-rio, J., Uskokovic, M., Helman, L.J., 1996. (1a,25)-dihydroxy-16-ene-23-yne-26,27-hexafluorocholecalciferol (Ro24-5531)modulation of insulin-like growth factor-binding protein-3 andinduction of differentiation and growth arrest in a human os-teosarcoma cell line. J. Clin. Endocrinol. Metab. 81, 93–99.
Villafuente, B.C., Zhang, W.-N., Phillips, L.S., 1996. Insulin-likegrowth factor-I regulate hepatic insulin-like growth factor bindingprotein-3 by different mechanisms. Mol. Endocrinol. 10, 622–630.
Wakao, Y., Hilliker, S., Baylink, D.J., Mohan, S., 1994. Studies onthe regulation of insulin-like growth factor binding protein-3secretion in human osteosarcoma cells in vitro. J. Bone Miner.Res. 9, 865–872.
Wergedal, J.E., Mohan, S., Lundy, M., Baylink, D.J., 1990. Skeletalgrowth factor and other growth factors known to be present inbone marix stimulate proliferation and protein synthesis in humanbone cells. J. Bone Miner. Res. 5, 179–186.

R. Rosato et al. / Molecular and Cellular Endocrinology 175 (2001) 15–27 27
Westley, B.R., May, F.E., 1994. Role of insulin-like growth factors insteroid modulated proliferation. J. Steroid Biochem. Mol. Biol.51, 1–9.
Yamanaka, Y., Fowlkes, J.L., Wilson, E.M., Rosenfeld, R.G., Oh,Y., 1999. Characterization of insulin-like growth factor bindingprotein-3 (IGFBP-3) binding to human breast cancer cells: kinet-ics of IGFBP-3 binding and identification of receptor bindingdomain on the IGFBP-3 molecule. Endocrinology 140, 1319–1328.
Yeh, L.C., Adamo, M.L., Kitten, A.M., Olson, M.S., Lee, J.C., 1996.Osteogenic protein-1-mediated insulin-like growth factor gene ex-pression in primary cultures of rat osteoblastic cells. Endocrinol-ogy 137, 1921–1931.
Zapf, J., Haury, C., Waldvogel, M., Futo, E., Hasler, H., Binz, K.,Guler, H.P., Schmid, C., Froesch, E.R., 1989. Recombinant in-sulin-like growth factor-I induces its own specific carrier proteinin hypophysectomized and diabetic rats. Proc. Natl. Acad. Sci.USA 86, 3813–3817.
Zezulak, K.M., Green, H., 1986. The generation of insulin-likegrowth factor-I-sensitive cells by growth hormone action. Science233, 551–553.
Zhou, Y., Mohan, S., Linkhart, T.A., Baylink, D.J., Strong, D.D.,1996. Retinoic acid regulates insulin-like growth factor-bindingprotein expression in human osteoblast cells. Endocrinology 137,975–983.
.




















