
The herpes simplex virus 1 latency-associated transcript promotes functional exhaustion of...
12
JOURNAL OF VIROLOGY, Sept. 2011, p. 9127–9138 Vol. 85, No. 17 0022-538X/11/$12.00 doi:10.1128/JVI.00587-11 Copyright © 2011, American Society for Microbiology. All Rights Reserved. The Herpes Simplex Virus 1 Latency-Associated Transcript Promotes Functional Exhaustion of Virus-Specific CD8 T Cells in Latently Infected Trigeminal Ganglia: a Novel Immune Evasion Mechanism Aziz A. Chentoufi, 1 † Elizabeth Kritzer, 1 Michael V. Tran, 1 Gargi Dasgupta, 1 Chang Hyun Lim, 1 David C. Yu, 1 Rasha E. Afifi, 1 Xianzhi Jiang, 1,2 Dale Carpenter, 2 Nelson Osorio, 1,2 Chinhui Hsiang, 1,2 Anthony B. Nesburn, 1 Steven L. Wechsler, 1,2,4 and Lbachir BenMohamed 1,2,3,5 * Laboratory of Cellular and Molecular Immunology, The Gavin Herbert Eye Institute and Department of Ophthalmology, University of California Irvine, School of Medicine, Irvine, California 92697 1 ; Virology Research, The Gavin Herbert Eye Institute and Department of Ophthalmology, University of California Irvine, School of Medicine, Irvine, California 92697 2 ; Chao Family Comprehensive Cancer Center, University of California Irvine Medical Center, Irvine, California 92868-3201 3 ; Department of Microbiology and Molecular Genetics and Center for Virus Research, University of California Irvine, Irvine, California 92697 4 ; and Institute for Immunology, University of California Irvine, School of Medicine, Irvine, California 92697 5 Received 23 March 2011/Accepted 3 June 2011 Following ocular herpes simplex virus 1 (HSV-1) infection of C57BL/6 mice, HSV-specific (HSV-gB 498–505 tetramer ) CD8 T cells are induced, selectively retained in latently infected trigeminal ganglia (TG), and appear to decrease HSV-1 reactivation. The HSV-1 latency-associated transcript (LAT) gene, the only viral gene that is abundantly transcribed during latency, increases reactivation. Previously we found that during latency with HSV-1 strain McKrae-derived viruses, more of the total TG resident CD8 T cells expressed markers of exhaustion with LAT virus compared to LAT virus. Here we extend these findings to HSV-1 strain 17syn-derived LAT and LAT viruses and to a virus expressing just the first 20% of LAT. Thus, the previous findings were not an artifact of HSV-1 strain McKrae, and the LAT function involved mapped to the first 1.5 kb of LAT. Importantly, to our knowledge, we show here for the first time that during LAT virus latency, most of the HSV-1-specific TG resident CD8 T cells were functionally exhausted, as judged by low cytotoxic function and decreased gamma interferon (IFN-) and tumor necrosis factor alpha (TNF-) pro- duction. This resulted in LAT TG having more functional HSV-gB 498–505 tetramer CD8 T cells compared to LAT TG. In addition, LAT expression, in the absence of other HSV-1 gene products, appeared to be able to directly or indirectly upregulate both PD-L1 and major histocompatibility complex class I (MHC-I) on mouse neuroblastoma cells (Neuro2A). These findings may constitute a novel immune evasion mechanism whereby the HSV-1 LAT directly or indirectly promotes functional exhaustion (i.e., dysfunction) of HSV- specific CD8 T cells in latently infected TG, resulting in increased virus reactivation. Herpes simplex virus 1 (HSV-1) establishes lifelong latency in its host. After a primary acute infection of the eye, HSV-1 travels up sensory neurons to the trigeminal ganglia (TG), where it maintains latent infections (2, 16, 18, 42, 48). Recur- rent corneal disease results from the reactivation of latent virus from neurons of the TG, anterograde transportation to nerve termini, and reinfection of the cornea (2, 42). In addition to latency-associated transcript (LAT) and other viral factors, recent reports suggest that CD8 T cells in the TG may con- tribute to the establishment, maintenance, and/or reactivation of latent HSV-1 in TG (2, 42). However, the mechanisms remain to be fully elucidated. During HSV-1 latency, the HSV-1 latency-associated tran- script (LAT) gene is the only viral gene that is consistently detected to be abundantly transcribed. LAT viruses (i.e., wild-type [wt] HSV-1) have a significantly higher reactivation phenotype (explant TG induced reactivation in mice and spon- taneous reactivation in rabbits) than their LAT mutant coun- terparts (32, 43, 54, 55, 67), and this appears to be due in large part to LAT’s antiapoptosis activity (1, 8, 22, 55, 57, 58, 60, 72). HSV-specific CD8 T cells are selectively activated and re- tained in latently infected TG (2, 16, 42). They can significantly reduce explant TG-induced reactivation in mice (42, 48), ap- parently by interfering with virus replication and spread fol- lowing the initial molecular events of reactivation. Thus, CD8 T cells may help to decrease or delay HSV-1 reactivations (2, 42, 48) but do not eliminate the latent virus. This CD8 T-cell activity is suggested by mouse, rabbit, and human studies (37, 42, 70, 75). How does HSV-1 manage to reactivate in the face of an often sizable pool of virus-specific CD8 T cells in the TG (16, 33, 42)? One possibility is CD8 T-cell exhaustion. CD8 T-cell exhaustion is usually the result of the prolonged high- * Corresponding author. Mailing address: Laboratory of Cellular and Molecular Immunology, The Gavin Herbert Eye Institute, Oph- thalmology Research Laboratories, Hewitt Hall, Room 232, 843 Health Sciences Rd., Irvine, CA 92697-4390. Phone: (949) 824-8937. Fax: (949) 824-9626. E-mail: [email protected]. † Present address: Pathology and Clinical Laboratory Medicine, De- partment of Immunology, King Fahad Medical City, Riyadh, Kingdom of Saudi Arabia. Published ahead of print on 29 June 2011. 9127
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The herpes simplex virus 1 latency-associated transcript promotes functional exhaustion of...

JOURNAL OF VIROLOGY, Sept. 2011, p. 9127–9138 Vol. 85, No. 170022-538X/11/$12.00 doi:10.1128/JVI.00587-11Copyright © 2011, American Society for Microbiology. All Rights Reserved.
The Herpes Simplex Virus 1 Latency-Associated Transcript PromotesFunctional Exhaustion of Virus-Specific CD8� T Cells in Latently
Infected Trigeminal Ganglia: a Novel Immune Evasion Mechanism�
Aziz A. Chentoufi,1† Elizabeth Kritzer,1 Michael V. Tran,1 Gargi Dasgupta,1 Chang Hyun Lim,1
David C. Yu,1 Rasha E. Afifi,1 Xianzhi Jiang,1,2 Dale Carpenter,2Nelson Osorio,1,2 Chinhui Hsiang,1,2 Anthony B. Nesburn,1
Steven L. Wechsler,1,2,4 and Lbachir BenMohamed1,2,3,5*Laboratory of Cellular and Molecular Immunology, The Gavin Herbert Eye Institute and Department of Ophthalmology, University of
California Irvine, School of Medicine, Irvine, California 926971; Virology Research, The Gavin Herbert Eye Institute andDepartment of Ophthalmology, University of California Irvine, School of Medicine, Irvine, California 926972;
Chao Family Comprehensive Cancer Center, University of California Irvine Medical Center, Irvine,California 92868-32013; Department of Microbiology and Molecular Genetics and Center for
Virus Research, University of California Irvine, Irvine, California 926974; and Institute forImmunology, University of California Irvine, School of Medicine, Irvine, California 926975
Received 23 March 2011/Accepted 3 June 2011
Following ocular herpes simplex virus 1 (HSV-1) infection of C57BL/6 mice, HSV-specific (HSV-gB498–505tetramer�) CD8� T cells are induced, selectively retained in latently infected trigeminal ganglia (TG), andappear to decrease HSV-1 reactivation. The HSV-1 latency-associated transcript (LAT) gene, the only viralgene that is abundantly transcribed during latency, increases reactivation. Previously we found that duringlatency with HSV-1 strain McKrae-derived viruses, more of the total TG resident CD8 T cells expressedmarkers of exhaustion with LAT� virus compared to LAT� virus. Here we extend these findings to HSV-1strain 17syn�-derived LAT� and LAT� viruses and to a virus expressing just the first 20% of LAT. Thus, theprevious findings were not an artifact of HSV-1 strain McKrae, and the LAT function involved mapped to thefirst 1.5 kb of LAT. Importantly, to our knowledge, we show here for the first time that during LAT� viruslatency, most of the HSV-1-specific TG resident CD8 T cells were functionally exhausted, as judged by lowcytotoxic function and decreased gamma interferon (IFN-�) and tumor necrosis factor alpha (TNF-�) pro-duction. This resulted in LAT� TG having more functional HSV-gB498–505 tetramer� CD8� T cells comparedto LAT� TG. In addition, LAT expression, in the absence of other HSV-1 gene products, appeared to be ableto directly or indirectly upregulate both PD-L1 and major histocompatibility complex class I (MHC-I) onmouse neuroblastoma cells (Neuro2A). These findings may constitute a novel immune evasion mechanismwhereby the HSV-1 LAT directly or indirectly promotes functional exhaustion (i.e., dysfunction) of HSV-specific CD8� T cells in latently infected TG, resulting in increased virus reactivation.
Herpes simplex virus 1 (HSV-1) establishes lifelong latencyin its host. After a primary acute infection of the eye, HSV-1travels up sensory neurons to the trigeminal ganglia (TG),where it maintains latent infections (2, 16, 18, 42, 48). Recur-rent corneal disease results from the reactivation of latent virusfrom neurons of the TG, anterograde transportation to nervetermini, and reinfection of the cornea (2, 42). In addition tolatency-associated transcript (LAT) and other viral factors,recent reports suggest that CD8� T cells in the TG may con-tribute to the establishment, maintenance, and/or reactivationof latent HSV-1 in TG (2, 42). However, the mechanismsremain to be fully elucidated.
During HSV-1 latency, the HSV-1 latency-associated tran-script (LAT) gene is the only viral gene that is consistentlydetected to be abundantly transcribed. LAT� viruses (i.e.,wild-type [wt] HSV-1) have a significantly higher reactivationphenotype (explant TG induced reactivation in mice and spon-taneous reactivation in rabbits) than their LAT� mutant coun-terparts (32, 43, 54, 55, 67), and this appears to be due in largepart to LAT’s antiapoptosis activity (1, 8, 22, 55, 57, 58, 60, 72).
HSV-specific CD8� T cells are selectively activated and re-tained in latently infected TG (2, 16, 42). They can significantlyreduce explant TG-induced reactivation in mice (42, 48), ap-parently by interfering with virus replication and spread fol-lowing the initial molecular events of reactivation. Thus, CD8�
T cells may help to decrease or delay HSV-1 reactivations (2,42, 48) but do not eliminate the latent virus. This CD8� T-cellactivity is suggested by mouse, rabbit, and human studies (37,42, 70, 75).
How does HSV-1 manage to reactivate in the face of anoften sizable pool of virus-specific CD8� T cells in the TG (16,33, 42)? One possibility is CD8� T-cell exhaustion. CD8�
T-cell exhaustion is usually the result of the prolonged high-
* Corresponding author. Mailing address: Laboratory of Cellularand Molecular Immunology, The Gavin Herbert Eye Institute, Oph-thalmology Research Laboratories, Hewitt Hall, Room 232, 843Health Sciences Rd., Irvine, CA 92697-4390. Phone: (949) 824-8937.Fax: (949) 824-9626. E-mail: [email protected].
† Present address: Pathology and Clinical Laboratory Medicine, De-partment of Immunology, King Fahad Medical City, Riyadh, Kingdomof Saudi Arabia.
� Published ahead of print on 29 June 2011.
9127

level presence of antigen (Ag), as occurs during productivechronic infections (49). Chronic viral infections, such as withlymphocytic choriomeningitis virus (LCMV), HIV-1, hepatitisB virus (HBV), hepatitis C virus (HCV), and human T-lym-photropic virus (HTLV), appear to sustain their productivityby inducing functional impairment (exhaustion) of virus-spe-cific CD8� T cells (4, 21, 49). Functional exhaustion is char-acterized by progressive loss of production of cytokines, suchas interleukin-2 (IL-2), tumor necrosis factor alpha (TNF-�),and gamma interferon (IFN-�), and other functions, such ascytotoxic activity (2, 38, 41, 49, 66, 74). Phenotypic exhaustionrefers to the presence of markers of exhaustion on CD8 T cells,most commonly PD-1 (4, 25, 38, 62, 65, 66, 78). We recentlyshowed that markers of exhaustion are present on CD8 T cellsduring HSV-1 latent infection of mouse trigeminal ganglia(TG) (2) and that more CD8 T cells have these markers in TGof mice latently infected with wild-type (wt) LAT� virus com-pared to mutant LAT� virus.
In this article, we present the following findings. (i) Duringlatency, LAT� viruses caused significantly more functional ex-haustion of total and HSV-1-specific TG resident CD8� T cellsthan LAT� viruses. (ii) This was not due to an artifact of theHSV-1 strain used, since similar results were obtained withLAT� viruses from two different HSV-1 strains, McKrae and17syn�. (iii) LAT3.3A, a mutant that expresses just the first 1.5kb of the primary 8.3-kb LAT gene, gave the same result as wtMcKrae and wt 17syn�, mapping the LAT “exhaustion” func-tion to the same region of LAT that is responsible for LAT’santiapoptosis activity and ability to support the wt reactivationphenotype (59). (iv) The number of functional (not exhausted)CD8� T cells was significantly higher during latency withLAT� compared to LAT� virus. (v) In mouse neuroblastomacells stably expressing LAT, in the absence of other viral genes(as occurs during latency), both PD-L1 and major histocom-patibility complex class I (MHC-I) were significantly upregu-lated. Our findings suggest that LAT may be involved in animmune evasion mechanism that reduces the number of func-tional HSV-specific CD8� T cells within latently infected TG,thus allowing for increased viral reactivation of LAT� viruscompared to LAT� mutants.
MATERIALS AND METHODS
Mice. Female C57BL/6 (B6) mice, 5 weeks old, were purchased from theJackson Laboratory (Bar Harbor, ME). Animal studies conformed to the Guidefor the Care and Use of Laboratory Animals (50a).
Virus. Throughout the study, wild-type HSV-1 strains and LAT-rescued mu-tants expressing LAT (e.g., dLAT2903R) are designated LAT�. LAT-null virusmutants are designated LAT�. Plaque-purified HSV-1 strains McKrae (wildtype) and its derived mutants, LAT� dLAT2903, LAT� dLAT2903R, and LAT�
LAT3.3A, were grown in rabbit skin (RS) cell monolayers in minimal essentialmedium (MEM) containing 5% fetal calf serum (FCS), as described previously(39, 55, 58). The wild-type HSV-1 strain 17syn� and its LAT� mutant and LAT�
rescuants were generously provided by Nigel Fraser (10).Ocular infection. Mice were infected ocularly with 2 � 105 PFU of either
LAT� or LAT� virus. All McKrae-derived viruses were suspended in 5 �l oftissue culture medium and administered as an eye drop without prior cornealscarification. For 17syn� strain-derived viruses, corneal scarification was per-formed just prior to infection (10). Mock-infected mice received sterile tissueculture media and were used as negative controls in all experiments.
Preparation of single-cell suspensions from mouse TG. Mice were euthanizedand TG were harvested, pooled, and digested in DMEM–5% fetal bovine serum(FBS) containing collagenase I (Life Technologies, Carlsbad, CA), as we previ-ously described (5, 16, 17, 37). The digested tissue suspension was passed through
a 70-�m nylon cell strainer followed by a 40-�m nylon cell strainer. Cell suspen-sions were spun down at 1,400 rpm for 5 min at 4°C and then washed andsuspended in fluorescence-activated cell sorter (FACS) buffer (phosphate-buff-ered saline [PBS]–0.01% NaN3–0.1% bovine serum albumin [BSA], 2 mMEDTA) for FACS acquisition and analysis. Ten TG were pooled and digestedwith collagenase type I. To obtain the number of cells per TG, the total numberof cells obtained from a 10-TG cell suspension was divided by 10. Cells werestained with monoclonal antibodies (MAbs) and tetramers, as described below.The lymphocyte populations in the TG suspension were gated from other non-lymphocyte cells (i.e., glial cells, Schwann cells, neurons, fibroblasts, etc.) basedon their specific size and granularity. The total number of CD8� T cells wascalculated based on the number of gated lymphocytes and used for normaliza-tion. The percentage of CD8� T cells that were tetramer� or PD-1� was calcu-lated based on the following formula: [(no. of events in CD3� CD8� tetramer�/PD-1� cells) � (no. of events in gated lymphocytes)/(no. of total eventsacquired)] � 100.
Tetramer assay. TG cell suspensions were analyzed for the frequency of CD8�
T cells specific to the HSV-gB498–505 epitope using the HSV-gB498–505 tetramer,which is H-2Kb restricted, as we previously described (82). The cells were firstincubated with 2 �l of phycoerythrin (PE)-labeled HSV-gB498-505/H-2Kb
tetramer at 4°C for 30 to 45 min. The cells were washed twice and stained with1 �l fluorescein isothiocyanate (FITC)-conjugated rat anti-mouse CD8 antibody(clone 53-6.7; eBioscience). After two additional washings, cells were fixed with1% formaldehyde in PBS. A total of 400,000 events were acquired by FACScan(Becton Dickinson) followed by analysis using Cell Quest software (BD Biosci-ences, San Jose, CA). The absolute numbers of HSV-gB498-505-specific CD8� Tcells were calculated using the following formula: no. of CD8� tetramer� cells inthe test � no. of CD8� tetramer� cells in the negative control. The anti-mouseCD8 MAb was pretested prior to use to confirm that it did not interfere withaltered tetramer binding.
Intracellular cytokine assay. Intracellular cytokines produced by HSV-gB498–505
CD8� T cells were measured using an intracellular cytokine assay kit (BD-Biosciences). Freshly isolated TG-derived CD8� T cells were either unstimu-lated or were stimulated in triplicate with HSV-gB498–505 peptide or phorbolmyristate acetate plus ionomycin (PMA � Iono) (positive control) at 37°C for6 h, as recommended by the manufacturer. The cells were surface stained withperidinin chlorophyll protein (PerCP)-conjugated anti-mouse CD8 for 30 min at4°C. For subsequent intracellular staining, cells were permeabilized and stainedwith PE-conjugated anti-mouse IFN-� and TNF-� MAbs, as recommended bythe manufacturer. The cells were washed twice and analyzed using a FACScan(BD Biosciences).
CD107 cytotoxicity assay. The CD107 assay was performed as recently de-scribed (15–18, 37) with a few modifications. Freshly isolated TG-derived cellswere left unstimulated or were stimulated with gB498–505 or phytohemagglutinin(PHA) (positive control) at 37°C for 5 to 6 h in the presence of BD GolgiStop(BD Biosciences) and 10 �l of FITC-conjugated CD107 antibody. Cells werewashed and stained with 2 �l of PE-labeled HSV-gB498–505/H-2Kb tetramer and1 �l of PerCP-conjugated anti-mouse CD8 for 30 min at 4°C. The cells were thenwashed again and analyzed using a FACScan (BD Biosciences).
Neuronal cell lines. The mouse neuroblastoma cell lines C1300 and Neuro2Ahave been widely used as neuronal tissue culture models to study various aspectsof herpes simplex virus. The LAT� DC-LAT6 and LAT� DC-�LAT311 C1300-derived cell lines and the LAT� JWLAT and LAT� JW-�LAT Neuro2A-de-rived cell lines have been described previously (12, 13, 37). JWLAT expresses ashorter region of LAT, LAT nucleotides (nt) 1 to 1499, compared to LAT nt 1to 3225 for DC-LAT6. C1300-derived cell lines were grown in MEM containing10% (vol/vol) heat-inactivated fetal calf serum, penicillin (10 U/ml), streptomy-cin (100 mg/ml), 1 mM sodium pyruvate, 1� MEM nonessential amino acids, and1 mg/ml G-418. The Neuro2A-derived cell lines were grown in the same medium,but with 1 �g/ml puromycin in place of the G-418.
RNA expression levels. For real-time PCR, the RNeasy minikit from Qiagenwas used to isolate RNA from Neuro2A cells according to the manufacturer’sinstructions. The quality (optical density at 260/280 nm [OD260/280]) and quantity(concentration) of RNA were analyzed by Nanodrop spectrophotometer (ND-1000). To prepare cDNA, the RNA was reverse transcribed by reverse trans-criptase following the instructions provided in the Qiagen Sensiscript synthesiskit. The cDNA was stored at �20°C until use. cDNA from each sample (equiv-alent to 50 to 100 ng input RNA) was diluted with RNase-free water andsubjected to PCR using specific primers. The primer pair PD-L1 (catalog no.MP201906) and PD-L2 (catalog no. MP211705) were purchased from OriGeneTechnologies, Inc., and were used as recommended by the manufacturer. SYBRgreen master mix (Applied Biosystems) was used in a final volume of 25 �l. Thecycling parameters were 95°C for 10 min, followed by 40 cycles of 95°C for 30s
9128 CHENTOUFI ET AL. J. VIROL.

and 60°C for 60s. Samples were normalized to 16S RNA and were amplified intriplicate using an MJ Research Opticon thermal cycler. PCR controls withoutreverse transcriptase (water control) or with normal human genomic DNA as atemplate were routinely negative. The levels of cytokine expression were deter-mined using a threshold cycle (2�[supb]��CT) method as described previously(51). The parental Neuro2A cell line was used as the reference for the expressionof PD-L1 and PD-L2 in the LAT� and LAT� cell lines.
Western blotting. Total proteins from whole-cell lysates of LAT� and LAT�
Neuro2A cell lines were separated on NuPAGE 4 to 12% Bis-Tris gels. AfterWestern transfer, the membrane was probed with 1 �g/ml anti-PD-L1 (catalogno. AP30655PU-N; Acris Antibodies). Horseradish peroxidase (HRP)-conju-gated goat anti-rabbit IgG (Invitrogen) at a 1:2,000 dilution was used as thesecondary antibody. Super Signal West Femto maximum sensitivity substrate(Thermo Scientific) was used to detect PD-L1.
Statistical analysis. Student’s t test and analysis of variance (ANOVA) wereperformed using the computer program Instat (GraphPad, San Diego). Resultswere considered statistically significant when the P value was �0.05.
RESULTS
Increased number of total TG resident CD8 T cells on days11 and 35 postocular infection of mice with LAT� compared toLAT� virus. In a pilot experiment, we compared the percent-ages and numbers of total T cells in TG of mice latentlyinfected with wild-type HSV-1 (strain McKrae) that had beenperfused or not perfused prior to euthanasia. The numbers andpercentages of HSV-1-specific CD4� and CD8� T cells weresimilar in perfused and nonperfused TG (not shown). This wasexpected, since during HSV-1 latency there are few if anyHSV-1-specific T cells in the circulating blood. Subsequentexperiments were carried out using nonperfused mice.
Groups of five B6 mice were infected ocularly with 2 � 105
PFU/eye of the LAT-null virus mutant dLAT2903 (LAT�) orits marker-rescued virus, dLAT2903R (LAT�) (55). Both vi-ruses are derived from HSV-1 strain McKrae. TG were har-vested 11 days postinfection (p.i.) (just after acute infection) or35 days p.i. (i.e., after latency was well established). The per-centage and number of CD3� CD8� T cells in TG were de-termined on both day 11 (Fig. 1A) and day 35 (Fig. 1B) postin-fection. Representative experiments, each consisting of cellspooled from 10 TG, show the percentage of total TG residentCD8� T cells (i.e., CD3� CD8�) (left panels). The average standard deviation (SD)/TG of seven independent experimentsis shown in the right panels. Both the percentage and averagenumber of CD8� T cells were higher in LAT� TG comparedto LAT� TG. As expected, more TG resident CD8� T cellswere present on day 11 than on day 35. On both day 11 and day35, there were no differences in the percentages or numbers oftotal CD3� CD8� T cells in spleens from LAT�- compared toLAT�-infected mice (data not shown), indicating that the ef-fect of LAT on CD8� T cells was TG specific.
High numbers of CD8� T cells were detected in LAT� TGregardless of the mutant or strain of the virus. To verify thatthe higher number of CD8� T cells in LAT� TG seen in Fig.1 was not due to an artifact linked to the mutant virus used, weinfected additional groups of 5 B6 mice with a different LAT�
mutant that has the first 1.5 kb of LAT expressed from anectopic location in an otherwise LAT-null mutant (LAT3.3A)(58). LAT3.3A is a McKrae strain mutant and has a wild-typelatency and reactivation phenotype similar to those of bothLAT� dLAT2903R and wild-type McKrae (58). Again, onboth day 11 (Fig. 2A, left panel) and day 35 (Fig. 2A, rightpanel), the cells from 10 TG/group were pooled. The average
number of CD8� T cells per TG from five independent exper-iments was significantly higher with LAT� virus than withLAT� virus (P � 0.05). These results indicate that the first 1.5kb of the 8.3-kb LAT primary transcript is sufficient to directlyor indirectly increase the CD8� T-cell pool in TG. Similar tothe results shown in Fig. 1, more CD3� CD8� T cells weredetected on day 11 than during latency on day 35.
To confirm that the above results were not specific to HSV-1strain McKrae and its mutants, an additional set of experi-ments was conducted using HSV-1 17syn� (LAT�) and itsLAT� mutant, 17N/H (44) (Fig. 2B). As described above, twogroups of five B6 mice were ocularly infected with 2 � 105
PFU/eye of either HSV-1 17syn� (LAT�) or 17N/H (LAT�).The numbers of TG resident CD3� CD8� T cells were com-pared between the two groups during latency (35 days p.i.),using FACS as in Fig. 2A. Again, the average number of CD3�
CD8� T cells/TG from five independent experiments was sig-nificantly higher in latently infected LAT� TG compared toLAT� TG (Fig. 2B). These results indicate that the increase in
FIG. 1. Higher percentages and numbers of total CD8� T cellsdetected in LAT� TG compared to LAT� TG during latent herpes-virus infection. Three groups of B6 mice (n 10) were either leftuninfected (Mock) or were ocularly infected with 2 � 105 PFU/eye of(i) LAT-null virus mutant dLAT2903 (LAT�) or with (ii) its marker-rescued virus, dLAT2903R (LAT�) (53). Both TG were harvestedfrom each mouse either 11 days postinfection (5 mice) or 35 dayspostinfection (5 mice). The 10 TG were pooled and treated withcollagenase I, and the numbers (nbr) and percentages of CD3� T cellsand CD3� CD8� T cells were determined in TG cell suspensions byFACS. (A). Dot plot showing percentages of CD3� CD8� T cells (leftpanel) and the average number SD/TG (right panel) of CD3� CD8�
T cells per TG on day 11 postinfection. (B) Dot plot of the percentagesof CD3� CD8� T cells (left panel) and the average number SD/TG(right panel) of CD3� CD8� T cells per TG on day 35 postinfection.Each histogram and dot plot represents seven consecutive independentexperiments. The bar graphs show the means and SD of the sevenindependent experiments at each time point. *, P � 0.05 for cellnumbers in LAT� TG versus LAT� TG.
VOL. 85, 2011 CD8� T-CELL EXHAUSTION IN LATENT HERPESVIRUS INFECTION 9129
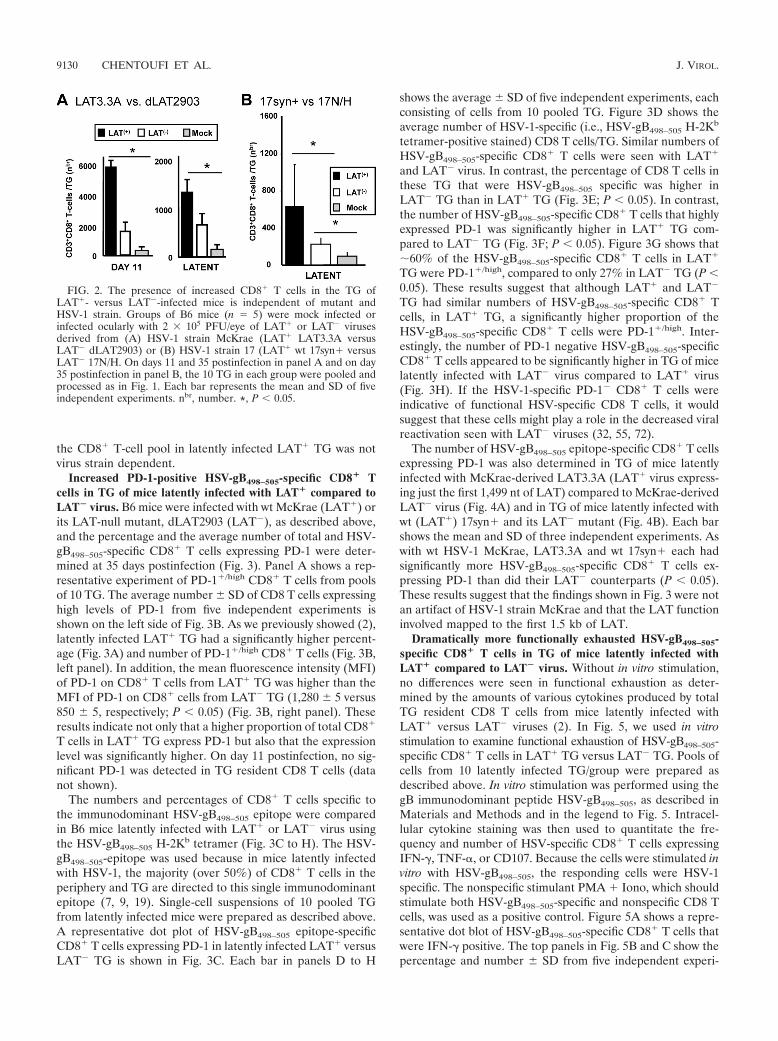
the CD8� T-cell pool in latently infected LAT� TG was notvirus strain dependent.
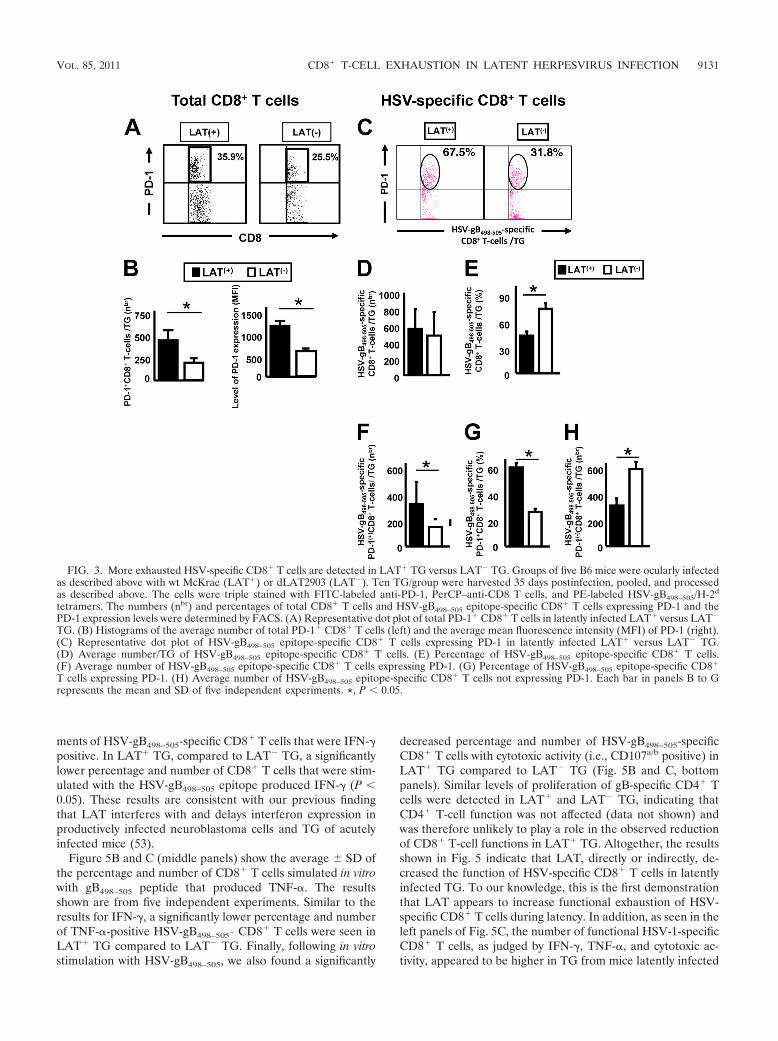
Increased PD-1-positive HSV-gB498–505-specific CD8� Tcells in TG of mice latently infected with LAT� compared toLAT� virus. B6 mice were infected with wt McKrae (LAT�) orits LAT-null mutant, dLAT2903 (LAT�), as described above,and the percentage and the average number of total and HSV-gB498–505-specific CD8� T cells expressing PD-1 were deter-mined at 35 days postinfection (Fig. 3). Panel A shows a rep-resentative experiment of PD-1�/high CD8� T cells from poolsof 10 TG. The average number SD of CD8 T cells expressinghigh levels of PD-1 from five independent experiments isshown on the left side of Fig. 3B. As we previously showed (2),latently infected LAT� TG had a significantly higher percent-age (Fig. 3A) and number of PD-1�/high CD8� T cells (Fig. 3B,left panel). In addition, the mean fluorescence intensity (MFI)of PD-1 on CD8� T cells from LAT� TG was higher than theMFI of PD-1 on CD8� cells from LAT� TG (1,280 5 versus850 5, respectively; P � 0.05) (Fig. 3B, right panel). Theseresults indicate not only that a higher proportion of total CD8�
T cells in LAT� TG express PD-1 but also that the expressionlevel was significantly higher. On day 11 postinfection, no sig-nificant PD-1 was detected in TG resident CD8 T cells (datanot shown).
The numbers and percentages of CD8� T cells specific tothe immunodominant HSV-gB498–505 epitope were comparedin B6 mice latently infected with LAT� or LAT� virus usingthe HSV-gB498–505 H-2Kb tetramer (Fig. 3C to H). The HSV-gB498–505-epitope was used because in mice latently infectedwith HSV-1, the majority (over 50%) of CD8� T cells in theperiphery and TG are directed to this single immunodominantepitope (7, 9, 19). Single-cell suspensions of 10 pooled TGfrom latently infected mice were prepared as described above.A representative dot plot of HSV-gB498–505 epitope-specificCD8� T cells expressing PD-1 in latently infected LAT� versusLAT� TG is shown in Fig. 3C. Each bar in panels D to H
shows the average SD of five independent experiments, eachconsisting of cells from 10 pooled TG. Figure 3D shows theaverage number of HSV-1-specific (i.e., HSV-gB498–505 H-2Kb
tetramer-positive stained) CD8 T cells/TG. Similar numbers ofHSV-gB498–505-specific CD8� T cells were seen with LAT�
and LAT� virus. In contrast, the percentage of CD8 T cells inthese TG that were HSV-gB498–505 specific was higher inLAT� TG than in LAT� TG (Fig. 3E; P � 0.05). In contrast,the number of HSV-gB498–505-specific CD8� T cells that highlyexpressed PD-1 was significantly higher in LAT� TG com-pared to LAT� TG (Fig. 3F; P � 0.05). Figure 3G shows that�60% of the HSV-gB498–505-specific CD8� T cells in LAT�
TG were PD-1�/high, compared to only 27% in LAT� TG (P �0.05). These results suggest that although LAT� and LAT�
TG had similar numbers of HSV-gB498–505-specific CD8� Tcells, in LAT� TG, a significantly higher proportion of theHSV-gB498–505-specific CD8� T cells were PD-1�/high. Inter-estingly, the number of PD-1 negative HSV-gB498–505-specificCD8� T cells appeared to be significantly higher in TG of micelatently infected with LAT� virus compared to LAT� virus(Fig. 3H). If the HSV-1-specific PD-1� CD8� T cells wereindicative of functional HSV-specific CD8 T cells, it wouldsuggest that these cells might play a role in the decreased viralreactivation seen with LAT� viruses (32, 55, 72).
The number of HSV-gB498–505 epitope-specific CD8� T cellsexpressing PD-1 was also determined in TG of mice latentlyinfected with McKrae-derived LAT3.3A (LAT� virus express-ing just the first 1,499 nt of LAT) compared to McKrae-derivedLAT� virus (Fig. 4A) and in TG of mice latently infected withwt (LAT�) 17syn� and its LAT� mutant (Fig. 4B). Each barshows the mean and SD of three independent experiments. Aswith wt HSV-1 McKrae, LAT3.3A and wt 17syn� each hadsignificantly more HSV-gB498–505-specific CD8� T cells ex-pressing PD-1 than did their LAT� counterparts (P � 0.05).These results suggest that the findings shown in Fig. 3 were notan artifact of HSV-1 strain McKrae and that the LAT functioninvolved mapped to the first 1.5 kb of LAT.
Dramatically more functionally exhausted HSV-gB498–505-specific CD8� T cells in TG of mice latently infected withLAT� compared to LAT� virus. Without in vitro stimulation,no differences were seen in functional exhaustion as deter-mined by the amounts of various cytokines produced by totalTG resident CD8 T cells from mice latently infected withLAT� versus LAT� viruses (2). In Fig. 5, we used in vitrostimulation to examine functional exhaustion of HSV-gB498–505-specific CD8� T cells in LAT� TG versus LAT� TG. Pools ofcells from 10 latently infected TG/group were prepared asdescribed above. In vitro stimulation was performed using thegB immunodominant peptide HSV-gB498–505, as described inMaterials and Methods and in the legend to Fig. 5. Intracel-lular cytokine staining was then used to quantitate the fre-quency and number of HSV-specific CD8� T cells expressingIFN-�, TNF-�, or CD107. Because the cells were stimulated invitro with HSV-gB498–505, the responding cells were HSV-1specific. The nonspecific stimulant PMA � Iono, which shouldstimulate both HSV-gB498–505-specific and nonspecific CD8 Tcells, was used as a positive control. Figure 5A shows a repre-sentative dot blot of HSV-gB498–505-specific CD8� T cells thatwere IFN-� positive. The top panels in Fig. 5B and C show thepercentage and number SD from five independent experi-
FIG. 2. The presence of increased CD8� T cells in the TG ofLAT�- versus LAT�-infected mice is independent of mutant andHSV-1 strain. Groups of B6 mice (n 5) were mock infected orinfected ocularly with 2 � 105 PFU/eye of LAT� or LAT� virusesderived from (A) HSV-1 strain McKrae (LAT� LAT3.3A versusLAT� dLAT2903) or (B) HSV-1 strain 17 (LAT� wt 17syn� versusLAT� 17N/H. On days 11 and 35 postinfection in panel A and on day35 postinfection in panel B, the 10 TG in each group were pooled andprocessed as in Fig. 1. Each bar represents the mean and SD of fiveindependent experiments. nbr, number. *, P � 0.05.
9130 CHENTOUFI ET AL. J. VIROL.
ments of HSV-gB498–505-specific CD8� T cells that were IFN-�positive. In LAT� TG, compared to LAT� TG, a significantlylower percentage and number of CD8� T cells that were stim-ulated with the HSV-gB498–505 epitope produced IFN-� (P �0.05). These results are consistent with our previous findingthat LAT interferes with and delays interferon expression inproductively infected neuroblastoma cells and TG of acutelyinfected mice (53).
Figure 5B and C (middle panels) show the average SD ofthe percentage and number of CD8� T cells simulated in vitrowith gB498–505 peptide that produced TNF-�. The resultsshown are from five independent experiments. Similar to theresults for IFN-�, a significantly lower percentage and numberof TNF-�-positive HSV-gB498–505� CD8� T cells were seen inLAT� TG compared to LAT� TG. Finally, following in vitrostimulation with HSV-gB498–505, we also found a significantly
decreased percentage and number of HSV-gB498–505-specificCD8� T cells with cytotoxic activity (i.e., CD107a/b positive) inLAT� TG compared to LAT� TG (Fig. 5B and C, bottompanels). Similar levels of proliferation of gB-specific CD4� Tcells were detected in LAT� and LAT� TG, indicating thatCD4� T-cell function was not affected (data not shown) andwas therefore unlikely to play a role in the observed reductionof CD8� T-cell functions in LAT� TG. Altogether, the resultsshown in Fig. 5 indicate that LAT, directly or indirectly, de-creased the function of HSV-specific CD8� T cells in latentlyinfected TG. To our knowledge, this is the first demonstrationthat LAT appears to increase functional exhaustion of HSV-specific CD8� T cells during latency. In addition, as seen in theleft panels of Fig. 5C, the number of functional HSV-1-specificCD8� T cells, as judged by IFN-�, TNF-�, and cytotoxic ac-tivity, appeared to be higher in TG from mice latently infected
FIG. 3. More exhausted HSV-specific CD8� T cells are detected in LAT� TG versus LAT� TG. Groups of five B6 mice were ocularly infectedas described above with wt McKrae (LAT�) or dLAT2903 (LAT�). Ten TG/group were harvested 35 days postinfection, pooled, and processedas described above. The cells were triple stained with FITC-labeled anti-PD-1, PerCP–anti-CD8 T cells, and PE-labeled HSV-gB498–505/H-2d
tetramers. The numbers (nbr) and percentages of total CD8� T cells and HSV-gB498–505 epitope-specific CD8� T cells expressing PD-1 and thePD-1 expression levels were determined by FACS. (A) Representative dot plot of total PD-1� CD8� T cells in latently infected LAT� versus LAT�
TG. (B) Histograms of the average number of total PD-1� CD8� T cells (left) and the average mean fluorescence intensity (MFI) of PD-1 (right).(C) Representative dot plot of HSV-gB498–505 epitope-specific CD8� T cells expressing PD-1 in latently infected LAT� versus LAT� TG.(D) Average number/TG of HSV-gB498–505 epitope-specific CD8� T cells. (E) Percentage of HSV-gB498–505 epitope-specific CD8� T cells.(F) Average number of HSV-gB498–505 epitope-specific CD8� T cells expressing PD-1. (G) Percentage of HSV-gB498–505 epitope-specific CD8�
T cells expressing PD-1. (H) Average number of HSV-gB498–505 epitope-specific CD8� T cells not expressing PD-1. Each bar in panels B to Grepresents the mean and SD of five independent experiments. *, P � 0.05.
VOL. 85, 2011 CD8� T-CELL EXHAUSTION IN LATENT HERPESVIRUS INFECTION 9131

with LAT� compared to LAT� virus. This suggests that thehigher number of functional HSV-1-specific CD8 T cells in TGof mice latently infected with LAT� virus may play an impor-tant role in the decreased viral reactivation seen with LAT�
viruses (32, 55, 72).Neuronally derived cells stably expressing LAT have ele-
vated MHC-I and PD-L1. Approximately 30% to over 50% ofneurons in TG from mice latently infected with LAT� (wt)virus harbor latent virus, as judged by the presence of viralDNA or LAT RNA (46, 63, 64). However, less than 1 neu-ron/TG has detectable viral Ag by immunostaining (2, 23, 24,45, 80). The amount of latency and detectable viral Ag in TGfrom mice latently infected with LAT� virus is even smallerthan with LAT� virus (31). As expected, we obtained similarresults in these studies (not shown). In addition, spontaneousreactivation in mice is extremely rare (28). Thus, HSV-1 la-tency in mice represents an almost completely dormant state.The very low levels of viral Ag detected during latency suggestthat the continuous high-level exposure to Ag that is thought tobe necessary to produce CD8 T-cell exhaustion may not occurduring HSV-1 latency in mouse TG. It was therefore of interestto determine if LAT alone, in the absence of other viral genes,could contribute to the observed CD8� T-cell exhaustion, sowe examined two mouse neuroblastoma cell lines (Neuro2Acells and C1300 cells) that we had previously independentlyengineered to stably express LAT (12, 13, 37). JWLAT cells
FIG. 4. PD-1 expression in LAT3.3A and in LAT� and LAT�
HSV-1 strain 17syn� viruses. Groups of five B6 mice were ocularlyinfected as described above with 17syn� (LAT�), 17N/H (LAT�),LAT3.3A (LAT�), or dLAT2903 (LAT�). Ten TG/group were har-vested 35 days postinfection, pooled, and processed as describedabove. The cells were triple stained with FITC-labeled anti-PD-1,PerCP–anti-CD8, and PE-labeled HSV-gB498–505/H-2d tetramers andanalyzed by FACS as described above. The average numbers (nbr) ofHSV-gB498–505 epitope-specific CD8� T cells expressing PD-1 areshown. (A) McKrae-derived LAT� LAT3.3A (expresses LAT nt 1 to1499) versus McKrae-derived LAT� dLAT2903. (B) 17syn�-derivedLAT� wt versus 17syn�-derived LAT� 17N/H. Each bar shows themean and SD of three independent experiments. *, P � 0.05.
FIG. 5. HSV-gB498–505-specific CD8� T cells from LAT� TG have reduced cytokine production and cytolytic activity. Two groups of B6 mice(n 5) were ocularly infected with wt McKrae (LAT�) or dLAT2903 (LAT�) as described above. At 35 days postinfection, the 10 TG in eachgroup were pooled, and single-cell suspensions were prepared as described above. For intracellular cytokine staining, the cells were stimulated withgD498–505 peptide (10 �g/ml) or PMA � Iono (1 �g/ml) (positive control) or mock stimulated (negative control) at 37°C for 6 h in the presenceof Golgi plug (BD Biosciences). The cells were then stained with PerCP-conjugated anti-mouse CD8 for 30 min, permeabilized, and stained withPE-labeled anti-mouse IFN-� or TNF-� antibodies, as recommended by the manufacturer. For the CD107 degranulation assay, the cells were mockstimulated or stimulated with HSV-gB498–505 peptide or PMA � Iono (positive control) at 37°C for 6 h in the presence of 0.7 �l/ml of BDGolgi-Stop and 10 �l of CD107a/b-FITC, washed with FACS buffer, and stained with PerCP-conjugated anti-mouse CD8 MAb at 4°C. Thepercentages and average numbers (Nbr)/TG of HSV-gB498–505 epitope-specific CD8� T cells producing IFN-�, TNF-�, or expressing CD107a/b weredetermined by FACS. (A) Representative dot plots of intracellular IFN-�� CD8�T cells. (B) Percentage of HSV-gB498–505 epitope-specific CD8�
T cells producing IFN-�, TNF-�, or CD107a/b per TG. (C) Average number/TG of HSV-gB498–505-epitope-specific CD8� T cells producing IFN-�,TNF-�, or CD107a/b. In panels B and C, each bar represents the average SD of five experiments. *, P � 0.05.
9132 CHENTOUFI ET AL. J. VIROL.
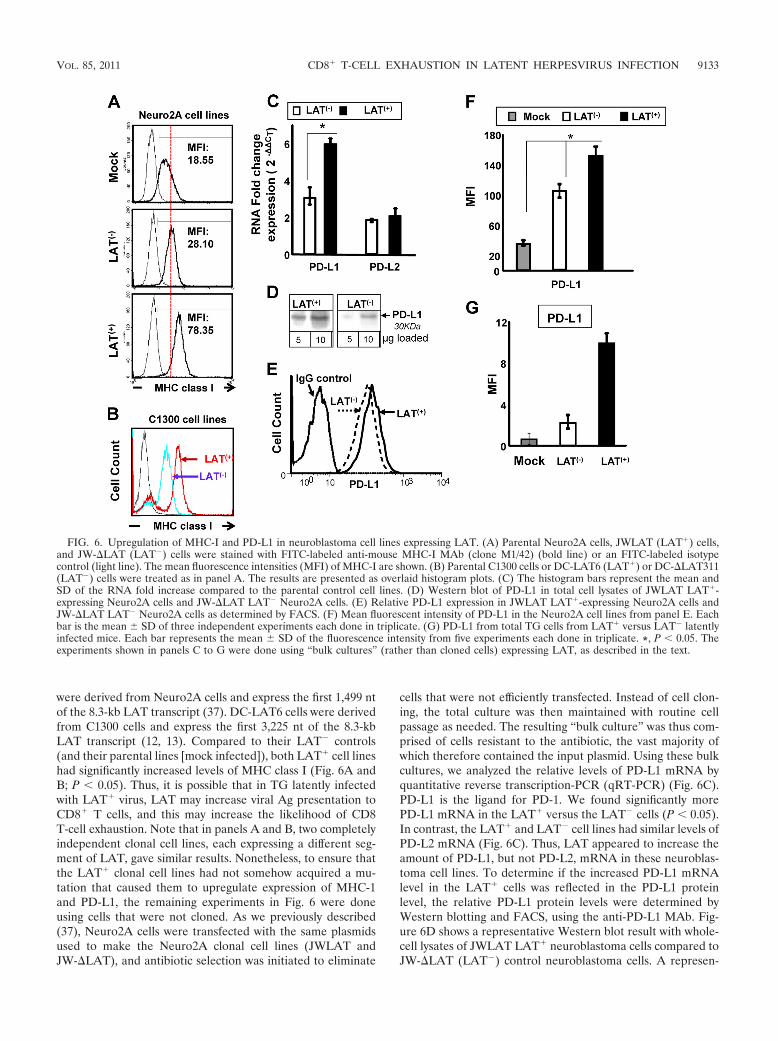
were derived from Neuro2A cells and express the first 1,499 ntof the 8.3-kb LAT transcript (37). DC-LAT6 cells were derivedfrom C1300 cells and express the first 3,225 nt of the 8.3-kbLAT transcript (12, 13). Compared to their LAT� controls(and their parental lines [mock infected]), both LAT� cell lineshad significantly increased levels of MHC class I (Fig. 6A andB; P � 0.05). Thus, it is possible that in TG latently infectedwith LAT� virus, LAT may increase viral Ag presentation toCD8� T cells, and this may increase the likelihood of CD8T-cell exhaustion. Note that in panels A and B, two completelyindependent clonal cell lines, each expressing a different seg-ment of LAT, gave similar results. Nonetheless, to ensure thatthe LAT� clonal cell lines had not somehow acquired a mu-tation that caused them to upregulate expression of MHC-1and PD-L1, the remaining experiments in Fig. 6 were doneusing cells that were not cloned. As we previously described(37), Neuro2A cells were transfected with the same plasmidsused to make the Neuro2A clonal cell lines (JWLAT andJW-�LAT), and antibiotic selection was initiated to eliminate
cells that were not efficiently transfected. Instead of cell clon-ing, the total culture was then maintained with routine cellpassage as needed. The resulting “bulk culture” was thus com-prised of cells resistant to the antibiotic, the vast majority ofwhich therefore contained the input plasmid. Using these bulkcultures, we analyzed the relative levels of PD-L1 mRNA byquantitative reverse transcription-PCR (qRT-PCR) (Fig. 6C).PD-L1 is the ligand for PD-1. We found significantly morePD-L1 mRNA in the LAT� versus the LAT� cells (P � 0.05).In contrast, the LAT� and LAT� cell lines had similar levels ofPD-L2 mRNA (Fig. 6C). Thus, LAT appeared to increase theamount of PD-L1, but not PD-L2, mRNA in these neuroblas-toma cell lines. To determine if the increased PD-L1 mRNAlevel in the LAT� cells was reflected in the PD-L1 proteinlevel, the relative PD-L1 protein levels were determined byWestern blotting and FACS, using the anti-PD-L1 MAb. Fig-ure 6D shows a representative Western blot result with whole-cell lysates of JWLAT LAT� neuroblastoma cells compared toJW-�LAT (LAT�) control neuroblastoma cells. A represen-
FIG. 6. Upregulation of MHC-I and PD-L1 in neuroblastoma cell lines expressing LAT. (A) Parental Neuro2A cells, JWLAT (LAT�) cells,and JW-�LAT (LAT�) cells were stained with FITC-labeled anti-mouse MHC-I MAb (clone M1/42) (bold line) or an FITC-labeled isotypecontrol (light line). The mean fluorescence intensities (MFI) of MHC-I are shown. (B) Parental C1300 cells or DC-LAT6 (LAT�) or DC-�LAT311(LAT�) cells were treated as in panel A. The results are presented as overlaid histogram plots. (C) The histogram bars represent the mean andSD of the RNA fold increase compared to the parental control cell lines. (D) Western blot of PD-L1 in total cell lysates of JWLAT LAT�-expressing Neuro2A cells and JW-�LAT LAT� Neuro2A cells. (E) Relative PD-L1 expression in JWLAT LAT�-expressing Neuro2A cells andJW-�LAT LAT� Neuro2A cells as determined by FACS. (F) Mean fluorescent intensity of PD-L1 in the Neuro2A cell lines from panel E. Eachbar is the mean SD of three independent experiments each done in triplicate. (G) PD-L1 from total TG cells from LAT� versus LAT� latentlyinfected mice. Each bar represents the mean SD of the fluorescence intensity from five experiments each done in triplicate. *, P � 0.05. Theexperiments shown in panels C to G were done using “bulk cultures” (rather than cloned cells) expressing LAT, as described in the text.
VOL. 85, 2011 CD8� T-CELL EXHAUSTION IN LATENT HERPESVIRUS INFECTION 9133

tative FACS result from PD-L1 expression on JWLAT LAT�
neuroblastoma cells compared to JW-�LAT (LAT�) controlneuroblastoma cells is shown in Fig. 6E. The mean SD ofthree independent FACS experiments, each done in triplicate,is shown in Fig. 6F. LAT� and LAT� cells both had morePD-L1 than the normal control Neuro2A cells (mock). Moreimportantly, there was significantly more PD-L1 expression onthe LAT� cells compared to the LAT� cells (Fig. 6F; P �0.05). Thus, both PD-L1 mRNA and protein were elevated inthe LAT� compared to the LAT� Neuro2A cell lines. To ourknowledge, this is the first report of LAT alone, in the absenceof other viral genes or gene products, upregulating eitherMHC-I or PD-L1. PD-L1 protein also appeared to be upregu-lated in total cell extracts of TG from mice latently infectedwith LAT� compared to LAT� virus (Fig. 6G; P � 0.05). Theresults shown in Fig. 6 suggest that LAT can upregulate bothMHC-I and PD-L1 on neuronally derived cells (and possiblyother LAT� TG resident cells—for example, dendritic cells).This might increase CD8� T-cell exhaustion by providing in-creased antigen presentation (via increased MHC-I) and/or byrendering them dysfunctional through the PD-L1/PD-1 nega-tive regulatory pathway.
DISCUSSION
This report extends and expands recent findings regardingthe role of HSV-1 LAT in CD8 T-cell exhaustion in TG ofmice latently infected with HSV-1 (2). Using only HSV-1 strainMcKrae-derived viruses, the previous report showed that, com-pared to LAT� virus, in mice latently infected with LAT�
virus, there is increased phenotypic exhaustion of total TGresident CD8 T cells. In this report, we have extended thosefindings to LAT� and LAT� HSV-1 strain 17syn� viruses,thus demonstrating that the previous findings were not anartifact of HSV-1 strain McKrae. We also showed that a mu-tant expressing just the first 1,499 nt of the 8.3-kb LAT wascapable of producing phenotypic exhaustion of CD8 T cells.This mapped the LAT region involved to the beginning of theLAT transcript, the same region to which we previouslymapped LAT’s antiapoptosis activity (39) and LAT’s ability toenhance spontaneous reactivation in rabbits (35, 58). In addi-tion, we have now found that on day 11 postinfection (just afterthe acute infection has cleared, a time not previously exam-ined), there were significantly more total CD8 T cells withLAT� versus LAT� virus. This is similar to what was previ-ously found during latency on day 30 postinfection (2) andwhat we found here on day 35 postinfection. However, incontrast to days 30 and 35, on day 11, we did not find significantCD8 T-cell exhaustion in either group (data not shown). Thus,the increase in CD8 T-cell numbers in LAT� TG on day 11 wasnot the result of increased exhaustion, which had not yet oc-curred. More importantly, we now show that compared toLAT� virus, in mice latently infected with LAT� virus, (i)HSV-1-specific CD8 T cells, as well as total CD8 T cells, werephenotypically exhausted; (ii) HSV-1-specific CD8 T cells werefunctionally exhausted; and (iii) LAT alone, in the absence ofall other HSV-1 gene products, increased expression of bothMHC-I and PD-L1 on neuronally derived cells. To our knowl-edge, this is the first report showing LAT-related functionalexhaustion of HSV-1-specific CD8 T cells in TG of latently
infected mice and the first report showing that LAT by itselfcan increase two different proteins (MHC-I and PD-L1), bothof which could logically increase CD8 T-cell exhaustion.
During HSV-1 neuronal latency in TG of mice and humans,some neurons that may or may not be latently infected aresurrounded by CD8� T cells (34, 42, 68–70, 76). Since CD8� Tcells are presumably attracted to these neurons by viral Ags, itis assumed that the neurons surrounded by CD8� T cells arethose in which the virus has initiated the early stages of reac-tivation from latency. In mice, experimental reactivation ofHSV-1 from latency is typically accomplished by explanting TGinto tissue culture media for up to 14 days and testing for theappearance of infectious (i.e., reactivated) virus. In this TGexplant-induced reactivation model, depletion of CD8� T cellswith a specific MAb leads to the detection of more reactivatedvirus (42, 47). Conversely, addition of exogenous CD8� T cellsreduces detection of reactivated virus (16, 33, 42). Thus, withwild-type HSV-1, CD8� T cells in the TG are apparently ableto reduce the detection of infectious reactivated virus.
Induced HSV-1 reactivation in mice and spontaneousHSV-1 reactivation in rabbits are both significantly reduced inanimals latently infected with LAT� mutants compared toLAT� viruses (32, 55, 72). Combining this fact with the aboveconcept that increased numbers of CD8� T cells in the TGresult in decreased detection of HSV-1 reactivation, we hy-pothesized that TG from mice latently infected with LAT�
viruses would have increased numbers of CD8� T cells com-pared to TG from mice latently infected with LAT� viruses.Unexpectedly, in the previous study (2) and here, we found thereverse. Both the absolute number of CD8� T cells per mouseTG and the percentage of CD8� T cells among all CD3� Tcells were significantly higher in mice infected with LAT�
compared to LAT� viruses. In addition, we now show that thiswas also the case on day 11 post-ocular infection (just afterclearance of the acute infection) as well as when latency waswell established (day 30 or 35).
Many persistent and/or chronic viral infections generatefunctionally impaired antigen-specific T-cell populations (4,30, 38, 79). It should be noted that PD-1 expression by itselfdoes not demonstrate exhaustion. However, it is well estab-lished that activated memory T cells and exhausted T cells havedifferent levels of PD-1 expression, with exhausted CD8� Tcells expressing higher levels of PD-1 compared to activatedmemory T cells (4, 6, 79). With LCMV, PD-1 is markedlyupregulated on exhausted T cells but only transiently expressedon effector T cells during acute infections and is not present onfunctionally competent memory T cells (4, 26, 36). These dif-ferent levels of PD-1 can help distinguish between exhaustionand activation. In addition, in this study, exhausted (dysfunc-tional) CD8� T cells were confirmed by impaired cytotoxicityactivity and decreased IFN-� and TNF-� production.
In the previous report (2), exhaustion was studied with totalTG resident CD8 T cells. In this report, we studied exhaustionof HSV-specific CD8 T cells, since these are the CD8 T cellsthat would affect HSV-1 latency and reactivation. In the pre-vious report, qRT-PCR of total TG RNA did not reveal anysignificant decreases in the amount of IL-2, IFN-�, or TNF-�mRNAs in LAT� versus LAT� latently infected TG (see Fig.5 in reference 2). Thus, that experiment did not provide evi-dence of functional exhaustion in TG latently infected with
9134 CHENTOUFI ET AL. J. VIROL.

LAT� compared to LAT� virus. In addition, a FACS analysisin which the CD8 T cells were not stimulated in vitro did notreveal any obvious functional exhaustion of CD8� T cells fromLAT� versus LAT� TG (see Table 2 in reference 2). In con-trast, here we found that the percentage and number of IFN-�HSV-specific CD8 T cells, TNF-� HSV-specific CD8 T cells,and CD107� HSV-specific CD8 T cells were all significantlydecreased in TG of mice latently infected with LAT� com-pared to LAT� virus. This indicated functional exhaustion ofboth cytokine expression and cytotoxic activity of these CD8 Tcells. To our knowledge, this is the first report to show signif-icantly more phenotypically and functionally exhausted HSV-specific CD8� T cells in TG from mice latently infected withLAT� virus compared to LAT� virus. In addition, since wefound here that most of the HSV-1-specific CD8� T cells in theTG of mice latently infected with LAT� virus appeared to beboth phenotypically and functionally exhausted, there wereactually significantly more functional (as judged by both cyto-toxic activity and cytokine production [Fig. 5C]) CD8� T cellsin LAT� TG. Thus, our original hypothesis that one mecha-nism by which LAT enhances reactivation is by reducing thenumber of CD8� T cells in the TG was correct, as long as wereplace “CD8� T cells” with “HSV-specific functional CD8� Tcells.”
In well-studied CD8� T-cell exhaustion systems (6, 21, 49,73), the exhaustion appears to results from overstimulation ofCD8� T cells by the presence of sustained high-level Ag. Thisseems unlikely to account for the increased CD8� T-cell ex-haustion in TG of mice latently infected with LAT� viruscompared mice latently infected with LAT� virus. Studiesfrom several labs indicate that LAT suppresses lytic gene tran-scription during acute and latent infection (14, 27, 77). If thisis correct, then there would be less viral Ag with LAT� virus,and this could not account for the increased CD8� T-cellexhaustion. In contrast, elegant reports from the Margolis lab(23, 24, 45, 80), using immunostaining to examine sufficientsections from each mouse TG to allow thorough examinationof the entire TG for HSV-1 Ags, found more HSV-1 Ag in TGfrom mice latently infected with LAT� compared to LAT�
virus. This result could help explain our finding of more ex-hausted CD8� T cells in LAT� TG. However, the amount ofviral Ag was quite low—less than 1 positive neuron/TG withLAT� virus. We have also found less than 1 positive neu-ron/TG with LAT� virus and less with LAT� virus, doingsimilar but less rigorous studies (2). Even when we extendedthe window for positive neurons to include even those withminimal staining, only 76 neurons in 60 LAT� TG and only 21neurons in 88 LAT� TG were scored as positive (not shown).In addition, if spontaneous reactivation occurs in mouse TG, itis minimal (28). Thus, even though CD8� T cells are muchmore sensitive to Ag than the antibodies used for immuno-staining, the very low unsustained Ag level that appears to bethe situation in mouse TG is unlikely to result in exhaustion ofCD8� T cells, which in other systems requires high levels ofcontinuous Ag exposure (6, 21, 49, 73). Thus, it seems unlikelythat the CD8� T-cell exhaustion detected was due to viral Agload, unless additional factors contributed to immune stimu-lation. For example, TG resident HSV-specific CD8� T cellscould have a higher functional avidity (ability to respond to lowepitope density) than their counterparts in the periphery (42).
Alternatively, CD8� T-cell exhaustion may suggest that thereis a lot more viral Ag present in TG of mice latently infectedwith LAT� virus than has previously been detected or thatthere is a lot of undetected viral reactivation in TG of micelatently infected with LAT� viruses.
Various studies, including ours, have estimated that theamount of latency with LAT� viruses is anywhere from notsignificantly different from that of LAT� virus to as high as6-fold more than that of LAT� virus, with an �2-fold increaseof LAT� to LAT� being common (55, 61, 71). In an indepen-dent study done as part of this report, we found an approxi-mately 2-fold increase in HSV-1 genomic DNA levels by qRT-PCR in TG of mice latently infected with LAT� versus LAT�
virus (data not shown). Thus, the above described exhaustionof TG resident HSV-specific CD8� T cells in LAT� TG wasassociated with an average �2-fold increase in the viral ge-nome copy number. Note that this increased latent HSV-1genome load does not imply increased viral Ag load. In addi-tion, as discussed in the previous paragraph, the amount ofviral Ag detectable by immunostaining in LAT� and LAT�
TG is probably too low, by itself, to account for the CD8 T-cellexhaustion.
We have previously shown that for acute infection with wtMcKrae and dLAT2903 (the LAT� and LAT� viruses usedhere), levels of virus replication in eyes and TG are identicalboth kinetically and for peak titers (55, 56). Since day 11represents the end of the acute infection, the CD8� T-cellresponse at this time is likely due to the acute infection and notthe amount of latent viral DNA in the TG on day 11. The onlydifference between LAT� and LAT� virus during the acuteinfection is the presence of LAT. This suggests that LAT�
neurons may play a role in modulating CD8� T-cell responsesin TG. This was supported by our in vitro finding that LAT�
neuroblastoma cells had elevated levels of PD-L1 and MHC-I.Our finding that neuroblastoma cells expressing LAT had
elevated MHC class I and elevated PD-L1 (the ligand forPD-1) may be relevant to why there was increased CD8� T-cellexhaustion in TG of mice latently infected with LAT� com-pared to LAT� viruses. Increased levels of MHC-I would beexpected to increase presentation of viral Ag to CD8� T cells.Thus, responding CD8� T cells in LAT� TG might be “seeing”more viral antigen because of an ability to interact with in-fected MHC-Ihigh neuronal cells. Even very low Ag levelsmight then be capable of leading to CD8� T-cell exhaustion. Inaddition, elevated PD-L1 would result in increased binding toPD-1 on CD8� T cells, which in turn could trigger exhaustion.Thus, it is possible that the continued presence of LAT duringlatency acts as just described and/or through some additionalyet to be determined mechanism(s) to decrease the function ofCD8� T cells in the TG. This decreased CD8� T-cell functionwould in turn help neurons in which virus is beginning toreactivate to avoid being killed by HSV-1-specific CD8� Tcells, resulting in increased HSV-1 reactivation. Such a sce-nario may be one mechanism by which the HSV-1 LAT geneworks to increase successful viral reactivation from latency,thus allowing for recurrent ocular infection, recurrent ocularvirus shedding in tears, and viral spread to new hosts. Consis-tent with this, we previously showed that neuronally derivedcell lines expressing LAT were resistant to cytotoxic CD8 T-cell killing (37).
VOL. 85, 2011 CD8� T-CELL EXHAUSTION IN LATENT HERPESVIRUS INFECTION 9135

We examined several different LAT� and LAT� mutantsfrom two different strains of HSV-1. These results showed thatour findings were not virus strain dependent. In addition, sinceone of the LAT� viruses expresses just the first 1.5 kb of theprimary 8.3-kb LAT transcript, our results indicate that theability of LAT to directly or indirectly interfere with CD8�
T-cell function in latently infected TG resides within the first20% of the LAT gene. This region of LAT is known to beresponsible for LAT’s antiapoptosis activity. Whether and howthis may be involved in how LAT interferes with CD8� T-cellfunction remains to be determined. Additionally, this region ofLAT does not appear to encode an abundant protein, althoughlow levels of one small protein appear to be expressed (31).Also note that CD4� T cells appeared unaffected in LAT� TGcompared to LAT� TG, indicating that the effect of LAT wasCD8� T-cell specific.
The HSV-gB498–505-epitope and B6 mice were chosen todetect HSV-1 specific CD8� T cells in this study because in thismouse strain the majority (over 50%) of CD8� T cells in theperiphery as well as in latently infected TG are directed to thissingle immunodominant epitope (7, 9, 19). Whether the func-tions of CD8� T cells specific to other mouse HSV-1 epitopesor human CD8� T-cell epitopes in humans are also affected byLAT expression remains to be determined. Using our HLAtransgenic rabbit model (16, 20), we are currently in the pro-cess of assessing whether LAT affects the number and/or func-tion of CD8� T cells specific to the set of human CD8� T-cellHSV-1 epitopes that we have identified as being strongly rec-ognized by these animals.
HSV-1 has a broad tropism and high degree of infectivity,but during latency, sensory neurons are the main cell typeharboring the virus. The number and type (i.e., neurons, den-dritic cells, etc.) of TG cells that are infected will likely have asignificant impact on the total antigenic burden seen by re-sponding CD8� T cells. In this study, we found an increase inMHC-I and PD-L1 expression on LAT-transfected neuroblas-toma cells, which may reduce the function of CD8� T cells.Furthermore, we observed a significant increase in the level ofPD-L1 expressed in total TG from mice latently infected withLAT� compared to LAT� viruses (Fig. 6). Following PD-L1/PD-L2 ligation, PD-1 inhibits activation and expansion ofCD8� T cells, rendering them dysfunctional (81). Under invivo conditions, different neighboring cell types, such as stro-mal cells (50), microglial cells (3), and even dendritic cells (40),may also contribute to local concentrations of antigen-MHCcomplexes and influence the function of HSV-specific CD8� Tcells in latently infected TG. In addition, it is likely that otherinhibitory signals, such as Galectin9-TIM3 ligand (29, 52) orIL-10 (4, 11, 49), or local concentrations of antigen-MHCcomplexes can influence CD8� T-cell responses. This couldexplain why in these studies the amount of functional exhaus-tion appeared higher than the amount of phenotypic exhaus-tion based on PD-1 alone. In addition, since LAT is expressedin mouse TG during and throughout the acute infection andlatency, it is possible that LAT could impact the immune re-sponse in the TG within a few days of ocular infection.
In summary, we show here for the first time that TG frommice latently infected with LAT� viruses have more exhaustedHSV-specific CD8� T cells and fewer fully functional HSV-specific CD8� T cells than do TG from mice latently infected
with LAT� virus. Since functional HSV-specific CD8� T cellsappear to be important in decreasing reactivation from latency(42), the higher level of such cells during LAT� latency mayhelp explain why LAT� viruses have reduced reactivation com-pared to wild-type LAT� viruses. To our knowledge, the pres-ent study is also the first to report that LAT expression, in theabsence of other virus genes, can produce elevated MHC-I andPD-L1, both of which could logically lead to generation ofdysfunctional HSV-specific CD8� T cells in latently infectedTG. This would represent a novel immune evasion mechanismwhereby LAT decreases HSV-specific CD8� T-cell numberand function in latently infected TG, thus allowing more reac-tivation. Blockade of the PD-1 pathway in combination withtherapeutic vaccination may have great therapeutic promise.
ACKNOWLEDGMENTS
This work was supported by Public Health Service NIH grantsEY14900, EY14017, and EY019896 to L.B.M., EY013191 andEY018171 to S.L.W., The Discovery Eye Foundation, The Henry L.Guenther Foundation, and a Research to Prevent Blindness Challengegrant. L.B.M. is an RPB Special Award Investigator. S.L.W. is an RPBSenior Scientific Investigator.
We thank Nigel W. Fraser for providing HSV-1 strain 17 and de-rived mutants and Alison Deckhut Augustine and Amy K. Stout fromthe NIH Tetramer Facility for providing tetramers used in this study.
REFERENCES
1. Ahmed, M., M. Lock, C. G. Miller, and N. W. Fraser. 2002. Regions of theherpes simplex virus type 1 latency-associated transcript that protect cellsfrom apoptosis in vitro and protect neuronal cells in vivo. J. Virol. 76:717–729.
2. Allen, S. R., et al. 2011. The role of LAT in increased CD8� T cell exhaus-tion in trigeminal ganglia of mice latently infected with herpes simplex virustype 1. J. Virol. 85:4184–4197.
3. Aravalli, R. N., S. Hu, T. N. Rowen, G. Gekker, and J. R. Lokensgard. 2006.Differential apoptotic signaling in primary glial cells infected with herpessimplex virus 1. J. Neurovirol. 12:501–510.
4. Barber, D. L., et al. 2006. Restoring function in exhausted CD8 T cells duringchronic viral infection. Nature 439:682–687.
5. Bettahi, I., et al. 2007. Protective immunity against ocular herpes infectionand disease induced by highly immunogenic self-adjuvanting glycoprotein Dlipopeptide vaccines. Invest. Ophthalmol. Vis. Sci. 48:4643–4653.
6. Blackburn, S. D., et al. 2009. Coregulation of CD8� T cell exhaustion bymultiple inhibitory receptors during chronic viral infection. Nat. Immunol.10:29–37.
7. Blaney, J. E., Jr., et al. 1998. Immunization with a single major histocom-patibility complex class I-restricted cytotoxic T-lymphocyte recognitionepitope of herpes simplex virus type 2 confers protective immunity. J. Virol.72:9567–9574.
8. Block, T. M., et al. 1990. A herpes simplex virus type 1 latency-associatedtranscript mutant reactivates with normal kinetics from latent infection.J. Virol. 64:3417–3426.
9. Bonneau, R. H., L. A. Salvucci, D. C. Johnson, and S. S. Tevethia. 1993.Epitope specificity of H-2Kb-restricted, HSV-1-, and HSV-2-cross-reactivecytotoxic T lymphocyte clones. Virology 195:62–70.
10. Branco, F. J., and N. W. Fraser. 2005. Herpes simplex virus type 1 latency-associated transcript expression protects trigeminal ganglion neurons fromapoptosis. J. Virol. 79:9019–9025.
11. Brooks, D. G., et al. 2006. Interleukin-10 determines viral clearance orpersistence in vivo. Nat. Med. 12:1301–1309.
12. Carpenter, D., et al. 2008. Introducing point mutations into the ATGs of theputative open reading frames of the HSV-1 gene encoding the latency as-sociated transcript (LAT) reduces its anti-apoptosis activity. Microb. Pathog.44:98–102.
13. Carpenter, D., et al. 2007. Stable cell lines expressing high levels of theherpes simplex virus type 1 LAT are refractory to caspase 3 activation andDNA laddering following cold shock induced apoptosis. Virology 369:12–18.
14. Chen, S. H., M. F. Kramer, P. A. Schaffer, and D. M. Coen. 1997. A viralfunction represses accumulation of transcripts from productive-cycle genesin mouse ganglia latently infected with herpes simplex virus. J. Virol. 71:5878–5884.
15. Chentoufi, A. A., and L. BenMohamed. 2010. Future viral vectors for thedelivery of asymptomatic herpes epitopes-based immunotherapeutic vac-cines. Future Virol. 5:1–4.
9136 CHENTOUFI ET AL. J. VIROL.

16. Chentoufi, A. A., et al. 2010. A novel HLA (HLA-A*0201) transgenic rabbitmodel for preclinical evaluation of human CD8� T cell epitope-based vac-cines against ocular herpes. J. Immunol. 184:2561–2571.
17. Chentoufi, A. A., et al. 2010. Nasolacrimal duct closure modulates ocularmucosal and systemic CD4(�) T-cell responses induced following topicalocular or intranasal immunization. Clin. Vaccine Immunol. 17:342–353.
18. Chentoufi, A. A., et al. 2008. HLA-A*0201-restricted CD8� cytotoxic Tlymphocyte epitopes identified from herpes simplex virus glycoprotein D.J. Immunol. 180:426–437.
19. Cose, S. C., J. M. Kelly, and F. R. Carbone. 1995. Characterization of diverseprimary herpes simplex virus type 1 gB-specific cytotoxic T-cell responseshowing a preferential V beta bias. J. Virol. 69:5849–5852.
20. Dasgupta, G., A. A. Chentoufi, A. B. Nesburn, S. L. Wechsler, and L.BenMohamed. 2009. New concepts in herpes simplex virus vaccine develop-ment: notes from the battlefield. Expert Rev. Vaccines 8:1023–1035.
21. Day, C. L., et al. 2006. PD-1 expression on HIV-specific T cells is associatedwith T-cell exhaustion and disease progression. Nature 443:350–354.
22. Devi-Rao, G. B., et al. 1997. Herpes simplex virus genome replication andtranscription during induced reactivation in the rabbit eye. J. Virol. 71:7039–7047.
23. Ellison, A. R., L. Yang, C. Voytek, and T. P. Margolis. 2000. Establishmentof latent herpes simplex virus type 1 infection in resistant, sensitive, andimmunodeficient mouse strains. Virology 268:17–28.
24. Feldman, L. T., et al. 2002. Spontaneous molecular reactivation of herpessimplex virus type 1 latency in mice. Proc. Natl. Acad. Sci. U. S. A. 99:978–983.
25. Fourcade, J., et al. 2010. Upregulation of Tim-3 and PD-1 expression isassociated with tumor antigen-specific CD8� T cell dysfunction in mela-noma patients. J. Exp. Med. 207:2175–2186.
26. Freeman, G. J., et al. 2000. Engagement of the PD-1 immunoinhibitoryreceptor by a novel B7 family member leads to negative regulation of lym-phocyte activation. J. Exp. Med. 192:1027–1034.
27. Garber, D. A., P. A. Schaffer, and D. M. Knipe. 1997. A LAT-associatedfunction reduces productive-cycle gene expression during acute infection ofmurine sensory neurons with herpes simplex virus type 1. J. Virol. 71:5885–5893.
28. Gebhardt, B. M., and W. P. Halford. 2005. Evidence that spontaneousreactivation of herpes virus does not occur in mice. Virol. J. 2:67.
29. Golden-Mason, L., et al. 2009. Negative immune regulator Tim-3 is overex-pressed on T cells in hepatitis C virus infection and its blockade rescuesdysfunctional CD4� and CD8� T cells. J. Virol. 83:9122–9130.
30. Ha, S. J., et al. 2008. Enhancing therapeutic vaccination by blocking PD-1-mediated inhibitory signals during chronic infection. J. Exp. Med. 205:543–555.
31. Henderson, G., T. Jaber, D. Carpenter, S. L. Wechsler, and C. Jones. 2009.Identification of herpes simplex virus type 1 proteins encoded within the first1.5 kb of the latency-associated transcript. J. Neurovirol. 15:439–448.
32. Hill, J. M., F. Sedarati, R. T. Javier, E. K. Wagner, and J. G. Stevens. 1990.Herpes simplex virus latent phase transcription facilitates in vivo reactiva-tion. Virology 174:117–125.
33. Hoshino, Y., L. Pesnicak, J. I. Cohen, and S. E. Straus. 2007. Rates ofreactivation of latent herpes simplex virus from mouse trigeminal ganglia exvivo correlate directly with viral load and inversely with number of infiltrat-ing CD8� T cells. J. Virol. 81:8157–8164.
34. Hufner, K., et al. 2006. Latency of alpha-herpes viruses is accompanied by achronic inflammation in human trigeminal ganglia but not in dorsal rootganglia. J. Neuropathol Exp. Neurol. 65:1022–1030.
35. Inman, M., et al. 2001. Region of herpes simplex virus type 1 latency-associated transcript sufficient for wild-type spontaneous reactivation pro-motes cell survival in tissue culture. J. Virol. 75:3636–3646.
36. Ishida, Y., Y. Agata, K. Shibahara, and T. Honjo. 1992. Induced expressionof PD-1, a novel member of the immunoglobulin gene superfamily, uponprogrammed cell death. EMBO J. 11:3887–3895.
37. Jiang, X., et al. 2011. The herpes simplex virus type 1 latency associatedtranscript can protect neuronal derived C1300 and Neuro2A cells fromgranzyme B induced apoptosis and CD8 T-cell killing. J. Virol. 85:2325–2332.
38. Jin, H. T., et al. 2010. Cooperation of Tim-3 and PD-1 in CD8 T-cellexhaustion during chronic viral infection. Proc. Natl. Acad. Sci. U. S. A.107:14733–14738.
39. Jin, L., et al. 2003. Identification of herpes simplex virus type 1 latency-associated transcript sequences that both inhibit apoptosis and enhance thespontaneous reactivation phenotype. J. Virol. 77:6556–6561.
40. Kather, A., et al. 2010. Herpes simplex virus type 1 (HSV-1)-induced apop-tosis in human dendritic cells as a result of downregulation of cellularFLICE-inhibitory protein and reduced expression of HSV-1 antiapoptoticlatency-associated transcript sequences. J. Virol. 84:1034–1046.
41. Klenerman, P., and A. Hill. 2005. T cells and viral persistence: lessons fromdiverse infections. Nat. Immunol. 6:873–879.
42. Knickelbein, J. E., et al. 2008. Noncytotoxic lytic granule-mediated CD8� Tcell inhibition of HSV-1 reactivation from neuronal latency. Science 322:268–271.
43. Leib, D. A., et al. 1989. A deletion mutant of the latency-associated transcriptof herpes simplex virus type 1 reactivates from the latent state with reducedfrequency. J. Virol. 63:2893–2900.
44. Maggioncalda, J., A. Mehta, Y. H. Su, N. W. Fraser, and T. M. Block. 1996.Correlation between herpes simplex virus type 1 rate of reactivation fromlatent infection and the number of infected neurons in trigeminal ganglia.Virology 225:72–81.
45. Margolis, T. P., et al. 2007. Spontaneous reactivation of herpes simplex virustype 1 in latently infected murine sensory ganglia. J. Virol. 81:11069–11074.
46. Mehta, A., et al. 1995. In situ DNA PCR and RNA hybridization detectionof herpes simplex virus sequences in trigeminal ganglia of latently infectedmice. Virology 206:633–640.
47. Miyajima, K., M. B. Knickelbein, and A. Nakajima. 2008. Stern-Gerlachstudy of multidecker lanthanide-cyclooctatetraene sandwich clusters. J. Phys.Chem. A 112:366–375.
48. Mott, K. R., et al. 2009. Level of herpes simplex virus type 1 latency corre-lates with severity of corneal scarring and exhaustion of CD8� T cells intrigeminal ganglia of latently infected mice. J. Virol. 83:2246–2254.
49. Mueller, S. N., and R. Ahmed. 2009. High antigen levels are the cause of Tcell exhaustion during chronic viral infection. Proc. Natl. Acad. Sci. U. S. A.106:8623–8628.
50. Nagahara, K., et al. 2008. Galectin-9 increases Tim-3� dendritic cells andCD8� T cells and enhances antitumor immunity via galectin-9-Tim-3 inter-actions. J. Immunol. 181:7660–7669.
50a.National Research Council. 1996. Guide for the care and use of laboratoryanimals. National Academy Press, Washington, DC.
51. Nesburn, A. B., et al. 2007. Functional Foxp3� CD4� CD25(Bright�)“natural” regulatory T cells are abundant in rabbit conjunctiva and suppressvirus-specific CD4� and CD8� effector T cells during ocular herpes infec-tion. J. Virol. 81:7647–7661.
52. Oikawa, T., et al. 2006. Preferential involvement of Tim-3 in the regulationof hepatic CD8� T cells in murine acute graft-versus-host disease. J. Im-munol. 177:4281–4287.
53. Peng, W., et al. 2005. The locus encompassing the latency-associated tran-script of herpes simplex virus type 1 interferes with and delays interferonexpression in productively infected neuroblastoma cells and trigeminal gan-glia of acutely infected mice. J. Virol. 79:6162–6171.
54. Perng, G. C., et al. 1996. The region of the herpes simplex virus type 1 LATgene that is colinear with the ICP34.5 gene is not involved in spontaneousreactivation. J. Virol. 70:282–291.
55. Perng, G. C., et al. 1994. The latency-associated transcript gene of herpessimplex virus type 1 (HSV-1) is required for efficient in vivo spontaneousreactivation of HSV-1 from latency. J. Virol. 68:8045–8055.
56. Perng, G. C., et al. 2001. Three herpes simplex virus type 1 latency-associatedtranscript mutants with distinct and asymmetric effects on virulence in micecompared with rabbits. J. Virol. 75:9018–9028.
57. Perng, G. C., H. Ghiasi, R. Kaiwar, A. B. Nesburn, and S. L. Wechsler. 1994.An improved method for cloning portions of the repeat regions of herpessimplex virus type 1. J. Virol. Methods 46:111–116.
58. Perng, G. C., H. Ghiasi, S. M. Slanina, A. B. Nesburn, and S. L. Wechsler.1996. The spontaneous reactivation function of the herpes simplex virus type1 LAT gene resides completely within the first 1.5 kilobases of the 8.3-kilobase primary transcript. J. Virol. 70:976–984.
59. Perng, G. C., et al. 2000. Virus-induced neuronal apoptosis blocked by theherpes simplex virus latency-associated transcript. Science 287:1500–1503.
60. Perng, G. C., et al. 2002. A gene capable of blocking apoptosis can substitutefor the herpes simplex virus type 1 latency-associated transcript gene andrestore wild-type reactivation levels. J. Virol. 76:1224–1235.
61. Perng, G. C., et al. 2000. The latency-associated transcript gene enhancesestablishment of herpes simplex virus type 1 latency in rabbits. J. Virol.74:1885–1891.
62. Petrovas, C., et al. 2007. SIV-specific CD8� T cells express high levels ofPD1 and cytokines but have impaired proliferative capacity in acute andchronic SIVmac251 infection. Blood 110:928–936.
63. Ramakrishnan, R., M. Levine, and D. J. Fink. 1994. PCR-based analysis ofherpes simplex virus type 1 latency in the rat trigeminal ganglion establishedwith a ribonucleotide reductase-deficient mutant. J. Virol. 68:7083–7091.
64. Ramakrishnan, R., P. L. Poliani, M. Levine, J. C. Glorioso, and D. J. Fink.1996. Detection of herpes simplex virus type 1 latency-associated transcriptexpression in trigeminal ganglia by in situ reverse transcriptase PCR. J. Virol.70:6519–6523.
65. Rodrigue-Gervais, I. G., et al. 2010. Dendritic cell inhibition is connected toexhaustion of CD8� T cell polyfunctionality during chronic hepatitis C virusinfection. J. Immunol. 184:3134–3144.
66. Sakuishi, K., et al. 2010. Targeting Tim-3 and PD-1 pathways to reverse Tcell exhaustion and restore anti-tumor immunity. J. Exp. Med. 207:2187–2194.
67. Sawtell, N. M., and R. L. Thompson. 1992. Rapid in vivo reactivation ofherpes simplex virus in latently infected murine ganglionic neurons aftertransient hyperthermia. J. Virol. 66:2150–2156.
68. Shimeld, C., et al. 1995. Immune cell infiltration and persistence in the
VOL. 85, 2011 CD8� T-CELL EXHAUSTION IN LATENT HERPESVIRUS INFECTION 9137

mouse trigeminal ganglion after infection of the cornea with herpes simplexvirus type 1. J. Neuroimmunol. 61:7–16.
69. Simmons, A., and D. C. Tscharke. 1992. Anti-CD8 impairs clearance ofherpes simplex virus from the nervous system: implications for the fate ofvirally infected neurons. J. Exp. Med. 175:1337–1344.
70. Theil, D., et al. 2003. Latent herpesvirus infection in human trigeminalganglia causes chronic immune response. Am. J. Pathol. 163:2179–2184.
71. Thompson, R. L., and N. M. Sawtell. 1997. The herpes simplex virus type 1latency-associated transcript gene regulates the establishment of latency.J. Virol. 71:5432–5440.
72. Trousdale, M. D., et al. 1991. In vivo and in vitro reactivation impairment ofa herpes simplex virus type 1 latency-associated transcript variant in a rabbiteye model. J. Virol. 65:6989–6993.
73. Vali, B., et al. 2010. HCV-specific T cells in HCV/HIV co-infection showelevated frequencies of dual Tim-3/PD-1 expression that correlate with liverdisease progression. Eur. J. Immunol. 40:2493–2505.
74. van Baarle, D., et al. 2008. Progressive telomere shortening of Epstein-Barrvirus-specific memory T cells during HIV infection: contributor to exhaus-tion? J. Infect. Dis. 198:1353–1357.
75. van Lint, A. L., et al. 2005. Latent infection with herpes simplex virus isassociated with ongoing CD8� T-cell stimulation by parenchymal cellswithin sensory ganglia. J. Virol. 79:14843–14851.
76. Verjans, G. M., et al. 2007. Selective retention of herpes simplex virus-specific T cells in latently infected human trigeminal ganglia. Proc. Natl.Acad. Sci. U. S. A. 104:3496–3501.
77. Wang, Q. Y., et al. 2005. Herpesviral latency-associated transcript genepromotes assembly of heterochromatin on viral lytic-gene promoters in la-tent infection. Proc. Natl. Acad. Sci. U. S. A. 102:16055–16059.
78. Wherry, E. J., J. N. Blattman, K. Murali-Krishna, R. van der Most, and R.Ahmed. 2003. Viral persistence alters CD8 T-cell immunodominance andtissue distribution and results in distinct stages of functional impairment.J. Virol. 77:4911–4927.
79. Wherry, E. J., et al. 2007. Molecular signature of CD8� T cell exhaustionduring chronic viral infection. Immunity 27:670–684.
80. Yang, L., C. C. Voytek, and T. P. Margolis. 2000. Immunohistochemicalanalysis of primary sensory neurons latently infected with herpes simplexvirus type 1. J. Virol. 74:209–217.
81. Yao, S., and L. Chen. 2006. Reviving exhausted T lymphocytes duringchronic virus infection by B7-H1 blockade. Trends Mol. Med. 12:244–246.
82. Zhang, X., et al. 2005. Th-cytotoxic T-lymphocyte chimeric epitopes ex-tended by Nε-palmitoyl lysines induce herpes simplex virus type 1-specificeffector CD8� Tc1 responses and protect against ocular infection. J. Virol.79:15289–15301.
9138 CHENTOUFI ET AL. J. VIROL.




















