
THE GENUS STEINMANELLA (BIVALVIA, TRIGONIOIDA) IN THE LOWER MEMBER OF THE AGRIO FORMATION (LOWER...
17
1069 J. Paleont., 77(6), 2003, pp. 1069–1085 Copyright q 2003, The Paleontological Society 0022-3360/03/0077-1069$03.00 THE GENUS STEINMANELLA (BIVALVIA, TRIGONIOIDA) IN THE LOWER MEMBER OF THE AGRIO FORMATION (LOWER CRETACEOUS), NEUQUE ´ N BASIN, ARGENTINA DARIO G. LAZO Departamento de Ciencias Geolo ´gicas, Universidad de Buenos Aires, Ciudad Universitaria, 1428 Buenos Aires, Argentina, CONICET, ,[email protected]. ABSTRACT—Myophorellid bivalves (Trigonioida, Myophorellacea) with conspicuously tuberculate shells are abundant in Lower Cre- taceous rocks of the Neuque ´n Basin represented by the Gondwanian genus Steinmanella Crickmay, 1930. Publications concerning Neuque ´n trigonioids deal only with taxonomy, and until now the sedimentary environments of the Steinmanella bearing facies have not been interpreted. The present study analyzes morphology, taxonomy, stratigraphy, and paleobiology of Steinmanella species collected from five sections of the Lower Member of the Agrio Formation, including the type locality. Two species are differentiated: Steinmanella pehuenmapuensis (Leanza, 1998) from the lower part of the member (Pseudofavrella angulatiformis Zone) and Steinmanella transitoria (Steinmann, 1881) from the middle and upper part of the member (Holcoptychites neuquensis, Hoplitocrioceras gentilii, and Weaver- iceras vacaensis Zones). Specimens of both species have been recorded in intermediate to high-energy shoreface and low-energy offshore deposits. The inferred paleoecologic model is that Steinmanella lived semi-infaunally and burrowed on sandy substrates in the shoreface and on muddy substrates in the offshore. Colonization of low-energy subenvironments in the offshore probably occurred during increased sea- floor oxygenation and reduced net sedimentation or brief omission periods. INTRODUCTION T HE TRIGONIOIDS constitute a monophyletic group of marine bivalves that have been morphologically distinct since the Middle Ordovician (Pojeta, 1978). They radiated during the Me- sozoic (Stanley, 1977), especially during the Jurassic and Creta- ceous, and underwent a rapid decline at the Cretaceous-Tertiary boundary. During the Cenozoic, the group was restricted to Aus- tralia and New Guinea and represented only by two genera, Eotri- gonia (Upper Paleocene-Miocene) and Neotrigonia (Middle Mio- cene-Recent) which contains all the living trigonioid species (Fleming, 1964; Darragh, 1986). The main trigonioid diagnostic characters are: 1) burrowing life habit; 2) bilateral symmetry; 3) tendency to the trigonal shape; 4) absence of siphons (except some Upper Paleozoic examples), free mantle margins, and entire pallial line; 5) filibranchs and suspen- sion feeding in Recent species; 6) external opisthodetic parivin- cular ligament on nymph; 7) two or three strong and stable hinge teeth; 8) isomyarian condition; 9) completely aragonitic shell in- cluding the internal fibrous layer of the ligament (Taylor et al., 1969; Newell and Boyd, 1975). The relationships of trigonioids with other groups are poorly known. Traditionally, these bivalves were grouped into the Su- perfamily Trigoniacea, which included five families: Lyrodes- matidae?, Myophoriidae, Scaphellinidae?, Trigoniidae, and Tri- gonioididae? (Newell and Boyd, 1965; Cox, 1969b). This subdi- vision seems inappropriate because it implies radiation devoid of phyletic lineages and five families equally related. Thus, Cooper (1991) made a phylogenetic analysis of the whole order and iden- tified two suborders: Trigoniina (paraphyletic) and Myophorellina (monophyletic), and four superfamilies: Myophoriacea, Trigoni- acea, Myophorellacea, and Megatrigoniacea (Fig. 1.1). According to this suprageneric classification, the Superfamily Myophorella- cea includes the myophorellids with nodose and mainly oblique flank ribs. The Myophorellidae in turn are subdivided into the Subfamilies Myophorellinae (trigonal-suborbicular, gracile shell) and Steinmanellinae (quadrate, massive and robust shell) (Fig. 1.2). The later group was originally defined by Steinmann (1882) under the name Pseudoquadratae. It has been typified by Stein- manella (Transitrigonia) transitoria (Steinmann, 1881) from South America and S.(Macrotrigonia) herzogi (Goldfuss, 1837) from South Africa. Steinmanellinae ranges from the Upper Juras- sic to the Upper Cretaceous, and contains the genus Steinmanella Crickmay, 1930, and now includes also the North Pacific genus Yaadia Crickmay, 1930 (Cooper, 1991). The stratigraphic record of the genus Steinmanella ranges from the Upper Jurassic up to the Lower Cretaceous (Camacho and Oliv- ero, 1985; Cooper, 1991). It has been described from South Amer- ica (Argentina, Chile, and Peru ´), South and East Africa, and India (Fig. 2). There is no record from Antarctica, Australia, and New Zealand (Kelly, 1995a–1995b). Thus, the Steinmanella records are restricted to Southwest Gondwana and primarily are derived from Lower Cretaceous rocks (Camacho and Olivero, 1985). In Argentina, the species of Steinmanella occur in two Mesozoic basins located in the Andes foothills in Patagonia: the Neuque ´n Basin (368–408S) and the Austral Basin (448–558S) (Fig. 3). It is interesting to note that despite the fact that these closely situated basins being open to the Pacific Ocean at the same time during the Mesozoic (separated only by a non marine gap of 250 km), there are no known Steinmanella affinities between these basins. In fact, there is an overall lack of faunal affinities between their coeval marine invertebrate fauna (Aguirre-Urreta and Rawson, 1998). Although the presence in Argentina of the genus Steinmanella is unquestioned, the stratigraphic distribution of the several de- scribed species is poorly documented. Also, many of these species are based on a few specimens that do not account for intra-pop- ulation variability and ontogenetic change. They have been col- lected and studied since the end of the nineteenth century (Beh- rendsen, 1892; Burckhardt, 1900, 1903; Weaver, 1931; Leanza, 1941, 1993, 1998; Lambert, 1944; Leanza and Castellaro, 1955; Levy de Caminos, 1969; Damborenea et al., 1979; Aguirre-Urreta and Ramos, 1981; Camacho and Olivero, 1985; Leanza and Gar- ate Zubillaga, 1987; Lo Forte, 1988). The great majority of the previous works focused only on taxonomy or merely mention the bivalves, and there is little information about the facies containing them and the paleoenvironment where they lived has not been interpreted. The objectives of the present contribution are: 1) describe the species of Steinmanella present in the Lower Member of the Agrio Formation in the Neuque ´n Basin based on new material; 2) study the facies distribution of these bivalves along one de- tailed stratigraphic profile of the Lower Member of the Agrio
Transcript of THE GENUS STEINMANELLA (BIVALVIA, TRIGONIOIDA) IN THE LOWER MEMBER OF THE AGRIO FORMATION (LOWER...

1069
J. Paleont., 77(6), 2003, pp. 1069–1085Copyright q 2003, The Paleontological Society0022-3360/03/0077-1069$03.00
THE GENUS STEINMANELLA (BIVALVIA, TRIGONIOIDA) IN THE LOWERMEMBER OF THE AGRIO FORMATION (LOWER CRETACEOUS),
NEUQUEN BASIN, ARGENTINADARIO G. LAZO
Departamento de Ciencias Geologicas, Universidad de Buenos Aires, Ciudad Universitaria, 1428 Buenos Aires, Argentina, CONICET,,[email protected].
ABSTRACT—Myophorellid bivalves (Trigonioida, Myophorellacea) with conspicuously tuberculate shells are abundant in Lower Cre-taceous rocks of the Neuquen Basin represented by the Gondwanian genus Steinmanella Crickmay, 1930. Publications concerningNeuquen trigonioids deal only with taxonomy, and until now the sedimentary environments of the Steinmanella bearing facies havenot been interpreted. The present study analyzes morphology, taxonomy, stratigraphy, and paleobiology of Steinmanella species collectedfrom five sections of the Lower Member of the Agrio Formation, including the type locality. Two species are differentiated: Steinmanellapehuenmapuensis (Leanza, 1998) from the lower part of the member (Pseudofavrella angulatiformis Zone) and Steinmanella transitoria(Steinmann, 1881) from the middle and upper part of the member (Holcoptychites neuquensis, Hoplitocrioceras gentilii, and Weaver-iceras vacaensis Zones).
Specimens of both species have been recorded in intermediate to high-energy shoreface and low-energy offshore deposits. Theinferred paleoecologic model is that Steinmanella lived semi-infaunally and burrowed on sandy substrates in the shoreface and onmuddy substrates in the offshore. Colonization of low-energy subenvironments in the offshore probably occurred during increased sea-floor oxygenation and reduced net sedimentation or brief omission periods.
INTRODUCTION
THE TRIGONIOIDS constitute a monophyletic group of marinebivalves that have been morphologically distinct since the
Middle Ordovician (Pojeta, 1978). They radiated during the Me-sozoic (Stanley, 1977), especially during the Jurassic and Creta-ceous, and underwent a rapid decline at the Cretaceous-Tertiaryboundary. During the Cenozoic, the group was restricted to Aus-tralia and New Guinea and represented only by two genera, Eotri-gonia (Upper Paleocene-Miocene) and Neotrigonia (Middle Mio-cene-Recent) which contains all the living trigonioid species(Fleming, 1964; Darragh, 1986).
The main trigonioid diagnostic characters are: 1) burrowing lifehabit; 2) bilateral symmetry; 3) tendency to the trigonal shape; 4)absence of siphons (except some Upper Paleozoic examples), freemantle margins, and entire pallial line; 5) filibranchs and suspen-sion feeding in Recent species; 6) external opisthodetic parivin-cular ligament on nymph; 7) two or three strong and stable hingeteeth; 8) isomyarian condition; 9) completely aragonitic shell in-cluding the internal fibrous layer of the ligament (Taylor et al.,1969; Newell and Boyd, 1975).
The relationships of trigonioids with other groups are poorlyknown. Traditionally, these bivalves were grouped into the Su-perfamily Trigoniacea, which included five families: Lyrodes-matidae?, Myophoriidae, Scaphellinidae?, Trigoniidae, and Tri-gonioididae? (Newell and Boyd, 1965; Cox, 1969b). This subdi-vision seems inappropriate because it implies radiation devoid ofphyletic lineages and five families equally related. Thus, Cooper(1991) made a phylogenetic analysis of the whole order and iden-tified two suborders: Trigoniina (paraphyletic) and Myophorellina(monophyletic), and four superfamilies: Myophoriacea, Trigoni-acea, Myophorellacea, and Megatrigoniacea (Fig. 1.1). Accordingto this suprageneric classification, the Superfamily Myophorella-cea includes the myophorellids with nodose and mainly obliqueflank ribs. The Myophorellidae in turn are subdivided into theSubfamilies Myophorellinae (trigonal-suborbicular, gracile shell)and Steinmanellinae (quadrate, massive and robust shell) (Fig.1.2). The later group was originally defined by Steinmann (1882)under the name Pseudoquadratae. It has been typified by Stein-manella (Transitrigonia) transitoria (Steinmann, 1881) fromSouth America and S. (Macrotrigonia) herzogi (Goldfuss, 1837)
from South Africa. Steinmanellinae ranges from the Upper Juras-sic to the Upper Cretaceous, and contains the genus SteinmanellaCrickmay, 1930, and now includes also the North Pacific genusYaadia Crickmay, 1930 (Cooper, 1991).
The stratigraphic record of the genus Steinmanella ranges fromthe Upper Jurassic up to the Lower Cretaceous (Camacho and Oliv-ero, 1985; Cooper, 1991). It has been described from South Amer-ica (Argentina, Chile, and Peru), South and East Africa, and India(Fig. 2). There is no record from Antarctica, Australia, and NewZealand (Kelly, 1995a–1995b). Thus, the Steinmanella records arerestricted to Southwest Gondwana and primarily are derived fromLower Cretaceous rocks (Camacho and Olivero, 1985).
In Argentina, the species of Steinmanella occur in two Mesozoicbasins located in the Andes foothills in Patagonia: the NeuquenBasin (368–408S) and the Austral Basin (448–558S) (Fig. 3). It isinteresting to note that despite the fact that these closely situatedbasins being open to the Pacific Ocean at the same time during theMesozoic (separated only by a non marine gap of 250 km), thereare no known Steinmanella affinities between these basins. In fact,there is an overall lack of faunal affinities between their coevalmarine invertebrate fauna (Aguirre-Urreta and Rawson, 1998).
Although the presence in Argentina of the genus Steinmanellais unquestioned, the stratigraphic distribution of the several de-scribed species is poorly documented. Also, many of these speciesare based on a few specimens that do not account for intra-pop-ulation variability and ontogenetic change. They have been col-lected and studied since the end of the nineteenth century (Beh-rendsen, 1892; Burckhardt, 1900, 1903; Weaver, 1931; Leanza,1941, 1993, 1998; Lambert, 1944; Leanza and Castellaro, 1955;Levy de Caminos, 1969; Damborenea et al., 1979; Aguirre-Urretaand Ramos, 1981; Camacho and Olivero, 1985; Leanza and Gar-ate Zubillaga, 1987; Lo Forte, 1988). The great majority of theprevious works focused only on taxonomy or merely mention thebivalves, and there is little information about the facies containingthem and the paleoenvironment where they lived has not beeninterpreted.
The objectives of the present contribution are: 1) describe thespecies of Steinmanella present in the Lower Member of theAgrio Formation in the Neuquen Basin based on new material;2) study the facies distribution of these bivalves along one de-tailed stratigraphic profile of the Lower Member of the Agrio

1070 JOURNAL OF PALEONTOLOGY, V. 77, NO. 6, 2003
FIGURE 1—1, Hypothetical relationships within the proposed superfam-ilies of the Order Trigonioida. The position of the analyzed FamilyMyophorellidae is in bold; 2, Subfamilies and genera within the FamilyMyophorellidae. Cladogram from Cooper (1991).
FIGURE 2—Early Cretaceous paleogeographic distribution of the genusSteinmanella (Crickmay, 1930). This genus has been recorded inSouthwest Gondwana: Peru (Lisson, 1930), North Chile (Steinmann,1881), Central Chile (Steinmann, 1882; Philippi, 1899; Reyes Bianchiand Perez D’Angelo, 1979), South Chile (Reyes Bianchi, 1970, ReyesBianchi and Perez D’Angelo, 1979); Neuquen Basin in Argentina(Burckhardt, 1900, 1903; Weaver, 1931; A. F. Leanza, 1941; Levy deCaminos, 1969; Damborenea et al., 1979; H. A. Leanza, 1993), AustralBasin in Argentina (Camacho and Olivero, 1985), South Africa (Kitch-in, 1908; Cooper, 1991), Tanzania (Camacho and Olivero, 1985), andIndia (Kitchin, 1903; Cooper, 1979). Early Cretaceous stereographicmap from Gilbert Smith et al. (1973); paleogeographic pole in thecenter and paleoequator at the edge.
Formation at the type locality; 3) interpret the paleoenvironmentwhich the bivalves inhabited; and 4) correlate the Steinmanella-bearing beds with the associated ammonite fauna to establish abiostratigraphic framework.
This study is part of an integrated approach to understand thedepositional environments and the paleoecology of the fossil as-semblages of the Agrio Formation.
GEOLOGY AND PALEONTOLOGY OF THE AGRIO FORMATION
The Neuquen Basin is an important Meso-Cenozoic depocenterlocated in West-Central Argentina (Fig. 3.2), and surrounded bythe North Patagonian massif, the Sierra Pintada, and the Andeanvolcanic arc. The tectonic and stratigraphic evolution of this basinbegan during the Late Triassic and continued until the Late Ter-tiary (Vergani et al., 1995).
During the Jurassic and Early Cretaceous, thick marine and con-tinental deposits accumulated due to periodic transgressions of thePacific Ocean. Particularly in the Late Valanginian, a relative risein sea level extended the coastline quickly toward the continent,forming the Neuquen Embayment. During this transgression, therocks of the Agrio Formation (Weaver, 1931) were deposited (Le-garreta and Gulisano, 1989). This unit crops out extensively in theNeuquen Province as a series of meridian anticlines along the AgrioFold and Thrust Belt (Bracaccini, 1970).
The Agrio Formation is a 1,200 m sequence and is subdividedinto three members: Lower, Middle (or Avile Sandstone), andUpper members (Weaver, 1931). The Upper and Lower membersconsist of intercalated shales, sandstones, and bioclastic lime-stones of marine origin. The 20 to 100 m Middle Member is acontinental sandstone that accumulated discontinuously during aregression in the middle Hauterivian (Gulisano and GutierrezPleimling, 1988; Ryer, 1991).
The Lower and Upper members of the Agrio Formation repre-sent a mixed clastic-carbonate sedimentation and was interpretedas shoreface to offshore paleoenvironments on a storm-dominated,shallow-marine ramp (Brinkmann, 1994; Spalletti et al., 2001).
Weaver (1931) first described in detail the abundant fauna of theAgrio Formation. It is composed principally of marine invertebrates
(bivalves, gastropods, nautiloids, ammonites, corals, crustaceans,echinoderms and serpulids), diverse trace-fossil assemblages, andscarce vertebrate remains (elasmosaurid plesiosaurs and pycnodon-tid fishes). Subsequent paleontological studies focused mainly onammonoid biostratigraphy (Leanza, 1945; Leanza and Wiedmann,1980, 1992; Aguirre-Urreta, 1998; Aguirre-Urreta and Rawson,1997, 1998, 1999, 2001a–2001b) and trigonioid taxonomy (Leanzaand Garate Zubillaga, 1987; Leanza, 1993).
The study area is located in central-north Neuquen Provincebetween Chos Malal and Las Lajas (Fig. 3.2). In this area, theAgrio Formation is well exposed. From north to south, the local-ities where Steinmanella material has been collected are Pichai-hue, Agua de la Mula, Salado Sur, Bajada del Agrio, and CerroMesa (Fig. 3.2, localities 1–5). At Bajada del Agrio, which is thetype locality of the Agrio Formation, a bed-by-bed analysis of theLower Member was made to study the stratigraphic and faciesdistribution of Steinmanella.
STEINMANELLA FACIES AND PALEOENVIRONMENTS
Field studies of fossils, in conjunction with taphonomic andsedimentological analyses, allow us to infer paleobiologic modelsfor extinct bivalve species. Rocks containing Steinmanella bi-valves in the Bajada del Agrio section are grouped into two facies(Fig. 4). The facies are defined by grain size, sedimentary struc-tures, and paleontologic and taphonomic features. Both facies arelaterally continuous and can be easily correlated between Agua

1071LAZO—CRETACEOUS BIVALVES FROM ARGENTINA
FIGURE 3—1, Location map of the Neuquen and Austral Basins in Argentina. 2, The Neuquen Basin in West-Central Argentina showing the locationof the five fossil localities (shaded region): 1, Pichaihue; 2, Agua de la Mula; 3, Salado Sur; 4, Bajada del Agrio (type locality of the AgrioFormation); 5, Cerro Mesa. In locality 4 a bed-by-bed analysis of the Lower Member of the Agrio Formation was done. Base map from DiGregorioet al. (1984).
de la Mula, Salado Sur, and Bajada del Agrio (Fig. 3). Generalizedenvironmental interpretations are made for each facies.
DARK-GRAY CLAYSTONE FACIES
Description.Dark gray to medium dark gray, mainly massiveclaystone. It forms units 1 to 45 m thick. It is the dominant li-thology in Bajada del Agrio. It is overlain by and grades intosiltstone, sandstone or lenticular to flaser bedding. Sedimentarystructures are poorly developed. Fissility is partly recorded. Fine-grained, carbonate concretions are common and form discrete lev-els. Their shape can vary from flattened to ovoid and their di-ameter ranges from 3 to 30 cm. Full-bodied fossils are commonlyfound in concretions.
Fossils are sparse and locally abundant. They occur as thin shellbeds (0.05 to 0.1 m) and pavements. Shell beds are fine-grained,internally homogenous, and dense to disperse packed. Shells areprimarily concordant, convex-down and convex-up. The degreeof articulation is high while degrees of abrasion and fragmentationare very low. Articulated valves are fill with fine-grained sedi-ment. Size-sorting is very poor, shell size ranges from 5 to 30cm. Encrustation of shells by Parsimonia antiquata (Sowerby)
and Ceratostreon sp. is common. The taxonomic compositionvaries from monospecific to polyspecific (up to seven benthic gen-era). The dominant group is suspension-feeding Bivalvia followedby ammonoids, gastropods, and serpulid worm tubes. The mostabundant bivalve genera are Astarte, Ceratostreon, Cucullaea,Modiolus, Panopea, and Steinmanella.
Steinmanella specimens occurred at eight levels of dark-grayclaystone facies (Fig. 4, beds labeled with *). Large and smallspecimens were present with ‘‘perfect’’ preservation quality.
Interpretation.The monotonous claystones resulted from fall-out processes interpreted as deposited in lower-energy subenvi-ronments at the offshore zone below fairweather wave base. Taph-onomic data also point to a quiet environment and indicate thatlateral transport was insignificant. The claystone’s fossil associa-tions are regarded to represent the autochthonous relict of ancientcommunities. They are time-averaged and do not reflect short-term fluctuations but general environmental conditions that per-sisted over longer time periods (Fursich and Aberhan, 1990;Aberhan, 1993).
An episodic sedimentation probably encouraged the formation

1072 JOURNAL OF PALEONTOLOGY, V. 77, NO. 6, 2003
FIGURE 4—Detailed lithological column of the Lower Member of the Agrio Formation at Bajada del Agrio locality showing the stratigraphicdistribution of Steinmanella pehuenmapuensis (Leanza, 1998) and Steinmanella transitoria (Steinmann, 1881). To simplify, only densely packedshell beds’ compositions are indicated. Left vertical scale in meters.

1073LAZO—CRETACEOUS BIVALVES FROM ARGENTINA
FIGURE 4—Continued
of concretionary horizons, which formed during pauses in depo-sition or during periods of greatly reduced sedimentation at thesediment-water interface slightly above the horizon of growth(Canfield and Raiswell, 1991).
The sparse presence of benthic organisms, abundant organiccontent (inferred from dark color), and relatively low diversityassemblages is probably related to low levels of benthic oxygen.
The Steinmanella beds are interpreted as colonization events ofthe muddy bottom that occurred periodically during increased ox-ygen levels and reduced net sedimentation or brief omission pe-riods. The low net sedimentation would have allowed benthicsuspension-feeders to filter without clogging, and perhapschanged the consistency of the substrate after its initial deposition(synsedimentary modification) with a gradual decrease in water
content allowing it to become firmer and more suitable for bur-rowing (Walker and Diehl, 1986; Wignall, 1993).
SHELL DEBRIS DOMINATED FACIES
Description.Fine to coarse gray sandstone alternating withabundant shell beds. This facies is coarsening-upwards and rangesin thickness from 0.08 to 8 m. It overlies the dark-gray claystonefacies and includes the coarsest terms of the section. Contacts aresharp and irregular. It has low to high amalgamation degree.Wave-ripple cross lamination and occasionally hummocky cross-stratification are observed. Bioturbation is usually present (Lazo,2001).
Shell beds up to 0.5 m thick are multi-event in character. Theytypically comprise highly disarticulated and highly fragmented

1074 JOURNAL OF PALEONTOLOGY, V. 77, NO. 6, 2003
FIGURE 5—Descriptive terminology and measured morphologic characters of the genus Steinmanella. 1, Left lateral view; 2, Anterior view; 3, Dorsalview; 4, Internal view of right valve. Abbreviations are as follows: L: length, H: height, W: width of both valves together, Le: length of escutcheon,Ln: length of nymph, Av: angle between the anterior and ventral margins; Pv: angle between the posterior and ventral margins.
shells of poorly size-sorted. Shell debris usually dominates theshell beds. Shells that are more complete are dispersed in thedebris. The degree of abrasion is usually high. Bioclasts havemixed orientation, but large ammonoids and ramose corals areinvariably concordant. The taxonomic composition is polyspecific(up to 15 benthic genera). The associations are dominated bysuspension-feeding bivalves, but coral colonies are locally abun-dant. Ammonoids, gastropods, and serpulid worm tubes are alsorecorded. The most abundant bivalve genera are Aphrodina, Cer-atostreon, Cucullaea, Eriphyla, Myoconcha, Panopea, Ptycho-mya, and Steinmanella.
Steinmanella specimens occurred at 15 levels of shell debrisdominated facies (Fig. 4, beds labeled with ●).
Interpretation.Sedimentologic data and abundant shell debrispoint to intermediate to high levels of energy with influence offair- and storm-weather waves. Fossil associations are regardedas time-averaged and parautochthonous to allochthonous. Con-centration of shells is largely due to physical processes.
High disarticulation, fragmentation, and abrasion indicate highenvironmental energy and turbulence and significant lateral trans-port.
Relatively high diversity associations and abundant bioturba-tion is suggestive of well-oxygenated substrate.
In sum, the shell debris dominated facies is interpreted as shal-lowing-upwards (lower to upper) shoreface deposits above fair-weather wave base influenced by storm processes.
SYSTEMATIC PALEONTOLOGY
Repository.The described and figured specimens are depos-ited in the following collection under the catalogue number:CPBA-Catedra de Paleontologıa, Departamento de Ciencias Geo-logicas, Facultad de Ciencias Exactas y Naturales, Universidad deBuenos Aires, Pabellon II, Ciudad Universitaria, 1428, BuenosAires, Argentina. Each catalogue number refers to a sample ofspecimens of given section and stratigraphic position. Suffix num-ber means the number of specimens discussed of a given cata-logue number.
Synonymy.Only those citations with new and figured speciesof Steinmanella are listed in the synonymy.
Terminology and measurements.Complex sculpture of tri-goniid bivalves have given rise to an abundance of morpholog-ical terms. Descriptive terms used in this work are those listedand defined by Cox (1969a–1969b). They are as follows: es-cutcheon is the lozenge-shaped dorsal differentiated area ex-tending posteriorly from beaks and bordered by ridge in eachvalva; escutcheon ridge or carina is the ridge extending poste-riorly from beak in each valve and forming border of escutch-eon; posterior area or corselet is the portion of the shell dorsalto the marginal carina, it is divided into a dorsal wedge or innercorselet and a ventral wedge or outer corselet by a mediangroove, it marks the respiratory margins of the animal, the outercorselet valve margins indicate the position of the inhalant cur-rent, the inner corselet valve margins indicate the position of the

1075LAZO—CRETACEOUS BIVALVES FROM ARGENTINA
TABLE 1—Measurements (in mm) of Steinmanella pehuenmapuensis (Leanza, 1998). MU: Agua de la Mula, BA: Bajada del Agrio, and SS: Salado Sur. Forabbreviations see Systematic Paleontology.
Specimen Locality-Bed Length nymphLength
escutcheon Length Height Width L/H H/W
CPBA 19686.4CPBA 19686.6CPBA 19686.7CPBA 19686.8CPBA 19686.9
MU 9-C2MU 9-C2MU 9-C2MU 9-C2MU 9-C2
22.124.327.5
——
50.951.264.562.374.2
85.982.6
120.194.6
135.5
62.363.278.367.481.0
40.742.850.348.558.6
1.381.311.531.401.67
1.531.431.561.391.38
CPBA 19686.10CPBA 19686.10CPBA 19686.10CPBA 19686.10CPBA 19686.10
MU 9-C2MU 9-C2MU 9-C2MU 9-C2MU 9-C2
—14.3
———
63.730.3
——
51.7
101.344.3
103.695.581.6
79.236.562.669.663.0
44.324.440.445.936.9
1.281.211.651.371.30
1.791.501.551.521.71
CPBA 19687.1CPBA 19688.1CPBA 19688.4CPBA 19688.5CPBA 19691.2
MU 6-C1MU 6/9MU 6/9MU 6/9BA 13
—24.122.6
——
62.161.151.451.569.8
110.5111.291.586.6
118.6
74.871.062.567.078.5
42.148.439.741.251.3
1.481.571.461.291.51
1.781.471.571.631.53
CPBA 19691.8CPBA 19691.9CPBA 19691.14CPBA 19691.16CPBA 19691.17
BA 13BA 13BA 13BA 13BA 13
——
24.5—
26.0
73.658.554.0
—51.3
114.6101.880.652.686.8
90.073.663.543.065.6
52.848.337.324.240.7
1.271.381.271.221.32
1.701.521.701.781.61
CPBA 19691.20CPBA 19691.22CPBA 19691.23CPBA 19691.24CPBA 19691.25
BA 13BA 13BA 13BA 13BA 13
—15.630.910.2
—
58.342.476.2
—62.7
90.162.4
109.147.1
100.4
65.946.884.637.179.5
42.330.151.822.547.4
1.371.331.291.271.26
1.561.551.631.651.68
CPBA 19691.27CPBA 19691.28CPBA 19691.30CPBA 19694.4CPBA 19694.5CPBA 19695.1
BA 13BA 13BA 13BA 10BA 10SS 3
26.0—8.5—
11.5—
68.764.318.763.125.372.7
119.5106.630.296.536.8
110.0
80.575.125.167.531.071.0
80.444.617.744.617.740.3
1.481.421.201.431.191.55
1.001.681.421.511.751.76
MeanStandard deviationCoefficient variation
20.587.13
34.66
56.8314.3925.32
90.6026.1928.91
65.0516.1724.85
41.8812.5129.88
1.380.139.47
1.580.16
10.01
exhalant current; marginal carina or posterior ridge is the ridgethat runs from umbo to posteroventral angle of shell and delimitsposterior area; flank is the median part of surface of valve lim-ited by marginal carina; lunule is the cordate depression presentanterior to beaks; nymph is a narrow lunate platform extendingposteriorly from beak along dorsal margin and serving for at-tachment of ligament.
Measurements were made using a digital caliper (accurate to0.2 mm) and recorded in mm. Abbreviations are as follows: L 5length; H 5 height; W 5 width of both valves together; L/H 5elongation; H/W 5 inflation; Le 5 length of escutcheon; Ln 5length of nymph.
Angular measurements are accurate to the nearest degree: Av5 angle between the anterior and ventral margins; Pv 5 anglebetween the posterior and ventral margins.
For details in terminology and measured characters see Figure5, and for the measurements see Tables 1, 2.
Shell size is said to be small, medium or large as comparedwith other specimens of the same species.
Occurence.Stratigraphic range of Steinmanella species fromthe Agrio Formation is based on the ammonoid zonation proposedby Aguirre-Urreta and Rawson (1997, 1998, 2001b).
Order TRIGONIOIDA Dall, 1889Suborden MYOPHORELLINA Cooper, 1991
Superfamily MYOPHORELLACEA Kobashayi, 1954Family MYOPHORELLIDAE Kobashayi, 1954Subfamily STEINMANELLINAE Cooper, 1991
Genus STEINMANELLA Crickmay, 1930Type species.Trigonia holubi KITCHIN, 1908.Diagnosis.Steinmanellines with massive and robust shell; es-
cutcheon and corselet ornamented with commarginal tuberculate
ribs; flank ornamented with coarsely nodate oblique ribs, gener-ally weakly curved, and uninterrupted (slightly modified fromCooper, 1991).
Discussion.Since Crickmay (1930) created the genus Stein-manella, it has had different taxonomic meanings and the rela-tionships between the species and consequently the space-tem-poral distribution of the genus is still under discussion (Camachoand Olivero, 1985). The main problems seem to be the underes-timation of the intra-population variation and the lack of detailedstratigraphic data.
Reyes Bianchi et al. (1981), using only previously describedSouth American forms, have made the only known phylogeneticstudy of this genus. They concluded that in South America theSteinmanella species followed two evolutionary trends. The firstone consists of the plexus S. transitoria characterized by mod-erately convex margins, wide corselet, well-developed carinae andconcave flank ribs. The second, the S. erycina plexus, containsforms with straight margins, low corselet lacking carinae and bun-dled flank ribs.
The characters used for the identification of this genus varyaccording to author (Camacho and Olivero, 1985; Cooper, 1991;Leanza, 1993) making it difficult to assign new fossil material toit. This situation demonstrates the necessity of detailed systematicstudies where the myophorellid genera are compared with eachother, assessing the characters within a solid stratigraphic context.
The subgenera of Steinmanella have also been discussed. Cox(1969c) synonymized S. (Steinmanella) and S. (Transitrigonia),and included Northern Hemisphere forms: S. (Litschkovitrigonia),S. (Setotrigonia) and S. (Yeharella). Camacho and Olivero (1985)separated S. (Steinmanella) from S. (Transitrigonia) revalidatingboth subgenera. In addition, they created S. (Macrotrigonia) and

1076 JOURNAL OF PALEONTOLOGY, V. 77, NO. 6, 2003
TABLE 2—Measurements (in mm) of Steinmanella transitoria (Steinmann, 1881). MU: Agua de la Mula, BA: Bajada del Agrio, SS: Salado Sur, and PI:Pichaihue. For abbreviations see Systematic Paleontology.
Specimen Locality-BedLengthnymph
Lengthescutcheon Length Height Width L/H H/W
CPBA 19710.0CPBA 19704.1CPBA 19704.3CPBA 19705.2CPBA 19706.1
MU 41-C11SS 36SS 36SS 37SS 63
22.023.420.8
—10.5
62.850.1
—63.725.7
89.184.372.5
108.637.8
74.164.360.871.830.7
38.636.342.449.014.9
1.201.311.191.511.23
1.921.771.431.472.06
CPBA 19707.1CPBA 19708.1CPBA 19709.1CPBA 19712.1CPBA 19712.2
SS 65SS 79SS 83MU O. (O.) laticosta SuboneMU O. (O.) laticosta Subone
18.221.921.6
—24.5
50.557.944.963.159.2
78.795.471.790.293.3
56.271.953.672.768.3
42.149.038.944.746.0
1.401.331.341.241.37
1.331.471.381.631.48
CPBA 19713.1CPBA 19714CPBA 19715.1CPBA 19715.5CPBA 19716.11CPBA 19717.1
MU w. vacaensis ZoneBA 38BA 57BA 57BA 77PI H. gentilii Zone
18.220.623.5
——
22.1
47.154.059.362.753.949.5
70.782.191.0
102.081.280.7
56.664.370.177.164.457.2
40.540.645.046.938.934.0
1.251.281.301.321.261.41
1.401.581.561.641.661.68
MeanStandard deviationCoefficient variation
20.613.72
18.03
53.639.92
18.50
83.0816.1119.39
63.3811.2917.81
40.498.10
20.00
1.310.086.48
1.590.20
12.51
→
FIGURE 6—Steinmanella pehuenmapuensis (Leanza, 1998). 1, CPBA 19691.2 in left lateral view; 2 , CPBA 19691.29 in anterior view; 3, CPBA19691.30 in left lateral view; 4, 6, CPBA 19686.5 in dorsal and anterior views; 5, 9, CPBA 19691.27 in dorsal and left lateral views; 7, 8, CPBA19694.5 in dorsal and left lateral views; 10, CPBA 19694.5 in anterior view; 11, CPBA 19686.9 in anterior view. Figures 1–3, 5–7, 9, 10 fromBajada del Agrio; 4, 8, 11 from Agua de la Mula. All figures 31.
excluded the boreal forms mentioned above. Finally Leanza(1993) followed Camacho and Olivero (1985), but added the sub-genus S. (Splenditrigonia).
According to Camacho and Olivero (1985), one of the maindifferences between the subgenera S. (Steinmanella)) and S.(Transitrigonia) is the position of tubercles on the flank ribs inthe proximity of the ventral margin. Nevertheless, the arrange-ment seems to change through ontogeny and therefore is not adiagnostic character.
S. (splenditrigonia) is characterized by flank ribs arranged innarrow or wide bundles (Leanza, 1993).
S. (Macrotrigonia) can be easily identified because it is moreelongated (length/height . 1.65) than S. (Steinmanella), S. (Tran-sitrigonia), and S. (Splenditrigonia) (all with length/height ,1.65) (Camacho and Olivero, 1985; Leanza, 1993).
Therefore, the separation of S. (Steinmanella) and S. (Transitri-gonia) does not seem valid, but S. (Macrotrigonia) and S. (Splen-ditrigonia) can be readily distinguished from those subgenera.
In the present work, subgenera of Steinmanella will not beused, awaiting a detailed revision of the systematics of this genus.
Occurrence.Tithonian-Hauterivian, Barremian?, Aptian?: Ar-gentina, Chile, Peru, Tanzania, South Africa, and India (Lisson,1930; Camacho and Olivero, 1985) (Fig. 2).
STEINMANELLA PEHUENMAPUENSIS (Leanza, 1998)Figure 6.1–11
Trigonia transitoria STEINMANN. DOUVILLE, 1910, p. 20, fig. 5; WEAVER,1931, p. 242–245, pl. 21, figs. 106–108; LAMBERT, 1944, p. 374–377,pl. 6, fig. 2, pl. 7, figs. 1, 2.
Steinmanella vacaensis (WEAVER, 1931). LEANZA AND GARATE ZUBIL-LAGA, 1987, p. 215–216, pl. 10, figs. 1–3.
Steinmanella (Macrotrigonia) pehuenmapuensis sp. nov. LEANZA, 1998,p. 57–67, pl. 1, figs. 1–4.
Description.Equivalved subtriangular shell elongated anteropos-teriorly. Beaks are opisthogyrous. The anterior margin is straight toweakly convex in lateral view. It is flat and wide in anterior view.
It shows a lunule and extensions of flank ribs with asymmetric crosssection (Fig. 6.2, 6.6, 6.10–11). The posterior margin is slightly con-vex. The dorsal margin is straight. The ventral margin is convex.The ventral and anterior margins form of an angle of approximately908. The ventral and posterior margins form an acute angle of 708,which offers an acuminate aspect (Fig. 6.1, 6.9).
The flank is ornamented with conspicuously tuberculate obliqueribs. The interspaces increase in width from dorsal to ventral. Thetubercles have rounded surface and irregular shape. It is delimitedby a marginal carina, with tubercles in the anterior portion andcommarginal ribs in the posterior region.
The corselet has commarginal ribs (Fig. 6.1, 6.5, 6.7, 6.9). Itis delimited by the escutcheon carina, with tubercles in the an-terior portion and commarginal ribs in the posterior region. Amedian groove divides the corselet in lower and upper wedges.
The escutcheon has few irregular tuberculate ribs prolongedfrom the ribs of corselet. The nymph has a lanceolated outlineand lies within the escutcheon.
Internally approximately isomyarian adductor muscle scars andentire pallial line are recognized. In addition, folds in the posteriorregion, that externally coincide with the medium groove and theescutcheon carina, are present.
Material examined.94 articulate bivalved specimens all col-lected by the author in the Lower Member of the Agrio Formationin Neuquen: 47 from Agua de la Mula (CPBA 19686.1–29,19686.31–37, 19687.1–2, 19688.1, 19688.3–7; 19688.9–10 fromthe C. ornatum Subzone; CPBA 19689.1 from the Neocomites sp.Subzone); 10 from Salado Sur (CPBA 19695.1–3, 19696.1–7from the C. ornatum Subzone); and 37 from Bajada del Agrio(CPBA 19698.1, 19698.8, 19699.1–3 from the P. angulatiformisSubzone; CPBA 19691.1–9, 19691.11–14, 19691.16–30,19694.4–5 from C. ornatum Subzone; CPBA 19701.1, 19702.1from the Neocomites sp. Subzone);
Additional material includes one articulate bivalved specimen(CPBA 7571; Lambert, 1944, pl. 7, figs. 1, 2) collected by F.Pastore in the Lower Cretaceous of Loncopue (Neuquen); three

1077LAZO—CRETACEOUS BIVALVES FROM ARGENTINA
1078 JOURNAL OF PALEONTOLOGY, V. 77, NO. 6, 2003
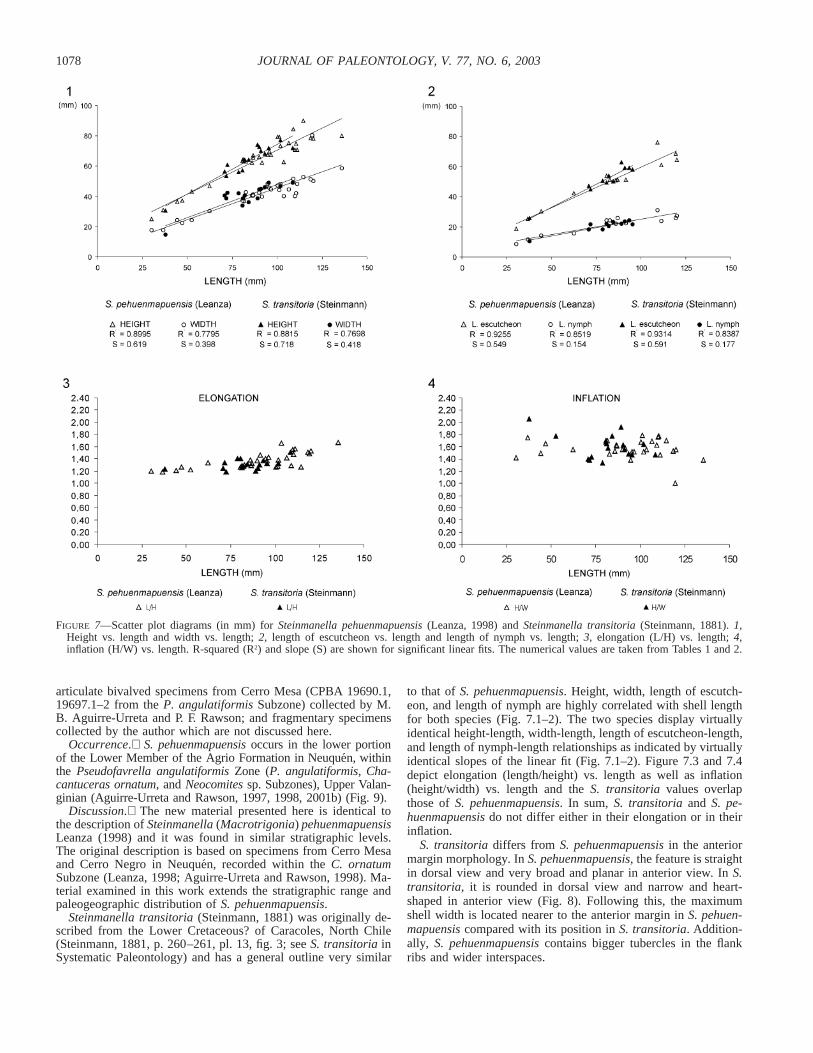
FIGURE 7—Scatter plot diagrams (in mm) for Steinmanella pehuenmapuensis (Leanza, 1998) and Steinmanella transitoria (Steinmann, 1881). 1,Height vs. length and width vs. length; 2, length of escutcheon vs. length and length of nymph vs. length; 3, elongation (L/H) vs. length; 4,inflation (H/W) vs. length. R-squared (R2) and slope (S) are shown for significant linear fits. The numerical values are taken from Tables 1 and 2.
articulate bivalved specimens from Cerro Mesa (CPBA 19690.1,19697.1–2 from the P. angulatiformis Subzone) collected by M.B. Aguirre-Urreta and P. F. Rawson; and fragmentary specimenscollected by the author which are not discussed here.
Occurrence.S. pehuenmapuensis occurs in the lower portionof the Lower Member of the Agrio Formation in Neuquen, withinthe Pseudofavrella angulatiformis Zone (P. angulatiformis, Cha-cantuceras ornatum, and Neocomites sp. Subzones), Upper Valan-ginian (Aguirre-Urreta and Rawson, 1997, 1998, 2001b) (Fig. 9).
Discussion.The new material presented here is identical tothe description of Steinmanella (Macrotrigonia) pehuenmapuensisLeanza (1998) and it was found in similar stratigraphic levels.The original description is based on specimens from Cerro Mesaand Cerro Negro in Neuquen, recorded within the C. ornatumSubzone (Leanza, 1998; Aguirre-Urreta and Rawson, 1998). Ma-terial examined in this work extends the stratigraphic range andpaleogeographic distribution of S. pehuenmapuensis.
Steinmanella transitoria (Steinmann, 1881) was originally de-scribed from the Lower Cretaceous? of Caracoles, North Chile(Steinmann, 1881, p. 260–261, pl. 13, fig. 3; see S. transitoria inSystematic Paleontology) and has a general outline very similar
to that of S. pehuenmapuensis. Height, width, length of escutch-eon, and length of nymph are highly correlated with shell lengthfor both species (Fig. 7.1–2). The two species display virtuallyidentical height-length, width-length, length of escutcheon-length,and length of nymph-length relationships as indicated by virtuallyidentical slopes of the linear fit (Fig. 7.1–2). Figure 7.3 and 7.4depict elongation (length/height) vs. length as well as inflation(height/width) vs. length and the S. transitoria values overlapthose of S. pehuenmapuensis. In sum, S. transitoria and S. pe-huenmapuensis do not differ either in their elongation or in theirinflation.
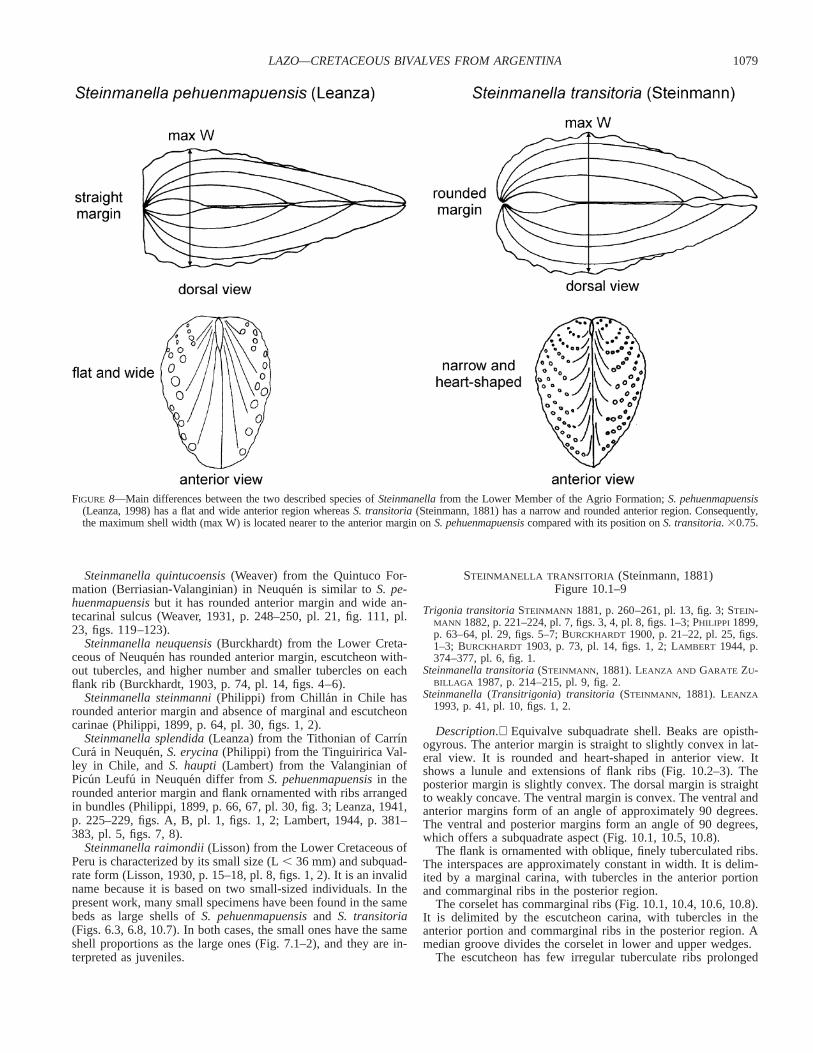
S. transitoria differs from S. pehuenmapuensis in the anteriormargin morphology. In S. pehuenmapuensis, the feature is straightin dorsal view and very broad and planar in anterior view. In S.transitoria, it is rounded in dorsal view and narrow and heart-shaped in anterior view (Fig. 8). Following this, the maximumshell width is located nearer to the anterior margin in S. pehuen-mapuensis compared with its position in S. transitoria. Addition-ally, S. pehuenmapuensis contains bigger tubercles in the flankribs and wider interspaces.
1079LAZO—CRETACEOUS BIVALVES FROM ARGENTINA
FIGURE 8—Main differences between the two described species of Steinmanella from the Lower Member of the Agrio Formation; S. pehuenmapuensis(Leanza, 1998) has a flat and wide anterior region whereas S. transitoria (Steinmann, 1881) has a narrow and rounded anterior region. Consequently,the maximum shell width (max W) is located nearer to the anterior margin on S. pehuenmapuensis compared with its position on S. transitoria. 30.75.
Steinmanella quintucoensis (Weaver) from the Quintuco For-mation (Berriasian-Valanginian) in Neuquen is similar to S. pe-huenmapuensis but it has rounded anterior margin and wide an-tecarinal sulcus (Weaver, 1931, p. 248–250, pl. 21, fig. 111, pl.23, figs. 119–123).
Steinmanella neuquensis (Burckhardt) from the Lower Creta-ceous of Neuquen has rounded anterior margin, escutcheon with-out tubercles, and higher number and smaller tubercles on eachflank rib (Burckhardt, 1903, p. 74, pl. 14, figs. 4–6).
Steinmanella steinmanni (Philippi) from Chillan in Chile hasrounded anterior margin and absence of marginal and escutcheoncarinae (Philippi, 1899, p. 64, pl. 30, figs. 1, 2).
Steinmanella splendida (Leanza) from the Tithonian of CarrınCura in Neuquen, S. erycina (Philippi) from the Tinguiririca Val-ley in Chile, and S. haupti (Lambert) from the Valanginian ofPicun Leufu in Neuquen differ from S. pehuenmapuensis in therounded anterior margin and flank ornamented with ribs arrangedin bundles (Philippi, 1899, p. 66, 67, pl. 30, fig. 3; Leanza, 1941,p. 225–229, figs. A, B, pl. 1, figs. 1, 2; Lambert, 1944, p. 381–383, pl. 5, figs. 7, 8).
Steinmanella raimondii (Lisson) from the Lower Cretaceous ofPeru is characterized by its small size (L , 36 mm) and subquad-rate form (Lisson, 1930, p. 15–18, pl. 8, figs. 1, 2). It is an invalidname because it is based on two small-sized individuals. In thepresent work, many small specimens have been found in the samebeds as large shells of S. pehuenmapuensis and S. transitoria(Figs. 6.3, 6.8, 10.7). In both cases, the small ones have the sameshell proportions as the large ones (Fig. 7.1–2), and they are in-terpreted as juveniles.
STEINMANELLA TRANSITORIA (Steinmann, 1881)Figure 10.1–9
Trigonia transitoria STEINMANN 1881, p. 260–261, pl. 13, fig. 3; STEIN-MANN 1882, p. 221–224, pl. 7, figs. 3, 4, pl. 8, figs. 1–3; PHILIPPI 1899,p. 63–64, pl. 29, figs. 5–7; BURCKHARDT 1900, p. 21–22, pl. 25, figs.1–3; BURCKHARDT 1903, p. 73, pl. 14, figs. 1, 2; LAMBERT 1944, p.374–377, pl. 6, fig. 1.
Steinmanella transitoria (STEINMANN, 1881). LEANZA AND GARATE ZU-BILLAGA 1987, p. 214–215, pl. 9, fig. 2.
Steinmanella (Transitrigonia) transitoria (STEINMANN, 1881). LEANZA
1993, p. 41, pl. 10, figs. 1, 2.
Description.Equivalve subquadrate shell. Beaks are opisth-ogyrous. The anterior margin is straight to slightly convex in lat-eral view. It is rounded and heart-shaped in anterior view. Itshows a lunule and extensions of flank ribs (Fig. 10.2–3). Theposterior margin is slightly convex. The dorsal margin is straightto weakly concave. The ventral margin is convex. The ventral andanterior margins form of an angle of approximately 90 degrees.The ventral and posterior margins form an angle of 90 degrees,which offers a subquadrate aspect (Fig. 10.1, 10.5, 10.8).
The flank is ornamented with oblique, finely tuberculated ribs.The interspaces are approximately constant in width. It is delim-ited by a marginal carina, with tubercles in the anterior portionand commarginal ribs in the posterior region.
The corselet has commarginal ribs (Fig. 10.1, 10.4, 10.6, 10.8).It is delimited by the escutcheon carina, with tubercles in theanterior portion and commarginal ribs in the posterior region. Amedian groove divides the corselet in lower and upper wedges.
The escutcheon has few irregular tuberculate ribs prolonged

1080 JOURNAL OF PALEONTOLOGY, V. 77, NO. 6, 2003
FIGURE 9—Ranges of Steinmanella species of the Agrio Formation. The numerical time-scale and ammonoid zonation are based on Aguirre-Urretaand Rawson (1997, 1998, 2001b); lithology of the Upper Member from Brinkmann (1994); paleoenvironments from Uliana et al. (1977) andGulisano and Gutierrez Pleimling (1988); and unconformities from Zavala (2000).
→FIGURE 10—Steinmanella transitoria (Steinmann, 1881). 1, 4, CPBA 19708.1 in right lateral and dorsal views; 2, 6, CPBA 19714.1 in anterior and
dorsal views; 3, CPBA 19708.3 in anterior view; 5, CPBA 19707.1 in left lateral view; 7, CPBA 19706.1 in rigth lateral view; 8, 9, CPBA 19713.2in left lateral view and internal view of left valve. Figures 1, 3, 4, 5, 7 from Salado Sur; 8, 9 from Agua de la Mula; 2, 6 from Bajada del Agrio.All figures 31.
from the ribs of corselet. The nymph has a lanceolated contourand is within the escutcheon.
Internally approximately isomyarian adductor muscle scars andentire pallial line are recognized. In addition, folds in the posteriorregion, that externally coincide with the medium groove and theescutcheon carina, are present (Fig. 10.9).
Material examined.38 articulate bivalved specimens all col-lected by the author in the Lower Member of the Agrio Formationin Neuquen: one from Pichaihue (CPBA 19717.1 from the H.gentilii Zone); eight from Agua de la Mula (CPBA 19710.1–3,19710.7 from the H. compressum Subzone; CPBA 19712.1–2from the O. (O.) laticosta Subzone; CPBA 19711.2 from the H.
gentilii Zone; CPBA 19713.1 from the W. vacaensis Zone); 14from Salado Sur (CPBA 19704.1–5, 19705.1–2 from the H. com-pressum Subzone; CPBA 19706.1, 19707.1, 19708.1–3 from theH. gentilii Zone; CPBA 19709.1, 19709.3–4 from the W. vacaen-sis Zone); 15 from Bajada del Agrio (CPBA 19714.1 from the H.neuquensis Subzone; CPBA 19715.1, 19715.4–6, 19715.8–10,19715.12, 19715.14, 19715.17 from the H. compressum Subzone;CPBA 19716.5–6, 19716.10–11 from the H. gentilii Zone).
Additional material includes one left valve from Salado Sur(CPBA 19713.2, W. vacaensis Zone) collected by M. B. Aguirre-Urreta and P. F. Rawson, and fragmentary specimens collected bythe author which are not discussed here.

1081LAZO—CRETACEOUS BIVALVES FROM ARGENTINA

1082 JOURNAL OF PALEONTOLOGY, V. 77, NO. 6, 2003
Occurrence.This species was originally described from theLower Cretaceous? of Caracoles in North Chile (Steinmann,1881). It has been found in the Upper Jurassic and Lower Cre-taceous of Central Chile (Steinmann, 1882; Philippi, 1899; ReyesBianchi and Perez D’Angelo, 1979; Aguirre-Urreta and Lo Forte,1996) and Lower cretaceous of Argentina (Burckhardt, 1900,1903; Haupt, 1907; Lambert, 1944; Damborenea et al., 1979;Leanza and Garate Zubillaga, 1987; Leanza, 1993).
In this work it has been recorded in the middle and upper partsof the Lower Member of the Agrio Formation in Neuquen, withinthe Holcoptychites neuquensis (H. neuquensis, H. compressum,and Olcostephanus (O.) laticosta Subzones), Hoplitocriocerasgentilii and Weavericeras vacaensis Zones, Lower Hauterivian(Aguirre-Urreta and Rawson, 1997, 1998, 2001b) (Fig. 9).
Discussion.For differences with S. pehuenmapuensis see pre-vious discussion.
Steinmanella quintucoensis (Weaver) from the Quintuco For-mation (Berriasian-Valanginian) in Neuquen has a wide antecar-inal sulcus (Weaver, 1931, p. 248–250, pl. 21, fig. 111, pl. 23,figs. 119–123).
Steinmanella steinmanni (Philippi) from Chillan in Chile hasnot marginal and escutcheon carinae (PHILIPPI, 1899, p. 64, pl.30, figs. 1, 2).
Steinmanella neuquensis (Burckhardt) from the Lower Creta-ceous of Neuquen has escutcheon without tubercles (Burckhardt,1903, p. 74, pl. 14, figs. 4–6).
Steinmanella splendida (Leanza) from the Tithonian of CarrınCura in Neuquen, Steinmanella erycina (Philippi) from the Tin-guiririca Valley in Chile, and Steinmanella haupti (Lambert) fromthe Valanginian of Picun Leufu in Neuquen differ from S. tran-sitoria in the flank ornamented with ribs arranged in bundles (Phi-lippi, 1899, p. 66, 67, pl. 30, fig. 3; Leanza, 1941, p. 225–229, figs.A, B, pl. 1, figs. 1, 2; Lambert, 1944, p. 381–383, pl. 5, figs. 7, 8).
STEINMANELLA SPECIES OF THE AGRIO FORMATION
In the Lower Member of the Agrio Formation, two stratigraph-ically distinct Steinmanella species have been identified: S. pe-huenmapuensis (Leanza, 1998), from the lower part of the studiedsection (Fig. 9), and S. transitoria (Steinmann, 1881) from themiddle and upper part. S. pehuenmapuensis comes from the Pseu-dofavrella angulatiformis Zone, that comprises three ammonitesubzones: Pseudofavrella angulatiformis, Chacantuceras orna-tum, and Neocomites sp. These ammonites are assigned to theLate Valanginian. Moreover, S. transitoria is found within thethree successive ammonite zones (Fig. 9): Holcoptychites neu-quensis (H. neuquensis, H. compressum, and O. (O.) laticostaSubzones), Hoplitocrioceras gentilii and Weavericeras vacaensis,assigned to the early Hauterivian (Aguirre-Urreta and Rawson,1997). This is one of the first accurate studies of the stratigraphicdistribution of Lower Cretaceous bivalves of Argentina, based onsystematic revision and new collected material.
The identified Steinmanella species succeed each other alongthe analyzed sequence (Fig. 9). No co-occurrence levels were ob-served and no transitional forms were recorded. A future moredetailed sampling of the Neocomites sp. Subzone will probablyprovide new insights into possible morphological changes be-tween S. pehuenmapuensis and S. transitoria.
The presence of at least one Steinmanella species, S. vacaensis(Weaver, 1931) in the Upper Member of the Agrio Formation(Fig. 9) has been documented (Weaver, 1931, p. 251–254, pl. 24,figs. 126–130). It is an elongated form (length/height .1.65) andit seems to be very closely related to Steinmanella (Macrotrigon-ia) katterfeldensis Camacho and Olivero, S. (M.) posadaensis Ca-macho and Olivero, and S. (M.) maxima Camacho and Oliverorecorded in the Austral Basin (Camacho and Olivero, 1985).
Thus, it will be necessary to revise these forms and their strati-graphic range in future studies to comprehend the affinities ofSteinmanella species known in the Neuquen and Austral basins.
STEINMANELLA LIFE HABIT
Stanley (1977) demonstrated that the tuberculate flank ribs ofthe closely related genus Yaadia facilitates excavation in cohesivesediments (with high percentage of mud) where adhesion of thesediment to tubercles occurs. These results are also applicable tothe genus Steinmanella which is recorded in situ in claystones ofthe Agrio Formation.
Like modern Neotrigonia, the genus Steinmanella has an entirepallial line and is presumed, therefore, to be a nonsiphonate bivalve.Nevertheless, the presence of two internal radial folds on bothvalves in coincidence with the position of the escutcheon carinaand median groove would have permitted the separation of theinhalant and exhalant currents as a pseudo-siphon. This separationwould have increased respiratory function and suspension feeding.
The use of epibionts and endobionts for the reconstruction ofthe life habit of fossil trigonioids is supported by the fact thatsome recent species of Neotrigonia have filamentous seaweed andothers epibionts on the posterior region (Tevesz, 1975). Never-theless, the distribution of epi- and endobionts on shells must bestatistically tested on a reasonable number of specimens, as pos-sible random effects can not be discarded (Alexander, 1993). Be-sides, it is necessary to assess post-mortem colonization of theshells or movement of the hosts once the animal died (Saul,1978). In such cases, the host’s distribution on shells does notreflect bivalve life position.
Villamil et al. (1998) used the epibiont orientation four Stein-manella species to interpret life position and depth of burial. Theydeduced a semi-infaunal life habit, with anteroposterior axis per-pendicular to the substrate in an almost vertical position, and with40 to 70 percent of the shell located above the sediment- waterinterface. However, in the four analyzed species only one indi-vidual of each was observed, being impossible to test statisticallythe results. Moreover, as foot protrusion would have taken placeat the intersection of the anterior and ventral margins or at theventral margin, it would have been very difficult for the animalto acquire a vertical position when digging. On the other hand,the large proportion of the animal above the substrate would causethe shell to become unstable and susceptible to be reworked bycurrents. Statistical tests of epibiont’s distribution on the speci-mens collected here will probably be analyzed in detail in futureworks.
Here a semi-infaunal to shallow-infaunal life habit for the spe-cies of Steinmanella is assumed similar to the proposed mode oflife of Yaadia by other authors (Stanley, 1977; Kennedy, 1978;Tashiro and Matsuda, 1988) (Fig. 11). The commisural plane isplaced perpendicular to the sediment-water interface and, becausethey are nonsiphonate bivalves the posterior margin (respiratory)should at least align with the interface and not be oriented belowit. However, it is very difficult to asses the exact angle betweenthe dorsal margin and the interface, and probably it was variabledepending on the depth to which the bivalve burrowed.
CONCLUSIONS
1. In the Lower Member of the Agrio Formation, two strati-graphically distinct species of the genus Steinmanella wereidentified: Steinmanella pehuenmapuensis (Leanza, 1998) ofUpper Valanginian age, and Steinmanella transitoria (Stein-mann, 1881) of Lower Hauterivian age.
2. Based on functional morphology analysis a semi-infaunal toshallow infaunal life habit for the species of the genus Stein-manella is assumed.

1083LAZO—CRETACEOUS BIVALVES FROM ARGENTINA
FIGURE 11—Proposed semi-infaunal to shallow infaunal life habit for thegenus Steinmanella based on classic functional morphology studies, themode of life and soft anatomy of the living genus Neotrigonia and newfield data about the facies bearing Steinmanella. 1, Steinmanella spec-imen burrowing on a muddy bottom; 2, Approximate life positionreached after burrowing with the respiratory region just above the sed-iment-water interface (SWI). The angle formed by the hinge axis andthe SWI may be variable. M: site of pedal emergence at maximumshell gape from Stanley, 1970. 30.75.
3. Steinmanella bivalves were recorded in intermediate to high-energy shoreface and low-energy offshore of a storm-domi-nated, shallow-marine ramp located on the Southeastern Pa-cific.
4. Steinmanella bivalves periodically colonized low-energymuddy bottoms in the offshore during increased oxygen lev-els and reduced net sedimentation or brief omission periods.
ACKNOWLEDGMENTS
I am most grateful to M. B. Aguirre-Urreta (Universidad deBuenos Aires) for her constant support and helpful revision ofthe manuscript, and to P. F. Rawson (University College London)for his advice and supervision during field trips. E. Olivero as-sisted with valuable bibliography. I appreciate helpful commentsand suggestions from P. Harries, A. Dhondt, and anonymous re-viewer. Also, I wish to thank G. A. Concheyro, M. Cichowolski,and R. Sanci for their valuable help during field work. This re-search was supported by PIP CONICET 360/98 and UBACYTX155 grants to M. B. Aguirre-Urreta.
REFERENCES
ABERHAN, M. 1993. Benthic macroinvertebrate associations on a carbon-ate-clastic ramp in segments of the Early Jurassic back-arc basin ofnorthern Chile (26–298S). Revista Geologica de Chile, 20:105–136.
AGUIRRE-URRETA, M. B. 1998. The ammonites Karakaschiceras andNeohoploceras (Valanginian Neocomitidae) from the Neuquen Basin,West-Central Argentina. Journal of Paleontology, 72:39–59.
AGUIRRE-URRETA, M. B., AND G. L. LO FORTE. 1996. Los depositosTithoneocomianos, p. 179–229. In V. A. Ramos, M. B. Aguirre-Urreta,P. P. Alvarez, M. I. Cegarra, E. O. Cristallini, S. M. Kay, G. L. Forte,F. X. Pereyra, and D. J. Perez (eds.), Geologıa de la region del Acon-cagua. Provincias de San Juan y Mendoza, Republica Argentina. Di-reccion Nacional del Servicio Geologico Subsecretarıa de Minerıa dela Nacion Anales 24, Buenos Aires.
AGUIRRE-URRETA, M. B., AND V. A. RAMOS. 1981. Estratigrafıa y pa-leontologıa de la Alta cuenca del rıo Noble, Cordillera Patagonica,provincia de Santa Cruz. Actas del VIII Geologico Argentino, SanLuis, 3:101–138.
AGUIRRE-URRETA, M. B., AND P. F. RAWSON. 1997. The ammonite se-quence in the Agrio Formation (Lower Cretaceous), Neuquen Basin,Argentina. Geological Magazine, 134(4):449–458.
AGUIRRE-URRETA, M. B., AND P. F. RAWSON. 1998. The Early Cretaceous(Valanginian) ammonite Chacantuceras gen. nov.—a link between theNeuquen and Austral basins. Revista de la Asociacion Geologica Ar-gentina, 53:354–364.
AGUIRRE-URRETA, M. B., AND P. F. RAWSON. 1999. Lower Cretaceousammonites from the Neuquen Basin, Argentina: Viluceras, a new Va-langinian subgenus of Olcostephanus. Cretaceous Research, 20:343–357.
AGUIRRE-URRETA, M. B., AND P. F. RAWSON. 2001a. Lower Cretaceousammonites from the Neuquen Basin, Argentina: the Hauterivian neo-comitid genus Hoplitocrioceras (Giovine, 1950). Cretaceous Research,22:201–218.
AGUIRRE-URRETA, M. B., AND P. F. RAWSON. 2001b. Lower Cretaceousammonites from the Neuquen Basin, Argentina: a Hauterivian Olcos-tephanus fauna. Cretaceous Research, 22:763–778.
ALEXANDER, R. R. 1993. Correlation of shape and habit with sedimentgrain size for selected species of the bivalve Anadara. Lethaia, 26:153–162.
BEHRENDSEN, O. 1892. Zur Geologie des Ostabhanges der argentinischenCordillere. II. Theil. Zeitschrift der Deutschen geologischen Gesell-schaft, 44:1–42, 4 pls.
BRACACCINI, O. I. 1970. Rasgos tectonicos de las acumulaciones meso-zoicas en las provincias de Mendoza y Neuquen, Republica Argentina.Revista de la Asociacion Geologica Argentina, 25:275–284.
BRINKMANN, H.-D. 1994. Facies and sequences of the Agrio Formation(Lower Cretaceous) in the central and southern Neuquen Basin, Ar-gentina. Zentralblatt fur Geologie und Palaeontologie, Teil, I (1/2):309–317.
BURCKHARDT, C. 1900. Coupe geologique de la Cordillere entre LasLajas et Curacautin. Anales del Museo de La Plata, Seccion Geologicay Mineralogica, 3:1–100, 26 pls.
BURCKHARDT, C. 1903. Beitrage zur Kenntniss der Jura- und Kreidefor-mation der Cordillere. Palaeontographica, 50(1–3):1–144.
CAMACHO, H. H., AND E. B. OLIVERO. 1985. El genero SteinmanellaCrickmay, 1930 (Bivalvia, Trigoniidae) en el Cretacico Inferior delsudoeste gondwanico. Anales de la Academia Nacional de CienciasExactas, Fısicas y Naturales, Buenos Aires, 37:41–62.
CANFIELD, D. E., AND R. RAISWELL. 1991. Carbonate precipitation anddissolution. Its relevance to fossil preservation, p. 412–453. In P. A.Allison and D. E. G. Briggs (eds.),Taphonomy: Releasing the DataLocked in the Fossil Record, Volume 9 of Topics in Geobiology. Ple-num Press, New York.
COOPER, M. R. 1979. Cretaceous Trigoniidae (Mollusca, Bivalvia) fromthe Brenton Formation, Knysna. Annals of the South African Museum,78:49–67.
COOPER, M. R. 1991. Lower Cretaceous Trigonioida (Mollusca, Bivalvia)from the Algoa Basin, with a revised classification of the order. Annalsof the South African Museum, 100(1):1–52.
COX, L. R. 1969a. General features of Bivalvia, p. N3–N129. In R. C.Moore and C. Teichert (eds.), Treatise on Invertebrate Paleontology,Pt. N, Mollusca 6, Bivalvia 1. Geological Society of America and Uni-versity of Kansas Press, Lawrence.
COX, L. R. 1969b. Superfamily Trigoniacea Lamarck, 1819, p. N471. InR. C. Moore and C. Teichert (eds.), Treatise on Invertebrate Paleon-tology, Pt. N, Mollusca 6, Bivalvia 1. Geological Society of Americaand University of Kansas Press, Lawrence.
COX, L. R. 1969c. Family Trigoniidae Lamarck, 1819, p. N476–N488.In R. C. Moore and C. Teichert (eds.), Treatise on Invertebrate Pale-ontology, Pt. N, Mollusca 6, Bivalvia 1. Geological Society of Americaand University of Kansas Press, Lawrence.
CRICKMAY, C. H. 1930. Fossils from Harrison Lake area, British Colum-bia. National Museum of Canada Bulletin, 63:33–66.
DALL, W. H. 1889. On the hinge of pelecypods and its development, withan attempt toward a better subdivision of the group. American Journalof Science, 38(3):445–462.
DAMBORENEA, S. E., M. MANCENIDO, AND A. C. RICCARDI. 1979. Es-tudio paleontologico de la Formacion Chachao. Informe Interno YPF,152 p., 19 pls (unpublished).
DARRAGH, T. A. 1986. The Cainozoic Trigoniidae of Australia. Alcher-inga, 10:1–34.

1084 JOURNAL OF PALEONTOLOGY, V. 77, NO. 6, 2003
DIGREGORIO, R. E., C. A. GULISANO, A. R. GUTIERREZ PLEIMLING, AND
S. A. MINNITI. 1984. Esquema de la evolucion geodinamica de laCuenca Neuquina y sus implicancias paleogeograficas. Actas del IXCongreso Geologico Argentino, San Carlos de Bariloche, 2:147–162.
DOUVILLE, R. 1910. Cephalopodes argentins. Memoires de la SocieteGeologique de France, 43, 24 p.
FLEMING, C. A. 1964. History of the Bivalve Family Trigoniidae in theSouth-West Pacific. The Geological Background to an Australian ‘Liv-ing Fossil’. Australian Journal of Science, 26(7):196–204.
FURSICH, F. T., AND M. ABERHAN. 1990. Siginificance of time-averagingfor palaeocommunity analysis. Lethaia, 23:143–152.
GILBERT SMITH, A., J. C. BRIDEN, AND G. E. DREWRY. 1973. Phanero-zoic world maps. Special Papers in Palaeontology, 12:1–42.
GOLDFUSS, G. A. 1826–1844. Petrefacta Germaniae tam ea, quae in mu-seo universitatis regiae Borussicae Friedericiae Wilhelminae Rhenanaeservantur quam alia quaecunque in museis Hoeninghusiano Muenster-iano aliisque extant, iconibus et descriptionibus illustrata. Abbildungenund Beschreibungen der Petrefacten Deytschlands un der angranzendenLander unter Mitwirkung des Herrn Grafen Georg zu Munster. Dus-seldorf, Arnz & Co, 2:1–312.
GULISANO, C. A., AND A. R. GUTIERREZ PLEIMLING. 1988. Depositoseolicos del Miembro Avile (Formacion Agrio, Cretacico Inferior) en elnorte del Neuquen, Argentina. Actas de la Segunda Reunion Argentinade Sedimentologıa, Buenos Aires, 1:120–124.
HAUPT, O. 1907. Beitrage zur Fauna des oberen Malm und der unterenKreide in der argentinischen Cordillere. Neues Jahrbuch fur Mineral-ogie, Geologie und Palaontologie, Beilage-Band, 23:187–236, 7–10pls.
KELLY, S. R. A. 1995a. New Trigonioid bivalves from the Early Jurassicto Earliest Cretaceous of the Antarctic Peninsula region: systematicsand Austral Paleobiogeography. Journal of Paleontology, 69:66–84.
KELLY, S. R. A. 1995b. New Trigonioid bivalves from the Albian (EarlyCretaceous) of Alexander Island, Antarctic Peninsula: systematics, Pa-leoecology, and Austral Cretaceous Paleobiogeography. Journal of Pa-leontology, 69:264–279.
KENNEDY, W. J. 1978. Cretaceous, p. 280–322. In W. S. McKerrow (ed.),The Ecology of Fossils, an Illustrated Guide. MIT Press, Cambridge,Massachusetts.
KITCHIN, F. L. 1903. The Jurassic fauna of Cutch. The Lamellibranchiata.Genus Trigonia. Memoirs of the Geological Survey of India, Palaeon-tologia Indica, 9, 3(2):1–122.
KITCHIN, F. L. 1908. The invertebrate fauna and palaeontological relationsof the Uitenhage Series. Annals of the South African Museum, 7:21–250.
KOBASHAYI, T. 1954. Studies on the Jurassic trigonians in Japan, Pt. I,Preliminary notes. Japanese Journal of Geology and Geography, 15(1/2):61–80.
LAMBERT, L. R. 1944. Algunas Trigonias del Neuquen. Revista del Mu-seo de La Plata (Nueva Serie), Paleontologıa, 2(14):357–397, 13 pls.
LAZO, D. G. 2001. Distribucion ambiental de trazas fosiles en el MiembroInferior de la Formacion Agrio, Cretacico de Cuenca Neuquina. Actasde la IV Reunion Argentina de Icnologıa y II Reunion de Icnologıadel Mercosur, Tucuman, p. 52.
LEANZA, A. F. 1941. Dos nuevas Trigonias del Titoniense de Carrin-Cura,en el territorio del Neuquen. Notas del Museo de La Plata, Paleonto-logıa, 6(31):225–233, 1 pl.
LEANZA, A. F. 1945. Amonites del Jurasico Superior y del CretacicoInferior de la Sierra Azul, en la parte meridional de la provincia deMendoza. Anales del Museo de La Plata (Nueva Serie), 1:1–99.
LEANZA, A. F., AND H. A. CASTELLARO. 1955. Algunos fosiles cretacicosde Chile. Revista de la Asociacion Geologica Argentina, 10:179–211,4 pls.
LEANZA, H. A. 1993. Jurassic and Cretaceous Trigoniid bivalves fromWest-Central Argentina. Bulletins of American Paleontology, 105(343):1–95.
LEANZA, H. A. 1998. Una nueva especie de Steinmanella Crickmay (Biv-alvia, Trigonioida) del Cretacico Inferior de la Cuenca Neuquina, Ar-gentina. Revista Geologica de Chile, 25:57–67.
LEANZA, H. A., AND J. I. GARATE ZUBILLAGA. 1987. Fauna de Trigonias(Bivalvia) del Jurasico y Cretacico Inferior de la provincia del Neu-quen, Argentina, conservadas en el Museo Juan Olsacher de Zapala, p.
201–255. In W. Volkheimer (ed.), Bioestratigrafıa de los Sistemas Re-gionales del Jurasico y Cretacico de America del Sur. Comite Suda-mericano del Jurasico y Cretacico, Mendoza, Argentina.
LEANZA, H. A., AND J. WIEDMANN. 1980. Ammoniten des Valangin undHauterive (Unterkreide) von Neuquen und Mendoza, Argentinien.Eclogae Geologicae Helvetiae, 73:941–981.
LEANZA, H. A., AND J. WIEDMANN. 1992. Nuevos Holcodiscidae (Ceph-alopoda-Ammonoidea) del Barremiano de la Cuenca Neuquina, Argen-tina, y su significado estratigrafico. Neues Jahrbuch fur Geologie undPalaontologie, Monatshefte 1992, Heft 1:24–38.
LEGARRETA, L., AND C. A. GULISANO. 1989. Analisis estratigrafico se-cuencial de la Cuenca Neuquina (Triasico Superior-Terciario Inferior),p. 221–243. In G. A. Chebli and L. A. Spalletti (eds.), Cuencas Sedi-mentarias Argentinas, Correlacion Geologica 6. Universidad Nacionalde Tucuman, Tucuman.
LEVY DE CAMINOS, R. 1969. Revision de las Trigonias de la Argentina.Parte V: El grupo de las Pseudoquadratae. Ameghiniana, 6:65–68.
LISSON, C. I. 1930. Contribucion al conocimiento de algunas trigoniasneocomicas del Peru. Boletın de Minas. Industrias y Construcciones,Serie 2, 20:3–26, 10 pls.
LO FORTE, G. L. 1988. La fauna de trigonias (Mollusca; Bivalvia) delTithoniano-Neocomiano de la Quebrada Blanca, Alta Cordillera deMendoza. Actas del V Congreso Geologico Chileno, Santiago de Chile,2:C277–C293.
NEWELL, N. D., AND D. W. BOYD. 1965. Classification of the Bivalvia.American Museum Novitates, 2206, 25 p.
NEWELL, N. D., AND D. W. BOYD. 1975. Parallel evolution in early Tri-goniacean bivalves. Bulletin of the American Museum of Natural His-tory, 154(2):53–162.
PHILIPPI, R. A. 1899. Los fosiles Secundarios de Chile. Santiago de Chile,104 p., 42 pls.
POJETA, J., JR. 1978. The origin and early taxonomic diversification ofpelecypods. Philosophical Transactions of the Royal Society of LondonB, 284:225–246.
REYES BIANCHI, R. 1970. La fauna de trigonias de Aisen. Instituto deInvestigaciones Geologicas de Chile Boletın, 26:5–39.
REYES BIANCHI, R., AND E. PEREZ D’ANGELO 1979. Estado actual delconocimiento de la Familia Trigoniidae (Mollusca; Bivalvia) en Chile.Revista Geologica de chile, 8:13–64.
REYES BIANCHI, R., E. PEREZ D’ANGELO, AND I. SEREY E.1981. Estudiosistematico y filogenetico de las especies sudamericanas del generoSteinmanella (Trigoniidae; Bivalvia). Revista Geologica de Chile, 12:25–47.
RYER, T. A. 1991. Lower Cretaceous Avile Sandstone, Neuquen Basin,Argentina-Exploration model for a Lowstand Clastic Wedge in a Back-Arc Basin. American Association of Petroleum Geologists, Bulletin,75:665.
SAUL, L. R. 1978. The North Pacific Cretaceous trigoniid genus Yaadia.University of California Publications in Geological Sciences, Volume119, 65 p., 12 pls.
SPALLETTI, L. A., D. G. POIRE, D. PIRRIE, S. D. MATHEOS, AND P.DOYLE. 2001. Respuesta sedimentologica a cambios en el nivel de baseen una secuencia mixta clastica-carbonatica del Cretacico de la CuencaNeuquina, Argentina. Revista Sociedad Geologica Espana, 14(1–2):57–74.
STANLEY, S. M. 1970. Relations of shell form to life habits of the Biv-alvia (Mollusca). Geological Society of America Memoir, 125.
STANLEY, S. M. 1977. Coadaptation in the Trigoniidae, a remarkablefamily of burrowing bivalves. Palaeontology, 20:866–899.
STEINMANN, G. 1881. Zur Kenntniss der Jura- und Kreideformation vonCaracoles (Bolivia). Neues Jahrbuch fur Mineralogie, Geologie undPalaeontologie, Beilage-Band I, 239–301, pls. 9–14.
STEINMANN, G. 1882. Die Gruppe der Trigoniae pseudo-quadratae. NeuesJahrbuch fur Mineralogie, Geologie und Palaeontologie 1882, Band I,Abhandlungen, 219–228, pls. 7–9.
TASHIRO, M., AND T. MATSUDA. 1988. Mode of life in Cretaceous tri-gonians. Fossils, 45:9–21. (In Japanese with English abstract)
TAYLOR, J. D., W. J. KENNEDY, AND A. HALL. 1969. The shell structureand mineralogy of the Bivalvia. Introduction. Nuculacea-Trigoniacea.Bulletin of the British Museum (Natural History), Zoology Supple-ment, 3:1–125.
TEVESZ, M. J. S. 1975. Structure and habits of the ‘living fossil’ pelec-ypod Neotrigonia. Lethaia, 8:321–327.

1085LAZO—CRETACEOUS BIVALVES FROM ARGENTINA
ULIANA, M. A., D. A. DELLAPE, AND G. A. PANDO. 1977. Analisis es-tratigrafico y evaluacion del potencial petrolıfero de las FormacionesMulichinco, Chachao y Agrio. Cretacico Inferior de las provincias deNeuquen y Mendoza. Petrotecnia, Enero-Febrero 1977, 31–46; Marzo1977, 25–33.
VERGANI, G. D., A. J. TANKARD, H. J. BELOTTI, AND H. J. WELSINK.1995. Tectonic Evolution and Paleogeography of the Neuquen Basin,Argentina. In A. J. Tankard, R. Suarez S., and H. J. Welsink (eds.),Petroleum Basins of South America. American Association of Petro-leum Geologists Memoir, 62:383–402.
VILLAMIL, T., E. G. KAUFFMAN, AND H. A. LEANZA. 1998. Epibionthabitation patterns and their implications for life habits and orientationamong trigoniid bivalves. Lethaia, 31:43–56.
WALKER, K. R., AND W. W. DIEHL. 1986. The effect of synsedimentarysubstrate modification on the composition of paleocommunities: paleo-ecologic sucession revisited. Palaios, 1:65–74.
WEAVER, C. 1931. Paleontology of the Jurassic and Cretaceous of WestCentral Argentina. Memoirs of the University of Washington, 1:1–595.
WIGNALL, P. B. 1993. Distinguishing between oxygen and substrate con-trol in fossil benthic assemblages. Journal of the Geological Society ofLondon, 150:193–196.
ZAVALA, C. 2000. New advances in the sequence stratigraphy and sedi-mentology of the Mulichinco Formation, Neuquen Basin. Boletın deInformaciones Petroleras, Tercera Epoca, 63:40–54.
ACCEPTED 20 FEBRUARY 2003




















