
The Exonucleolytic and Endonucleolytic Cleavage Activities of Human Exonuclease 1 Are Stimulated by...
11
The Exonucleolytic and Endonucleolytic Cleavage Activities of Human Exonuclease 1 Are Stimulated by an Interaction with the Carboxyl-terminal Region of the Werner Syndrome Protein* Received for publication, December 16, 2002, and in revised form, March 26, 2003 Published, JBC Papers in Press, April 18, 2003, DOI 10.1074/jbc.M212798200 Sudha Sharma‡§, Joshua A. Sommers‡, Henry C. Driscoll‡, Laura Uzdilla¶, Teresa M. Wilson¶, and Robert M. Brosh, Jr.‡ From the ‡Laboratory of Molecular Gerontology, NIA, National Institutes of Health, Baltimore, Maryland 21224, the ¶Radiation Oncology Research Laboratory, Department of Radiation Oncology, University of Maryland, Baltimore, Maryland 21201, and the §Department of Biochemistry, School of Life Sciences, University of Hyderabad, Hyderabad 500 046 India Exonuclease 1 (EXO-1), a member of the RAD2 family of nucleases, has recently been proposed to function in the genetic pathways of DNA recombination, repair, and replication which are important for genome integrity. Although the role of EXO-1 is not well understood, its 5 to 3-exonuclease and flap endonuclease activities may cleave intermediates that arise during DNA metabolism. In this study, we provide evidence that the Werner syn- drome protein (WRN) physically interacts with human EXO-1 and dramatically stimulates both the exonucleo- lytic and endonucleolytic incision functions of EXO-1. The functional interaction between WRN and EXO-1 is mediated by a protein domain of WRN which interacts with flap endonuclease 1 (FEN-1). Thus, the genomic instability observed in WRN/ cells may be at least partially attributed to the lack of interactions between the WRN protein and human nucleases including EXO-1. Werner syndrome (WS) 1 is a hereditary premature aging disorder characterized by genome instability (1). WS cells dis- play elevated chromosomal aberrations (2– 4), replication de- fects (3, 5– 8), abnormal recombination (9, 10), altered telomere dynamics (11), and hypersensitivity to DNA-damaging agents (12–16). The gene defective in WS, designated WRN (17), en- codes a protein with DNA helicase (18, 19) and exonuclease (20 –22) activities which presumably functions in DNA metab- olism to preserve genome integrity. To understand the basis of WS, a number of groups have investigated WRN protein inter- actions (for review, see Ref. 23). The collective work indicates that WRN interacts functionally with proteins implicated in replication and DNA repair including replication protein A (RPA), p53, Ku, and polymerase . These studies have enabled researchers to speculate about pathways of DNA metabolism in which WRN might participate; however, the precise functions of the WRN gene product in vivo are not well understood. Recently we reported a novel interaction of WRN protein with the human 5-flap endonuclease/5 to 3-exonuclease (FEN-1) (24), a DNA structure-specific nuclease implicated in DNA replication, recombination, and repair (25). WRN protein stimulates FEN-1 cleavage activity by a physical interaction with a COOH-terminal domain of the WRN protein (24). An- other member of the RAD2 family to which FEN-1 belongs is human exonuclease 1 (EXO-1) (26). Like FEN-1, EXO-1 is a structure-specific endonuclease as well as an exonuclease (27, 28). EXO-1 has been implicated in a number of DNA metabolic pathways including mismatch correction, mitotic and meiotic recombination, and double strand break repair (29 –34). A role for EXO-1 in Okazaki fragment processing during DNA repli- cation has been suggested based on its structure-specific endo- nuclease and RNase H activities that are similar to those of FEN-1 (30). Functional overlap between EXO-1 and FEN-1 (Rad27 in Saccharomyces cerevisiae) has been proposed from observations in yeast that exoI: rad27 double mutants are inviable (35) and that overexpression of yeast EXO-1 or human EXO-1 complements cellular phenotypes of rad27 mutants (30, 35, 36). The evident sequence homology and similar biochemical ac- tivities of EXO-1 and FEN-1, as well as the potential functional redundancy of the two nucleases, suggested to us that WRN might also interact functionally with EXO-1. We have found this to be the case. Evidence is presented that WRN interacts physically with human EXO-1 and stimulates the endonucleo- lytic and exonucleolytic cleavage activities of EXO-1. The func- tional interaction is independent of WRN catalytic function and mediated by a COOH-terminal region of WRN which also in- teracts with FEN-1. Thus, WRN modulates the cleavage activ- ities of both human EXO-1 and FEN-1 by a direct protein interaction, suggesting that either structure-specific nuclease may act together with WRN during DNA replication, recombi- nation, or repair. The physical and functional interaction of WRN with these human nucleases is likely to be important for the cellular role of WRN in the maintenance of genome integrity. MATERIALS AND METHODS Proteins—Hexahistidine-tagged recombinant human EXO-1 protein was overexpressed using a baculovirus/insect system and purified as described elsewhere. 2 The purified EXO-1 protein was judged to be 97% pure from analysis on Coomassie-stained SDS-polyacrylamide gels (see Fig. 2A). Baculovirus constructs for full-length recombinant * The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. To whom correspondence should be addressed: Laboratory of Molec- ular Gerontology, NIA, National Institutes of Health, 5600 Nathan Shock Dr., Baltimore, MD 21224. Tel.: 410-558-8578; Fax: 410-558- 8157; E-mail: [email protected]. 1 The abbreviations used are: WS, Werner syndrome; dsDNA, double strand DNA; EXO-1, exonuclease 1; FEN-1, 5-flap endonuclease/5 to 3-exonuclease; nt, nucleotide; PCNA, proliferating cell nuclear antigen; PVDF, polyvinylidene difluoride; RPA, replication protein A; ssDNA, single strand DNA. 2 L. A. Uzdilla, J. P. Carney, P. Hungspreugs, D. M. Wilson, and T. M. Wilson, manuscript in preparation. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 26, Issue of June 27, pp. 23487–23496, 2003 Printed in U.S.A. This paper is available on line at http://www.jbc.org 23487 by guest on June 28, 2016 http://www.jbc.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The Exonucleolytic and Endonucleolytic Cleavage Activities of Human Exonuclease 1 Are Stimulated by...

The Exonucleolytic and Endonucleolytic Cleavage Activities ofHuman Exonuclease 1 Are Stimulated by an Interaction with theCarboxyl-terminal Region of the Werner Syndrome Protein*
Received for publication, December 16, 2002, and in revised form, March 26, 2003Published, JBC Papers in Press, April 18, 2003, DOI 10.1074/jbc.M212798200
Sudha Sharma‡§, Joshua A. Sommers‡, Henry C. Driscoll‡, Laura Uzdilla¶, Teresa M. Wilson¶,and Robert M. Brosh, Jr.‡�
From the ‡Laboratory of Molecular Gerontology, NIA, National Institutes of Health, Baltimore, Maryland 21224, the¶Radiation Oncology Research Laboratory, Department of Radiation Oncology, University of Maryland, Baltimore,Maryland 21201, and the §Department of Biochemistry, School of Life Sciences, University of Hyderabad,Hyderabad 500 046 India
Exonuclease 1 (EXO-1), a member of the RAD2 familyof nucleases, has recently been proposed to function inthe genetic pathways of DNA recombination, repair, andreplication which are important for genome integrity.Although the role of EXO-1 is not well understood, its 5�to 3�-exonuclease and flap endonuclease activities maycleave intermediates that arise during DNA metabolism.In this study, we provide evidence that the Werner syn-drome protein (WRN) physically interacts with humanEXO-1 and dramatically stimulates both the exonucleo-lytic and endonucleolytic incision functions of EXO-1.The functional interaction between WRN and EXO-1 ismediated by a protein domain of WRN which interactswith flap endonuclease 1 (FEN-1). Thus, the genomicinstability observed in WRN�/� cells may be at leastpartially attributed to the lack of interactions betweenthe WRN protein and human nucleases including EXO-1.
Werner syndrome (WS)1 is a hereditary premature agingdisorder characterized by genome instability (1). WS cells dis-play elevated chromosomal aberrations (2–4), replication de-fects (3, 5–8), abnormal recombination (9, 10), altered telomeredynamics (11), and hypersensitivity to DNA-damaging agents(12–16). The gene defective in WS, designated WRN (17), en-codes a protein with DNA helicase (18, 19) and exonuclease(20–22) activities which presumably functions in DNA metab-olism to preserve genome integrity. To understand the basis ofWS, a number of groups have investigated WRN protein inter-actions (for review, see Ref. 23). The collective work indicatesthat WRN interacts functionally with proteins implicated inreplication and DNA repair including replication protein A(RPA), p53, Ku, and polymerase �. These studies have enabledresearchers to speculate about pathways of DNA metabolism inwhich WRN might participate; however, the precise functionsof the WRN gene product in vivo are not well understood.
Recently we reported a novel interaction of WRN proteinwith the human 5�-flap endonuclease/5� to 3�-exonuclease(FEN-1) (24), a DNA structure-specific nuclease implicated inDNA replication, recombination, and repair (25). WRN proteinstimulates FEN-1 cleavage activity by a physical interactionwith a COOH-terminal domain of the WRN protein (24). An-other member of the RAD2 family to which FEN-1 belongs ishuman exonuclease 1 (EXO-1) (26). Like FEN-1, EXO-1 is astructure-specific endonuclease as well as an exonuclease (27,28). EXO-1 has been implicated in a number of DNA metabolicpathways including mismatch correction, mitotic and meioticrecombination, and double strand break repair (29–34). A rolefor EXO-1 in Okazaki fragment processing during DNA repli-cation has been suggested based on its structure-specific endo-nuclease and RNase H activities that are similar to those ofFEN-1 (30). Functional overlap between EXO-1 and FEN-1(Rad27 in Saccharomyces cerevisiae) has been proposed fromobservations in yeast that exoI: rad27 double mutants areinviable (35) and that overexpression of yeast EXO-1 or humanEXO-1 complements cellular phenotypes of rad27 mutants(30, 35, 36).
The evident sequence homology and similar biochemical ac-tivities of EXO-1 and FEN-1, as well as the potential functionalredundancy of the two nucleases, suggested to us that WRNmight also interact functionally with EXO-1. We have foundthis to be the case. Evidence is presented that WRN interactsphysically with human EXO-1 and stimulates the endonucleo-lytic and exonucleolytic cleavage activities of EXO-1. The func-tional interaction is independent of WRN catalytic function andmediated by a COOH-terminal region of WRN which also in-teracts with FEN-1. Thus, WRN modulates the cleavage activ-ities of both human EXO-1 and FEN-1 by a direct proteininteraction, suggesting that either structure-specific nucleasemay act together with WRN during DNA replication, recombi-nation, or repair. The physical and functional interaction ofWRN with these human nucleases is likely to be important forthe cellular role of WRN in the maintenance of genomeintegrity.
MATERIALS AND METHODS
Proteins—Hexahistidine-tagged recombinant human EXO-1 proteinwas overexpressed using a baculovirus/insect system and purified asdescribed elsewhere.2 The purified EXO-1 protein was judged to be�97% pure from analysis on Coomassie-stained SDS-polyacrylamidegels (see Fig. 2A). Baculovirus constructs for full-length recombinant
* The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
� To whom correspondence should be addressed: Laboratory of Molec-ular Gerontology, NIA, National Institutes of Health, 5600 NathanShock Dr., Baltimore, MD 21224. Tel.: 410-558-8578; Fax: 410-558-8157; E-mail: [email protected].
1 The abbreviations used are: WS, Werner syndrome; dsDNA, doublestrand DNA; EXO-1, exonuclease 1; FEN-1, 5�-flap endonuclease/5� to3�-exonuclease; nt, nucleotide; PCNA, proliferating cell nuclear antigen;PVDF, polyvinylidene difluoride; RPA, replication protein A; ssDNA,single strand DNA.
2 L. A. Uzdilla, J. P. Carney, P. Hungspreugs, D. M. Wilson, and T. M.Wilson, manuscript in preparation.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 26, Issue of June 27, pp. 23487–23496, 2003Printed in U.S.A.
This paper is available on line at http://www.jbc.org 23487
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from

hexahistidine-tagged WRN proteins (wild-type, WRN-K577M, WRN-E84A) or a truncated version of WRN containing only the amino-terminal 368 amino acids of the protein (designated N-WRN1–368) werekindly provided by Drs. M. Gray (University of Washington, Seattle)and J. Campisi (Lawrence Berkeley National Laboratory, Berkeley,CA). Amplified WRN-encoding baculovirus was used to infect Sf9 cellsfor overexpression of WRN protein as described elsewhere (37). A re-combinant hexahistidine-tagged carboxyl-terminal fragment of WRN(residues 940–1432, designated C-WRN940–1432) was overexpressed inEscherichia coli and purified as described previously (38). Recombinanthuman FEN-1 was purified as described previously (24). Human PCNAand RPA were graciously provided by Dr. M. Kenny (Albert EinsteinCollege of Medicine, Bronx, NY).
WRN-EXO-1 Coimmunoprecipitation Experiments—HeLa cells weregrown in Dulbecco’s modified Eagle’s medium supplemented with 10%fetal bovine serum at 37 °C in 5% CO2. Nuclear extract was prepared asdescribed previously (39). For coimmunoprecipitation, HeLa nuclearextract (1.5 mg of protein) was incubated with goat anti-WRN poly-clonal antibody (1:40; Santa Cruz Biotechnology) in buffer D (50 mM
HEPES pH 7.5, 100 mM KCl, 10% glycerol) for 4 h at 4 °C and tumbledwith 20 �l of protein G-agarose (Roche Applied Science) at 4 °C over-night. Beads were washed three times with buffer D supplemented with0.1% Tween 20. Proteins were eluted by boiling treatment in SDSsample buffer, resolved on 8% polyacrylamide Tris-glycine SDS gels,and transferred to PVDF membranes (Amersham Biosciences). Themembranes were blocked with 5% nonfat dry milk in phosphate-buff-ered saline containing 0.1% Tween 20 and probed using rabbit poly-clonal antibody against a recombinant human EXO-1 nuclease domainfragment characterized previously (27) (1:5,000; courtesy of Dr. D. M.Wilson III (NIA, NIH)) followed by goat anti-rabbit IgG conjugated tohorseradish peroxidase (Santa Cruz Biotechnology) or probed for WRNusing mouse anti-WRN monoclonal antibody (1:250; BD Pharmingen)followed by goat-anti-mouse IgG-horseradish peroxidase (Vector Labo-ratories). EXO-1 or WRN on immunoblot was detected using ECL Plus(Amersham Biosciences).
Coimmunoprecipitation of purified WRN and EXO-1 was performedin the presence of binding buffer (50 mM Tris, pH 8.0, 10% glycerol, 100mM NaCl, 0.01% Nonidet P-40) containing protease inhibitors (0.1 mM
phenylmethylsulfonyl fluoride, 1 �g/ml aprotinin, and 5 �g/ml leupep-tin). In a 100-�l reaction volume, 500 ng of purified WRN and 500 ng ofEXO-1 were incubated with goat anti-WRN polyclonal antibody (1:40)in binding buffer for 4 h at 4 °C. The protein complex was adsorbed onto protein G-agarose beads by incubating the mixture overnight at 4 °Cwith gentle rotation. The beads were washed three times with bindingbuffer, eluted by boiling with SDS sample buffer, and resolved on8–16% gradient SDS-polyacrylamide Tris-glycine gels. After transfer-ring the proteins to PVDF membranes, the membranes were blockedwith 5% nonfat dry milk in phosphate-buffered saline containing 0.1%Tween 20 and probed with either anti-EXO-1 or anti-WRN antibodiesas described above.
Far Western Blotting—Far Western blotting was conducted as de-scribed previously (40). Briefly, 0.2–1.0 �g of each protein was electro-phoresed on 8–16% SDS-polyacrylamide gels, transferred to PVDFmembranes, and processed as described previously (40) with the excep-tion that specified blocked membranes were incubated in the presenceof 0.5 �g/ml EXO-1. Membranes were washed, and conventional West-
ern analysis was then performed to detect the presence of EXO-1 usingrabbit anti-EXO-1 polyclonal antibody (described above) (1:5,000). Goatanti-rabbit IgG-horseradish peroxidase conjugate was used as second-ary antibody at a 1:10,000 dilution and detected using ECL Plus.
Oligonucleotide Substrates—PAGE-purified oligonucleotides (Mid-land Certified Reagent Co.) (Table I) were used for preparation ofsubstrates. Substrates were prepared as described previously (41).Briefly, 10 pmol of the appropriate flap oligonucleotide was 5�-radiola-beled with [�-32P]ATP and T4 polynucleotide kinase (New EnglandBiolabs) using the manufacturer’s protocol. Unincorporated nucleotidewas removed using a Sephadex G-25 spin column (Amersham Bio-sciences). The radiolabeled oligonucleotide was annealed to 25 pmol ofthe appropriate template oligonucleotide by heating at 95 °C, thencooling down from 70 to 24 °C. If necessary, 50 pmol of an upstreamoligonucleotide was then annealed to the duplex substrate by heating at37 °C for 1 h and slowly cooling to 24 °C. Two nicked duplex substrateswere prepared: 1) nicked duplex A using FLAP00A, TSTEM, and U25;and 2) nicked duplex B using FLAP00B, TSTEM2, and U21. The mo-nodeoxynucleotide (nt) flap was prepared with FLAP01, TSTEM, andU25. For the monoribodeoxynucleotide flap, FLAP01RNA, TSTEM, andU25 were used. The 10-nt flap was created with FLAP10, TSTEM2,and U21. The blunt ended duplex substrate was made using TSTEM-COMP and TSTEM.
For the nicked duplex substrate with a 3�-end label on the upstreamprimer, 10 pmol of U24 was annealed to 25 pmol of TSTEM and endlabeled with [�-32P]dCTP and Klenow fragment (New England Biolabs)at 25 °C for 20 min followed by an additional incubation for 20 min at25 °C with 50 �M unlabeled dCTP. Unincorporated nucleotide wasremoved by passing over two Sephadex G-25 columns. Klenow was thenheat inactivated at 95 °C followed by slow cooling to permit the oligo-nucleotides to reanneal. 50 pmol of FLAP00 was then added and an-nealed by heating at 37 °C for 1 h followed by slow cooling.
EXO-1 Incision Assay—20-�l reactions contained 0.5 nM DNA sub-strate (unless otherwise noted) and the indicated concentrations ofWRN and/or EXO-1 in 30 mM HEPES pH 7.6, 5% glycerol, 40 mM KCl,0.1 mg/ml bovine serum albumin, and 8 mM MgCl2. WRN was mixedwith the substrate on ice prior to the addition of EXO-1. Reactions wereincubated at 37 °C for 15 min (unless indicated otherwise), terminatedwith the addition of 10 �l of formamide dye (80% formamide (v/v), 0.1%bromphenol blue, and 0.1% xylene cyanol), and heated to 95 °C for 5min. Products were resolved on 20% polyacrylamide, 7 M urea denatur-ing gels. A PhosphorImager was used for detection, and the Image-Quant software (Molecular Dynamics) was used for quantitation of thereaction products. The percent incision was calculated as describedpreviously (24).
WRN Helicase Assay—20-�l reactions contained 0.5 nM 10-nt 5�-flapDNA substrate (except where indicated) and the indicated concentra-tions of WRN and/or EXO-1 in the same reaction buffer as used forEXO-1 incision assays except for the additional presence of 2 mM ATP.Reactions were incubated at 37 °C for 15 min, terminated with theaddition of 20 �l of helicase stop solution (50 mM EDTA, 40% glycerol,0.1% bromphenol blue, 0.1% xylene cyanol) containing a 10-fold excessof unlabeled oligonucleotide with the same sequence as the labeledstrand. The products of the helicase reactions were resolved on nonde-naturing 12% polyacrylamide gels. Radiolabeled DNA species in poly-acrylamide gels were visualized using a PhosphorImager and quanti-
TABLE IOligonucleotide sequences for DNA substrates (5� to 3�)
All oligonucleotides were PAGE-purified. The sequence in bold is single stranded when annealed to the proper template oligonucleotide. For theoligonucleotide with a single ribonucleotide, the ribonucleotide is indicated with an underline below the corresponding base.
Template Length Sequence
nt
TSTEM 44 GCACTGGCCGTCGTTTTACGGTCGTGACTGGGAAAACCCTGGCGTSTEM2 42 CCAGTGAATTCGAGCTCGGTACCCGCTAGCGGGGATCCTCTA
FlapsFLAP00A 19 GTAAAACGACGGCCAGTGCFLAP00B 21 TACCGAGCTCGAATTCACTGGFLAP01 20 AGTAAAACGACGGCCAGTGCFLAP01RNA 20 AGTAAAACGACGGCCAGTGCFLAP10 31 ATTGGTTATTTACCGAGCTCGAATTCACTGG
PrimersTSTEMCOMP 44 CGCCAGGGTTTTCCCAGTCACGACCGTAAAACGACGGCCAGTGCU21 21 TAGAGGATCCCCGCTAGCGGGU24 24 CGCCAGGGTTTTCCCAGTCACGACU25 25 CGCCAGGGTTTTCCCAGTCACGACC
Activation of EXO-1 Cleavage by WRN23488
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from
tated using the ImageQuant software. The percent helicase substrateunwound was calculated as described previously (42).
RESULTS
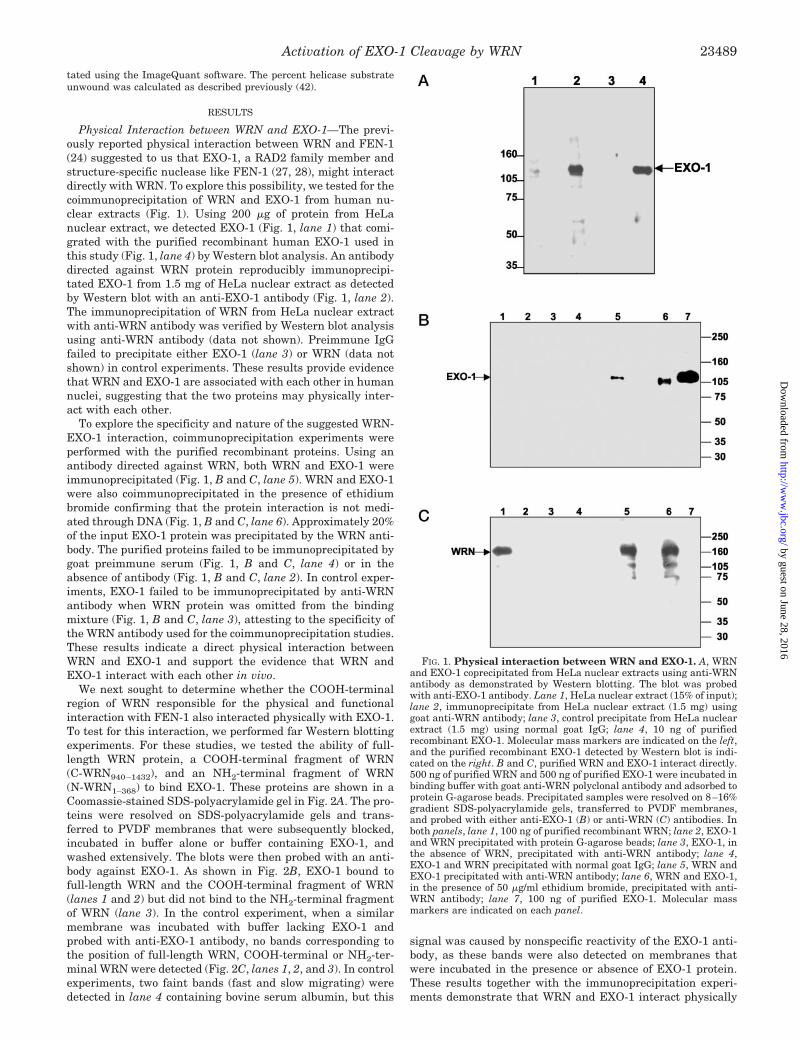
Physical Interaction between WRN and EXO-1—The previ-ously reported physical interaction between WRN and FEN-1(24) suggested to us that EXO-1, a RAD2 family member andstructure-specific nuclease like FEN-1 (27, 28), might interactdirectly with WRN. To explore this possibility, we tested for thecoimmunoprecipitation of WRN and EXO-1 from human nu-clear extracts (Fig. 1). Using 200 �g of protein from HeLanuclear extract, we detected EXO-1 (Fig. 1, lane 1) that comi-grated with the purified recombinant human EXO-1 used inthis study (Fig. 1, lane 4) by Western blot analysis. An antibodydirected against WRN protein reproducibly immunoprecipi-tated EXO-1 from 1.5 mg of HeLa nuclear extract as detectedby Western blot with an anti-EXO-1 antibody (Fig. 1, lane 2).The immunoprecipitation of WRN from HeLa nuclear extractwith anti-WRN antibody was verified by Western blot analysisusing anti-WRN antibody (data not shown). Preimmune IgGfailed to precipitate either EXO-1 (lane 3) or WRN (data notshown) in control experiments. These results provide evidencethat WRN and EXO-1 are associated with each other in humannuclei, suggesting that the two proteins may physically inter-act with each other.
To explore the specificity and nature of the suggested WRN-EXO-1 interaction, coimmunoprecipitation experiments wereperformed with the purified recombinant proteins. Using anantibody directed against WRN, both WRN and EXO-1 wereimmunoprecipitated (Fig. 1, B and C, lane 5). WRN and EXO-1were also coimmunoprecipitated in the presence of ethidiumbromide confirming that the protein interaction is not medi-ated through DNA (Fig. 1, B and C, lane 6). Approximately 20%of the input EXO-1 protein was precipitated by the WRN anti-body. The purified proteins failed to be immunoprecipitated bygoat preimmune serum (Fig. 1, B and C, lane 4) or in theabsence of antibody (Fig. 1, B and C, lane 2). In control exper-iments, EXO-1 failed to be immunoprecipitated by anti-WRNantibody when WRN protein was omitted from the bindingmixture (Fig. 1, B and C, lane 3), attesting to the specificity ofthe WRN antibody used for the coimmunoprecipitation studies.These results indicate a direct physical interaction betweenWRN and EXO-1 and support the evidence that WRN andEXO-1 interact with each other in vivo.
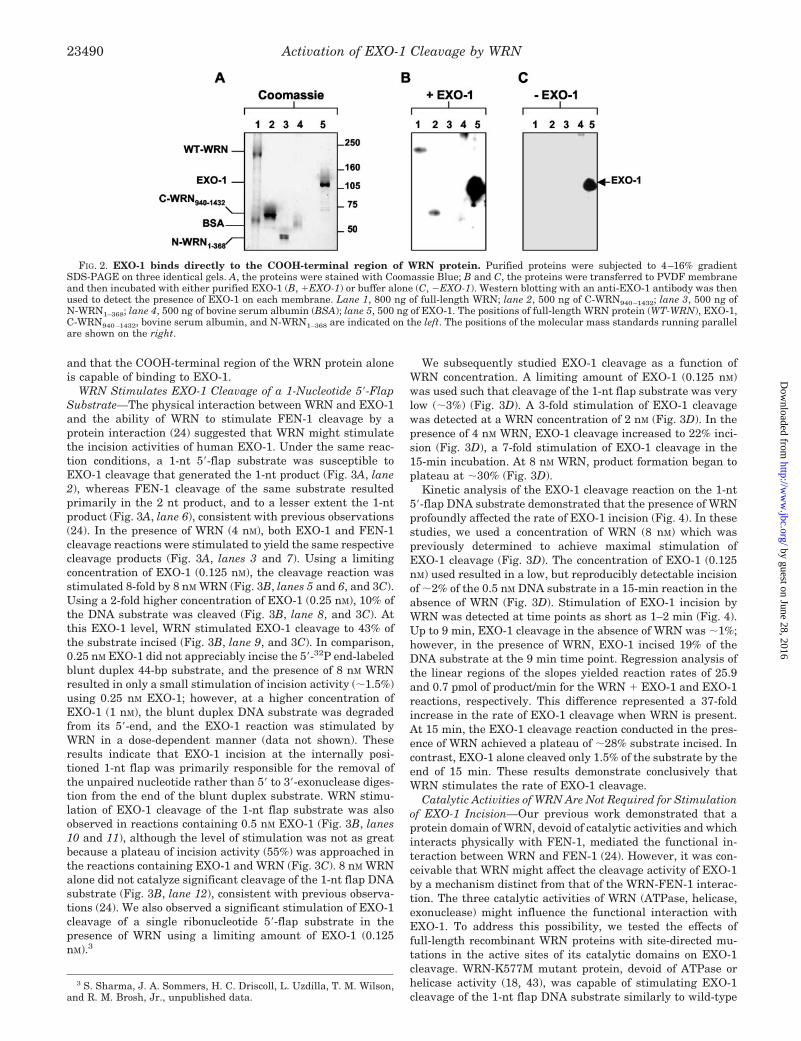
We next sought to determine whether the COOH-terminalregion of WRN responsible for the physical and functionalinteraction with FEN-1 also interacted physically with EXO-1.To test for this interaction, we performed far Western blottingexperiments. For these studies, we tested the ability of full-length WRN protein, a COOH-terminal fragment of WRN(C-WRN940–1432), and an NH2-terminal fragment of WRN(N-WRN1–368) to bind EXO-1. These proteins are shown in aCoomassie-stained SDS-polyacrylamide gel in Fig. 2A. The pro-teins were resolved on SDS-polyacrylamide gels and trans-ferred to PVDF membranes that were subsequently blocked,incubated in buffer alone or buffer containing EXO-1, andwashed extensively. The blots were then probed with an anti-body against EXO-1. As shown in Fig. 2B, EXO-1 bound tofull-length WRN and the COOH-terminal fragment of WRN(lanes 1 and 2) but did not bind to the NH2-terminal fragmentof WRN (lane 3). In the control experiment, when a similarmembrane was incubated with buffer lacking EXO-1 andprobed with anti-EXO-1 antibody, no bands corresponding tothe position of full-length WRN, COOH-terminal or NH2-ter-minal WRN were detected (Fig. 2C, lanes 1, 2, and 3). In controlexperiments, two faint bands (fast and slow migrating) weredetected in lane 4 containing bovine serum albumin, but this
signal was caused by nonspecific reactivity of the EXO-1 anti-body, as these bands were also detected on membranes thatwere incubated in the presence or absence of EXO-1 protein.These results together with the immunoprecipitation experi-ments demonstrate that WRN and EXO-1 interact physically
FIG. 1. Physical interaction between WRN and EXO-1. A, WRNand EXO-1 coprecipitated from HeLa nuclear extracts using anti-WRNantibody as demonstrated by Western blotting. The blot was probedwith anti-EXO-1 antibody. Lane 1, HeLa nuclear extract (15% of input);lane 2, immunoprecipitate from HeLa nuclear extract (1.5 mg) usinggoat anti-WRN antibody; lane 3, control precipitate from HeLa nuclearextract (1.5 mg) using normal goat IgG; lane 4, 10 ng of purifiedrecombinant EXO-1. Molecular mass markers are indicated on the left,and the purified recombinant EXO-1 detected by Western blot is indi-cated on the right. B and C, purified WRN and EXO-1 interact directly.500 ng of purified WRN and 500 ng of purified EXO-1 were incubated inbinding buffer with goat anti-WRN polyclonal antibody and adsorbed toprotein G-agarose beads. Precipitated samples were resolved on 8–16%gradient SDS-polyacrylamide gels, transferred to PVDF membranes,and probed with either anti-EXO-1 (B) or anti-WRN (C) antibodies. Inboth panels, lane 1, 100 ng of purified recombinant WRN; lane 2, EXO-1and WRN precipitated with protein G-agarose beads; lane 3, EXO-1, inthe absence of WRN, precipitated with anti-WRN antibody; lane 4,EXO-1 and WRN precipitated with normal goat IgG; lane 5, WRN andEXO-1 precipitated with anti-WRN antibody; lane 6, WRN and EXO-1,in the presence of 50 �g/ml ethidium bromide, precipitated with anti-WRN antibody; lane 7, 100 ng of purified EXO-1. Molecular massmarkers are indicated on each panel.
Activation of EXO-1 Cleavage by WRN 23489
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from
and that the COOH-terminal region of the WRN protein aloneis capable of binding to EXO-1.
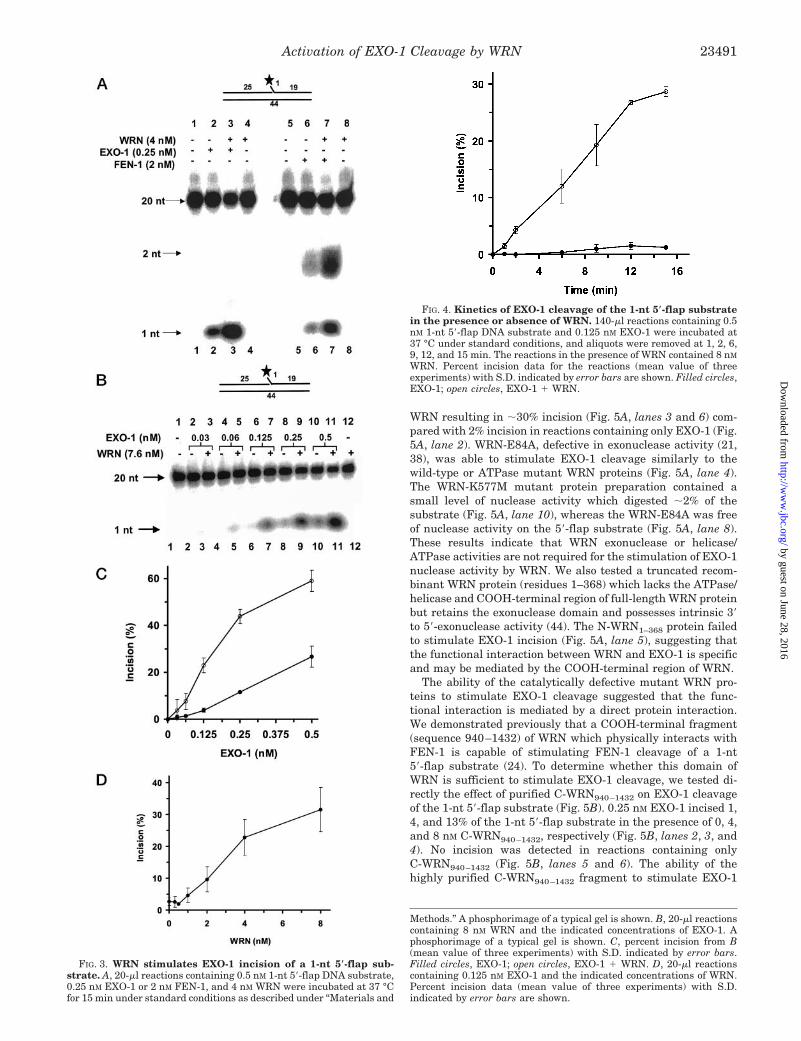
WRN Stimulates EXO-1 Cleavage of a 1-Nucleotide 5�-FlapSubstrate—The physical interaction between WRN and EXO-1and the ability of WRN to stimulate FEN-1 cleavage by aprotein interaction (24) suggested that WRN might stimulatethe incision activities of human EXO-1. Under the same reac-tion conditions, a 1-nt 5�-flap substrate was susceptible toEXO-1 cleavage that generated the 1-nt product (Fig. 3A, lane2), whereas FEN-1 cleavage of the same substrate resultedprimarily in the 2 nt product, and to a lesser extent the 1-ntproduct (Fig. 3A, lane 6), consistent with previous observations(24). In the presence of WRN (4 nM), both EXO-1 and FEN-1cleavage reactions were stimulated to yield the same respectivecleavage products (Fig. 3A, lanes 3 and 7). Using a limitingconcentration of EXO-1 (0.125 nM), the cleavage reaction wasstimulated 8-fold by 8 nM WRN (Fig. 3B, lanes 5 and 6, and 3C).Using a 2-fold higher concentration of EXO-1 (0.25 nM), 10% ofthe DNA substrate was cleaved (Fig. 3B, lane 8, and 3C). Atthis EXO-1 level, WRN stimulated EXO-1 cleavage to 43% ofthe substrate incised (Fig. 3B, lane 9, and 3C). In comparison,0.25 nM EXO-1 did not appreciably incise the 5�-32P end-labeledblunt duplex 44-bp substrate, and the presence of 8 nM WRNresulted in only a small stimulation of incision activity (�1.5%)using 0.25 nM EXO-1; however, at a higher concentration ofEXO-1 (1 nM), the blunt duplex DNA substrate was degradedfrom its 5�-end, and the EXO-1 reaction was stimulated byWRN in a dose-dependent manner (data not shown). Theseresults indicate that EXO-1 incision at the internally posi-tioned 1-nt flap was primarily responsible for the removal ofthe unpaired nucleotide rather than 5� to 3�-exonuclease diges-tion from the end of the blunt duplex substrate. WRN stimu-lation of EXO-1 cleavage of the 1-nt flap substrate was alsoobserved in reactions containing 0.5 nM EXO-1 (Fig. 3B, lanes10 and 11), although the level of stimulation was not as greatbecause a plateau of incision activity (55%) was approached inthe reactions containing EXO-1 and WRN (Fig. 3C). 8 nM WRNalone did not catalyze significant cleavage of the 1-nt flap DNAsubstrate (Fig. 3B, lane 12), consistent with previous observa-tions (24). We also observed a significant stimulation of EXO-1cleavage of a single ribonucleotide 5�-flap substrate in thepresence of WRN using a limiting amount of EXO-1 (0.125nM).3
We subsequently studied EXO-1 cleavage as a function ofWRN concentration. A limiting amount of EXO-1 (0.125 nM)was used such that cleavage of the 1-nt flap substrate was verylow (�3%) (Fig. 3D). A 3-fold stimulation of EXO-1 cleavagewas detected at a WRN concentration of 2 nM (Fig. 3D). In thepresence of 4 nM WRN, EXO-1 cleavage increased to 22% inci-sion (Fig. 3D), a 7-fold stimulation of EXO-1 cleavage in the15-min incubation. At 8 nM WRN, product formation began toplateau at �30% (Fig. 3D).
Kinetic analysis of the EXO-1 cleavage reaction on the 1-nt5�-flap DNA substrate demonstrated that the presence of WRNprofoundly affected the rate of EXO-1 incision (Fig. 4). In thesestudies, we used a concentration of WRN (8 nM) which waspreviously determined to achieve maximal stimulation ofEXO-1 cleavage (Fig. 3D). The concentration of EXO-1 (0.125nM) used resulted in a low, but reproducibly detectable incisionof �2% of the 0.5 nM DNA substrate in a 15-min reaction in theabsence of WRN (Fig. 3D). Stimulation of EXO-1 incision byWRN was detected at time points as short as 1–2 min (Fig. 4).Up to 9 min, EXO-1 cleavage in the absence of WRN was �1%;however, in the presence of WRN, EXO-1 incised 19% of theDNA substrate at the 9 min time point. Regression analysis ofthe linear regions of the slopes yielded reaction rates of 25.9and 0.7 pmol of product/min for the WRN � EXO-1 and EXO-1reactions, respectively. This difference represented a 37-foldincrease in the rate of EXO-1 cleavage when WRN is present.At 15 min, the EXO-1 cleavage reaction conducted in the pres-ence of WRN achieved a plateau of �28% substrate incised. Incontrast, EXO-1 alone cleaved only 1.5% of the substrate by theend of 15 min. These results demonstrate conclusively thatWRN stimulates the rate of EXO-1 cleavage.
Catalytic Activities of WRN Are Not Required for Stimulationof EXO-1 Incision—Our previous work demonstrated that aprotein domain of WRN, devoid of catalytic activities and whichinteracts physically with FEN-1, mediated the functional in-teraction between WRN and FEN-1 (24). However, it was con-ceivable that WRN might affect the cleavage activity of EXO-1by a mechanism distinct from that of the WRN-FEN-1 interac-tion. The three catalytic activities of WRN (ATPase, helicase,exonuclease) might influence the functional interaction withEXO-1. To address this possibility, we tested the effects offull-length recombinant WRN proteins with site-directed mu-tations in the active sites of its catalytic domains on EXO-1cleavage. WRN-K577M mutant protein, devoid of ATPase orhelicase activity (18, 43), was capable of stimulating EXO-1cleavage of the 1-nt flap DNA substrate similarly to wild-type
3 S. Sharma, J. A. Sommers, H. C. Driscoll, L. Uzdilla, T. M. Wilson,and R. M. Brosh, Jr., unpublished data.
FIG. 2. EXO-1 binds directly to the COOH-terminal region of WRN protein. Purified proteins were subjected to 4–16% gradientSDS-PAGE on three identical gels. A, the proteins were stained with Coomassie Blue; B and C, the proteins were transferred to PVDF membraneand then incubated with either purified EXO-1 (B, �EXO-1) or buffer alone (C, �EXO-1). Western blotting with an anti-EXO-1 antibody was thenused to detect the presence of EXO-1 on each membrane. Lane 1, 800 ng of full-length WRN; lane 2, 500 ng of C-WRN940–1432; lane 3, 500 ng ofN-WRN1–368; lane 4, 500 ng of bovine serum albumin (BSA); lane 5, 500 ng of EXO-1. The positions of full-length WRN protein (WT-WRN), EXO-1,C-WRN940–1432, bovine serum albumin, and N-WRN1–368 are indicated on the left. The positions of the molecular mass standards running parallelare shown on the right.
Activation of EXO-1 Cleavage by WRN23490
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from
WRN resulting in �30% incision (Fig. 5A, lanes 3 and 6) com-pared with 2% incision in reactions containing only EXO-1 (Fig.5A, lane 2). WRN-E84A, defective in exonuclease activity (21,38), was able to stimulate EXO-1 cleavage similarly to thewild-type or ATPase mutant WRN proteins (Fig. 5A, lane 4).The WRN-K577M mutant protein preparation contained asmall level of nuclease activity which digested �2% of thesubstrate (Fig. 5A, lane 10), whereas the WRN-E84A was freeof nuclease activity on the 5�-flap substrate (Fig. 5A, lane 8).These results indicate that WRN exonuclease or helicase/ATPase activities are not required for the stimulation of EXO-1nuclease activity by WRN. We also tested a truncated recom-binant WRN protein (residues 1–368) which lacks the ATPase/helicase and COOH-terminal region of full-length WRN proteinbut retains the exonuclease domain and possesses intrinsic 3�to 5�-exonuclease activity (44). The N-WRN1–368 protein failedto stimulate EXO-1 incision (Fig. 5A, lane 5), suggesting thatthe functional interaction between WRN and EXO-1 is specificand may be mediated by the COOH-terminal region of WRN.
The ability of the catalytically defective mutant WRN pro-teins to stimulate EXO-1 cleavage suggested that the func-tional interaction is mediated by a direct protein interaction.We demonstrated previously that a COOH-terminal fragment(sequence 940–1432) of WRN which physically interacts withFEN-1 is capable of stimulating FEN-1 cleavage of a 1-nt5�-flap substrate (24). To determine whether this domain ofWRN is sufficient to stimulate EXO-1 cleavage, we tested di-rectly the effect of purified C-WRN940–1432 on EXO-1 cleavageof the 1-nt 5�-flap substrate (Fig. 5B). 0.25 nM EXO-1 incised 1,4, and 13% of the 1-nt 5�-flap substrate in the presence of 0, 4,and 8 nM C-WRN940–1432, respectively (Fig. 5B, lanes 2, 3, and4). No incision was detected in reactions containing onlyC-WRN940–1432 (Fig. 5B, lanes 5 and 6). The ability of thehighly purified C-WRN940–1432 fragment to stimulate EXO-1
Methods.” A phosphorimage of a typical gel is shown. B, 20-�l reactionscontaining 8 nM WRN and the indicated concentrations of EXO-1. Aphosphorimage of a typical gel is shown. C, percent incision from B(mean value of three experiments) with S.D. indicated by error bars.Filled circles, EXO-1; open circles, EXO-1 � WRN. D, 20-�l reactionscontaining 0.125 nM EXO-1 and the indicated concentrations of WRN.Percent incision data (mean value of three experiments) with S.D.indicated by error bars are shown.
FIG. 3. WRN stimulates EXO-1 incision of a 1-nt 5�-flap sub-strate. A, 20-�l reactions containing 0.5 nM 1-nt 5�-flap DNA substrate,0.25 nM EXO-1 or 2 nM FEN-1, and 4 nM WRN were incubated at 37 °Cfor 15 min under standard conditions as described under “Materials and
FIG. 4. Kinetics of EXO-1 cleavage of the 1-nt 5�-flap substratein the presence or absence of WRN. 140-�l reactions containing 0.5nM 1-nt 5�-flap DNA substrate and 0.125 nM EXO-1 were incubated at37 °C under standard conditions, and aliquots were removed at 1, 2, 6,9, 12, and 15 min. The reactions in the presence of WRN contained 8 nM
WRN. Percent incision data for the reactions (mean value of threeexperiments) with S.D. indicated by error bars are shown. Filled circles,EXO-1; open circles, EXO-1 � WRN.
Activation of EXO-1 Cleavage by WRN 23491
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from
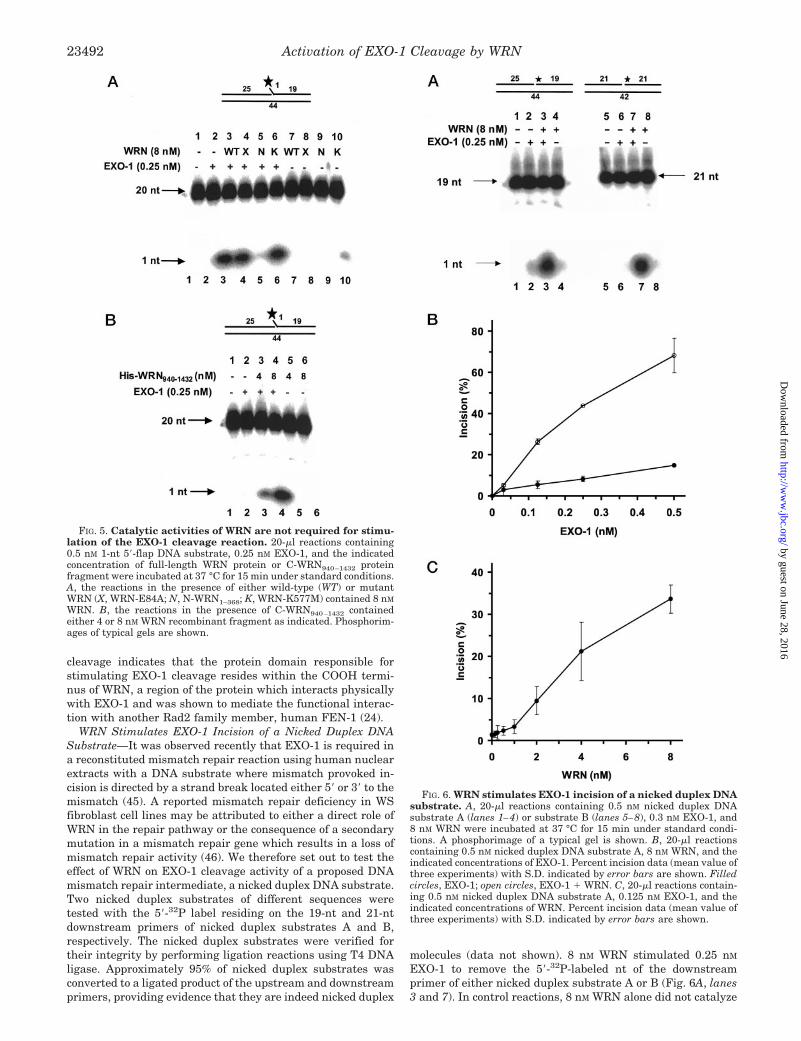
cleavage indicates that the protein domain responsible forstimulating EXO-1 cleavage resides within the COOH termi-nus of WRN, a region of the protein which interacts physicallywith EXO-1 and was shown to mediate the functional interac-tion with another Rad2 family member, human FEN-1 (24).
WRN Stimulates EXO-1 Incision of a Nicked Duplex DNASubstrate—It was observed recently that EXO-1 is required ina reconstituted mismatch repair reaction using human nuclearextracts with a DNA substrate where mismatch provoked in-cision is directed by a strand break located either 5� or 3� to themismatch (45). A reported mismatch repair deficiency in WSfibroblast cell lines may be attributed to either a direct role ofWRN in the repair pathway or the consequence of a secondarymutation in a mismatch repair gene which results in a loss ofmismatch repair activity (46). We therefore set out to test theeffect of WRN on EXO-1 cleavage activity of a proposed DNAmismatch repair intermediate, a nicked duplex DNA substrate.Two nicked duplex substrates of different sequences weretested with the 5�-32P label residing on the 19-nt and 21-ntdownstream primers of nicked duplex substrates A and B,respectively. The nicked duplex substrates were verified fortheir integrity by performing ligation reactions using T4 DNAligase. Approximately 95% of nicked duplex substrates wasconverted to a ligated product of the upstream and downstreamprimers, providing evidence that they are indeed nicked duplex
molecules (data not shown). 8 nM WRN stimulated 0.25 nM
EXO-1 to remove the 5�-32P-labeled nt of the downstreamprimer of either nicked duplex substrate A or B (Fig. 6A, lanes3 and 7). In control reactions, 8 nM WRN alone did not catalyze
FIG. 5. Catalytic activities of WRN are not required for stimu-lation of the EXO-1 cleavage reaction. 20-�l reactions containing0.5 nM 1-nt 5�-flap DNA substrate, 0.25 nM EXO-1, and the indicatedconcentration of full-length WRN protein or C-WRN940–1432 proteinfragment were incubated at 37 °C for 15 min under standard conditions.A, the reactions in the presence of either wild-type (WT) or mutantWRN (X, WRN-E84A; N, N-WRN1–368; K, WRN-K577M) contained 8 nM
WRN. B, the reactions in the presence of C-WRN940–1432 containedeither 4 or 8 nM WRN recombinant fragment as indicated. Phosphorim-ages of typical gels are shown.
FIG. 6. WRN stimulates EXO-1 incision of a nicked duplex DNAsubstrate. A, 20-�l reactions containing 0.5 nM nicked duplex DNAsubstrate A (lanes 1–4) or substrate B (lanes 5–8), 0.3 nM EXO-1, and8 nM WRN were incubated at 37 °C for 15 min under standard condi-tions. A phosphorimage of a typical gel is shown. B, 20-�l reactionscontaining 0.5 nM nicked duplex DNA substrate A, 8 nM WRN, and theindicated concentrations of EXO-1. Percent incision data (mean value ofthree experiments) with S.D. indicated by error bars are shown. Filledcircles, EXO-1; open circles, EXO-1 � WRN. C, 20-�l reactions contain-ing 0.5 nM nicked duplex DNA substrate A, 0.125 nM EXO-1, and theindicated concentrations of WRN. Percent incision data (mean value ofthree experiments) with S.D. indicated by error bars are shown.
Activation of EXO-1 Cleavage by WRN23492
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from
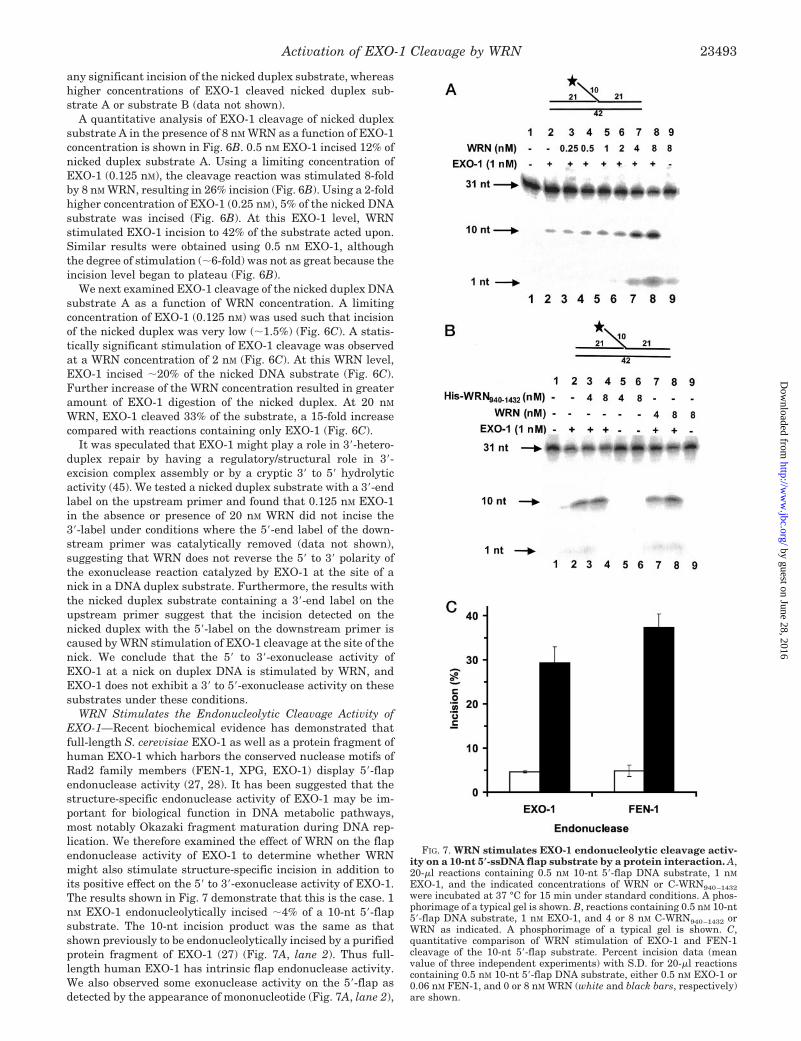
any significant incision of the nicked duplex substrate, whereashigher concentrations of EXO-1 cleaved nicked duplex sub-strate A or substrate B (data not shown).
A quantitative analysis of EXO-1 cleavage of nicked duplexsubstrate A in the presence of 8 nM WRN as a function of EXO-1concentration is shown in Fig. 6B. 0.5 nM EXO-1 incised 12% ofnicked duplex substrate A. Using a limiting concentration ofEXO-1 (0.125 nM), the cleavage reaction was stimulated 8-foldby 8 nM WRN, resulting in 26% incision (Fig. 6B). Using a 2-foldhigher concentration of EXO-1 (0.25 nM), 5% of the nicked DNAsubstrate was incised (Fig. 6B). At this EXO-1 level, WRNstimulated EXO-1 incision to 42% of the substrate acted upon.Similar results were obtained using 0.5 nM EXO-1, althoughthe degree of stimulation (�6-fold) was not as great because theincision level began to plateau (Fig. 6B).
We next examined EXO-1 cleavage of the nicked duplex DNAsubstrate A as a function of WRN concentration. A limitingconcentration of EXO-1 (0.125 nM) was used such that incisionof the nicked duplex was very low (�1.5%) (Fig. 6C). A statis-tically significant stimulation of EXO-1 cleavage was observedat a WRN concentration of 2 nM (Fig. 6C). At this WRN level,EXO-1 incised �20% of the nicked DNA substrate (Fig. 6C).Further increase of the WRN concentration resulted in greateramount of EXO-1 digestion of the nicked duplex. At 20 nM
WRN, EXO-1 cleaved 33% of the substrate, a 15-fold increasecompared with reactions containing only EXO-1 (Fig. 6C).
It was speculated that EXO-1 might play a role in 3�-hetero-duplex repair by having a regulatory/structural role in 3�-excision complex assembly or by a cryptic 3� to 5� hydrolyticactivity (45). We tested a nicked duplex substrate with a 3�-endlabel on the upstream primer and found that 0.125 nM EXO-1in the absence or presence of 20 nM WRN did not incise the3�-label under conditions where the 5�-end label of the down-stream primer was catalytically removed (data not shown),suggesting that WRN does not reverse the 5� to 3� polarity ofthe exonuclease reaction catalyzed by EXO-1 at the site of anick in a DNA duplex substrate. Furthermore, the results withthe nicked duplex substrate containing a 3�-end label on theupstream primer suggest that the incision detected on thenicked duplex with the 5�-label on the downstream primer iscaused by WRN stimulation of EXO-1 cleavage at the site of thenick. We conclude that the 5� to 3�-exonuclease activity ofEXO-1 at a nick on duplex DNA is stimulated by WRN, andEXO-1 does not exhibit a 3� to 5�-exonuclease activity on thesesubstrates under these conditions.
WRN Stimulates the Endonucleolytic Cleavage Activity ofEXO-1—Recent biochemical evidence has demonstrated thatfull-length S. cerevisiae EXO-1 as well as a protein fragment ofhuman EXO-1 which harbors the conserved nuclease motifs ofRad2 family members (FEN-1, XPG, EXO-1) display 5�-flapendonuclease activity (27, 28). It has been suggested that thestructure-specific endonuclease activity of EXO-1 may be im-portant for biological function in DNA metabolic pathways,most notably Okazaki fragment maturation during DNA rep-lication. We therefore examined the effect of WRN on the flapendonuclease activity of EXO-1 to determine whether WRNmight also stimulate structure-specific incision in addition toits positive effect on the 5� to 3�-exonuclease activity of EXO-1.The results shown in Fig. 7 demonstrate that this is the case. 1nM EXO-1 endonucleolytically incised �4% of a 10-nt 5�-flapsubstrate. The 10-nt incision product was the same as thatshown previously to be endonucleolytically incised by a purifiedprotein fragment of EXO-1 (27) (Fig. 7A, lane 2). Thus full-length human EXO-1 has intrinsic flap endonuclease activity.We also observed some exonuclease activity on the 5�-flap asdetected by the appearance of mononucleotide (Fig. 7A, lane 2),
FIG. 7. WRN stimulates EXO-1 endonucleolytic cleavage activ-ity on a 10-nt 5�-ssDNA flap substrate by a protein interaction. A,20-�l reactions containing 0.5 nM 10-nt 5�-flap DNA substrate, 1 nM
EXO-1, and the indicated concentrations of WRN or C-WRN940–1432were incubated at 37 °C for 15 min under standard conditions. A phos-phorimage of a typical gel is shown. B, reactions containing 0.5 nM 10-nt5�-flap DNA substrate, 1 nM EXO-1, and 4 or 8 nM C-WRN940–1432 orWRN as indicated. A phosphorimage of a typical gel is shown. C,quantitative comparison of WRN stimulation of EXO-1 and FEN-1cleavage of the 10-nt 5�-flap substrate. Percent incision data (meanvalue of three independent experiments) with S.D. for 20-�l reactionscontaining 0.5 nM 10-nt 5�-flap DNA substrate, either 0.5 nM EXO-1 or0.06 nM FEN-1, and 0 or 8 nM WRN (white and black bars, respectively)are shown.
Activation of EXO-1 Cleavage by WRN 23493
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from

a result consistent with that observed when the purified nucle-ase domain fragment of EXO-1 was incubated with the sameflap substrate (27). At low concentrations of WRN (0.5 and 1nM), a small stimulation of EXO-1 incision (�1.5-fold) wasobserved (Fig. 7A, lanes 4 and 5). Using 2 nM WRN, EXO-1cleaved 9% of the 10-nt 5�-flap DNA substrate (Fig. 7A, lane 6).EXO-1 cleavage was elevated further in the presence of 4 or 8nM WRN, yielding 21 and 31% incision, respectively (Fig. 7A,lanes 7 and 8). Importantly, in control reactions 8 nM WRNalone did not incise the 10-nt 5�-flap structure (Fig. 7A, lane 9),as reported previously (42). At high concentrations of WRN (8nM) (lane 9), a very low level of 5�-nuclease activity could bedetected which was also evident in reactions containing exonu-clease-defective WRN (data not shown), suggesting that a mi-nor contaminant in the WRN protein preparation is responsiblefor this activity. Thus WRN is competent to stimulate theEXO-1 endonucleolytic cleavage reaction, as great as 8-fold, onthe 10-nt 5�-flap substrate. We also observed stimulation of theEXO-1 5� to 3�-exonuclease activity on the 5�-ssDNA flap byWRN, resulting in 5 and 17% incision at WRN concentrationsof 4 and 8 nM, respectively (Fig. 7A, lanes 7 and 8). The abilityof WRN to stimulate 5� to 3�-exonuclease activity of EXO-1 onthe 5�-ssDNA flap oligonucleotide is consistent with our obser-vation that EXO-1 incision of 5�-32P-labeled ssDNA oligonu-cleotide is also stimulated by WRN (data not shown).
We next tested the purified COOH-terminal protein frag-ment of WRN (C-WRN940–1432), devoid of the helicase andexonuclease domains, for its ability to stimulate EXO-1 cleav-age of the 10-nt 5�-flap substrate. Using 1 nM EXO-1, thecleavage reaction was stimulated �15-fold by 4 nM
C-WRN940–1432 (Fig. 7B, lane 3). Slightly higher EXO-1 cleav-age (37%) was observed using 8 nM C-WRN940–1432 (Fig. 7B,lane 4). The 10-nt product from reactions containingC-WRN940–1432 and EXO-1 were also detected from reactionscontaining EXO-1 � full-length WRN (Fig. 7A) or higher con-centrations of EXO-1 (data not shown), indicating that thecleavage site specificity of EXO-1 is not altered by the WRNCOOH-terminal protein fragment. Purified N-WRN proteinfragment failed to stimulate endonucleolytic or exonucleolyticincision by EXO-1 (data not shown), adding further evidencethat the functional interaction between WRN and EXO-1 is theresult of a protein interaction mediated by the COOH-terminalregion of WRN. The levels of stimulation of EXO-1 endonucleo-lytic cleavage by C-WRN940–1432 were comparable with thoseobserved with the full-length WRN protein (Fig. 7B, lanes 7and 8). These results indicate that the protein domain of WRNwhich physically binds to EXO-1 is capable of stimulatingEXO-1 cleavage of the 10-nt 5�-flap substrate.
WRN Stimulates EXO-1 Cleavage as Effectively as FEN-1Cleavage—Because WRN can stimulate either EXO-1 or FEN-1endonucleolytic incision of 5�-flap substrates, we wanted tocompare the -fold stimulation by WRN of the two human Rad2nucleases. Using the 10-nt 5�-flap substrate, WRN was testedfor its effect on EXO-1 or FEN-1 cleavage under the identicalreaction conditions. The concentrations of EXO-1 (0.5 nM) andFEN-1 (0.06 nM) were adjusted to achieve a similar level ofendonucleolytic cleavage (�5%) of the 5�-flap substrate (Fig.7C). In the presence of 8 nM WRN, EXO-1 cleavage was in-creased 6-fold, and FEN-1 cleavage was increased 7-fold (Fig.7C). These results indicate that the stimulatory effect of WRNon flap cleavage by the two human endonucleases is verysimilar.
Both RPA and PCNA stimulate FEN-1 cleavage (41, 47),raising the possibility that these human nuclear proteins mightstimulate EXO-1 cleavage; however, there is no previous reportof the effect of RPA or PCNA on human EXO-1 cleavage. At
molar concentrations of PCNA homotrimer or RPA heterotri-mer equal to WRN monomer (8 nM) that displayed a 16-foldstimulation of EXO-1 endonucleolytic cleavage of the 10-nt flapsubstrate, we did not detect any significant stimulation ofEXO-1 cleavage (endonucleolytic or exonucleolytic) by eitherPCNA or RPA (data not shown).
WRN Stimulates EXO-1 Endonucleolytic Cleavage underConditions Favorable for Helicase-catalyzed DNA Unwind-ing—Recently, we reported that WRN helicase efficiently un-winds a 10-nt 5�-flap substrate, releasing the 5�-flap oligonu-cleotide, leaving the duplex species containing the upstreamoligonucleotide intact (42). To determine the outcome of 5�-flapprocessing by EXO-1 on a substrate that is also unwound byWRN, we examined both DNA unwinding (release of the 5�-flapoligonucleotide) and EXO-1 cleavage of the 5�-flap oligonucleo-tide in the presence of ATP, which fuels the WRN helicasereaction. The products were analyzed on nondenaturing poly-acrylamide gels, and the results of a typical experiment areshown in Fig. 8. 4 or 8 nM WRN stimulated cleavage of the flapsubstrate by 0.5 nM EXO-1 �10-fold and 6-fold, respectively(Fig. 8, lanes 3 and 4) as detected by the increased amount ofthe 10-nt incision product that comigrated with the radiola-beled 10-nt marker indicated on the left. Under these condi-tions in the presence of 0.5 nM EXO-1, 4 or 8 nM WRN unwound8 and 39% of the flap substrate, respectively (Fig. 8, lanes 3 and4). The oligonucleotide released by WRN helicase activity comi-grated with the intact 31-mer (FLAP10) used in the prepara-tion of the 5�-flap substrate. Similar levels of WRN helicaseactivity (14 and 44%) were detected in reactions containingonly 4 or 8 nM WRN (lanes 5 and 6). When EXO-1 was increasedto 2 nM, 7% of the 5�-flap substrate was endonucleolyticallyincised in the absence of WRN (lane 7). In the presence of WRN,endonucleolytic incision by EXO-1 was again increased signif-icantly as detected by the appearance of the 10-nt incisionproduct (lanes 8 and 9). The percent endonucleolytic incision by2 nM EXO-1 in the presence of 4 or 8 nM WRN was 41 and 18%,respectively. The reduction in EXO-1 10-nt cleavage product inthe presence of 8 nM WRN compared with 4 nM WRN is accom-panied by a greater amount of the intact flap oligonucleotidereleased. WRN also stimulates EXO-1 degradation of the re-
FIG. 8. WRN stimulates EXO-1 endonucleolytic incision of a5�-flap substrate under conditions suitable for DNA unwinding.20-�l reactions containing 0.5 nM 10-nt 5�-flap DNA substrate, 0.5 or 2nM EXO-1, and 4 or 8 nM WRN were incubated at 37 °C for 15 min understandard conditions in the presence of 2 mM ATP. Reaction productswere resolved on 12% polyacrylamide nondenaturing gels. A phospho-rimage of a typical gel is shown. Filled triangle, heat-denatured DNAsubstrate control. Markers shown on the left are: 5�-32P-labeled FLAP10oligonucleotide, 5�-32P-labeled incised 10-mer, and [32P]AMP.
Activation of EXO-1 Cleavage by WRN23494
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from

leased 5�-ssDNA flap and/or the 5�-ssDNA end of the intact flapstructure as evidenced by comigration of the fast migratingspecies (lanes 3, 4, 8, and 9) with [32P]AMP. Only 19% of theintact 31-mer was released by 8 nM WRN in the presence of 2nM EXO-1 compared with 44% flap oligonucleotide released byWRN alone, indicating a 2.3-fold reduction in the release ofintact flap oligonucleotide. These results indicate that underconditions in which WRN is active as a helicase (presence ofATP), WRN can stimulate EXO-1 endonucleolytic incision orunwind the 5�-flap oligonucleotide.
DISCUSSION
In this study we have demonstrated that WRN and EXO-1interact physically as evidenced by their coimmunoprecipita-tion using either human extracts or purified recombinant pro-teins. The region of WRN which interacts physically withEXO-1 was mapped to the COOH-terminal region of WRN byfar Western analysis. WRN robustly stimulates EXO-1 cleav-age by a direct protein interaction that is independent of en-ergy and catalytic activity. WRN exerts a marked enhancement(37-fold) in the rate of EXO-1 incision. Although a greateramount of WRN monomer compared with EXO-1 was neces-sary to achieve maximal stimulation of EXO-1 nuclease activ-ity, the functional assembly state of WRN is not known andmay assume an oligomeric state as proposed for some helicases.On a stoichiometric basis, WRN monomer is far superior toPCNA homotrimer or RPA heterotrimer in stimulating EXO-1cleavage of short and long 5�-flap substrates. The results fromthese biochemical studies clearly indicate that WRN proteinstrongly stimulates the endonucleolytic and exonucleolytic in-cision activities of EXO-1 on proposed DNA intermediates ofreplication and repair. Importantly, WRN is the first proteinidentified that modulates the catalytic activities of EXO-1.
The ability of WRN to stimulate the incision reactions cata-lyzed by the sequence-related human nucleases EXO-1 andFEN-1 on short (1-nt) and long (10-nt) 5�-ssDNA flap sub-strates suggests that the functional interaction may proceed bya similar mechanism. In both cases, the stimulation of incisionis mediated by a COOH-terminal domain of the WRN protein.Although the WRN interaction site on FEN-1 is not known, itwill be of interest to determine whether the site of WRN phys-ical interaction is the same for EXO-1 and FEN-1. A number ofsimilarities exist between EXO-1 and FEN-1, but there aresome noticeable differences in their DNA substrate specificitiesand cleavage products (27, 28). For the structures tested thusfar, the DNA substrate specificities of EXO-1 and FEN-1 arenot altered by the presence of WRN. WRN stimulates EXO-1cleavage of ssDNA and blunt duplex dsDNA, whereas FEN-1acts poorly on these substrates, and WRN does not appreciablystimulate FEN-1 on either of these structures (24). AlthoughWRN is able to stimulate structure-specific endonucleolyticcleavage of 5�-flap structures by EXO-1 and FEN-1, the nucle-ase reactions display a subtle difference in the products gener-ated. WRN stimulation of EXO-1 yields a 1-nt product from the1-nt 5�-flap substrate, consistent with the cleavage productsobtained from reactions containing higher levels of EXO-1alone. Thus EXO-1 cleaves primarily at the base of the flap,yielding a nicked duplex intermediate that can be readilysealed by ligase. In contrast, FEN-1 (in the absence or presenceof WRN) incises primarily 1 nt into the downstream annealedregion, and to a lesser extent at the base of the flap, resultingin primarily a 1-nt gap duplex and a lower amount of nickedduplex, respectively (24, 41). This difference may be importantduring Okazaki fragment processing of double flap substratesin which the upstream primer has a single unannealed nt, apreferred DNA substrate for FEN-1 cleavage (48). By the pref-erential FEN-1 incision of the 5�-flap 1 nt into the annealed
downstream region, the unannealed (but complementary) nt atthe 3�-end of the upstream primer may anneal to form a ligat-able nicked DNA duplex substrate. It will be of interest todetermine whether human EXO-1 also prefers a double flapstructure as a cleavage substrate; alternatively, the cleavagespecificity of EXO-1 may be well suited to process 5�-flap struc-tures that lack 3�-flaps, a reaction that may also be importantduring eukaryotic DNA replication or some other pathway ofDNA metabolism.
In which cellular pathways is the functional interaction ofWRN and EXO-1 likely to be important? Yeast genetic studieshave implicated EXO-1 in spontaneous mitotic and meioticrecombination between direct repeats (29). In one model, the 5�to 3�-exonuclease activity of EXO-1 may act to generate 3�-ssDNA tails that can be used by homologous pairing proteinsfor recombination. EXO-1 has also been implicated in the proc-essing of complementary double strand ends in a recombina-tion pathway of double strand break repair. More specifically,it was recently proposed that the 5� to 3�-exonuclease activity ofEXO-1 partially compensates for a deficiency in the function ofthe Rad50�Mre11�Xrs2 complex in homologous recombinationrepair (32, 34). Although a precise role of the WRN protein in aspecific recombination pathway remains to be defined, thereare several lines of evidence suggesting that a role of WRN inrecombination is likely. 1) WS cells display severe genomicinstability, elevated homologous recombination (9), and aber-rant mitotic recombination (10, 49). 2) the human WRN genecan suppress the increased homologous and illegitimate recom-bination in the yeast WRN homolog sgs1 (50). 3) WRN helicasecan recognize and unwind a number of DNA structures withjunctions (42) and catalyzes branch fork migration of Hollidayjunctions (51), a key DNA intermediate of homologous recom-bination. The results presented in this report suggest thatWRN may function to modulate the cleavage activity of EXO-1which is thought to be essential to the role of the enzyme inhomologous recombination.
EXO-1 has been implicated in mismatch repair by its re-quirement in mutation avoidance (30) and interactions withmismatch repair proteins (33, 35). Very recently, EXO-1 wasdemonstrated to be required in mismatch repair in a reconsti-tuted system using extracts from human cells (45). HumanEXO-1 was shown to perform mismatch-provoked excision di-rected by a strand break located either 5� or 3� to the mispair.Evidence that WRN protein may play a role in mismatch repairwas provided by the demonstration that extracts from threeWS fibroblastoid cell lines were deficient in repair of base-baseand insertion/deletion mismatches (46). However, in the samestudy, extracts from four WS lymphoblastoid cell lines were allproficient in mismatch repair, suggesting that WRN proteincould have a cell type- and/or tissue-specific role in mismatchrepair. It was reported subsequently that introduction of hu-man chromosome 8 encoding the WRN gene to WS fibroblastcell lines deficient in mismatch repair resulted in expression ofWRN protein but did not restore mismatch repair activity (52).Thus, it is unclear whether or not WRN plays a direct role inmismatch repair. Based on our results that WRN and EXO-1interact functionally, it is possible that WRN modulates EXO-1activity in a pathway of mismatch repair. The elevated fre-quency of spontaneous mutations in the HPRT gene (4) and thepronounced genomic instability of WS cells (2, 3) may reflect atleast in part a defect in a mutation avoidance pathway thatrelies on the mismatch repair machinery, including EXO-1.
It has been suggested that EXO-1 plays a role in Okazakifragment processing in yeast that is at least partially redun-dant to FEN-1 (RAD27). On a biochemical level, EXO-1 is ableto remove RNA primers from Okazaki fragment model sub-
Activation of EXO-1 Cleavage by WRN 23495
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from

strates efficiently, suggesting that the enzyme may function inRNA primer removal during lagging strand synthesis (30). Theability of human EXO-1 to suppress the lethality of a rad27mutant at the restrictive temperature is consistent with thisnotion (30). WRN stimulation of EXO-1 5�-flap cleavage mayserve a function during Okazaki fragment maturation or theprocessing of other DNA structures at the replication fork.Evidence presented here demonstrates that WRN is able tostimulate EXO-1 cleavage of flap structures with 5�-ssDNAtails up to at least 10 nt, an activity that may be important ina FEN-1 independent pathway of 5�-flap processing (53). Alter-natively, EXO-1 may play a role in the removal of the terminalribonucleotide at the RNA-DNA junction during Okazaki frag-ment processing in a WRN-stimulated reaction. WRN mayserve as a general factor to promote replication intermediateprocessing through its interaction with structure-specificendonucleases.
Very recently it was reported that EXO-1-dependent ssDNAat telomeres is required for cell cycle arrest in budding yeastku70 deletion mutants (54). The observation that EXO-1 affectsthe metabolism of damaged telomeres and checkpoint re-sponses is interesting in light of circumstantial evidence thatWRN may also play a role in telomere metabolism. WS fibro-blasts that express a transfected human telomerase (hTERT)gene have an extended lifespan and can become immortalized(55), suggesting that the absence of WRN protein confers ac-celerated cellular senescence in a pathway dependent on telom-erase. However, WRN may also function in an alternate telom-erase-independent mechanism, as suggested by studies of theyeast WRN homolog Sgs1 (56, 57). Although no direct role ofWRN protein in telomere metabolism has been established, itis conceivable that WRN stimulation of EXO-1 nuclease activ-ity in subtelomeric regions is important for the replication andstability of telomeres and/or the activation of the appropriateDNA damage and signal checkpoint pathways to tolerate dam-aged telomeres.
Acknowledgments—We thank our colleagues in the Department ofRadiation Oncology (University of Maryland, Baltimore) and the Lab-oratory of Molecular Gerontology (NIA, National Institutes of Health)for helpful discussion. We thank Drs. Patricia Gearhart and DavidWilson III (NIA, National Institutes of Health) for a critical reading ofthe manuscript. We are grateful to Dr. David Wilson III for the antibodyagainst a purified recombinant human EXO-1 fragment. We also thankDr. Mark Kenny for providing human PCNA and RPA purified proteins.
REFERENCES
1. Martin, G. M. (1978) Birth Defects Orig. Artic. Ser. 14, 5–392. Salk, D., Au, K., Hoehn, H., and Martin, G. M. (1981) Cytogenet. Cell Genet. 30,
92–1073. Salk, D., Bryant, E., Hoehn, H., Johnston, P., and Martin, G. M. (1985) Adv.
Exp. Med. Biol. 190, 305–3114. Fukuchi, K., Martin, G. M., and Monnat, R. J. J. (1989) Proc. Natl. Acad. Sci.
U. S. A. 86, 5893–58975. Martin, G. M., Sprague, C. A., and Epstein, C. J. (1970) Lab. Invest. 23, 86–926. Takeuchi, F., Hanaoka, F., Goto, M., Akaoka, I., Hori, T., Yamada, M., and
Miyamoto, T. (1982) Hum. Genet. 60, 365–3687. Hanaoka, F., Yamada, M., Takeuchi, F., Goto, M., Miyamoto, T., and Hori, T.
(1985) Adv. Exp. Med. Biol. 190, 439–4578. Poot, M., Hoehn, H., Runger, T. M., and Martin, G. M. (1992) Exp. Cell Res.
202, 267–2739. Cheng, R. Z., Murano, S., Kurz, B., and Shmookler, R. R. (1990) Mutat. Res.
237, 259–26910. Prince, P. R., Emond, M. J., and Monnat, R. J. J. (2001) Genes Dev. 15, 933–93811. Schulz, V. P., Zakian, V. A., Ogburn, C. E., McKay, J., Jarzebowicz, A. A.,
Edland, S. D., and Martin, G. M. (1996) Hum. Genet. 97, 750–75412. Ogburn, C. E., Oshima, J., Poot, M., Chen, R., Hunt, K. E., Gollahon, K. A.,
Rabinovitch, P. S., and Martin, G. M. (1997) Hum. Genet. 101, 121–12513. Lebel, M., and Leder, P. (1998) Proc. Natl. Acad. Sci. U. S. A. 95, 13097–13102
14. Poot, M., Gollahon, K. A., and Rabinovitch, P. S. (1999) Hum. Genet. 104,10–14
15. Pichierri, P., Franchitto, A., Mosesso, P., and Palitti, F. (2000) Mutat. Res. 456,45–57
16. Poot, M., Yom, J. S., Whang, S. H., Kato, J. T., Gollahon, K. A., andRabinovitch, P. S. (2001) FASEB J. 15, 1224–1226
17. Yu, C. E., Oshima, J., Fu, Y. H., Wijsman, E. M., Hisama, F., Alisch, R.,Matthews, S., Nakura, J., Miki, T., Ouais, S., Martin, G. M., Mulligan, J.,and Schellenberg, G. D. (1996) Science 272, 258–262
18. Gray, M. D., Shen, J. C., Kamath-Loeb, A. S., Blank, A., Sopher, B. L., Martin,G. M., Oshima, J., and Loeb, L. A. (1997) Nat. Genet. 17, 100–103
19. Suzuki, N., Shimamoto, A., Imamura, O., Kuromitsu, J., Kitao, S., Goto, M.,and Furuichi, Y. (1997) Nucleic Acids Res. 25, 2973–2978
20. Shen, J. C., Gray, M. D., Oshima, J., Kamath-Loeb, A. S., Fry, M., and Loeb,L. A. (1998) J. Biol. Chem. 273, 34139–34144
21. Huang, S., Li, B., Gray, M. D., Oshima, J., Mian, I. S., and Campisi, J. (1998)Nat. Genet. 20, 114–116
22. Kamath-Loeb, A. S., Shen, J. C., Loeb, L. A., and Fry, M. (1998) J. Biol. Chem.273, 34145–34150
23. Brosh, R. M., Jr., and Bohr, V. A. (2002) Exp. Gerontol. 37, 491–50624. Brosh, R. M., Jr., von Kobbe, C., Sommers, J. A., Karmakar, P., Opresko, P. L.,
Piotrowski, J., Dianova, I., Dianov, G. L., and Bohr, V. A. (2001) EMBO J.20, 5791–5801
25. Lieber, M. R. (1997) Bioessays 19, 233–24026. Wilson, D. M., Carney, J. P., Coleman, M. A., Adamson, A. W., Christensen,
M., and Lamerdin, J. E. (1998) Nucleic Acids Res. 26, 3762–376827. Lee, B. I., and Wilson, D. M. (1999) J. Biol. Chem. 274, 37763–3776928. Tran, P. T., Erdeniz, N., Dudley, S., and Liskay, R. M. (2002) DNA Repair 1,
895–91229. Fiorentini, P., Huang, K. N., Tishkoff, D. X., Kolodner, R. D., and Symington,
L. S. (1997) Mol. Cell. Biol. 17, 2764–277330. Qiu, J., Qian, Y., Chen, V., Guan, M. X., and Shen, B. (1999) J. Biol. Chem.
274, 17893–1790031. Tsubouchi, H., and Ogawa, H. (2000) Mol. Biol. Cell 11, 2221–223332. Moreau, S., Morgan, E. A., and Symington, L. S. (2001) Genetics 159,
1423–143333. Schmutte, C., Sadoff, M. M., Shim, K. S., Acharya, S., and Fishel, R. (2001)
J. Biol. Chem. 276, 33011–3301834. Lewis, L. K., Karthikeyan, G., Westmoreland, J. W., and Resnick, M. A. (2002)
Genetics 160, 49–6235. Tishkoff, D. X., Boerger, A. L., Bertrand, P., Filosi, N., Gaida, G. M., Kane,
M. F., and Kolodner, R. D. (1997) Proc. Natl. Acad. Sci. U. S. A. 94,7487–7492
36. Tishkoff, D. X., Amin, N. S., Viars, C. S., Arden, K. C., and Kolodner, R. D.(1998) Cancer Res. 58, 5027–5031
37. Orren, D. K., Brosh, R. M., Jr., Nehlin, J. O., Machwe, A., Gray, M. D., andBohr, V. A. (1999) Nucleic Acids Res. 27, 3557–3566
38. Cooper, M. P., Machwe, A., Orren, D. K., Brosh, R. M., Jr., Ramsden, D., andBohr, V. A. (2000) Genes Dev. 14, 907–912
39. Dignam, J. D., Lebovitz, R. M., and Roeder, R. G. (1983) Nucleic Acids Res. 11,1475–1489
40. Brosh, R. M., Jr., Li, J. L., Kenny, M. K., Karow, J. K., Cooper, M. P.,Kureekattil, R. P., Hickson, I. D., and Bohr, V. A. (2000) J. Biol. Chem. 275,23500–23508
41. Tom, S., Henricksen, L. A., and Bambara, R. A. (2000) J. Biol. Chem. 275,10498–10505
42. Brosh, R. M., Jr., Waheed, J., and Sommers, J. A. (2002) J. Biol. Chem. 277,23236–23245
43. Brosh, R. M., Jr., Orren, D. K., Nehlin, J. O., Ravn, P. H., Kenny, M. K.,Machwe, A., and Bohr, V. A. (1999) J. Biol. Chem. 274, 18341–18350
44. Brosh, R. M., Jr., Karmakar, P., Sommers, J. A., Yang, Q., Wang, X. W.,Spillare, E. A., Harris, C. C., and Bohr, V. A. (2001) J. Biol. Chem. 276,35093–35102
45. Genschel, J., Bazemore, L. R., and Modrich, P. (2002) J. Biol. Chem. 277,13302–13311
46. Bennett, S. E., Umar, A., Oshima, J., Monnat, R. J., Jr., and Kunkel, T. A.(1997) Cancer Res. 57, 2956–2960
47. Biswas, E. E., Zhu, F. X., and Biswas, S. B. (1997) Biochemistry 36, 5955–596248. Kao, H. I., Henricksen, L. A., Liu, Y., and Bambara, R. A. (2002) J. Biol. Chem.
277, 14379–1438949. Saintigny, Y., Makienko, K., Swanson, C., Emond, M. J., and Monnat, R. J., Jr.
(2002) Mol. Cell. Biol. 22, 6971–697850. Yamagata, K., Kato, J., Shimamoto, A., Goto, M., Furuichi, Y., and Ikeda, H.
(1998) Proc. Natl. Acad. Sci. U. S. A. 95, 8733–873851. Constantinou, A., Tarsounas, M., Karow, J. K., Brosh, R. M., Jr., Bohr, V. A.,
Hickson, I. D., and West, S. C. (2000) EMBO Reports 1, 80–8452. Bennett, S. E., Umar, A., Kodoma, S., Barett, J. C., Monnat, R. J., Jr., and
Kunkel, T. A. (1999) Mol. Biol. Aging 44, 214–22453. Bae, S. H., Bae, K. H., Kim, J. A., and Seo, Y. S. (2001) Nature 412, 456–46154. Maringele, L., and Lydall, D. (2002) Genes Dev. 16, 1919–193355. Wyllie, F. S., Jones, C. J., Skinner, J. W., Haughton, M. F., Wallis, C.,
Wynford-Thomas, D., Faragher, R. G., and Kipling, D. (2000) Nat. Genet.24, 16–17
56. Huang, P., Pryde, F. E., Lester, D., Maddison, R. L., Borts, R. H., Hickson,I. D., and Louis, E. J. (2001) Curr. Biol. 11, 125–129
57. Wu, L., and Hickson, I. D. (2001) Science 292, 229–230
Activation of EXO-1 Cleavage by WRN23496
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from

and Robert M. Brosh, Jr.Sudha Sharma, Joshua A. Sommers, Henry C. Driscoll, Laura Uzdilla, Teresa M. Wilson
Werner Syndrome Protein1 Are Stimulated by an Interaction with the Carboxyl-terminal Region of the
The Exonucleolytic and Endonucleolytic Cleavage Activities of Human Exonuclease
doi: 10.1074/jbc.M212798200 originally published online April 18, 20032003, 278:23487-23496.J. Biol. Chem.
10.1074/jbc.M212798200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/278/26/23487.full.html#ref-list-1
This article cites 57 references, 35 of which can be accessed free at
by guest on June 28, 2016http://w
ww
.jbc.org/D
ownloaded from




















