
The Escherichia coli heat shock response and bacteriophage λ development
11
ELSEVIER FEMS Microbiology Reviews 17 (1995) 159-169 MICROBIOLOGY REVIEWS The Escherichia coli heat shock response and bacteriophage A development Alessandra Polissi *, Laurence Goffin, Costa Georgopoulos D~partement de Bioehimie M~dicale, Centre M~dical Universitaire, Universit$ de Genbve, 1211 Geneva 4, Switzerland Abstract Tile Escherichia coli/bacteriophage A genetic interaction system has been used to uncover the existence of various biological machines. The starting point of all these studies was the isolation and characterization of E. coli mutants that blocked A growth, and the corresponding A compensatory mutations. In this manner, the AN-promoted transcriptional anti-termination machine was discovered composed of the NusA/NusB/NusE/NusG host proteins. In addition, the DnaK and GroEL chaperone machines were discovered composed of DnaK/DnaJ/GrpE and GroES/GroEL heat shock proteins. The individual members of the DnaK and GroEL chaperone machines have been conserved throughout evolution in both function and structure. Their biological roles include a direct involvement in A DNA replication and morphogenesis, the protection of proteins from aggregation, the disaggregation of various protein aggregates, the manipulation of protein structure and function, as well as the autoregulation of the heat shock response. The evolution of A to extensively rely on the status of the heat shock response of E. coli is likely linked to its lytic versus lysogenic choice of lifestyle. The bacteriophage 1"4 gp31 protein has been purified and shown to substitute for many of GroES' ¢o-chaperonin activities. Keywords: GroEL chaperone machine; DnaK chaperone machine; Heat shock response; Bacteriophage A; Bacteriophage T4 gp31 co-chaperonin; Regulation response of heat shock 1. Introduction The studies of bacteriophage A/Escherichia coli genetic interactions were initiated in the early 1970s and resulted in the identification of a variety of bacterial host functions, which turned out to be of fundamental biological importance (reviewed in de- tail in [1,2]). The start point of all these studies was the isola- tion of E. coil mutants that could not propagate bacteriophage A and the characterization of the cot- • Corresponding aathor. Tel.: +41 (22) 702 5510; Fax: +41 (22) 347 3334 responding bacteriophage compensatory mutations. Many E. coli genes required for a specific step in development were thus isolated. The elucidation of the molecular mechanisms of the action of the corre- sponding gene products revealed that, in addition to being necessary for bacteriophage A growth, they also performed functions essential to host physiology and growth. For example, the E. coli NusA/NusB system, which in conjunction with the AN gene product positively controls early A development, has been shown to act at the level of anti-termination of mRNA transcription ([3,4]; see below). The danK, dnaJ and grpE genes (whose gene products form the DnaK 'chaperone machine'; see below) were origi- 0168-6445/95/$29.00 © 1995 Federation of European Microbiological Societies. All fights reserved SSDI 0168-6445(94)00071-9
Transcript of The Escherichia coli heat shock response and bacteriophage λ development

ELSEVIER FEMS Microbiology Reviews 17 (1995) 159-169
MICROBIOLOGY REVIEWS
The Escherichia coli heat shock response and bacteriophage A development
Alessandra Polissi *, Laurence Goffin, Costa Georgopoulos D~partement de Bioehimie M~dicale, Centre M~dical Universitaire, Universit$ de Genbve, 1211 Geneva 4, Switzerland
Abstract
Tile Escherichia coli/bacteriophage A genetic interaction system has been used to uncover the existence of various biological machines. The starting point of all these studies was the isolation and characterization of E. coli mutants that blocked A growth, and the corresponding A compensatory mutations. In this manner, the AN-promoted transcriptional anti-termination machine was discovered composed of the NusA/NusB/NusE/NusG host proteins. In addition, the DnaK and GroEL chaperone machines were discovered composed of DnaK/DnaJ /GrpE and GroES/GroEL heat shock proteins. The individual members of the DnaK and GroEL chaperone machines have been conserved throughout evolution in both function and structure. Their biological roles include a direct involvement in A DNA replication and morphogenesis, the protection of proteins from aggregation, the disaggregation of various protein aggregates, the manipulation of protein structure and function, as well as the autoregulation of the heat shock response. The evolution of A to extensively rely on the status of the heat shock response of E. coli is likely linked to its lytic versus lysogenic choice of lifestyle. The bacteriophage 1"4 gp31 protein has been purified and shown to substitute for many of GroES' ¢o-chaperonin activities.
Keywords: GroEL chaperone machine; DnaK chaperone machine; Heat shock response; Bacteriophage A; Bacteriophage T4 gp31 co-chaperonin; Regulation response of heat shock
1. Introduction
The studies o f bacteriophage A/ Es cher i ch i a coli genetic interactions were initiated in the early 1970s and resulted in the identification of a variety of bacterial host functions, which turned out to be o f fundamental biological importance ( reviewed in de- tail in [1,2]).
The start point of all these studies was the isola- tion o f E. coil mutants that could not propagate bacter iophage A and the characterization o f the cot-
• Corresponding aathor. Tel.: +41 (22) 702 5510; Fax: +41 (22) 347 3334
responding bacteriophage compensatory mutations. Many E. coli genes required for a specific step in development were thus isolated. The elucidation of the molecular mechanisms o f the action o f the corre- sponding gene products revealed that, in addition to being necessary for bacteriophage A growth, they also performed functions essential to host physiology and growth. For example, the E. coli N u s A / N u s B system, which in conjunction with the AN gene product positively controls early A development , has been shown to act at the level o f anti-termination o f m R N A transcription ([3,4]; see below). The danK, d n aJ and grpE genes (whose gene products fo rm the D n a K 'chaperone machine ' ; see below) were origi-
0168-6445/95/$29.00 © 1995 Federation of European Microbiological Societies. All fights reserved SSDI 0168-6445(94)00071-9
160 A. Polissi et a L / FEMS Microbiology Reviews 17 (I 995) 159-169
nally identified as functions essential for A DNA replication [1,2]. The groEL and groES genes (whose gene products form the GroEL chaperone machine; see below) were originaUy shown to be indispens- able for the head morphogenesis of bacteriophage A [1,2]. Much later, it was shown that the DnaK, DnaJ, GrpE, GroEL and GroES are all universally con- served proteins, belonging to the so-called 'heat shock', or 'stress' class of proteins, and play funda- mental and indispensable roles in protein folding, polypeptide transport, protein disaggregation and regulation of the heat shock response [5,6].
2. Transcr ipt ional ant i - terminat ion
Following infection of E. coil by bacteriophage A, the bacteriophage-encoded N protein prevents transcriptional termination at multiple sites (called tR
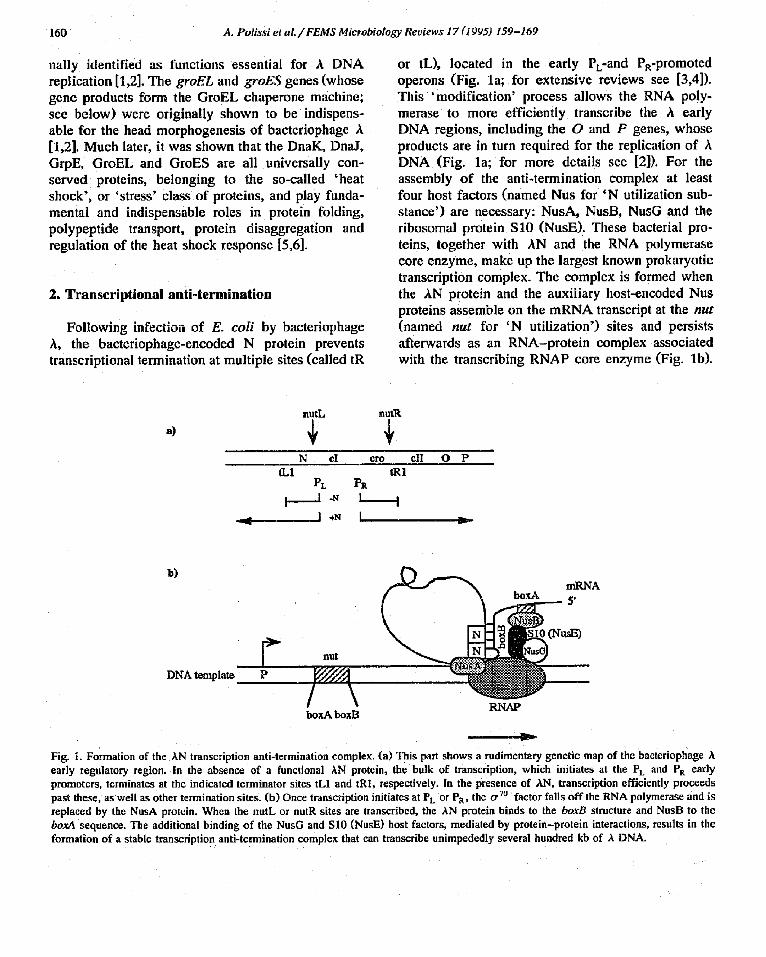
or tL), located in the early PL-and PR-promoted operons (Fig. la; for extensive reviews see [3,4]). This 'modification' process allows the RNA poly- merase to more efficiently transcribe the A early DNA regions, including the O and P genes, whose products are in turn required for the replication of A DNA (Fig. la; for more details see [2]). For the assembly of the anti-termination complex at least four host factors (named Nus for 'N utilization sub- stance') are necessary: NusA, NusB, NusG and the ribosomal protein $10 (NusE). These bacterial pro- teins, together with AN and the RNA polymerase core enzyme, make up the largest known prokaryotic transcription complex. The complex is formed when the AN protein and the auxiliary host-encoded Nus proteins assemble on the mRNA transcript at the nut (named nut for 'N utilization') sites and persists afterwards as an RNA-protein complex associated with the transcribing RNAP core enzyme (Fig. lb).
a)
b)
DNA template
nutL
N tL1
I
r P
nunR
d ero ell O P tR1
PL PR I-N I I I + N I
boxA 5'
)
nut
W///A / \ bor, A boxB
Fig. 1. Formation of the AN transcription anti-termination complex. (a) This part shows a rudimentary genetic map of the bacteriophage A early regulatory region. In the absence of a functional kN protein, the balk of transcription, which initiates at the PL and PR early promoters, terminates at the indicated terminator sites tL1 and tR1, respectively. In the presence of AN, transcription efficiently proceeds past these, as well as other termination sites. (b) Once transcription initiates at PL or Pa, the tr TM factor falls off the RNA polymerase and is replaced by the NusA protein. When the nutL or nutR sites are transcribed, the AN protein binds to the boxB structure and NusB to the boxA sequence. The additional binding of the NusG and $10 (NusE) host factors, mediated by protein-protein interactions, results in the formation of a stable transcription anti-termination complex that can transcribe unimpededly several hundred kb of A DNA.

A, Polissi et al. / FEMS Microbiology Reviews 17 (1995) 159-169 161
This complex is believed to prevent pausing of RNA polymerase at specific transcriptional sites, thus pre- venting premature release of the nascent mRNA transcript, and thus termination of transcription.
The nut sites are comprised of two elements, termed boxA and boxB; boxB is a stem-loop struc- ture'in which most of the nut mutations have been localized [7]. The AN protein binds to the boxB RNA sequence as a dimer, by utilizing an RNA recognition domain whose most important structural element is a cluster of arginines [4]. N-like proteins are very poorly conserved among bacteriophages. For example, the corresponding N proteins from bacteriophage 21 and P22, which are close relatives of A, cannot substitute for the AN protein, because they can only associate with their own corresponding nut site sequences [8]. Interestingly, the Tat protein of the human immunodeficiency virus (HIV) func- tions in a manner reminiscent of that of AN. Like AN, Tat possesses an arginine-rich motif, binds to a specific mRNA hairpin-containing element, called Tar (located proximal to the unique HIV promoter), and prevents premature termination of transcription at that site, thus allowing processive transcript elon- gation throughout the HIV genome [4]. The Tat protein is thus essential for HIV development, and is being used as target for drug therapy.
The isolation of E. coli nus mutants that do not support anti-termination by AN were complemented by the isolation of A bacteriophage compensatory
mutants (termed nin; 'N independent'; see Table 1; [9]) that no longer require AN protein for growth. The convergence of ;hese two type~ of experiments has been invaluable in the elucidation of the details of the mechanism of AN protein action. In vitro work with crude cell-free extracts, as well as purified protein components, has determined the exact role for the Nus factors in the anti-termination process (for a review see [4]). Affinity chromatographic stud- ies coupled with band shift experiments have al- lowed the identification of the following specific protein-protein interactions: NusA with both AN and RNA polymerase, NusG and S10 (NusE) with RNA polymerase core, and NusB with S10 (NusE) [3,4]. These protein-protein and protein-RNA inter- actions form the basis for the assembly and stability of the large anti-termination complex depicted in Fig. lb.
3. The GroEL chaperone machine
Initially identified because of its essentiality for bacteriophage A and T4 head assembly [10,11], the groE operon was subsequently shown to be essential for bacteriophage T5 tail assembly and for E. coli growth under all-growth conditions as well [10]. Further studies established that the two GroE pro- teins, GroES and GroEL, are universally conserved both in structure and function (see below).
Table 1 Plating efficiency o f bacteriophage A on various E. coil mutant hosts
Bacteriophages A Anin AP " AnusB ÷ ,~ groEL ~ hdnaK + T4 T431"
Bacterial host wild-type 1.0 1.0 1.0 1.0 1.0 1.0 1.0 1.0 nusA1 < 10 -6 = 1.0 < 10 - 6 < 10 - 6 < 10 -6 < 10 -6 1.0 1.0
nusB5 < 10 -6 -- 1.0 < 10- ~ 1.0 < 10 -6 < 10 -~ 1.0 1.0 groES619 < 10 -6 < 10-6 < 10- ~ < 10 -6 < 10 -6 < 10-6 1.0 1.0 groEL44 < 1 0 -6 < 1 0 -6 < 1 0 - 6 < 1 0 - 6 1.0 < 1 0 -6 < 1 0 - 6 ~ 1 . 0 dnaK756 < 10 -6 < 10-6 = 1.0 < 10 -6 < 10 -6 1.0 1.0 1.0 dnaJ259 < 10 -6 < 10-6 ~ 1.0 < 10 -6 < 10 -6 < 10-6 1.0 1.0 grpE280 < 10 -6 < 10- ~ = 1.0 < 10 -6 < 10 -6 < 10-6 1.0 1.0
The plating efficiency o f each bacteriophage strain is relative to its efficiency on E. coli wild-type (taken as 1.0). ' Ihe Anin mutant was originally isolated as able to form plaques in the absence of AN protein function. The AP * mutant was isolated as capable of forming a plaque on dunK756 mutant bacteria. The AnusB ~, AgroEL +, and AdnaK ÷, are transducing bacteriophages which carry a copy o f the corresponding wild-type E. coil gene on their chromosome. Their ability to plate is due to complementation of the corresponding mutated function, encoded by the host. The T431" mutant was isolated as capable o f forming a plaque on groEL44 mutant bacteria.

162 A. Polissi et al, /FEMS Microbiology Reviews 17 (1995) 159-169
3.1. Bacteriophage head assembly
Bacteriophage A head assembly and DNA encap- sidation occur in a succession of steps that involve at least ten different gene products [1,12,13]. The for- mation of the 'pre-connector' complex, made up of approximately 12 subunits of gpB [14], is essential for allowing the subsequent correct assembly of the major capsid protein, gpE, into a functional prohead. At this stage, this prohead structure is composed of approximately 12 gpB, 12 gpC, 420 gpE and 200 Nu3 molecules [12]. Additional steps, including pro- tein fusion and cleavage, DNA encapsidation and structure stabilization lead to the formation of a mature head [13]. Finally, a spontaneous interaction between the DNA-filled head and the newly synthe- sized tail gives rise to an infectious A particle. The groE genes were thus named because the first A compensator), mutations, called ~, were mapped to the E gene [11]. However, subsequent studies estab- lished that the primary block exerted by the groE mutations was at the level of the pre-connector as- sembly, whose function is absolutely needed for the further correct assembly of the prohead [14].
An extensive genetic analysis has shown that the groE operon is composed of two genes: a small one, groES (that codes for a M r 10 368 polypeptide) and a larger one, groEL (that codes for a M r 57259 polypeptide). The GroEL protein is composed of two heptameric rings, arranged one on top of the other to form a central cavity, as characterized by electron microscopy [15,16]. The GroES protein is composed of seveo subunits that form a single ring with a hole in the middle [17].
The characterization of the groE gene product requirement for h bacteriophage growth led to the early realization that the biochemical functions of the GroE proteins was a participation in the assembly or oligomerization of other host proteins as well. Subse- quently, Ellis and co-workers characterized a pea chloroplast protein, necessary for the assembly of the oligomeric enzyme RuBisCo. It turned out that this so-called 'RaBisCo binding protein' shares approxi- mately 50% amino acid identity with E. coli's GroEL [18]. Moreover, it was shown that GroEL could replace the RuBi~Co bindit,~g protein function in vivo [19]. From thes:~' results, as well as other experiments carried out in a wide variety of organisms, a new
class of highly conserved proteins of general and essential function emerged and given the name 'molecular chaperones'. Unlike other members ~f the universally conserved heat shock proteins, how- ever, GroES and GroEL are only found in eubaete- ria, chloroplasts and mitochondria. Because of this unique distribution in nature, J. Ellis coined the term 'ehaperonins' to apply exclusively to these proteins [201.
3.2. Chaperonin function
Proteins are thought to be capable of acquiring their proper three-dimensional, functional states by a spontaneous process that depends entirely on their specific amino acid sequence [21]. Although this overall conclusion is correct, the high intracellular concentration of proteins (thought to be approxi- mately 150 mg/ml ) , should favor protein aggrega- tion instead. Hydrophobic interactions, which lead to protein aggregation, are enhanced at higher tempera- tures. As a consequence, in most cells, intracellular protein aggregation should readily occur, especially on nascent polypeptides as they emerge from the ribosome, or in the process of being translocated across biological membranes. In order to avoid such wholesale protein aggregation, all living organisms have evolved the molecular chaperone proteins, whose role is to prevent premature interactions that may lead to aggregation. The GroES/GroEL pro- teins form such a 'chaperone machine" [22], which helps to minimize overall intracellular protein aggre- gation. The biological role of this universally con- served chaperone machine is exemplified by the fact that mutations in either the creES or groEL genes lead to wholesale protein aggregation [23,24].
It turns out that the GroEL protein has the ability to recognize and bin0. ~=J certain denatured proteins, while GroES plays the role of a 'cohort' helping to release G.roEL-bound polypeptides [22]. In vitro studies have shown that GroEL does not bind to native proteins. Rather, it binds to an intermediate structure of the folding process, between the fully extended and the native protein, called 'molten glob- ule' [5]. The molten globule structure exhibits inter- mediate tryptophan fluorescence properties and an intermediate exposure of hydrophobie groups. Elec- tron microscopy shows that the unfolded protein

A. Polissi et aL /FEMS Microbiology Reviews 17 (1995) 159-169 163
binds the GroEL within its central cavity [25,26]. The GroEL central cavity can accommodate a pro- tein as large as M r 100 000. Thus, it is thought that the GroEL-bound polypeptide is isolated from the rest of the proteins and can proceed to fold, unim- peded by other protein folding events in the cell [5].
Because both the strength and quality of GroEL binding to the various polypeptide substrates varies, the requirements for GroEL-bound substrate release also vary. For example, for some bound polypep- tides, the simple addition of ATP or non-hydrolysa- ble ATP causes release, for others, ATP hydrolysis is needed and for still others the combined action of ATP hydrolysis and GroES is essential [5]. GroES' helpful role in substrate release is exerted by these three different, yet not mutually exclusive rneeha- nisms. GroES affects GroEL's ATPase in two ways: on the one hand, it inhibits its overall ATPase activ- ity [5,6,17], while on the other, it increases the cooperativity of ATP hydrolysis by the GroEL sub- units [5,6]. Such cooperative ATP hydrolysis may synchronize the disappearance of hydrophobic sur- faces from GroEL, thus leading to the simultaneous release of the bound substrate.
Langer and co-workers [27] have recently pro- posed a 'chaperone relay' team that handles the maturation of certain polypeptide substrates. Such a chaperone relay action is exemplified in the folding of denatured rhodanese and is graphically depicted in Fig. 2. The DnaK chaperone machine "traps' the unfolded rhodanese upon dilution from a denaturant, and subsequently 'hands it off' to the GroEL chaper- one machine for its final maturation stages (see below). Finally, the GroEL chaperone machine has been shown in vitro to both protect certain substrates from heat denaturation, and to disaggregate such aggregates upon formation [28].
3.3. The bacteriophage T4-encoded gp31 protein is a functional substitute for GroES
a } 5' ~ 3'
D n a J ~ ATP"~ ADP+Pi
b) ~ m -" GrpE
ATP - . ~ GroEI./ES ~ A D P + P i
d'
AlP ADP+Pi
e) monomer - - ~ ~ Oiigomerizatlon
Fig. 2. A role of the DnaK and GroEL chaperone machines in polypeptide folding. Shown are various steps at which chaperones are thought to assist in proper polypeptide maturation. (a) The DnaK and DnaJ chaperones bind to their corresponding preferred polypeptide substrate regions, as they emerge from the ribosome. (b) Following ATP hydrolysis, a very stable DnaK substrate-DnaJ complex forms. (c) The GrpE protein, by associating with the DnaK-substrate-DnaJ complex, destabilizes it so that the bound substrate is more readily released. (d) The polypeptide chain thus released can be trapped by the GroEL chaperone machine. (e) Continuous cycles of substrate binding and release, promoted by GroES and ATP hydrolysis allow the polypeptide to either fold properly or at least be on the proper pathway to correct folding. (O The released monomer can oligomerize with itself or other polypeptides, thus leading to the formation of more complex protein structures (see text for details). The scheme and figure are essentially those originally proposed by l.anger et ai. [27].
It was shown many years ago that some of the original groE mutations blocked bacteriophage T4 head assembly as well [1,10]. T4 compensatory mu- tations mapped in the 31 morphogenetic gene sug- gesting that the E. colt-encoded GroE system inter- acted with the gene M-encoded product, gp31, to ensure the correct polymerization of the T4 head.
When subsequently the groE mutations were divided into either the groES or groEL complementation groups, it became apparent that none of the groES mutations blocked T4 growth. This key observation led to the proposal that the gp31 protein is a func- tional analogue of GroES, thus leading to T4 's ap- parent independence of GroES function [29].

164 A. Polissi et al. / FEMS Microbiology Reuiews 17 (1995) 159-169
Recently, Van der Vies et al. [30] have proved that the gp31 polypeptide of "1"4 indeed encodes for a GroES analogue. This conclusion is based on the following lines of genetic and biochemical evidence: (i) overproduction of gp31 reverts some of the in vivo phenotypes or groES mutations, including bac- teriophage A growth, and colony-forming ability at otherwise non-permissive temperatures; (ii) overpro- duction of gp31 restores to groES mutant bacteria the ability to assemble in vivo the cyanobacterial RuBisCo enzyme; (iii) purified gp31 is an oligomer (most likely an heptamer) that binds to GroEL in the presence of ATP; (iv) purified gp31 inhibits GroEL's ATPase activity to the same extent as does GroES; (v) purified gp31 and GroEL function together to promote the folding of denatured dimeric RuBisCo from Rhodospirillum rubrum, with the same rate and final yield as GroES/GroEL. It is interesting that GroES and gp31 are functional analogues, because they share no obvious amino acid sequence homol- ogy. However, preliminary experiments by Van der Vies (personal communication) suggest that gp31 possesses a segment that corresponds to that of the 'mobile loop" of GroES [31]. In analogy with GroES, this could be the region of gp31 that interacts with GroEL [31]. In support of this conclusion, a limited amino acid sequence homology can be detected be- tween these two mobile loops, and many of the T4 gene 31 mutations, that compensate for the mutated groEL function, map in this region [29].
4. The DnaK chaperone machine
One of the reasons why DnaK, DnaJ and GrpE are referred to as a 'machine' is because they work together to perform a variety of important biochemi- cal functions [22]. For example, temperature-sensi- tive mutations in any of the corresponding genes were originally shown to block A growth at non-per- missive temperatures [1,6,22]. Later it was found that such mutations block even E. coli DNA replication at the non-permissive temperature [1,6]. The precise role of DnaK, DnaJ and GrpE in A DNA replication is the disassembly of a AO-AP-DnaB multiprotein complex at the origin of A replication (see Table 1) [6,32]. Such a synergistic action of DnaK, DnaJ and GrpE is also observed in the reactivation of heat-in-
activated RNA polymerase. In the absence of DnM and GrpE more than 100 molecules of the DnaK chaperone protein are required to reactivate 1 molecule of aggregated RNA polymerase, whereas in the presence of DnaJ or DnaJ and GrpE, progres- sively fewer molecules of DnaK are required [28].
4.1. The DnaK protein
The E. coli M r 67 121 DnaK protein belongs to the universally conserved Hsp70 family of stress proteins [5,6]. The dnaK gene cannot be deleted in most E. coli genetic backgrounds. The rare dnaK null mutants isolated grow very slowly and readily accumulate extragenic suppressors [33]. A gene en- coding a DnaK homologue in E. coli, named hsc, has been recently found. The product of hsc, Hsc66, shows 40% sequence identity with DnaK and other Hsp70 class of proteins. However, unlike DnaK, Hsc66 expression is not induced by elevated temper- atures. The function of Hse66 is still unknown [34].
DnaK possesses an extremely weak ATPase activ- ity, and the ability to bind peptides and unfolded polypeptides in their extended conformation [5,6]. The presence of DnaJ and GrpE stimulates approxi- mately 50-fold the ATPase activity of DnaK [35]. In DnaK, as well as in the Hsp70 homologues, two domains can be distinguished: an amino-terminal domain containing the ATP binding site and a car- boxyl-terminai domain which carries the substrate binding fi,~nction [5,6]. The binding of ATP to DnaK changes the conformation of the protein and this conformational change results in a reduced affinity of the protein for the substrate polypeptide. Recently, it has been shown that the dissociation of the bound peptide is triggered by the binding of ATP, and does not require its hydrolysis [36].
4.2. The DnaJ protein
The DnaJ protein is a constitutively expressed stressed protein of M r 40 973 whose synthesis in- creases after heat shock [6]. The dnaJ gene is always found in the same operon as the dnaK gene, and is not absolutely essential for E. coil viability: dnaJ null mutants grow very slowly up to 42°C, but die at 43°C [6]. As with DnaK, DnaJ homologues have been found in all organisms, ranging from yeast to

A. Polissi et aL / FEMS Microbiology Reviews 17 (1995) 159-16~ 165
plants and humans [37]. All DnaJ-like proteins share a highly conserved 70 amino acid sequence, called the 'J domain', which is proposed to mediate DnaJ's interactions with DnaK (Hsp70).
A homologue of DnaJ has been recently identified in E. colt [38]. Overexpression of this gene, called cbpA (named so because it was originally identified as a curved DNA binding protein), can support both A growth and bacterial growth in a dnaJ null mutant background. These findings suggest a functional rela- tionship between the DnaJ and CbpA proteins of E. colt.
Recent results have shown that DnaJ itself is a major chaperone protein. In fact it binds some dena- tured proteins, like rhodanese and luciferase, better than DnaK does. Unlike DnaK, but like GroEL, DnaJ binds polypeptides in their 'molten globule' conformation and in so doing prevents their aggrega- tion [5]. However, DnaJ binds better to some of its substrates by acting synergistically with DnaK [5,6,27].
4.3. The GrpE protein
The E. colt grpE gene is monoeistronic. Like DnaK and DnaJ, the M r 21 688 GrpE protein is constitutively expressed, and its rate of synthesis is increased following a heat shock [6]. The grpE gene is essential for E. colt viability, since it can not be deleted under all conditions in all wild-types back- ground tested [39]. However, the grpE gene can be deleted in an E. colt background which has been previously 'adapted' to the loss of dnaK by the accumulation of extragenic suppressors [39]. Thus, the sole biological role of the GrpE protein is to provide assistance to the DnaK chaperone protein. The yeast mitochondrial GrpE, whose presence was detected by its ability to bind to the mitochondrial Hsp70 homologue, is likely to be involved in protein translocation into the mitochondrial matrix [40]. GrpE performs its biological function through its influence on DnaK's activities. For example, GrpE's binding to DnaK triggers the release of bound nucleotide [35,36]. In the DnaK-substrate interaction, GrpE has been shown to play varied roles, i.e. it helps the release of rhodanese from the DnaK-rhodanese- DnaJ complex, whereas it "targets' DnaK, so that it can preferentially bind to the DnaJ-AP complex, as opposed to free ,~P protein [41].
4.4. Cellular j~mction~ of the DnaK chaperone ma- chine
The DnaK chaper0~e machine carries out many important biological functions, such as protecting proteins against denaturation, de novo protein fold- ing and protein disaggregati0n. As an example, it has been shown recently that the DnaK chaperone ma- chine is essential for the efficient in vivo repair of thermally denatured lueiferase [42]. In fact, in either dunK, or dnaJ or grpE mutant bacteria, heat-in- activated luciferase cannot be reactivated. DnaJ plays a crucial role in preventing luciferase aggregation by binding to the denatured protein and thus targeting it to DnaK, whereas GriPE is required for the subse- quent dissociation of the DnaK-DnaJ-luciferase complex. Interestingl~¢, the in vitro experiments showed that tile DnaK chaperone machine is suffi- cient for the renaturation of heat-inactivated lu- ciferase and that the GroEL chaperone machine is not seemingly required in this refolding reaction. These and other data show that the DnaK and the GroEL chaperone machines can act either separately or, as shown for rhodanese [25], jointly as in the refolding of denatured substrates.
The DnaK chaperone machine is also involved in de novo protein folding. Using cell-free translation extracts, it has been shown that DnaJ binds to either chloramphenicol acety]-transferase or luciferase dur- ing the early phases o f their translation. This binding can lead to proper folding of both proteins to the native state, upon addition of DnaK and GrpE [5,43].
5. Auteregulation of |he heat-shock response
When E. colt cells are exposed to high tempera- tures, the rate of synthesis of the heat-shock genes is transiently induced. Early work from different labo- ratories [44,45] showed that the E. colt heat-shock response is under the transcriptional control of the rpoH gene product. KpoH protein is the ~r 32 tran- scription factor that complexes with the RNA poly- merase core (E), to constitute the Err 32 holoenzyme. The E r r 32 holoenzyme promotes transcription from a set of promoters, containing a specific consensus sequence in their - 3 5 and - 1 0 promoter regions, the so-called heat shock promoters. Following a

166 A. Polissi e ta l . /FEMS Microbiology Reviews 17 (1995) 159-169
temperature shift, the intracellular levels o f 0 .32 increase transiently with a concomitant transient in- crease in the expression o f the heat shock genes. The o" 32 protein is very unstable (half life approx. 1 min) and the level o f active o ~32 in the cell is modula ted by transcriptional as well as post- transcriptional mechan i sms (see below).
The transcription of the rpoH gene is controlled by four promoters: P1, P4, and PS, which are under the control o f the 0-7o transcription factor, and P3 which is controlled by o "E [44,45]. Transcr ipt ion
f rom the P3 promoter increases gradual ly with tem- perature, and at 50°C it is the only rpoH promoter that is functional [44,45].
By using rpoH-lacZ protein fusions and by analysing their rate o f synthesis, it has been shown that the rpoH m R N A contains two cis-acting regula- tory elements, A and B, which are involved in its post- transcriptional control [45]. The A element acts as a posi t ive regulator, since its deletion leads to an increase in 0 -32 expression at low temperatures. E lement A performs its posi t ive action, most likely,
Pl a) r-"
0,7O
P3
oE o~ o,° I~ / I~t I ' - - - - A B C
b} ( Degradation },
o"e 7
C DegradaUon ]1 1[ "rang "ep~snO,
( Inactivation )
~ (DnaK, DnaJ, GrpE,h
0 .32 9
rahSk~t [orta| ~ pression
31
folded unfolded protein protei~
Fig. 3. Proposed steps at which the autoregalation of the heat shock response operates. (a) An abbreviated designation of the positions of the four known promoters of the rpoH gone, Pl , P3, 1)4 and I)5. The arrows point to each start site and direction of transcription. The 0 -70 or 0"E symbols below the start sites indicate the transcription factor that, in association with the RNA polymerase core (E), helps to initiate transcription from these sites. The structural part of the rpoH gone is represented by the boxed area. The hatched areas A, B and C refer to cis-acting regions, either at the RNA or the protein level, that positively or negatively regulate various aspects of rpoH gone expression (see text for details). (b) A unified scheme, depicting various levels at which the DnaK, DnaJ and GrpE heat shock proteins autoregulate their own synthesis. The DnaK/DnaJ/GrpE proteins can either block translation of or 3z, 'offer' the unfinished or finished 0 '32 polypeptide to proteascs for degradation, or sequester 0-32 away from the RNA polymerasc core enzyme (E), (sec text for details). This figure is essentially that shown in the review by Yura et al. [45].

A. Polissi et al. / FEMS Microbiology Reviews 17 (1995) 159-169 167
through a base-pairing with the 16S rRNA. The B 32 element is a negative regulator that represses ¢r
synthesis at low temperatures. Computer analysis predicts the formation of a secondary structure be- tween the A and B cis-acting elements. It is thought that this RNA secondary structure is likely to be stabilized at low temperatures, thus limiting the translation of rpoH mRNA, and to be disrupted at high temperatures, allowing the synthesis of 0 .32 . Using the same r p o H - l a c Z protein fusions, a third element, named C, was identified. Unlike the A and B elements, the C element operates at the level of shutting of f 0 ̀ -32 synthesis, and controlling 0 -32
protein stability [45] (see below). It appears that the 0"32 peptide terminated by the C element sequence triggers the shut-off of translation and causes en- hanced instability of the o -32 mature protein. It is likely that the DnaK chaperone machine "captures' this peptide as it exits f rom the ribosome, thus resulting in a halt o f 0. 32 translation, a n d / o r presen- tation to proteases for degradation.
The interaction between 0-32 and the DnaK chap- erone machine represents an additional mechanism of modulation o f ,7 32 activity in the cell. Originally, it was observed that in dnaK, dnaJ or grpE mutants the heat-shock response was upregulated and that in these mutants the stability of 0 -32 was greatly in- creased (half life approx. 20 rain) [44-46]. Later, in vitro experiments using purified 0.32, DnaK, DnaJ, GrpE and RNA polymerase proteins showed that: (i) the DnaK and DnaJ proteins can both bind separately to 0. 32; (ii) in the presence o f ATP a stable D n a K - DnaJ-0 . 32 complex is formed; (iii) in the presence of ATP and or 7°, DnaK and DnaJ can efficiently compete with R N A polymerase for the binding to 0-32; and (iv) in the presence o f ATP and 0.70, DnaK and DnaJ can 'strip off ' 0.32 bound to RNA poly- merase [46,47]. In this purified system, the GrpE protein neither helped the formation nor interfered with the D n a K - D n a J - o "32 complex [47]. It is not known how DnaK and DnaJ control the stability of o" 32. A plausible explanation is that the formation of D n a K - D n a J - o -32 complex is necessary for the proper 'presentation' of 0.32 ~,, an as yet unidenti- fied protease complex. On the basis of these data, a global model o f regulation for the heat shock re- sponse, proposed by Yura et al. [45], is shown in Fig. 3.
6. Conclusion
In order to prevent premature protein aggregation and to ensure the solubilization o f protein aggre- gates, various universally conserved chaperone ma- chines have evolved. In the ease of E. coli, all members of the two major chaperone machines, GroEL and DnaK are heat shock proteins. The DnaK and GroEL chaperone systems complement each other by preferentially l'~inding to different protein substrates. In some cases a synergistic action o f both chaperone machines is needed in order to ensure the correct folding of certain polypeptides. In addition, the DnaK chaperone machine has evolved to nega- tively autoregulate its own synthesis, as well as that of the rest o f the heat shock proteins. Bacteriophage
has taken advantage of the existence of these two chaperone machines, to ensure that its DNA replica- tion cycle and its morphogenesis are carried out efficiently. This dependence led to the identification of the DnaK and GroEL chaperone machines.
References
[1] Friedman, D.I., Olson, E.R., Tiily, K., Georgopoulos, C., Herskowitz, I. and Banuett, F. (1984) Interactions of baete- riophage A and host macromolecules in the growth of bacte- riophage A. Microbiol. Rev. 48, 299-325.
[2] Friedman, D.I. (1992) Interaction between bacteriophage A and its Esel2erichia coli host. Curr. Biol. 2, 727-738.
[3] Das, A. (1993) Control of transcription termination by RNA-binding proteins. Annu. Rev. Biochem. 62, 893-930.
[4] Greenblatt, J., Nodwell, J.R. and Mason, S.W. (1993) Tran- seriptional antitermination. Nature 364, 401-406.
[5] Hendrick, LP. and Hartl, F.-U. (1993) Molecular chaperone functions of heat-shock proteins. Annu. Rev. Biochem. 62, 349-384.
[6] Georgopoulos, C. and Welch, WJ. (1993) Role of major heat shock proteins as molecular chaperones. Annu. Rev. Cell. Biol. 9, 601-635.
[7] Doelling, J.H. and Franklin, N.C. (1989) Effects of all single base substitutions in the loop of box B on antitermination of transcription by bacteriophage A's N protein. Nucleic Acids Res. I7, 5565-5577.
[8] Lazinski, D., Grzadzielska, E. and Das, A. (1989) Sequence- specific recognition of RNA hairpins by bacteriophage an- titerminators requires a conserved arginine-rich motif. Cell 59, 207-218.
[9] Bunler, B. and Echols, H. (1970) Regulation of bacterio- phage A development by gene N: properties of a mutation that bypasses N control of late protein synthesis. Virology 40, 212-222.

168 A. Polissi etal. /FEMS Microbiology Reviews 17 (1995) 159-169
[10] Georgupoulos, C., Hendrix, R.W., Kaiser, A.D. and Wood, W.B. (1972) Role of the host cell in bacteriophage morpho- genesis: effects of a bacterial mutation on T4 head assembly. Nature New Biol. 239, 38-41.
[11] Georgopoulos, C., Hen&ix, R.W., Casjens, S.R. and Kaiser, A.D. (1973) Host participation in bacteriophage lambda head assembly. J. Mol. Biol. 76, 45-60.
[12] Georgopoulos, C., Tiliy, K. and Casjens, S. (1983) Lambdoid phage head assembly. In: Lambda II (Hendrix, R.W., Roberts, J.W., Sthal, F.W. and Weisberg, R.A., Eds.), pp. 279-304. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
[13] Feiss, M. and Becket, A. (1983) DNA packaging and cutting. In: Lambda II (Hen&ix, R,W., Roberts, J.W., Sthal, F.W. and Weisberg, R.A., Eds.), pp. 305-330. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
[14] Kochan, J., Carrascosca, J. and Murialdo, H. (1984) Bacte- riophage lambda preconneetors. J. Mol. Biol. 174, 433-447
[15] Hendrix, R.W. (1979) Purification and properties of GroE, a host protein involved in bacteriophage assembly. J. Mol. Biol. 129, 375-392.
[16] Hohn, T., Hohn, B., Engei, A. and Wurtz, M. (1979) Isola- tion and characterization of the host protein GroE involved in bacteriophage lambda assembly. J. Mol. Biol. 129, 359-373.
[17] Chandrasekhar, G.N., Woolford, C., Hen&ix, R. and Geor- gopoulos, C. (1986) Purification and properties of the groES morphogunetic protein of Escherichia call J. Biol. Chem. 261, 12414-12419.
[18] Hemmingsen, S.M., Woolford, C., van tier Vies, S.M., Tilly, K., Dennis, D.T., Georgopoulos, C., Hendrix, R.W. and Ellis, J.P. (1988) Homologous plant and bacterial proteins chaperone oligomerie protein assembly. Nature 333, 330- 334.
[19] Goloubinoff, P., Christeller, J.T., Gatenby, A.A. and Lorimer, G.H. (1989) Reconstitution of active dimerie ribulose bispho- sphate carboxylase from an unfolded state depends on two chaperonin proteins and Mg-ATP. Nature 342, 884-889.
[20] Ellis, J. (1987) Proteins as molecular chaperones. Nature 328, 378-379.
[21] Anfinsen, C.B. (1973) Principles that govern the folding of protein chains. Science 181, 223-230.
[22] Georgupoulos, C. (1992) The emergence of the chaperone machines. Trends Biochem. Sei. 17, 295-299.
[23] Gragerov, A., Nudler, E., Komissarova, N., Gaitanaris, G.A., Gottesman, M.E. and Nikiforov, V. (1992) Cooperation of GroEL/GroES and DnaK/DnaJ heat shock proteins in pre- venting protein misfolding in Escherichia coli. Proc. Natl. Acad. Sci. USA 89, 10341-10344.
[24] Horwich, A.L., Low, K.B., Fenton, W.A., Hirshfield, I.N. and Furtak, K. (1993) Folding in vivo of bacterial cytoplas- mic proteins: role of GroEL. Cell 74, 909-917.
[25] Langer, T., Pfeifer, G., Martin, J., Baumeister, W. and Harti, F.-U. (1992) Chaperonin-mediated protein folding: GroES binds to one end of the GroEL cylinder, which accommo- dates the protein substrate within its central cavity. EMBO J. 11, 4757-4765.
[26] Braig, K., Simon, M., Furuya, F., Hainfeld, J. and Horwieh, A.L. (1993) A polypeptide bound by the chaperonin GroEL
is localized within a central cavity. Proc. Natl. Acad. Sci. USA 90, 3978-3982.
[27] Langer, T., Lu, C., Eehols, H., Flanagan, J., Hayer, M.K. and Hartl, F.-U. (1992) Successive action of DnaK, DnaJ and GroEL along the pathway of chaperone-mediated protein folding. Nature 356, 683-689.
[28] Ziemienowiez, A., Skowyra, D., Zeilstra-Ryalls, J., Fayet, O., Georgopoulos, C. and Zyliez, M. (1993) Either of the Escherichia coli GroEL/GroES and DnaK/DnaJ/GrpE chaperone machines can reactivate heat-treated RNA poly- merase: different mechanisms for the same activity. J. Biol. Chem. 268, 25425-25431.
[29] Keppel, F., Lipinska, B., Aug, D. and Georgopoulos, C. (1990) Mutational analysis of the phage T4 morphogenetie gone 31, whose product interacts with the E. coli groEL protein. Gone 90, 19-25.
[30] Van der Vies, S.M., Gatenby, A,A. and Georgopoulos, C. (1994) Bacteriophage T4 encodes a co-chaperonin that can substitute for Escherichia coli GroES in protein folding. Nature 368, 654-656.
[31] Landry, S.J., Zeirstra-Ryalls, J., Fayet, O., Georgopoulos, C. and Gierasch, L.M. (1993) Characterization of a functionally important mobile domain of GroES. Nature 364, 255-258,
[32] Zylicz, M., Aug, D., Liberek, K. and Georgopoulos, C. (1989) Initiation of a DNA replication with purified host-and bacteriophage-encoded proteins: the role of the DnaK, DnaJ and GrpE heat shock proteins. EMBO J. 8, 1601-1608.
[33] Bukau, B. and Walker, G.C. (1990) Mutations altering heat shock specific subuuit of RNA polymerase suppress major cellular defects of E. coli mutants lacking the DnaK chaper- one. E M B O J. 9, 4027-4036.
[34] Seaton, B.L. and Viekery, L.E. (1994) A gone encoding a DnaK/hspT0 homolog in Escherichia coll. Proc. Natl. Acad. Sci. USA 91, 2066-2070.
[35] Liberek, K., Marszalek, J., Ang, D., Georgopoulos, C. and Zylicz, M. (1991) The Escfierichia coli'DnaJ and GrpE heat shock proteins jointly stimulate DnaK's ATPase activity. Proc. Natl. Acad. Sci. USA 88, 2874-2878.
[36] PaUeros, D.R., Reid, K.L., Shi, L., Welch, W.J. and Fink, A.L. (1993) ATP-induced protein-HspT0 complex dissocia- tion requires K + but not ATP hydrolysis. Nature 365, 664-666.
[37] Cyr, D.M., Langer, T. and Douglas, M.G. (1994) DnaJ-like proteins: molecular chaperones and specific regulators of Hsp70. Trends Biochem. Sei. 19, 176-181.
[38] Ueguchi, C., Kakeda, M., Yamada, H. and Mizuno, T. (1994) An analogue of the DnaJ molecular chaperone in Escherichia coli. Proe. Natl. Acad. Sci. USA 91, 1054-1058.
[39] Aug, D. and Georgopoulos, C. (1989) The heat shock-regu- lated grpE gone of Escherichia coli is required for bacterial growth at all temperatures, but is dispensable in certain mutant backgrounds. J. Bacteriol. t71, 2748-2755.
[40] Bolliger, L., Deloche, O., Glick, B.S., Georgopoulos, C., Jeng, P., Kronidou, N., Horst, M., Morishima, N. and Sehatz, G. (1994) A mitochondrial homolog of bacterial GrpE inter- acts with mitoehondrial hsp70 and is essential for viability. EMBO J. 13, 1998-2006.

A. Polissi et al. / FEMS Microbiology Reviews 17 (199~) 159-169 169
[41] Osipiuk, J., Georgopoulos, C. and Zylicz~, M. (1993) Initia- tion of A DNA replicalion: the Escherichia coil small heat- shock proteins, DnaJ and GrpE, increase DnaK's affinity for the AP protein. J. Biol. Chem. 268, 4821-4827,
[42] Schr6der, H., Langer, T., Hartl, F.-U. and Bukau, B. (1993) DnaK, DnaJ and GrpE form a cellular chaperone machinery capable of repairing heat-induced protein damage. EMBO .L 12, 4137-4144.
[43] Hendriek, J.P., Langer, T., Davis, T.A., Hartl, F.-U. and Wiedman, M. (1993) Control of folding and membrane translocatinn by binding of the chaperone DnaJ to nascent polypeptides. Prec. Natl. Acad. Sci. USA 90, 10216-10220.
[44] Gross, C.A., Straus, D.B., Erickson, J.W. and Yura, T. (1990) The function and regulation of heat shock proteins in
[451
[46]
[47]
Escherichia coil In: Stress Proteins in Biology and Medicine (Morimoto, R.I., Tissieres, A. and Georgopoulos, C., Eds.), pp. 167-189. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. Yura, T., Nagai, H. and Mori, H. (1993) Regulation of the heat-shock response in bacteria. Annu. Rev. Microbiol. 47, 321-350. Bukau, B. (1993) Regulation of the Eseherichia coli heat- shock response. Mol. Microbioi. 9, 671-680. Liberek, K. and Georgopoulos, C. (1993) Autoregulation of the Escherichia colt heat shock response by the DnaK and DnaJ heat shock proteins. Prec. Natl. Acad. Sci. USA 90, 11019-11023.




















