
The effects of nitrous oxide on the central endogenous pro-opiomelanocortin system in the rat
9
Brain Research, 420 (1987) 57-65 57 Elsevier BRE 12833 The effects of nitrous oxide on the central endogenous pro-opiomelanocortin system in the rat John R. Zuniga, Shirley A. Joseph and Karl M. Knigge Neuroendocrine Unit, University of Rochester, Rochester, NY 14642 (U.S.A.) (Accepted 3 February 1987) Key words: Nitrous oxide; fl-Endorphin; a-Melanocyte stimulating hormone; Adrenocorticotropic hormone; Medial basal hypothalamus; Periaqueductal gray The hypothesis that nitrous oxide stimulates the central pro-opiomelanocortin system in vivo was explored in this study. A concen- tration-dependent stimulation of central pro-opiomelanocortin neuropeptides was demonstrated after exposures to variable concen- trations of nitrous oxide with oxygen. Rats exposed to 60% and 80% nitrous oxide with oxygen demonstrated an elevation offi-endor- phin concentration along the neuraxis involved with analgesia; no similar effect was observed in a-MSH concentration, neither dura- tion of exposure nor acclimation to the enclosed environment altered this stimulation. The discontinuation of nitrous oxide exposure resulted in the diminution of fl-endorphin concentration to pre-exposure levels in 15-30 min. With an ACTH t ~,~antisera, a semi- quantitative increase in opiocortin immunoreactivity after exposures to nitrous oxide was demonstrated. In conclusion, the increase in fl-endorphin concentration and immunoreactive ACTHI_39 staining in the cells of origin, areas of fiber projection and terminal fields suggest that nitrous oxide stimulates the central pro-opiomelanocortin system in vivo in the rat. INTRODUCTION The inhalation of nitrous oxide in concentrations well below those required for anesthesia produce analgesia in humans and animals. Biobehavioral studies have suggested a common mechanism be- tween the analgesia produced by nitrous oxide and the opiates; nitrous oxide analgesia was partially at- tenuated by narcotic antagonists in rats, mice and hu- mans ~ 3,6,2(L30,62. This led to a supposition that nitrous oxide analgesia may in part be mediated by the en- dogenous endorphinergic or enkephalinergic sys- tems. Nitrous oxide increased immunoreactive Met- enkephalin levels in brain and cerebrospinal fluid (CSF) perfusates in rats 46~47, however, failed to alter Met-enkephalin levels in discrete areas of the brain associated with analgesia 39. Plasma and brain fl-en- dorphin levels in rats increased after nitrous oxide exposure6~'63; however, neither CSF fl-endorphin levels in the surgical arena 58 nor plasma levels in un- stressed volunteers increased after nitrous oxide ex- posures 1°. The endogenous pro-opiomelanocortin (POMC) precursor synthesize fl-endorphin, adrenocorticotro- pic hormone (ACTH), and a-melanocyte stimulating hormone (a-MSH) in two distinctly separate neuro- anatomical and physiological systems, one of pitu- itary origin and the other of hypothalamic ori- gin 12"27'43'5°. Recent studies provide evidence that analgesia is produced by the central hypothalamic system 5'35'36'as'49. The arcuate nucleus, of the medial basal hypthalamus (MBH), contain the POMC-pro- ducing neurons of this s y s t e m 4"12"23"26273343"57. Jo- seph et al. 2a have described a second region of opio- cortin cells within the nucleus tractus solitarius. It is believed that these medullary opiocortin neurons may participate in cardiovascular and respiratory ho- meostasis; however, the contribution of these medul- lary opiocortin cells to analgesia remains to be deter- mined. Fiber tracts from the arcuate POMC neurons Correspondence: J.R. Zuniga, Department of Oral and Maxillofacial Surgery, University of North Carolina, School of Dentistry 209H, Chapel Hill, NC 27514, U.S.A. 0006-8993/87/$03.50 © 1987 Elsevier Science Publishers B.V. (Biomedical Division)
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of The effects of nitrous oxide on the central endogenous pro-opiomelanocortin system in the rat

Brain Research, 420 (1987) 57-65 57 Elsevier
BRE 12833
The effects of nitrous oxide on the central endogenous pro-opiomelanocortin system in the rat
John R. Zuniga, Shirley A. Joseph and Karl M. Knigge Neuroendocrine Unit, University of Rochester, Rochester, NY 14642 (U.S.A.)
(Accepted 3 February 1987)
Key words: Nitrous oxide; fl-Endorphin; a-Melanocyte stimulating hormone; Adrenocorticotropic hormone; Medial basal hypothalamus; Periaqueductal gray
The hypothesis that nitrous oxide stimulates the central pro-opiomelanocortin system in vivo was explored in this study. A concen- tration-dependent stimulation of central pro-opiomelanocortin neuropeptides was demonstrated after exposures to variable concen- trations of nitrous oxide with oxygen. Rats exposed to 60% and 80% nitrous oxide with oxygen demonstrated an elevation offi-endor- phin concentration along the neuraxis involved with analgesia; no similar effect was observed in a-MSH concentration, neither dura- tion of exposure nor acclimation to the enclosed environment altered this stimulation. The discontinuation of nitrous oxide exposure resulted in the diminution of fl-endorphin concentration to pre-exposure levels in 15-30 min. With an ACTH t ~,~ antisera, a semi- quantitative increase in opiocortin immunoreactivity after exposures to nitrous oxide was demonstrated. In conclusion, the increase in fl-endorphin concentration and immunoreactive ACTHI_39 staining in the cells of origin, areas of fiber projection and terminal fields suggest that nitrous oxide stimulates the central pro-opiomelanocortin system in vivo in the rat.
INTRODUCTION
The inhalat ion of nitrous oxide in concentrat ions
well below those required for anesthesia produce
analgesia in humans and animals. Biobehavioral
studies have suggested a common mechanism be-
tween the analgesia produced by nitrous oxide and
the opiates; nitrous oxide analgesia was part ial ly at-
tenuated by narcotic antagonists in rats, mice and hu-
mans ~ 3,6,2(L30,62. This led to a supposi t ion that nitrous
oxide analgesia may in part be media ted by the en-
dogenous endorphinergic or enkephal inergic sys-
tems. Nitrous oxide increased immunoreact ive Met-
enkephalin levels in brain and cerebrospinal fluid
(CSF) perfusates in rats 46~47, however , failed to alter
Met-enkephal in levels in discrete areas of the brain
associated with analgesia 39. Plasma and brain fl-en-
dorphin levels in rats increased after nitrous oxide
exposure6~'63; however, nei ther CSF f l-endorphin
levels in the surgical arena 58 nor p lasma levels in un-
stressed volunteers increased after nitrous oxide ex-
posures 1°.
The endogenous pro-opiomelanocor t in (POMC)
precursor synthesize f l -endorphin, adrenocor t icot ro-
pic hormone ( A C T H ) , and a -melanocyte st imulating
hormone ( a -MSH) in two distinctly separate neuro-
anatomical and physiological systems, one of pitu-
itary origin and the other of hypothalamic ori-
gin 12"27'43'5°. Recent studies provide evidence that
analgesia is produced by the central hypothalamic system 5'35'36'as'49. The arcuate nucleus, of the medial
basal hypthalamus (MBH), contain the POMC-pro- ducing neurons of this system 4"12"23"26273343"57. Jo-
seph et al. 2a have described a second region of opio-
cortin cells within the nucleus tractus solitarius. It is
believed that these medul lary opiocort in neurons
may part icipate in cardiovascular and respiratory ho-
meostasis; however, the contr ibution of these medul-
lary opiocort in cells to analgesia remains to be deter-
mined. Fiber tracts from the arcuate POMC neurons
Correspondence: J.R. Zuniga, Department of Oral and Maxillofacial Surgery, University of North Carolina, School of Dentistry 209H, Chapel Hill, NC 27514, U.S.A.
0006-8993/87/$03.50 © 1987 Elsevier Science Publishers B.V. (Biomedical Division)

58
descend through the diencephalon to terminal fields in the periaqueductal gray (PAG) of the midbrain. Here, these endorphinergic fibers influence sermon- ergic and catecholaminergic nuclei which in turn in- hibit transmission of nociceptive afferents ascending in the spinal cord 5"11"15"22"32"45'61. The hypothesis that
nitrous oxide stimulates the central endogenous POMC system along the neuraxis involved with anal-
gesia was explored in this study.
MATERIALS AND METHODS
Adult male Sprague-Dawley rats were housed in temperature- and light-controlled rooms and fed ad libitum. Experiments were conducted between 10.00
and 14:00 h EST to avoid intertrial variability sec- ondary to diurnal influences on POMC neuropep- tides 13.
Fifty-four rats weighing between 300 and 400 g
were placed into six independent groups. Concentra- tions of 15%, 30%, 60% and 80% nitrous oxide bal- anced with oxygen at a 10 liters per minute flow rate
was delivered into a 20 liter sealed chamber housing groups of 3 rats for 60 rain beyond T99 (time required for 99% confidence that the desired concentration
has been reached: 9.21 rain21). Balanced concentra- tions of nitrous oxide and oxygen were delivered into a sealed anesthetic chamber via a Narkomed (North American Drager) anesthesia delivery system. Gas
flow rates were determined and maintained by a guarded rotameter and mechanic volumetric accura-
cy of delivered concentrations were determined by infrared spectrometer. An open circuit system deliv- ery of gases was established through a low flow ex-
haust outlet exiting from the sealed chamber to: (1) prevent from positive pressure atmosphere; (2) pre- vent from rebreathing; (3) maintain stable tempera- ture and gas mixtures; and (4) eliminate waste gases.
A 'naive' group (rats not exposed in the chamber) and an 'air ' group (rats exposed to a 10 liters per mi- nute flow of air in the chamber for 60 rain beyond T99) served as controls. Animals were decapitated imme- diately after exposure, their brains removed, divided into forebrain and brainstem and frozen on dry-ice. Under a microscope the MBH including the median eminence (ME) was removed. The MBH was bound by the posterior margin of the optic chiasm, the anterior margin of the mamillary bodies, 4 mm on
either side of the midline and 3 mm dorsally. The re- mainder of the diencephalon was removed up to the lateral ventricle; this block was bound by the optic chiasm, the mamillary bodies and 1 mm medial to tile inferior temporal gyri. The diencephalon block in- cluded the thalamus, fiber tracts of the dorsal longi- tudinal fasciculus of Schutz (DLF) and the hypotha- lamic proper minus the MBH and ME. The midbram
was serially sectioned at 300 um in a cryostat at -20 °C with a special adapter for thick sectioning. Sections were transferred to a trozen slide main- tained at --20 °C. Using a 14-gauge stainless steel cannula the PAG region was ~punched-out" as
described by Palkovits 42. This tissue, the MBH sam- ple and the remaining block of diencephalon were
homogenized separately in ice-cold, acidified ace- tone. A 101) gl aliquot of homogenate was analyzed
for protein using the modified Lowry technique 3a. The remaining homogenate was centrifuged at 12,000 g for 3(I rain. Predetermined atiquots of this su-
pernatant were dried under nitrogen and stored at -20 °C. Samples were reconstituted and fl-endorphin and a-MSH content determined in a late addition radioimmunoassay 17
Radioimmunoassay Tracer for fl-endorphin was prepared by a modifi-
cation of chloramine-T method of Hunter and Greenwood t~j and purified on an anionic ion-ex-
change QAE Sephadex column (QAE Sephadex, Q-25-120) eluted 0.1 M borate, 1% gelatin buffer (pH 9.2). Serial dilutions of synthetic camel fl-endor~
phin (4-4000 pg) in RIA buffer (0.05 M phosphate buf- fer with saline and 0.05% BSA at pH 7.6) were used for the standards. Antisera generated against camel fl-endorphin was used at an initial dilution of l:9000 and a final dilution of 1:45,000. Following a 24 h incu- bation at 4 °C, 100/~1 of tracer (10,000 cpm) was added and incubated for 48 h. An immunoabsorbant, Pansorbin (Calbiochem Chemicals, LaJolla, CA), was added and incubated for 24 h. Antibody-bound tracer was separated from unbound tracer by centri- fugation for 45 min at 4500 g. The supernatant was aspirated and the pellet counted in a Micromedic 4/200 gamma counter. Tracer for a-MSH was pre- pared similar to the fl-endorphin iodination with the exception that purification was performed on a G-10 Sephadex column eluted with 0.1)5 M PBS, 1% gela-
tin, 0.05% Triton X-100 (pH 7.4). Serial dilutions of
synthetic a-MSH (4000-8000 pg) in buffer were used for standard curves. Antisera generated against syn- thetic a-MSH was used at a final dilution of 1:20,000.
The polyclonal antisera for camel fl-endorphin (cBE-4-D) was generated in a rabbit. Cross-reactivi-
ty studies indicate that the antibody is directed against the mid-region of amino acid sequence of fl- endorphin; it does not bind Met-enkephalin, ACTH,
a-MSH, vasopressin, oxytocin and substance P, and cross-reacts in equimolar ratios with fl-lipotropin and
the 31 kDa precursor 44. At a final dilution of 1:45,000
the sensitivity of the assay was 8 pg, the intra-assay co- efficient of variance was 2.1% and the interassay coef- ficient of variance, 5.6%. The polyclonal antisera for a-MSH (a-MSH-97) was generated in a rabbit. Cross-reactivity studies indicate that the antibody
does not bind ACTH, fl-endorphin, fl-MSH, LHRH, TRH, substance P, oxytocin, somatostatin, and Met- enkephalin 2s. At a final dilution of 1:20,000, the sen-
sitivity of the assay was 4 pg.
Immunocytochemistry For anatomic studies the distribution of ACTH l 39
was examined along the arcuate POMC system of control rats and rats exposed to 60% and 80% nitrous oxide. The characterization of the ACTHI_39 anti- body has been described previously23; specificity
tests revealed no cross-reactivity with LHRH, SRIF,
fl-endorphin, a-MSH, fl-MSH and ACTHI_u~. Na- ive, air exposed, 60% and 80% nitrous oxide ex- posed rats were anesthetized with pentobarbitol and perfused intracardially with Bouins fixative (n = 3
per group). Following perfusion the brains were re- moved and postfixed overnight at 4 °C in the same fixative. The brains were identified by notching, blocked in frontal planes, and 50 #m sections serially cut on an Oxford vibratome. Sections from the region of the optic chiasm were made from all 4 groups and transferred to one well. Similarly, sections were
made from the retrochiasmatic areas, median emi- nence, supramamillary nuclei, and PAG. Repre- sentative control and experimental sections were pooled into the same well and incubated with ACTHj_39 antisera at a dilution of 1:8000 for 48 h. The presence of ACTH was demonstrated using the ABC technique (Vectastain, Vector Laboratories, Burlingame, CA)'8. Sections were mounted on a glass
59
slide, air-dried, cleared in xylene and coverslipped.
Statistics fl-Endorphin and ct-MSH hormone levels were
analyzed by multiple level one-way analysis of vari- ance (ANOVA) followed by the S tudent -Newman-
Keuls multiple comparisons test. The two-tailed Stu- dent's t-test was used for comparison between two means. P values were considered significant when less than 0.05. In all experiments, the control hor-
mone levels (both air and naive) were compared with levels at various nitrous oxide concentrations.
RESULTS
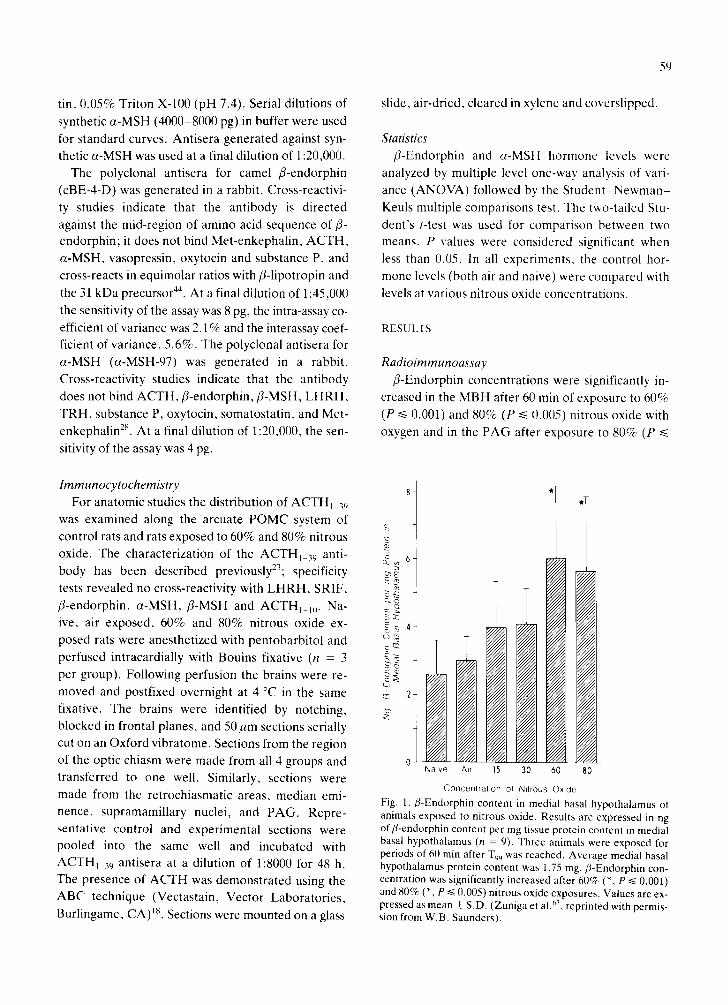
Radioimmunoassay fl-Endorphin concentrations were significantly in-
creased in the MBH after 60 rain of exposure to 60%
(P ~< 0.001) and 80% (P ~< 0.005) nitrous oxide with oxygen and in the PAG after exposure to 80% (P ~<
8 ~
~ 6 - c~ E E ~
O.©
o~ 4
g ~
~ 2 ~ '
Naive A,r 15 30 60 80
Concentration of N,trous Oxide
Fig. 1. ,8-Endorphin content in medial basal hypothalamus of animals exposed to nitrous oxide. Results are expressed in ng of fl-endorphin content per mg tissue protein content in medial basal hypothalamus (n = 9). Three animals were exposed for periods of 60 rain after T99 was reached. Average medial basal hypothalamus protein content was 1.75 rag. f l-Endorphin con- centration was significantly increased after 6 0 ~ (% P ~< 0.001) and 80% (*, P ~< 0.005) nitrous oxide exposures. Values are ex- pressed as mean _+ S.D. (Zuniga et al. ~, reprinted with permis- sion from W.B. Saunders).

60
E
Q.
-.~ 2
Jz:
c)
[] DIENCEP~ E] PERIAGUEI)tIETAL ~Y
Naive Air 30 60 80
Concentration of Nitrous Oxide
Fig. 2. fl-Endorphin content in the remaining diencephalon and periaqueductal gray of animals exposed to nitrous oxide. Re- suits are expressed in ng of fl-endorphin in the diencephalon (n = 9) and pg of fl-endorphin in the periaqueductal gray (n = 9) per mg tissue protein content. The average diencephalon pro- tein content was 2.56 mg and average periaqueductal gray pro- tein content was 0.71 mg. fl-Endorphin concentration was in- creased (*, P ~< 0.05) in periaqueductal gray but not in the dien- cephalon. Values are expressed as mean _+ S.D. (Zuniga et al.63, reprinted with permission from W.B. Saunders).
£ Q.
E
Q_
o = C3
"5
5 -
4--
3 "
[ 7 Med,al Basal HypOlhalarnus
[ ] Dlencel~alon
• Peflaqueduclal Grey
I i
± k Naive Atr
£
6O
f
80
Concentration of Nitrous Ox ide
Fig. 3. a-Melanocyte stimulating hormone (a-MSH) in medial basal hypothalamus, remaining diencepbalon and periaque- ductal gray of rats exposed to nitrous oxide, a-MSH concen- tration was increased (*, P ~< 0.01) in medial basal hypothala- mus but not in the diencephalon or periaqueductal gray. Values are expressed as mean _ S.D.
Cb Q_
C~
g (O
E~
[ ] N.,.,,
Naive Atr 60
Concentration of Ni t rous Ox ide
A
6 o q_
E 5
Q.
E 4
LD :Z: 3 09
2
E ] ~at.,
] '~c:hr, m'{:
Nawe A~r 6O
B
80
C o n c e n t r a t i o n of N i t rous C)xlde
Fig. 4. fl-Endorphin (A) and a-MSH (B) concentrations in the medial basal hypothalamus of naive rats (n = 12) and accli- mated rats (n = 12). Results are expressed in ng of a-MSH con- tent and ng of fl-endorphin content per mg tissue protein con- tent. In both acclimated and naive animals 60% and 80% ni- trous oxide increased fl-endorphin and 80% nitrous oxide in- creased a-MSH in the MBH compared to control. No statistical difference was demonstrated in the responses between accli- mated and naive rats.
0.05) nitrous oxide compared to both controls (Figs.
1 and 2). Nitrous oxide (80%) had a significant effect
on ct-MSH concentration in the MBH but not the PAG (Fig. 3). To determine if the increase in central
POMC neuropeptides in gas-exposed rats was a re-
sponse to the stress of enclosure, animals were accli-
mated to the chamber before exposure. Twelve rats
were exposed to air in the chamber for 15 min daily
for 10 days: 12 rats served as non-acclimated 'naive"
controls. On the eleventh day, acclimated and naive
rats were: (1) immediately sacrificed; (2) exposed to
air for 60 rain: or (3) exposed to 60% and 80% ni-
trous oxide for 60 min. In both acclimated and naive
animals 60% and 80% nitrous oxide increased fl-en-
dorphin and 80% nitrous oxide increased a-MSH in
the' MBH, with no statistical difference in the re-
sponse of acclimated and naive rats (Fig. 4A and B).
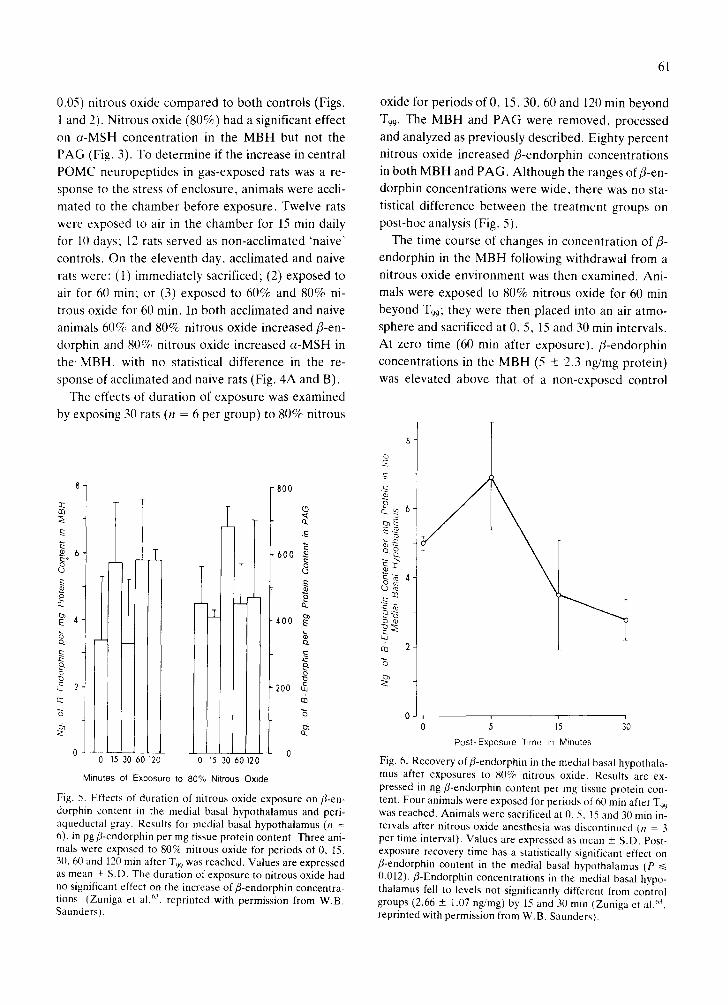
The effects of duration of exposure was examined
by exposing 30 rats (n = 6 per group) to 80% nitrous
8
Q3
6
£ CL
~4
CL
2
2
1
800
0 0- 15 30 60 120 0 15 30 60 12
Minutes of Exposure to 80% Nitrous Oxide
600
400
200
(3
a .
£
cl.
Fig. 5. Effects of duration of nitrous oxide exposure on fl-en- dorphin content in the medial basal hypothalamus and peri- aqueductal gray. Results for medial basal hypothalamus (n = 6), in pg fl-endorphin per mg tissue protein content. Three ani- mals were exposed to 80% nitrous oxide for periods of 0, 15, 30, 60 and 120 min after T99 was reached. Values are expressed as mean _+ S.D. The duration of exposure to nitrous oxide had no significant effect on the increase of f l-endorphin concentra- tions (Zuniga et al. 63, reprinted with permission from W.B. Saunders).
61
oxide for periods of 0, 15, 30, 60 and 120 min beyond
T99. The MBH and PAG were removed, processed
and analyzed as previously described. Eighty percent
nitrous oxide increased fl-endorphin concentrations in both MBH and PAG. Although the ranges offl-en-
dorphin concentrations were wide, there was no sta-
tistical difference between the treatment groups on post-hoc analysis (Fig. 5).
The time course of changes in concentration of fi-
endorphin in the MBH following withdrawal from a
nitrous oxide environment was then examined. Ani-
mals were exposed to 80% nitrous oxide for 60 rain
b e y o n d T99; they were then placed into an air atmo- sphere and sacrificed at 0, 5, 15 and 30 rain intervals.
At zero time (60 min after exposure), fl-endorphin
concentrations in the MBH (5 _+ 2.3 ng/mg protein)
was elevated above that of a non-exposed control
_o c) ~ . ~ 6 . ~ E E~
Q Co
t.U
~ 2
0 0 5 15 30
Post*Exposure Time in Minutes
Fig. 6. Recovery off l -endorphin in the medial basal hypothala- mus after exposures to 80% nitrous oxide. Results are ex- pressed in ng fl-endorphin content per mg tissue protein con- tent. Four animals were exposed for periods of 60 rain after T99 was reached. Animals were sacrificed at 0, 5, 15 and 30 rain in- tervals after nitrous oxide anesthesia was discontinued (n = 3 per time interval). Values are expressed as mean +_ S.D. Post- exposure recovery time has a statistically significant effect on fl-endorphin content in the medial basal hypothalamus (P ~< 0.012). f l-Endorphin concentrat ions in the medial basal hypo- thalamus fell to levels not significantly different from control groups (2.66 + 1.07 ng/mg) by 15 and 30 rain (Zuniga et al/'3, reprinted with permission from W.B. Saunders).
62
group (2.5 + 1 ng /mg pro te in ) ; by 15 and 30 min af ter
exposu re , f l - endorph in in the M B H fell to levels not
s ignif icantly d i f ferent f rom con t ro l g roups (Fig. 6).
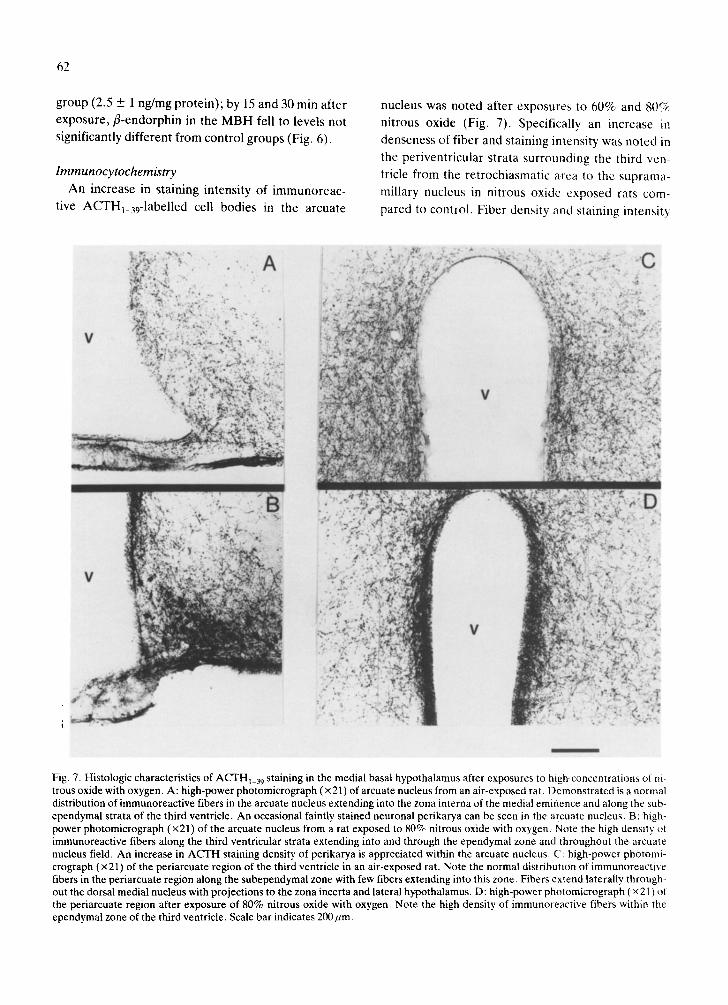
Immunocytochemistry A n increase in s taining intensi ty of i m m u n o r e a c -
t ive ACTHl_39-1abel led cell bod ies in the a rcua te
nucleus was no ted af ter exposures to 60% and 8( )~
ni t rous oxide (Fig. 7). Specif ical ly an increase in
denseness of f iber and s taining intensi ty was no t ed in
the pe r iven t r i cu la r s t rata su r round ing the third ven-
tricle f rom the r e t roch iasmat i c area to the sup rama-
mil lary nucleus in ni t rous oxide exposed rats com-
pared to control . F ibe r densi ty and s taining intensi ty
Fig. 7. Histologic characteristics of ACTHi_39 staining in the medial basal hypothalamus after exposures to high concentrations of ni- trous oxide with oxygen. A: high-power photomicrograph (x 21) of arcuate nucleus from an air-exposed rat. Demonstrated is a normal distribution of immunoreactive fibers in the arcuate nucleus extending into the zona interna of the medial eminence and along the sub- ependymal strata of the third ventricle. An occasional faintly stained neuronal perikarya can be seen in the arcuate nucleus. B: high- power photomicrograph (x21) of the arcuate nucleus from a rat exposed to 80% nitrous oxide with oxygen. Note the high density of immunoreactive fibers along the third ventricular strata extending into and through the ependymal zone and throughout the arcuate nucleus field. An increase in ACTH staining density of perikarya is appreciated within the arcuate nucleus. C: high-power photomi- crograph (× 21) of the periarcuate region of the third ventricle in an air-exposed rat. Note the normal distribution of immunoreactive fibers in the periarcuate region along the subependymal zone with few fibers extending into this zone. Fibers extend laterally through- out the dorsal medial nucleus with projections to the zona incerta and lateral hypothalamus. D: high-power photomicrograph (x 2 L) of the periarcuate region after exposure of 80% nitrous oxide with oxygen. Note the high density of immunoreactive fibers within the ependymal zone of the third ventricle. Scale bar indicates 200 urn.

were increased also in both the DLF and PAG in ni- trous oxide exposed animals.
DISCUSSION
The present study suggests that nitrous oxide has a
stimulatory effect on the central endogenous POMC neuropeptide system. The influence of 60% and 80%
nitrous oxide with oxygen was shown to generate a significant increase in fl-endorphin content in the MBH. Stimulation by nitrous oxide was shown to be
concentration dependent and not dependent upon the duration of exposure or environmental acclima-
tion. Increase offl-endorphin content in the PAG ter- minal fields of the arcuate system suggest that in ad- dition to a potential increase in synthesis by arcuate
neurons an increase in transport also exists. Since fl- endorphin and a-MSH are derivatives of the same POMC precursor in the basal hypothalamus, the dis-
parity between fl-endorphin and a-MSH response after exposures suggest that nitrous oxide stimulates post-translational processing of the POMC molecule through some selective mechanism. With an
ACTHI_39 antisera, a semi-quantitative increase in opiocortin immunoreactivity after exposures to high concentrations of nitrous oxide was demonstrated. Especially prominent was the increase in staining of cell bodies and periventricular fibers.
Upon cessation of nitrous oxide exposure fl-endor- phin concentrations diminished to control levels within 15 min. Evidence has shown that in humans and animals, nitrous oxide levels in brain fall rapidly (10-15 min) following cessation of its application due to its low solubility in blood and low tissue:blood par-
tition coefficient 14"4~,52. The normal clearance of ni-
trous oxide from the brain as implicated above may play a prominent role in the rapid recovery of fl-en- dorphin content in the basal hypothalamus demon- strated in this study.
Numerous studies have suggested similarities be- tween the pharmacological effects of nitrous oxide and the opiates 1-3,6'29,30A6.47,60,62 and support the hy-
pothesis of an instigative stimulation of the endoge-
63
nous opioids by nitrous oxide. In contrast, a number
of studies dispute that any behavioral or physiologic
similarity exists between the opiates and nitrous oxide 1°'31'39'55'56'58'59. Several studies do not support
the hypothesis that nitrous oxide may act by inter- acting with the endogenous POMC peptides since ni-
trous oxide failed to increase fl-endorphin levels in human plasma 1°'58. Plasma opiomelanocortin pep-
tides may reflect peripheral and not central POMC neuronal activity since evidence supports two POMC systems, a peripheral one which is of pituitary origin
and a central system of hypothalamic origin. To date the central POMC system, in contrast to the periph- eral system, appears to be related to a mechanism promoting analgesia 5' 15,35,36,45,49.
Nitrous oxide appears to have a stimulatory effect
on the central endogenous POMC system in vivo in the rat. Most anesthetics are believed to produce nar-
cosis by suppressing cortical and subcortical activity; however, mechanisms whereby anesthetic agents
promote central nervous system (CNS) depression appears to be selective at subanesthetic levels. Anes- thetic agents such as thiopental, halothane and
chloral hydrate depress CNS metabolic and neuronal activity 7"8'21'3~'51'53'54 whereas, metabolic and neu-
ronal activity increase in discrete CNS sites during ni- trous oxide exposure 7,821'25'29'37,3s'5~. Among these
sites are regions closely associated with analgesia. In
this study, elevation of fl-endorphin concentration in the cells of origin, areas of fiber projection and termi- nal fields suggest that nitrous oxide has the potential
to increase synthesis of fl-endorphin in arcuate neu- rons and enhance the transport for release and there-
by may possibly be involved in part in the production of analgesia.
ACKNOWLEDGEMENT
This research was supported in part by the East-
man Dental Center Foundation Teacher Training Grant and the American Association of Oral and Maxillofacial Surgery Research Fellowship Award.

~4
REFERENCES
1 Beneditti, C., Chapman, C.R., Colpitts, Y.H. and Chen. A.C.. Effects of nitrous oxide concentration on event-re- lated potentials during painful tooth stimulation, Anesthe- siology, 56 (1982) 360-364.
2 Berkowitz, B.A., Ngai. S.H. and Finck, A.D., Nitrous oxide 'analgesia': resemblance to opiate action, Science, 194 (1976) 967-968.
3 Berkowitz, B.A., Finck, A.D. and Ngai, S.H., Nitrous oxide analgesia: reversal by naloxone and development of tolerance, J. Pharmacol. Exp. Ther., 203 (1977) 539-547.
4 Bloom, F., Battenberg, E., Rossier, J., Ling, N. and Guile- min. R., Neurons containing fl-endorphin in rat brain exist separately from those containing enkephalin: immunocyto- chemical studies, Proc. Natl. Acad. ScL U.S.A.. 75 (1978) 1591-1595.
5 Carr, K D . and Uysal, S., Evidence of a supraspinal opioid analgesia mechanism engaged by lateral hypothalamic elec- trical stimulation, Brain Research, 335 (1985) 55-62.
6 Chapman, C.R. and Beneditti, C., Nitrous oxide effects on cerebral evoked potential pain; partial reversal with a nar- cotic antagonist, Anesthesiology, 51 (1979) 135-138.
7 Crosby, G., Crane, A.M., Jehle, J. and Sokoloff, L., The local metabolic effects of somatosensory stimulation in the central nervous system of rats given pentobarbital or ni- trous oxide, Anesthesiology, 58 (1983) 38-43.
8 Crosb~, G., Crane, A.M. and Sokoloff, L., A comparison of local rates of glucose utilization in spinal cord and brain in conscious and nitrous oxide- or pentobarbital-treated rats, Anesthesiology, 61 (1984) 434-438.
9 Eger, E.I., II, Nitrous Oxide~N20, Elsevier, New York, 1985, pp. 81-103.
10 Evans, S.F., Stringer, M., Bukht, M.D.G., Thomas, W.A. and Tomlin, S.J., Nitrous oxide inhalation does not influ- ence plasma concentrations of fl-endorphin or Met-enke- phalin like immunoreactivity. Br. J. Anaesthesiol.. 57 (1985) 624-628.
I 1 Fardin, V., Oliveras, J-L. and Besson, J-M., A reinvestiga- tion of the analgesic effects induced by stimulation of the periaqueductal gray matter in the rat. II. Differential characteristics of the analgesia induced by ventral and dor- sal PAG stimulation, Brain Research, 306 (1984) 125-139.
12 Finley, J.C., Lindstr6m, P. and Petrusz, P., Immunocyto- chemical localization of/3-endorphin containing neurons in the rat brain, Neuroendocrinology, 33 (1981) 28-42.
13 Frederickson, R.C.A., Burgis, V. and Edwards, J.D., Hy- peralgesia induced by naloxone follows diurnal rhythm in responsivity to painful stimuli, Science. 198 (1977) 756-758.
14 Frumin, M.J., Salanitre, E, and Rackow, H., Excretion of nitrous oxide in anesthetized man, J. Appl. Physiol., 16 (1961) 720-722.
15 Giesler. G.J., Gerhart, K.D., Yezierski, R.P., Wilcox, T.K. and Willis, W.D., Postsynaptic inhibition of primate spinothalamic neurons by stimulation of nucleus raphe magnus, Brain Research, 204 (1981) 184-188.
16 Grome, J.J. and McCulloch, J., The effects of apomor- phine upon local cerebral glucose utilization in conscious rats and in rats anesthetized with chloral hydrate, J. Neuro- chem., 40 (1983) 569-576.
17 Guillemin, R., Ling, N. and Vargo, T. Radioimmunoassays for a-endorphin and j3-endorphin, Biochem. Biophys. Res. Commun.. 77 (1977) 361-366.
18 Hsu, S.M., Raine, L. and Fanger, Ft., Use of avidm-bio- tin-peroxidase complex (ABC) in immunoperoxidase techniques: a comparison between ABC and unlabeled an- tibody (PAP) procedures, J. Histochem. Cytochem., 29 (1981) 577-580.
19 Hunter, W. and Greenwood. F., Preparation of iodine-131 labeled human growth hormone of high specific activity, Nature (London), 194 (1962) 495-497
20 Hynes, M.D. and Berkowitz, B.A., Nitrous oxide stimula- tion of locomotor activity: evidence for an opiate-like be- havioral effect, J. Pharmacol. Exp. Ther., 209 (1979) 304-309.
21 Ingvar, M., Abdul-Rahman, A. and Sicslo, B.K., Local ce- rebral glucose consumption in the artificially ventilated rat: influence of nitrous oxide analgesia and of phenobarbital anesthesia, Acta Physiol. Scand.. 109 (1980) 177-185.
22 Jensen, T.S. and Yaksh, T.L.. I., Comparison of antinoci- ceptive action of morphine in the periaqueductal gray, me- dial and paramedial medulla in rat, Brain Research, 363 (1986) 99-113.
23 Joseph, S.A., Immunoreactive adrenocorticotropin in rat brain: a neuroanatomical study using antisera generated against synthetic ACTI<I~ ~, Am. J, Anat.. 158 (1980) 533-548.
24 Joseph, S.A., Pilcher, W. and Bennett-Clarke, C., lmmu- nocytochemical Localization of ACTH perikarya in nucleus tractus solitarius: evidence for a second opiocortin neu- ronal system, Neurosci. Lett., 38 (1983) 221-225.
25 Kitahara, L., McAIlister, R. and Taub, A., Identification of central trigeminal nociception and the effects of nitrous oxide, Anesthesiology, 38 (1973) t2-19.
26 Knigge, K.M. and Joseph, S.A., Relationship of the central ACTH in opiocortin system to the median eminence and the pituitary gland in the rat, Cell Tiss. Res., 215 (1981) 333-341.
27 Knigge, K.M. and Joseph, S.A., Anatomy of the opioid- systems of the brain, Can. ]. Neurol Sci., 11 (1984) 14-23.
28 Knigge, K.M., Piekut, D.T. and Joseph, S.A., The alpha- MSH-specific and arcuate pro-opiomelanocortin neuronal systems of the hypothalamus: observations on either con- nectivity and response to colchicine treatment, Psvclto. pharmacol. Bull., submitted.
29 Komatsu, T., Shingu, K., Tomemori, N., Urabe, N. and Mori, K., Nitrous oxide activates the supraspinal pain in- hibition system, Acta AnaesthesioL Scand.. 25 (1981) 519-522.
30 Lawrence, D. and Livingston, P,, Opiate-like analgesic ac- tivity in general anesthetics, Br. J Pharmacol., 73 (198l) 435-442.
31 Levine, J.D., Gordon, N.C. and Fields, H.L., Naloxone fails to antagonize nitrous oxide analgesia for clinical pain, Pain, 13 ( 1985 ) 165-170.
32 Lewis, V.A. and Gebhart, G.F., Evaluation of the peri- aqueductal gray (PAG) as a morphine-specific locus of ac- tion and examination of morphine-induced and stimulation- produced analgesia at coincident PAG loci. Brain Re, search, 124 (1977) 283-303.
33 Liotta, A., Loudes, C., McKelvy, J.F. and Kreiger, D.T., Biosynthesis of precursor corticotropin/endorphin-, corti- cotropin-, ~2-melanotropin-, fl-endorphin-like material by cultured neonatal rat hypothalamic neurons, Proc. Natl. Acad. Sci. U.S.A., 77 (1980) 1880-1884.
34 Markwell, M., Haas, S., Tolbert, N. and Bieber, L., Pro- tein determination in membrane and lipoprotein samples:

manual and automated procedures, Meth. Enzymol., 72 (1981) 296-303.
35 Mayer, D.J. and Liebeskind, J.C., Pain reduction by focal electrical stimulation of the brain: an anatomical and be- havioral analysis, Brain Research, 68 (1974) 73-93.
36 Mayer, D.J. and Price, D.D., Central nervous system mechanisms of analgesia, Pain, 2 (1976) 379-404.
37 Mori, K., Winters, W.D. and Spooner, C.E., Comparison of reticular and multiple unit activity with auditory evoked responses during wtrious stages induced by anesthetic agents, Electroencephalogr. Clin. Neurophysiol., 24 (1968) 242-248.
38 Mori, K., Kawamata, M., Miyajima, S. and Fujita, M., The effects of several anesthetic agents on the neuronal reactive properties of thalamic relay nuclei in the cat, Anesthesiolo- gy, 36 (1972) 550-557.
39 Morris, B. and Livingston, A., Effects of nitrous oxide ex- posure on Met-enkephalin levels in discrete areas of rat brain, Neurosci. Lett., 45 (1984) 11-14.
40 Munson, E.S., Eger, E.I., Tham, M.K. and Embro, W.J., Increase in anesthetic uptake, excretion and blood solubili- ty in man after eating, Anesthesiol. Analgesia, 57 (1978) 224-231.
41 Nelson. G.O., Flow-rate and Volume Measurements. In Controlled Test Atmosphere. Principles and Techniques, Ann Arbor Science Publishers, Ann Arbor, MI, 1982, pp. 21-81.
42 Palkovits, M., Isolated removal of hypothalamic or other brain nuclei of the rat, Brain Research, 59 (1973) 449-450.
43 Pelletier, G., LeClerc, R., Saavedra, J.M., Brownstein, M.J.. Vaudry, H., Ferland, L. and Labrie, F., Distribution of fl-lipotropin (fl-LPH), adrenocorticotropin (ACTH), and cx-melanocyte-stimulating hormone (a-MSH) in the rat brain. I. Origin of the extrahypothalamic fibers, Brain Re- search, 192 (1980) 433-440.
44 Pilcher, W.H., Radioimmunoassay for human and ratfl-en- dorphin: identification and characterization of species-spe- cific antisera. In Studies of Opiocortin and Corticotropin-RF (CRF) Neuropeptide Systems in Brain, Thesis, University of Rochester, 1983, part II, pp. 12-24.
45 Pittman, Q.J., Blume, H.W., Kearney, R.E. and Ranaud, L.P., Influence of midbrain stimulation on the excitability of neurons in the medial hypothalamus of the rat, Brain Re- search, 174 (I979) 39-53.
46 Quock, R.M., Kouchich, F.J. and Tseng, L.-F., Does ni- trous oxide induce release of brain opioid peptides? Phar- macology, 30 (19851 95-99.
47 Quock, R.M., Kouchich. F.J. and Tseng, L.-F., Influence of nitrous oxide upon regional bran levels of metbionine- enkephalin-like immunoreactivity in rats, Brain Res. Bull.. 16 (1986) 321-323.
48 Rhodes, D.L. and Liebeskind, J.C., Analgesia from rostral brain stem stimulation in the rat, Brain Research, 143 (19781 521-532.
49 Rhodes, D.L., Periventricular system lesions and stimula-
65
tion-produced analgesia, Pain, 7 (1979) 51-63. 50 Rossier, J., Vargo, T.M., Minick, S., Ling, N., Bloom,
F.E. and Guillemin, R., Regional dissociation of the fl-en- dorphin and enkephalin contents in the rat brain and pitu- itary, Proc. Natl. Acad. Sci. U.S.A., 74 (1977) 5162-5165.
51 Sakabe, T., Tsutsui, T., Mackawa, T., Ishikawa, T. and Takeshita, H., Local cerebral glucose utilization during hi- trous oxide and pentobarbital anesthesia in rats, Anesthe- siology, 63 (1982) 262-266.
52 Salanitre, E., Rackow, H., Greene, L.T., Klonymus, D. and Epstein, R.M., Uptake and excretion of subanesthetic concentrations of nitrous oxide in man, Anesthesiology. 23 (1962) 814-822.
53 Savaki, H.E., Desban, M., Gtowinski, J. and Besson, M.- J., Local glucose consumption in the rat. I. Effects of halo- thane anesthesia, J. Comp. Neurol., 213 (1983) 36-45.
54 Shimoji, K., Matsuki, M., Schimizu, H.. Maruyama, Y. and Aida, A., Dishabituation of mesencephalic reticular neurons by anesthetics, Anesthesiology, 47 (19771 349-352.
55 Shingu, K., Osawa, M., Fukada, K. and Mori, K., Acute tolerance to analgesic action of nitrous oxide does not de- velop in rats, Anesthesiology, 62 (1985) 5(12-504.
56 Smith, R.A., Wilson, M., Miller, K.W. and Phil, D., Na- loxone has no effect on nitrous oxide anesthesia, Anesthe- siology, 49 (1978) 6-8.
57 Watson, S.J., Brachus, J.D. and Li, C.H., fl-Lipotropin: localization of cells and axons in rat brain by immunocyto- chemistry, Proc. Natl. Acad. Sci. U.S.A., 74 (1977) 5155-5158.
58 Way, W.L., Hosobuchi, Y., Johnson, B.H., Eger, E.I. and Bloom, F.E., Anesthesia does not increase opioid peptides in cerebrospinal fluid in human, Anesthesiology, 60 (19841 43-45.
59 Willer, J.C., Bergeret, S., Gaudy, J.-H. and Dauthier, C., Failure of naloxone to reverse the nitrous oxide-induced depression of a brain stem reflex: an electrophysiologic and double-blind study in humans, Anesthesiology, 63 (1985) 467-472.
60 Williard, K.F., Gillmor, S.T., Stanley, T.H., Meuleman, J.R. and Pace, N.L., The influence of nitrous oxide and no- ciceptive stimuli on rat plasma and brain endogenous con- centrations, Anesthesiology, 57 (1982) A3(12.
61 Yaksh, T.L., Yeung, J.C. and Rudy, T.A., Systematic ex- amination in the rat of brain sites sensitive to the direct ap- plication of morphine: observation and differential effects within the periaqueductal gray, Brain Research, 114 (1976) 83-103.
62 Yang, J.C., Clark, W.C. and Ngai, S.H., Antagonism of ni- trous oxide analgesia by naloxone in man, Anesthesiology, 52 (1980) 414-417.
63 Zuniga, J.R., Knigge, K.M. and Joseph, S.A., Central fl- endorphin release and recovery after exposure to nitrous oxide in rats, J. Oral Maxillofacial Surg., 44 (1986) 714-718.




















