
The effect of baseline metabolic rate on pulmonary O₂ uptake kinetics during very heavy intensity...
7
Respiratory Physiology & Neurobiology 180 (2012) 223–229 Contents lists available at SciVerse ScienceDirect Respiratory Physiology & Neurobiology j our nal ho me p age: www.elsevier.com/locate/resphysiol The effect of baseline metabolic rate on pulmonary O 2 uptake kinetics during very heavy intensity exercise in boys and men Brynmor C. Breese a , Alan R. Barker a , Neil Armstrong a , Andrew M. Jones b , Craig A. Williams a,∗ a Children’s Health and Exercise Research Centre, College of Life and Environmental Sciences, University of Exeter, UK b Sport and Health Sciences, College of Life and Environmental Sciences, University of Exeter, UK a r t i c l e i n f o Article history: Accepted 23 November 2011 Keywords: Phase II time constant ˙ VO 2 slow component Electromyogram Youth a b s t r a c t This study tested the hypothesis that pulmonary ˙ V O 2 kinetics would be slowed during ‘work-to-work’ exercise in adults but not in children. Eight boys (mean age = 12.5 ± 0.5 years) and nine men completed very heavy step transitions initiated from either ‘unloaded’ pedalling (U → VH) or unloaded-to-moderate cycling (i.e. U → M to M → VH). The phase II was significantly (p < 0.05) lengthened in M → VH compared to U → M and U → VH in boys (30 ± 5 vs. 19 ± 5 vs. 21 ± 5 s) and men (49 ± 14 vs. 30 ± 5 vs. 34 ± 8 s). In U → VH, a greater relative ˙ V O 2 slow component temporally coincided with an increased linear iEMG slope in men compared boys ( ˙ V O 2 slow component: 16 ± 3 vs. 11 ± 4%; iEMG slope: 0.19 ± 0.24 vs. −0.06 ± 0.14%, p < 0.05). These results suggest that an age-linked modulation of ˙ V O 2 kinetics might be influenced by alterations in muscle fibre recruitment following the onset of exercise. © 2011 Elsevier B.V. All rights reserved. 1. Introduction Following the onset of step exercise, the arrival of more deoxy- genated blood from the contracting muscles at the pulmonary capillaries signals an exponential rise in pulmonary O 2 uptake ( ˙ V O 2 ) that has been shown to reflect the kinetics of muscle O 2 con- sumption (m ˙ V O 2 ) in humans (Grassi et al., 1996; Krustrup et al., 2009) and is reported to be speeded in children relative to adult or teenage counterparts [see Armstrong and Barker (2009) for a review]. Furthermore, there is an elevated O 2 cost or ‘gain’ during the initial exponential phase that coincides with an attenuated slow rise in ˙ V O 2 above the primary phase amplitude (i.e. the ˙ V O 2 slow component) in young people exercising above the gas exchange threshold (GET) (Armon et al., 1991; Fawkner and Armstrong, 2004; Williams et al., 2001). The physiological factors that mediate these age differences in ˙ V O 2 kinetics are, however, unresolved with few experimental data available. Previous studies using muscle biopsies has revealed that cycling at higher work rates mandates the recruitment of muscle fibre pools with increasing metabolic diversity (i.e. types I, IIa and IIx) in order to meet the requirements for muscle force production (Gollnick et al., 1974; Krustrup et al., 2004; Vollestad and Blom, 1985). For example, type II muscle fibres are purported to have a ∗ Corresponding author at: Children’s Health and Exercise Research Centre, Sport and Health Sciences, University of Exeter, St. Luke’s Campus, Heavitree Road, Exeter EX1 2LU, UK. Tel.: +44 01392 264890; fax: +44 01392 724726. E-mail address: [email protected] (C.A. Williams). reduced microvascular O 2 pressure head, slower ˙ V O 2 kinetics and an increased metabolic cost of force production compared to type I fibres (Behnke et al., 2003; Crow and Kushmerick, 1982; Krustrup et al., 2008; McDonough et al., 2005). The recruitment of type II muscle fibres has therefore been implicated in extending the over- all ˙ V O 2 and gain during step transitions spanning the heavy and very heavy domains compared to moderate exercise [see Jones et al. (2005) for a review]. Conversely, the characteristic ˙ V O 2 pro- file reported in young people during heavy-intensity exercise is similar to conditions in which type I fibre activation is presumably enhanced, for example, following selective glycogen depletion of type II fibres (Carter et al., 2004), during pedalling at slow cadences (Breese et al., 2011; Pringle et al., 2003b), and in adult subjects with a greater proportion of type I fibres (Barstow et al., 1996; Pringle et al., 2003a). However, the possibility that children’s faster phase II ˙ V O 2 kinetics and reduced ˙ V O 2 slow component might be linked to differences in muscle fibre recruitment compared to older counterparts has yet to be examined experimentally. In adults, it has been suggested that a lengthened phase II and increased ˙ V O 2 gain when step exercise transitions are initiated from an elevated baseline work rate (i.e. work-to-work exercise) might reflect the metabolic properties of higher-order (type II) muscle fibres that are revealed on the pulmonary O 2 signal under these circumstances (Brittain et al., 2001; DiMenna et al., 2008, 2010; Wilkerson and Jones, 2006, 2007). This supposition is based on the size principle (Henneman and Mendell, 1981) which posits an orderly recruitment from smaller motor units with a greater muscle fibre oxidative capacity (i.e. type I) to larger motor units (i.e. types IIa and IIx) as the requirement for muscle force production is 1569-9048/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.resp.2011.11.013
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The effect of baseline metabolic rate on pulmonary O₂ uptake kinetics during very heavy intensity...

Tv
Ba
b
a
AA
KPVEY
1
gcts2ortrctWae
api(1
aE
1d
Respiratory Physiology & Neurobiology 180 (2012) 223– 229
Contents lists available at SciVerse ScienceDirect
Respiratory Physiology & Neurobiology
j our nal ho me p age: www.elsev ier .com/ locate / resphys io l
he effect of baseline metabolic rate on pulmonary O2 uptake kinetics duringery heavy intensity exercise in boys and men
rynmor C. Breesea, Alan R. Barkera, Neil Armstronga, Andrew M. Jonesb, Craig A. Williamsa,∗
Children’s Health and Exercise Research Centre, College of Life and Environmental Sciences, University of Exeter, UKSport and Health Sciences, College of Life and Environmental Sciences, University of Exeter, UK
r t i c l e i n f o
rticle history:ccepted 23 November 2011
eywords:
a b s t r a c t
This study tested the hypothesis that pulmonary V̇O2 kinetics would be slowed during ‘work-to-work’exercise in adults but not in children. Eight boys (mean age = 12.5 ± 0.5 years) and nine men completedvery heavy step transitions initiated from either ‘unloaded’ pedalling (U → VH) or unloaded-to-moderate
hase II time constant˙ O2 slow componentlectromyogramouth
cycling (i.e. U → M to M → VH). The phase II � was significantly (p < 0.05) lengthened in M → VH comparedto U → M and U → VH in boys (30 ± 5 vs. 19 ± 5 vs. 21 ± 5 s) and men (49 ± 14 vs. 30 ± 5 vs. 34 ± 8 s). InU → VH, a greater relative V̇O2 slow component temporally coincided with an increased linear iEMG slopein men compared boys (V̇O2 slow component: 16 ± 3 vs. 11 ± 4%; iEMG slope: 0.19 ± 0.24 vs. −0.06 ± 0.14%,p < 0.05). These results suggest that an age-linked modulation of V̇O2 kinetics might be influenced byalterations in muscle fibre recruitment following the onset of exercise.
. Introduction
Following the onset of step exercise, the arrival of more deoxy-enated blood from the contracting muscles at the pulmonaryapillaries signals an exponential rise in pulmonary O2 uptake (V̇O2 )hat has been shown to reflect the kinetics of muscle O2 con-umption (mV̇O2 ) in humans (Grassi et al., 1996; Krustrup et al.,009) and is reported to be speeded in children relative to adultr teenage counterparts [see Armstrong and Barker (2009) for aeview]. Furthermore, there is an elevated O2 cost or ‘gain’ duringhe initial exponential phase that coincides with an attenuated slowise in V̇O2 above the primary phase amplitude (i.e. the V̇O2 slowomponent) in young people exercising above the gas exchangehreshold (GET) (Armon et al., 1991; Fawkner and Armstrong, 2004;
illiams et al., 2001). The physiological factors that mediate thesege differences in V̇O2 kinetics are, however, unresolved with fewxperimental data available.
Previous studies using muscle biopsies has revealed that cyclingt higher work rates mandates the recruitment of muscle fibreools with increasing metabolic diversity (i.e. types I, IIa and IIx)
n order to meet the requirements for muscle force productionGollnick et al., 1974; Krustrup et al., 2004; Vollestad and Blom,985). For example, type II muscle fibres are purported to have a
∗ Corresponding author at: Children’s Health and Exercise Research Centre, Sportnd Health Sciences, University of Exeter, St. Luke’s Campus, Heavitree Road, ExeterX1 2LU, UK. Tel.: +44 01392 264890; fax: +44 01392 724726.
E-mail address: [email protected] (C.A. Williams).
569-9048/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.resp.2011.11.013
© 2011 Elsevier B.V. All rights reserved.
reduced microvascular O2 pressure head, slower V̇O2 kinetics andan increased metabolic cost of force production compared to type Ifibres (Behnke et al., 2003; Crow and Kushmerick, 1982; Krustrupet al., 2008; McDonough et al., 2005). The recruitment of type IImuscle fibres has therefore been implicated in extending the over-all V̇O2 � and gain during step transitions spanning the heavy andvery heavy domains compared to moderate exercise [see Joneset al. (2005) for a review]. Conversely, the characteristic V̇O2 pro-file reported in young people during heavy-intensity exercise issimilar to conditions in which type I fibre activation is presumablyenhanced, for example, following selective glycogen depletion oftype II fibres (Carter et al., 2004), during pedalling at slow cadences(Breese et al., 2011; Pringle et al., 2003b), and in adult subjectswith a greater proportion of type I fibres (Barstow et al., 1996;Pringle et al., 2003a). However, the possibility that children’s fasterphase II V̇O2 kinetics and reduced V̇O2 slow component might belinked to differences in muscle fibre recruitment compared to oldercounterparts has yet to be examined experimentally.
In adults, it has been suggested that a lengthened phase II �and increased V̇O2 gain when step exercise transitions are initiatedfrom an elevated baseline work rate (i.e. work-to-work exercise)might reflect the metabolic properties of higher-order (type II)muscle fibres that are revealed on the pulmonary O2 signal underthese circumstances (Brittain et al., 2001; DiMenna et al., 2008,2010; Wilkerson and Jones, 2006, 2007). This supposition is based
on the size principle (Henneman and Mendell, 1981) which positsan orderly recruitment from smaller motor units with a greatermuscle fibre oxidative capacity (i.e. type I) to larger motor units (i.e.types IIa and IIx) as the requirement for muscle force production is
2 logy & Neurobiology 180 (2012) 223– 229
iith
aiV
iemtprweBolot
wriarpnepboa
2
2
1aved(TbcpapwPwtet
2
ct1
24 B.C. Breese et al. / Respiratory Physio
ncreased. It is therefore conceivable that child–adult differencesn the phase II � and gain might be amplified in the upper stepransition of work-to-work exercise due to the recruitment ofigher-order (type II) muscle fibres.
It has also been proposed, at least in adults (Jones et al., 2011)nd recently in teenagers (Breese et al., 2011), that alterationsn muscle fibre recruitment might be involved in modulating the˙ O2 slow component during exercise above the GET. For example,ncreases in V̇O2 from the 3rd to 6th minute of constant work ratexercise has been shown to temporally coincide with markers ofuscle activity (Endo et al., 2007; Saunders et al., 2000) and addi-
ional muscle fibre recruitment (Krustrup et al., 2004). It is thereforeertinent that an increased primary V̇O2 amplitude and subsequenteduction in the V̇O2 slow component has been reported to coincideith reciprocal changes in the measured profile of the integrated
lectromyogram (iEMG) in quadriceps muscle (Bailey et al., 2009;urnley et al., 2002; cf. Scheuermann et al., 2001). However, tour knowledge, no previous study has quantified whether simi-ar alterations in motor unit recruitment might contribute to thebserved differences in the primary and slow component ampli-udes between adults and children.
The primary purpose of this study was to therefore utilise theork-to-work exercise model in combination with surface iEMG
ecordings in order to explore the influence of putative alterationsn muscle fibre recruitment on V̇O2 kinetics during exercise in boysnd men. It was hypothesised that increasing the baseline workate prior to very heavy-intensity transitions would lengthen thehase II V̇O2 � and increase the total V̇O2 gain in men, but wouldot alter the same V̇O2 kinetic parameters in boys. We also hypoth-sised that unloaded-to-very heavy transitions would increase therimary V̇O2 gain and reduce the relative V̇O2 slow component inoys compared to men, and this will coincide with a lower ratef change in iEMG activity over time in the boys compared to thedults.
. Methods
.1. Participants
Eight boys (mean ± SD age 12.5 ± 0.5 years, stature.49 ± 0.06 m, body mass 38.9 ± 4.1 kg) and nine men (mean ± SDge 26.0 ± 2.9 years, stature 1.77 ± 0.09 m, body mass 72.1 ± 8.6 kg)olunteered to participate in this study. Somatic maturity wasstimated in the boys using sex-specific regression algorithms toetermine a maturity offset score from age at peak height velocityPHV) using anthropometric measurements (Mirwald et al., 2002).hese estimates yielded a mean offset score from PHV in theoys of −2.3 ± 0.5 years (range −1.6 to −2.9). Written, informedonsent was obtained from each participant and the children’sarent(s)/guardian(s) prior to the commencement of the studyfter verbal and written explanations of the study’s aims, risks, androcedures were given. The procedures employed in this studyere approved by the Institutional Research Ethics Committee.
articipants were instructed to visit the laboratory in a rested andell hydrated state having also abstained from food or caffeine in
he preceding 3 h. In total, boys and men completed nine and fivexercise tests respectively over a 4 week period at approximatelyhe same time of day (±2 h).
.2. Experimental procedures
On the first visit, subjects performed a ramp incremental exer-ise test to voluntary exhaustion for determination of peak V̇O2 andhe gas exchange threshold (GET). After 3-min baseline cycling at5 W, the work rate increased continuously by 15 W min−1 in boys
Fig. 1. Schematic illustration of the experimental protocol including each exercisecondition.
and 30 W min−1 in men to attain a test duration of approximately8–12 min in each individual. Participants were instructed to select apreferred pedal rate of between 70 and 80 rev min−1 and maintainthis cadence throughout the test. The peak V̇O2 was taken as thehighest recorded 10-s stationary average value during the incre-mental test which has been shown recently to reflect a maximumV̇O2 in 93% of young people performing ramp exercise (Barker et al.,2011). The GET was determined using the V-slope method (Beaveret al., 1986) as the first disproportionate increase in CO2 produc-tion (V̇CO2 ) relative to the increase in V̇O2 , and subsequently verifiedfrom visual inspection of the increase in the ventilatory equivalentfor V̇O2 (V̇E/V̇O2 ) with no increase in V̇E/V̇CO2 .
The power outputs that would require 90% of the GET(moderate-intensity exercise) and 60% of the difference (�)between the GET and peak V̇O2 (very heavy-intensity exercise,�60%) were estimated for each participant. Each participant thenreturned to the laboratory to perform one of two step exercise pro-tocols: (1) 3-min of cycling at 15 W, followed by 6-min of veryheavy-intensity cycling (U → VH); and (2) 3-min of cycling at 15 W,followed by 4-min of moderate-intensity cycling (U → M), followedby 6-min of very heavy-intensity cycling (M → VH). A schematicillustration of the experimental protocol is shown in Fig. 1. Eachchild and adult completed four and two repetitions of protocols (1)and (2), respectively. Each protocol was presented to participantsin random order with each laboratory visit separated by ≥48 h.
All exercise tests were performed on an electronically brakedcycle ergometer (Lode Excalibur Sport, Groningen, the Netherlands)with the seat height, handlebar height, and crank length adapted toeach subject and subsequently maintained throughout the testingperiod. Pulmonary gas exchange and ventilation were measuredand displayed breath-by-breath during each exercise trial (Metal-yser 3B Cortex, Biophysik, Leipzig, Germany). Gas fractions ofO2 and CO2 were drawn continuously from a face mask-turbineassembly following calibration with gases of known concentration.Expired volume was measured using a DVT turbine digital trans-ducer which was manually calibrated using a 3-L syringe (HansRudolph, Kansas City, MO) before each test. All calibration pro-cedures were repeated before each experimental test. Heart rate(HR) was recorded every breath during all exercise tests usingshort-range radiotelemetry (Polar S610, Polar Electro Oy, Kempele,Finland).
Neuromuscular activity of the right leg m. vastus lateraliswas measured using surface EMG. The right leg was initially
abraded and cleaned to enhance the muscle electrical signal andgraphite snap electrodes (Unilect 40713, Unomedical, Stonehouse,UK) were adhered to the skin surface in a bipolar arrangement
logy &
(beapcT
ralMsLswTocn1pDdaclt
2
wudsvct
tIe
�
wairwdIiwtfioitaelCtp
B.C. Breese et al. / Respiratory Physio
interelectrode distance: 40 mm) positioned at the midway pointetween the greater trochanter and lateral epicondyle. A groundlectrode was also placed on the m. rectus femoris bisecting thective electrodes with an elastic bandage wrapped around thearticipant’s leg to prevent displacement of the electrodes duringycling. The EMG signal was recorded using a ME3000PB Muscleester (Mega Electronics).
EMG measurements at a sampling frequency of 1000 Hz wereecorded throughout all exercise tests. The bipolar signal wasmplified (amplifier input impedance >1 M�), and data were col-ected online in raw form and stored on a personal computer using
egaWin software (Mega Electronics). The raw EMG data wereubsequently exported as an ASCII file and digitally filtered usingabview 8.2 (National Instruments, Newbury, UK). Initially, theignals were filtered with a 20-Hz high-pass, second-order Butter-orth filter to remove contamination from movement artefacts.
he signal was then rectified and low-pass filtered at a frequencyf 50 Hz to produce a linear envelope. The average iEMG was cal-ulated for 15-s time bins throughout exercise with these valuesormalised to the average measured during 15–180 s of cycling at5 W before the initial step transition. Therefore, all iEMG data areresented as a percentage of the initial ‘unloaded’ cycling phase.ata from repeat trials were averaged, with the �iEMG3-bl responseefined as the difference between the average from 165 to 180 snd the average value recorded during baseline pedalling for eachondition. We also used regression analysis in order to quantify theinear time vs. iEMG slope from the V̇O2 slow component time delayo end-exercise (iEMg6−TD 2) during U → VH and M → VH exercise.
.3. Data analysis procedures
The breath-by-breath V̇O2 data from each step exercise boutere initially examined to exclude errant breaths by removing val-es lying more than four standard deviations from the local meanetermined using a 5-breath rolling average. Filtered V̇O2 data wereubsequently linearly interpolated to provide second-by-secondalues and, for each individual, identical repetitions of each exerciseondition were time aligned to the start of exercise and averagedogether to form a single data set for analysis.
The first 15 s of data after the onset of exercise were deletedo remove the phase I (cardio-dynamic) response, and the phaseI portion of the V̇O2 response was modelled using the non-linearquation:
V̇O2 (t) = �V̇O2 · (1 − e−(t−TD/�) (1)
here �V̇O2 (t), �V̇O2 , TD and � represent the value of V̇O2 at given time (t), the amplitude change in V̇O2 from baseline tots asymptote, time delay and the time constant of the response,espectively. In order to identify an optimal fitting window withhich to estimate parameters of the phase II response, a purpose-esigned software programme developed with LabVIEW (National
nstruments, Newbury, UK) was used. The fitting window wasteratively widened by 1 s intervals, starting from a 60 s fitting
indow and finishing with a fitting window that encompassedhe entire data set (Rossiter et al., 2001). The estimated � for eachtting window was plotted against time to allow the beginningf the V̇O2 slow component to be determined through visualnspection. The onset of the V̇O2 slow component was defined ashe point at which a plateau in the estimated � was followed by
progressive increase in the estimated �. The phase II parameterstimates from Eq. (1) were then resolved by least-squares non-
inear regression (GraphPad Prism, GraphPad Software, San Diego,A). The amplitude of the V̇O2 slow component was calculated ashe difference between the mean of the last 30 s of exercise and thehase II asymptote and expressed in relative terms against the V̇O2Neurobiology 180 (2012) 223– 229 225
at end exercise. To provide information on the ‘overall’ V̇O2 kinetics[mean response time (MRT)], Eq. (1) with TD constrained to 0 s (i.e.no delay term) was fit from the onset to the end of exercise. Forall conditions the functional gain of the primary V̇O2 response wascalculated by dividing the asymptotic primary phase amplitude bythe increment in work rate above baseline (�V̇O2 /�WR). Likewise,the end-exercise V̇O2 gain was calculated using a similar technique.
HR kinetics was also modelled for each condition within a pre-determined fitting window that excluded any slow phase in theHR response using the procedures as described above for V̇O2 data,with the TD parameter in Eq. (1). constrained to t = 0 s (i.e. mono-exponential model with no delay).
2.4. Statistical analysis
Mean differences in V̇O2 , iEMG, and HR parameters wereexamined using a two-way repeated-measures ANOVA withBonferroni adjusted post hoc tests used in order to locate statis-tically significant differences between and within groups. Pearsonproduct-moment correlation coefficients were used to investigaterelationships between the parameters of V̇O2 , HR, and iEMG. Allresults are presented as means ± SD with rejection of the nullhypotheses accepted at an alpha level of 0.05.
3. Results
The subjects’ peak V̇O2 was 1.94 ± 0.14 L min−1 in boys and3.25 ± 0.62 L min−1 in men, with the GET occurring at a similar frac-tion of peak V̇O2 in both groups (boys: 50 ± 3 vs. men: 45 ± 8%).Based on the responses to ramp exercise, the work rates calculatedto require 90% of the GET and �60% were 42 ± 4 and 115 ± 7 W inboys and 101 ± 19 and 230 ± 28 W in men.
3.1. V̇O2 kinetics
Table 1 presents the participants’ V̇O2 kinetic responses for eachcondition. An example V̇O2 response for a typical child and adultparticipant in each exercise condition is shown in Fig. 2. Groupmean V̇O2 responses at the onset of U → VH and M → VH exercisein boys are presented in Fig. 3. Step exercise was initiated from anelevated baseline V̇O2 in M → VH compared to U → M and U → VHexercise in both groups (p < 0.001). The phase II V̇O2 � was slowerin M → VH compared to the other exercise conditions in both boysand men (p < 0.028). However, phase II V̇O2 kinetics were not differ-ent between U → M and U → VH exercise in boys (p = 0.26) or men(p = 0.64). Boys had a faster phase II � compared to men in eachexercise condition (p < 0.007).
The primary V̇O2 gain was reduced in U → VH and M → VH com-pared to U → M in boys (p < 0.04) but there were no significantdifferences for this parameter across exercise conditions in men(p > 0.114). During U → VH exercise, the primary V̇O2 gain tendedto be elevated in boys (p = 0.10) and the relative V̇O2 slow com-ponent was lower compared to men (p = 0.022). The absolute V̇O2slow component amplitude was also reduced in M → VH comparedto U → VH within men (p = 0.033). The total V̇O2 gain increasedfrom U → M to U → VH in men (p = 0.006) with this value furtherextended in M → VH compared to other exercise conditions in adultsubjects (p = 0.003). By contrast, there were no differences in thetotal V̇O2 gain between conditions in boys (p > 0.99).
3.2. HR kinetics
Table 2 presents the mean HR responses to exercise in each con-dition. Group mean HR responses following the onset of U → VH andM → VH in boys are presented in Fig. 3. Baseline HR was increased

226 B.C. Breese et al. / Respiratory Physiology & Neurobiology 180 (2012) 223– 229
Table 1V̇O2 kinetics during U → M, U → VH, and M → VH exercise in boys and men.
Variable ANOVA Boys (n = 8) Men (n = 9)
U → M U → VH M → VH U → M U → VH M → VH
V̇O2 b1 (L min−1) P < 0.001c 0.57 ± 0.05† 0.59 ± 0.07† 0.91 ± 0.08*,#,† 0.83 ± 0.12 0.82 ± 0.12 1.54 ± 0.25*,#
TD1 (s) P = 0.040b 12 ± 3 11 ± 3 9 ± 2* 12 ± 6 12 ± 6 8 ± 6#
�1 (s) P < 0.001a,b 19 ± 5† 21 ± 5† 30 ± 5*,#,† 30 ± 5 34 ± 8 49 ± 14*,#
A1 (L min−1) P < 0.001c 0.34 ± 0.04† 0.99 ± 0.11*,† 0.64 ± 0.07*,#,† 0.70 ± 0.23 1.83 ± 0.28* 1.28 ± 0.27*,#
Gp (mL min−1 W−1) P = 0.008c 10.6 ± 1.1† 9.5 ± 1.0* 9.1 ± 0.8* 8.5 ± 1.4 8.7 ± 0.4 9.9 ± 1.4TD2 (s) NS – 172 ± 26 161 ± 44 – 155 ± 22 175 ± 30A′
2 (L min−1) P = 0.049c – 0.12 ± 0.05† 0.12 ± 0.06† – 0.36 ± 0.12 0.24 ± 0.07#
Rel. A′2 (%) P = 0.048a – 11 ± 4† 15 ± 7 – 16 ± 3 16 ± 6
V̇O2 tot (L min−1) P < 0.001c 0.91 ± 0.08† 1.70 ± 0.12*,† 1.67 ± 0.13*,† 1.53 ± 0.26 3.01 ± 0.43* 3.06 ± 0.44*
Gtot (mL min−1 W−1) P = 0.003c 10.6 ± 1.1† 10.7 ± 1.0 10.7 ± 1.2 8.5 ± 1.4 10.4 ± 0.6* 11.8 ± 1.0*,#
MRT (s) P = 0.046c 31 ± 9† 40 ± 9*† 54 ± 9*,#,† 46 ± 9 66 ± 15* 83 ± 17*,#
Data are presented as mean ± SD. AVOVA, two-way repeated measures ANOVA results.a Significant main effect for age.b Significant main effect for condition.c Significant age × condition interaction.
NS no significant differences found (p > 0.05). Bonferroni adjusted paired comparisons:* Significantly different from U → M condition: p < 0.05.# Significantly different from U → VH condition: p < 0.05.† Significant within-condition age difference: p < 0.05.
V̇O2 b1, mean V̇O2 during baseline pedalling; �1, TD1, A1, time constant, time delay and asymptotic amplitude of the phase II V̇O2 response (derived from Eq. (1)); TD2, A′2, time
delay and amplitude of the V̇O2 slow component; Gp, Gtot, gain (�V̇O2 /�WR) of the primary response and at end exercise; MRT, mean response time.
F ach ec model
ptd(�w(
3
c
TH
Dp
ig. 2. V̇O2 kinetic responses in a representative child (�) and adult (©) subject in eorrecting for the O2 cost of unloaded pedalling. Solid grey lines denote the phase IIine.
rior to the onset of M → VH exercise in boys and men comparedo the other conditions (p < 0.001). The HR � was not significantlyifferent between U → VH and M → VH exercise in either boysp = 0.51) or men (p = 0.63). Relative (%) changes in the phase II V̇O2
between U → VH and M → VH were not significantly correlatedith alterations in HR kinetics in boys (r = 0.55, p = 0.16) or men
r = −0.56, p = 0.19).
.3. iEMG activity
Group mean iEMG responses following the onset of step exer-ise are presented in Fig. 4 for each condition. Baseline pedalling at
able 2R kinetics during U → M, U → VH, and M → VH exercise in boys and men.
Variable ANOVA Boys (n = 8)
U → M U → VH
HRbl (b min−1) P = 0.010a; P < 0.001b 101 ± 8† 101 ± 9†
HR � (s) P < 0.001b 28 ± 13 35 ± 8*
HRtot (b min−1) P = 0.043a; P < 0.001b 123 ± 8† 179 ± 10*
�HR (b min−1) P < 0.001b 22 ± 3 79 ± 8*
ata are presented as mean ± SD. AVOVA, two-way repeated measures AVOVA results.
edalling; HR �, time constant for HR; HRtot, HR at end-exercise; �HR, difference betwee
xercise condition. Data are normalised relative to the end-exercise amplitude afterl fit in each condition. The onset of step exercise is indicated by the vertical dotted
M → VH significantly increased iEMG activity in both groups (boys:192 ± 69 vs. men: 237 ± 64%) compared to other exercise condi-tions (p < 0.022) and reduced the �iEMG3-bl response in M → VHrelative to U → VH in boys (205 ± 87 vs. 316 ± 105%, p = 0.021) andmen (206 ± 79 vs. 335 ± 148%, p = 0.006). There were no betweengroup differences in �iEMG3-bl within each condition (p > 0.05).Fig. 5 presents iEMG responses during U → VH exercise in a rep-resentative child and adult subject. Two-way ANOVA revealed
a significant age × time interaction effect for iEMG activity dur-ing U → VH exercise (p = 0.002). In men, follow-up post hoc testslocated significant increases in iEMG from 60 s to 180 s (406 ± 145vs. 434 ± 148%, p = 0.018) and from 180 s to 360 s (434 ± 148 vs.Men (n = 9)
M → VH U → M U → VH M → VH
122 ± 8*#† 83 ± 13 85 ± 13 110 ± 12*#
40 ± 11* 32 ± 12 39 ± 12 46 ± 16*179 ± 11* 109 ± 14 165 ± 16* 167 ± 15*
58 ± 9*# 26 ± 8 80 ± 9* 57 ± 5*#
See Table 1 for explanation of statistical analyses. HRbl, mean HR during baselinen HRbl and HRtot.
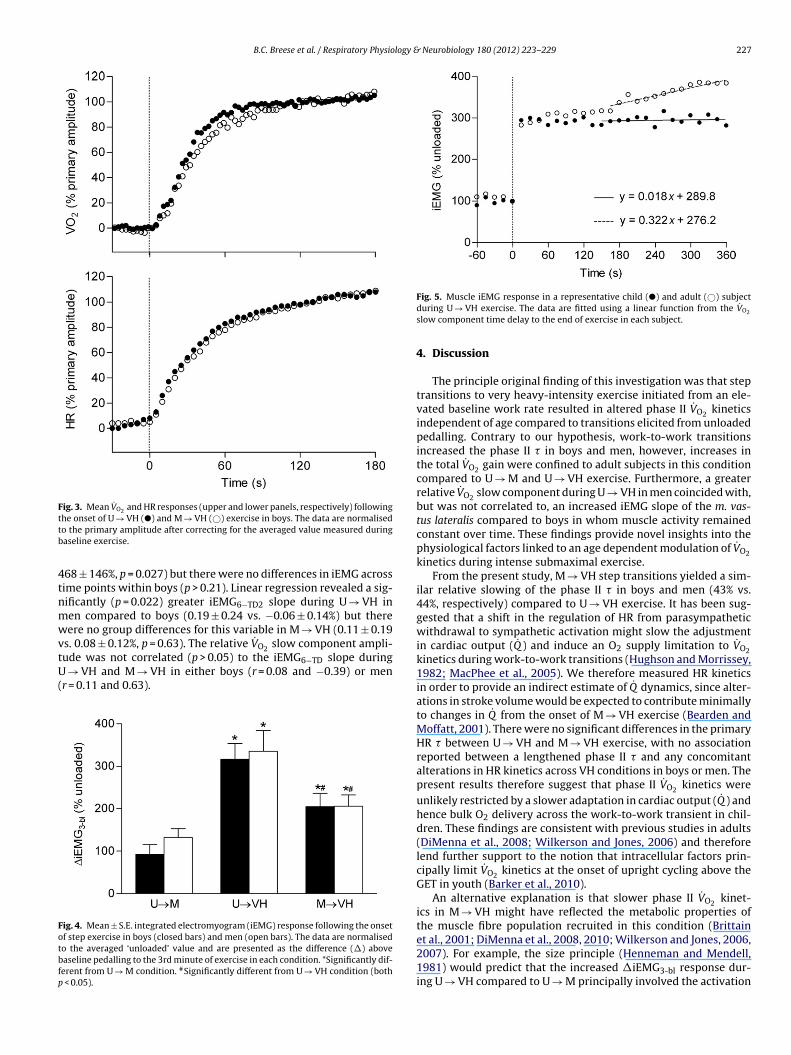
B.C. Breese et al. / Respiratory Physiology & Neurobiology 180 (2012) 223– 229 227
Fig. 3. Mean V̇O2 and HR responses (upper and lower panels, respectively) followingttb
4tnmwvtU(
Fotbfp
he onset of U → VH (�) and M → VH (©) exercise in boys. The data are normalisedo the primary amplitude after correcting for the averaged value measured duringaseline exercise.
68 ± 146%, p = 0.027) but there were no differences in iEMG acrossime points within boys (p > 0.21). Linear regression revealed a sig-ificantly (p = 0.022) greater iEMG6−TD2 slope during U → VH inen compared to boys (0.19 ± 0.24 vs. −0.06 ± 0.14%) but thereere no group differences for this variable in M → VH (0.11 ± 0.19
s. 0.08 ± 0.12%, p = 0.63). The relative V̇O slow component ampli-
2ude was not correlated (p > 0.05) to the iEMG6−TD slope during→ VH and M → VH in either boys (r = 0.08 and −0.39) or menr = 0.11 and 0.63).
ig. 4. Mean ± S.E. integrated electromyogram (iEMG) response following the onsetf step exercise in boys (closed bars) and men (open bars). The data are normalisedo the averaged ‘unloaded’ value and are presented as the difference (�) aboveaseline pedalling to the 3rd minute of exercise in each condition. *Significantly dif-erent from U → M condition. #Significantly different from U → VH condition (both
< 0.05).
Fig. 5. Muscle iEMG response in a representative child (�) and adult (©) subjectduring U → VH exercise. The data are fitted using a linear function from the V̇O2
slow component time delay to the end of exercise in each subject.
4. Discussion
The principle original finding of this investigation was that steptransitions to very heavy-intensity exercise initiated from an ele-vated baseline work rate resulted in altered phase II V̇O2 kineticsindependent of age compared to transitions elicited from unloadedpedalling. Contrary to our hypothesis, work-to-work transitionsincreased the phase II � in boys and men, however, increases inthe total V̇O2 gain were confined to adult subjects in this conditioncompared to U → M and U → VH exercise. Furthermore, a greaterrelative V̇O2 slow component during U → VH in men coincided with,but was not correlated to, an increased iEMG slope of the m. vas-tus lateralis compared to boys in whom muscle activity remainedconstant over time. These findings provide novel insights into thephysiological factors linked to an age dependent modulation of V̇O2kinetics during intense submaximal exercise.
From the present study, M → VH step transitions yielded a sim-ilar relative slowing of the phase II � in boys and men (43% vs.44%, respectively) compared to U → VH exercise. It has been sug-gested that a shift in the regulation of HR from parasympatheticwithdrawal to sympathetic activation might slow the adjustmentin cardiac output (Q̇ ) and induce an O2 supply limitation to V̇O2kinetics during work-to-work transitions (Hughson and Morrissey,1982; MacPhee et al., 2005). We therefore measured HR kineticsin order to provide an indirect estimate of Q̇ dynamics, since alter-ations in stroke volume would be expected to contribute minimallyto changes in Q̇ from the onset of M → VH exercise (Bearden andMoffatt, 2001). There were no significant differences in the primaryHR � between U → VH and M → VH exercise, with no associationreported between a lengthened phase II � and any concomitantalterations in HR kinetics across VH conditions in boys or men. Thepresent results therefore suggest that phase II V̇O2 kinetics wereunlikely restricted by a slower adaptation in cardiac output (Q̇ ) andhence bulk O2 delivery across the work-to-work transient in chil-dren. These findings are consistent with previous studies in adults(DiMenna et al., 2008; Wilkerson and Jones, 2006) and thereforelend further support to the notion that intracellular factors prin-cipally limit V̇O2 kinetics at the onset of upright cycling above theGET in youth (Barker et al., 2010).
An alternative explanation is that slower phase II V̇O2 kinet-ics in M → VH might have reflected the metabolic properties ofthe muscle fibre population recruited in this condition (Brittain
et al., 2001; DiMenna et al., 2008, 2010; Wilkerson and Jones, 2006,2007). For example, the size principle (Henneman and Mendell,1981) would predict that the increased �iEMG3-bl response dur-ing U → VH compared to U → M principally involved the activation
2 logy &
otcetttUa(aFttkeedt
2(VtnapbeidIpahWii(i
fidpitate2iUiseH�sinFob(i
28 B.C. Breese et al. / Respiratory Physio
f a greater proportion of the total muscle fibre pool (includingype I and II fibres) in order to meet the requirements for mus-le force production. Conversely, step transitions initiated from anlevated baseline work rate (i.e. M → VH) may have reduced theotal number of fibres recruited in this condition to those posi-ioned higher in the recruitment hierarchy. This is consistent withhe observed reduced �iEMG3-bl response in M → VH compared to
→ VH exercise. Higher-order (type II) fibres are reported to haven increased O2 cost per unit of force production in small mammalsCrow and Kushmerick, 1982) and in vivo following neural block-de of type I fibres in exercising humans (Krustrup et al., 2008).urthermore, there is a greater fractional O2 extraction requiredo increase V̇O2 in fast-twitch muscle that compromises capillary-o-myocyte O2 flux and would therefore be expected to slow V̇O2inetics (Behnke et al., 2003; McDonough et al., 2005). However, thextent to which work-to-work transitions influenced spatial het-rogeneities in matching O2 delivery-to-O2 utilisation as previouslyemonstrated for cycling exercise (Koga et al., 2007) is unclear fromhe present study.
Consistent with previous studies during cycling (Breese et al.,010; Fawkner and Armstrong, 2004) and treadmill runningWilliams et al., 2001), U → VH transitions yielded faster phase II˙ O2 kinetics in younger compared to older counterparts. Althoughhe mechanisms are unknown, the previous demonstration of aegative correlation between the percentage of type I muscle fibresnd the phase II � in adults (Pringle et al., 2003a) has been inter-reted to suggest that differences in muscle fibre recruitment mighte linked to age differences following the onset of heavy/very heavyxercise (Armstrong and Barker, 2009). From this perspective, it isnteresting to note that dividing a full U → VH transition into twoiscrete step bouts (i.e. U → M and M → VH) yielded faster phase
I V̇O2 kinetics in boys compared to men in each step. This mightredict a longer � in men if muscle fibre pools recruited in U → Mnd M → VH were to contribute to the external power output andence influence the rise in V̇O2 from the onset of U → VH exercise.hilst we were unable to determine muscle fibre recruitment dur-
ng exercise, the demonstration that glycogen content is reducedn single type I and II muscle fibres following the onset of intense80% peak V̇O2 ) cycling in adults (Krustrup et al., 2004) is, at leastn part, consistent with this proposal.
An implicit assumption based on the aforementioned musclebre recruitment profile is that the phase II � would be lengtheneduring exercise above compared to below the GET if the metabolicroperties of higher-order (type II) muscle fibres were involved
n modulating the V̇O2 response earlier into the transition. Con-rary to this notion, similar phase II V̇O2 kinetics between U → Mnd U → VH were found in boys (� = 19 s vs. 21 s). This is consis-ent with previous reports that the phase II � is independent ofxercise intensity in young people (Lai et al., 2008; Williams et al.,001), at least for work rates spanning the moderate-to-very heavy-
ntensity domains. We also reported an invariant phase II � between → M and U → VH in men, although it is conceivable that large
nter-subject variability in the parameter estimates combined withmall sample sizes might have reduced the statistical power nec-ssary to detect meaningful differences (Poole and Jones, 2005).owever, assuming that transitions from unloaded pedalling to60% recruited a large fraction of the total muscle fibre pool in our
ubjects (Sargeant, 1999), it is possible to reconcile V̇O2 responsesn the M → VH condition with a lower relative V̇O2 slow compo-ent amplitude during U → VH exercise in boys compared to men.or example, it has been suggested that the protracted response
˙
f earlier recruited muscle fibres with slower VO2 kinetics mighte involved in creating a V̇O2 slow component of delayed onsetWilkerson and Jones, 2007). It is therefore pertinent that M → VHncreased the total V̇O2 gain (39%) relative to U → M in men, butNeurobiology 180 (2012) 223– 229
there were no differences between conditions for this parameter inboys. This might predict that higher-order muscle fibres, if recruitedfollowing the onset of U → VH exercise, could have extended therise in V̇O2 above the phase II asymptote in men compared to boys.However, in accord with this model, faster V̇O2 kinetics in earlierrecruited fibres would be expected to shorten the V̇O2 slow com-ponent TD in boys compared to men which was not present in theU → VH condition.
It has been demonstrated that the V̇O2 slow component prin-cipally originates within contracting skeletal muscle (Poole et al.,1991) and temporally coincides with an increased PCr cost of exer-cise (Rossiter et al., 2002) and the recruitment of additional musclefibres (Krustrup et al., 2004). It is therefore pertinent that the devel-opment of the V̇O2 slow component was not attended by increasesin iEMG activity during U → VH exercise in boys. This contrastswith previous reports in adults (Burnley et al., 2002; Shinohara andMoritani, 1992) and might be interpreted to suggest that increasesin the metabolic requirements from earlier recruited muscle fibrescould be involved in increasing the O2 cost of the external work ratein young people. For example, the demonstration of a progressivelyincreased V̇O2 gain under conditions in which the total motor unitpool is recruited following the onset of contractions suggests thatthe development of the V̇O2 slow component might not obligateadditional muscle fibre recruitment in animal muscle (Zoladz et al.,2008) and more recently in humans (Vanhatalo et al., 2011).
In contrast, an elevated iEMG6−TD2 slope during U → VH isconsistent with the notion that an increased rate of fatigue devel-opment mandated either an increased firing frequency of motorunits and/or recruitment of previously inactive (higher-order) mus-cle fibres in order to sustain the target work rate in men. In the lattercase, a progressively increasing iEMG would, if reliant upon the acti-vation of less efficient type II fibres (Crow and Kushmerick, 1982;Krustrup et al., 2008), be expected to increase V̇O2 further above theprimary asymptote in adults compared to children as was shownin the present study. However, it is important to consider that theiEMG6−TD2 slope was not correlated to the relative V̇O2 slow compo-nent amplitude in boys or men. Furthermore, there were no groupdifferences in the �iEMG3-bl despite a 9% increase in the primarygain during U → VH in boys. Our findings therefore cohere with aprevious investigation (Koga et al., 2001) in which differences in theV̇O2 gain were dissociated from changes in quadriceps iEMG activityduring incremental exercise that recruited a smaller muscle masscompared to conventional cycle ergometry.
5. Conclusion
This study combined work-to-work exercise transitions withmeasurements of the integrated electromyogram (iEMG) in orderto explore the putative factors involved in modulating V̇O2 kineticsin boys and men. Step transitions to very heavy exercise initiatedfrom an elevated baseline work rate lengthened the phase II �in boys and men, however, the overall V̇O2 gain was increased inadults only compared to transitions elicited from unloaded ped-alling. Furthermore, the development of the V̇O2 slow componentduring unloaded-to-very heavy exercise was accompanied by anincreased rate of change in iEMG activity of the m. vastus later-alis in men but not in boys. These findings are consistent with thenotion that age related differences in V̇O2 kinetics may be linked toalterations in muscle fibre recruitment following the onset of veryheavy-intensity exercise.
Acknowledgements
We would like to thank the pupils and staff from Ivybridge Com-munity College for their participation in this research project.

logy &
R
A
A
B
B
B
B
B
B
B
B
B
B
B
C
C
D
D
E
F
G
G
H
H
J
B.C. Breese et al. / Respiratory Physio
eferences
rmon, Y., Cooper, D.M., Flores, R., Zanconato, S., Barstow, T.J., 1991. Oxygen uptakedynamics during high-intensity exercise in children and adults. J. Appl. Physiol.70, 841–848.
rmstrong, N., Barker, A.R., 2009. Oxygen uptake kinetics in children and adoles-cents: a review. Pediatr. Exerc. Sci. 21, 130–147.
ailey, S.J., Vanhatalo, A., Wilkerson, D.P., Dimenna, F.J., Jones, A.M., 2009. Optimizingthe priming effect: influence of prior exercise intensity and recovery durationon O2 uptake kinetics and severe-intensity exercise tolerance. J. Appl. Physiol.107, 1743–1756.
arker, A.R., Jones, A.M., Armstrong, N., 2010. The influence of priming exerciseon oxygen uptake, cardiac output, and muscle oxygenation kinetics duringvery heavy-intensity exercise in 9- to 13-yr-old boys. J. Appl. Physiol. 109,491–500.
arker, A.R., Williams, C.A., Jones, A.M., Armstrong, N., 2011. Establishing maximaloxygen uptake in young people during a ramp cycle test to exhaustion. Br. J.Sports Med. 45, 498–503.
arstow, T.J., Jones, A.M., Nguyen, P.H., Casaburi, R., 1996. Influence of muscle fibretype and pedal frequency on oxygen uptake kinetics of heavy exercise. J. Appl.Physiol. 81, 1642–1650.
earden, S.E., Moffatt, R.J., 2001. V̇O2 and heart rate kinetics in cycling: transitionsfrom an elevated baseline. J. Appl. Physiol. 90, 2081–2087.
eaver, W.L., Wasserman, K., Whipp, B.J., 1986. A new method for detecting anaer-obic threshold by gas exchange. J. Appl. Physiol. 60, 2020–2027.
ehnke, B.J., McDonough, P., Padilla, D.J., Musch, T.I., Poole, D.C., 2003. Oxygenexchange profile in rat muscles of contrasting fibre types. J. Physiol. 549,597–605.
reese, B.C., Williams, C.A., Barker, A.R., Welsman, J.R., Fawkner, S.G., Armstrong,N., 2010. Longitudinal changes in the oxygen uptake kinetic response toheavy-intensity exercise in 14- to 16-year-old boys. Pediatr. Exerc. Sci. 22,69–80.
reese, B.C., Armstrong, N., Barker, A.R., Williams, C.A., 2011. The effect of pedal rateon pulmonary O2 uptake kinetics during very heavy intensity exercise in trainedand untrained teenage boys. Respir. Physiol. Neurobiol. 177, 149–154.
rittain, C.J., Rossiter, H.B., Kowalchuk, J.M., Whipp, B.J., 2001. Effect of priormetabolic rate on the kinetics of oxygen uptake during moderate-intensity exer-cise. Eur. J. Appl. Physiol. 86, 125–134.
urnley, M., Doust, J.H., Ball, D., Jones, A.M., 2002. Effects of prior heavy exercise onV̇O2 kinetics during heavy exercise are related to changes in muscle activity. J.Appl. Physiol. 93, 167–174.
arter, H., Pringle, J.S., Boobis, L., Jones, A.M., Doust, J.H., 2004. Muscle glycogendepletion alters oxygen uptake kinetics during heavy exercise. Med. Sci. SportsExerc. 36, 965–972.
row, M.T., Kushmerick, M.J., 1982. Chemical energetics of slow- and fast-twitchmuscles of the mouse. J. Gen. Physiol. 79, 147–166.
iMenna, F.J., Wilkerson, D.P., Burnley, M., Jones, A.M., 2008. Influence ofpriming exercise on pulmonary O2 uptake kinetics during transitions tohigh-intensity exercise from an elevated baseline. J. Appl. Physiol. 105, 538–546.
iMenna, F.J., Bailey, S.J., Vanhatalo, A., Chidnok, W., Jones, A.M., 2010. Elevated base-line V̇O2 per se does not slow O2 uptake kinetics during work-to-work exercisetransitions. J. Appl. Physiol. 109, 1148–1154.
ndo, M.Y., Kobayakawa, M., Kinugasa, R., Kuno, S., Akima, H., Rossiter, H.B.,Miura, A., Fukuba, Y., 2007. Thigh muscle activation distribution and pul-monary V̇O2 kinetics during moderate, heavy, and very heavy intensity cyclingexercise in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 293, R812–R820.
awkner, S.G., Armstrong, N., 2004. Longitudinal changes in the kinetic response toheavy-intensity exercise in children. J. Appl. Physiol. 97, 460–466.
ollnick, P.D., Piehl, K., Saltin, B., 1974. Selective glycogen depletion pattern inhuman muscle fibres after exercise of varying intensity and at varying pedallingrates. J. Physiol. 241, 45–57.
rassi, B., Poole, D.C., Richardson, R.S., Knight, D.R., Erickson, B.K., Wagner, P.D., 1996.Muscle O2 uptake kinetics in humans: implications for metabolic control. J. Appl.Physiol. 80, 988–998.
enneman, E., Mendell, L.M., 1981. Functional organisation of a motorneuron pooland its inputs. In: Brooks, V.B. (Ed.), Handbook of Physiology I, vol. II, part 1.American Physiological Society, Bethesda, pp. 423–507.
ughson, R.L., Morrissey, M., 1982. Delayed kinetics of respiratory gas exchange in
the transition from prior exercise. J. Appl. Physiol. 52, 921–929.ones, A.M., Pringle, J.S., Carter, H., 2005. Influence of muscle fibre type and motorunit recruitment on V̇O2 kinetics. In: Jones, A.M., Poole, D.C. (Eds.), OxygenUptake Kinetics in Sport, Exercise, and Medicine. Routledge, London and NewYork, pp. 261–293.
Neurobiology 180 (2012) 223– 229 229
Jones, A.M., Grassi, B., Christensen, P.M., Krustrup, P., Bangsbo, J., Poole, D.C., 2011.Slow component of V̇O2 kinetics: mechanistic bases and practical applications.Med. Sci. Sports Exerc. 43, 2046–2062.
Koga, S., Barstow, T.J., Shiojiri, T., Takaishi, T., Fukuba, Y., Kondo, N., Shibasaki, M.,Poole, D.C., 2001. Effect of muscle mass on V̇O2 kinetics at the onset of work. J.Appl. Physiol. 90, 461–468.
Koga, S., Poole, D.C., Ferreira, L.F., Whipp, B.J., Kondo, N., Saitoh, T., Ohmae, E.,Barstow, T.J., 2007. Spatial heterogeneity of quadriceps muscle deoxygenationkinetics during cycle exercise. J. Appl. Physiol. 103, 2049–2056.
Krustrup, P., Soderlund, K., Mohr, M., Bangsbo, J., 2004. The slow component ofoxygen uptake during intense, sub-maximal exercise in man is associated withadditional fibre recruitment. Pflugers Arch. 447, 855–866.
Krustrup, P., Secher, N.H., Relu, M.U., Hellsten, Y., Soderlund, K., Bangsbo, J., 2008.Neuromuscular blockade of slow twitch muscle fibres elevates muscle oxygenuptake and energy turnover during submaximal exercise in humans. J. Physiol.586, 6037–6048.
Krustrup, P., Jones, A.M., Wilkerson, D.P., Calbet, J.A., Bangsbo, J., 2009. Muscularand pulmonary O2 uptake kinetics during moderate- and high-intensity sub-maximal knee-extensor exercise in humans. J. Physiol. 587, 1843–1856.
Lai, N., Nasca, M.M., Silva, M.A., Silva, F.T., Whipp, B.J., Cabrera, M.E., 2008. Influence ofexercise intensity on pulmonary oxygen uptake kinetics at the onset of exerciseand recovery in male adolescents. Appl. Physiol. Nutr. Metabol. 33, 107–117.
MacPhee, S.L., Shoemaker, J.K., Paterson, D.H., Kowalchuk, J.M., 2005. Kinetics ofO2 uptake, leg blood flow, and muscle deoxygenation are slowed in the uppercompared with lower region of the moderate-intensity exercise domain. J. Appl.Physiol. 99, 1822–1834.
McDonough, P., Behnke, B.J., Padilla, D.J., Musch, T.I., Poole, D.C., 2005. Control ofmicrovascular oxygen pressures in rat muscles comprised of different fibretypes. J. Physiol. 563, 903–913.
Mirwald, R.L., Baxter-Jones, A.D., Bailey, D.A., Beunen, G.P., 2002. An assessmentof maturity from anthropometric measurements. Med. Sci. Sports Exerc. 34,689–694.
Poole, D.C., Jones, A.M., 2005. Towards an understanding of the mechanistic bases ofV̇O2 kinetics. In: Jones, A.M., Poole, D.C. (Eds.), Oxygen Uptake Kinetics in Sport,Exercise and Medicine. Routledge, London and New York, pp. 294–328.
Pringle, J.S., Doust, J.H., Carter, H., Tolfrey, K., Campbell, I.T., Sakkas, G.K., Jones,A.M., 2003a. Oxygen uptake kinetics during moderate, heavy and severe inten-sity submaximal exercise in humans: the influence of muscle fibre type andcapillarisation. Eur. J. Appl. Physiol. 89, 289–300.
Pringle, J.S., Doust, J.H., Carter, H., Tolfrey, K., Jones, A.M., 2003b. Effect of pedal rateon primary and slow-component oxygen uptake responses during heavy-cycleexercise. J. Appl. Physiol. 94, 1501–1507.
Rossiter, H.B., Ward, S.A., Kowalchuk, J.M., Howe, F.A., Griffiths, J.R., Whipp, B.J., 2001.Effects of prior exercise on oxygen uptake and phosphocreatine kinetics duringhigh-intensity knee-extension exercise in humans. J. Physiol. 537, 291–303.
Rossiter, H.B., Ward, S.A., Howe, F.A., Kowalchuk, J.M., Griffiths, J.R., Whipp, B.J., 2002.Dynamics of intramuscular 31P-MRS Pi peak splitting and the slow componentsof PCr and O2 uptake during exercise. J. Appl. Physiol. 93, 2059–2069.
Sargeant, A.J., 1999. Neuromuscular determinants of human performance. In:Whipp, B.J., Sargeant, A.J. (Eds.), Physiological Determinants of Exercise Toler-ance in Humans. Portland Press, London, pp. 13–28.
Saunders, M.J., Evans, E.M., Arngrimsson, S.A., Allison, J.D., Warren, G.L., Cureton, K.J.,2000. Muscle activation and the slow component rise in oxygen uptake duringcycling. Med. Sci. Sports Exerc. 32, 2040–2045.
Scheuermann, B.W., Hoelting, B.D., Noble, M.L., Barstow, T.J., 2001. The slow compo-nent of O2 uptake is not accompanied by changes in muscle EMG during repeatedbouts of heavy exercise in humans. J. Physiol. 531, 245–256.
Shinohara, M., Moritani, T., 1992. Increase in neuromuscular activity and oxygenuptake during heavy exercise. Ann. Physiol. Anthropol. 11, 257–262.
Vanhatalo, A., Poole, D.C., Dimenna, F.J., Bailey, S.J., Jones, A.M., 2011. Muscle fiberrecruitment and the slow component of O2 uptake: constant work rate vs. all-outsprint exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 300, R700–R707.
Vollestad, N.K., Blom, P.C., 1985. Effect of varying exercise intensity on glycogendepletion in human muscle fibres. Acta Physiol. Scand. 125, 395–405.
Wilkerson, D.P., Jones, A.M., 2006. Influence of initial metabolic rate on pulmonaryO2 uptake on-kinetics during severe intensity exercise. Respir. Physiol. Neuro-biol. 152, 204–219.
Wilkerson, D.P., Jones, A.M., 2007. Effects of baseline metabolic rate on pulmonary O2
uptake on-kinetics during heavy-intensity exercise in humans. Respir. Physiol.Neurobiol. 156, 203–211.
Williams, C.A., Carter, H., Jones, A.M., Doust, J.H., 2001. Oxygen uptake kinetics duringtreadmill running in boys and men. J. Appl. Physiol. 90, 1700–1706.
Zoladz, J.A., Gladden, L.B., Hogan, M.C., Nieckarz, Z., Grassi, B., 2008. Progressiverecruitment of muscle fibers is not necessary for the slow component of V̇O2kinetics. J. Appl. Physiol. 105, 575–580.




















