
The ecology of the invasive cyanobacterium Cylindrospermopsis raciborskii (Nostocales, Cyanophyta)...
11
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Kokocinski, Mikolaj] On: 21 September 2010 Access details: Access Details: [subscription number 927163748] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK European Journal of Phycology Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713725516 The ecology of the invasive cyanobacterium Cylindrospermopsis raciborskii (Nostocales, Cyanophyta) in two hypereutrophic lakes dominated by Planktothrix agardhii (Oscillatoriales, Cyanophyta) Mikołaj Kokociński ab ; Karolina Stefaniak b ; Joanna Mankiewicz-Boczek c ; Katarzyna Izydorczyk c ; Janne Soininen d a Collegium Polonicum, Adam Mickiewicz University, 69-100 Słubice, Poland b Faculty of Biology, Adam Mickiewicz University, Poland c International Institute of the Polish Academy of Sciences - European Regional Centre for Ecohydrology u/a UNESCO, Tylna 3, 90-364 Łódź, Poland d Department of Environmental Sciences, University of Helsinki, Finland First published on: 21 September 2010 To cite this Article Kokociński, Mikołaj , Stefaniak, Karolina , Mankiewicz-Boczek, Joanna , Izydorczyk, Katarzyna and Soininen, Janne(2010) 'The ecology of the invasive cyanobacterium Cylindrospermopsis raciborskii (Nostocales, Cyanophyta) in two hypereutrophic lakes dominated by Planktothrix agardhii (Oscillatoriales, Cyanophyta)', European Journal of Phycology, 45: 4, 365 — 374, First published on: 21 September 2010 (iFirst) To link to this Article: DOI: 10.1080/09670262.2010.492916 URL: http://dx.doi.org/10.1080/09670262.2010.492916 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of The ecology of the invasive cyanobacterium Cylindrospermopsis raciborskii (Nostocales, Cyanophyta)...

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Kokocinski, Mikolaj]On: 21 September 2010Access details: Access Details: [subscription number 927163748]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
European Journal of PhycologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713725516
The ecology of the invasive cyanobacterium Cylindrospermopsisraciborskii (Nostocales, Cyanophyta) in two hypereutrophic lakesdominated by Planktothrix agardhii (Oscillatoriales, Cyanophyta)Mikołaj Kokocińskiab; Karolina Stefaniakb; Joanna Mankiewicz-Boczekc; Katarzyna Izydorczykc; JanneSoininend
a Collegium Polonicum, Adam Mickiewicz University, 69-100 Słubice, Poland b Faculty of Biology,Adam Mickiewicz University, Poland c International Institute of the Polish Academy of Sciences -European Regional Centre for Ecohydrology u/a UNESCO, Tylna 3, 90-364 Łódź, Poland d Departmentof Environmental Sciences, University of Helsinki, Finland
First published on: 21 September 2010
To cite this Article Kokociński, Mikołaj , Stefaniak, Karolina , Mankiewicz-Boczek, Joanna , Izydorczyk, Katarzyna andSoininen, Janne(2010) 'The ecology of the invasive cyanobacterium Cylindrospermopsis raciborskii (Nostocales,Cyanophyta) in two hypereutrophic lakes dominated by Planktothrix agardhii (Oscillatoriales, Cyanophyta)', EuropeanJournal of Phycology, 45: 4, 365 — 374, First published on: 21 September 2010 (iFirst)To link to this Article: DOI: 10.1080/09670262.2010.492916URL: http://dx.doi.org/10.1080/09670262.2010.492916
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Eur. J. Phycol. (2010) 45(4): 365–374
The ecology of the invasive cyanobacterium Cylindrospermopsis
raciborskii (Nostocales, Cyanophyta) in two hypereutrophic
lakes dominated by Planktothrix agardhii (Oscillatoriales,
Cyanophyta)
MIKOLAJ KOKOCINSKI1,2, KAROLINA STEFANIAK2, JOANNA MANKIEWICZ-BOCZEK3,
KATARZYNA IZYDORCZYK3 AND JANNE SOININEN4
1Collegium Polonicum, Adam Mickiewicz University, ul. Kosciuszki 1, 69-100 Slubice, Poland2Faculty of Biology, Adam Mickiewicz University, Poland3International Institute of the Polish Academy of Sciences – European Regional Centre for Ecohydrology u/a UNESCO, Tylna 3,
90-364 L=odz, Poland4Department of Environmental Sciences, University of Helsinki, Finland
(Received 23 October 2009; accepted 10 May 2010)
Biological invasions have attracted particular attention since they often result in serious consequences for natural ecosystems.
One planktonic invasive species is Cylindrospermopsis raciborskii, a cyanobacterium originally reported to occur exclusively
in the tropics. Over the last few decades its range has extended to temperate regions and it occupies shallow highly eutrophic
lakes previously dominated by other cyanobacteria. The purpose of this study was to examine the ecology of C. raciborskii
during Planktothrix agardhii blooms in two shallow lakes in western Poland and to determine whether these species have
different environmental preferences. Multiple linear regression showed that the biomass of P. agardhii was significantly
negatively related to Secchi depth in Lake Bninskie. In Lake Bytynskie, P. agardhii was significantly positively related to
concentrations of PO3�4 , chlorophyll a and total phosphorus and negatively related to Secchi depth, NO�3 , and total nitrogen.
Cylindrospermopsis raciborskii was significantly positively related only to concentrations of NHþ4 . There was a negative
correlation between the biomass of P. agardhii and C. raciborskii perhaps showing different responses to environmental
variables. Moreover, the biomass of P. agardhii was negatively correlated with Shannon–Wiener diversity of the phyto-
plankton assemblages. Our results support the concept that these cyanobacterial species have different environmental pref-
erences and their niches differ from each other. These results suggest that light is an important driver of phytoplankton
community structure resulting in shifts from a community dominated by P. agardhii in very turbid waters to more diverse
communities perhaps including the invasive C. raciborskii in clearer waters.
Key words: biological invasions, Cylindrospermopsis raciborskii, environmental factors, phytoplankton, Planktothrix agardhii,
shallow lakes
Introduction
Biological invasions are considered a major threatto global biodiversity and are an important part ofongoing global climate change. The number ofinvasions has grown considerably in recent dec-ades, many of them driven by human activitiesthat diminish natural barriers to dispersal (Wellset al., 1986; Dukes & Mooney, 1999). At presentonly a few habitats remain free of species intro-duced by humans (Fridriksson & Magnusson,1992). Ecological consequences of biotic invasionsvary enormously, but they often result in the
altered identity and functioning of natural commu-nities (Mack et al., 2000). Invaders may severelyaffect the ecosystems by modifying the speciesdominance structure, nutrient dynamics andlevels of primary productivity (Bertness, 1984;Vitousek, 1990).Successful invasions accompany the major com-
ponents of global change that include increasingtemperatures (Walther, 2003), nitrogen deposition,and CO2 concentration, plus changing patterns ofland use or disturbance regimes (Dukes &Mooney, 1999). Aquatic ecosystems are especiallysensitive to these changes, which often includemore frequent cyanobacterial blooms. Rising tem-peratures favour cyanobacteria both directly, as
Correspondence to: Mikolaj Kokocinski. E-mail:
ISSN 0967-0262 print/ISSN 1469-4433 online/10/04000365–374 � 2010 British Phycological Society
DOI: 10.1080/09670262.2010.492916
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010

they grow faster at higher temperatures during per-iods of water stratification, and indirectly bychanging patterns of precipitation and drought(Paerl & Huisman, 2008). These changes mayalso allow for the establishment of invasive cyano-bacterial species.Among the many examples of biological inva-
sions is the expansion of the cyanobacteriumCylindrospermopsis raciborskii (Woloszynska)Seenayya & Subba Raju in temperate aquatic eco-systems. This species, common in the tropics, hasbeen observed in temperate regions over the lasttwo decades (Padisak, 1997; Wiedner et al., 2002;Saker et al., 2003; Manti et al., 2005). Recently, itwas found in shallow, eutrophic, polymictic lakesof the Wielkopolska region in western Poland,often co-occurring with the native speciesPlanktothrix agardhii (Gomont) Anagnostidis &Komarek (Stefaniak & Kokocinski, 2005). Thesespecies are planktonic, filamentous cyanobacteriabelonging to the Nostocales and Oscillatoriales,respectively. Both species are of great concerndue to their ability to produce toxins harmful tohumans and animals. Planktothrix agardhii isknown for producing a variety of microcystins(Carmichael, 1994; Chorus, 2001), while C. raci-borskii is able to synthesize the neurotoxin saxi-toxin (Lagos et al., 1999; Castro et al., 2004) andthe alkaloid hepatotoxin cylindrospermopsin(Ohtani et al., 1992; Li et al., 2001).Although C. raciborskii and P. agardhii are able
to grow under varied conditions, their environmen-tal requirements probably differ. Temperature hasbeen considered the major limiting factor forgrowth of C. raciborskii (Padisak, 1997; Wiedneret al., 2007), with its optimal growth rate in labo-ratory experiments being at 25–35�C (Saker &Griffiths, 2000; Briand et al., 2004; Castro et al.,2004; Chonudomkul et al., 2004). Furthermore, itsakinete germination is temperature dependent, andgermination occurs primarily from 22–23.5�C(Padisak, 1997). In contrast, P. agardhii can growwell at much lower temperatures, 6–16�C (Berger,1989; Dokulil & Teubner, 2000) and long-lastingpopulations have been observed in highly eutro-phic reservoirs even during winter (Berger, 1975;Post et al., 1985; Nixdorf, 1994; Poulickovaet al., 2004). Nonetheless, it is well known thatP. agardhii populations develop optimally in latesummer or autumn, because it prefers warm watersabove 20�C (Van Liere & Mur, 1980; Ramberg,1988).Population densities of phytoplankton species
are also affected by light intensity. Based on earlierfindings, it was suggested that C. raciborskii canbenefit from low light intensities (Dokulil &Mayer, 1996; Padisak & Reynolds, 1998; Bouvyet al., 1999; Briand et al., 2002b; Mischke &
Nixdorf, 2003). However, recent experimentalstudies on tropical and temperate C. raciborskiiisolates showed no significant differences ingrowth rates at different light intensities (Briandet al., 2004). Planktothrix agardhii is also associ-ated with turbid waters, and is a common species inshallow, polymictic lakes with high disturbance fre-quency (Rucker et al., 1997; Scheffer et al., 1997;Nixdorf et al., 2003). Light and temperature andtheir interactions are currently gaining attentionsince global warming has increased water temper-atures in lakes during spring in addition to alteringphysical conditions (George et al., 2000; Gerten &Adrian, 2000). Moreover, relationships betweentemperature and photoperiod linked to climatewarming may influence phytoplankton composi-tion (Nicklisch et al., 2008).Nutrient concentrations also have a role in
structuring phytoplankton communities.Cylindrospermopsis raciborskii is characterized byits association with high TN/TP ratios (Briandet al., 2002b), while P. agardhii appears to befavoured by low TN/TP ratios (Rucker et al.,1997). Moreover, both species are able to developvery abundant populations or even form intenseblooms (Berger, 1975, 1989; De Souza et al.,1998; Fonseca & De M. Bicudo, 2008).Occurrence of C. raciborskii with mass develop-ment of P. agardhii is known in Polish lakes(Stefaniak & Kokocinski, 2005; Pelechata et al.,2006; Kokocinski et al., 2009) but documentedexamples of abundant coexistence of these speciesare sparse (Mischke & Nixdorf, 2003; Crossetti &De M. Bicudo, 2008).Although the autecology of these species is now
better known, the relationships between their envi-ronment and their occurrence in temperate lakes isless understood. Therefore, the objectives of thisstudy are: (i) to examine which environmental fac-tors drive the temporal patterns in phytoplanktoncommunities in the two lakes studied; (ii) to exam-ine which environmental factors control C. raci-borskii and P. agardhii concentrations andwhether the species respond differently to environ-mental factors; and (iii) determine whether thesetwo species coexist or show clear negative correla-tions in their abundances. We predict that growthof C. raciborskii will be favoured by high temper-atures and high light intensities, while P. agardhii isassociated with reduced light intensities and lowertemperatures (Fig. 1).
Materials and methods
Sampling sites
The study was conducted in two lakes, Bninskie (here-after ‘Lake BN’) and Bytynskie (hereafter ‘Lake BY’),
M. Kokocinski et al. 366
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010

located near the city of Poznan in the Wielkopolskaregion of western Poland. Both lakes are shallow (max-imum depth 58.5m), polymictic and highly eutrophic,surrounded by agricultural catchments, with public rec-reational usage. Their average depths are 4.2 and 3.5mrespectively, with other features given in Table 1.
Sampling
Sampling was carried out twice a month from July toOctober each year from 2004 to 2007. Altogether 32 sam-ples were collected from each lake during the study. As nothermal or oxygenation stratification was observed, andfollowing previous studies in shallow lakes (Berger, 1975;Romo & Miracle, 1993, 1994; Briand et al., 2002a; Ottet al., 2003) phytoplankton samples were collected from0.5m below the surface of the water. Samples for phyto-plankton and chemical analysis were collected using a 5 lLimnos water sampler at the deepest point of thereservoir. The phytoplankton samples were fixed withacidified Lugol’s solution (Wetzel & Likens, 2000)immediately after sampling. Then, formaldehyde wasadded as a preservative and samples were stored undercool and dark conditions until counted. Before counting,samples were sedimented in a 1 l glass cylinder for 48 h,after which the overlying water was gently decanted offand the lower layer (volume 40ml) was used for phyto-plankton analyses.
Phytoplankton analysis
Phytoplankton identification and counts were conductedusing a Zeiss light microscope (magnification 400�).
The phytoplankton counts were carried out in 160fields of the Fuchs-Rosenthal chamber, which ensuredthat at least 400 specimens were counted to reduce theerror to less than 10% (P¼ 0.05; Javornicky, 1958).Single cells, coenobia or filaments were considered asone specimen in this study. We assumed 100mm to bethe standard length for filaments and 100 cells the sizeof large spherical colonies. The biovolume of each specieswas determined via volumetric analysis of cells using geo-metric approximation and expressed as wet weight fol-lowing Hindak (1978) and Wetzel & Likens (2000). Foreach species geometrical dimensions of at least 30 indi-viduals were measured to obtain amean value. The diver-sity was calculated using the Shannon–Wiener diversityindex (Shannon, 1948) based on phytoplankton biomass.
Chemical and physical analysis
Water samples for chemical analyses were collected atthe same time as the phytoplankton samples, and wereanalysed for ammonium nitrogen (NHþ4 ), nitrate nitro-gen (NO�3 ), nitrite nitrogen (NO�2 ), total nitrogen (TN),orthophosphate phosphorus (PO3�
4 ) and total phospho-rus (TP) by using a HACH Spectrophotometer. Forchlorophyll a (chl a) analysis, 200ml of water was fil-tered through a GF/C Whatman filter. The concentra-tion was determined spectrophotometrically followingthe Lorenzen method after 90% acetone extractionand corrected for phaeopigments (Wetzel & Likens,2000). During field sampling, water temperature, pHand conductivity were determined using a multipara-meter Elmetron CPC-401 probe. Water transparencywas measured each time using a Secchi disc.
Statistical analyses
Prior to analyses, rare phytoplankton taxa, i.e. thosethat occurred in only one sample, were excluded fromthe ordinations. Environmental variables (except pH)were log-transformed to better approximate multivariatenormality. We first used canonical correspondence anal-ysis (CCA) to examine the temporal patterns of theentire phytoplankton community along major environ-mental gradients. CCA is a direct gradient analysis thatuses both species and environmental data by combiningordination and regression techniques (Ter Braak, 1986;Legendre & Legendre, 1998). We used forward selectionof environmental variables. At each step, only variablessignificantly (P50.05; Monte Carlo randomization testwith 499 permutations) related to assemblage structurewere included in the model. CCA was run usingCANOCO version 4.0 (Ter Braak & Smilauer, 1998).
We then analysed more closely which of the environ-mental variables were controlling the P. agardhii andC. raciborskii populations by using stepwise multipleregression. We used the probability level of 0.05 as thelimit for entry into the model. Before analysis, weexcluded variables that showed strongest (r40.80)co-linearity with other variables.
Spearman correlation analysis was carried out usingStatistica 7.1 to examine the correlation between bio-mass of P. agardhii and C. raciborskii. We then related
dominates
P. agardhiidominates
Temperature
Lig
ht
Coexistence
C. raciborskiidominates
P. agardhiidominates
Fig. 1. Conceptual model of the relationship betweenPlanktothrix agardhii and Cylindrospermopsis raciborskiiwith regard to temperature and light.
Table 1. Physical characteristics of Lake Bninskie (BN)
and Lake Bytynskie (BY) in western Poland.
Lake BN Lake BY
Latitude 52�12.7 52�30.5
Longitude 17�06.8 16�30.1
Area (ha) 225.9 308.8
Max. depth (m) 8.5 7.0
Average depth (m) 4.2 3.5
Max. width (m) 655 1490
Max. length (m) 4545 4160
Length of shore line (m) 11830 17475
Ecology of Cylindrospermopsis raciborskii in two hypereutrophic lakes 367
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010
the biomass of P. agardhii to phytoplankton speciesrichness using linear regression analysis.
Results
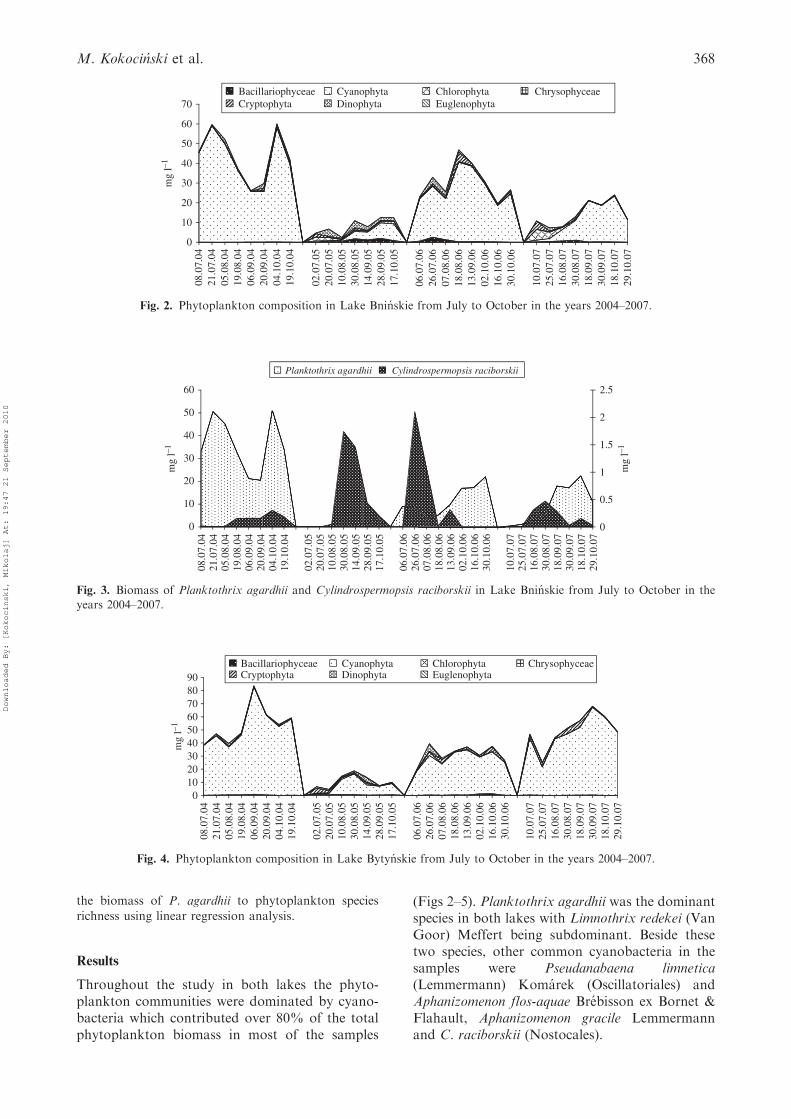
Throughout the study in both lakes the phyto-plankton communities were dominated by cyano-bacteria which contributed over 80% of the totalphytoplankton biomass in most of the samples
(Figs 2–5). Planktothrix agardhii was the dominantspecies in both lakes with Limnothrix redekei (VanGoor) Meffert being subdominant. Beside thesetwo species, other common cyanobacteria in thesamples were Pseudanabaena limnetica(Lemmermann) Komarek (Oscillatoriales) andAphanizomenon flos-aquae Brebisson ex Bornet &Flahault, Aphanizomenon gracile Lemmermannand C. raciborskii (Nostocales).
0
10
20
30
40
50
60
08.0
7.04
21.0
7.04
05.0
8.04
19.0
8.04
06.0
9.04
20.0
9.04
04.1
0.04
19.1
0.04
02.0
7.05
20.0
7.05
10.0
8.05
30.0
8.05
14.0
9.05
28.0
9.05
17.1
0.05
06.0
7.06
26.0
7.06
07.0
8.06
18.0
8.06
13.0
9.06
02.1
0.06
16.1
0.06
30.1
0.06
10.0
7.07
25.0
7.07
16.0
8.07
30.0
8.07
18.0
9.07
30.0
9.07
18.1
0.07
29.1
0.07
mg
l–1
mg
l–1
0
0.5
1
1.5
2
2.5
Planktothrix agardhii Cylindrospermopsis raciborskii
Fig. 3. Biomass of Planktothrix agardhii and Cylindrospermopsis raciborskii in Lake Bninskie from July to October in theyears 2004–2007.
0
10
20
30
40
50
60
70
08.0
7.04
21.0
7.04
05.0
8.04
19.0
8.04
06.0
9.04
20.0
9.04
04.1
0.04
19.1
0.04
02.0
7.05
20.0
7.05
10.0
8.05
30.0
8.05
14.0
9.05
28.0
9.05
17.1
0.05
06.0
7.06
26.0
7.06
07.0
8.06
18.0
8.06
13.0
9.06
02.1
0.06
16.1
0.06
30.1
0.06
10.0
7.07
25.0
7.07
16.0
8.07
30.0
8.07
18.0
9.07
30.0
9.07
18.1
0.07
29.1
0.07
mg
l–1
Bacillariophyceae Cyanophyta Chlorophyta ChrysophyceaeCryptophyta Dinophyta Euglenophyta
Fig. 2. Phytoplankton composition in Lake Bninskie from July to October in the years 2004–2007.
0102030405060708090
08.0
7.04
21.0
7.04
05.0
8.04
19.0
8.04
06.0
9.04
20.0
9.04
04.1
0.04
19.1
0.04
02.0
7.05
20.0
7.05
10.0
8.05
30.0
8.05
14.0
9.05
28.0
9.05
17.1
0.05
06.0
7.06
26.0
7.06
07.0
8.06
18.0
8.06
13.0
9.06
02.1
0.06
16.1
0.06
30.1
0.06
10.0
7.07
25.0
7.07
16.0
8.07
30.0
8.07
18.0
9.07
30.0
9.07
18.1
0.07
29.1
0.07
mg
l–1
Bacillariophyceae Cyanophyta Chlorophyta ChrysophyceaeCryptophyta Dinophyta Euglenophyta
Fig. 4. Phytoplankton composition in Lake Bytynskie from July to October in the years 2004–2007.
M. Kokocinski et al. 368
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010

Chlorophyta, Bacillariophyceae and other algalgroups were less abundant, never dominating thecommunities. Higher concentrations of algae otherthan cyanobacteria were observed only in 2005when the total phytoplankton biomass was thelowest over the study period. During the beginningof the sampling period, there was an increased totalbiomass of Chlorophyta and Dinophyta in LakeBN and of Chlorophyta and Cryptophyta inLake BY. Similarly in 2007, Chlorophyta andCryptophyta were relatively common in LakeBN. The dominant species were Coelastrum micro-porumNag. in A. Braun, Oocystis lacustris Chodat,Scenedesmus ecornis (Ehr. ex Ralfs) Chodat,Monoraphidium griffithii (Berk.) Kom.-Legn.(Chlorophyta), Cryptomonas erosa Ehrenberg,Cryptomonas reflexa (M. Marsson.) Skuja,Cryptopmonas marsonii Skuja, Rhodomonasminuta Skuja (Cryptophyta) and Peridiniopsis ber-olinense (Lemm.) Bourrelly (Dinophyta).The biomass of C. raciborskii in Lake BY was
generally slightly higher than in Lake BN duringthe period studied, with the exception of 2007. Thehighest concentration of C. raciborskii in bothlakes occurred in 2006 (Figs 2–5) but the highestcontribution of C. raciborskii to the total phyto-plankton biomass was in 2005 when it contributed18.5% of the total biomass in Lake BN and 9.3%in Lake BY.In Lake BN, the eigenvalues of the first two CCA
axes were 0.399 and 0.338 and significant (P50.05,Table 2). The first two CCA axes jointly explained31.8% of the total variance (2.00) in the phyto-plankton communities. The species–environmentcorrelations for CCA axes were relatively high, indi-cating a strong relationship between phytoplanktonand the environmental variables. Secchi depth, chla, and the TN/TP ratio were the most significantcontributors to CCA axis 1 (Fig. 6). CCA axis 2was most strongly correlated with PO3�
4 concentra-tions, temperature and water pH.
In Lake BY the eigenvalues of the first two CCAaxes were 0.462 and 0.208 and significant (P50.05,Table 2). The first two axes jointly explained35.2% of the total variance (1.53) in the commu-nities. The species-environment correlations forCCA axes were also high in Lake BY, indicatinga relatively strong relationship between phyto-plankton and the environmental variables. Secchidepth, pH, chl a, and the amount of total nutrientswere the most significant contributors to CCA axis1, with PO3�
4 , chl a and TN/TP ratio contributingmost to CCA axis 2 (Fig. 7).According to multiple regression analyses, bio-
mass of P. agardhii was significantly (P50.001)negatively related to Secchi depth in Lake BN(Table 3). In Lake BN, C. raciborskii was notrelated to any of the measured environmental var-iables. In Lake BY P. agardhii was significantly
0
10
20
30
40
50
60
70
80
08.0
7.04
21.0
7.04
05.0
8.04
19.0
8.04
06.0
9.04
20.0
9.04
04.1
0.04
19.1
0.04
02.0
7.05
20.0
7.05
10.0
8.05
30.0
8.05
14.0
9.05
28.0
9.05
17.1
0.05
06.0
7.06
26.0
7.06
07.0
8.06
18.0
8.06
13.0
9.06
02.1
0.06
16.1
0.06
30.1
0.06
10.0
7.07
25.0
7.07
16.0
8.07
30.0
8.07
18.0
9.07
30.0
9.07
18.1
0.07
29.1
0.07
mg
l–1
mg
l–1
0
0.5
1
1.5
2
2.5
3
3.5
Planktothrix agardhii Cylindrospermopsis raciborskii
Fig. 5. Biomass of Planktothrix agardhii and Cylindrospermopsis raciborskii in Lake Bytynskie from July to October in theyears 2004–2007.
Table 2. The results of Canonical CorrespondenceAnalyses (CCA) with forward selected variables (P50.05)for Lake Bninskie (BN) and Lake Bytynskie (BY).
Lake BN Lake BY
Axis 1 Axis 2 Axis 1 Axis 2
Eigenvalue 0.399 0.338 0.462 0.208
Species-environment
correlation
0.890 0.935 0.963 0.762
Cumulative
percentage variance
of species data
17.2 31.8 24.3 35.2
Inter-set correlations
with axes
Temperature �0.060 0.390 0.345 �0.002
pH �0.044 �0.335 �0.727 0.122
SD �0.817 0.153 0.767 0.077
PO3�4 �0.218 �0.730 �0.028 0.323
NHþ4 0.216 0.235 0.590 �0.142
NO�3 �0.357 �0.241 0.586 �0.121
TP �0.154 �0.186 0.626 �0.096
TN 0.147 �0.255 0.667 �0.135
TN:TP 0.379 �0.007 �0.015 �0.160
Chl a 0.559 �0.112 �0.700 �0.219
Ecology of Cylindrospermopsis raciborskii in two hypereutrophic lakes 369
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010

(P50.001) positively related to PO3�4 , chl a and
TP; and negatively related to Secchi depth, NO�3 ,and TN. C. raciborskii was also significantly(P50.001) positively related to NHþ4 .According to the Spearman correlation analysis,
P. agardhii was significantly negatively correlated(r¼ 0.149, p50.05) with C. raciborskii in Lake BN.Planktothrix agardhii was significantly negativelycorrelated (r¼ 0.438, P50.05) with C. raciborskiiin Lake BY. In addition, P. agardhii was signifi-cantly negatively correlated (r¼ 0.942, P50.05)with Shannon–Wiener diversity in Lake BN andin Lake BY (r¼ 0.781, P50.05).Cylindrospermopsis raciborskii was significantlypositively correlated (r¼ 0.542) with Shannon–Wiener diversity in Lake BY. In Lake BN, the cor-relation of C. raciborskii with Shannon–Wienerdiversity was non-significant (r¼ 0.208).
Discussion
Oscillatoriales dominated the phytoplankton com-munities in both lakes. Although communitieswere strongly dominated by P. agardhii, L. redekeiwas also abundant, thus supporting findings byRucker et al. (1997) that both species are oftenpresent in shallow and eutrophic waters, and com-pete successfully with other filamentous cyanobac-teria since they tolerate very low light conditions.In addition to the Oscillatoriales, Nostocales such
as Aphanizomenon flos-aquae, Aphanizomenongracile and C. raciborskii were relatively abundant.Cylindrospermopsis raciborskii was often presentand its abundance increased slightly after 2003(Stefaniak & Kokocinski, 2005). Its increasedabundance is typical for many eutrophic temperatelakes (Nixdorf et al., 2003; Stuken et al., 2006).Cylindrospermopsis raciborskii reached its highestbiomass during 2005, but biomass was nonethelessrelatively low in comparison with other groups ofalgae.The CCA indicated that turnover of the phyto-
plankton composition was controlled moststrongly by changes in Secchi depth (suggestingdifferences in light intensity), water temperature,water pH and nutrient concentrations. The resultsare highly congruent with other studies where theimportance of nutrient concentrations, tempera-ture and light has been documented (Forsstromet al., 2005; Grover & Chrzanowski, 2006;Kokocinski & Soininen, 2008). Although P. agard-hii and C. raciborskii often co-occurred, multipleregression analyses confirmed that these speciesresponded differently to several environmental var-iables. For example, the importance of light inten-sity for regulating P. agardhii was supported giventhat Secchi depth was a significant explanatoryvariable of its biomass in both lakes.More specifically, the biomass of P. agardhii wasnegatively related to Secchi depth supporting
Aphanizomenon flos-aquae
Aphanizomenongracile
Cylindrospermopsisraciborskii
Limnothrixredekei
Planktothrixagardhii
Pseudanabaena limnetica
Cryptomonaserosa
Cryptomonasmarssonii
Cryptomonasreflexa
Ceratium hirundinellaTemp
pH
SD
PO4
NH4
NO3 Tot P Tot N
N/P
Chl-a 11–3–
–1
1
Axis 1
Axi
s 2
Fig. 6. Ordination diagram for Canonical Correspondence Analysis of the phytoplankton assemblages in Lake Bninskie.
M. Kokocinski et al. 370
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010

earlier findings that it is able to grow exceedinglywell under low light intensities (Rucker et al., 1997;Scheffer et al., 1997; Nixdorf et al., 2003).Moreover, the finding that the biomass ofC. raciborskii was not related to Secchi depth wascongruent with our initial expectation that C. raci-borskii can grow under different light intensitiesand at high temperature (Fig. 1). Our findingsalso concur with Briand et al. (2004) since therewere no significant differences in growth rates ofC. raciborskii at different light intensities.
Both species preferred nutrient-rich waters, withP. agardhii being positively related to TP and PO3�
4
concentrations (in Lake BY), and C. raciborskii toNHþ4 concentrations. The fact that C. raciborskiiwas not related to phosphorus may suggest that ithas a high uptake and high storage capacity forphosphorus allowing it to grow under relativelylow phosphate supply (Branco & Senna, 1994;Presing et al., 1996; Isvanovics et al., 2000;Briand et al., 2002b). Recent experimental studiesalso showed that C. raciborskii is able to grow
Fragilaria ulna
Aphanizomenon gracile
Aphanizomenonissatschenkoi
Cylindrospermopsis raciborskii
Limnothrixredekei
Planktothrixagardhii
Pseudanabaena limnetica
Botryococcus braunii
Cryptomonas reflexa
Peridinium elpatiewskii
TemppH
SD
PO4
NH4
NO3
Tot PTot NN/PChl-a–2 2
Axis 1
Axi
s 2
Fig. 7. Ordination diagram for Canonical Correspondence Analysis of the phytoplankton assemblages in Lake Bytynskie.
Table 3. The results of multiple regression analyses for Lake Bninskie (BN) and Lake Bytynskie (BY) showing the main
environmental factors related to the abundance of P. agardhii and C. raciborskii.
Model R2 P
Lake BN
Planktothrix agardhii Y¼ 54.81� 64.6 � SD 0.517 0.000
Cylindrospermopsis raciborskii No significant variables in the model
Lake BY
Planktothrix agardhii Y¼ 54.16� 50.6 � SD� 0.04 �NO�3 þ 0.088 �PO3�4 þ 0.119 �Chla-a� 0.007 �TNþ 0.032 �TP 0.918 0.000
Cylindrospermopsis raciborskii Y¼ 0.122þ 0.003 �NHþ4 0.318 0.001
Ecology of Cylindrospermopsis raciborskii in two hypereutrophic lakes 371
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010

faster and sustain a high biomass under phospho-rus limitation during a sufficient supply of NHþ4 orNO�3 (Kenesi et al., 2009). In addition, NHþ4 wasshown to be effectively assimilated by C. racibors-kii in previous experimental studies by Spro
00
beret al. (2003). A positive relationship betweenC. raciborskii and NHþ4 in our study seems to sup-port these findings. In contrast, the growth ofP. agardhii is favoured by higher concentrationsof phosphorus and its dominance is especiallymarked in shallow, turbid lakes with intensiveinteraction between the sediment and watercolumn (Nixdorf et al., 2003; Noges & Jensen,2003).The different environmental requirements of
C. raciborskii and P. agardhii were indicated by theregression analyses where their abundances had anegative relationship. At low light intensities andlow temperature P. agardhii dominated the com-munity, whereas C. raciborskii was favoured byhigher temperatures and greater light intensities.The latter was evident, however, only in CCAwhere C. raciborskii was positively related toSecchi depth and water temperature. These find-ings are congruent with our initial predictions(Fig. 1). The finding that P. agardhii grows bestin turbid waters is also in agreement with studies byNixdorf et al. (2003) who place P. agardhii in thephytoplankton functional group S1 (sensuReynoldset al., 2002) of photoadapting filamentous cyano-bacteria common in shallow, wind-exposed lakeswith reduced light availability and intense sedi-ment–water interactions. Although C. raciborskiinever reached dominance, it increased in abundancewhen P. agardhii was less abundant.It is feasible that in this situation C. raciborskii
was able to out-compete P. agardhii, and theincreasing number of successful invasions byC. raciborskii in temperate regions supports thisview. This finding supports the hypothesis ofReynolds et al. (2002) that P. agardhii and C. raci-borskii belong to different functional groups.According to Padisak & Reynolds (1998) andReynolds et al. (2002) C. raciborskii has been allo-cated to the SN phytoplankton functional groupcharacteristic of warm, mixed waters enrichedwith phosphorus. The Shannon–Wiener diversitywas also consistently low when there was a highbiomass of P. agardhii. In contrast, C. raciborskiiwas positively correlated with Shannon–Wienerdiversity in Lake BY. These results indicate thatP. agardhii can be considered as a steady-state spe-cies during the Oscillatoriaceae-dominated steadystate in shallow lakes (Dokulil & Teubner, 2000;Scheffer et al., 1997; Scheffer, 1998) when few spe-cies are able to coexist (Nixdorf et al., 2003).To conclude, the abundances of P. agardhii and
the invasive C. raciborskii are negatively related,
possibly because they have different environmentalniches. As we initially predicted, P. agardhii isfavoured by turbid conditions with elevated phos-phorus concentrations, while the abundance ofC. raciborskii is directly related to ammoniumnitrogen and it prefers less turbid warmer waters.Our results may therefore shed more light onmechanisms promoting the invasion of C. racibors-kii in temperate regions. Future studies are neededto examine if C. raciborskii is able to sustain largepopulations in temperate lakes and to out-competethe native cyanobacteria.
Acknowledgements
Financial support was provided by the PolishMinistry of Science and Higher Education throughresearch grant No. N 304 051 31/1855 and by theAcademy of Finland and Kone foundation (to JS).We thank Dr Harold G. Marshall for his Englishlanguage revision and helpful comments on themanuscript.
References
BERGER, C. (1975). Occurrence of Oscillatoria agardhii Gom. in
some shallow eutrophic lakes. Verh. Internat. Verein. Limnol.,
19: 2689–2697.
BERGER, C. (1989). In situ primary production, biomass and light
regime in the Wolderwijd, the most stable Oscillatoria agardhii
lake in the Netherlands. Hydrobiologia, 188: 233–344.
BERTNESS, M.D. (1984). Habitat and community modification by
an introduced herbivorous snail. Ecology, 65: 370–381.
BOUVY, M., MOLICA, R., DE, OLIVEIRA, S., MARINHO, M. &
BEKER, B. (1999). Dynamics of a toxic cyanobacterial bloom
(Cylindrospermopsis raciborskii) in a shallow reservoir in the
semi-arid region of northern Brazil. Aquat. Microb. Ecol., 20:
285–297.
BRANCO, C.W.C. & SENNA, P.A.C. (1994). Factors influencing the
development of Cylindrospermopsis raciborskii and Microcystis
aeruginosa in the Paranoa Reservoir, Brasilia, Brazil. Algol.
Stud., 75: 85–96.
BRIAND, J.F., ROBILLOT, C., QUIBLIER-LLOBERAS, C. & BERNARD, C.
(2002a). A perennial bloom of Planktothrix agardhii
(Cyanobacteria) in a shallow eutrophic French lake: limnological
and microcystin production studies. Arch. Hydrobiol., 153:
605–622.
BRIAND, J.F., ROBILLOT, C., QUIBLIER-LIOBERAS, C., HUMBERT, J.F.,
COUTE, A. & BERNARD, C. (2002b). Environmental context of
Cylindrospermopsis raciborskii (Cyanobacteria) blooms in shal-
low pond in France. Water Res., 36: 3183–3192.
BRIAND, J.F., LEBOULANGER, C., HUMBERT, J.F., BERNARD, C. &
DUFOUR, P. (2004). Cylindrospermopsis raciborskii
(Cyanobacteria) invasion at mid-latitudes: selection, wide phy-
siological tolerance, or global warming? J. Phycol., 40: 231–238.
CASTRO, D., VERA, D., LAGOS, N., GARCIA, C. & VASQUEZ, M.
(2004). The effect of temperature on growth and production of
paralytic shellfish poisoning toxins by the cyanobacterium
Cylindrospermopsis raciborskii C10. Toxicon, 44: 483–489.
CARMICHAEL, W.W. (1994). The toxins of Cyanobacteria. Sci. Am.,
270: 64–70.
CHONUDOMKUL, D., YONGMANITCHAI, W., THEERAGOOL, G.,
KAWACHI, M., KASAI, F., KAYA, K. & WATANABE, M.M.
(2004). Morphology, genetic diversity, temperature tolerance
and toxicity of Cylindrospermopsis raciborskii (Nostocales,
M. Kokocinski et al. 372
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010

Cyanobacteria) strains from Thailand and Japan. FEMS
Microbiol. Ecol., 48: 345–355.
CHORUS, I. (2001). Cyanotoxins: Occurrence, Causes, Consequences.
Springer-Verlag KG, Berlin.
CROSSETTI, L.O. & DE M. BICUDO, C. (2008). Adaptations in phy-
toplankton life strategies to imposed change in a shallow urban
tropical eutrophic reservoir, Garcas Reservoir, over 8 years.
Hydrobiologia, 614: 91–105.
DE SOUZA, R.C.R., CARVALHO, M.C. & TRUZZI, A.C. (1998).
Cylindrospermopsis raciborskii (Wolosz.) Seenaya and Subba
Raju (Cyanophyceae) dominance and a contribution to the
knowledge of Rio Pequeno arm, Billings Reservoir, Brazil.
Environ. Toxicol. Water Qual., 13: 73–81.
DOKULIL, M.T. & MAYER, J. (1996). Population dynamics and
photosynthetic rates of a Cylindrospermopsis–Limnothrix asso-
ciation in highly eutrophic urban lake, Alte Donau, Austria.
Algol. Stud., 83: 179–195.
DOKULIL, M.T. & TEUBNER, K. (2000). Cyanobacterial dominance
in lakes. Hydrobiologia, 438: 1–12.
DUKES, J.S. & MOONEY, H.A. (1999). Does global change increase
the success of biological invaders? Trends Ecol. Evol., 14:
135–139.
FONSECA, B.M. & DE M. BICUDO, C.E. (2008). Phytoplankton sea-
sonal variation in a shallow stratified eutrophic reservoir (Garcas
Pond, Brazil). Hydrobiologia, 600: 267–282.
FORSSTROM, L., SORVARI, S. & KORHOLA, A. (2005). Seasonality of
phytoplankton in subarctic Lake Saanajarvi in NW Finnish
Lapland. Polar Biol., 28: 846–861.
FRIDRIKSSON, S. & MAGNUSSON, B. (1992). Development of the
ecosystem on Surtsey with references to Anak Krakatau.
GeoJournal, 28: 287–291.
GEORGE, D.G., TALLING, J.F. & RIGG, E. (2000). Factors influen-
cing the temporal coherence of five lakes in the English Lake
District. Freshwat. Biol., 43: 449–461.
GERTEN, D. & ADRIAN, R. (2000). Climate-driven changes in spring
plankton dynamics and the sensitivity of shallow polymictic lakes
to the North Atlantic Oscillation. Limnol. Oceanogr., 45:
1058–1066.
GROVER, J.P. & CHRZANOWSKI, T.H. (2006). Seasonal dynamics of
phytoplankton in two warm temperate reservoirs: association of
taxonomic composition with temperature. J. Plankton Res., 28:
1–17.
HINDAK, F., editor (1978). Freshwater Algae. Slovenske
Pedagogicke Nakladelstvo, Bratislava.
ISVANOVICS, V., SHAFIK, H.M., PRESING, M. & JUHOS, S. (2000).
Growth and phosphate uptake kinetics of the cyanobacterium,
Cylindrospermopsis raciborskii (Cyanophyceae) in throughflow
cultures. Freshwat. Biol., 2: 257–275.
JAVORNICKY, P. (1958). Revise nekterych method pro
zjist’ovanıkvantity fitoplantonu (Revision of some methods for
the phytoplankton quantity estimation). Scientific Paper 2,
Faculty of Technology of Fuel and Water, Institute of
Chemical Technology, Prague, pp. 283–267.
KENESI, G., SHAFIK, H.M., KOVACS, A.W., HERODEK, S. &
PRESING, M. (2009). Effect of nitrogen forms on growth, cell
composition and N2 fixation of Cylindrospermopsis raciborskii
in phosphorus-limited chemostat cultures. Hydrobiologia, 623:
191–202.
KOKOCINSKI, M., DZIGA, D., SPOOF, L., STEFANIAK, K., JURCZAK, T.,
MANKIEWICZ-BOCZEK, J. & MERILUOTO, J. (2009). First report of
the cyanobacterial toxin cylindrospermopsin in the shallow,
eutrophic lakes of western Poland. Chemosphere, 74: 669–675.
KOKOCINSKI, M. & SOININEN, J. (2008). Temporal variation in phy-
toplankton in two lakes with contrasting disturbance regimes.
Fundam. Appl. Limnol., 171: 39–48.
LAGOS, N., ONODERA, H., ZAGATTO, P.A., ANDRINOLO, D.,
AZEVEDO, S.M.F.Q. & OSHIMA, Y. (1999). The first evidence of
paralytic shellfish toxins in the freshwater cyanobacterium
Cylindrospermopsis raciborskii, isolated from Brazil. Toxicon,
37: 1359–1373.
LEGENDRE, P. & LEGENDRE, L. (1998). Numerical ecology. 2nd
English ed. Elsevier Science BV, Amsterdam.
LI, R., CARMICHAEL, W.W., BRITTAIN, S., EAGLESHAM, G.K.,
SHAW, G.R., NOPARATNARAPORN, A.M.N., YONGMANITCHAI, W.,
KAYA, K. & WATANABE, M.M. (2001). Detection of
cylindrospermopsin from a strain of Cylindrospermopsis
raciborskii (Cyanobacteria) isolated from Thailand. Toxicon,
39: 973–980.
MACK, R.N., SIMBERLOFF, D., LONSDALE, W.M., EVANS, H.,
CLOUT, M. & BAZZAZ, F.A. (2000). Biotic invasions: causes, epi-
demiology, global consequences and control. Ecol. Appl., 10:
689–710.
MANTI, G., MATTEI, D., MESSINEO, V., MELCHIORRE, S.,
BOGIALLI, S., SECHI, N., CASIDDU, P., LUGLIO, L., DI BRIZIO, M.
& BRUNO, M. (2005). First report of Cylindrospermopsis racibors-
kii (Nostocales, Cyanophyta) in Italy. Harmful Algae News, 28:
8–9.
MISCHKE, U. & NIXDORF, B. (2003). Equilibrium phase
conditions in shallow German lakes: how Cyanoprocaryota
species establish a steady state phase in late summer.
Hydrobiologia, 502: 123–132.
NICKLISCH, A., SHATWELL, T. & KOHLER, J. (2008). Analysis and
modelling of the interactive effects of temperature and light on
phytoplankton growth and relevance for the spring bloom.
J. Plankton Res., 30: 75–91.
NIXDORF, B. (1994). Polymixis of a shallow lake (Großer
Muggelsee, Berlin) and its influence on seasonal phytoplankton
dynamics. Hydrobiologia, 275/276: 173–186.
NIXDORF, B., MISCHKE, U. & RUCKER, J. (2003). Phytoplankton
assemblages and steady state in deep and shallow eutrophic
lakes – an approach to differentiate the habitat properties of
Oscillatoriales. Hydrobiologia, 502: 111–121.
NO00
GES, P. & JENSEN, J.P. (2003). Occurrence, coexistence and
competition of Limnothrix redekei and Planktothrix agardhii:
analysis of Danish–Estonian lake database. Algol. Stud., 109
(Cyanobacterial Research 4): 429–441.
OHTANI, I., MOORE, R.E. & RUNNEGAR, M.T.C. (1992).
Cylindrospermopsin: a potent hepatotoxin from the blue-green
alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc., 114:
7941–7942.
OTT, I., NOGES, P., LAUGASTE, R. & KOIV, T. (2003). Occurrence of
Limnothrix redekei Van Goor in Estonian lakes. Algol. Stud., 109
(Cyanobacterial Research 4): 455–468.
PADISAK, J. (1997). Cylindrospermopsis raciborskii (Woloszynska)
Seenayya et Subba Raju, an expanding, highly adaptative cya-
nobacterium: worldwide distribution and review of its ecology.
Arch. Hydrobiol., Suppl., 107: 563–593.
PADISAK, J. & REYNOLDS, C.S. (1998). Selection of phytoplankton
associations in Lake Balaton, Hungary, in response to eutrophi-
cation and restoration measures, with special reference to the
cyanoprokaryotes. Hydrobiologia, 384: 41–53.
PAERL, H.W. & HUISMAN, J. (2008). Blooms like it hot. Science,
320: 57–58.
PELECHATA, A., PELECHATY, M. & PUKACZ, A. (2006).
Cyanoprokaryota of shallow lakes of Lubuskie Lakeland (mid-
western Poland). Ocean. Hydrob. Stud., 35: 3–14.
POST, A., DE WIT, R. & MUR, L.R. (1985). Interactions between
temperature and light intensity on growth and photosynthesis of
the cyanobacterium Oscillatoria agardhii. J. Plankton Res, 7:
487–495.
POULICKOVA, A., HASLER, P. & KITNER, M. (2004). Annual
cycle of Planktothrix agardhii (Gommont) Anagnostidis &
Komarek nature population. Int. Rev. Hydrobiol, 89:
278–288.
PRESING, M., HERODEK, S., VOROS, L. & KOBOR, I. (1996). Nitrogen
fixation, ammonium and nitrate uptake during a bloom of
Cylindrospermopsis raciborskii in Lake Balaton. Arch.
Hydrobiol, 136: 553–562.
RAMBERG, L. (1988). Relations between planktic blue-green algal
dynamics and environmental factors in four eutrophic Swedish
lakes. Arch. Hydrobiol, 112: 161–175.
REYNOLDS, C.S., HUSZAR, V., KRUK, C., NASELLI-FLORES, L. &
MELO, S. (2002). Towards a functional classification of the fresh-
water phytoplankton. J. Plankton Res., 24: 417–428.
Ecology of Cylindrospermopsis raciborskii in two hypereutrophic lakes 373
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010

ROMO, S. & MIRACLE, M.R. (1993). Long-term periodicity of
Planktothrix agardhii, Pseudanabaena galeata and Geitlerinema
sp. in a shallow hypertrophic lagoon, the Albufera of Valencia
(Spain). Arch. Hydrobiol., 126: 469–486.
ROMO, S. & MIRACLE, M.R. (1994). Long-term phytoplankton
changes in a shallow hypertrophic lake, Albufera of Valencia
(Spain). Hydrobiologia, 275: 153–164.
RUCKER, J., WIEDNER, C. & ZIPPEL, P. (1997). Factors controlling
the dominance of Planktothrix agardhii and Limnothrix redekei
in eutrophic shallow lakes. Hydrobiologia, 342/343: 107–115.
SAKER, M.L. & GRIFFITHS, D.J. (2000). The effect of temperature
on growth and cylindrospermopsin content of seven isolates of
the cyanobacterium Cylindrospermopsis raciborskii
(Woloszynska) Seenayya and Subba Raju from water bodies in
northern Australia. Phycologia, 39: 349–354.
SAKER, M.L., NOGUEIRA, I.C.G., VASCONCELOS, V.M.,
NEILAN, B.A., EAGLESHAM, G.K. & PEREIRA, P. (2003). First
report and toxicological assessment of the cyanobacterium
Cylindrospermopsis raciborskii from Portuguese freshwaters.
Ecotoxicol. Environ. Safety, 55: 243–250.
SCHEFFER, M. (1998). Ecology of Shallow Lakes. Chapman & Hall,
London.
SCHEFFER, M., RINALDI, S., GRAGNAMI, A., MUR, L.R. &
VAN NES, E.H. (1997). On the dominance of filamentous
Cyanobacteria in shallow, turbid lakes. Ecology, 78: 272–282.
SHANNON, C.E. (1948). A mathematical theory of communication.
Bell System Technical Journal, 27: 626–656.
SPRO00
BER, P., SHAFIK, M.H., PRESING, M., KOVACS, A.W. &
HERODEK, S. (2003). Nitrogen uptake and fixation in the cyano-
bacterium Cylindrospermopsis raciborskii under different nitro-
gen conditions. Hydrobiologia, 506–509: 169–174.
STEFANIAK, K. & KOKOCINSKI, M. (2005). Occurrence of invasive
Cyanobacteria species in polimictic lakes of the Wielkopolska
region (Western Poland). Ocean. Hydrob. Stud., 34, Suppl, 3:
137–148.
STUKEN, A., RUCKER, J., ENDRULAT, T., PREUSSEL, K., HEMM, M.,
NIXDORF, B., KARSTEN, U. & WIEDNER, C. (2006). Distribution of
three alien cyanobacterial species (Nostocales) in northeast
Germany: Cylindrospermopsis raciborskii, Anabaena bergii and
Aphanizomenon aphanizomenoides. Phycologia, 45: 696–703.
TER BRAAK, C.J.F. (1986). Canonical correspondence analysis: a
new eigenvector technique for multivariate direct gradient ana-
lysis. Ecology, 67: 1167–1178.
TER BRAAK, C.J.F. & SMILAUER, P. (1998). Program CANOCO,
version 4.0. Centre for Biometry, Wageningen.
VAN LIERE, L. & MUR, L.R. (1980). Occurrence of Oscillatoria
agardhii and some related species, a survey. In Developments in
Hydrobiology, Vol. 2 (Barica, J. & Mur, L.E., editors), 67–77. Dr.
W. Junk BV, The Hague.
VITOUSEK, P.M. (1990). Biological invasions and ecosystem pro-
cess – towards an integration of population biology and ecosys-
tem studies. Oikos, 57: 7–13.
WALTHER, G.-R. (2003). Plants in a warmer world. Perspect. Plant
Ecol. Evol. System., 6: 169–185.
WELLS, M.J., POYNTON, A.A., BALSINHAS, K.J., MUSIL, H., VAN
HOEPEN, J.E. & ABBOTT, S.K. (1986). The history of introduction
of invasive alien plants to southern Africa. In The Ecology and
Management of Biological Invasions in Southern Africa
(Macdonald, A.W., Kruger, F.J. & Ferrar, A.A., editors),
21–35. Oxford University Press, Cape Town.
WETZEL, R.G. & LIKENS, G.F. (2000). Limnological Analyses. 3rd
ed. Springer-Verlag, New York.
WIEDNER, C., NIXDORF, B., HEINZE, R., WIRSING, B., NEUMANN, U.
& WECKESSER, J. (2002). Regulation of Cyanobacteria and micro-
cystin dynamics in polymictic shallow lakes. Arch. Hydrobiol.,
155: 383–400.
WIEDNER, C., RUCKER, J., BRUGGEMANN, R. & NIXDORF, B. (2007).
Climate change affects timing and size of populations of inva-
sive cyanobacterium in temperate regions. Oecologia, 152:
473–484.
M. Kokocinski et al. 374
Downloaded By: [Kokocinski, Mikolaj] At: 19:47 21 September 2010




















