
Mechanism of colonic permeation of inulin: Is rat colon more permeable than small intestine
Upload
independentCategory
view
0download
0
J. Comp. Path. 1999 Vol. 121, 249–263
The Distribution of Lymphocytes Expressing ab andcd T-cell Receptors, and the Expression of MucosalAddressin Cell Adhesion Molecule-1 in the Canine
Intestine
A. J. German∗†, E. J. Hall∗, P.F. Moore‡, D. J. Ringler§,W. Newman§ and M. J. Day†
∗ Department of Clinical Veterinary Science and † Department of Pathology and Microbiology,University of Bristol, Langford House, Langford, Bristol, BS40 5DU, UK, ‡Department of
Pathology, Microbiology and Immunology, School of Veterinary Medicine, University of California(Davis), CA 95616, USA and § Leukocyte Inc., 215 First Street, Cambridge, MA 02142, USA
Summary
The present study defined the distribution of ab and cd T-cell subsets, andthe expression of mucosal addressin cell adhesion molecule-1 (MAdCAM-1)in the intestinal tract of an outbred population of dogs with no evidence ofgastrointestinal disease. A panel of monoclonal antibodies reactive with thecanine ab and cd T-cell receptors (TCRs) and human MAdCAM-1 was usedin a series of immunoperoxidase or immunofluorescence studies. ab T cellswere predominant within the villous lamina propria with a significantlydecreasing linear trend from upper villus to crypt (P<0·0001). A proportionof intraepithelial lymphocytes (IELs) also expressed the ab T-cell receptor,with significantly greater numbers in villous than in crypt epithelium(P<0·0001). However, there were no significant differences in the numbersof either lamina propria or epithelial ab T cells between different intestinalregions. cd T cells were rare in the lamina propria, but a prominent cd IELpopulation was present and shown by double-colour immunofluorescencestudies to be principally of the CD4−CD8a− phenotype. MAdCAM-1 wasexpressed by endothelial cells in the mucosa, sub-mucosa and muscularislayers of all levels of the intestinal tract. In the mucosa, significantly moreMAdCAM-1 positive endothelium was present in regions of crypt than villouslamina propria (P<0·0001), but there were no significant differences betweenexpression in different intestinal regions. The quantitative and qualitativedata will enable comparisons of these parameters to be made with those indogs suffering from inflammatory bowel diseases.
1999 Harcourt Publishers Ltd
Introduction
The mucosal immune system comprises the largest population of lymphocytesin the body and is constantly exposed to a wide array of environmentalantigens. It must be capable of mounting protective immune responses topathogens and simultaneously remain tolerant to harmless environmentalantigens such as commensal bacteria and food. The systemic and mucosalimmune systems have many characteristics in common; there are, however,important differences, particularly with respect to the phenotype and function
0021–9975/99/070249+15 $12.00 1999 Harcourt Publishers Ltd

250 A. J. German et al.
of constituent lymphocytes and the recirculation pathways utilized by thesecells.
One unusual feature of the mucosal immune system is the presence of asubstantial population of T lymphocytes that express the cd rather than abT-cell receptor (TCR). Such cells may be extrathymically derived (Poussierand Julius, 1994) and, in the mouse, cd T cells have a restricted repertoire ofTCRs (Abreu-Martin and Targan, 1996). These cells have been shown toexpress constitutive cytotoxic activity (Goodman and Lefrancois, 1988, 1989)to produce a wide array of either Th1 or Th2 cytokines (Taguchi et al., 1990;Beagley et al., 1995), and to use alternative activation pathways (Trejdosiewicz,1992). It is thought that such cells provide an important rapid responsemechanism against pathogen invasion, representing a component of a moreprimitive immune system (Abreu-Martin and Targan, 1996). cd T cells havebeen documented in most species examined to date, including mice, humans,chickens, rats, sheep, cattle and pigs (Haas et al., 1993). Further, they haverecently been documented in both normal and diseased canine skin (Cannonet al., 1998) and may account for approximately 1–4% of the T cells inperipheral blood (Moore et al., 1998).
To mount an effective mucosal immune response, lymphocytes must “traffic”between the inductive (Peyer’s patch and mesenteric lymph node) and theeffector sites (lamina propria and intestinal epithelium) of the gastrointestinal-associated lymphoid tissue (GALT). Fundamental to this migration is theinteraction of the ligand a4b7 (expressed by lymphocytes of mucosal specificity)with mucosal addressin cell adhesion molecule-1 (MAdCAM-1) (Berlin et al.,1993), expressed by vascular endothelium in mucosal tissues (Streeter et al.,1988). The anatomical distribution of MAdCAM-1 has been well characterizedin man (Briskin et al., 1997), rodents (Streeter et al., 1988) and sheep (Mackayet al., 1996) but, to date, the distribution in the dog is not known.
Immunohistochemistry is a powerful means of assessing the distribution ofimmune cell populations (Elwood et al., 1997; German et al., 1999) andimmunologically relevant molecules (German et al., 1998) within the canineintestine, but application of the technique has been hampered by the limitedavailability of both specific and cross-reactive immunological reagents for thisspecies. Monoclonal antibodies (MAbs) have recently been developed to boththe ab and cd T-cell receptors (Moore et al., 1994, 1998) and used to definethe distributions of both subsets of T cell in both normal and diseased canineskin (Olivry et al., 1996; Moore et al., 1998). Furthermore, a panel of MAbsto human MAdCAM-1 has recently been developed (Briskin et al., 1997) buthas yet to be tested for cross-reactivity in other species. The aim of the presentstudy was to describe the qualitative and quantitative distribution of MAdCAM-1 and of ab and cd T cells within the canine intestine.
Materials and MethodsCollection and Processing of Material
Intestinal tissue was obtained from 14 dogs of various breeds submitted for routinepost-mortem examination (n=6) or humanely killed at an animal shelter (n=8)
251T-cell Subsets and MAdCAM-1 in the Canine Gut
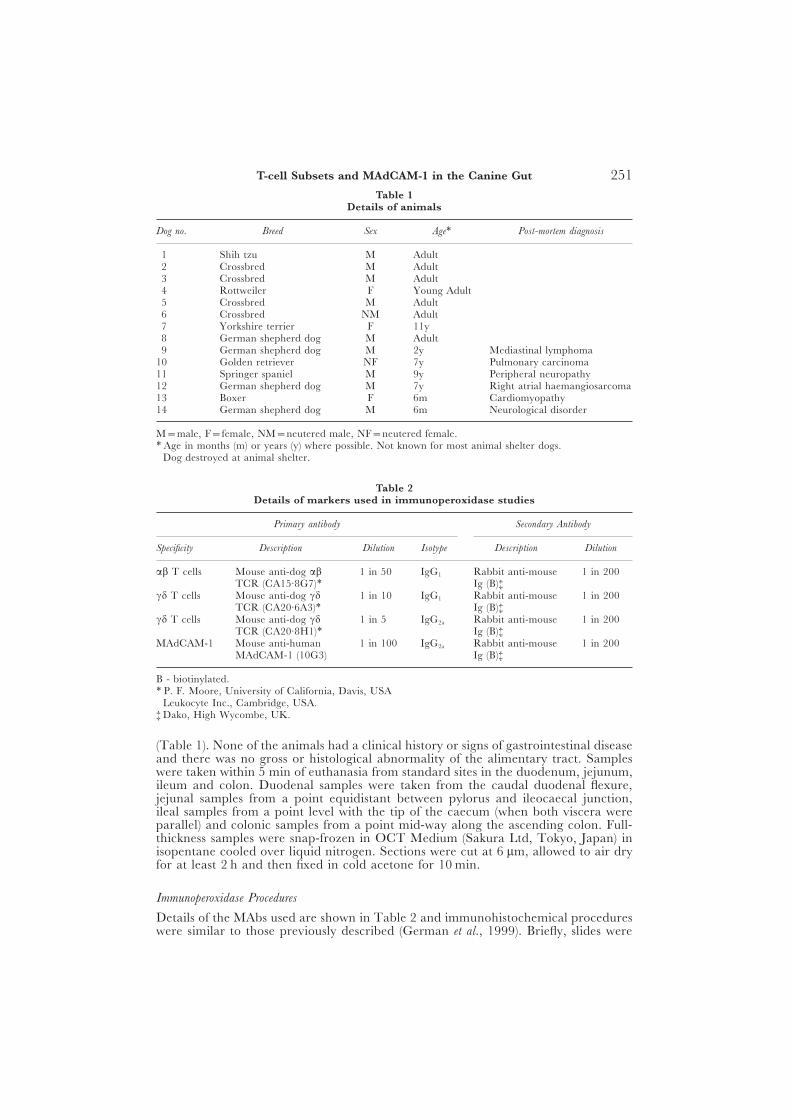
Table 1Details of animals
Dog no. Breed Sex Age∗ Post-mortem diagnosis
1 Shih tzu M Adult †2 Crossbred M Adult †3 Crossbred M Adult †4 Rottweiler F Young Adult †5 Crossbred M Adult †6 Crossbred NM Adult †7 Yorkshire terrier F 11y †8 German shepherd dog M Adult †9 German shepherd dog M 2y Mediastinal lymphoma
10 Golden retriever NF 7y Pulmonary carcinoma11 Springer spaniel M 9y Peripheral neuropathy12 German shepherd dog M 7y Right atrial haemangiosarcoma13 Boxer F 6m Cardiomyopathy14 German shepherd dog M 6m Neurological disorder
M=male, F=female, NM=neutered male, NF=neutered female.∗ Age in months (m) or years (y) where possible. Not known for most animal shelter dogs.† Dog destroyed at animal shelter.
Table 2Details of markers used in immunoperoxidase studies
Primary antibody Secondary Antibody
Specificity Description Dilution Isotype Description Dilution
ab T cells Mouse anti-dog ab 1 in 50 IgG1 Rabbit anti-mouse 1 in 200TCR (CA15·8G7)∗ Ig (B)‡
cd T cells Mouse anti-dog cd 1 in 10 IgG1 Rabbit anti-mouse 1 in 200TCR (CA20·6A3)∗ Ig (B)‡
cd T cells Mouse anti-dog cd 1 in 5 IgG2a Rabbit anti-mouse 1 in 200TCR (CA20·8H1)∗ Ig (B)‡
MAdCAM-1 Mouse anti-human 1 in 100 IgG2a Rabbit anti-mouse 1 in 200MAdCAM-1 (10G3)† Ig (B)‡
B - biotinylated.∗ P. F. Moore, University of California, Davis, USA† Leukocyte Inc., Cambridge, USA.‡ Dako, High Wycombe, UK.
(Table 1). None of the animals had a clinical history or signs of gastrointestinal diseaseand there was no gross or histological abnormality of the alimentary tract. Sampleswere taken within 5 min of euthanasia from standard sites in the duodenum, jejunum,ileum and colon. Duodenal samples were taken from the caudal duodenal flexure,jejunal samples from a point equidistant between pylorus and ileocaecal junction,ileal samples from a point level with the tip of the caecum (when both viscera wereparallel) and colonic samples from a point mid-way along the ascending colon. Full-thickness samples were snap-frozen in OCT Medium (Sakura Ltd, Tokyo, Japan) inisopentane cooled over liquid nitrogen. Sections were cut at 6 lm, allowed to air dryfor at least 2 h and then fixed in cold acetone for 10 min.
Immunoperoxidase Procedures
Details of the MAbs used are shown in Table 2 and immunohistochemical procedureswere similar to those previously described (German et al., 1999). Briefly, slides were
252 A. J. German et al.
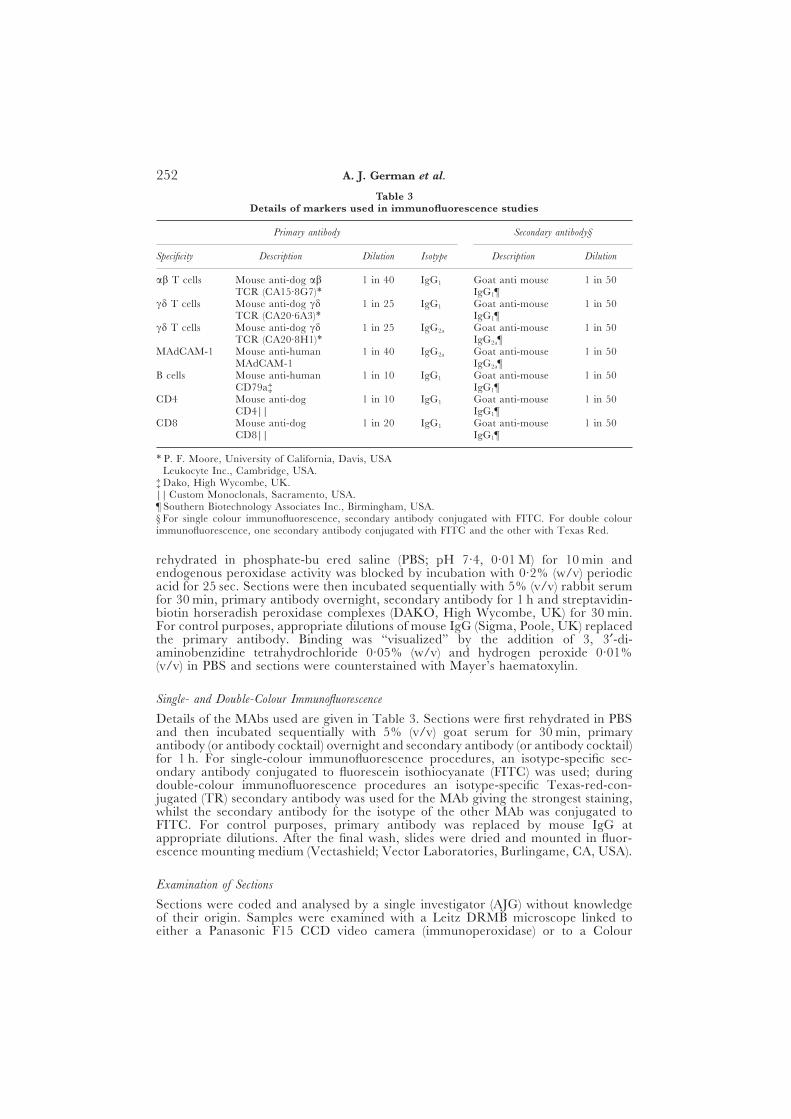
Table 3Details of markers used in immunofluorescence studies
Primary antibody Secondary antibody§
Specificity Description Dilution Isotype Description Dilution
ab T cells Mouse anti-dog ab 1 in 40 IgG1 Goat anti mouse 1 in 50TCR (CA15·8G7)∗ IgG1¶
cd T cells Mouse anti-dog cd 1 in 25 IgG1 Goat anti-mouse 1 in 50TCR (CA20·6A3)∗ IgG1¶
cd T cells Mouse anti-dog cd 1 in 25 IgG2a Goat anti-mouse 1 in 50TCR (CA20·8H1)∗ IgG2a¶
MAdCAM-1 Mouse anti-human 1 in 40 IgG2a Goat anti-mouse 1 in 50MAdCAM-1† IgG2a¶
B cells Mouse anti-human 1 in 10 IgG1 Goat anti-mouse 1 in 50CD79a‡ IgG1¶
CD4 Mouse anti-dog 1 in 10 IgG1 Goat anti-mouse 1 in 50CD4|| IgG1¶
CD8 Mouse anti-dog 1 in 20 IgG1 Goat anti-mouse 1 in 50CD8|| IgG1¶
∗ P. F. Moore, University of California, Davis, USA† Leukocyte Inc., Cambridge, USA.‡ Dako, High Wycombe, UK.|| Custom Monoclonals, Sacramento, USA.¶ Southern Biotechnology Associates Inc., Birmingham, USA.§ For single colour immunofluorescence, secondary antibody conjugated with FITC. For double colourimmunofluorescence, one secondary antibody conjugated with FITC and the other with Texas Red.
rehydrated in phosphate-buffered saline (PBS; pH 7·4, 0·01 M) for 10 min andendogenous peroxidase activity was blocked by incubation with 0·2% (w/v) periodicacid for 25 sec. Sections were then incubated sequentially with 5% (v/v) rabbit serumfor 30 min, primary antibody overnight, secondary antibody for 1 h and streptavidin-biotin horseradish peroxidase complexes (DAKO, High Wycombe, UK) for 30 min.For control purposes, appropriate dilutions of mouse IgG (Sigma, Poole, UK) replacedthe primary antibody. Binding was “visualized” by the addition of 3, 3′-di-aminobenzidine tetrahydrochloride 0·05% (w/v) and hydrogen peroxide 0·01%(v/v) in PBS and sections were counterstained with Mayer’s haematoxylin.
Single- and Double-Colour Immunofluorescence
Details of the MAbs used are given in Table 3. Sections were first rehydrated in PBSand then incubated sequentially with 5% (v/v) goat serum for 30 min, primaryantibody (or antibody cocktail) overnight and secondary antibody (or antibody cocktail)for 1 h. For single-colour immunofluorescence procedures, an isotype-specific sec-ondary antibody conjugated to fluorescein isothiocyanate (FITC) was used; duringdouble-colour immunofluorescence procedures an isotype-specific Texas-red-con-jugated (TR) secondary antibody was used for the MAb giving the strongest staining,whilst the secondary antibody for the isotype of the other MAb was conjugated toFITC. For control purposes, primary antibody was replaced by mouse IgG atappropriate dilutions. After the final wash, slides were dried and mounted in fluor-escence mounting medium (Vectashield; Vector Laboratories, Burlingame, CA, USA).
Examination of Sections
Sections were coded and analysed by a single investigator (AJG) without knowledgeof their origin. Samples were examined with a Leitz DRMB microscope linked toeither a Panasonic F15 CCD video camera (immunoperoxidase) or to a Colour

253T-cell Subsets and MAdCAM-1 in the Canine Gut
Fig. 1. A diagramatic representation of small intestinal and colonic lamina propria to demonstrate theareas used for ab T cell counts. Area 1, villous tip; area 2, villous base; area 3, base of crypt.
CoolView video camera (Photonic Science, Robertsbridge, UK) (immunofluorescence).For peroxidase sections, subjective analysis of the distribution of positive cells (ab-and cd-TCR MAb) or endothelial staining (MAdCAM-1 MAb), was initially madewith ×4·5, ×10 and ×40 objectives. Quantitative determinations included thenumber of ab T cells within the lamina propria, the number of ab and cd T cellswithin the epithelium, and the amount of MAdCAM-1-positive endothelium withinthe lamina propria.
The numbers of small intestinal lamina propria ab T cells were determined instandardized areas at the tip of the villus (area 1), base of the villus (area 2) and baseof the crypts (area 3) (Fig. 1), as previously described (German et al., 1999). Resultswere expressed as cells/10 000 lm2. Similar lamina propria cell counts were madefor the crypt areas of colonic sections (Fig. 1).
ab Intra-epithelial leucocytes (IELs) were assessed in immunoperoxidase sectionsby counting positive cells in four to six regions (each of 100–300 enterocytes) forboth villous and crypt epithelium (small intestine) and crypt epithelium (colon).Immunofluorescence sections were used to quantify cd T cells within villous epithelium.Images from small intestinal sections of seven representative dogs were transferred tocomputer with Image Pro-Plus software (Media Cybernetics, Maryland, USA). IELswere again counted relative to enterocyte numbers and the results expressed as positivecells/100 enterocytes.
For quantitative determination of MAdCAM-1 staining, ×10 objective images ofimmunoperoxidase sections were transferred to computer (as before) and the amountof positively stained endothelium determined with image analysis software (GaugeGem; ME Electronics, Reading, UK). Five appropriate areas of either villous (includingareas at both the tip and base) or crypt lamina propria were selected at random anddelineated as for the cell counting procedure. Areas of positively stained endothelium(excluding vessel lumen) were then outlined and results expressed as the proportionof MAdCAM-1-positive area to total lamina propria area.
254 A. J. German et al.
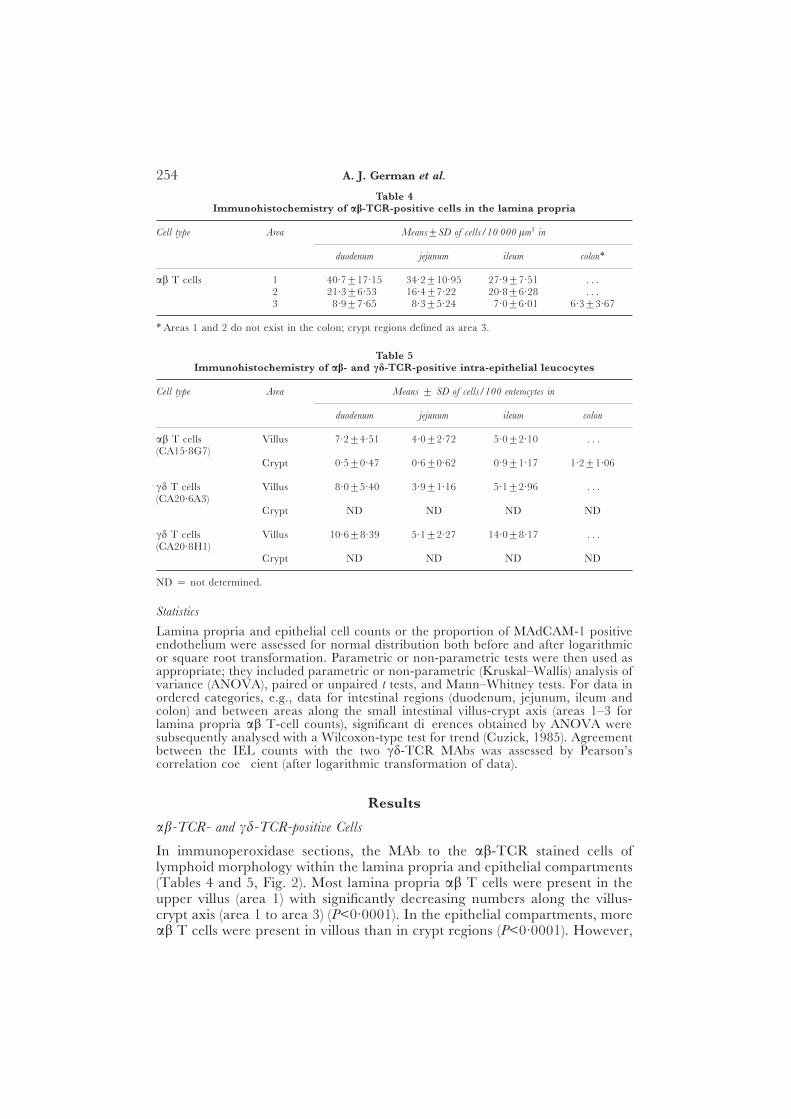
Table 4Immunohistochemistry of ab-TCR-positive cells in the lamina propria
Cell type Area Means±SD of cells/10 000 lm2 in
duodenum jejunum ileum colon∗
ab T cells 1 40·7±17·15 34·2±10·95 27·9±7·51 . . .2 21·3±6·53 16·4±7·22 20·8±6·28 . . .3 8·9±7·65 8·3±5·24 7·0±6·01 6·3±3·67
∗ Areas 1 and 2 do not exist in the colon; crypt regions defined as area 3.
Table 5Immunohistochemistry of ab- and cd-TCR-positive intra-epithelial leucocytes
Cell type Area Means ± SD of cells/100 enterocytes in
duodenum jejunum ileum colon
ab T cells Villus 7·2±4·51 4·0±2·72 5·0±2·10 . . .(CA15·8G7)
Crypt 0·5±0·47 0·6±0·62 0·9±1·17 1·2±1·06
cd T cells Villus 8·0±5·40 3·9±1·16 5·1±2·96 . . .(CA20·6A3)
Crypt ND ND ND ND
cd T cells Villus 10·6±8·39 5·1±2·27 14·0±8·17 . . .(CA20·8H1)
Crypt ND ND ND ND
ND = not determined.
Statistics
Lamina propria and epithelial cell counts or the proportion of MAdCAM-1 positiveendothelium were assessed for normal distribution both before and after logarithmicor square root transformation. Parametric or non-parametric tests were then used asappropriate; they included parametric or non-parametric (Kruskal–Wallis) analysis ofvariance (ANOVA), paired or unpaired t tests, and Mann–Whitney tests. For data inordered categories, e.g., data for intestinal regions (duodenum, jejunum, ileum andcolon) and between areas along the small intestinal villus-crypt axis (areas 1–3 forlamina propria ab T-cell counts), significant differences obtained by ANOVA weresubsequently analysed with a Wilcoxon-type test for trend (Cuzick, 1985). Agreementbetween the IEL counts with the two cd-TCR MAbs was assessed by Pearson’scorrelation coefficient (after logarithmic transformation of data).
Results
ab-TCR- and cd-TCR-positive Cells
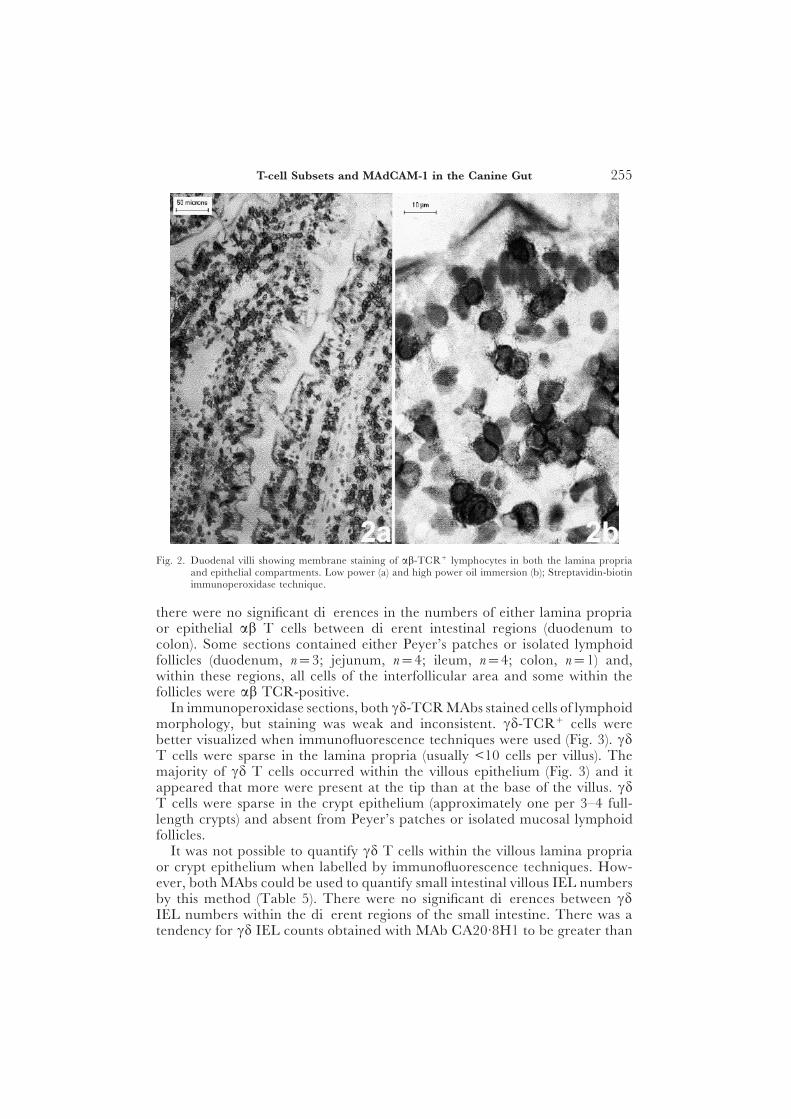
In immunoperoxidase sections, the MAb to the ab-TCR stained cells oflymphoid morphology within the lamina propria and epithelial compartments(Tables 4 and 5, Fig. 2). Most lamina propria ab T cells were present in theupper villus (area 1) with significantly decreasing numbers along the villus-crypt axis (area 1 to area 3) (P<0·0001). In the epithelial compartments, moreab T cells were present in villous than in crypt regions (P<0·0001). However,
255T-cell Subsets and MAdCAM-1 in the Canine Gut
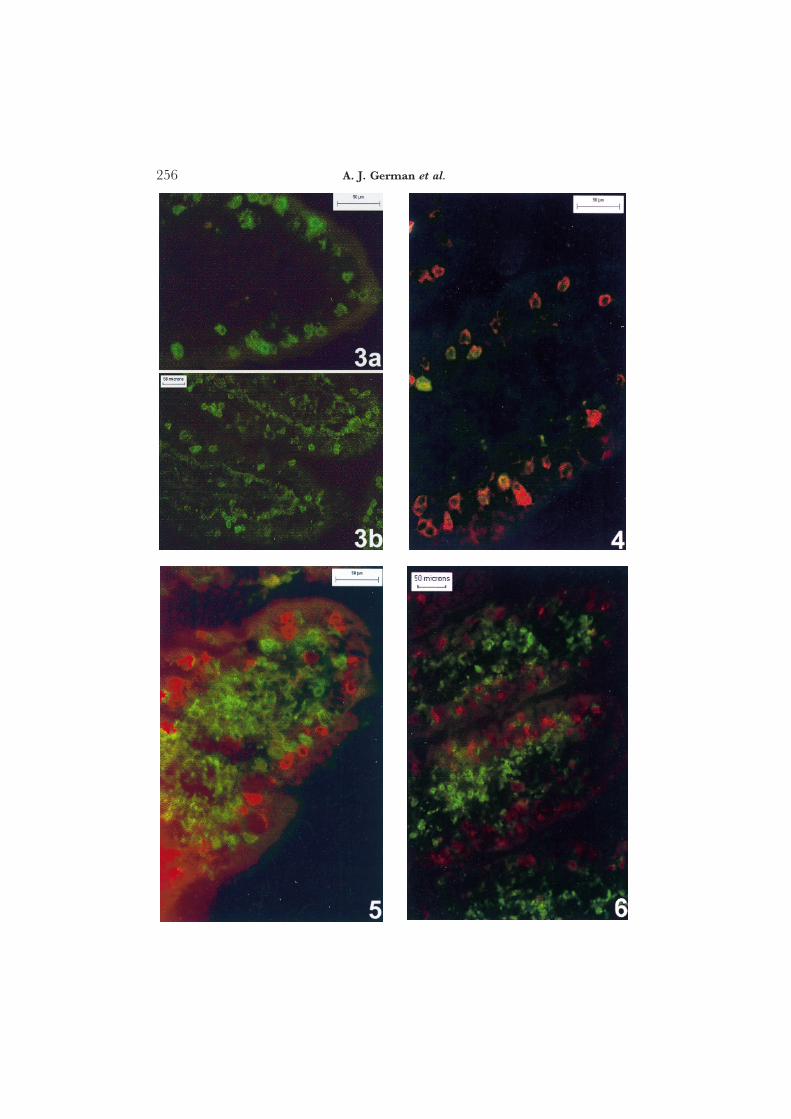
Fig. 2. Duodenal villi showing membrane staining of ab-TCR+ lymphocytes in both the lamina propriaand epithelial compartments. Low power (a) and high power oil immersion (b); Streptavidin-biotinimmunoperoxidase technique.
there were no significant differences in the numbers of either lamina propriaor epithelial ab T cells between different intestinal regions (duodenum tocolon). Some sections contained either Peyer’s patches or isolated lymphoidfollicles (duodenum, n=3; jejunum, n=4; ileum, n=4; colon, n=1) and,within these regions, all cells of the interfollicular area and some within thefollicles were ab TCR-positive.
In immunoperoxidase sections, both cd-TCR MAbs stained cells of lymphoidmorphology, but staining was weak and inconsistent. cd-TCR+ cells werebetter visualized when immunofluorescence techniques were used (Fig. 3). cdT cells were sparse in the lamina propria (usually <10 cells per villus). Themajority of cd T cells occurred within the villous epithelium (Fig. 3) and itappeared that more were present at the tip than at the base of the villus. cdT cells were sparse in the crypt epithelium (approximately one per 3–4 full-length crypts) and absent from Peyer’s patches or isolated mucosal lymphoidfollicles.
It was not possible to quantify cd T cells within the villous lamina propriaor crypt epithelium when labelled by immunofluorescence techniques. How-ever, both MAbs could be used to quantify small intestinal villous IEL numbersby this method (Table 5). There were no significant differences between cdIEL numbers within the different regions of the small intestine. There was atendency for cd IEL counts obtained with MAb CA20·8H1 to be greater than
256 A. J. German et al.

257T-cell Subsets and MAdCAM-1 in the Canine Gut
when MAb CA20·6A3 was used, but the difference was not statisticallysignificant (P=0·45) and good correlation was demonstrated between IELcounts determined with the two MAbs (Rp=0·829, P<0·02). Further, mostpositive cells were “dual positive” when double-colour immunofluorescencewas used (Fig. 4), contrasting with the “single-positive” cells seen when ab-and cd-TCR MAbs (CA15·8G7 and CA20·8H1, respectively) were usedsimultaneously (Fig. 5).
The phenotype of cd IELs was examined further by double-colour immuno-fluorescence. Sections from all parts of the intestine from representative dogswere co-stained with MAb CA20·8H1 and either anti-dog CD4 or anti-dog-CD8 a chain. CD4+ cells predominated in the lamina propria rather than inthe epithelium, and CD4 was not expressed by any of the cd-TCR+ cells.CD8a+ cells were present within both lamina propria and epithelial com-partments, but this marker did not co-stain the majority of cd TCR+ cells(Fig. 6).
MAdCAM-1 Endothelial Staining
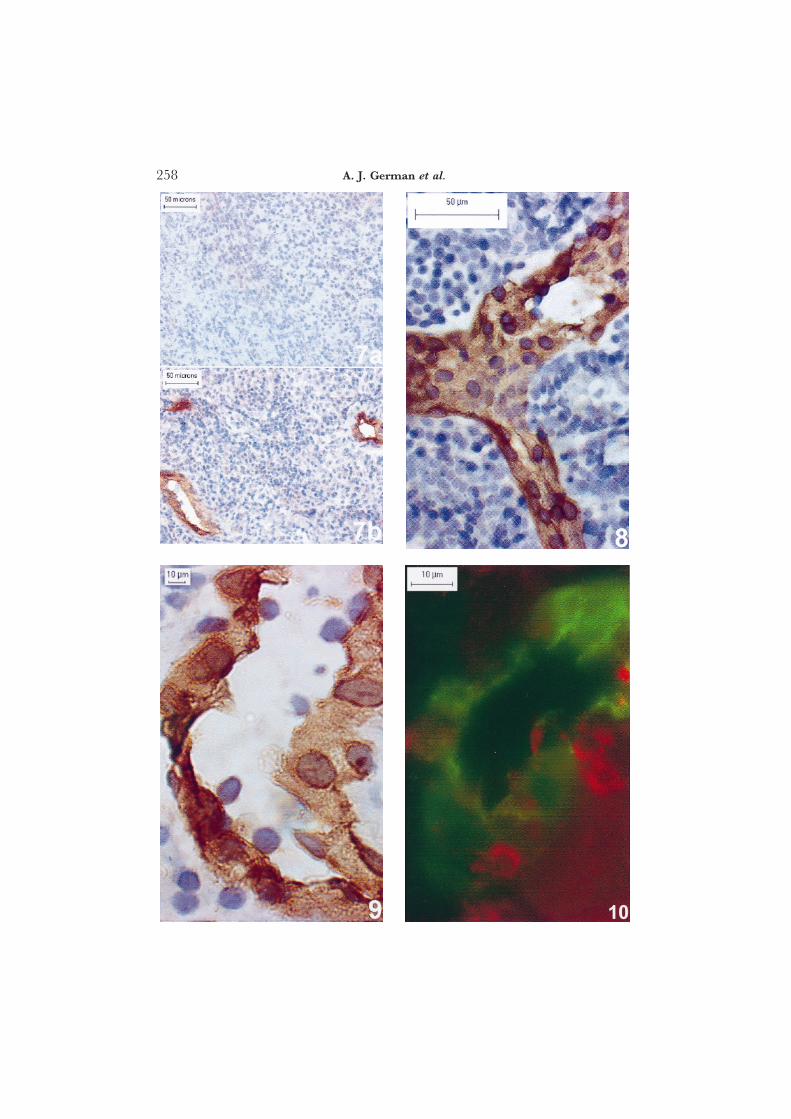
In preliminary studies, a panel of three anti-human MAdCAM-1 antibodies(Briskin et al., 1997) was tested for cross-reactivity with canine intestinal tissues;only MAb 10G3 showed positive reactivity and further studies were thereforeconducted with this MAb alone. In subsequent studies the specificity of theMAb 10G3 for the GALT was examined. Samples of tissue, collected fromthree dogs, included duodenum (n=2), colon (n=2), skin (n=2), muscle (n=2), liver (n=1), spleen (n=1), mesenteric lymph node (n=2), popliteal lymphnode (n=1), and prescapular lymph node (n=2). Duodenum, colon andmesenteric lymph node contained MAdCAM-1 positive endothelium, but allother tissues were negative (Fig. 7).
The distribution of MAdCAM-1 was examined in sections of duodenum,jejunum, ileum and colon from 14 dogs without intestinal disease (Figs 8 and9). MAdCAM-1 staining, which was present on endothelial cells in all regionsof the intestinal tract (duodenum to colon), was seen in the lamina propria,sub-mucosa and muscularis. Further, MAdCAM-1+ endothelium could befound in close proximity to, or within, Peyer’s patches and isolated lymphoidfollicles. MAdCAM-1+ endothelium was often cuboidal and lymphocytes were
Fig. 3. Single-colour immunofluorescence of duodenal villous tip showing cd-TCR+ lymphocytes withinepithelial compartment. Occasional positive cells are also seen within the lamina propria. (a) MAbCA20·6A3, (b) MAb CA20·8H1.
Fig. 4 Double-colour immunofluorescence showing duodenal villous tip stained with MAbs CA20·6A3(FITC) and CA20·8H1 (Texas red) to canine cd-TCR. The majority of cells are within the epithelialcompartment and are dual positive.
Fig. 5. Double-colour immunofluorescence showing duodenal villous tip stained with MAbs to ab-TCR(CA15·8G7, FITC) and to cd-TCR (CA20·8H1, Texas red) All cells are single positive, with ab Tcells predominating in the lamina propria; a mixture of ab and cd T cells is present within theepithelium.
Fig. 6. Double-colour immunofluorescence showing duodenal villous tip stained with MAbs to CD8a chain(FITC) and to cd-TCR (CA20·8H1, Texas red) Most cells are single positive, but occasional cellsare dual positive.
258 A. J. German et al.

259T-cell Subsets and MAdCAM-1 in the Canine Gut
Table 6Immunohistochemistry of MAdCAM-1-positive endothelium within the lamina propria
Immunohistochemical Area Means ± SD of MAdCAM-1+ endothelium/lamina propria area inmarker
duodenum jejunum ileum colon∗
MAdCAM-1 Villus 0·04±0·025 0·04±0·046 0·03±0·021 . . .Crypt 0·15±0·047 0·15±0·052 0·16±0·050 0·12±0·044
∗ Areas 1 and 2 do not exist in the colon; crypt regions defined as area 3.
seen in close apposition to their luminal surface (Fig. 9). Double-colourimmunofluorescence procedures showed that such cells were most commonlyab TCR+ cells (Fig. 10) or B cells (CD79a+) (not shown); some were alsopositive for CD4 (not shown).
Computer-aided morphometry was used to quantify the amount of MAd-CAM-1 positive endothelium within the villous and crypt lamina propria(Table 6). Significantly more MAdCAM-1 staining was found in the cryptthan villous areas (P<0·0001) but there were no significant differences betweenthe regions of the intestine (P=0·84).
Discussion
In the present study, immunohistochemical markers were used to determinethe distribution of cells expressing different TCRs and the expression ofMAdCAM-1 in the canine intestinal tract. Immunofluorescence techniqueswere preferred to immunoperoxidase procedures for the cd-TCR MAbs, sincestaining with the latter proved unreliable. The reasons for this were unclearbut may have been associated with the use of periodic acid to block endogenousperoxidase, which may have destroyed the antigenic epitopes bound by theseMAbs. Alternative peroxidase-blocking procedures, such as the use of H2O2
in 50% methanol, might have been more appropriate, but these have giveninferior staining when used in conjunction with other MAbs (unpublishedobservations). Alternatively, it is possible that some MAbs are best suited toperoxidase procedures, but that others give stronger staining when fluorescencetechniques are used. In either case, thorough standardization is required whendetermining the optimum immunohistochemical procedure for any MAb.
ab and cd T lymphocytes were present in both the intestinal laminapropria and epithelium. ab T cells predominated in the lamina propria, theirdistribution and number being similar to those of both CD3+ and CD5+
Fig. 7. Sections of canine prescapular (a) and mesenteric (b) lymph node stained with anti-human MAdCAM-1 MAb 10G3. MAdCAM-1+ endothelium is present only within mesenteric lymph node. Streptavidin-biotin immunoperoxidase technique.
Fig. 8. Jejunal crypt regions stained with anti-human MAdCAM-1 MAb and demonstrating MAdCAM-1positive endothelium. Streptavidin-biotin peroxidase technique.
Fig 9. High power oil immersion photomicrograph showing MAdCAM-1+ endothelium with lymphocytesin close apposition to the luminal surface. Streptavidin-biotin peroxidase technique.
Fig. 10. Double-colour immunofluorescence of jejunal crypts stained with MAb to MAdCAM-1 (FITC)and ab-TCR (Texas red). ab-T cells can be seen in close apposition to MAdCAM-1+ endothelium.

260 A. J. German et al.
lamina propria cells documented in previous work that included intestinalsections from most of the dogs of the present study (German et al., 1999). Thisfinding suggests that most intestinal lamina propria T cells in the dog use theab-TCR, consistent with TCR usage by lamina propria T lymphocytes inother species (Abreu-Martin and Targan, 1996).
Positively stained cells in the intestinal epithelial compartment were countedrelative to the numbers of enterocytes. Similar numbers of ab and cd T cellswere noted in villous epithelium, but direct comparisons should be madecautiously, given that enumeration was performed on peroxidase- and fluor-escence-stained sections, respectively. However, the finding suggests that, aswith other species, the IEL population in the dog is heterogeneous (Abreu-Martin and Targan, 1996). Further, it would appear that the cd IEL populationin the dog is larger than that documented in man, but of a size equivalent tothat of the mouse (Poussier and Julius, 1994). A previous study demonstratedthe distribution of CD3+T lymphocytes within the intraepithelial compartmentof the canine intestine (German et al., 1999). The number of CD3+ IELsexceeds that of either ab or cd IELs alone, but when the numbers of ab andcd IELs are combined the total exceeds the CD3+ IEL count. The first ofthese observations implies that canine cd IELs express CD3, as reported inother species (Abreu-Martin and Targan, 1996). The second observationrepresents an apparent discrepancy that might be explained by the fact thatthe CD3+ counts in the previous study were performed on sections (4 lm) offormalin-fixed tissue, not on sections (6 lm) of frozen tissue as in the presentstudy. Given that the staining efficacy of different MAbs may vary, directcomparisons between absolute numbers of cells stained by different markersshould be interpreted with caution. It is also noted that the number of CD5+
IELs previously determined (German et al., 1999) exceeds that of either ab orcd IELs. This suggests that canine cd IELs express the CD5 molecule, anobservation that contrasts with findings in other species (Abreu-Martin andTargan, 1996).
As mentioned above, when cd IEL counts were performed on tissues sectionslabelled by the two different MAbs (CA20·6A3 and CA20·8H1), those obtainedwith the latter MAb tended to be greater, although the difference was notstatistically significant. The difference, if real, may have reflected non-specificbinding of MAb CA20·8H1 to another cell population, for example byinteraction with Fc receptors. Against this, our negative control slides did notreveal any non-specific binding, and therefore the observed difference maysimply have reflected variability in the cell counting technique.
Double-colour immunofluorescence was used to characterize further thephenotype of canine intestinal cd T cells, and the results suggested that fewof these cells expressed either CD4 or CD8 cell surface molecules. Thiscontrasts with the situation in other species, in which a large proportion cdIELs express an unusual CD8 molecule, formed by the homodimerization oftwo CD8 a chains (Lefrancois, 1991). This CD8aa homodimer has also beendocumented on ab IELs and its presence provides circumstantial evidence foran extrathymic pathway of development for both ab and cd T cells withinthe intestinal epithelium (Poussier et al., 1992; Poussier and Julius, 1994).

261T-cell Subsets and MAdCAM-1 in the Canine Gut
This study also demonstrated the distribution of MAdCAM-1 in variouscanine tissues and showed that expression was restricted to endothelial cellsin the GALT, including Peyer’s patches, mesenteric lymph nodes and theintestinal mucosa, submucosa and muscularis. This distribution was similarto that recorded in other species, including mice (Streeter et al., 1988), man(Briskin et al., 1997) and sheep (Mackay et al., 1996), but there was no stainingof follicular dendritic cells as previously reported in mice (Szabo et al., 1997).MAdCAM-1 expression is also present on endothelium in murine mammarytissue but absent from the endothelium of other mucosal sites including oraltissues, lung and uterus (Streeter et al., 1988). In future studies we hope todetermine whether MAdCAM-1 expression is increased in canine inflammatorybowel disease (IBD), as reported in both human IBD (Briskin et al., 1997) andmurine models of intestinal inflammation (Picarella et al., 1997).
The process by which lymphocytes migrate from the circulation into tissueshas been well documented (Springer, 1994). Fundamental to this process arethe interactions between the “homing receptors” (selections and integrins) onthe lymphocyte cell surface and “vascular addressins” (selections, mucin-like molecules, chemoattractant receptors and integrins) on the vascularendothelium (Springer, 1994). These molecules interact in a systematic wayto allow lymphocytes sequentially to roll, attach and extravasate throughthe endothelium. In the present study, immunoperoxidase-stained sectionsdemonstrated circulating lymphocytes in close apposition to MAdCAM-1-positive cuboidal endothelium, suggesting that these cells were in the processof rolling, adhering or extravasating through high endothelial venules. Further,double-colour immunofluorescence revealed that these cells were a mixtureof B cells, ab-TCR-positive T cells and CD4+ T cells, and it has been shownin man that subsets of both B cells and CD4+ T cells express the a4b7 integrinthat binds preferentially to MAdCAM-1 (Rott et al., 1996). The presence ofa4b7 on the surface of canine lymphocytes has yet to be demonstrated. Finally,most of the endothelium expressing MAdCAM-1 in the lamina propria ispresent in crypt rather than villous regions; most mucosal T cells, however,are present within the villus. This suggests that, after extravasation, T cellsmigrate from crypt areas towards the tips of the villi, perhaps in response tolocalized production of chemokines or cytokines.
In summary, this study demonstrated the distribution of MAdCAM-1 and ofab and cd T lymphocytes in the intestinal tract of dogs without enteric disease.Future comparisons with dogs suffering from inflammatory bowel diseases maydetermine whether intestinal inflammation is associated with any alterationin the proportions of T-cell subsets, or in the expression of MAdCAM-1.
AcknowledgmentsThis study was funded by the Kennel Club Charitable Trust, and Biotechnology andBiological Sciences Research Council grant S09425.
ReferencesAbreu-Martin, M. T. and Targan, S. R. (1996). Regulation of immune responses in
the intestinal mucosa. Critical Reviews in Immunology, 16, 277–309.

262 A. J. German et al.
Beagley, K. W., Fujihashi, K., Lagoo, A. S., Lagoodeenadaylan, S., Black, C. A.,Murray, A. M., Sharmanov, A. T., Yamamoto, M., McGhee, J. R., Elson, C.O. and Kiyono, H. (1995). Differences in intraepithelial lymphocyte T cell subsetsisolated from murine small intestine versus large intestine. Journal of Immunology,154, 5611–5619.
Berlin, C., Berg, E. L., Briskin, M.J., Andrew, D. P., Kilshaw, P. J., Holzmann, B.,Weissman, I. L., Hamann, A and Butcher, E. C. (1993). a4b7 Integrin mediateslymphocyte binding to the mucosal vascular addressin MAdCAM-1. Cell, 74,185–195.
Briskin, M., Winsor-Hines, D., Shyjan, A., Cochran, N., Bloom, S., Wilson, J.,McEvoy, L. M., Butcher, E. C., Kassam, N., Mackay, C. R., Newman, W. andRingler, D. J. (1997). Human mucosal addressin cell adhesion molecule-1 ispreferentially expressed in intestinal tract and associated lymphoid tissue. AmericanJournal of Pathology, 151, 97–110.
Cannon, A. G., Olivry, T., Ihrke, P. J., Naydan, D. K. and Moore, P. F. (1998).Gamma delta T cells in normal and diseased canine skin. In: Advances in VeterinaryDermatology, Vol. 3, K. W. Kwochka, T. Willemse and C. von Tscharner, Eds,Butterworth Heinemann, Oxford, pp. 137–143.
Cuzick, J. (1985). A Wilcoxon-type test for trend. Statistics in Medicine, 4, 87–90.Elwood, C. M., Hamblin, A. S. and Batt, R. M. (1997). Quantitative and qualitative
immunohistochemistry of T cell subsets and MHC class II expression in thecanine intestine. Veterinary Immunology and Immunopathology, 58, 195–207.
German, A. J., Bland, P. W., Hall, M. J. and Day, M. J. (1998). Expression of majorhistocompatibility complex class II antigens in the canine intestine. VeterinaryImmunology and Immunopathology, 61, 171–180.
German, A. J., Hall, E. J. and Day, E. J. (1999). Analysis of leucocyte subsets in thecanine intestine. Journal of Comparative Pathology, 120, 129–145.
Goodman, T. and Lefrancois, L. (1988). Expression of the gamma-delta T-cell receptoron intestinal CD8+ intraepithelial lymphocytes. Nature, 333, 855–858.
Goodman, T. and Lefrancois, L. (1989). Intraepithelial lymphocytes. Anatomical site,not T cell receptor form dictates phenotype and function. Journal of ExperimentalMedicine, 170, 1569–1581.
Haas, W., Pereira, P. and Tonegawa, S. (1993). Gamma/delta cells. Annual Review ofImmunology, 11, 637–685.
Lefrancois, L. (1991). Phenotypic complexity of intraepithelial lymphocytes of thesmall intestine. Journal of Immunology, 147, 1746–1751.
Mackay, C. R., Andrew, D. P., Briskin, M., Ringler, D. J. and Butcher, E. C. (1996).Phenotype and migration of three major subsets of tissue homing T cells in sheep.European Journal of Immunology, 26, 2433–2439.
Moore, P. F., Affolter, V. K., Olivry, T. and Schrenzel, M. D. (1998). The use ofimmunological reagents in defining the pathogenesis of canine skin diseasesinvolving proliferation of leukocytes. In: Advances in Veterinary Dermatology, Vol. 3, K.W. Kwochka, T. Willemse and C. von Tscharner, Eds, Butterworth Heinemann,Oxford, pp. 78–94.
Moore, P. F., Rossitto, P. V. and Olivry, T. (1994). Development of monoclonalantibodies to canine T cell receptor-1 (TCR-cd) and their utilization in thediagnosis of epidermotropic cutaneous T cell lymphoma. Veterinary Pathology, 31,597.
Olivry, T., Moore, P. F., Affolter, V. K. and Naydan, D. K. (1996). Langerhans cellhyperplasia and IgE expression in canine atopic dermatitis. Archives of DermatologicalResearch, 288, 579–585.
Picarella, D., Hurlbut, P., Rottman, J., Shi, X., Butcher, E. J. and Ringler, D. J.(1997). Monoclonal antibodies specific for b7-integrin and mucosal addressin celladhesion molecule-1 (MAdCAM-1) reduce inflammation in the colon of scidmice reconstituted with CD45RBhighCD4+ T cells. Journal of Immunology, 158,2099–2106.

263T-cell Subsets and MAdCAM-1 in the Canine Gut
Poussier, P., Edouard, P., Lee, C., Binnie, M. and Julius, M. (1992). Thymus-independent development and negative selection of T cells expressing T cellreceptor a/b in the intestinal epithelium: evidence for distinct circulation patternsof gut- and thymus-derived T lymphocytes. Journal of Experimental Medicine, 176,187–199.
Poussier, P. and Julius, M. (1994). Thymus-independent T cell development andselection in the intestinal epithelium. Annual Review of Immunology, 12, 521–553.
Rott, L. S., Briskin, M. J., Andrew, D. P., Berg, E. L. and Butcher, E. C. (1996). Afundamental subdivision of circulating lymphocytes defined by adhesion tomucosal addressin cell adhesion molecule-1. Comparison with vascular celladhesion molecule-1 and correlation with b7 integrins and memory differentiation.Journal of Immunology, 156, 3727–3736.
Springer, T. A. (1994). Traffic signals for lymphocyte recirculation and leukocyteemigration: the multistep paradigm. Cell, 76, 301–314.
Streeter, T. A., Lakey-Berg, E., Rouse, B. T. N., Bargatze, R. F. and Butcher, E. C.(1988). A tissue-specific endothelial cell molecule involved in lymphocyte homing.Nature, 331, 41–46.
Szabo, M. C., Butcher, E. C. and McEvoy, L. M. (1997). Specialization of mucosalfollicular dendritic cells revealed by mucosal addressin-cell adhesion molecule-1display. Journal of Immunology, 158, 5584–5588.
Taguchi, T., McGhee, J. R., Coffman, R. L., Beagley, K. W., Eldridge, J. H., Takatsu,K. and Kiyono, H. (1990). Analysis of Th1 and Th2 cells in murine gut-associatedtissues. Frequencies of CD4+ and CD8+ T cells that secrete IFN- gamma andIL-5. Journal of Immunology, 145, 68–77.
Trejdosiewicz, L. K. (1992). Intestinal intraepithelial lymphocytes and lympho-epithelial interactions in the human gastrointestinal mucosa. Immunology Letters,32, 13–20.
CReceived, March 8th, 1999Accepted, May 17th, 1999D
Copyright © 2022 FDOKUMEN




















