
The defensive secretion of Carabus lefebvrei Dejean 1826 pupa (Coleoptera, Carabidae): Gland...
11
The Defensive Secretion of Carabus lefebvrei Dejean 1826 Pupa (Coleoptera, Carabidae): Gland Ultrastructure and Chemical Identification ANITA GIGLIO, 1 * PIETRO BRANDMAYR, 1 RENATO DALPOZZO, 2 GIOVANNI SINDONA, 2 ANTONIO TAGARELLI, 2 FEDERICA TALARICO, 1 TULLIA ZETTO BRANDMAYR, 1 AND ENRICO A. FERRERO 3 1 Department of Ecology, University of Calabria, I-87036 Arcavacata di Rende (CS), Calabria, Italy 2 Department of Chemistry, University of Calabria , I-87036 Arcavacata di Rende (CS), Calabria, Italy 3 Department of Life Sciences, University of Trieste, I-34127 Trieste, Calabria, Italy KEY WORDS abdominal glands; ultrastructure; chemical defense; monoterpenes ABSTRACT This study documents the defensive function of flavored humor secreted by the ab- dominal glands of Carabus lefebvrei pupae. The morphology and the ultrastructure of these glands were described and the volatile compounds of glands secretion were identified by gas chromatogra- phy/mass spectrometry. The ultrastructure analysis shows an acinose complex formed by about 50 clusters. Each cluster has 20 glandular units and the unit—composed of one secretory and one canal cell lying along a duct—belongs to the class 3 cell type of Quennedey (1998). In the cyto- plasm, the secretory cell contains abundant rough endoplasmatic reticula, glycogen granules, numerous mitochondria, and many well-developed Golgi complexes producing electron-dense se- cretory granules. Mitochondria are large, elongated, and often adjoining electronlucent vesicles. The kind and the origin of secretory granules varying in size and density were discussed. The chemical analysis of the gland secretion revealed the presence of a mixture of low molecular weight terpenes, ketones, aldehydes, alcohols, esters, and carboxylic acids. Monoterpenes, especially linal- ool, were the major products. We supposed that ketones, aldehydes, alcohols, esters, and carboxylic acids have a deterrent function against the predators and monoterpenes provide a prophylaxis function against pathogens. Microsc. Res. Tech. 72:351–361, 2009. V V C 2008 Wiley-Liss, Inc. INTRODUCTION A wide variety of insect species have evolved chemi- cal defense mechanisms—involved in interspecific interactions—that contributed to their widespread suc- cess in ecosystems (Thiele, 1977). In general, insect de- fensive substances are relatively low in molecular weight and are highly volatile, such as alkanes, ter- penes, alcohols, ketones, esters, aldehydes, organic acids, phenols, and quinones. They are secreted by exo- crine glands as blends and have repellent, toxicant, or antimicrobic function. The products of exocrine glands are well known in Isoptera, Thysanoptera, Hymenop- tera, Hemiptera, Lepidoptera, and Coleoptera (Burse et al., 2007; Chapman, 1998; Dettner, 1987; Whitman et al., 1990). According to our current knowledge, the order Coleoptera shows the largest number of species with the greatest variety of defensive glands. Allo- mones are extensively investigated in coleopteran adult beetles, their larvae and eggs but no attention has been paid to the pupae (Chapman, 1998; Dettner, 1987). Defensive chemicals have a high adaptive value in the pupal stage because they do not show antipreda- tion strategies such as behavior or structural modifica- tions to protect themselves against natural enemies. The ground beetles (Coleoptera, Carabidae) make up the largest adephagan family with about 40,000 species and their distribution is cosmopolitan. They are well known as bioindicators and natural enemies of agricul- tural pests. Because of their bio-indication and econom- ical importance, the role of biotic and abiotic factors in carabid population dynamics has been studied exten- sively. Abiotic factors and predation represent the main causes of mortality for all life-cycle stages of cara- bid beetles; pathogens and parasites can be important for some developmental stages (Lo ¨vei and Sunderland, 1996). Therefore, in response to predation pressure they have evolved morphological, behavioral, and chemical mechanisms against predation. Carabid bee- tles produce a large variety of defensive allomones (Andrew et al., 2000; Moore, 1979). Although exocrine glands (pygidial glands) and their defensive secretion were well investigated in adults, no information is available about larvae and pupae which are the most vulnerable stages of life cycle. This study is focused on gland secretion at the pupal stage of carabid beetle Carabus lefebvrei Dejean, 1826. We chose this species because it is easy to rear under laboratory conditions and easy to manipulate during experimental protocols. C. lefebvrei is a snail eating predatory Italian endemic species that lives in the beech, oak, chestnut, and pine forests of the Central and Southern Apennines, from lower altitudes to about *Correspondence to: Anita Giglio, Department of Ecology, University of Cala- bria, via P. Bucci cubo 4B, I-87036 Arcavacata di Rende (CS), Calabria, Italy. E-mail: [email protected] Received 10 July 2008; accepted in revised form 18 October 2008 DOI 10.1002/jemt.20660 Published online 9 December 2008 in Wiley InterScience (www.interscience. wiley.com). V V C 2008 WILEY-LISS, INC. MICROSCOPY RESEARCH AND TECHNIQUE 72:351–361 (2009)
Transcript of The defensive secretion of Carabus lefebvrei Dejean 1826 pupa (Coleoptera, Carabidae): Gland...

The Defensive Secretion of Carabus lefebvrei Dejean 1826Pupa (Coleoptera, Carabidae): Gland Ultrastructure andChemical IdentificationANITA GIGLIO,1* PIETRO BRANDMAYR,1 RENATO DALPOZZO,2 GIOVANNI SINDONA,2
ANTONIO TAGARELLI,2 FEDERICA TALARICO,1 TULLIA ZETTO BRANDMAYR,1 AND ENRICO A. FERRERO3
1Department of Ecology, University of Calabria, I-87036 Arcavacata di Rende (CS), Calabria, Italy2Department of Chemistry, University of Calabria , I-87036 Arcavacata di Rende (CS), Calabria, Italy3Department of Life Sciences, University of Trieste, I-34127 Trieste, Calabria, Italy
KEY WORDS abdominal glands; ultrastructure; chemical defense; monoterpenes
ABSTRACT This study documents the defensive function of flavored humor secreted by the ab-dominal glands of Carabus lefebvrei pupae. The morphology and the ultrastructure of these glandswere described and the volatile compounds of glands secretion were identified by gas chromatogra-phy/mass spectrometry. The ultrastructure analysis shows an acinose complex formed by about50 clusters. Each cluster has 20 glandular units and the unit—composed of one secretory and onecanal cell lying along a duct—belongs to the class 3 cell type of Quennedey (1998). In the cyto-plasm, the secretory cell contains abundant rough endoplasmatic reticula, glycogen granules,numerous mitochondria, and many well-developed Golgi complexes producing electron-dense se-cretory granules. Mitochondria are large, elongated, and often adjoining electronlucent vesicles.The kind and the origin of secretory granules varying in size and density were discussed. Thechemical analysis of the gland secretion revealed the presence of a mixture of low molecular weightterpenes, ketones, aldehydes, alcohols, esters, and carboxylic acids. Monoterpenes, especially linal-ool, were the major products. We supposed that ketones, aldehydes, alcohols, esters, and carboxylicacids have a deterrent function against the predators and monoterpenes provide a prophylaxisfunction against pathogens.Microsc. Res. Tech. 72:351–361, 2009. VVC 2008 Wiley-Liss, Inc.
INTRODUCTION
A wide variety of insect species have evolved chemi-cal defense mechanisms—involved in interspecificinteractions—that contributed to their widespread suc-cess in ecosystems (Thiele, 1977). In general, insect de-fensive substances are relatively low in molecularweight and are highly volatile, such as alkanes, ter-penes, alcohols, ketones, esters, aldehydes, organicacids, phenols, and quinones. They are secreted by exo-crine glands as blends and have repellent, toxicant, orantimicrobic function. The products of exocrine glandsare well known in Isoptera, Thysanoptera, Hymenop-tera, Hemiptera, Lepidoptera, and Coleoptera (Burseet al., 2007; Chapman, 1998; Dettner, 1987; Whitmanet al., 1990). According to our current knowledge, theorder Coleoptera shows the largest number of specieswith the greatest variety of defensive glands. Allo-mones are extensively investigated in coleopteranadult beetles, their larvae and eggs but no attentionhas been paid to the pupae (Chapman, 1998; Dettner,1987). Defensive chemicals have a high adaptive valuein the pupal stage because they do not show antipreda-tion strategies such as behavior or structural modifica-tions to protect themselves against natural enemies.
The ground beetles (Coleoptera, Carabidae) make upthe largest adephagan family with about 40,000 speciesand their distribution is cosmopolitan. They are wellknown as bioindicators and natural enemies of agricul-tural pests. Because of their bio-indication and econom-ical importance, the role of biotic and abiotic factors in
carabid population dynamics has been studied exten-sively. Abiotic factors and predation represent themain causes of mortality for all life-cycle stages of cara-bid beetles; pathogens and parasites can be importantfor some developmental stages (Lovei and Sunderland,1996). Therefore, in response to predation pressurethey have evolved morphological, behavioral, andchemical mechanisms against predation. Carabid bee-tles produce a large variety of defensive allomones(Andrew et al., 2000; Moore, 1979). Although exocrineglands (pygidial glands) and their defensive secretionwere well investigated in adults, no information isavailable about larvae and pupae which are the mostvulnerable stages of life cycle.
This study is focused on gland secretion at the pupalstage of carabid beetle Carabus lefebvrei Dejean, 1826.We chose this species because it is easy to rear underlaboratory conditions and easy to manipulate duringexperimental protocols. C. lefebvrei is a snail eatingpredatory Italian endemic species that lives in thebeech, oak, chestnut, and pine forests of the Centraland Southern Apennines, from lower altitudes to about
*Correspondence to: Anita Giglio, Department of Ecology, University of Cala-bria, via P. Bucci cubo 4B, I-87036 Arcavacata di Rende (CS), Calabria, Italy.E-mail: [email protected]
Received 10 July 2008; accepted in revised form 18 October 2008
DOI 10.1002/jemt.20660
Published online 9 December 2008 in Wiley InterScience (www.interscience.wiley.com).
VVC 2008 WILEY-LISS, INC.
MICROSCOPY RESEARCH AND TECHNIQUE 72:351–361 (2009)

1500 m a.s.l. It is a spring reproducing species, activefrom April until September and hibernates as adult.Eggs are laid in humus rich soils in April to June andlarvae are active on the surface of the leaf litter fromJune to August (Thiele, 1977; Turin, 2003). Larvaemoult to the pupal stage in a cell in the ground dug bythe 3rd instar larva. The pupa is exarate which wasdescribed by Sturani (1962), who also mentions a ‘‘fla-vored humor’’ produced by the pupa. Sturani (1962)supposed that this excretion has a waterproofing or anantipredatory function.
The aim of this study is to describe the ultrastruc-ture of the exocrine glands of C. lefebvrei pupa and toidentify the volatile constituents of gland secretion bygas chromatography/mass spectrometry (GC/MS).
MATERIALS AND METHODSExperimental Animals
C. lefebvrei males and females were hand collectedfrom under rotten bark of pine trees in the Catena Cos-tiera mountains (Southern Italy, Calabria) duringspring 2007 (900–1000 m a.s.l.). The beetles were keptin groups of three (two females and one male) in humidi-fied soil in 10 L plastic boxes. Females readily laid eggs,and the emerging larvae were transferred singly into150 mL glass jars. Adults and larvae were reared at alight regime of L/D5 16/8 h, 70% RH and at a day/nighttemperature of 23/188C. All specimens were fed withsnails. Ten days old pupae were used for this study.
Scanning Electron Microscopy
Five pupae were anesthetized and immersed in 100%chloroform solution for 1 h at 408C to clean the glandsecretion. After this specimens were rinsed with distilledwater, dehydrated in a graded ethanol series and criticalpoint-dried. After dehydration, they were mounted onstubs, gold coated and examined under a scanning elec-tron microscope (Cambridge Stereoscan 100).
Light and Transmission Electron Microscopy
Specimens were briefly anesthetized with chloroformand immediately immersed in 3% glutaraldehyde fixa-tive in 0.1 M cacodylate buffer with 5% sucrose pH 7.2–7.3 and kept overnight at 48C. Subsequently, they wererinsed in cacodylate buffer, postfixed with 1% osmiumtetroxide in 0.1 M cacodylate buffer for 1 h at 48C andrinsed in the same buffer. Dehydration in a gradedethanol series was followed by embedding in Spurrresin (Electron Microscopy Sciences). For transmissionelectron microscopy, thin sections were cut with anUltratome Leica Ultracut UCT ultramicrotome, subse-quently stained with uranyl acetate and lead citrateand examined with a Zeiss EM900 electron microscopeat 80 kV. For transmission electron microscopy, nega-tive plates were digitised with an Epson Photo Perfec-tion 2450 PHOTO scanner at 1200 dpi (optical resolu-tion) and saved as a tagged image format file.
For light microscopy, semithin sections were stainedwith methylene blue and observed under an AXIO-SKOP Zeiss light microscope. Images were acquiredwith a Nikon Coolpix 4500 photo camera.
Measurements were taken with Image-Pro Plus ver-sion 4.5 software (Media Cybernetics) on digitisedimage and processed as means6 Standard Deviation.
Glands are described and classified according toQuennedey (1998).
Gas Chromatography/Mass Spectrometry
For the GC/MS analyses of volatile compounds, thesolid phase-microextraction (SPME) technique wasused. SPME was performed with a 50/30 lm divinyl-benzene/carboxen/polydimethylsiloxane fiber (Supelco,Bellefonte, PA) which was introduced into a vial con-taining a single pupa; the fiber was lowered so that theextraction was performed in the headspace volume(�8 mL) at room temperature for 30 min and thenwithdrawn into the needle. The adsorbed analyteswere thermally desorbed by introducing the fiber for3 min into the injector port, set in splitless mode, of aVarian CP-3800 gas-chromatograph equipped with a30 m varian factor four capillary column (0.25 mm i.d.,0.25-lm thickness). Helium was used as carrier gas(1.2 mL/min flow). The injection port and the transferline were set at 2808C and 2308C, respectively. Theoven was heated at 408C for 1 min, and then the tem-perature was increased to 1808C at a rate of 58C/min,increased again at 108C/min to 2208C, and held for2 min. The GC was interfaced with a Varian SaturnMS-2000 ion trap (emission current 10 lA, ionizationtime 2 ms, scan range 40–300 amu). The MS systemwas operated in electron ionization (EI) mode. A blankanalysis of the fiber did not display any peak becauseof the analytes under investigation.
Compounds were identified based on alkane linearretention indices (C5 to C25) on the DB-5 column andcomparison of mass spectra with NIST 98 library spec-tra. Analyses were carried out on five pupae.
RESULTSMorphological Analysis
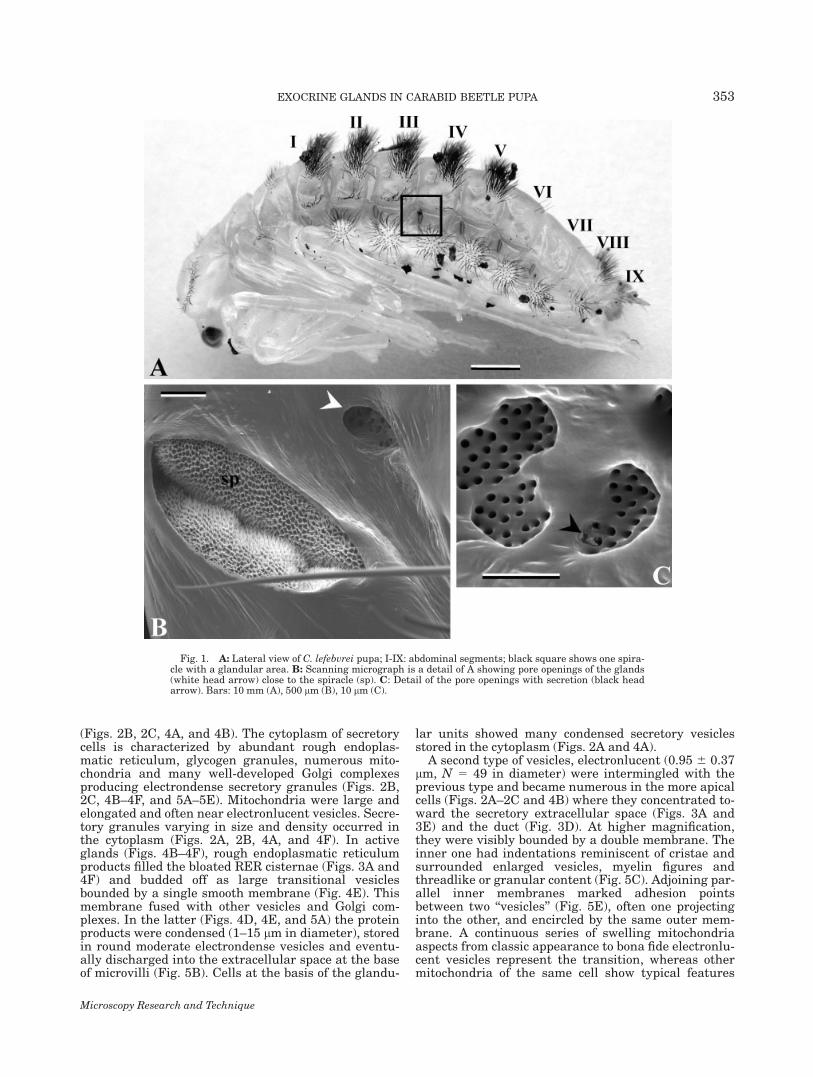
There was a deep invagination of the integument(0.6 mm 3 0.3 mm in diameter) with many pores,located on either side of the body near the spiracle fromI to VII abdominal segment (Figs. 1A–1C). A cross-sec-tion shows that the glands inside were grouped inabout 50 clusters at the side of each trachea, in closeassociation with tracheoles (Figs. 2A and 2B).
About 20 glandular units formed each cluster andthe unit was composed of two cells—a secretory and acanal cell—lying along a duct (Figs. 2A–2C and 3A–3F).
The gland’s duct was about 1 lm 3 1.6 lm in diame-ter and 400–600 lm long and had two parts: a conduct-ing (Figs. 3A, 3E, and 3F) and a receiving canal (Figs.3A–3D). The conducting canal was comprised of cuticu-lin and inner epicuticle (0.27 6 0.3 lm thick, N 5 10)and was surrounded by one canal cell (Figs. 3A–3D). Avery thin cytoplasm layer lined the conducting canalthat ran towards the cuticle through the epithelium(Figs. 2A and 3C). The receiving canal, 15–20 lm long,is located in the wide extracellular space of the secre-tory cell and is surrounded by microvilli (Figs. 3C, 3E,and 3F). Its epicuticular wall is perforated by curvedsplits (Figs. 3E and 3F). Septate junctions fastened theduct cell to the secretory cell (Fig. 3A).
The subspherical secretory cell had a diameter ofabout 60–70 lm. The nucleus, about 15–20 lm in diam-eter, is located in the basal portion of cytoplasm, waslargely euchromatic and had an evident nucleolus
Microscopy Research and Technique
352 A. GIGLIO ET AL.
(Figs. 2B, 2C, 4A, and 4B). The cytoplasm of secretorycells is characterized by abundant rough endoplas-matic reticulum, glycogen granules, numerous mito-chondria and many well-developed Golgi complexesproducing electrondense secretory granules (Figs. 2B,2C, 4B–4F, and 5A–5E). Mitochondria were large andelongated and often near electronlucent vesicles. Secre-tory granules varying in size and density occurred inthe cytoplasm (Figs. 2A, 2B, 4A, and 4F). In activeglands (Figs. 4B–4F), rough endoplasmatic reticulumproducts filled the bloated RER cisternae (Figs. 3A and4F) and budded off as large transitional vesiclesbounded by a single smooth membrane (Fig. 4E). Thismembrane fused with other vesicles and Golgi com-plexes. In the latter (Figs. 4D, 4E, and 5A) the proteinproducts were condensed (1–15 lm in diameter), storedin round moderate electrondense vesicles and eventu-ally discharged into the extracellular space at the baseof microvilli (Fig. 5B). Cells at the basis of the glandu-
lar units showed many condensed secretory vesiclesstored in the cytoplasm (Figs. 2A and 4A).
A second type of vesicles, electronlucent (0.95 6 0.37lm, N 5 49 in diameter) were intermingled with theprevious type and became numerous in the more apicalcells (Figs. 2A–2C and 4B) where they concentrated to-ward the secretory extracellular space (Figs. 3A and3E) and the duct (Fig. 3D). At higher magnification,they were visibly bounded by a double membrane. Theinner one had indentations reminiscent of cristae andsurrounded enlarged vesicles, myelin figures andthreadlike or granular content (Fig. 5C). Adjoining par-allel inner membranes marked adhesion pointsbetween two ‘‘vesicles’’ (Fig. 5E), often one projectinginto the other, and encircled by the same outer mem-brane. A continuous series of swelling mitochondriaaspects from classic appearance to bona fide electronlu-cent vesicles represent the transition, whereas othermitochondria of the same cell show typical features
Fig. 1. A: Lateral view of C. lefebvrei pupa; I-IX: abdominal segments; black square shows one spira-cle with a glandular area. B: Scanning micrograph is a detail of A showing pore openings of the glands(white head arrow) close to the spiracle (sp). C: Detail of the pore openings with secretion (black headarrow). Bars: 10 mm (A), 500 lm (B), 10 lm (C).
Microscopy Research and Technique
353EXOCRINE GLANDS IN CARABID BEETLE PUPA

Fig. 2. A: Semithin section of subepitelial glands. Black rectanglehighlights as detail of a cluster of glands units; bl, basal lamina; blackhead arrow, duct; cu, cuticle; e, epithelial cell; od, opening ducts; s,secretion; sc, secretory cell; l, lipid droplet. B: Longitudinal thin sec-tion of active secretory cell shows a nucleus (n) with a large nucleolus
(nu), abundant rough endoplasmatic reticulum (rer) and vesicle ofsecretion (sv2). Tracheoles (t) occur between the secretory cells. C:Longitudinal thin section of secretory cell showing many cytoplasmicevaginations (mi) and electronlucent vesicles (sv2); n, nucleus. Bars:100 lm (A), 2.5 lm (B, C).
Microscopy Research and Technique
354 A. GIGLIO ET AL.
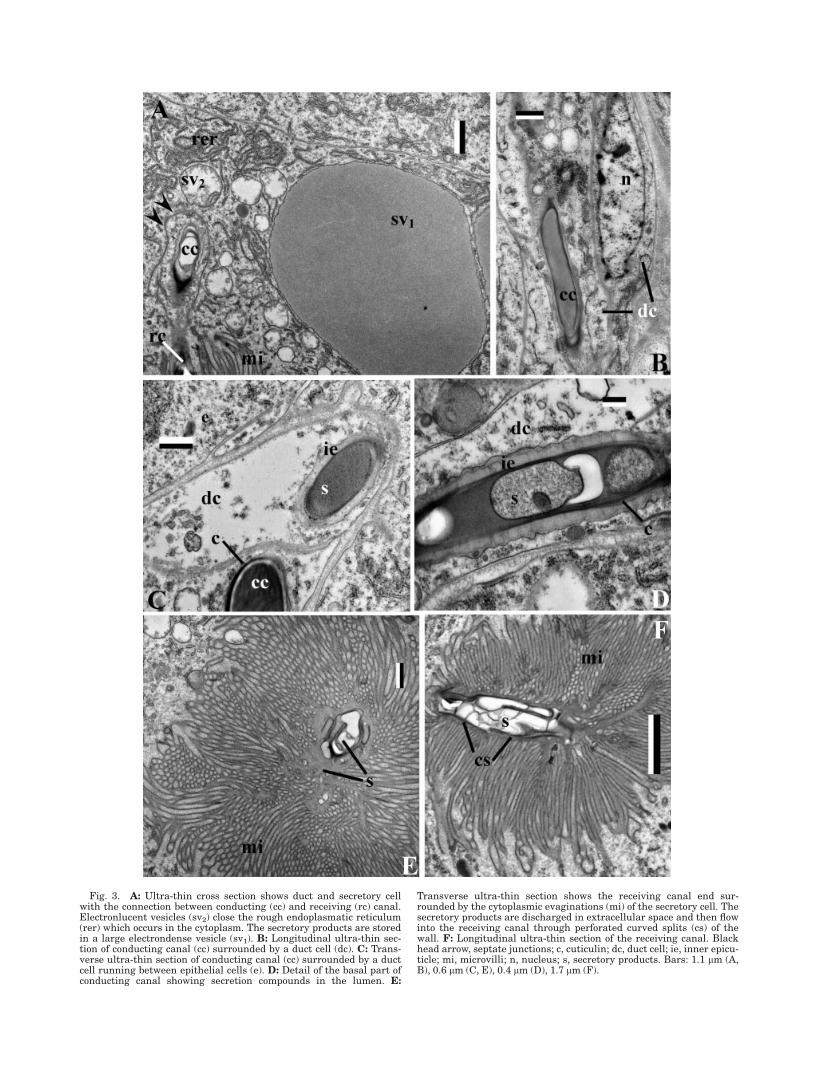
Fig. 3. A: Ultra-thin cross section shows duct and secretory cellwith the connection between conducting (cc) and receiving (rc) canal.Electronlucent vesicles (sv2) close the rough endoplasmatic reticulum(rer) which occurs in the cytoplasm. The secretory products are storedin a large electrondense vesicle (sv1). B: Longitudinal ultra-thin sec-tion of conducting canal (cc) surrounded by a duct cell (dc). C: Trans-verse ultra-thin section of conducting canal (cc) surrounded by a ductcell running between epithelial cells (e). D: Detail of the basal part ofconducting canal showing secretion compounds in the lumen. E:
Transverse ultra-thin section shows the receiving canal end sur-rounded by the cytoplasmic evaginations (mi) of the secretory cell. Thesecretory products are discharged in extracellular space and then flowinto the receiving canal through perforated curved splits (cs) of thewall. F: Longitudinal ultra-thin section of the receiving canal. Blackhead arrow, septate junctions; c, cuticulin; dc, duct cell; ie, inner epicu-ticle; mi, microvilli; n, nucleus; s, secretory products. Bars: 1.1 lm (A,B), 0.6 lm (C, E), 0.4 lm (D), 1.7 lm (F).
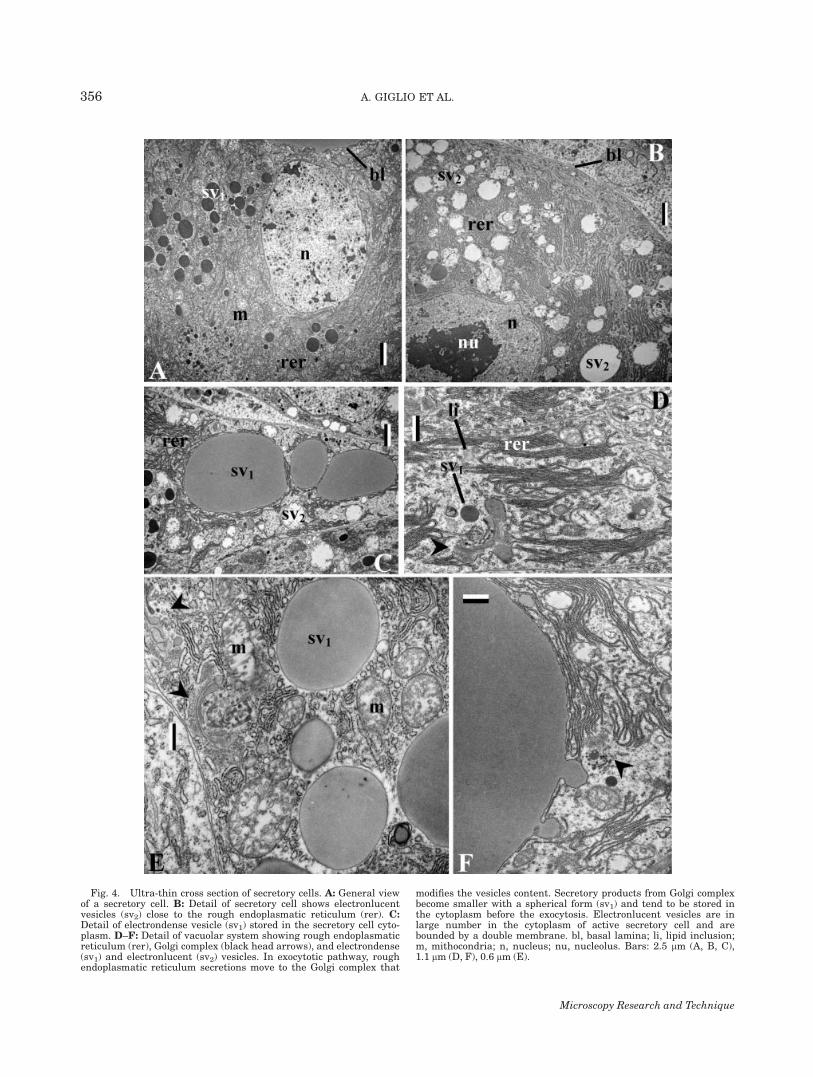
Fig. 4. Ultra-thin cross section of secretory cells. A: General viewof a secretory cell. B: Detail of secretory cell shows electronlucentvesicles (sv2) close to the rough endoplasmatic reticulum (rer). C:Detail of electrondense vesicle (sv1) stored in the secretory cell cyto-plasm. D–F: Detail of vacuolar system showing rough endoplasmaticreticulum (rer), Golgi complex (black head arrows), and electrondense(sv1) and electronlucent (sv2) vesicles. In exocytotic pathway, roughendoplasmatic reticulum secretions move to the Golgi complex that
modifies the vesicles content. Secretory products from Golgi complexbecome smaller with a spherical form (sv1) and tend to be stored inthe cytoplasm before the exocytosis. Electronlucent vesicles are inlarge number in the cytoplasm of active secretory cell and arebounded by a double membrane. bl, basal lamina; li, lipid inclusion;m, mithocondria; n, nucleus; nu, nucleolus. Bars: 2.5 lm (A, B, C),1.1 lm (D, F), 0.6 lm (E).
356 A. GIGLIO ET AL.
Microscopy Research and Technique
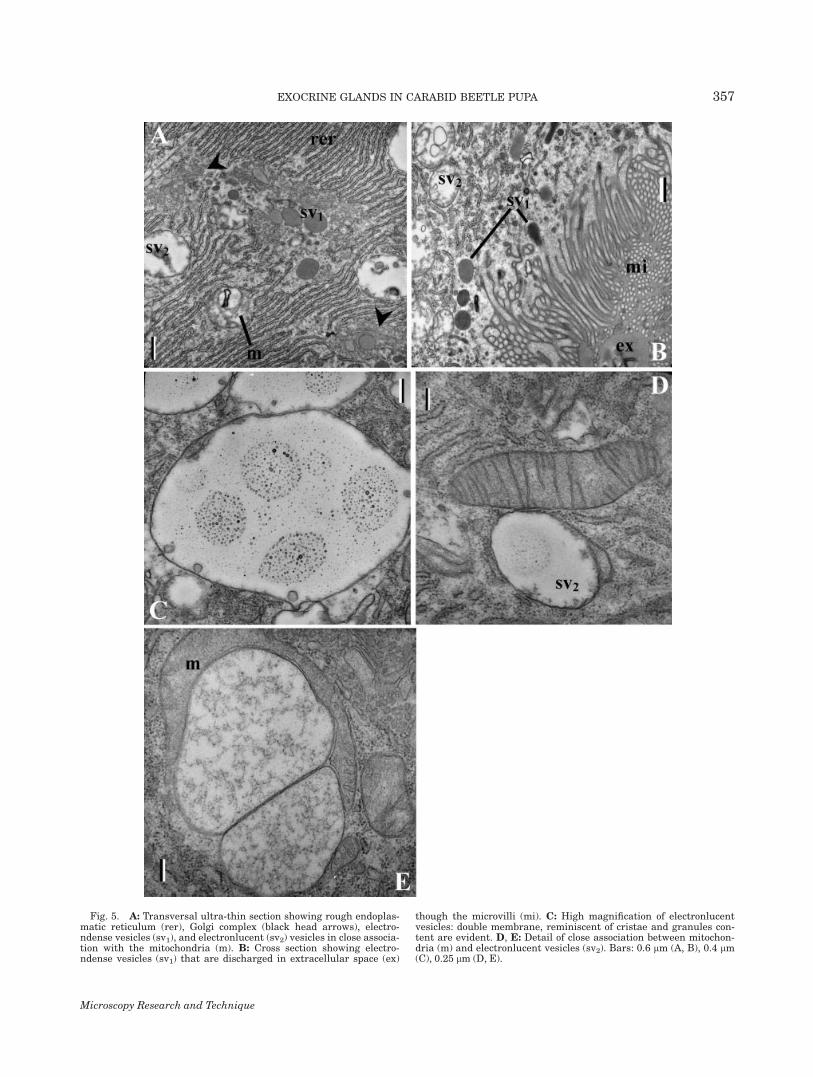
Fig. 5. A: Transversal ultra-thin section showing rough endoplas-matic reticulum (rer), Golgi complex (black head arrows), electro-ndense vesicles (sv1), and electronlucent (sv2) vesicles in close associa-tion with the mitochondria (m). B: Cross section showing electro-ndense vesicles (sv1) that are discharged in extracellular space (ex)
though the microvilli (mi). C: High magnification of electronlucentvesicles: double membrane, reminiscent of cristae and granules con-tent are evident. D, E: Detail of close association between mitochon-dria (m) and electronlucent vesicles (sv2). Bars: 0.6 lm (A, B), 0.4 lm(C), 0.25 lm (D, E).
Microscopy Research and Technique
357EXOCRINE GLANDS IN CARABID BEETLE PUPA
(Fig. 5D). In addition, mitochondria frequentlyembrace electronlucent vesicles and share their outermembrane (Fig. 5E).
Occasional lipid droplets were not bounded by amembrane although they had a densely stained inter-face with the cytosol (Figs. 2A and 4D).
The proximal portion of the cell showed a clearextracellular space with many cytoplasmic evagina-tions that increased the cell’s secretion surface (Figs.2A, 2C, 3E, and 3F). The secretory cell discharges loosematerial, through the microvilli, into their extracellu-lar space which then flows in the receiving canal.
Chemical Analysis
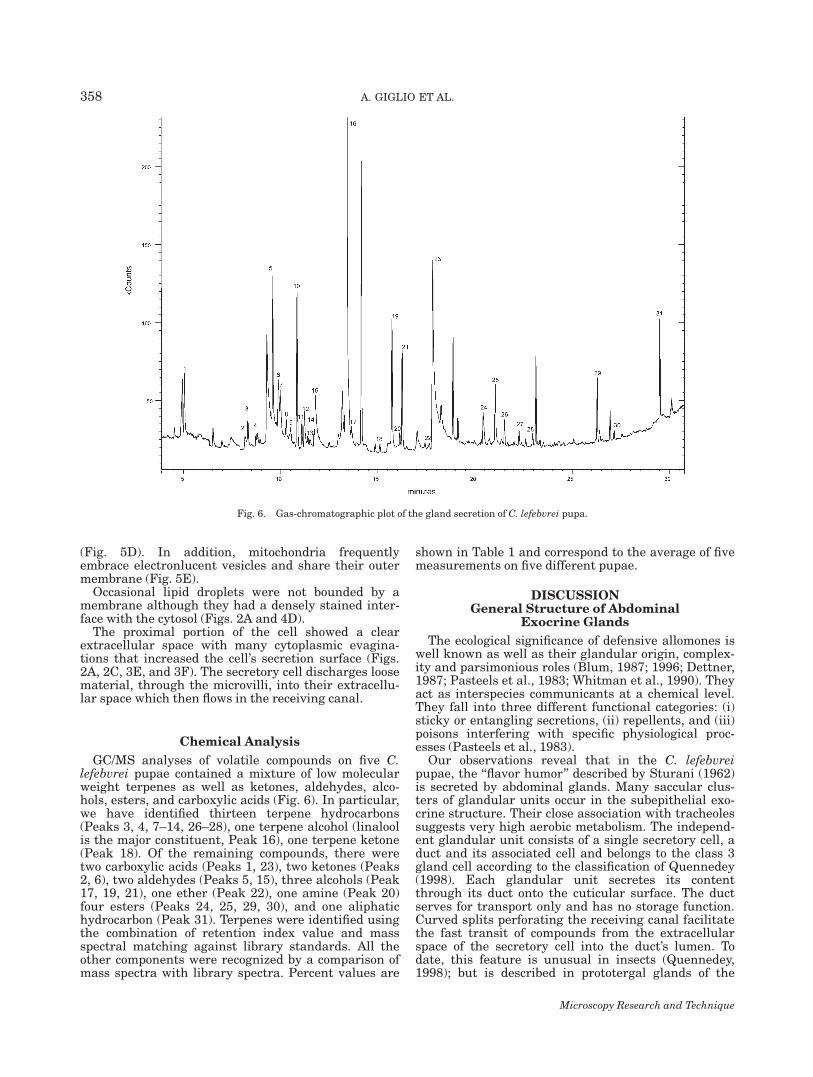
GC/MS analyses of volatile compounds on five C.lefebvrei pupae contained a mixture of low molecularweight terpenes as well as ketones, aldehydes, alco-hols, esters, and carboxylic acids (Fig. 6). In particular,we have identified thirteen terpene hydrocarbons(Peaks 3, 4, 7–14, 26–28), one terpene alcohol (linaloolis the major constituent, Peak 16), one terpene ketone(Peak 18). Of the remaining compounds, there weretwo carboxylic acids (Peaks 1, 23), two ketones (Peaks2, 6), two aldehydes (Peaks 5, 15), three alcohols (Peak17, 19, 21), one ether (Peak 22), one amine (Peak 20)four esters (Peaks 24, 25, 29, 30), and one aliphatichydrocarbon (Peak 31). Terpenes were identified usingthe combination of retention index value and massspectral matching against library standards. All theother components were recognized by a comparison ofmass spectra with library spectra. Percent values are
shown in Table 1 and correspond to the average of fivemeasurements on five different pupae.
DISCUSSIONGeneral Structure of Abdominal
Exocrine Glands
The ecological significance of defensive allomones iswell known as well as their glandular origin, complex-ity and parsimonious roles (Blum, 1987; 1996; Dettner,1987; Pasteels et al., 1983; Whitman et al., 1990). Theyact as interspecies communicants at a chemical level.They fall into three different functional categories: (i)sticky or entangling secretions, (ii) repellents, and (iii)poisons interfering with specific physiological proc-esses (Pasteels et al., 1983).
Our observations reveal that in the C. lefebvreipupae, the ‘‘flavor humor’’ described by Sturani (1962)is secreted by abdominal glands. Many saccular clus-ters of glandular units occur in the subepithelial exo-crine structure. Their close association with tracheolessuggests very high aerobic metabolism. The independ-ent glandular unit consists of a single secretory cell, aduct and its associated cell and belongs to the class 3gland cell according to the classification of Quennedey(1998). Each glandular unit secretes its contentthrough its duct onto the cuticular surface. The ductserves for transport only and has no storage function.Curved splits perforating the receiving canal facilitatethe fast transit of compounds from the extracellularspace of the secretory cell into the duct’s lumen. Todate, this feature is unusual in insects (Quennedey,1998); but is described in prototergal glands of the
Fig. 6. Gas-chromatographic plot of the gland secretion of C. lefebvrei pupa.
Microscopy Research and Technique
358 A. GIGLIO ET AL.

adult rove beetle Philonthus varians (Quennedeyet al., 2002).
The secretory cell is very large and shows a widearray of microvilli increasing the exocytotic cell sur-face. Large amounts of rough endoplasmatic reticulumwith enlarged cisternae were found in the cytoplasm ofsecretory cells in close association with many Golgicomplexes. This suggests that the secretion containsproteins likely associated with lipids and/or glucids.Secretory products show great variability in theirshape and electron density. Moderately electrondensevesicles derive from the rough endoplasmatic reticu-lum and their contents are modified in the Golgi com-plexes. Electrondense vesicles are condensing materialincluding a lipid moiety and derived from the Golgicomplexes (Locke, 1998)
Electronlucent vesicles are commonly described inmany insect gland systems (e.g., Costa-Leonardo,2006; Happ et al., 1966; Sobotnık and Weyda, 2003;Quennedey, 1998), and are often associated with glyco-gen breakdown and lipid production (Locke, 1998).Some of the features like close association with mito-chondria, granular or myelin content are already men-tioned in the literature, but no high power magnifica-tion illustration nor explicit reference is given aboutthe presence of double membrane walled intracellularvesicles, unless for autophagic processes under way.Transient double membrane vesicles released in theextracellular space are generated through a peculiarexocytotic budding pattern from the microvillar tip of amidgut cell of lepidopteran mouth (Jordao et al., 1999).
Happ et al. (1966) report the occurrence of myelinfigures and degenerating mitochondria during the
functional cycle of the terpene secretory cell. Sobotnikand Weyda (2003) show ‘‘lucent vesicles containingirregular electrondense component’’ resulting ‘‘fromthe transformation of normal mitochondria’’ and statethat ‘‘another way of lucent vesicle decomposing is thechange into one or several myelin figures.’’ This processoccurs, however, in specialized duct cells and not in themain glands. Dividing mitochondria to compensateautophagic loss are reported by Locke (1998) in the fatbody, and they share the outer membrane. Actually,Burse et al. (2007) report in the fat body of monoterpe-noid producing Chrysomelina larvae increased 3-hydroxy-3-methylglutaryl-CoA reductase expressionand high enzymatic activity leading to accumulation ofgeraniol intermediates indicating geranyl diphosphatesynthase activity observed only there. However, thecell compartment involved is yet undefined.
Happ et al. (1966) show the granular localization inthe secretory cell of the mevalonate pathway enzymesinvolved in terpene biosynthesis of stick insects,whereas Oldham et al. (1996) demonstrate that D-la-beled mevalonate is incorporated into terpene synthe-sizing glands of a leaf beetle larva. Isoprenoid deriva-tive synthesis through mevalonate occurs in the cyto-plasm, but is dependent on a continuous massivesupply of acetyl residues from mitochondria—throughtricarboxylic acids cycle, citrate export and ATP-citratelyase—to bind cytoplasmic CoA (Garrett and Grisham,1995). Therefore, the presence of proliferating, hyper-functioning mitochondria is expected. In the end oftheir stressing hyperactivity their inner membranecan be fragmented and degenerate, turning into myelinfigures. The outer membrane makes up a single vesicle,
TABLE 1. Peaks identification of gland secretion of Carabus lefebvrei pupae (N 5 5)
PeaksRetentiontime (min) % N 5 5 Identification
1 4.95 0.18 Pivalic acid2 8.175 0.08 p-Benzoquinone3 8.318 0.48 a-Pinene4 8.819 0.20 Camphene5 9.291 0.13 Benzaldehyde6 9.882 0.79 6-Methyl-5-hepten-2-one7 9.976 2.97 b-Pinene8 10.29 0.69 2-Carene9 10.517 0.60 a-Phellandrene
10 10.84 5.42 a-Terpinene11 11.093 0.88 orto-Cimene12 11.222 0.96 Limonene13 11.398 0.46 cis-Ocimene14 11.722 0.50 trans-Ocimene15 11.793 0.79 2-Hydroxy benzaldehyde16 13.365 77.98 Linalool17 13.655 0.10 3-Nonen-1-ol (nonanal)18 14.853 0.10 Camphor19 15.728 1.24 4,8-Dimethyl-3,7-nonadien-2-ol20 16.128 0.14 1-(1-Oxobutyl)-1,2-dihydropyridine21 16.249 1.06 3,4-Di(1-butenyl)-tetrahydrofuran-2-ol22 17.437 0.36 Thymol methyl ether23 17.824 0.03 Phenylacetic acid24 20.424 0.72 Propanoic acid, 2-methyl-, 2,2-dimethyl-
1-(2-hydroxy-1-methylethyl)propyl ester25 21.04 1.01 Propanoic acid, 2-methyl-, 3-hydroxy-2,
4,4-trimethylpentyl ester26 21.514 0.90 b-Elemene27 22.266 0.40 a-Santalene28 22.603 0.23 trans-a-Bergamotene29 26.287 0.50 Pentanoic acid, 2,2,4-trimethyl-3-carboxyisopropyl, isobutyl ester30 27.157 0.05 Dodecanoic acid, 1-methylethyl ester31 29.508 0.06 Sclarene
Microscopy Research and Technique
359EXOCRINE GLANDS IN CARABID BEETLE PUPA

containing mainly lipids (Locke, 1998), which mayundergo the lytic or the secretory exocytotic pathway,including fusion with Golgi complex, eventuallyextruding its content.
Secretory Compounds
In the gas chromatographic analysis of volatile com-pounds of C. lefebvrei pupa we have identified a total of31 components. The main components of gland secre-tion were monoterpenes that are oligomeres of isoprenehydrocarbon (2-methyl-1,3 butadiene). Terpenes aremostly present in plants as constituents of essentialoils but are not unknown in insect secretions(Gershenzon and Dudareva, 2007).
In plants, monoterpenes (C10 compounds) areinvolved in antifungal and antimicrobial activity(Angioni et al., 2004; Ishida, 2005; Letchamo et al.,2004) as well as host plant attractants (Cunninghamet al., 2006; Halitschke et al., 2008); or repellents(Cheng et al., 2007; Guillet et al., 1998; Keeling andBohlmann, 2006; Lucia et al., 2007) or as a pheromoneor synergist to the pheromone (Erbilgin et al., 2003).
In insects, monoterpenes occur in gland secretions ofmany taxa (Isoptera, Hemiptera, Hymenoptera, Cole-optera), and are chemical deterrents, trail markers,mate attractants or alarm pheromones (Billen et al.,2000; Blum, 1987; Chapman, 1998; Dettner, 1987;Gnanasunderam et al., 1981; Whitman et al., 1990).Alfa- and b-pinene, camphene and limonene are foundin the Dufour’s glands secretion of worker ants (Billenet al., 2000) and in the pygidial defensive secretion oftwo tenebrionid species (Geiselhardt et al., 2006). Lin-alool is a mate attractant pheromone component in thebee Colletes cunicularius (Borg-Karlson et al., 2003)whereas in pentatomids it is an aggregation phero-mone secreted from dorsal abdominal glands of adults(Aldrich et al., 1984).
The biosynthetic origin of these volatile compoundshas been a controversial issue. The biochemical path-way of the terpene biosynthesis is outlined by Happet al. (1966). They suggest that the terpenoid toxicantin a phasmid gland is produced from the lipid reservesvia the mevalonic acid pathway. Blum (1987) and Erbil-gin et al. (2003) demonstrated that bark beetles canmetabolize host-derivate monoterpenes into phero-mones. In addition Oldham et al. (1996) gave experi-mental proof of the ability of terpene de novo synthesisin a leaf beetle Phaedon armoraciae defensive glands.Laurent et al. (2003) have summarized the biosyn-thetic pathway operating for the production of defen-sive compounds in some beetle and ant species. How-ever, knowledge is very insufficient to understand thedifferent selective pressures under which insects haveevolved the metabolic pathway for the biosynthesis ofdefensive volatile compounds. Moreover, no informa-tion is in reports about metabolite trafficking betweensubcellular compartments and the mechanism of therelease process.
Carboxylic acids, ketones, aldehydes, alcohols, ethers,esters, and amine are aromatic compounds that occurin smaller amount in gland secretion of C. lefebvreipupa. These compounds are commonly found in thesecretion of ants’ mandibular, pygidial, and Doufour’sglands (Holldobler and Wilson, 1990). Many speciesof beetles possess abdominal, thoracic and pygidial
glands which emit hydrocarbons, aldehydes, organicacids, esters (Dettner, 1987; Whitman et al., 1990). Inarthropods, the biosynthesis of many compoundsbelonging to these classes is reviewed by Blum (1987).
In carabid beetles, defensive chemicals from adultpygidial gland system have been well known so thatthey seem to be of considerable taxonomic value atthe genus and species levels (Attygalle et al., 2007;Dettner, 1987; Moore, 1979; Will et al., 2000). Amongthe compounds found in the pygidial gland secretion,carboxylic acids, hydrocarbons, ketones, esters, andaldehydes have been regarded as the deterrent againstpredators.
The present study is apparently the first report onchemical defenses in pupal stage. Further investiga-tions are needed to clarify the exact function of the C.lefebvrei pupa’s gland secretions. However, we sup-posed that this gland secretion has a deterrent functionagainst the predators and a prophylaxis functionagainst pathogens. The biological importance of themixture in gland secretions has been demonstrated inmany reports. Indeed, the nonpolar compounds act tofacilitate the spreading of volatile polar compounds inthe predator cuticle (Dettner, 1987; Pasteels et al.,1983). Moreover, mixtures of monoterpenes, such as inconifer resin, may act synergistically in defense. Theexarate pupal stages of carabid beetles have no physi-cal protection and then the chemical protection pro-vided by the abdominal glands are very important forindividual fitness. The soils of their habitat are veryrich in bacterial and fungal micro-organisms, some ofwhich are possible pathogens. Besides, the highly lipo-philic nature of monoterpene compounds suggests thattheir principal targets are bacteria and/or fungal cellmembranes.
ACKNOWLEDGMENTS
The authors thank Sig. Enrico Perrotta for the tech-nical assistance in electron microscopy, Dr. AntonioMazzei for the photo preparation of pupa (Fig. 1A), andthe anonymous reviewers for their useful comments.
REFERENCES
Aldrich JR, Lusby WR, Kochansky JP, Abrams CB. 1984. Volatilecompounds from the predatory insect Podisus maculiventris (Hemi-ptera: Pentatomidae). J Chem Ecol 10:561–568.
Andrew DJ, Henderson KD, Seshaiah P. 2000. Salivary gland devel-opment in Drosophila melanogaster. Mech Dev 92:5–17.
Angioni A, Barra A, Cereti E, Barile D, Coisson JD, Arlorio M, DessiS, Coroneo V, Cabras P. 2004. Chemical composition, plant geneticdifferences, antimicrobial and antifungal activity investigation ofthe essential oil of Rosmarinus officinalis L. J Agric Food Chem52:3530–3535.
Attygalle AB, Wu X, Will KW. 2007. Biosynthesis of tiglic, ethacrylic,and 2-methylbutyric acids in a carabid beetle, Pterostichus(Hypherpes) californicus. J Chem Ecol 33:963–970.
Billen J, Ito F, Tsuji K, Schoeters E, Maile R, Morgan ED. 2000. Struc-ture and chemistry of the Dufour’s gland in Pristomyrmex ants (Hy-menoptera, Formicidae). Acta Zool 81:159–166.
Blum MS. 1987. Biosynthesis of arthropod exocrine compounds. AnnuRev Entomol 32:381–413.
Blum MS. 1996. Semiochemical parsimony in the Arthropoda. AnnuRev Entomol 41:353–374.
Borg-Karlson A-K, Tengo J, Valterova I, Unelius CR, Taghizadeh T,Tolasch T, Francke W. 2003. (S)-(1)-Linalool, a mate attractantpheromone component in the bee Colletes cunicularius. J ChemEcol 29:1–14.
Burse A, Schmidt A, Frick S, Kuhn J, Gershenzon J, Boland W. 2007.Iridoid biosynthesis in Chrysomelina larvae: Fat body producesearly terpenoid precursors. Insect Biochem Mol Biol 37:255–265.
360 A. GIGLIO ET AL.
Microscopy Research and Technique

Chapman RF. 1998. The insects: structure and function. Cambridge,UK: Cambridge University Press.770 p.
Cheng A-X, Lou Y-G, Mao Y-B, Lu S, Wang L-J, Chen X-Y. 2007. Plantterpenoids: Biosynthesis and ecological functions. J Integr PlantBiol 49:179–186.
Costa-Leonardo AM. 2006. Morphology of the sternal gland in work-ers of Coptotermes gestroi (Isoptera, Rhinotermitidae). Micron 37:551–556.
Cunningham JP, Moore CJ, Zalucki MP, Cribb BW. 2006. Insect odourperception: Recognition of odour components by flower foragingmoths. Proc R Soc B Biol Sci 273:2035–2040.
Dettner K. 1987. Chemosystematics and evolution of beetle chemicaldefenses. Annu Rev Entomol 32:17–48.
Erbilgin N, Powell JS, Raffa KF. 2003. Effect of varying monoterpeneconcentrations on the response of Ips pini (Coleoptera: Scolytidae)to its aggregation pheromone: Implications for pest managementand ecology of bark beetles. Agric For Entomol 5:269–274.
Garrett RH, Grisham CM. 1995. Biochemistry. New York: SaundersCollege Publishing.
Geiselhardt S, Szepat T, Rasa O, Peschke K. 2006. Defensive secre-tion components of the host Parastizopus armaticeps as kairomonesfor the cleptoparasite Eremostibes opacus. J Chem Ecol 32:767–778.
Gershenzon J, Dudareva N. 2007. The function of terpene naturalproducts in the natural world. Nat Chem Biol 3:408–414.
Gnanasunderam C, Young H, Hutchins RFN. 1981. Defensive secre-tions of New Zealand tenebrionids. J Chem Ecol 7:889–894.
Guillet G, Belanger A, Arnason JT. 1998. Volatile monoterpenes inPorophyllum gracile and P. ruderale (Asteraceae): Identification,localization and insecticidal synergism with a-terthienyl. Phyto-chemistry 49:423–429.
Halitschke R, Stenberg JA, Kessler D, Kessler A, Baldwin IT. 2008.Shared signals-‘‘alarm calls’’ from plants increase apparency to her-bivores and their enemies in nature. Ecol Lett 11:24–34.
Happ GM, Strandberg JD, Happ CM. 1966. The terpene-producingglands of a phasmid insect cell morphology and histochemistry.J Morphol 119:143–159.
Holldobler B, Wilson EO. 1990. The Ants. Berlin Heidelberg:Springer-Verlag.732 p.
Ishida T. 2005. Biotransformation of terpenoids by mammals, micro-organisms, and plant-cultured cells. Chem Biodivers 2:569–590.
Jordao BP, Capella AN, Terra WR, Ribeiro AF, Ferreira C. 1999. Na-ture of the anchors of membrane-bound aminopeptidase, amylase,and trypsin and secretory mechanisms in Spodoptera frugiperda(Lepidoptera) midgut cells. J Insect Physiol 45:29–37.
Keeling CI, Bohlmann J. 2006. Genes, enzymes and chemicals of ter-penoid diversity in the constitutive and induced defence of conifersagainst insects and pathogens. New Phytol 170:657–675.
Laurent P, Braekman J-C, Daloze D, Pasteels J. 2003. Biosynthesis ofdefensive compounds from beetles and ants. European J Org Chem2003:2733–2743.
Letchamo W, Ward W, Heard B, Heard D. 2004. Essential oil of Valeri-ana officinalis L. Cultivars and their antimicrobial activity as influ-enced by harvesting time under commercial organic cultivation.J Agric Food Chem 52:3915–3919.
Locke M. 1998. The fat body. In: Harrison FW, Locke M, editors.Microscopic anatomy of invertebrates. New York: Wiley-Liss.pp. 641–686.
Lovei GL, Sunderland KD. 1996. Ecology and behavior of GroundBeetles (Coleoptera: Carabidae). Annu Rev Entomol 41:231–256.
Lucia A, Gonzalez Audino P, Seccacini E, Licastro S, Zerba E, MasuhH. 2007. Larvicidal effect of Eucalyptus grandis essential oil andturpentine and their major components on Aedes aegypti larvae.J AmMosq Control Assoc 23:299–303.
Moore BP. 1979. Chemical defense in carabids and its bearing on phy-logeny. In: Erwin TL, Ball GE, Whitehead DR, editors. Carabid bee-tles: Their evolution, natural history and classification. London: DrJunk W. pp. 193–203.
Oldham NJ, Veith M, Boland W, Dettner K. 1996. Iridoid mono-terpene biosynthesis in insects: Evidence for a de novo path-way occurring in the defensive glands of Phaedon armoraciae(Chrysomelidae) leaf beetle larvae. Naturwissenschaften 83:470–473.
Pasteels JM, Gregoire JC, Rowell-Rahier M. 1983. The chemicalecology of defense in arthropods. Annu Rev Entomol 28:263–289.
Quennedey A. 1998. Insect epidermal glands cells: Ultrastructureand morphogenesis. In: Harrison FW, Locke M, editors. Micro-scopic anatomy of invertebrates. New York: Wiley-Liss. pp. 177–207.
Quennedey A, Drugmand D, Deligne J. 2002. Morphology and ultra-structure of paired prototergal glands in the adult rove beetle Phil-onthus varians (Coleoptera, Staphylinidae). Arthropod Struct Dev31:173–183.
Sobotnık J, Weyda F. 2003. Ultrastructural ontogeny of the labialgland apparatus in termite Prorhinotermes simplex (Isoptera, Rhi-notermitidae). Arthropod Struct Dev 31:255–270.
Sturani M. 1962. Osservazioni e ricerche biologiche sul genere Cara-bus Linneus (sensu lato). Memorie della Societa Entomologica Ital-iana XLI:85–202.
Thiele H-U. 1977. Carabid beetles in their environments. Berlin Hei-delberg: Springer-Verlag. 369 p.
Turin H, Penev L, Casale A. 2003. The genus Carabus in Europe. Asynthesis. Sofia Moscov: Pensoft. 512 p.
Whitman WD, Blum MS, Alsop DW. 1990. Allomones: Chemicals fordefense. In: Evans DL, Schmidt OJ, editors. Insect defenses. Adapt-ive mechanisms and strategies of prey and predators. Albany: Uni-versity of New York Press. pp. 289–351.
Will KW, Attygalle AB, Herath K. 2000. New defensive chemical datafor ground beetles (Coleoptera: Carabidae): Interpretations in aphylogenetic framework. Biol J Linn Soc 71:459–481.
361EXOCRINE GLANDS IN CARABID BEETLE PUPA
Microscopy Research and Technique




















