
Indirect effects of species interactions on habitat provisioning
Upload
independentCategory
view
0download
0
Maternal provisioning of sequestered defensive steroids by the Asian snakeRhabdophis tigrinus
Deborah A. Hutchinson1, Alan H. Savitzky1, Akira Mori2, Jerrold Meinwald3, and Frank C. Schroeder4
1 Old Dominion University, Department of Biological Sciences, Norfolk, VA 23529, USA2 Kyoto University, Department of Zoology, Graduate School of Science, Sakyo, Kyoto 606-8502, Japan3 Cornell University, Department of Chemistry and Chemical Biology, Ithaca, NY 14853, USA4 Boyce Thompson Institute, Cornell University, Ithaca, NY 14853, USA
Summary. Rhabdophis tigrinus obtains defensive steroids(bufadienolides) from its diet and sequesters those com-pounds in specialized structures on its neck known asnuchal glands. Hatchling snakes lacking these steroidsmust acquire them from toads consumed as prey. Here weshow that females provision bufadienolides to their off-spring in amounts correlated to the quantity in their ownnuchal glands; thus, chemically protected mothers pro-duce defended offspring. Bufadienolides can be provi-sioned to embryos via deposition in yolk and by transferacross the egg membranes within the oviducts. Maternallyprovisioned bufadienolides persist in the nuchal glands ofjuvenile snakes from the time of hatching in late summeruntil the following spring, when toads of ingestible sizebecome abundant. Therefore, maternal provisioning mayprovide chemical protection from predators for young R.tigrinus in the absence of dietary sources of bufadieno-lides.
Key words. Dietary toxins – Bufo – bufadienolide – nu-chal glands – antipredator defense
Introduction
Chemically defended animals may synthesize their de-fensive toxins from nontoxic precursors or sequester in-tact toxins from an exogenous source, such as their diet(Duffey 1980; Brower 1984; Daly et al. 1994; Eisner etal. 1997; Nishida 2002; Dumbacher et al. 2004; Saporito etal. 2004; Williams et al. 2004). Regardless of how adultsbecome toxic, their newly emergent offspring could enjoya selective advantage if they were defended by maternalprovisioning of defensive chemicals. When toxins are
deposited in the eggs of oviparous species, those toxinsmay serve as deterrents against egg predation (oophagy).Subsequently, the toxins may be incorporated into thedeveloping embryos� tissues. If embryos incorporate thetoxins, the young also may be chemically defended for aperiod of time after hatching.
There are many examples of defensive toxins in theeggs of chemically defended species. Eggs of rough-skinned newts (Taricha granulosa) and a Costa Ricanharlequin frog (Atelopus chiriquiensis) are provisionedwith tetrodotoxin, a potent neurotoxin (Pavelka etal. 1977; Hanifin et al. 2003). Toxic eggs are known fromtoads (Licht 1968) and moths in the genus Utethesia(Eisner and Meinwald 1995; Eisner et al. 2000). Eggs offireflies (Photuris) are endowed with betaine synthesizedby the female, and also with sequestered lucibufaginswhen available (Gonz�lez et al. 1999a).
Toxins that provide chemical defense for eggs may ormay not become incorporated into the developing em-bryos. Among invertebrates, provisioning of toxins ineggs can result in chemically defended larvae (Pasteels etal. 1986; Blum and Hilker 2002; Eisner et al. 2002).Among vertebrates, some chemically defended eggsgradually lose their toxicity as development ensues(Phisalix 1922; Twitty 1937). Larvae of Anaxyrus boreas(formerly Bufo boreas ; see Frost et al. 2006) have beenshown to lack defensive bufadienolides (Benard andFordyce 2003), demonstrating a loss of provisioned toxinsthat had protected the eggs.
Rhabdophis tigrinus is an oviparous Asian snake(Colubridae: Natricinae) that possesses unusual defen-sive glands in the skin of its neck (Nakamura 1935; Smith1938). Each individual has a series of 10 – 20 pairs of thesenuchal glands that extends caudally behind the head(Nakamura 1935; Smith 1938; Toriba and Sawai 1990).The contents of the nuchal glands irritate mucous mem-branes, cause corneal injuries (Nakamura 1935; Kitaz-ume 1953; Kawashima 1957, 1959; Suzuki 1960; Asahi et
Correspondence to: Deborah A. Hutchinson, email: [email protected] and Alan H. Savitzky, email: [email protected]
Chemoecology 18: 181 – 190 (2008)0937-7409/08/030181-10� Birkh�user Verlag, Basel, 2008DOI 10.1007/s00049-008-0404-5
CHEMOECOLOGY

al. 1985), and contain cardiotonic steroids known as buf-adienolides (Akizawa et al. 1985a,b; Hutchinson etal. 2007). Bufadienolides act by inhibiting the sodium-potassium pump in a digitalis-like fashion (Melero etal. 2000). The steroidal toxins in the nuchal glands aresimilar to those found in the integumentary secretions oftoads (Bufonidae; Erspamer 1994), which often are con-sumed, in addition to other anuran prey, by R. tigrinus(Mori and Moriguchi 1988). Whereas toads are capable ofsynthesizing their defensive bufadienolides from choles-terol (Doull et al. 1951; Siperstein et al. 1957; Porto andGros 1971; Porto et al. 1972), R. tigrinus is dependent on adiet of toads from which it can sequester these com-pounds (Hutchinson et al. 2007). Although toads are theultimate source of bufadienolides in R. tigrinus, a previousexperiment suggested that hatchlings may either lack orpossess these defensive compounds, depending upon thelevel of toxins in the mother�s nuchal glands (Hutchinsonet al. 2007).
Having demonstrated previously that R. tigrinus de-pends upon a diet of toads to supply its bufadienolides(Hutchinson et al. 2007), we sought to determine whetherchemically defended dams (mothers) could provisionsequestered bufadienolides to their offspring. In thepresent study, we conducted a dietary experiment on 12gravid, wild-caught R. tigrinus and their offspring to an-swer this question. We maintained eight gravid R. tigrinuson nontoad diets and fed toads to four other gravid fe-males. We analyzed samples of nuchal gland fluid fromthe gravid females and their unfed, fish-fed, and toad-fedoffspring, as well as selected samples of yolk. We presentevidence that maternal provisioning of toxins in R. tigri-nus occurs through two mechanisms: by deposition inyolk and by transfer across the egg membranes within theoviduct, late in gestation. Furthermore, maternal provi-sioning of bufadienolides results in a gradation of chem-ical defense among clutches, in accordance with the levelof toxins in the dams.
Methods and Materials
Experimental Design and Sample Collection. Twelve gravid Rhabdo-phis tigrinus were collected in May and June, 2005 on the Japaneseislands of Honshu, Kyushu, and Shikoku. Nine of the snakes werecollected on Honshu, from Okayama Prefecture: Tomi-son (n = 1),Kagamino-cho (n = 1), Mikamo-son (n = 2), Shinjo-son (n = 1); Hi-roshima Prefecture: Shohbara City (n = 1); and Kyoto Prefecture:Miyama-cho (n = 3). On Kyushu, one snake was collected from Ku-mamoto Prefecture: Yamato-cho. Two snakes were collected on Shi-koku from Tokushima Prefecture: Naka-gun (n = 1) and Ishima Island(n = 1).
The gravid females were fed nonbufonid frogs, primarily Pelo-phylax nigromaculatus (formerly Rana ; Frost et al. 2006; Che etal. 2007), but also Hyla japonica and Rhacophorus arboreus. The fe-males also were fed pre-killed fishes (Plecoglossus altivelis, Sardinopssagax, Oncorhynchus mykiss, Hypomesus nipponensis, and Pseudo-rasbora parva). Additionally, four dams were fed a single, live adulttoad (Bufo japonicus) from Niijima (Tokyo Prefecture: Izu Islands)prior to oviposition, and two other dams were fed a live toad shortlyafter oviposition.
Samples of nuchal gland fluid were collected from the R. tigrinusdams upon capture (2–6 weeks prior to oviposition), and again afteroviposition, at which point they had been fed in the laboratory. Thenuchal gland fluid was collected by expressing a few glands onto aportion of Kimwipe (Kimwipes wipers S-200; Kimberly-Clark, Dallas,TX, USA) while wearing powder-free NBR nitrile gloves (AS ONE,Edobori, Nishiku, Osaka, Japan). The Kimwipe was then placed in avial of methanol with forceps and covered with a teflon-lined cap. Wewere careful to express only a few glands to ensure that we could res-ample individuals at later dates; nuchal glands do not appear to re-generate, at least initially, after they have ruptured. We attempted tostandardize the number of glands expressed (and thus the volume offluid that was collected), so that samples could be compared throughtime and across individuals. Gloves were changed and forceps wererinsed in methanol between samplings. A control vial consisting of aKimwipe in methanol was prepared at the end of each sampling sessionto ensure that cross contamination did not occur. All samples werestored at –208C.
Oviposition took place at Kyoto University from June through July2005; eggs hatched in August. The temperature of the enclosurehousing the eggs and snakes was maintained between 25 and 308C. Fiveoviposited eggs from clutches 3, 10, 11, and 12 were dissected to collectyolk for chemical analyses. Yolk samples either were extracted inmethanol and frozen, or were frozen without solvent, lyophilized, andlater extracted in methanol.
We assigned feeding treatments (toads or fish) to the hatchlings in abalanced, randomized design, so that each clutch was represented asequally as possible among the two treatment groups. However, forclutches with only two hatchlings (clutches 5 and 12), we designated alloffspring to the fish-fed group. Hatchlings were housed individually inscreen-topped plastic containers with a paper towel as a substrate.Water was available ad libitum. Some offspring that hatched early werefed fish (Oryzias latipes) to sustain them until the assignment of feedingtreatments.
During the feeding trial, all hatchlings (3–6 weeks of age) wereoffered food on the same days. On each feeding day, hatchlings in thefish-fed group were given one live fish (O. latipes), which typicallyweighed between 0.07 and 0.14 g; hatchlings in the toad-fed group wereoffered three thawed metamorphic toads (Bufo japonicus), whichweighed a total of 0.10–0.16 g. The metamorphic toads were raised fromtadpoles collected in Chiba Prefecture and were frozen in the spring of2005. The snakes were left with their food overnight, and any snakesthat had not eaten by the following day were force-fed. Food was of-fered to the hatchlings on three dates, with any necessary force-feedingstaking place on the following day.
Samples of nuchal gland fluid were collected from 36 unfedhatchlings, 15 of which were resampled four days after the end of thefeeding trial. Additionally, samples of nuchal gland fluid from 42previously unsampled hatchlings were collected after the feedingtrial to provide samples from a total of 29 fish-fed and 28 toad-fedsnakes.
To test for the persistence of bufadienolides, juvenile snakes thathad not been exhausted of nuchal gland fluid by the end of the feedingtrial (n = 22) were maintained in the laboratory on a diet of fish(Oryzias latipes and Pseudorasbora parva). Rhabdophis tigrinus is no-toriously difficult to maintain for extended periods, and the average ageat the time of death for the juveniles was 135� 26 days; the longest livedjuvenile died at 184 days of age. Additionally, a single dam was main-tained on a diet of fish after being fed frogs (including two Bufo japo-nicus) in the laboratory.
Samples of prey items fed to hatchling and adult R. tigrinus wereanalyzed chemically. Bufo japonicus metamorphs (Chiba Prefecture)and O. latipes, both of which were used as prey for hatchling R. tigrinus,were prepared as whole-body extracts in methanol. Skin secretionsfrom Pelophylax nigromaculatus, individuals of which were fed to adultR. tigrinus, were collected using a transcutaneous amphibian stimulator(Grant and Land 2002). Parotoid gland secretions from adult B. japo-nicus (from Niijima Island) were collected by manually squeezing theglands. Skin secretions from anurans were collected on sections of Ki-mwipes and stored in methanol at –20 8C. Our animal use protocolsconformed to institutional policies and practices.
182 D. A. Hutchinson et al. CHEMOECOLOGY

Chemical Analyses. Samples of nuchal gland fluid and prey extractswere shipped from Japan to the United States on dry ice for chemicalanalysis. Samples of nuchal gland fluid and skin secretions from anuransrequired no preparation prior to evaporation. The whole-body samplesof prey in methanol (fish and metamorphic toads) were homogenizedwith a glass tissue grinder and filtered prior to chemical analysis. Allsamples were evaporated to dryness, reconstituted in deuteratedmethanol (CD3OD), and analyzed with 1H-NMR on a Unity INOVA600 MHz spectrometer (Varian, Palo Alto, CA, USA) equipped with anOxford magnet (Oxford Instruments, Eynsham, Witney, Oxon, UK)and a 5 mm inverse-detection HCN probe.
Following analysis by 1H-NMR spectroscopy, we evaporated allsamples to dryness and reconstituted them in a standardized volume ofmethanol for analysis with HPLC. Most samples were placed in 300 mLmethanol, but this amount occasionally resulted in samples that weretoo concentrated or inadequately dissolved. To correct for this, we used900 mL of methanol for yolk samples and 1500 mL for parotoid glandsecretions of toads and nuchal gland fluid of adult R. tigrinus thatcontained large quantities of bufadienolides. Samples were analyzedwith an Agilent (Santa Clara, CA, USA) 1100 Series HPLC equippedwith a quaternary pump, diode array detector, and autosampler. Thesamples were fractionated through a reversed phase 25 cm x 10 mmSupelco (Bellefonte, PA, USA) Discovery HS C18 column. We used asolvent gradient starting with a mixture of methanol and water, themethanol content of which was held at 20% (v/v) for two minutes andthen increased linearly over a period of 38 minutes until reaching 100%methanol. After two minutes at 100% methanol, the solvent compo-sition changed back to the initial 20: 80 methanol/water mixture. A flowrate of 3.4 ml/min was used. The injection volumes of the samples were25 mL each, and each run lasted 53 minutes.
To identify bufadienolides in nuchal gland fluid, we compared theretention times of peaks to those produced by the 17 previously iden-tified compounds from nuchal gland fluid of R. tigrinus (Fig. 1b;Hutchinson et al. 2007). Isolated samples of the previously identifiedcompounds were used as standards (Fig. 2). Of the 17 bufadienolides,compounds 6 and 7 co-eluted and thus could not be distinguished fromone another.
To summarize the data for hatchlings with the same feeding his-tories, only those compounds that were present in every individual aredisplayed in the results. Individual bufadienolides were consideredpresent only when the area of their HPLC peak measured greater than15.0 mAU*s (milli-absorbance units times seconds).
To determine the quantities of bufadienolides present in eachsample, we calibrated our HPLC system using a series of dilutions oftelocinobufagin. The equation obtained from the linear trend line fittedto the dilution curve was used to convert the areas (mAU*s) of buf-adienolide peaks at 280 nm into quantities of bufadienolides. Thewavelength of 280 nm was selected because bufadienolides have ab-sorbance maxima at approximately 200 and 300 nm; the latter exhibits amore stable baseline and experiences less interference from the UVabsorbance of methanol. Peaks produced by bufadienolides (bothstructurally known and unknown) were identified by their character-istic UV absorbance spectra, and only those peaks with areas greaterthan 15.0 mAU*s were used to calculate the total quantity of bufadie-nolides per sample of nuchal gland fluid.
We identified the major bufadienolides in parotoid gland secre-tions from adult B. japonicus by using dqCOSY NMR and HPLC.Chain lengths of bufotoxins were confirmed by direct injection into anEsquire-LC electrospray ion trap mass spectrometer (Bruker, Billerica,MA, USA). We compared our findings to structures reported in theliterature (Shimada et al. 1977). Compounds are referred to by numbersin bold type, and those found in toads are identified by the prefix T.
Statistical Analysis. To analyze the relationship of bufadienolides innuchal gland fluid of unfed R. tigrinus dams and their unfed offspring,we performed a nonparametric Spearman correlation analysis usingSPSS version 11.0 (Chicago, IL, USA). The data were plotted usingSigmaPlot version 9.0 (SYSTAT, San Jose, CA, USA).
Results
Prey Chemistry and Preference. The presence or absenceof bufadienolides in species commonly fed to captiveRhabdophis tigrinus was confirmed by 1H-NMR spec-troscopy and HPLC. Bufadienolides were absent in thefish (Oryzias latipes) fed to hatchlings and in a nonbufo-nid frog (Pelophylax nigromaculatus) fed to adults. Buf-adienolides were abundant in the parotoid gland secre-tions of adult Bufo japonicus, the main components ofwhich are shown in Fig. 1a, as determined by dqCOSYNMR, mass spectroscopy, and HPLC. Small quantities(less than 20 mg) of bufadienolides were present in themetamorphic B. japonicus fed to hatchlings.
Most hatchlings in the toad-fed group readily con-sumed the toads, but hatchlings in the fish-fed group oftenrejected the fish and had to be force-fed. Overall, toadswere rejected only 5.6% of the time, whereas fish wererejected in 70 % of feedings. This preference for bufonidprey, as opposed to fish, is consistent with previous find-ings for R. tigrinus (Mori 2004).
Dams Fed Nonbufonid Prey. Six gravid, wild-caught R.tigrinus females were fed only nonbufonid prey in thelaboratory. Dams 7 and 9 possessed less than 5 mg ofbufadienolides per sample of nuchal gland fluid andproduced offspring that lacked these compounds (Fig. 1b,Table 1). Samples of nuchal gland fluid from dams 1 and 4contained bufadienolides in quantities less than 50 mg and0.25 mg, respectively. Unfed hatchlings from clutches 1and 4 possessed less than 50 mg of bufadienolides persample of nuchal gland fluid (Table 1). Samples fromdams 2 and 6 at the time of their capture contained morethan 1.1 mg (up to 1.9 mg) of bufadienolides. Unfedhatchlings from clutches 2 and 6 possessed more than0.35 mg (up to 0.60 mg) of these compounds per sample ofnuchal gland fluid. It is important to note that the abso-lute quantities of bufadienolides reported here representonly a portion of the total amount of bufadienolides in asnake. We expressed only a few nuchal glands at any onetime to allow for repeated sampling of each individual.
Among unfed and fish-fed offspring, compound 10(gamabufotalin) was the principal bufadienolide in allbufadienolide-containing samples (Table 1). Among thedams, however, compound 10 was the most prevalentbufadienolide in only three of nine samples.
Dams Fed Bufonid Prey. Four gravid, wild-caught dams(3, 5, 10, and 11) were fed one toad (Bufo japonicus) each,9–19 days prior to oviposition. Samples of nuchal glandfluid from dams 10 and 11 at the time of their capturecontained 8.3 and 2.4 mg of bufadienolides, respectively(Table 2). Unfed hatchlings from clutch 10 possessedmore than 0.75 mg (up to 1.4 mg) of bufadienolides persample of nuchal gland fluid, whereas unfed hatchlingsfrom dam 11 possessed more than 0.25 mg of bufadieno-lides per sample (Table 2). At the time of their capture,dams 3 and 5 possessed more than 0.30 and 0.50 mg of
Maternal Provisioning in RhabdophisVol. 18, 2008 183

bufadienolides, respectively, per sample of nuchal glandfluid (Table 2). Unfed hatchlings from clutches 3 and 5possessed more than 0.15 mg (up to 0.30 mg) of bufadie-nolides per sample (Table 2).
Some of the gravid dams that were fed a toad se-questered bufadienolides from that prey item that theydid not possess previously. One of these newly accumu-
lated bufadienolides (compound 14) was detected in thenuchal gland fluid of their offspring as well. Dams 3 and11 accumulated compound 14 from a toad consumed12–19 days prior to oviposition; this bufadienolide wasdetected in the nuchal gland fluid of all of their offspringtested (Fig. 3, Table 2). Compound 14 was lacking in thesample of nuchal gland fluid from dam 5 at the time of her
Fig. 1 (a) Major bufadienolides inthe parotoid gland secretion ofBufo japonicus, as determined byNMR, HPLC, and MS. Com-pounds T3, T5, and T7 are knownalso from skin secretions of NorthAmerican toads. Note the correct-ed chain length of T5 (Hutchinsonet al. 2007). (b) Bufadienolides inthe nuchal gland fluid of Rhabdo-phis tigrinus. Compound 8 is 11a-hydroxytelocinobufagin, com-pound 10 is gamabufotalin, com-pound 13 hellebrigenin, and com-pound 17 is telocinobufagenin.
184 D. A. Hutchinson et al. CHEMOECOLOGY
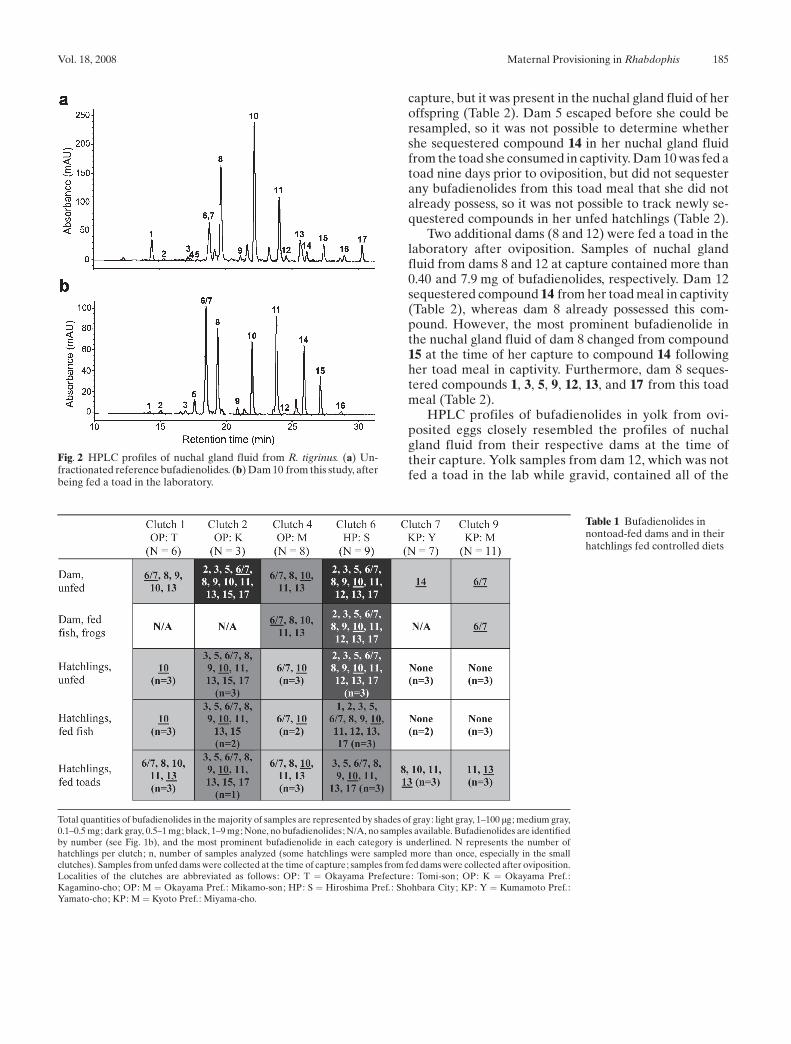
capture, but it was present in the nuchal gland fluid of heroffspring (Table 2). Dam 5 escaped before she could beresampled, so it was not possible to determine whethershe sequestered compound 14 in her nuchal gland fluidfrom the toad she consumed in captivity. Dam 10 was fed atoad nine days prior to oviposition, but did not sequesterany bufadienolides from this toad meal that she did notalready possess, so it was not possible to track newly se-questered compounds in her unfed hatchlings (Table 2).
Two additional dams (8 and 12) were fed a toad in thelaboratory after oviposition. Samples of nuchal glandfluid from dams 8 and 12 at capture contained more than0.40 and 7.9 mg of bufadienolides, respectively. Dam 12sequestered compound 14 from her toad meal in captivity(Table 2), whereas dam 8 already possessed this com-pound. However, the most prominent bufadienolide inthe nuchal gland fluid of dam 8 changed from compound15 at the time of her capture to compound 14 followingher toad meal in captivity. Furthermore, dam 8 seques-tered compounds 1, 3, 5, 9, 12, 13, and 17 from this toadmeal (Table 2).
HPLC profiles of bufadienolides in yolk from ovi-posited eggs closely resembled the profiles of nuchalgland fluid from their respective dams at the time oftheir capture. Yolk samples from dam 12, which was notfed a toad in the lab while gravid, contained all of the
Fig. 2 HPLC profiles of nuchal gland fluid from R. tigrinus. (a) Un-fractionated reference bufadienolides. (b) Dam 10 from this study, afterbeing fed a toad in the laboratory.
Total quantities of bufadienolides in the majority of samples are represented by shades of gray: light gray, 1–100 mg; medium gray,0.1–0.5 mg; dark gray, 0.5–1 mg; black, 1–9 mg; None, no bufadienolides; N/A, no samples available. Bufadienolides are identifiedby number (see Fig. 1b), and the most prominent bufadienolide in each category is underlined. N represents the number ofhatchlings per clutch; n, number of samples analyzed (some hatchlings were sampled more than once, especially in the smallclutches). Samples from unfed dams were collected at the time of capture; samples from fed dams were collected after oviposition.Localities of the clutches are abbreviated as follows: OP: T = Okayama Prefecture: Tomi-son; OP: K = Okayama Pref.:Kagamino-cho; OP: M = Okayama Pref.: Mikamo-son; HP: S = Hiroshima Pref. : Shohbara City; KP: Y = Kumamoto Pref.:Yamato-cho; KP: M = Kyoto Pref.: Miyama-cho.
Table 1 Bufadienolides innontoad-fed dams and in theirhatchlings fed controlled diets
Maternal Provisioning in RhabdophisVol. 18, 2008 185

bufadienolides found in her nuchal gland fluid (Fig. 4).Most of these bufadienolides also were detected in thenuchal gland fluid of her unfed hatchlings (Fig. 4). Theyolks of the other sampled clutches (3, 10, and 11) con-tained all of the major bufadienolides found in the nu-chal gland fluid of the respective unfed dams; thesecompounds were detected in the nuchal gland fluid oftheir offspring as well. Dams 3 and 11 sequesteredcompound 14 from a toad meal in captivity (consumedduring gestation), and small quantities of this bufadie-nolide were detected in samples of yolk taken from theiroviposited eggs.
Comparison of Bufadienolides in Dams and Their Off-spring. A Spearman correlation analysis revealed a sig-nificant, positive relationship between the quantities ofbufadienolides in the nuchal gland fluid from unfeddams and their unfed offspring (r = 0.939; p < 0.0005;Fig. 5).
The dominant bufadienolide in the dams was morevariable than that among the hatchlings, and the bufadie-nolides sequestered from adult and metamorphic B. japo-nicus differed. The major bufadienolide in most unfeddams was compound 6 or 7 (n = 6), and the second mostprominent bufadienolide was compound 10 (gamabufota-lin; n = 3). In contrast, the dominant bufadienolide in
hatchlings, regardless of feeding treatment, was almostalways compound 10 (72 of 83 samples; see Fig. 4). Amongthe dams that were fed a toad in captivity, compound 14was the most frequently sequestered bufadienolide thatwas not possessed previously. In hatchlings, compounds 8,11, and 13 were the most common newly acquired buf-adienolides sequestered from metamorphic toads.
Persistence of Bufadienolides in Hatchling and AdultSnakes. Samples collected from fish-fed juvenile snakesthat were maintained in the laboratory for extended pe-riods revealed that bufadienolides persist in the nuchalglands for at least six months. The two longest-livedhatchlings, which were fed only fish, were from a clutchlaid by a female from toad-rich Ishima Island (clutch 12);they lived for 184 and 180 days. These individuals had upto 3.1 mg of bufadienolides in their nuchal glands postmortem. Snakes from mainland Shikoku and Honshu alsoexhibited lengthy persistence of bufadienolides in theirnuchal gland fluid. Fish-fed juveniles from these locationsthat died at 107–150 days of age possessed up to 1.3 mg ofbufadienolides.
Despite the lengthy persistence of bufadienolides ingeneral, compounds present in small quantities at thetime of hatching diminished within a matter of weeks.Furthermore, the total quantity of bufadienolides in the
Total quantities of bufadienolides in the majority of samples are represented by shades of gray: light gray, 1–100 mg; medium gray,0.1–0.5 mg; dark gray, 0.5 –1 mg; black, 1–9 mg; N/A, no samples available. Bufadienolides are identified by number (see Fig. 1b),and the most prominent bufadienolide per category is underlined (inconsistent for clutch 12). N represents the number ofhatchlings per clutch; n, number of samples analyzed (some hatchlings were sampled more than once, especially in the smallclutches). The only adults fed toads prior to oviposition were dams 3, 5, 10, and 11; the number of days prior to oviposition that thetoad was consumed is indicated in the top row for these four dams. Samples from unfed dams were collected at the time of capture;samples from fed dams were collected after oviposition. Localities of the clutches are abbreviated as follows: OP: M = OkayamaPrefecture: Mikamo-son; OP: S = Okayama Pref.: Shinjo-son; KP: M = Kyoto Pref.: Miyama-cho; TP: N = Tokushima Pref.:Naka-gun; TP: I = Tokushima Pref.: Ishima Island.
Table 2 Bufadienolides intoad-fed dams and in theirhatchlings fed controlled diets
186 D. A. Hutchinson et al. CHEMOECOLOGY

nuchal gland fluid either remained the same or decreasedafter the offspring were fed fish. Among defendedhatchlings, toad-fed individuals typically possessed thesame quantity of bufadienolides as the fish-fed hatchlings,although the diversity of bufadienolide compounds oftenwas greater in the toad-fed group.
Hatchlings that were fed nonbufonid diets did notsynthesize bufadienolides over time. For example, fish-fed hatchlings from clutch 9, an undefended clutch, stilllacked bufadienolides at the time of their deaths at up to155 days of age.
Bufadienolides also showed persistence in the singleadult snake from Ishima Island (dam 12) that was main-tained in captivity. A sample collected from this individ-ual 286 days after she consumed her last toad meal in thelaboratory contained 16 mg of bufadienolides. Anothersample taken 405 days after her last toad meal contained9.9 mg of bufadienolides.
Discussion
Our results show that Rhabdophis tigrinus females pro-vision their offspring with sequestered bufadienolides.Hatchlings from dams with less than 100 mg of bufadie-
nolides per sample of nuchal gland fluid either lackedbufadienolides completely or possessed small quantitiesthat are unlikely to be biologically relevant in antipre-dator defense. In contrast, dams with large quantities ofbufadienolides in their own nuchal gland fluid consis-tently produced offspring with high levels of these de-fensive compounds.
Despite the positive relationship in the total quanti-ties of bufadienolides between dams and their offspring,maternal provisioning in R. tigrinus appears to be a se-lective process. Selectivity in the sequestration of de-fensive compounds has been reported in chrysomelidbeetles that store specific pyrrolizidine alkaloids fromplants (Hartmann et al. 1997). Although quantities ofindividual bufadienolides in adult R. tigrinus serve as
Fig. 3 Aromatic region of 1H-NMR spectra of nuchal gland fluid fromdam 3 and her offspring. The gray bars highlight the three regionsdiagnostic of bufadienolides. Nineteen days prior to oviposition, thedam was fed a toad, from which she sequestered two bufadienolidesthat she did not possess previously. One of the newly acquired buf-adienolides (arrows; compound 14) was provisioned to her unfed off-spring along with compounds that she possessed prior to that toad meal.
Fig. 4 HPLC profiles of nuchal gland fluid and yolk from clutch 12. Thedam�s nuchal gland fluid contained a large quantity of bufadienolides,all of which were found in the yolks of her oviposited eggs. Most of thesebufadienolides also were evident in the nuchal gland fluid of her unfedhatchlings. Note the prominence of compound 10 (gamabufotalin) inthe nuchal gland fluid of the hatchling, as compared to its prevalence inthe yolk and nuchal gland fluid of the dam.
Maternal Provisioning in RhabdophisVol. 18, 2008 187

general predictors of the compounds that will appear inhatchlings, the provisioning of gamabufotalin (com-pound 10) followed an atypical pattern. Gamabufotalinwas the dominant bufadienolide in 87 % of samples fromhatchlings, whereas this compound typically was not theprincipal bufadienolide in the nuchal gland fluid of thedams.
There are several possible explanations for theprominence of gamabufotalin in the hatchlings. Gama-bufotalin may be provisioned preferentially in yolk overother compounds. However, our data do not support thisexplanation because samples of yolk tended to possessthe same relative quantities of individual bufadienolidesas the nuchal gland fluid of the dams. Another possibleexplanation is that the most prominent bufadienolide ofhatchlings represents the most abundant compound in thedam at the time of vitellogenesis, perhaps reflecting herdiet at that time. However, it is unlikely that all of thedams would have had gamabufotalin as their mostprominent bufadienolide during vitellogenesis, given thevariation in the composition of nuchal gland fluid amongdams in this study. A more likely hypothesis is that gam-abufotalin may be absorbed from the yolk and trans-ported to the nuchal glands of embryos more readily thanother bufadienolides. Other processes cannot be ruledout, however, such as the modification of provisionedbufadienolides by embryos, resulting in the conversion ofother compounds to gamabufotalin. Regardless of thereason for the pattern, it may be beneficial to hatchling R.tigrinus to have gamabufotalin as their dominant buf-adienolide. This compound is approximately 10 timesmore effective at inhibiting the sodium-potassium pumpin canine kidney than ouabain (Azuma et al. 1986), andthus may be more effective as an antipredator compoundthan other bufadienolides. However, the physiologicalactivities of most of the other bufadienolides remain to bedetermined.
Sequestration and provisioning of bufadienolides byR. tigrinus may involve chemical modification, which hasbeen reported for other species that rely on sequestereddefensive compounds. Sequestered lucibufagins are hy-droxylated by fireflies (Gonz�lez et al. 1999b), and thetoxicity of sequestered alkaloids is enhanced by hy-droxylation in dendrobatid frogs (Daly et al. 2003). In R.tigrinus, gamabufotalin (T7 and 10 ; Fig. 1) may be se-questered unaltered by adults and hatchlings, or it may bederived from side-chain hydrolysis of a bufotoxin (T3).Similarly, compound 14, which was sequestered by sev-eral dams after their toad meal in the laboratory, mayresult from side-chain hydrolysis and subsequent hy-droxylation of T11. Side-chain hydrolysis and hydroxy-lation of T5 may give rise to compounds 7 and 11; all threeare hydroxyketones.
Bufadienolides present in the nuchal gland fluid offish-fed hatchlings, representing maternally provisionedcompounds, persist for at least six months. This findingsuggests that maternal provisioning alone can providechemical protection from the time of hatching in latesummer until the following spring. By the time R. tigrinushatch, the toads that had metamorphosed the previousspring may be too large to be consumed (Urano andIshihara 1987). Recent field observations suggest thathatchlings can, in at least some cases, consume juveniletoads (DAH, AHS and AM, personal observation).However, hatchling snakes that are unable to locate orconsume juvenile toads must rely on bufadienolidesprovisioned by the dam for chemical defense until newlymetamorphosed toads become available the followingspring. Such reliance would be possible based upon thepersistence of bufadienolides demonstrated here. Fur-thermore, our results indicate that metamorphic toadscan serve as a source of bufadienolides for young R. ti-grinus, although the quantities of those compounds inmetamorphs are small.
Importantly, our results demonstrate that toads con-sumed by a female during gestation can supply bufadie-nolides to her late-stage embryos, as well as to her ownnuchal gland fluid. The eggs of oviparous squamatesgenerally undergo about one-third of their developmentin the oviducts, prior to oviposition (Shine 1983; Andrewsand Mathies 2000). In the present study, newly acquiredbufadienolides were provisioned to offspring 12–19 daysbefore oviposition. At that stage of gestation, the eggspresumably are in the oviducts of the female, so anytransfer of toxins at that time would likely occur by dif-fusion across the egg membranes, and probably the intacteggshell, which is laid down during passage through theoviducts (Andrews and Mathies 2000). The presence oflarge quantities of bufadienolides in yolks of fertile eggsindicates that those compounds are provisioned by de-position in yolk as well, affording two opportunities forprovisioning to occur.
Our results suggest that maternal provisioning of se-questered bufadienolides is a common phenomenon, butnot a universal one, among R. tigrinus. Provisioned buf-
Fig. 5 Quantities of bufadienolides in nuchal gland fluid samples ofdams (prior to feeding in the laboratory) and their unfed hatchlings,showing mean and standard deviation for each clutch. The Spearmancorrelation coefficient is 0.939 (p < 0.0005).
188 D. A. Hutchinson et al. CHEMOECOLOGY

adienolides become incorporated into embryonic tissues,so that hatchlings are likely to be chemically defendedagainst predators. The presence of bufadienolides in yolkindicates that dams can transmit dietary bufadienolides totheir offspring during vitellogenesis, although it is un-known whether the compounds must be ingested duringvitellogenesis or whether they can be stored from earliermeals and mobilized later, when the yolk is deposited inthe developing ova. In addition, some provisioning canoccur long after vitellogenesis is complete, when the fullyyolked eggs pass through the oviducts. Hatchlings arecapable of sequestering additional bufadienolides fromingested toads, if such prey items are available to them. Iftoads are unavailable, maternally provisioned bufadie-nolides will play an especially important role in the abilityof juvenile snakes to deter predators. Given the vulner-ability of hatchling snakes to predation (Burger 1998), thepresence of defensive toxins presumably is an importantcontributor to offspring survival, and thus maternal pro-visioning is expected to be subject to strong selectivepressure.
Acknowledgments
We thank Mark J. Butler IV, Dayanand N. Naik, andChristopher A. Binckley for statistical advice; Stephen T.Deyrup for assistance with mass spectroscopy; Thomas F.Spande and John W. Daly for supplying telocinobufagin;Yohei Kadota, Akira Katayama, Noriko Kidera, ToshiroKuroki, Masami Hasegawa, Koji Mochida, Aya Nakadai,Sumio Okada, and Hirohiko Takeuchi for collecting an-imals in the field; Yohei Kadota, Noriko Kidera, TakashiHaramura, and Koji Tanaka for collecting some samplesof nuchal gland fluid; Tatsuya Hishida and Koji Tanakafor husbandry; R. James Swanson for use of a vacuumpump; and two anonymous reviewers for helpful com-ments that greatly improved the manuscript. This workwas supported by the National Science Foundation (IBN-0429223 and IOB-0519458 to AHS and JM); Old Do-minion University Dissertation Fellowship (to DAH); theSociety for Integrative and Comparative Biology andSigma Xi, The Scientific Research Society (Grants-in-Aidof Research to DAH); The Honor Society of Phi KappaPhi (Love of Learning Award to DAH); Kyoto UniversityGrants for the 21st Century COE A14 and the GlobalCOE Program A06 (to AM); and the Japan Society forthe Promotion of Science (to AM).
References
Akizawa T, Yasuhara T, Azuma H, Nakajima T (1985a) Chemicalstructures and biological activities of bufodienolides in the nucho-dorsal glands of Japanese snake, Rhabdophis tigrinus. J Pharma-cobio-Dyn 8: s–60
Akizawa T, Yasuhara T, Kano R, Nakajima T (1985b) Novel poly-hydroxylated cardiac steroids in the nuchal glands of the snake,Rhabdophis tigrinus. Biomed Res 6: 437–441
Andrews RM, Mathies T (2000) Natural history of reptilian develop-ment: Constraints on the evolution of viviparity. BioScience 50:227–238
Asahi H, Kohtari Y, Chiba K, Mishima A (1985) Effect of the nucho-dorsal gland venom of the yamakagashi snake on the eye. FoliaOphthalmol Japon 36: 379–383
Azuma H, Sekizaki S, Akizawa T, Yasuhara T, Nakajima T (1986)Activities of novel polyhydroxylated cardiotonic steroids purifiedfrom nuchal glands of the snake, Rhabdophis tigrinus. J PharmPharmacol 38: 388–390
Benard MF, Fordyce JA (2003) Are induced defenses costly? Conse-quences of predator-induced defenses in western toads, Bufo bo-reas. Ecology 84: 68–78
Blum MS, Hilker M (2002) Chemical protection of insect eggs. Pp 61–90in Hilker M, Meiners T (eds) Chemoecology of Insect Eggs andEgg Deposition. Malden: Blackwell Publishing
Brower LP (1984) Chemical defence in butterflies. Pp 109–134 in Vane-Wright RI, Ackery PR (eds) The Biology of Butterflies. London:Academic Press
Burger J (1998) Antipredator behaviour of hatchling snakes: Effects ofincubation temperature and simulated predators. Anim Behav 56:547–553
Che J, Pang J, Zhao H, Wu GF, Zhao EM, Zhang YP (2007) Phylogenyof Raninae (Anura: Ranidae) inferred from mitochondrial andnuclear sequences. Mol Phylogenet Evol 43: 1–13
Daly JW, Garraffo HM, Spande TF, Clark VC, Ma J, Ziffer H, Cover JFJr (2003) Evidence for an enantioselective pumiliotoxin 7-hy-droxylase in dendrobatid poison frogs of the genus Dendrobates.Proc Natl Acad Sci USA 100: 11092–11097
Daly JW, Garaffo HM, Spande T F, Jaramillo C, Rand AS (1994) Di-etary source for skin alkaloids of poison frogs (Dendrobatidae)? JChem Ecol 20: 943–955
Doull J, Dubois KP, Geiling EMK (1951) The biosynthesis of radioac-tive bufagin. Arch Int Pharmacodyn Ther 86: 454–464
Duffey SS (1980) Sequestration of plant natural products by insects.Annu Rev Entomol 25: 447–477
Dumbacher JP, Wako A, Derrickson SR, Samuelson A, Spande TF,Daly JW (2004) Melyrid beetles (Choresine): A putative source forthe batrachotoxin alkaloids found in poison-dart frogs and toxicpasserine birds. Proc Natl Acad Sci USA 101: 15857–15860
Eisner T, Eisner M, Rossini C, Iyengar VK, Roach BL, Benedikt E,Meinwald J (2000) Chemical defense against predation in an insectegg. Proc Natl Acad Sci USA 97: 1634–1639
Eisner T, Goetz MA, Hill DE, Smedley SR, Meinwald J (1997) Firefly“femmes fatales” acquire defensive steroids (lucibufagins) fromtheir firefly prey. Proc Natl Acad Sci USA 94: 9723–9728
Eisner T, Meinwald J (1995) The chemistry of sexual selection. ProcNatl Acad Sci USA 92: 50–55
Eisner T, Rossini C, Gonz�lez A, Iyengar VK, Siegler MVS, SmedleySR (2002) Paternal investment in egg defence. Pp 91–116 in HilkerM, Meiners T (eds) Chemoecology of Insect Eggs and Egg Depo-sition. Malden: Blackwell Publishing
Erspamer V (1994) Bioactive secretions of the amphibian integument.Pp 178–350 in Heatwole H, Barthalmus GT, Heatwole AY (eds)Amphibian Biology. Chipping Norton: Surrey Beatty and Sons
Frost DR, Grant T, Faivovich J, Bain RH, Haas A, Haddad CFB, DesaRO, Channing A, Wilkinson M, Donnellan SC, Raxworthy CJ,Campbell JA, Blotto BL, Moler P, Drewes RC, Nussbaum RA,Lynch JD, Green DM, Wheeler WC (2006) The amphibian tree oflife. Bull Am Mus Nat Hist 297: 1–291
Gonz�lez A, Hare JF, Eisner T (1999a) Chemical egg defense in Pho-turis firefly ”femmes fatales.” Chemoecology 9: 177–185
Gonz�lez A, Schroeder FC, Attygalle AB, Svatos A, Meinwald J, Eis-ner T (1999b) Metabolic transformations of acquired lucibufaginsby firefly ”femmes fatales.” Chemoecology 9: 105–112
Grant JB, Land B (2002) Transcutaneous amphibian stimulator (TAS):A device for collection of amphibian skin secretions. Herpetol Rev33: 38–41
Hanifin CT, Brodie ED III, Brodie ED Jr (2003) Tetrodotoxin levels ineggs of the rough-skin newt, Taricha granulosa, are correlated withfemale toxicity. J Chem Ecol 29: 1729–1739
Maternal Provisioning in RhabdophisVol. 18, 2008 189

Hartmann T, Witte L, Ehmke A, Theuring C, Rowell-Rahier M,Pasteels JM (1997) Selective sequestration and metabolism of plantderived pyrrolizidine alkaloids by chrysomelid leaf beetles. Phy-tochemistry 45: 489–497
Hutchinson DA, Mori A, Savitzky AH, Burghardt GM, Wu X, Mein-wald J, Schroeder FC (2007) Dietary sequestration of defensivesteroids in nuchal glands of the Asian snake Rhabdophis tigrinus.Proc Natl Acad Sci USA 104: 2265–2270
Kawashima J (1957) Disturbance of the eye by snake venom (Natrixtigrina). Jpn Rev Clin Ophthalmol 50: 837–839 (in Japanese)
Kawashima J (1959) Disturbance of the eye by snake venom (Natrixtigrina) II. Jpn Rev Clin Ophthalmol 53: 834–837 (in Japanese)
Kitazume Y (1953) On the increasing action of nuchal gland inclusionsof the snake, Natrix tigrina, upon the blood pressure of the rabbit.Zool Mag 62: 225–227
Licht LE (1968) Unpalatability and toxicity of toad eggs. Herpetologica24: 93–98
Melero CP, Medarde M, San Feliciano A (2000) A short review oncardiotonic steroids and their aminoguanidine analogues. Mole-cules 2000: 51–81
Mori A (2004) Another venom gland in snakes: Function, ultrastruc-ture, and physiology of the nuchal glands of Rhabdophis tigrinus.Bull Herpetol Soc Japan 2004: 29–33 (In Japanese)
Mori A, Moriguchi H (1988) Food habits of snakes in Japan: A criticalreview. Snake 20: 98–113
Nakamura K (1935) On a new integumental poison gland found in thenuchal region of a snake, Natrix tigrina. Mem Coll Sci Kyoto ImpUniv 10B: 229–240
Nishida R (2002) Sequestration of defensive substances from plants byLepidoptera. Annu Rev Entomol 47: 57–92
Pasteels JM, Daloze D, Rowell-Rahier M (1986) Chemical defence inchrysomelid eggs and neonate larvae. Physiol Entomol 11: 29–37
Pavelka LA, Kim YH, Mosher HS (1977) Tetrodotoxin and tetrodo-toxin-like compounds from the eggs of the Costa Rica frog, Atel-opus chiriquiensis. Toxicon 15: 135–139
Phisalix M (1922) Animaux Venimeux et Venins. Paris: Masson CieEditeurs
Porto AM, Baralle FE, Gros EG (1972) Biosynthesis of bufadienolidesin toads III – Experiments with [2– 14C]mevalonic acid, [20– 14C]3b-hydroxy-5-pregnen-20-one and [20– 14C]cholesterol. J SteroidBiochem 3: 11–17
Porto AM, Gros EG (1971) Biosynthesis of the bufadienolide mari-nobufagin in toads Bufo paracnemis from cholesterol-20–14C.Experientia 27: 506
Saporito RA, Garraffo, HM, Donnelly, MA, Edwards, AL, Longino JT,Daly JW (2004) Formicine ants: An arthropod source for thepumiliotoxin alkaloids of dendrobatid poison frogs. Proc Natl AcadSci USA 101: 8045–8050
Shimada K, Fujii Y, Yamashita E, Niizaki Y, Sato Y, Nambara T (1977)Studies on cardiotonic steroids from the skin of Japanese toad.Chem Pharmaceutical Bull (Tokyo) 25: 714–730
Shine R (1983) Reptilian reproductive modes: The oviparity-viviparitycontinuum. Herpetologica 39: 1–8
Siperstein MD, Murray AW, Titus E (1957) Biosynthesis of cardiotonicsterols from cholesterol in the toad, Bufo marinus. Arch BiochemBiophys 67: 154–160
Smith MA (1938). The nucho-dorsal glands of snakes. Proc Zool SocLond B 107: 575–583, pl I
Suzuki R (1960) Disturbance of the eye by snake venoms (Natrix tigrina(Boie)). J Clin Ophthal 14: 1384–1387
Toriba M, Sawai Y (1990) Venomous snakes of medical importance inJapan, pp 323–347 in Gopalakrishnakone P, Chou LM (eds) Snakesof Medical Importance (Asia-Pacific Region). Venom and ToxinResearch Group, Singapore: National University of Singapore
Twitty VC (1937) Experiments on the phenomenon of paralysis pro-duced by a toxin occurring in Triturus embryos. J Exp Zool 76:67–104
Urano A, Ishihara K (eds) (1987) Biology of Toads. Tokyo: Shokabo (InJapanese)
Williams BL, Brodie ED Jr, Brodie ED III (2004) A resistant predatorand its toxic prey: Persistence of newt toxin leads to poisonous (notvenomous) snakes. J Chem Ecol 30: 1901–1919
Received 20 January 2008; accepted 4 March 2008Published Online First 19 March 2008
To access this journal online:www.birkhauser.ch/chemo
190 D. A. Hutchinson et al. CHEMOECOLOGY
Copyright © 2022 FDOKUMEN




















