
The cytotoxic lipid peroxidation product 4-hydroxy-2-nonenal covalently modifies a selective range...
12
The Cytotoxic Lipid Peroxidation Product 4-Hydroxy-2-nonenal Covalently Modifies a Selective Range of Proteins Linked to Respiratory Function in Plant Mitochondria * □ S Received for publication, March 20, 2007, and in revised form, October 17, 2007 Published, JBC Papers in Press, October 18, 2007, DOI 10.1074/jbc.M702385200 Alison M. Winger ‡1 , Nicolas L. Taylor ‡2 , Joshua L. Heazlewood ‡2 , David A. Day § , and A. Harvey Millar ‡3 From the ‡ Australian Research Council Centre of Excellence in Plant Energy Biology, the University of Western Australia, 35 Stirling Highway, Crawley 6009, Western Australia and § Australian Research Council Centre of Excellence in Plant Energy Biology, School of Biological Sciences, University of Sydney, 2006 New South Wales, Australia Plants encounter a variety of environmental stresses that affect their cellular machinery and that they adapt to on a daily basis. Lipid peroxidation is one consequence, at the cel- lular level, of such stresses and yields cytotoxic lipid alde- hydes, including 4-hydroxy-2-nonenal (HNE), that react with specific sites in proteins, leading to diverse changes in protein function and/or stability. We have assessed the sensitivity of plant mitochondrial proteins to HNE modification, using one-dimensional and two-dimensional denaturing PAGE and blue native-PAGE coupled to immunological detection and tandem mass spectrometry identification. A select range of proteins was modified by exogenous application of HNE to mitochondria isolated from Arabidopsis cell cultures. These included a number of proteins that directly interact with the ubiquinone pool, as well as a number of soluble matrix pro- teins. Mitochondria isolated from cell cultures following hydrogen peroxide, antimycin A, or menadione treatment had significantly reduced respiratory capacity and elevated levels of HNE adduction to specific subsets of proteins. Tar- gets identified included the proteins affected by direct appli- cation of HNE but also some new proteins, including a num- ber of matrix dehydrogenases, the inner membrane adenine nucleotide translocator, and the outer membrane voltage-de- pendent anion channel. Degradation products of some pro- teins were also found to be HNE adducted, suggesting a link between HNE adduction and protein turnover. Some of the major enzyme complexes that were HNE adducted did not show demonstrable changes in their maximal activity meas- ured with artificial acceptors, but changes did occur in asso- ciations between respiratory chain complexes following stress treatments. The polyunsaturated fatty acids of membrane phospholipids are highly susceptible to peroxidation by reactive oxygen spe- cies (ROS), 4 and a self-propagating chain of free radical reac- tions can produce various aldehydes, alkenals, and hydroxyalk- enals (1, 2). These aldehydes are cytotoxic, generally more stable than ROS, and can cause extensive damage to proteins and other cellular constituents. HNE is the most abundant and toxic aldehyde generated through ROS-mediated peroxidation of abundant lipids in plants such as linoleic acid (1, 3). HNE is a highly reactive electrophile, with the primary reactivity of the molecule lying at the unsaturated bond of the C-3 atom. HNE has been shown to form Michael adducts via this C-3 atom with the sulfhydryl group of Cys residues, the imidazole group of His residues, and the -amino group of Lys residues on a large num- ber of proteins (3). Recently it has been proposed that HNE can also modify Arg residues of proteins (4). In addition to Michael adduct formation, Lys residues also form Schiff bases and pen- tylpyrrole adducts with HNE via the C-1 aldehyde group (5). HNE has also been shown to react via the C-3 position with the sulfhydryl groups of lipoic acid moieties on proteins (6). Mod- ification of proteins by HNE has the potential to have serious detrimental effects in a cell because of modification of amino acids and the potential to form cross-links in proteins (1–5). Investigations of the effects of HNE in mammalian mito- chondria show that HNE can inhibit proteins involved in the respiratory process. Complex IV activity is inhibited by HNE in a dose-dependent manner in mammals (7, 8), predominantly by binding to subunit VIII of the complex (9). Cytochrome c (4) and the subunit of the succinate dehydrogenase (complex II) (10, 11) have also been shown to be inhibited by HNE adduction in mammals. Complex I subunits have been shown to be adducted by HNE in heart mitochondria (12), but although * This work was supported in part by grants from the Australian Research Council through the Centres of Excellence Program (to A. H. M. and D. A. D.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental Tables S1–S7. 1 Recipient of an Australian postgraduate award. 2 Supported as Australian Research Council postdoctoral fellow. 3 Supported as Australian Research Council professorial fellow. To whom cor- respondence should be addressed. Tel.: 61-8-6488-7245; Fax: 61-8-6488- 4401; E-mail: [email protected]. 4 The abbreviations used are: ROS, reactive oxygen species; ANT, adenine nucleotide translocator; AOX, alternative oxidase; DCPIP, dichlorophe- nylindophenol; FeCN, [Fe(CN) 6 ] 3 ; HNE, 4-hydroxy-2-nonenal; IEF, isoelec- tric focusing; KGDC, -ketoglutarate dehydrogenase complex; n-PG, n-propyl gallate; PDC, pyruvate dehydrogenase complex; TEMED, N,N,N,N-tetramethylethylenediamine; UQ, ubiquinone; UQH2, reduced ubiquinone; BisTris, 2-[bis(2-hydroxyethyl)amino]-2-(hydroxymethyl)pro- pane-1,3-diol; CI, complex I; HAE, hydroxyalkenal; MS/MS, tandem mass spectrometry; LC, liquid chromatography; TES, 2-{[2-hydroxy-1,1- bis(hydroxymethyl)ethyl]amino}ethanesulfonic acid; CHAPS, 3-[(3-chol- amidopropyl)dimethylammonio]-1-propanesulfonic acid; Tricine, N-[2-hy- droxy-1,1-bis(hydroxymethyl)ethyl]glycine; BN, blue native; NL, nonlinear. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 52, pp. 37436 –37447, December 28, 2007 © 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 37436 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 52 • DECEMBER 28, 2007 at Univ of Western Australia, on May 7, 2012 www.jbc.org Downloaded from http://www.jbc.org/content/suppl/2007/11/07/M702385200.DC1.html Supplemental Material can be found at:
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The cytotoxic lipid peroxidation product 4-hydroxy-2-nonenal covalently modifies a selective range...

The Cytotoxic Lipid Peroxidation Product4-Hydroxy-2-nonenal Covalently Modifies a SelectiveRange of Proteins Linked to Respiratory Functionin Plant Mitochondria*□S
Received for publication, March 20, 2007, and in revised form, October 17, 2007 Published, JBC Papers in Press, October 18, 2007, DOI 10.1074/jbc.M702385200
Alison M. Winger‡1, Nicolas L. Taylor‡2, Joshua L. Heazlewood‡2, David A. Day§, and A. Harvey Millar‡3
From the ‡Australian Research Council Centre of Excellence in Plant Energy Biology, the University of Western Australia, 35 StirlingHighway, Crawley 6009, Western Australia and §Australian Research Council Centre of Excellence in Plant Energy Biology, Schoolof Biological Sciences, University of Sydney, 2006 New South Wales, Australia
Plants encounter a variety of environmental stresses thataffect their cellular machinery and that they adapt to on adaily basis. Lipid peroxidation is one consequence, at the cel-lular level, of such stresses and yields cytotoxic lipid alde-hydes, including 4-hydroxy-2-nonenal (HNE), that react withspecific sites in proteins, leading to diverse changes in proteinfunction and/or stability. We have assessed the sensitivity ofplant mitochondrial proteins to HNE modification, usingone-dimensional and two-dimensional denaturing PAGE andblue native-PAGE coupled to immunological detection andtandem mass spectrometry identification. A select range ofproteins was modified by exogenous application of HNE tomitochondria isolated from Arabidopsis cell cultures. Theseincluded a number of proteins that directly interact with theubiquinone pool, as well as a number of soluble matrix pro-teins. Mitochondria isolated from cell cultures followinghydrogen peroxide, antimycin A, or menadione treatmenthad significantly reduced respiratory capacity and elevatedlevels of HNE adduction to specific subsets of proteins. Tar-gets identified included the proteins affected by direct appli-cation of HNE but also some new proteins, including a num-ber of matrix dehydrogenases, the inner membrane adeninenucleotide translocator, and the outer membrane voltage-de-pendent anion channel. Degradation products of some pro-teins were also found to be HNE adducted, suggesting a linkbetween HNE adduction and protein turnover. Some of themajor enzyme complexes that were HNE adducted did notshow demonstrable changes in their maximal activity meas-ured with artificial acceptors, but changes did occur in asso-ciations between respiratory chain complexes followingstress treatments.
The polyunsaturated fatty acids of membrane phospholipidsare highly susceptible to peroxidation by reactive oxygen spe-cies (ROS),4 and a self-propagating chain of free radical reac-tions can produce various aldehydes, alkenals, and hydroxyalk-enals (1, 2). These aldehydes are cytotoxic, generally morestable than ROS, and can cause extensive damage to proteinsand other cellular constituents. HNE is the most abundant andtoxic aldehyde generated through ROS-mediated peroxidationof abundant lipids in plants such as linoleic acid (1, 3). HNE is ahighly reactive electrophile, with the primary reactivity of themolecule lying at the unsaturated bond of the C-3 atom. HNEhas been shown to formMichael adducts via this C-3 atomwiththe sulfhydryl group of Cys residues, the imidazole group of Hisresidues, and the �-amino group of Lys residues on a large num-ber of proteins (3). Recently it has been proposed that HNE canalso modify Arg residues of proteins (4). In addition toMichaeladduct formation, Lys residues also form Schiff bases and pen-tylpyrrole adducts with HNE via the C-1 aldehyde group (5).HNE has also been shown to react via the C-3 position with thesulfhydryl groups of lipoic acid moieties on proteins (6). Mod-ification of proteins by HNE has the potential to have seriousdetrimental effects in a cell because of modification of aminoacids and the potential to form cross-links in proteins (1–5).Investigations of the effects of HNE in mammalian mito-
chondria show that HNE can inhibit proteins involved in therespiratory process. Complex IV activity is inhibited by HNE ina dose-dependentmanner inmammals (7, 8), predominantly bybinding to subunit VIII of the complex (9). Cytochrome c (4)and the � subunit of the succinate dehydrogenase (complex II)(10, 11) have also been shown to be inhibited byHNE adductionin mammals. Complex I subunits have been shown to beadducted by HNE in heart mitochondria (12), but although
* This work was supported in part by grants from the Australian ResearchCouncil through the Centres of Excellence Program (to A. H. M. andD. A. D.). The costs of publication of this article were defrayed in part bythe payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Tables S1–S7.
1 Recipient of an Australian postgraduate award.2 Supported as Australian Research Council postdoctoral fellow.3 Supported as Australian Research Council professorial fellow. To whom cor-
respondence should be addressed. Tel.: 61-8-6488-7245; Fax: 61-8-6488-4401; E-mail: [email protected].
4 The abbreviations used are: ROS, reactive oxygen species; ANT, adeninenucleotide translocator; AOX, alternative oxidase; DCPIP, dichlorophe-nylindophenol; FeCN, [Fe(CN)6]3�; HNE, 4-hydroxy-2-nonenal; IEF, isoelec-tric focusing; KGDC, �-ketoglutarate dehydrogenase complex; n-PG,n-propyl gallate; PDC, pyruvate dehydrogenase complex; TEMED,N�,N�,N�,N�-tetramethylethylenediamine; UQ, ubiquinone; UQH2, reducedubiquinone; BisTris, 2-[bis(2-hydroxyethyl)amino]-2-(hydroxymethyl)pro-pane-1,3-diol; CI, complex I; HAE, hydroxyalkenal; MS/MS, tandem massspectrometry; LC, liquid chromatography; TES, 2-{[2-hydroxy-1,1-bis(hydroxymethyl)ethyl]amino}ethanesulfonic acid; CHAPS, 3-[(3-chol-amidopropyl)dimethylammonio]-1-propanesulfonic acid; Tricine, N-[2-hy-droxy-1,1-bis(hydroxymethyl)ethyl]glycine; BN, blue native; NL, nonlinear.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 52, pp. 37436 –37447, December 28, 2007© 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
37436 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 52 • DECEMBER 28, 2007
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from
http://www.jbc.org/content/suppl/2007/11/07/M702385200.DC1.html Supplemental Material can be found at:

complex I-linked respiration is inhibited by HNE in centralnervous system mitochondria, direct assays of complex I usingn-decylubiquinone failed to measure a loss of enzyme complexactivity (10). Other mitochondrial localized proteins in mam-mals that are HNE targets include tricarboxylic acid cycleenzymes KGDC and PDC (6, 13, 14), the adenine nucleotidetranslocator (ANT) (15), heat shock protein 70 (16), and a pro-tein-disulfide isomerase (17). It is interesting to note that not allmodifications by HNE are detrimental or have a measurableeffect on function. For example, increasing modification ofKGDC byHNE in aged rat mitochondria leads to an increase inactivity that correlated with a lowered Km value of the enzymefor �-ketoglutarate (14). Also, HNE has been shown to activateuncoupling protein function in plant and animal mitochondriaby an undefined mechanism (18).Plant mitochondria are known to respond to oxidative stress
by the induction of respiratory bypasses of the phosphorylatingrespiratory chain (19), as well as the induction of antioxidantdefenses such as the ascorbate/glutathione pathway (20) and aspecific mitochondrial peroxiredoxin (21, 22). We have previ-ously shown during oxidative stress in Arabidopsis (21) andenvironmental stress in pea (23) that, in addition to thesedefense responses, specific mitochondrial proteins aredegraded under oxidative stress by an unknown pathway. Inboth cases this degradation was coincident with the loss oflipoic acid cofactors of decarboxylating dehydrogenases, whichwe had previously linked in potato mitochondria to modifica-tion by the lipid peroxidation product HNE (24). We have alsoshown in a detailed investigation of the impact of HNE on res-piratory chain terminal oxidase function in plants that thestress-responsive nonphosphorylating alternative oxidase wasmuch more sensitive to HNE inhibition than the cytochromepathway (25). In that study, a combination of lipid peroxidationassays and the use of antibodies to HNE adducts revealed thathydroxyalkenals (HAEs) are produced in a dark-grown Arabi-dopsis cell culture model in response to H2O2, antimycin A, ormenadione and that this led to an array of HNE-modified pro-tein bands appearing in isolated mitochondrial samples (25).Here we have undertaken a more systematic analysis of thetargets of HNE in plant mitochondria using antibodies to HNEand mass spectrometry analysis to identify the targets and todeterminewhether there are links betweenHNE adduction andmitochondrial response to, or damage by, oxidative stress.
EXPERIMENTAL PROCEDURES
Arabidopsis Cell Culture and Mitochondria Isolation—Aheterotrophic Arabidopsis thaliana cell culture, establishedfrom callus of ecotype Lansberg erecta stem explants, wascultured in Murashige and Skoog basal salt mixture (Phyto-Technology Laboratories) supplemented with 3% (w/v)sucrose, 0.05% (w/v) naphthaleneacetic acid, and 0.005% (w/v)kinetin. Cell cultures (120 ml) were maintained in 100 �molm�2 s�1 light at 22 °C with shaking (120 rpm). At 7 days, cells(20 ml) were subcultured into 100 ml of fresh media. Cell cul-tures used for experimental analysis in this work were main-tained in the dark with shaking (120 rpm) following subculture.Mitochondria were purified from 7-day-old Arabidopsis cellcultures as described previously (26).
HNE Treatment of Isolated Mitochondria and OxidativeStress Treatments of Cells—Cells from 7-day-old dark-grownArabidopsis cell cultures were treated with 88 mM H2O2 (pre-pared in water by Asia Pacific Specialty Chemicals, Seven Hills,New South Wales, Australia), 25 �M antimycin A (prepared inethanol), or 400 �M menadione (prepared in ethanol). Isolatedmitochondria were incubated with indicated concentrations ofHNE for 20 min at 25 °C with shaking (800 rpm). HNE treat-ment of mitochondria subsequently analyzed by oxygen elec-trodewas carried out by incubation ofHNEwith 200–300�g ofmitochondrial protein in 250 �l of oxygen electrode reactionmedium in the oxygen electrode chamber at 25 °C.Measurement of Oxygen Consumption—All oxygen con-
sumption measurements were performed using a Clark-typeoxygen electrode (Hansatech, UK) attached to a personal com-puter. Calibration of the electrode was carried out by setting100% to the current recorded in air-saturated water at 22 or25 °C and by setting 0% to the current recorded after addition ofsodium dithionite to the water to remove all oxygen in the elec-trode chamber. Various substrate, inhibitor, and effector stand-ard additions are made as required. Arabidopsis cell culture(300 �l) was suspended in cell culture media to give a finalvolume of 1ml. Total respiration of intact cells wasmeasured at22 °C following the addition of the uncoupler carbonyl cyanidem-chlorophenylhydrazone (4 �M). Respiration through thealternative pathway or the cytochrome pathway in intact cellswas measured following the additions of 1 mM KCN or 1 mMn-PG, respectively. Isolated mitochondria (200–300 �g of pro-tein) were suspended in 1 ml of oxygen electrode reactionmedium (0.3 M sucrose, 5 mM KH2PO4, 10 mM TES, 10 mMNaCl, 2 mMMgSO4, 0.1% (w/v) bovine serum albumin, pH 7.2)at 25 °C. Total isolated mitochondrial respiration was meas-ured at 25 °C in the presence of various substrates as follows: forsuccinate-dependent respiration, succinate (10 mM) and ATP(0.5 mM); for malate plus glutamate-dependent respiration,malate (10 mM), glutamate (10 mM), thiamine pyrophosphate(0.2mM),NAD (2mM) and coenzymeA (12�M). Respiration viathe alternative pathway by isolated mitochondria was meas-ured in the presence of the complex III inhibitor myxothia-zol (2.5 �M), the reductant dithiothreitol (5 mM), and theAOX activator pyruvate (5 mM). Respiration via the cyto-chrome pathway was measured in the presence of the AOXinhibitor n-PG (0.5 mM).Protein Electrophoresis—SDS-PAGE was performed accord-
ing to Laemmli (27). Following electrophoresis gels were eitherstained with colloidal Coomassie or transferred to nitrocellu-lose membrane for Western blot analysis. IEF/SDS-PAGE wasperformed using immobilized pH gradient strips (ImmobilineDryStrips, GE Healthcare) of pH range 3–10NL (nonlinear; 18cm) or 4–7 (linear; 24 cm). Proteins were precipitated by addi-tion of acetone to samples to a final concentration of 90% ace-tone. Samples were placed at�20 °C overnight to allow precip-itation of proteins. Precipitates were centrifuged at 20,800 � gfor 20 min and solubilized in either 350 �l (3–10NL) or 450 �l(4–7 linear) of IEF sample buffer (6 M urea, 2 M thiourea, 2%(w/v) CHAPS, 2% (w/v) IPGbuffer pH3–10NLor pH4–7). 2�l(3–10NL), or 2.6�l (4–7 linear) of tributylphosphine (1:9 in IEFsample buffer) was added to each sample. Samples were then
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
DECEMBER 28, 2007 • VOLUME 282 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 37437
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from

used to rehydrate either pH 3–10NL DryStrips, or pH 4–7DryStrips overnight at room temperature. Isoelectric focusingwas performed using a flatbed electrophoresis unit (MultiphorII, GE Healthcare) at 20 °C. Following isoelectric focusing,strips were removed and incubated for 50 min in IEF/SDS-PAGE transfer buffer (4 M urea, 20% (v/v) glycerol, 2% (w/v)SDS, trace bromphenol blue, 0.375 M Tris-HCl, pH 8.8). Thesecond SDS-PAGE dimensions were run by placing the stripson top of 1-mm thick (25 � 18 cm) polyacrylamide gels (Ettan,Dalt 6, GEHealthcare) and following standard procedures. Bluenative-PAGE was performed based on the method by Schaggerand von Jagow (28). Mitochondrial samples (500 �g) were pel-leted at 18,300 � g for 10 min at 4 °C. The pellet was resus-pended in BN solubilization buffer with digitonin (150mM ace-tate, 10% glycerol, 5% (w/v) digitonin, 30mMHEPES, pH 7.4) at10�l per 100�g of protein to solubilized themembranes. Sam-ples were incubated on ice for 20 min and then centrifuged at18,300 � g for 10 min to remove unsolubilized material. Thesupernatant was then transferred to a new tube containing 5%Serva Blue G at 1 �l per 20 �l of solubilization buffer. The firstdimension separationwas performed on 1.5-mm thick (16� 11cm) polyacrylamide gels using the Protean II system (Bio-Rad).The gel consisted of a stacking layer (0.25 M �-aminocaproicacid, 4% (w/v) acrylamide:bisacrylamide (33:1), 0.05% (w/v)ammonium persulfate, 0.1% (v/v) TEMED, 25mMBisTris-HCl,pH 7.0), and a gradient separating layer (0.25 M �-aminocaproicacid, 4.5–16% (w/v) acrylamide:bisacrylamide (33:1), 0.05%(w/v) ammonium persulfate, 0.1% (v/v) TEMED, 25 mM Bis-Tris-HCl, pH 7.0). The gels were run with a cathode buffer (50mM Tricine, 0.02% (w/v) Coomassie 250G, 15 mM BisTris-HCl,pH 7.0) in the upper chamber and an anode buffer in the lowerchamber (50 mM BisTris-HCl, pH 7.0) of the gel tank. The gelwas run initially at a constant voltage of 100 V for 45 min andthereafter with a constant current of 15 mA per gel, withvoltage limited at 500 V, for 10 h. Gel runs containing sam-ples used for in-gel assay analysis were paused after 3 h ofelectrophoresis, and the upper cathode buffer wasexchanged for cathode buffer minus Coomassie to allow fora clearer background on the gel. Gels were stained with col-loidal Coomassie, prepared for in-gel enzymatic analysis, orprepared for second dimension SDS-PAGE.Enzyme Assays and In-gel Activity Staining—All assays were
performed with mitochondrial samples ruptured by freeze/thaw thus allowing direct access of substrates to both sides ofthe inner membrane. Succinate dehydrogenase (complex II ofthe respiratory chain) was measured using a protocol modifiedfrom Singer et al. (29). 200 �g of protein was added to 1 ml ofsuccinate dehydrogenase reaction mixture (60 �M 2,6-dichlo-rophenylindophenol, 0.03% (w/v) phenazine methosulfate, 1mM KCN, 1 mM n-PG, 50 mM KH2PO4, pH 7.6). The reactionwas started by the addition of 40 mM succinate to the mixture.Succinate dehydrogenase activity was measured as DCPIPreduction at 600 nm. Calculation of amount of DCPIP reducedwas performedusing � � 15.7mM�1 cm�1. In-gelmeasurementof complex I activity was performed based on the methoddescribed in Zerbetto et al. (30). Following blue native separa-tion ofmitochondrial protein, gels werewashed four times for 5min each with double distilled H2O and incubated in 50 ml per
five lanes of NADH DH assay mixture (0.14 mM NADH, 1mg/ml nitro blue tetrazolium, 0.1 M Tris-HCl, pH 7.4) until thestained complex was visible (10 min to 1 h). The reaction wasstopped by rinsing the gel in water and transferring the gel tofixing solution (40% methanol, 10% acetic acid). NADH anddeamino-NADH to FeCN [Fe(CN)63�] was measured spectro-photometrically as NADH oxidation at 340 nm using � � 6.22mM�1 cm�1 (31). Pyruvate dehydrogenase complex andmalatedehydrogenase were measured spectrophotometrically follow-ing NADH reduction and oxidation, respectively, at 340 nmusing � � 6.22 mM�1 cm�1 (24).Western Blotting and Immunodetection—Proteins were
transferred from polyacrylamide gels to HybondTM-C extranitrocellulose blotting membrane according to the method ofTowbin et al. (32) using a Hoefer SemiPhor (GE Healthcare)transfer system using standard procedures. FollowingWesternblotting, the membrane was incubated in 1% blocking solution(Roche Applied Science), either for 1 h at room temperature ona rocker or overnight at 4 °C without rocking, to block nonspe-cific binding of the antibody. Following a brief rinse in TBS/Tween, the membrane was incubated with polyclonal HNEadduct antibodies (HNE11-S, Alpha Diagnostics InternationalInc.) at 1:2000 dilution overnight at 4 °C, with gentle rocking.Themembrane was washed by three quick rinses, a 15-min andtwo 5-min washes with TBS/Tween, incubated with a 1:20,000dilution of an anti-rabbit horseradish peroxidase-conjugatedsecondary antibody (GE Healthcare) in 1% Western blockingreagent for 1 h at room temperature with gentle shaking. Themembrane was washed as before and detected using the BMchemiluminescence blotting substrate (POD) kit (RocheApplied Science). Images of the membranes were obtainedusing a LAS 1000 CCD camera and ImageGauge software (Fuji,Tokyo, Japan).Peptide Extraction from Gels—Gel spots of interest were
excised from colloidal Coomassie-stained diagonal PAGE ortwo-dimensional gels, placed into wells of 96-well PCR platesand frozen at�80 °C until analyzed. Gel spots were thawed anddestained twice with de-stain solution (50% (v/v) acetonitrile,10 mM NH4HCO3) for 45 min each time on an orbital shaker.Gel spotswere thendried at 50 °C for 1 h anddigested overnightwith digestion solution (12.5 �g/ml trypsin, 10 mMNH4HCO3,0.00125% (v/v) trifluoroacetic acid) at 37 °C. Peptides wereextracted by incubation with 12 �l of 100% acetonitrile for 15min on an orbital shaker. The supernatant from each samplewas placed into a cleanwell of a new 96-well PCRplate. Each gelspot was incubated further with 10 �l of a solution containing50% acetonitrile and 5% formic acid for 15 min with shaking.The supernatant was taken and placed with the previouslyextracted supernatant, and the previous 15-min incubationwasrepeated. Samples were dried down in a vacuum centrifuge andresuspended in 15 �l of resuspension solution (5% acetonitrile,0.1% formic acid) immediately prior to mass spectrometry.Samples were analyzed using either an Applied BiosystemsQSTAR Pulsar i LC/MS/MS system or an XCT Ultra (three-dimensional ion trap) from Agilent Technologies.Protein Identification by TandemMass Spectrometry—Sam-
ples from HNE-treated isolated mitochondria (Fig. 2 and sup-plemental Table S1) were analyzed using an Agilent 1100 series
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
37438 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 52 • DECEMBER 28, 2007
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from
capillary LC system and an Applied Biosystems QSTAR Pulsari LC/MS/MS system equipped with the IonSpray source run-ning Analyst QS software (version 1.1) with the instrument inpositive ionmode. Each extracted peptide sample was loaded inturn with the Agilent 1100 series capillary LC system onto a0.5� 50mmC18 (5�m, 100 Å) reverse phase column (HigginsAnalytical) with a C18OPTI-GUARDguard column (OptimizeTechnologies) at 16 �l/min equilibrated with 5% acetonitrileand 0.1% formic acid. Peptides were eluted from the C18reverse phase column into the QSTAR Pulsar i by a 7-min ace-tonitrile gradient (5–80%) at 16 �l/min under constant formicacid concentrations of 0.1%. During the period of ion detection,eluted peptides were analyzed by the mass spectrometer at 8�l/min. The method used to analyze eluted ions employed theinformation-dependent acquisition capabilities of Analyst QSand the rolling collision energy feature for automated collisionenergy determination based on the ions m/z (Sciex/AB). Themethod employed a 1-s time of flight-MS scan, which automat-ically switched (using information-dependent acquisition) to a2-s product ion scan (MS/MS) when a target ion reached anintensity of greater than 30 counts, and its charge state wasidentified as 2�, 3�, or 4�. Time of flight-MS scanning wasundertaken in them/z range of 400–1600m/z using aQ2 trans-missionwindow of 380 atomicmass units (100%). Samples ana-lyzed from oxidative stress-treated samples (Figs. 4 and 5; sup-plemental Tables S2 and S3) used an Agilent 1100 seriescapillary LC system and an Agilent Technologies XCT UltraIonTrap equipped with an ESI source equipped with a low flownebulizer in positivemode controlled by Chemstation (revisionB.01.03, build 204, Agilent Technologies) and MSD Trap Con-trol version 6.0 Build 38.15 software (Bruker Daltonik GmbH).Each extracted peptide sample was loaded in turn with the Agi-lent 1100 series capillary LC system onto a 0.5� 50-mmC18 (5�m, 100 Å; Varian) homemade reverse phase column at 10�l/min equilibrated with 5% acetonitrile and 0.1% formic acidunder a regulated temperature of 50 °C. Peptides were elutedfrom the C18 reverse phase column into the XCT Ultra by a9-min acetonitrile gradient (5–60%) under constant formicacid concentrations of 0.1%. The method used for initial iondetection utilized a mass range of 200–1400 m/z with scanmode set to Standard (8100m/zper s) and an ion charge controlconditions set at 250,000 and three averages taken per scan.Smartmode parameter settingswere employed using a target of800m/z, a compound stability factor of 90%, a trap drive level of80%, and optimize set to normal. Ions were selected forMS/MSafter reaching an intensity of 80,000 cps, and two precursor ionswere selected from the initial MS scan. MS/MS conditionsemployed SmartFrag for ion fragmentation, a scan range of70–2200 m/z using an average of three scans, the exclusion ofsingly charged ions option, and ion charge control conditionsset to 200,000 in ultra scan mode (26,000m/z per s).Protein Identification Searches with Mascot—Data produced
by both these methods were used to search the Mascot searchengine version 2.1.04 (Matrix Biosciences) for protein identifi-cation. Search parameters at Mascot employed a peptide toler-ance of �1.2 Da and a MS/MS tolerance of �0.6 Da, with theuse of the variable modifications, oxidized methionine (oxida-tionM) and HNE (CHK). Searches were performed against the
TAIR6 Arabidopsis data base (30,700 sequences; 12,656,682residues, released from Arabidopsis website on July 9, 2006).Details of the peptide matches and the coverage of matchedproteins byMascot searches are in supplemental Tables S4–S6,respectively.
RESULTS
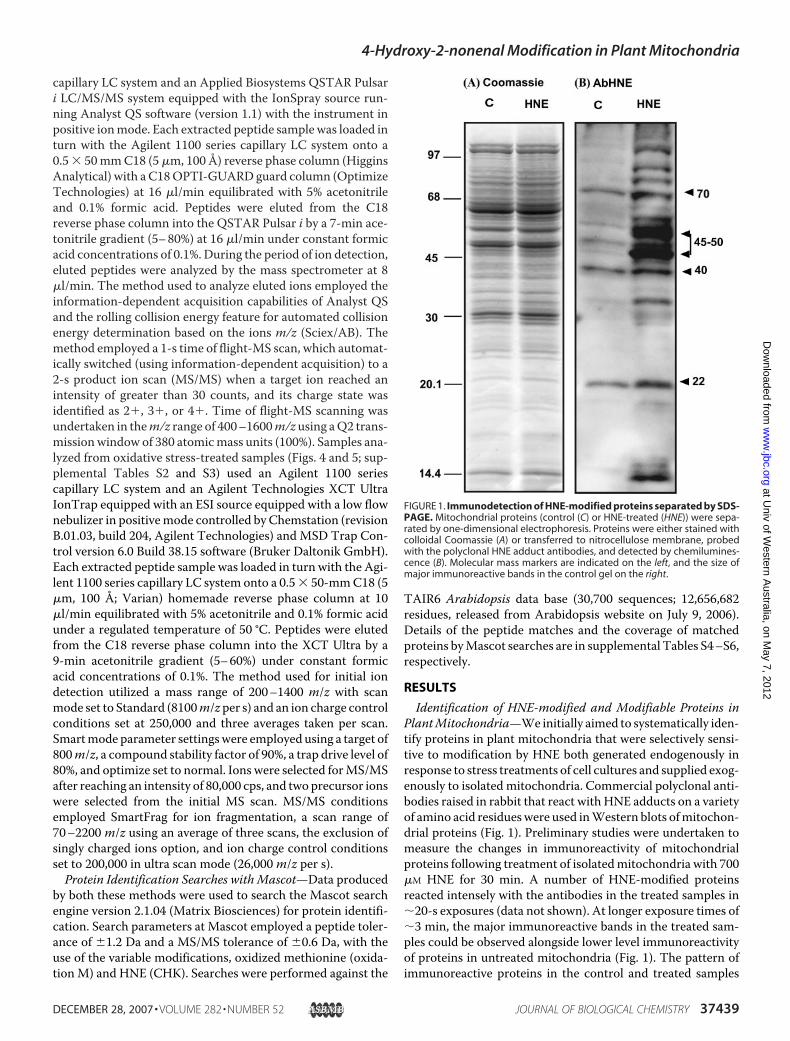
Identification of HNE-modified and Modifiable Proteins inPlantMitochondria—We initially aimed to systematically iden-tify proteins in plant mitochondria that were selectively sensi-tive to modification by HNE both generated endogenously inresponse to stress treatments of cell cultures and supplied exog-enously to isolated mitochondria. Commercial polyclonal anti-bodies raised in rabbit that react with HNE adducts on a varietyof amino acid residueswere used inWestern blots ofmitochon-drial proteins (Fig. 1). Preliminary studies were undertaken tomeasure the changes in immunoreactivity of mitochondrialproteins following treatment of isolatedmitochondria with 700�M HNE for 30 min. A number of HNE-modified proteinsreacted intensely with the antibodies in the treated samples in�20-s exposures (data not shown). At longer exposure times of�3 min, the major immunoreactive bands in the treated sam-ples could be observed alongside lower level immunoreactivityof proteins in untreated mitochondria (Fig. 1). The pattern ofimmunoreactive proteins in the control and treated samples
FIGURE 1. Immunodetection of HNE-modified proteins separated by SDS-PAGE. Mitochondrial proteins (control (C) or HNE-treated (HNE)) were sepa-rated by one-dimensional electrophoresis. Proteins were either stained withcolloidal Coomassie (A) or transferred to nitrocellulose membrane, probedwith the polyclonal HNE adduct antibodies, and detected by chemilumines-cence (B). Molecular mass markers are indicated on the left, and the size ofmajor immunoreactive bands in the control gel on the right.
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
DECEMBER 28, 2007 • VOLUME 282 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 37439
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from
contrasted with the total protein pattern observed in the colloi-dal Coomassie-stained control lanes (Fig. 1). Predominantbands at about 70, 45–50, 40, and 22 kDa reactedwith theHNE-protein adduct antibody in the control cells, whereas a muchgreater range of higher and lower molecular weight proteinsbecame immunoreactive following HNE treatment.As it was not feasible to identify the proteins reacting with
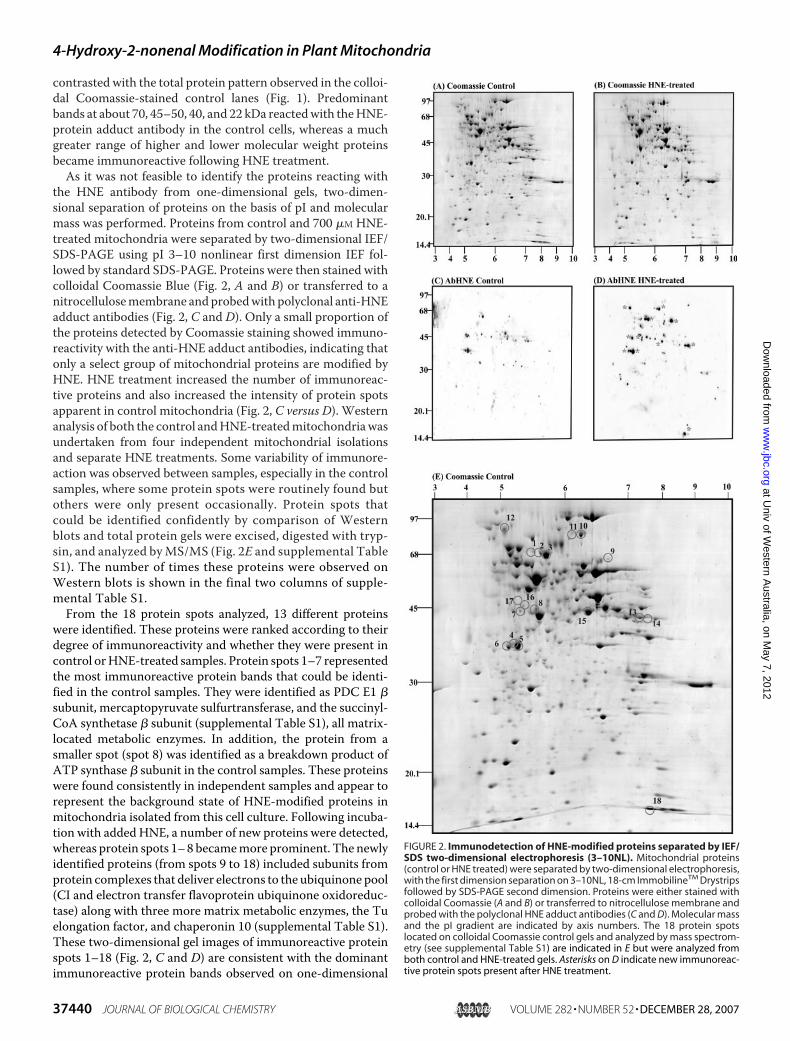
the HNE antibody from one-dimensional gels, two-dimen-sional separation of proteins on the basis of pI and molecularmass was performed. Proteins from control and 700 �M HNE-treated mitochondria were separated by two-dimensional IEF/SDS-PAGE using pI 3–10 nonlinear first dimension IEF fol-lowed by standard SDS-PAGE. Proteins were then stained withcolloidal Coomassie Blue (Fig. 2, A and B) or transferred to anitrocellulosemembrane andprobedwith polyclonal anti-HNEadduct antibodies (Fig. 2, C and D). Only a small proportion ofthe proteins detected by Coomassie staining showed immuno-reactivity with the anti-HNE adduct antibodies, indicating thatonly a select group of mitochondrial proteins are modified byHNE. HNE treatment increased the number of immunoreac-tive proteins and also increased the intensity of protein spotsapparent in control mitochondria (Fig. 2, C versus D). Westernanalysis of both the control andHNE-treatedmitochondriawasundertaken from four independent mitochondrial isolationsand separate HNE treatments. Some variability of immunore-action was observed between samples, especially in the controlsamples, where some protein spots were routinely found butothers were only present occasionally. Protein spots thatcould be identified confidently by comparison of Westernblots and total protein gels were excised, digested with tryp-sin, and analyzed byMS/MS (Fig. 2E and supplemental TableS1). The number of times these proteins were observed onWestern blots is shown in the final two columns of supple-mental Table S1.From the 18 protein spots analyzed, 13 different proteins
were identified. These proteins were ranked according to theirdegree of immunoreactivity and whether they were present incontrol orHNE-treated samples. Protein spots 1–7 representedthe most immunoreactive protein bands that could be identi-fied in the control samples. They were identified as PDC E1 �subunit, mercaptopyruvate sulfurtransferase, and the succinyl-CoA synthetase � subunit (supplemental Table S1), all matrix-located metabolic enzymes. In addition, the protein from asmaller spot (spot 8) was identified as a breakdown product ofATP synthase � subunit in the control samples. These proteinswere found consistently in independent samples and appear torepresent the background state of HNE-modified proteins inmitochondria isolated from this cell culture. Following incuba-tion with addedHNE, a number of new proteins were detected,whereas protein spots 1–8 becamemore prominent. The newlyidentified proteins (from spots 9 to 18) included subunits fromprotein complexes that deliver electrons to the ubiquinone pool(CI and electron transfer flavoprotein ubiquinone oxidoreduc-tase) along with three more matrix metabolic enzymes, the Tuelongation factor, and chaperonin 10 (supplemental Table S1).These two-dimensional gel images of immunoreactive proteinspots 1–18 (Fig. 2, C and D) are consistent with the dominantimmunoreactive protein bands observed on one-dimensional
FIGURE 2. Immunodetection of HNE-modified proteins separated by IEF/SDS two-dimensional electrophoresis (3–10NL). Mitochondrial proteins(control or HNE treated) were separated by two-dimensional electrophoresis,with the first dimension separation on 3–10NL, 18-cm ImmobilineTM Drystripsfollowed by SDS-PAGE second dimension. Proteins were either stained withcolloidal Coomassie (A and B) or transferred to nitrocellulose membrane andprobed with the polyclonal HNE adduct antibodies (C and D). Molecular massand the pI gradient are indicated by axis numbers. The 18 protein spotslocated on colloidal Coomassie control gels and analyzed by mass spectrom-etry (see supplemental Table S1) are indicated in E but were analyzed fromboth control and HNE-treated gels. Asterisks on D indicate new immunoreac-tive protein spots present after HNE treatment.
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
37440 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 52 • DECEMBER 28, 2007
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from
gels at apparent molecular masses of �70, 45–50, and 40 kDa(Fig. 1).Oxidative Stress Treatment of Intact Cells Leads to Changes
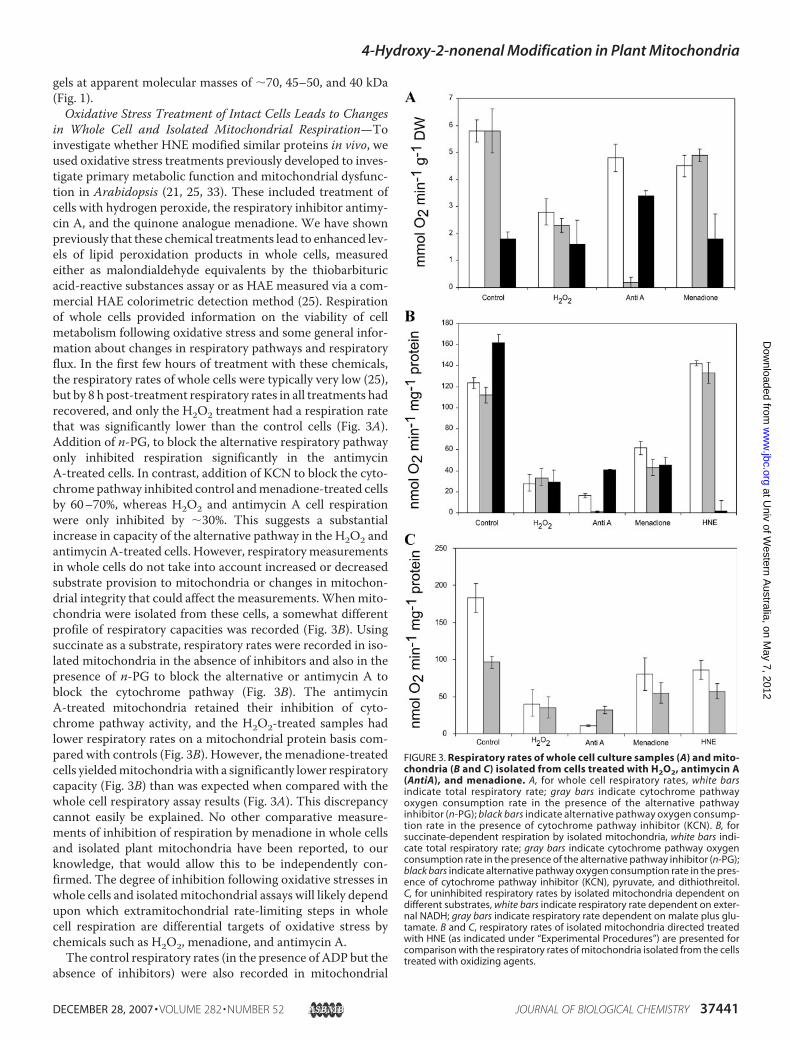
in Whole Cell and Isolated Mitochondrial Respiration—Toinvestigate whether HNE modified similar proteins in vivo, weused oxidative stress treatments previously developed to inves-tigate primary metabolic function and mitochondrial dysfunc-tion in Arabidopsis (21, 25, 33). These included treatment ofcells with hydrogen peroxide, the respiratory inhibitor antimy-cin A, and the quinone analogue menadione. We have shownpreviously that these chemical treatments lead to enhanced lev-els of lipid peroxidation products in whole cells, measuredeither as malondialdehyde equivalents by the thiobarbituricacid-reactive substances assay or as HAE measured via a com-mercial HAE colorimetric detection method (25). Respirationof whole cells provided information on the viability of cellmetabolism following oxidative stress and some general infor-mation about changes in respiratory pathways and respiratoryflux. In the first few hours of treatment with these chemicals,the respiratory rates of whole cells were typically very low (25),but by 8 h post-treatment respiratory rates in all treatments hadrecovered, and only the H2O2 treatment had a respiration ratethat was significantly lower than the control cells (Fig. 3A).Addition of n-PG, to block the alternative respiratory pathwayonly inhibited respiration significantly in the antimycinA-treated cells. In contrast, addition of KCN to block the cyto-chrome pathway inhibited control andmenadione-treated cellsby 60–70%, whereas H2O2 and antimycin A cell respirationwere only inhibited by �30%. This suggests a substantialincrease in capacity of the alternative pathway in the H2O2 andantimycin A-treated cells. However, respiratorymeasurementsin whole cells do not take into account increased or decreasedsubstrate provision to mitochondria or changes in mitochon-drial integrity that could affect the measurements.Whenmito-chondria were isolated from these cells, a somewhat differentprofile of respiratory capacities was recorded (Fig. 3B). Usingsuccinate as a substrate, respiratory rates were recorded in iso-lated mitochondria in the absence of inhibitors and also in thepresence of n-PG to block the alternative or antimycin A toblock the cytochrome pathway (Fig. 3B). The antimycinA-treated mitochondria retained their inhibition of cyto-chrome pathway activity, and the H2O2-treated samples hadlower respiratory rates on a mitochondrial protein basis com-pared with controls (Fig. 3B). However, the menadione-treatedcells yieldedmitochondriawith a significantly lower respiratorycapacity (Fig. 3B) than was expected when compared with thewhole cell respiratory assay results (Fig. 3A). This discrepancycannot easily be explained. No other comparative measure-ments of inhibition of respiration by menadione in whole cellsand isolated plant mitochondria have been reported, to ourknowledge, that would allow this to be independently con-firmed. The degree of inhibition following oxidative stresses inwhole cells and isolatedmitochondrial assays will likely dependupon which extramitochondrial rate-limiting steps in wholecell respiration are differential targets of oxidative stress bychemicals such as H2O2, menadione, and antimycin A.The control respiratory rates (in the presence of ADP but the
absence of inhibitors) were also recorded in mitochondrial
FIGURE 3. Respiratory rates of whole cell culture samples (A) and mito-chondria (B and C) isolated from cells treated with H2O2, antimycin A(AntiA), and menadione. A, for whole cell respiratory rates, white barsindicate total respiratory rate; gray bars indicate cytochrome pathwayoxygen consumption rate in the presence of the alternative pathwayinhibitor (n-PG); black bars indicate alternative pathway oxygen consump-tion rate in the presence of cytochrome pathway inhibitor (KCN). B, forsuccinate-dependent respiration by isolated mitochondria, white bars indi-cate total respiratory rate; gray bars indicate cytochrome pathway oxygenconsumption rate in the presence of the alternative pathway inhibitor (n-PG);black bars indicate alternative pathway oxygen consumption rate in the pres-ence of cytochrome pathway inhibitor (KCN), pyruvate, and dithiothreitol.C, for uninhibited respiratory rates by isolated mitochondria dependent ondifferent substrates, white bars indicate respiratory rate dependent on exter-nal NADH; gray bars indicate respiratory rate dependent on malate plus glu-tamate. B and C, respiratory rates of isolated mitochondria directed treatedwith HNE (as indicated under “Experimental Procedures”) are presented forcomparison with the respiratory rates of mitochondria isolated from the cellstreated with oxidizing agents.
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
DECEMBER 28, 2007 • VOLUME 282 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 37441
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from

samples using external NADHormalate plus glutamate as sub-strates (Fig. 3C). These data again showed lower respiratoryratesbyallmitochondriafromtreatedsamples.ExternalNADH-dependent respiration was the most significantly inhibited byboth direct HNE treatment and in vivo oxidative treatments,with all rates less than 35% of controls. Malate plus glutamate-dependent respiration was relatively less susceptible to inhibi-tion, with rates in treated samples being 40–60% of controlsand rates about 60% of controls after direct treatment withHNE.ASelective Set of HNE-modified ProteinsWere Identified in
Mitochondria from the Oxidative Stress-treated Cell Cul-tures—Using an approach similar to that adopted in Fig. 2,proteins from mitochondria isolated from chemical-treatedcell cultures at 8 h were separated by IEF/SDS-PAGE, followedby either staining for total protein or Western blotting andprobedwith the anti-HNEadduct antibodies. As themajority ofimmunoreactive protein spots from supplemental Table S1were in the pI 5–7 range, a narrower range 4–7 linear gradientIEF was used to maximize separation of immunoreactivespots. Antibody reaction in control samples was comparedon these gels with mitochondria from H2O2-, antimycin A-,and menadione-treated samples. We had previously usedthese antibodies to show that significant HNE modificationof mitochondrial proteins occurs under these conditions inone-dimensional gel comparisons (25), but we did not iden-tify the proteins concerned.In control samples several immunoreactive protein spots
were detected: protein spots 19, 21, 30, which were found onMS/MS analysis to be succinate dehydrogenase� subunit, ATPsynthase � subunit, and pyruvate dehydrogenase E1 � subunit(supplemental Table S2). All three were more prominent in thetreatment samples. This was broadly consistent with the iden-tifications made following direct HNE treatment (Fig. 2 andsupplemental Table S1). In addition, a range of new immuno-reactive protein spots was present in the samples from oxida-tively stressed cells (indicated by asterisks in Fig. 4). OnMS/MSanalysis, a number of these (protein spots 20 and 22–29) werefound to be breakdown products of two proteins, namely suc-cinate dehydrogenase� subunit andATP synthase� subunit. Afurther protein spot (no. 31) represented an additional presen-tation of pyruvate dehydrogenase E1 � subunit with a slightacidic shift from the main protein spot for this protein (30). Inaddition 10more proteinswere identified, including elongationfactor Tu, the 75-kDa subunit of complex I (also seen in supple-mental Table S1 after HNE treatment), and a selection of otherproteins over-represented in dehydrogenases and reductases(supplemental Table S2). Most of these new immunoreactiveproteinswere found in all three stress treatments butweremostpronounced and most commonly found in the menadionetreatments (supplemental Table S2).Because several of the identified proteins are membrane-
bound (supplemental Tables S1 and S2), blue native-PAGE sep-aration of the electron transport chain complexes coupled toSDS-PAGE separation of complex subunits was employed tofurther analyze HNEmodification ofmitochondrial membraneprotein samples (Fig. 5). Coomassie-stained gels of BN-PAGE/SDS-PAGE showed relatively similar abundance of oxidative
phosphorylation complexes in the treatments, with typicalbanding patterns for complex I, complex V, complex III, andcomplex IV present. Immunoreaction with the anti-HNE-pro-tein adduct antibodies in control samples identified proteinspots representing the � and � subunits of ATP synthase (sup-plemental Table S3, spots 42 and 43) and a protein spot con-taining the E3 dihydrolipoamide dehydrogenase (supplementalTable S3, spot 44). Following H2O2 treatment, several newintensely immunoreactive protein spots were apparent, notablyfour dehydrogenases (for glutamate, formate, glyeraldehyde-3-
FIGURE 4. Immunodetection of HNE-modified mitochondrial proteinsfrom H2O2-, antimycin A-, and menadione-treated cells, separated byIEF/SDS two-dimensional electrophoresis (4 –7NL). Mitochondrial pro-teins (control, H2O2-, antimycin A-, and menadione-treated) were separatedby two-dimensional electrophoresis, with the first dimension separation on4 –7NL, 18-cm ImmobilineTM Drystrips followed by SDS-PAGE second dimen-sion. Proteins were transferred to nitrocellulose membrane and probed withthe polyclonal HNE adduct antibodies (A–D). Molecular mass and the pI gra-dient are indicated by axis numbers. Asterisks on A–D indicate new immuno-reactive protein spots present in each treatment that are numbered on E. The23 protein spots located on colloidal Coomassie gels and analyzed by massspectrometry (see supplemental Table S2) are indicated on the stained con-trol gel in E but were analyzed from both control and treated samples.
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
37442 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 52 • DECEMBER 28, 2007
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from
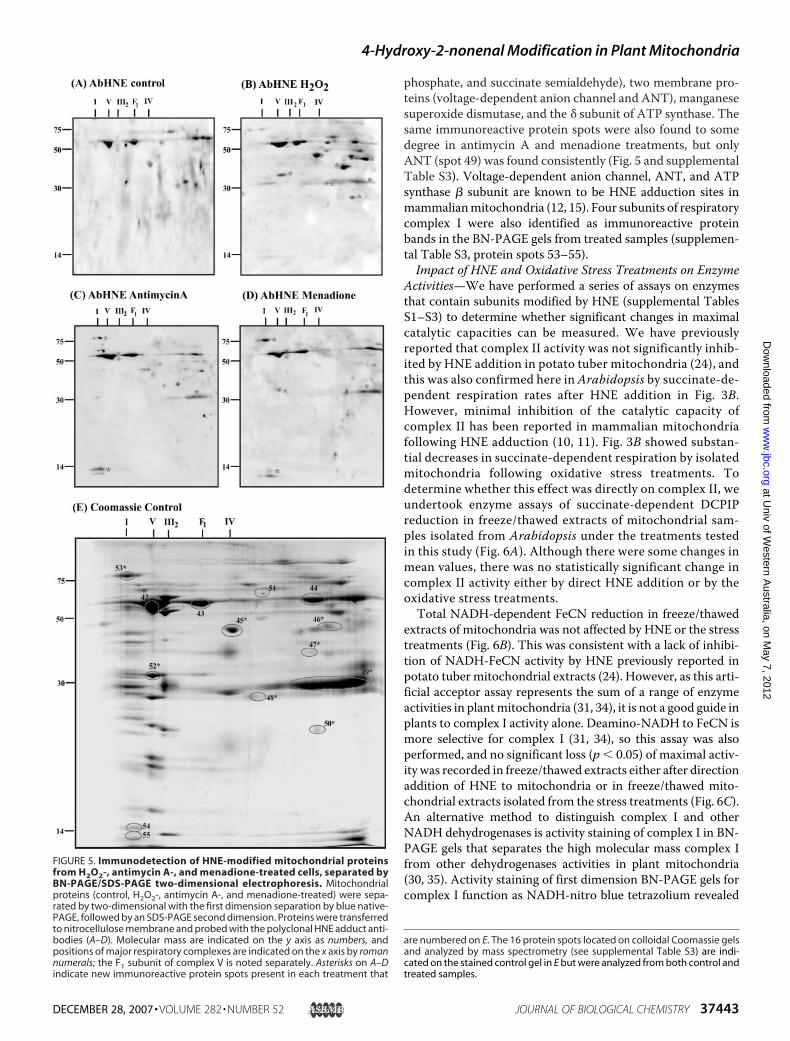
phosphate, and succinate semialdehyde), two membrane pro-teins (voltage-dependent anion channel and ANT), manganesesuperoxide dismutase, and the � subunit of ATP synthase. Thesame immunoreactive protein spots were also found to somedegree in antimycin A and menadione treatments, but onlyANT (spot 49) was found consistently (Fig. 5 and supplementalTable S3). Voltage-dependent anion channel, ANT, and ATPsynthase � subunit are known to be HNE adduction sites inmammalianmitochondria (12, 15). Four subunits of respiratorycomplex I were also identified as immunoreactive proteinbands in the BN-PAGE gels from treated samples (supplemen-tal Table S3, protein spots 53–55).Impact of HNE and Oxidative Stress Treatments on Enzyme
Activities—We have performed a series of assays on enzymesthat contain subunits modified by HNE (supplemental TablesS1–S3) to determine whether significant changes in maximalcatalytic capacities can be measured. We have previouslyreported that complex II activity was not significantly inhib-ited by HNE addition in potato tuber mitochondria (24), andthis was also confirmed here in Arabidopsis by succinate-de-pendent respiration rates after HNE addition in Fig. 3B.However, minimal inhibition of the catalytic capacity ofcomplex II has been reported in mammalian mitochondriafollowing HNE adduction (10, 11). Fig. 3B showed substan-tial decreases in succinate-dependent respiration by isolatedmitochondria following oxidative stress treatments. Todetermine whether this effect was directly on complex II, weundertook enzyme assays of succinate-dependent DCPIPreduction in freeze/thawed extracts of mitochondrial sam-ples isolated from Arabidopsis under the treatments testedin this study (Fig. 6A). Although there were some changes inmean values, there was no statistically significant change incomplex II activity either by direct HNE addition or by theoxidative stress treatments.Total NADH-dependent FeCN reduction in freeze/thawed
extracts of mitochondria was not affected by HNE or the stresstreatments (Fig. 6B). This was consistent with a lack of inhibi-tion of NADH-FeCN activity by HNE previously reported inpotato tuber mitochondrial extracts (24). However, as this arti-ficial acceptor assay represents the sum of a range of enzymeactivities in plantmitochondria (31, 34), it is not a good guide inplants to complex I activity alone. Deamino-NADH to FeCN ismore selective for complex I (31, 34), so this assay was alsoperformed, and no significant loss (p � 0.05) of maximal activ-ity was recorded in freeze/thawed extracts either after directionaddition of HNE to mitochondria or in freeze/thawed mito-chondrial extracts isolated from the stress treatments (Fig. 6C).An alternative method to distinguish complex I and otherNADH dehydrogenases is activity staining of complex I in BN-PAGE gels that separates the high molecular mass complex Ifrom other dehydrogenases activities in plant mitochondria(30, 35). Activity staining of first dimension BN-PAGE gels forcomplex I function as NADH-nitro blue tetrazolium revealed
FIGURE 5. Immunodetection of HNE-modified mitochondrial proteinsfrom H2O2-, antimycin A-, and menadione-treated cells, separated byBN-PAGE/SDS-PAGE two-dimensional electrophoresis. Mitochondrialproteins (control, H2O2-, antimycin A-, and menadione-treated) were sepa-rated by two-dimensional with the first dimension separation by blue native-PAGE, followed by an SDS-PAGE second dimension. Proteins were transferredto nitrocellulose membrane and probed with the polyclonal HNE adduct anti-bodies (A–D). Molecular mass are indicated on the y axis as numbers, andpositions of major respiratory complexes are indicated on the x axis by romannumerals; the F1 subunit of complex V is noted separately. Asterisks on A–Dindicate new immunoreactive protein spots present in each treatment that
are numbered on E. The 16 protein spots located on colloidal Coomassie gelsand analyzed by mass spectrometry (see supplemental Table S3) are indi-cated on the stained control gel in E but were analyzed from both control andtreated samples.
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
DECEMBER 28, 2007 • VOLUME 282 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 37443
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from
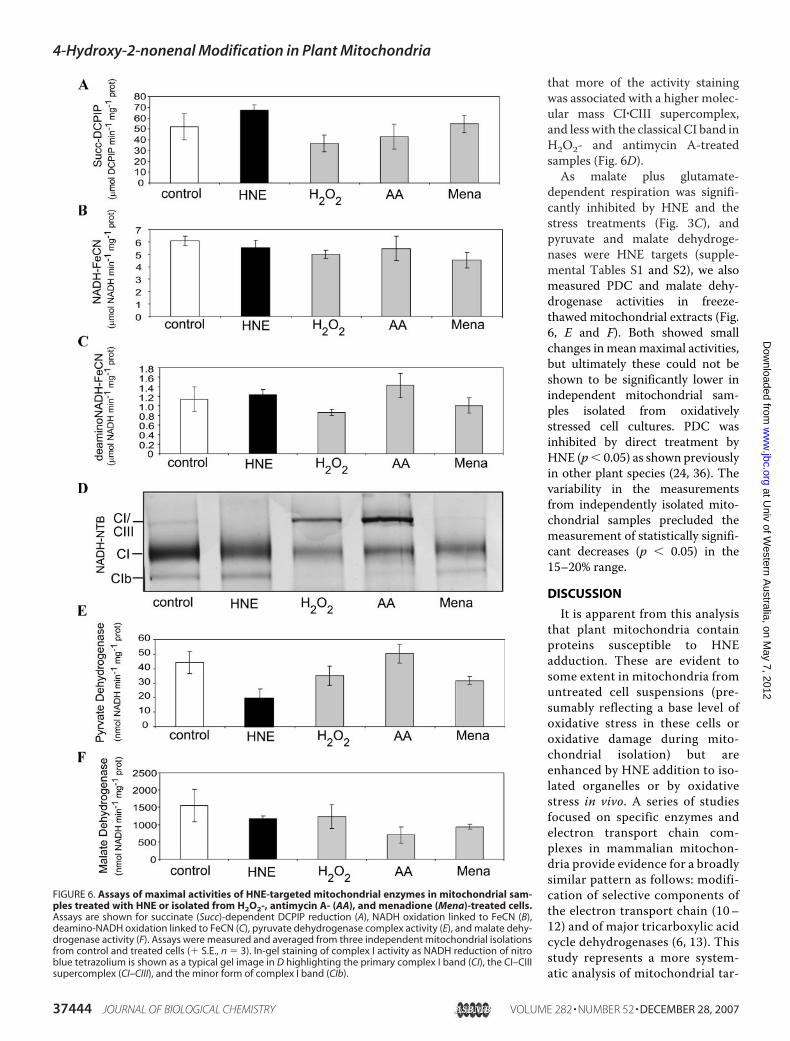
that more of the activity stainingwas associated with a higher molec-ular mass CI�CIII supercomplex,and less with the classical CI band inH2O2- and antimycin A-treatedsamples (Fig. 6D).As malate plus glutamate-
dependent respiration was signifi-cantly inhibited by HNE and thestress treatments (Fig. 3C), andpyruvate and malate dehydroge-nases were HNE targets (supple-mental Tables S1 and S2), we alsomeasured PDC and malate dehy-drogenase activities in freeze-thawed mitochondrial extracts (Fig.6, E and F). Both showed smallchanges in meanmaximal activities,but ultimately these could not beshown to be significantly lower inindependent mitochondrial sam-ples isolated from oxidativelystressed cell cultures. PDC wasinhibited by direct treatment byHNE (p� 0.05) as shown previouslyin other plant species (24, 36). Thevariability in the measurementsfrom independently isolated mito-chondrial samples precluded themeasurement of statistically signifi-cant decreases (p � 0.05) in the15–20% range.
DISCUSSION
It is apparent from this analysisthat plant mitochondria containproteins susceptible to HNEadduction. These are evident tosome extent in mitochondria fromuntreated cell suspensions (pre-sumably reflecting a base level ofoxidative stress in these cells oroxidative damage during mito-chondrial isolation) but areenhanced by HNE addition to iso-lated organelles or by oxidativestress in vivo. A series of studiesfocused on specific enzymes andelectron transport chain com-plexes in mammalian mitochon-dria provide evidence for a broadlysimilar pattern as follows: modifi-cation of selective components ofthe electron transport chain (10–12) and of major tricarboxylic acidcycle dehydrogenases (6, 13). Thisstudy represents a more system-atic analysis of mitochondrial tar-
FIGURE 6. Assays of maximal activities of HNE-targeted mitochondrial enzymes in mitochondrial sam-ples treated with HNE or isolated from H2O2-, antimycin A- (AA), and menadione (Mena)-treated cells.Assays are shown for succinate (Succ)-dependent DCPIP reduction (A), NADH oxidation linked to FeCN (B),deamino-NADH oxidation linked to FeCN (C), pyruvate dehydrogenase complex activity (E), and malate dehy-drogenase activity (F). Assays were measured and averaged from three independent mitochondrial isolationsfrom control and treated cells (� S.E., n � 3). In-gel staining of complex I activity as NADH reduction of nitroblue tetrazolium is shown as a typical gel image in D highlighting the primary complex I band (CI), the CI–CIIIsupercomplex (CI–CIII), and the minor form of complex I band (CIb).
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
37444 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 52 • DECEMBER 28, 2007
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from

gets in a single system. It also shows that protein modifica-tion by HNE can be relatively selective in the plantmitochondrial proteome and identifies the protein targets.It is quite notable from our results that a significant propor-
tion of the HNE adduction to mitochondrial membrane com-ponents focuses around the enzymes that deliver electrons tothe ubiquinone pool (Figs. 2, 4, and 5 and supplemental TablesS1–S3). Several complex I subunits were identified as HNE tar-gets, along with the FAD containing the � subunit of succinatedehydrogenase and the electron transfer flavoprotein ubiqui-none oxidoreductase recently identified in Arabidopsis for itsrole in delivering electrons to UQ from the products of thebranched chain amino acid degradation pathway (37).We havepreviously shown that AOX, which accepts electrons fromUQH2, is another target of HNE and is inactivated by the alde-hyde (25).It seems logical that these UQ-interacting enzymes might be
sites of HNE modification by oxidative stress in vivo as super-oxide production by the electron transport chain generallyarises from semiquinones in the membrane (38, 39). Thus,localized endogenous HNE production in the membrane andmodification of nearby UQ-interacting proteins are feasible.Although this is a reasonable working hypothesis for oxidativestresses generated within cells (Figs. 4 and 5 and supplementalTables S2 and S3), it does not readily explain such localizeddamage whenHNE is exogenously supplied tomitochondria invitro (Fig. 2 and supplemental Table S1). Rather, this suggests alink between the UQ pool, or at least proteins with a redoxpotential that allows them to interact with UQ, and susceptibil-ity to modification by HNE. Extensive studies of oxidativestress-induced changes in plant gene expression also show thatinduction of transcripts for the alternative external and internalNADH:UQ oxidoreductases and the UQH2:O2 alternative oxi-dase are the major transcription changes among all the genesfor mitochondrial proteins (40, 41).Thus it appears that a range of events are occurring during
oxidative stress focused on these nonphosphorylating bypassenzymes. First, their synthesis is being induced; second, theyare being damaged; and third, their catalytic action might helplower the ROS production via the mitochondrial electrontransport chain. The latter point is clear for AOX as it can helpavoid over-reduction of the UQ pool (19). The NADH:UQ oxi-doreductases could lower ROS production through avoidingreverse electron flow from succinate dehydrogenase to com-plex I when the UQ and/or matrix nucleotide redox poise isaltered.We currently do not have any data to suggest that theseprocesses are linked by feed-back or feed-forwardmechanisms.However, we can propose an hypothesis that links the observedevents. If modification by HNE leads to loss of activity,increased oxidative side reactions, or increased turnover of pro-teins in oxidative phosphorylation complexes, then coordi-nated changes in gene expression for nonphosphorylatingbypasses can be interpreted as a convenientway of increasing ormaintaining respiratory flux through the UQ pool by provisionof a short, readily assembled electron transport chain. In thiscontext, the inhibition of AOX activity by HNE may appearcontradictory, but this could trigger a signal to stimulate geneexpression resulting in increased compensatory AOX synthesis
aswell asNAD(P)Hdehydrogenase synthesis and the turn on ofother stress-related genes.HNE, which is likely to be present or produced in the mem-
brane, also appears to modify soluble enzymes of the matrixthat are likely to be in close proximity to the surface of themitochondrial inner membrane. Following the work of otherson HNE modification of lipoic acid cofactors in PDC andKGDC (6, 13), we have previously shown that the loss of lipoicacid inactivates these enzymes in potato, pea, and Arabidopsismitochondria (24, 36). Here we show that by directly monitor-ing HNE adduction of soluble proteins, two differences in per-spective are gained on HNE targets in the mitochondrialmatrix. First, even within the known susceptible protein com-plexes of PDC and KGDC, it is not only the E2 dihydrolipoam-ide acyltransferases, which contain bound lipoic acid, that aretargets. PDC E1 � subunit and the shared E3 lipoamide dehy-drogenase subunit are clearly major HNE adduction targetswithin these same protein complexes. Defining the precisemolecular mechanism of PDC and KGDC damage during oxi-dative stress will thus require a thorough analysis of each step inthe catalysis of these enzyme complexes and not simply a focuson lipoic acidmodification. Second, the susceptibility of matrixdehydrogenases extends beyond these classical lipoic acid-con-taining proteins to other major dehydrogenase classes (supple-mental Tables S1–S3). Glyceraldehyde-3-phosphate dehydro-genase (supplemental Table S3) is not a matrix enzyme but aglycolytic enzyme known to be associated with the outer mito-chondrial membrane (42) and to accumulate on mitochondriaduring oxidative stress (21). It has recently been highlighted asan oxidative stress-sensitive protein in plants (43), and its HNEadduction could be involved in this phenomenon.Interestingly, in a number of cases, proteins that are found to
be HNE targets are also identified by the HNE adduct antibod-ies as smaller stable degradation products (supplementalTables S1 and S2). We have found some of these products pre-viously in a quantitative analysis of changes in abundance ofproteins in mitochondria during the same stresses in cell cul-ture (21). This could be interpreted in a number of ways: first, alink may exist between HNE adduction and protein degrada-tion; second, HNE adduction may be blocking degradation,leading to accumulation of intermediates that would otherwisebe rapidly degraded. A case forHNE adduction limiting proteindegradation has been made in mammals (44, 45), but a case forspecific proteolysis of oxidatively damaged proteins in mito-chondria has also been made (46).It is not clear to what degree HNE adduction leads directly to
protein dysfunction. Although we have been able to observeinhibition of AOX activity by HNE (25), we have not observedsignificant inhibition of CI or CII by HNE adduction or oxida-tive stress (Fig. 6), but we do see significant losses in wholechain electron transport activity (Fig. 3B). It is also not possibleto determine the degree of HNE adduction of a protein fromthese data using anti-HNE adduct antibodies. If less than 20%ofa protein is adducted under these conditions, it is unlikely evenif the activity assay undertaken could measure the functionaleffect (for example on Vmax, substrate, cofactor, or product-binding constants) that a statistically significant change wouldbe recorded given technical and biological variability in activity
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
DECEMBER 28, 2007 • VOLUME 282 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 37445
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from

measurements. However, in combination with other oxidationdamage, inefficiency in a number of steps likely underlies theclearly measured losses in total respiratory rates observed bothin vivo and in isolatedmitochondria with a variety of substrates(Fig. 3). The degree of adduction could best be measured bydefining the peptide that is modified and assessing the abun-dance of the modified and unmodified peptides by mass spec-trometry. However, we have failed to identify any HNE-modi-fied peptides in our mass spectral analysis, despite extensivesearches of spectra for parent peptides �138 or �156 Da andsearches in MS/MS for the 139.1 dehydrated, protonated HNEmarker ion (data not shown). To our knowledge, there are veryfew cases where the sites of HNE adduction of mitochondrialproteins by in vivo modifications have been proven by massspectrometry. Almost all reports of HNE modification sites inmitochondria are from HNE addition to purified proteins orpeptides followed by immediate analysis by MS or MS/MS (4,47–50). This inability to identify HNE peptides might be due tothe reactivity of the aldehyde group of HNE, which has thepotential to cross-link peptides or cyclize the peptide duringsample handling prior to analysis (51).Taking a different approach to this issue, we have compared
the protein list generated in this report of HNE adducted pro-teins together with those generated in other reports in plantmitochondria looking at carbonyl group formation (52), tryp-tophan oxidation (53), and accessible thiols for thioredoxinbinding (54) with proteins known to be degraded during oxida-tive stress in plant mitochondria (21, 23). Remarkably, of the 31different proteins in this report, 24 have been listed in one ofthese categories (supplemental Table S7). It is even possiblethat the carbonyl group protein lists generated by Kristensen etal. (52)may be, at least in part, a direct analysis of HNE adducts.HNE adduction to amino acids by Michael addition from theC-3 unsaturation is the most common method of HNE adduc-tion (3), butwhen this occurs the adductedHNE retains theC-1aldehyde group, thus effectively introducing a carbonyl groupinto the protein. These HNE-Michael adducts can react with(2,4-dinitrophenyl)hydrazine via the carbonyl group on theHNE adduct (50, 51, 55).Considering supplemental Table S7 as a whole, it appears
that a common element in selective oxidative damage and deg-radation is the surface availability of thiols, which allows oxida-tive modification and generation of carbonyl groups, followedby protein degradation. The exact path of this process is notknown, and the role of HNE modification as an inhibitor ofenzyme activity, as a tag for degradation, and/or an inhibitor ofprotein turnover is likely to be complex and could be protein-specific. However, it is clear that HNEmodification is an activeprocess in plant mitochondria under oxidative stress and thattools to detect and localize this process could be used morebroadly in plants to gain insights into the mechanism of oxida-tive stress and the link between lipid peroxidation and proteindamage in plants.REFERENCES1. Schneider, C., Tallman, K. A., Porter, N. A., and Brash, A. R. (2001) J. Biol.
Chem. 276, 20831–208382. Moller, I. M., Jensen, P. E., and Hansson, A. (2007) Annu. Rev. Plant Biol.
58, 459–481
3. Esterbauer, H., Schaur, R. J., and Zollner, H. (1991) Free Radic. Biol. Med.11, 81–128
4. Isom,A. L., Barnes, S.,Wilson, L., Kirk,M., Coward, L., andDarley-Usmar,V. (2004) J. Am. Soc. Mass Spectrom. 15, 1136–1147
5. Sayre, L. M., Arora, P. K., Iyer, R. S., and Salomon, R. G. (1993) Chem. Res.Toxicol. 6, 19–22
6. Humphries, K.M., and Szweda, L. I. (1998)Biochemistry 37, 15835–158417. Chen, J., Henderson, G. I., and Freeman, G. L. (2001) J. Mol. Cell. Cardiol.
33, 1919–19278. Chen, J., Schenker, S., Frosto, T. A., and Henderson, G. I. (1998) Biochim.
Biophys. Acta 1380, 336–3449. Musatov, A., Carroll, C. A., Liu, Y. C., Henderson, G. I., Weintraub, S. T.,
and Robinson, N. C. (2002) Biochemistry 41, 8212–822010. Picklo, M. J., Amarnath, V., McIntyre, J. O., Graham, D. G., and Montine,
T. J. (1999) J. Neurochem. 72, 1617–162411. Lashin, O. M., Szweda, P. A., Szweda, L. I., and Romani, A. M. P. (2006)
Free Radic. Biol. Med. 40, 886–89612. Choksi, K. B., Boylston, W. H., Rabek, J. P., Widger, W. R., and Papacon-
stantinou, J. (2004) Biochim. Biophys. Acta 1688, 95–10113. Humphries, K. M., Yoo, Y., and Szweda, L. I. (1998) Biochemistry 37,
552–55714. Moreau, R., Heath, S. H., Doneanu, C. E., Lindsay, J. G., and Hagen, T. M.
(2003) Antioxid. Redox. Signal. 5, 517–52715. Vieira,H. L., Belzacq,A. S., Haouzi, D., Bernassola, F., Cohen, I., Jacotot, E.,
Ferri, K. F., El Hamel, C., Bartle, L. M., Melino, G., Brenner, C., Goldma-cher, V., and Kroemer, G. (2001) Oncogene 20, 4305–4316
16. Perluigi,M., Fai Poon,H.,Hensley, K., Pierce,W.M., Klein, J. B., Calabrese,V., De Marco, C., and Butterfield, D. A. (2005) Free Radic. Biol. Med. 38,960–968
17. Carbone, D. L., Doorn, J. A., Kiebler, Z., and Petersen, D. R. (2005) Chem.Res. Toxicol. 18, 1324–1331
18. Smith, A.M., Ratcliffe, R. G., and Sweetlove, L. J. (2004) J. Biol. Chem. 279,51944–51952
19. Vanlerberghe, G. C., and McIntosh, L. (1997) Annu. Rev. Plant Physiol.Plant Mol. Biol. 48, 703–734
20. Chew, O., Whelan, J., and Millar, A. H. (2003) J. Biol. Chem. 278,46869–46877
21. Sweetlove, L. J., Heazlewood, J. L., Herald, V., Holtzapffel, R., Day, D. A.,Leaver, C. J., and Millar, A. H. (2002) Plant J. 32, 891–904
22. Finkemeier, I., Goodman, M., Lamkemeyer, P., Kandlbinder, A., Sweet-love, L. J., and Dietz, K.-J. (2005) J. Biol. Chem. 280, 12168–12180
23. Taylor, N. L., Heazlewood, J. L., Day, D. A., and Millar, A. H. (2005)Mol.Cell. Proteomics 4, 1122–1133
24. Millar, A. H., and Leaver, C. J. (2000) FEBS Lett. 481, 117–12125. Winger, A. M., Millar, A. H., and Day, D. A. (2005) Biochem. J. 387,
865–87026. Millar, A. H., Liddell, A., and Leaver, C. J. (2001) Methods Cell Biol. 65,
53–7427. Laemmli, U. K. (1970) Nature 227, 680–68528. Schagger, H., and von Jagow, G. (1991) Anal. Biochem. 199, 223–23129. Singer, T. P., Oestreicher, G., and Hogue, P. (1973) Plant Physiol. 52,
616–62130. Zerbetto, E., Vergani, L., and Dabbeni-Sala, F. (1997) Electrophoresis 18,
2059–206431. Menz, R. I., Griffith, M., Day, D. A., and Wiskich, J. T. (1992) Eur. J. Bio-
chem. 208, 481–48532. Towbin, H., Staehelin, T., and Gordon, J. (1979) Proc. Natl. Acad. Sci.
U. S. A. 76, 4350–435433. Baxter, C. J., Redestig, H., Schauer, N., Repsilber, D., Patil, K. R., Nielsen, J.,
Selbig, J., Liu, J., Fernie, A. R., and Sweetlove, L. J. (2007)Plant Physiol. 143,312–325
34. Rasmusson, A. G., and Moller, I. M. (1991) Physiol. Plant. 83, 357–36535. Sabar, M., Balk, J., and Leaver, C. J. (2005) Plant J. 44, 893–90136. Taylor, N. L., Day, D. A., and Millar, A. H. (2002) J. Biol. Chem. 277,
42663–4266837. Ishizaki, K., Larson, T. R., Schauer, N., Fernie, A. R., Graham, I. A., and
Leaver, C. J. (2005) Plant Cell 17, 2587–260038. Raha, S., and Robinson, B. H. (2000) Trends Biochem. Sci. 25, 502–508
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
37446 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 52 • DECEMBER 28, 2007
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from

39. Moller, I.M. (2001)Annu. Rev. Plant Physiol. PlantMol. Biol. 52, 561–59140. Clifton, R., Lister, R., Parker, K., Sappl, P., Elhafez, D., Millar, A., Day, D.,
and Whelan, J. (2005) Plant Mol. Biol. 58, 193–21241. Clifton, R., Millar, A. H., and Whelan, J. (2006) Biochim. Biophys. Acta
1757, 730–74142. Giege, P., Heazlewood, J. L., Roessner-Tunali, U., Millar, A. H., Fernie,
A. R., Leaver, C. J., and Sweetlove, L. J. (2003) Plant Cell 15, 2140–215143. Hancock, J. T., Henson, D., Nyirenda, M., Desikan, R., Harrison, J.,
Lewis, M., Hughes, J., and Neill, S. J. (2005) Plant Physiol. Biochem. 43,828–835
44. Friguet, B., and Szweda, L. I. (1997) FEBS Lett. 405, 21–2545. Friguet, B., Szweda, L. I., and Stadtman, E. R. (1994) Arch. Biochem. Bio-
phys. 311, 168–17346. Marcillat, O., Zhang, Y., Lin, S. W., and Davies, K. J. (1988) Biochem. J.
254, 677–68347. Aldini, G., Dalle-Donne, I., Vistoli, G., Maffei Facino, R., and Carini, M.
(2005) J. Mass Spectrom. 40, 946–954
48. Carbone, D. L., Doorn, J. A., Kiebler, Z., Ickes, B. R., and Petersen, D. R.(2005) J. Pharmacol. Exp. Ther. 315, 8–15
49. Aldini, G., Gamberoni, L., Orioli, M., Beretta, G., Regazzoni, L., Facino,R. M., and Carini, M. (2006) J. Mass Spectrom. 41, 1149–1161
50. Carini, M., Aldini, G., and Facino, R. M. (2004) Mass Spectrom. Rev. 23,281–305
51. Fenaille, F., Guy, P. A., and Tabet, J. C. (2003) J. Am. Soc. Mass Spectrom.14, 215–226
52. Kristensen, B. K., Askerlund, P., Bykova, N. V., Egsgaard, H., and Moller,I. M. (2004) Phytochemistry 65, 1839–1851
53. Moller, I. M., and Kristensen, B. K. (2006) Free Radic. Biol. Med. 40,430–435
54. Balmer, Y., Vensel, W. H., Tanaka, C. K., Hurkman, W. J., Gelhaye, E.,Rouhier, N., Jacquot, J. P., Manieri, W., Schurmann, P., Droux, M., andBuchanan, B. B. (2004) Proc. Natl. Acad. Sci. U. S. A. 101, 2642–2647
55. Deighton, N., Magill, W. J., Bremner, D. H., and Benson, E. E. (1997) FreeRadic. Res. 27, 255–265
4-Hydroxy-2-nonenal Modification in Plant Mitochondria
DECEMBER 28, 2007 • VOLUME 282 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 37447
at Univ of W
estern Australia, on M
ay 7, 2012w
ww
.jbc.orgD
ownloaded from


![Drug Delivery System Based on Covalently Bonded Poly[N-Isopropylacrylamide-co-2-Hydroxyethylacrylate]-Based Nanoparticle Networks](https://static.fdokumen.com/doc/165x107/6340d5f6e0dac3b265042228/drug-delivery-system-based-on-covalently-bonded-polyn-isopropylacrylamide-co-2-hydroxyethylacrylate-based.jpg)

















