
Cardiac-Specific LIM Protein FHL2 Modifies the Hypertrophic Response to b-Adrenergic Stimulation
9
Bassel-Duby and R. Sanders Williams Yanfeng Kong, John M. Shelton, Beverly Rothermel, Xiangqing Li, James A. Richardson, Rhonda Stimulation -Adrenergic β Cardiac-Specific LIM Protein FHL2 Modifies the Hypertrophic Response to Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2001 American Heart Association, Inc. All rights reserved. is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Circulation doi: 10.1161/01.CIR.103.22.2731 2001;103:2731-2738 Circulation. http://circ.ahajournals.org/content/103/22/2731 Wide Web at: The online version of this article, along with updated information and services, is located on the World http://circ.ahajournals.org//subscriptions/ is online at: Circulation Information about subscribing to Subscriptions: http://www.lww.com/reprints Information about reprints can be found online at: Reprints: document. and Rights Question and Answer Permissions the middle column of the Web page under Services. Further information about this process is available in the the online version of the published article for which permission is being requested is located, click Request Permissions in can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once Circulation Requests for permissions to reproduce figures, tables, or portions of articles originally published in Permissions: by guest on July 22, 2014 http://circ.ahajournals.org/ Downloaded from by guest on July 22, 2014 http://circ.ahajournals.org/ Downloaded from
-
Upload
utsouthwestern -
Category
Documents
-
view
5 -
download
0
Transcript of Cardiac-Specific LIM Protein FHL2 Modifies the Hypertrophic Response to b-Adrenergic Stimulation

Bassel-Duby and R. Sanders WilliamsYanfeng Kong, John M. Shelton, Beverly Rothermel, Xiangqing Li, James A. Richardson, Rhonda
Stimulation-AdrenergicβCardiac-Specific LIM Protein FHL2 Modifies the Hypertrophic Response to
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2001 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/01.CIR.103.22.2731
2001;103:2731-2738Circulation.
http://circ.ahajournals.org/content/103/22/2731Wide Web at:
The online version of this article, along with updated information and services, is located on the World
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. and Rights Question and Answer
Permissionsthe middle column of the Web page under Services. Further information about this process is available in thethe online version of the published article for which permission is being requested is located, click Request Permissions in
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. OnceCirculation Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from

Cardiac-Specific LIM Protein FHL2 Modifies theHypertrophic Response tob-Adrenergic StimulationYanfeng Kong, PhD; John M. Shelton, BS; Beverly Rothermel, PhD; Xiangqing Li, PhD;
James A. Richardson, PhD; Rhonda Bassel-Duby, PhD; R. Sanders Williams, MD
Background—A deficiency of muscle LIM protein results in dilated cardiomyopathy, but the function of other LIMproteins in the heart has not been assessed previously. We have characterized the expression and function of FHL2, aheart-specific member of the LIM domain gene family.
Methods and Results—Expression of FHL2 mRNA and protein was examined by Northern blot, in situ hybridization, andWestern blot analyses of fetal and adult mice. FHL2 transcripts are present at embryonic day (E) 7.5 within the cardiaccrescent in a pattern that resembles that of Nkx2.5 mRNA. During later stages of cardiac development and in adultanimals, FHL2 expression is localized to the myocardium and absent from endocardium, cardiac cushion, outflow tract,or coronary vasculature. The gene encoding FHL2 was disrupted by homologous recombination, and knockout micedevoid of FHL2 were found to undergo normal cardiovascular development. In the absence of FHL2, however, cardiachypertrophy resulting from chronic infusion of isoproterenol is exaggerated (59% versus 20% increase in heartweight/body weight in FHL null versus wild-type mice;P,0.01).
Conclusions—FHL2 is an early marker of cardiogenic cells and a cardiac-specific LIM protein in the adult. FHL2 is notrequired for normal cardiac development but modifies the hypertrophic response tob-adrenergic stimulation.(Circulation. 2001;103:2731-2738.)
Key Words: hypertrophyn geneticsn molecular biologyn myocardium
The LIM protein superfamily comprises transcription fac-tors, protein kinases, and cytoskeletal proteins that are
involved in a wide range of cellular functions, including celllineage determination, differentiation, focal adhesion, andsignal transduction.1,2 The signature LIM motif is defined bythe presence of 1 or multiple consensus sequence motifs(CX2CX17–19HX2C2X2CX16–20CX2C), and LIM proteins can besubdivided into several subclasses based on the number ofLIM motifs and the presence or absence of additional motifsin the protein. LIM-homeodomain proteins include both LIMmotifs and a homeobox region, and several are known to beessential for development of skeletal, endocrine, or neuronalstructures in vertebrates.3–5 LIM-only (LMO) proteins lack ahomeodomain and contain variable numbers of LIM motifs.An LMO protein called muscle LIM protein (MLP) is apositive regulator of skeletal muscle differentiation6,7 and isrequired for maintenance of normal structure and function ofthe myocardium. Mice deficient in this LMO protein developdilated cardiomyopathy during postnatal life.8
A specific subclass of LMO proteins containfour and ahalfLIM motifs and are designated as FHL proteins. Human FHL2,originally known as DRAL (Down-Regulated in Rhabdomyo-sarcoma LIM protein), was first identified by subtractive hybrid-
ization of normal myoblasts versus a rhabdomyosarcoma cellline.9 We isolated a murine homologue of FHL2 from a cardiaccDNA library in a screen for proteins interacting with myocytenuclear factor (MNF), a forkhead/winged helix transcriptionfactor expressed in developing heart and skeletal muscles and inmyogenic progenitor cells (satellite cells) of adult mice.10–12 Inthis report, we describe the temporal and spatial pattern of FHL2expression pattern throughout mouse development. FHL2 ex-pression is evident in the earliest identifiable cardiac precursorcells of the embryo and is restricted primarily to cardiomyocytesduring fetal and adult life. Analysis of homozygous knockoutmice shows that FHL2 is not required for normal cardiacdevelopment. Cardiac mass is increased more, however, in adultmice devoid of FHL2 than in wild-type littermates after admin-istration of theb-adrenergic agonist isoproterenol. We concludethat the FHL2 is an early and persistent marker of the cardio-myocyte lineage in the mouse and can modify responses tocertain stresses in the adult heart.
MethodsCloning of FHL2 Gene and Construction of theTargeting VectorFHL2 cDNA was isolated from a murine cardiac library in a yeast2-hybrid screen using MNF10–12 as bait. Mouse genomic DNA
Received October 19, 2000; revision received December 31, 2000; accepted February 1, 2001.From the Departments of Internal Medicine, Molecular Biology, and Pathology (J.A.R.), University of Texas Southwestern Medical Center, Dallas,
Tex.Correspondence to R. Sanders Williams, University of Texas Southwestern Medical Center, 5323 Harry Hines Blvd, NB11.200, Dallas, TX
75390-8573. E-mail [email protected]© 2001 American Heart Association, Inc.
Circulation is available at http://www.circulationaha.org
2731
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from

encoding FHL2 was isolated from a 129SVJ genomic P1 library(Stratagene) by use of probes based on the cDNA sequence. TheFHL2LacZ/neo targeting vector was constructed in pBluescript (Strat-agene) to include 1.3 kb and 8 kb of homologous sequence regionslocated 59and 39, respectively, to the region of the FHL2 genealtered by homologous recombination. In the mutant FHL2 allele,protein coding sequences from exon 2 (E2) of the native gene,including the translation initiation codon ATG, the N-terminal halfLIM motif, part of the LIM1 domain, and 0.5 kb of intronic sequencedownstream of E2 are replaced with aLacZ reporter gene and aneoresistance cassette. LoxP sites flank theneo cassette. The finalconstruct was verified by restriction analysis and partial sequencing.
Generation of FHL2 Knockout MiceSouthern blot analysis performed on DNA isolated from neomycin-resistant ES clones showed that 8 of 500 ES clones had undergonehomologous recombination of the targeting vector into the FHL2locus. Three targeted ES clones transmitted the mutant allele throughthe germ line and generated FHL2LacZ/neo heterozygote (FHL21/2)animals, which were crossed to generate FHL22/2 mice. The pheno-typic consequences of FHL2 deficiency were equivalent in offspringof the 3 separately derived lines of transgenic mice.
Mouse Embryo AnalysisEmbryonic stage (days postcoitum) was estimated by timed preg-nancies and somite counts. The embryos and decidua were fixed in10% formalin, embedded in paraffin, sectioned, and stained withhematoxylin and eosin. Expression ofb-galactosidase in murineembryos was assessed as previously described.13
Expression of FHL2 mRNA and ProteinRNA was isolated from adult or fetal tissues, and Northern blotswere prepared and probed as previously described.14 Proteins (10mg/lane) were separated by electrophoresis through a 12% SDS-polyacrylamide gel, transferred to nylon filters, and probed with arabbit polyclonal antibody against FHL2. In situ hybridizations wereperformed as previously described.15
Chronic Infusion of IsoproterenolSeven-day osmotic minipumps (Alzet, model 2001) were loadedwith 0.2 mL of isoproterenol (28 mg/mL per 25 g body weight) andimplanted into the subcutaneous space of 6- to 7-month-old mice.After 10 days, hearts were harvested, weighed, and analyzedhistologically.
Statistical AnalysisThe hypertrophic responses to chronic isoproterenol administrationin FHL2 knockout and wild-type mice were compared by theWilcoxon rank sum test.
ResultsConservation of Human and Mouse FHL2cDNA SequencesThe cloned FHL2 cDNA encodes a 279-amino-acid protein thathas 91% homology to the human FHL2 protein9,16,17 (Figure1A). The FHL motifs are arrayed in tandem and separated by 8amino acid linkers that are conserved between human and mouseFHL2 proteins, as well as in FHL1/SLIM1 and FHL3/SLIM2.18,19 A list of currently known FHL and cysteine-richproteins (CRP proteins) is provided in Figure 1B.
Expression of FHL2 in Adult TissuesExpression of FHL2 mRNA in adult mice is detected byNorthern blot analysis only in heart, and not in skeletalmuscle, brain, spleen, liver, kidney, or testis (Figure 2A). Arabbit polyclonal antibody raised against recombinant murine
FHL2 protein detects a 33-kDa protein, approximately thesize predicted from the cDNA sequence, in extracts of adultmouse heart (Figure 2B), with trace levels also evident instomach. In situ hybridization performed with a35S-labeledantisense FHL2 probe on transverse sections of adult cardiacventricles (Figure 2C and 2D) demonstrates the presence ofFHL2 transcripts within cardiomyocytes throughout the ven-tricular free walls and prominently in the septum. In contrast,no FHL2 mRNA is present within cells of the coronaryvasculature (arrows in Figure 2D).
FHL2 Is an Early and Persistent Marker of theCardiomyocyte LineageFHL2 mRNA can be detected by Northern blot analysis asearly as embryonic day 7 (Figure 3A) and continues to bedetected in later-stage embryos. In situ hybridization (Figure3B through 3E) showed that FHL2 transcripts are presentduring heart development; in the common atrium, primitiveventricle and bulbus cordis of the looping heart tube at E9.5;in the common atrium, the developing septal primordium, andcompact and trabecular components of the bulbus cordis atE10.5; in the atria and ventricular free walls but not theatrioventricular cushion at E12.5; and in the myocardium,including the intraventricular septum, but not the outflowtract or atrioventricular cushions at E14.5.
Figure 1. A, Nucleotide sequence and predicted amino acidsequence of mouse and human FHL2 cDNA. Open readingframe codes for a protein of 279 amino acids. Amino acidscomprising LIM domains are bracketed. B, Schematic repre-sentations of secondary structures of FHL and CRP proteinsand summaries of their expression patterns and functions.1,2
2732 Circulation June 5, 2001
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from

FHL2 expression also can be detected in extracardiactissues at certain stages of development (Figure 3B through3E). From E9.5 to E12.5, FHL2 is expressed in undifferen-tiated cells of the head and gut mesenchyme, within theendothelium of the dorsal aorta and branchial arch arteries,and in a narrow subepithelial zone of the developing mid gutand bronchi. At E14.5, FHL2 expression is evident in the wallof the gut, in the urogenital sinus and associated umbilicalartery, and in mesenchyme surrounding the spinal cord.
The earliest stages of cardiac development in vertebratescan be defined by the expression of the homeodomain proteinNkx2.5/Csx20,21 in precardiac mesoderm. At'E7.5, Nkx2.5is expressed in cardiac progenitor cells located at the anteriorand lateral regions of the embryo, which fuse in the midlineto form the cardiac crescent.13,22–24 Whole-mount in situhybridizations performed on littermate embryos (Figure 4)show that FHL2 and Nkx2.5 transcripts are colocalizedwithin precardiac mesoderm of embryos at the early head-fold stage ('E7.5) and at subsequent stages of heartdevelopment.
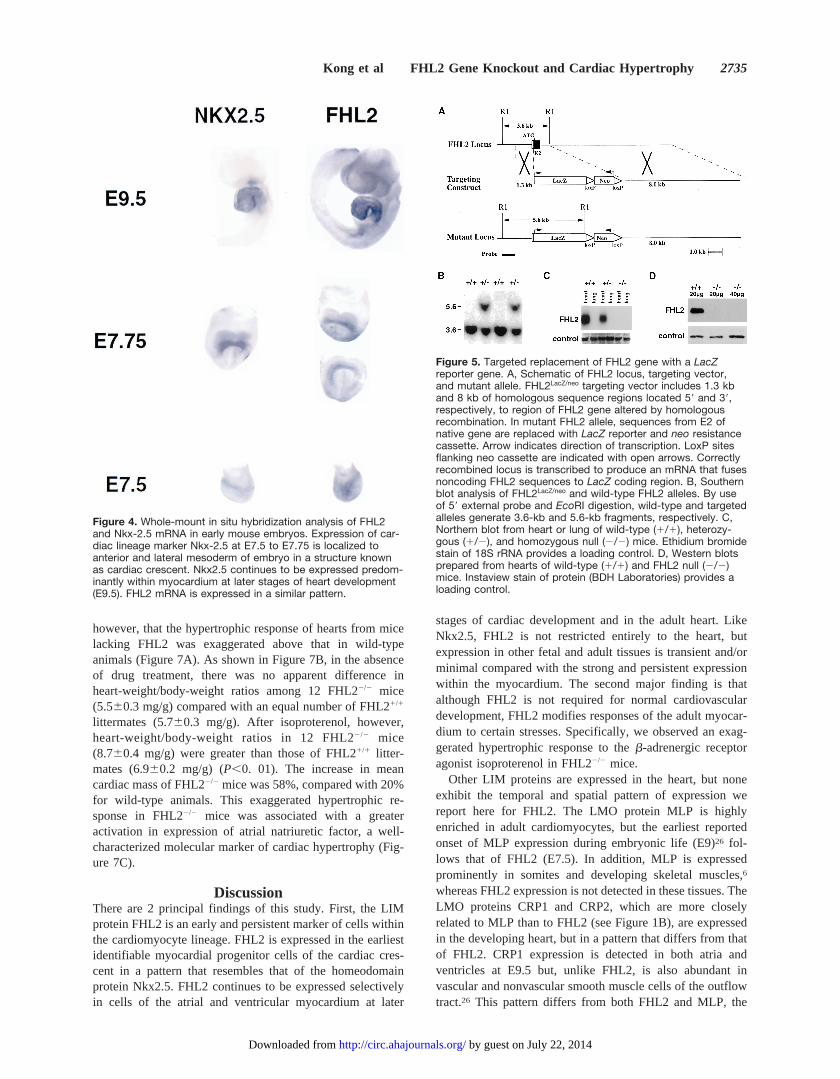
Expression of ab-Galactosidase (LacZ) ReporterGene Inserted Into the Mouse FHL2 Gene LocusHeterozygous FHL2LacZ/neo embryos were generated by ho-mologous recombination of a targeting vector into the FHL2
locus (Figure 5) and were analyzed forb-galactosidaseactivity at stages from E3.5 through E9.5 (Figure 6). In earlyembryos before heart development (E3.5 to E6.5), transcrip-tion of the LacZ transgene is evident in extraembryonictissues, such as polar trophectoderm around the blastocoeliccavity of the intact blastocyst (Figure 6A), and in extraem-bryonic ectoderm at the onset of gastrulation (Figure 6B).FHL2LacZ/neoexpression is observed in the chorion (part of thefuture placenta) of the extraembryonic tissue and in thechorioallantoic placenta at E9.5 (Figure 6A and 6B). At E7.5,b-galactosidase activity is detected in the cardiac crescent,corroborating results obtained with whole-mount in situhybridizations (Figure 4, E7.5). At later stages of heartdevelopment, strong FHL2LacZ/neo expression continues indeveloping cardiomyocytes, but no expression is seen in theendocardium or the bulbotruncal junction region of theoutflow tract of the primitive heart. Weak FHL2LacZ/neoexpres-sion can be detected in the stomach, fat pads, and spleen, butno detectable levels ofLacZexpression are observed in brain,lung, liver, and skeletal muscle (data not shown).
Normal Cardiovascular Development in MiceLacking FHL2The gene-targeting strategy used to insert aLacZ reportergene into the FHL2 locus was designed also to produce a null
Figure 2. Expression of FHL2 mRNA andprotein in adult tissues. A, Distribution ofFHL2 mRNA among tissues of adultmouse. A 1.4-kb FHL2 transcript wasdetected by Northern blot analysis only inheart. B, Distribution of FHL2 proteinamong tissues of adult mouse. A 33-kDaFHL2 protein, as detected by immunoblotanalysis, is most abundant in heart, andtrace amounts are present in stomach.Recombinant FHL2 (rFHL2) produced inEscherichia coli provides a positive con-trol. C, In situ hybridization analysis ofFHL2 mRNA in adult mouse heart. D,Inset of C shown at higher power.Bar5200 mm. FHL2 transcript is presentin cardiac myocytes but not within coro-nary vasculature (arrow).
Kong et al FHL2 Gene Knockout and Cardiac Hypertrophy 2733
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from

allele by eliminating E2 of the FHL2 gene (Figure 5).Genotyping revealed wild-type (FHL21/1), heterozygous(FHL21/2), and homozygous (FHL22/2) offspring close to themendelian prediction, indicating no survival disadvantage forembryos lacking FHL2. The null state was confirmed by theabsence of FHL2 mRNA and protein as assessed by Northern(Figure 5C) and Western (Figure 5D) blot analyses of heartsfrom FHL22/2 mice. Gross and microscopic anatomy ofhearts from adult mice without FHL2 were normal.
Hypertrophic Responses tob-Adrenergic StimulationOsmotic minipumps loaded with 0.2 mL of isoproterenol (28mg/mL per 25 g body weight) were implanted into mice tosubject these animals to the stress of sustainedb-adrenergicstimulation. This stimulus is known to provoke cardiachypertrophy in wild-type mice,25 and we confirmed thisfinding in wild-type littermates of FHL22/2 animals onharvesting the hearts 10 days after implantation. We noted,
Figure 3. Expression of FHL2 mRNA infetal tissues. A, Northern analysis of RNAisolated from mouse embryos at 7, 11,15, and 17 days postcoitum (p.c.). FHL2mRNA is detected as early as day 7 p.c.B through E, In situ hybridization analysisof FHL2 mRNA in mouse embryos.Bar5400 mm. High levels of expressionof FHL2 are evident within myocardiumat all stages from 9.5 to 14.5 days post-coitum (E9.5 to E14.5). Septal primor-dium has particularly high levels of FHL2expression at E14.5. No FHL2 transcriptis detected in cardiac cushion or in out-flow tracts (E). Low to moderate levels ofFHL2 expression are detected in gut andin epithelium around neural tube.
2734 Circulation June 5, 2001
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from
however, that the hypertrophic response of hearts from micelacking FHL2 was exaggerated above that in wild-typeanimals (Figure 7A). As shown in Figure 7B, in the absenceof drug treatment, there was no apparent difference inheart-weight/body-weight ratios among 12 FHL22/2 mice(5.560.3 mg/g) compared with an equal number of FHL21/1
littermates (5.760.3 mg/g). After isoproterenol, however,heart-weight/body-weight ratios in 12 FHL22/2 mice(8.760.4 mg/g) were greater than those of FHL21/1 litter-mates (6.960.2 mg/g) (P,0. 01). The increase in meancardiac mass of FHL22/2 mice was 58%, compared with 20%for wild-type animals. This exaggerated hypertrophic re-sponse in FHL22/2 mice was associated with a greateractivation in expression of atrial natriuretic factor, a well-characterized molecular marker of cardiac hypertrophy (Fig-ure 7C).
DiscussionThere are 2 principal findings of this study. First, the LIMprotein FHL2 is an early and persistent marker of cells withinthe cardiomyocyte lineage. FHL2 is expressed in the earliestidentifiable myocardial progenitor cells of the cardiac cres-cent in a pattern that resembles that of the homeodomainprotein Nkx2.5. FHL2 continues to be expressed selectivelyin cells of the atrial and ventricular myocardium at later
stages of cardiac development and in the adult heart. LikeNkx2.5, FHL2 is not restricted entirely to the heart, butexpression in other fetal and adult tissues is transient and/orminimal compared with the strong and persistent expressionwithin the myocardium. The second major finding is thatalthough FHL2 is not required for normal cardiovasculardevelopment, FHL2 modifies responses of the adult myocar-dium to certain stresses. Specifically, we observed an exag-gerated hypertrophic response to theb-adrenergic receptoragonist isoproterenol in FHL22/2 mice.
Other LIM proteins are expressed in the heart, but noneexhibit the temporal and spatial pattern of expression wereport here for FHL2. The LMO protein MLP is highlyenriched in adult cardiomyocytes, but the earliest reportedonset of MLP expression during embryonic life (E9)26 fol-lows that of FHL2 (E7.5). In addition, MLP is expressedprominently in somites and developing skeletal muscles,6
whereas FHL2 expression is not detected in these tissues. TheLMO proteins CRP1 and CRP2, which are more closelyrelated to MLP than to FHL2 (see Figure 1B), are expressedin the developing heart, but in a pattern that differs from thatof FHL2. CRP1 expression is detected in both atria andventricles at E9.5 but, unlike FHL2, is also abundant invascular and nonvascular smooth muscle cells of the outflowtract.26 This pattern differs from both FHL2 and MLP, the
Figure 4. Whole-mount in situ hybridization analysis of FHL2and Nkx-2.5 mRNA in early mouse embryos. Expression of car-diac lineage marker Nkx-2.5 at E7.5 to E7.75 is localized toanterior and lateral mesoderm of embryo in a structure knownas cardiac crescent. Nkx2.5 continues to be expressed predom-inantly within myocardium at later stages of heart development(E9.5). FHL2 mRNA is expressed in a similar pattern.
Figure 5. Targeted replacement of FHL2 gene with a LacZreporter gene. A, Schematic of FHL2 locus, targeting vector,and mutant allele. FHL2LacZ/neo targeting vector includes 1.3 kband 8 kb of homologous sequence regions located 59 and 39,respectively, to region of FHL2 gene altered by homologousrecombination. In mutant FHL2 allele, sequences from E2 ofnative gene are replaced with LacZ reporter and neo resistancecassette. Arrow indicates direction of transcription. LoxP sitesflanking neo cassette are indicated with open arrows. Correctlyrecombined locus is transcribed to produce an mRNA that fusesnoncoding FHL2 sequences to LacZ coding region. B, Southernblot analysis of FHL2LacZ/neo and wild-type FHL2 alleles. By useof 59 external probe and EcoRI digestion, wild-type and targetedalleles generate 3.6-kb and 5.6-kb fragments, respectively. C,Northern blot from heart or lung of wild-type (1/1), heterozy-gous (1/2), and homozygous null (2/2) mice. Ethidium bromidestain of 18S rRNA provides a loading control. D, Western blotsprepared from hearts of wild-type (1/1) and FHL2 null (2/2)mice. Instaview stain of protein (BDH Laboratories) provides aloading control.
Kong et al FHL2 Gene Knockout and Cardiac Hypertrophy 2735
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from

expression of which within the heart is restricted to cardio-myocytes. FHL1/SLIM1 and FHL3/SLIM2 are closely re-lated structurally to FHL2 and are expressed in skeletalmuscles and heart.16,18,19,27–30 Although FHL2 exhibits agreater degree of cardiac specificity than other known LIMproteins, it is possible that related FHL proteins can substitutefor developmental functions of FHL2 within in the develop-ing heart.
The present study did not address biochemical functions ofFHL2, but the literature provides several interesting leads forfuture investigations. The FHL protein ACT functions as atranscriptional coactivator with CREM and CREB proteins.31
FHL2 itself was recently shown to function as a coactivator inassociation with androgen receptors.32 It will be interesting todetermine whether FHL2 collaborates with nuclear receptorsor CREM/CREB proteins in the myocardium, particularlybecause transgenic mice expressing a dominant negative formof CREB develop dilated cardiomyopathy.33 Our own pre-liminary data showing a physical association between FHL2-and DNA-binding MNF proteins 10212 also lend credence tothe speculation that FHL2 may be a transcriptional modulatorin the heart.
Among the set of transcriptional regulators known to havean important role in cardiac development, FHL2 expressionmost closely parallels that of the homeodomain proteinNkx2.5. The colocalized expression of FHL2 and Nkx2.5suggests either that these 2 genes are responding to a commonset of upstream regulators or that one may serve to control theother. In either case, detailed examination of promoter andenhancer elements from the FHL2 gene, in parallel withsimilar studies of Nkx2.5 gene regulation, may shed light ontranscriptional events pertinent to the earliest steps of cardio-myocyte determination.
Finally, the observation that FHL22/2 mice have anexaggerated response to the hypertrophic effects of isopro-terenol suggests that FHL2 interacts with downstreameffectors ofb-adrenergic receptor signaling in the myocar-dium or with proteins shared among different hypertrophicsignaling pathways. As a gene that is not required fornormal development but is capable of modifying cardiacresponses to environmental stress, it is possible that allelicvariants of the FHL2 gene may be present within humanpopulations and influence the natural history of cardiovas-cular diseases in a clinical setting.
Figure 6. FHL2LacZ/neo expression in earlymouse embryos. LacZ staining of whole-mount preparations from embryos har-vested 3.5 to 9.5 days postcoitum. A, AtE3.5, embryo, LacZ-positive cells areseen around blastocoelic cavity. B,Expression was localized to extraembry-onic tissue at E6.5. C and D, As gastru-lation proceeds, FHL2LacZ/neo expressionis localized to chorion of extraembryonictissue and to cardiac crescent. E and F,LacZ staining is evident predominantly indeveloping heart, with some LacZ-positive cells seen within allantois. G, H,and I, Histological sections of embryosin C, D, and E. Bar5200 mm.
2736 Circulation June 5, 2001
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from

Acknowledgments
This work was supported by grants from the NIH (AR-40849,HL-61624, GM-62114, and HL-07360) and the D.W. ReynoldsFoundation. We thank S. Clay Williams for assistance with grossdissections and transcardial perfusions and Jeffrey M. Stark forpreparation of histological sections.
References1. Jurata LW, Gill GN. Structure and function of LIM domains.Curr Top
Microbiol Immunol. 1998;228:75–113.2. Bach I. The LIM domain: regulation by association.Mech Dev. 2000;
91:5–17.3. Shawlot W, Behringer RR. Requirement for Lim1 in head-organizer
function.Nature. 1995;374:425–430.
Figure 7. Hypertrophic response tob-adrenergic stimulation is augmented inmice lacking FHL2. A, Trichrome-stainedhistological sections of adult heart fromuntreated or isoproterenol-treatedFHL21/1 and FLH22/2 mice. Multifocalareas of myocardial fibrosis (blue) areevident within left ventricular free wall ofisoproterenol-treated FHL22/2 animal. B,Heart-weight–to–body-weight ratios areincreased by infusion of isoproterenolfrom osmotic minipumps in both wild-type (FHL21/1) and mutant (FHL22/2)mice. Magnitude of increase in FHL22/2
animals (59%), however, was greaterthan that observed in FHL21/1 mice(20%). C, Northern blot analysis showingincreased expression of hypertrophicmarker atrial natriuretic factor in micelacking FHL2 compared with wild-typemice after isoproterenol treatment.
Kong et al FHL2 Gene Knockout and Cardiac Hypertrophy 2737
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from

4. Sheng HZ, Moriyama K, Yamashita T, et al. Multistep control of pituitaryorganogenesis.Science. 1997;278:1809–1812.
5. Pfaff SL, Mendelsohn M, Stewart CL, et al. Requirement for LIMhomeobox gene Isl1 in motor neuron generation reveals a motor neuron-dependent step in interneuron differentiation.Cell. 1996;84:309–320.
6. Arber S, Halder G, Caroni P. Muscle LIM protein, a novel essentialregulator of myogenesis, promotes myogenic differentiation.Cell. 1994;79:221–231.
7. Kong YF, Flick MJ, Kudla AJ, et al. Muscle LIM protein promotesmyogenesis by enhancing the activity of myoD.Mol Cell Biol. 1997;17:4750–4760.
8. Arber S, Hunter JJ, Ross J, et al. MLP-deficient mice exhibit a disruptionof cardiac cytoarchitectural organization, dilated cardiomyopathy, andheart failure.Cell. 1997;88:393–403.
9. Genini M, Schwalbe P, Scholl FA, et al. Subtractive cloning and char-acterization of DRAL, a novel LIM-domain protein down-regulated inrhabdomyosarcoma.DNA Cell Biol. 1997;16:433–442.
10. Garry DJ, Yang Q, Bassel-Duby R, et al. Persistent expression of MNFidentifies myogenic stem cells in postnatal muscles.Dev Biol. 1997;188:280–294.
11. Yang Q, Bassel-Duby R, Williams RS. Transient expression of awinged-helix protein, MNF-beta, during myogenesis.Mol Cell Biol.1997;17:5236–5243.
12. Yang Q, Kong Y, Rothermel B, et al. The winged-helix/forkhead proteinmyocyte nuclear factor beta (MNF-beta) forms a co-repressor complexwith mammalian sin3B.Biochem J. 2000;345:335–343.
13. Reecy JM, Li X, Yamada M, et al. Identification of upstream regulatoryregions in the heart-expressed homeobox gene Nkx2-5.Development.1999;126:839–849.
14. Garry DJ, Bassel-Duby R, Richardson JA, et al. Postnatal developmentand plasticity of specialized muscle fiber characteristics in the hindlimb.Dev Genetics. 1996;19:146–156.
15. Shelton JM, Lee MH, Richardson JA, et al. Microsomal triglyceridetransfer protein expression during mouse development.J Lipid Res.2000;41:532–537.
16. Morgan MJ, Madgwick AJ. Slim defines a novel family of LIM-proteinsexpressed in skeletal muscle.Biochem Biophys Res Commun. 1996;225:632–638.
17. Chan KK, Tsui SK, Lee SM, et al. Molecular cloning and characterizationof FHL2, a novel LIM domain protein preferentially expressed in humanheart.Gene. 1998;210:345–350.
18. Morgan MJ, Madgwick AJ. The LIM proteins FHL1 and FHL3 areexpressed differently in skeletal muscle.Biochem Biophys Res Commun.1999;255:245–250.
19. Brown S, Biben C, Ooms LM, et al. The cardiac expression of striatedmuscle LIM protein 1 (SLIM1) is restricted to the outflow tract of thedeveloping heart.J Mol Cell Cardiol. 1999;31:837–843.
20. Lints TJ, Parsons LM, Hartley L, et al. Nkx-2.5: a novel murinehomeobox gene expressed in early heart progenitor cells and theirmyogenic descendants.Development. 1993;119:419–431.
21. Komuro I, Izumo S. Csx: a murine homeobox-containing gene specif-ically expressed in the developing heart.Proc Natl Acad Sci U S A.1993;90:8145–8149.
22. Tanaka M, Wechsler SB, Lee IW, et al. Complex modular cis-actingelements regulate expression of the cardiac specifying homeobox geneCsx/Nkx2.5.Development. 1999;126:1439–1450.
23. Searcy RD, Vincent EB, Liberatore CM, et al. A GATA-dependentnkx-2.5 regulatory element activates early cardiac gene expression intransgenic mice.Development. 1998;125:4461–4470.
24. Lien CL, Wu C, Mercer B, et al. Control of early cardiac-specifictranscription of Nkx2-5 by a GATA-dependent enhancer.Development.1999;126:75–84.
25. Kudej RK, Iwase M, Uechi M, et al. Effects of chronic beta-adrenergicreceptor stimulation in mice.J Mol Cell Cardiol. 1997;29:2735–2746.
26. Jain MK, Kashiki S, Hsieh CM, et al. Embryonic expression suggests animportant role for CRP2/SmLIM in the developing cardiovascularsystem.Circ Res. 1998;83:980–985.
27. Chu P, Ruiz-Lozano P, Zhou Q, et al. Expression patterns of FHL/SLIMfamily members suggest important functional roles in skeletal muscle andcardiovascular system.Mech Dev. 2000;95:259–265.
28. Brown S, McGrath MJ, Ooms LM, et al. Characterization of two isoforms ofthe skeletal muscle LIM protein 1, SLIM1: localization of SLIM1 at focaladhesions and the isoform slimmer in the nucleus of myoblasts andcytoplasm of myotubes suggests distinct roles in the cytoskeleton and innuclear-cytoplasmic communication.J Biol Chem. 1999;274:27083–27091.
29. Greene WK, Baker E, Rabbitts TH, et al. Genomic structure, tissueexpression and chromosomal location of the LIM-only gene, SLIM1.Gene. 1999;232:203–207.
30. Morgan MJ, Madgwick AJA. The LIM proteins FHL1 and FHL3 areexpressed differently in skeletal muscle.Biochem Biophys Res Commun.1999;255:245–250.
31. Fimia GM, De Cesare D, Sassone-Corsi P. CBP-independent activation ofCREM and CREB by the LIM-only protein ACT.Nature. 1999;398:165–169.
32. Muller JM, Isele U, Metzger E, et al. FHL2, a novel tissue-specificcoactivator of the androgen receptor.EMBO J. 2000;19:359–369.
33. Fentzke RC, Korcarz CE, Lang RM, et al. Dilated cardiomyopathy intransgenic mice expressing a dominant-negative CREB transcriptionfactor in the heart.J Clin Invest. 1998;101:2415–2426.
2738 Circulation June 5, 2001
by guest on July 22, 2014http://circ.ahajournals.org/Downloaded from




















