
Copulation modifies AR and ERα mRNA expression in the male rat brain
8
Copulation modifies AR and ERα mRNA expression in the male rat brain Alonso Fernández-Guasti ⁎, Paola Arteaga-López, Edwards Antonio-Cabrera Department of Pharmacobiology, CINVESTAV, Calzada de los Tenorios 235, Col. Granjas Coapa, Del. Tlalpan, México City, D. F. 14330 México abstract article info Article history: Received 22 January 2010 Received in revised form 13 August 2010 Accepted 23 August 2010 Keywords: Rat male sexual satiety Limbic areas Androgen- and estrogen alpha-receptor messenger ribonucleic acid In situ hybridization Background/Aims: One day after male sexual behavior [one ejaculation or copulation to satiety (ad libitum copulation during 4 h with the same female)] androgen receptor immunoreactivity (AR-ir) is decreased and estrogen receptor alpha immunoreactivity (ERα-ir) increased in various brain areas related with its control. Seven days after sexual satiety there was a limited recovery of sexual behavior accompanied by a partial recuperation in the AR-ir. In this study we evaluated if these changes in AR-ir and ERα-ir were paralleled by variations in their respective mRNA. Methods: Sexually experienced male rats were sacrificed at different intervals: immediately, 24 h or seven days after sexual satiety or 24 h after one ejaculation. The changes in AR and ERα mRNA were analyzed by in situ hybridization using digoxigenine-labeled oligonucleotide probes in the MPOA, LSV and the bed nucleus of the stria terminalis, medial division, anterior (BSTMA). Results: AR mRNA density was decreased in the MPOA and the LSV immediately and 24 h after one ejaculation or sexual satiety. Seven days after copulating to satiety, there was a recovery of AR mRNA. In the BSTMA the different behavioral conditions did not modify the AR mRNA expression. In the MPOA, LSV and BSTMA the ERα mRNA increased after a single ejaculation and at all intervals after sexual satiety. Conclusion: In some brain areas and after some intervals of sexual activity, the changes in steroid protein receptors expression seem to be consequence of parallel changes in the expression of the respective mRNA. © 2010 Elsevier Inc. All rights reserved. 1. Introduction Androgens regulate a wide variety of masculine physiological functions, including copulatory behavior [1–3] and, at least partly, this effect is mediated via their conversion to estrogens [4]. Most androgen and estrogen actions are exerted at their intracellular receptors, which expression is concentrated in limbic areas involved in the regulation of male sexual behavior, such as the medial preoptic area (MPOA), lateral septum (LS), bed nucleus of the stria terminals (BST), ventromedial hypothalamic nucleus (VMH), medial amygdala (MeA) and arcuate nucleus (Arc) [5–9]. The functional relationship between steroid sensitive neurons and male sexual behavior in these brain areas arises primarily from co-expression and direct steroid activation results. Regarding the former, a high percentage of neurons activated by sexual behavior (established by c-fos) expresses androgen receptor (AR), estrogen receptor alpha (ERα) or both [6]. In relation with the later, direct implant of androgens or estrogens in some of these brain areas facilitates copulatory behavior [5,10–12], and inversely, AR blockade in MPOA and VMH effectively inhibit sexual behavior in non-castrated males [2,13]. As aforementioned, there is evidence indicating that androgens promote male sexual activity via their aromatization to estrogens which act primarily on the ERα. Thus aromatase inhibition results in a loss of mating [14,15] and knock-out mice lacking either the aromatase or the ERα (but not the ERβ)-gene present severe deficits in mating behavior [16–18]. In male rats the sexual satiety phenomenon is defined as the inhibition of sexual behavior triggered by repetitive ejaculations [19– 21]. Recently we reported that in MPOA, VMH, LSV and nucleus accumbens (Acb) the sexual behavior inhibition was paralleled by a reduction in AR-immunoreactivity (AR-ir) [8,21]. However, one ejaculation or copulation to satiety did not modify the expression of AR-ir in the BST [8,9,21]. Interestingly, the reduction in AR-ir triggered by sexual behavior was not accompanied by changes in the levels of serum androgens [9,21]. The functional association between the reduction in AR-ir and the inhibition of sexual behavior was studied by analyzing whether the recovery from sexual satiety was accom- panied by an increase of the AR density. Remarkably, the inhibition of sexual behavior coincided with a decrease in AR-ir in the MPOA 24 and 48 h after sexual satiety, while seven days later, when there is a partial recovery of copulatory behavior -evidenced as the capacity of all animals to show at least two ejaculations-, the AR density returned to its basal levels [9]. Further functional evidence included the restoration of copulatory behavior in sexually satiated males by the administration of a mix of anabolic androgens that induce an over- expression of AR [22]. As to the ERα, we recently reported that 24 h after sexual satiety there was an increase in ERα density in the BST, LSV, MeA, MPOA and Acb. A single ejaculation was related to an increase in ERα density only in the BST and MeA. ERα density in the Physiology & Behavior 101 (2010) 738–745 ⁎ Corresponding author. Tel.: +52 55 5483 2870; fax: +52 55 5483 2863. E-mail address: [email protected] (A. Fernández-Guasti). 0031-9384/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.physbeh.2010.08.015 Contents lists available at ScienceDirect Physiology & Behavior journal homepage: www.elsevier.com/locate/phb
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Copulation modifies AR and ERα mRNA expression in the male rat brain

Physiology & Behavior 101 (2010) 738–745
Contents lists available at ScienceDirect
Physiology & Behavior
j ourna l homepage: www.e lsev ie r.com/ locate /phb
Copulation modifies AR and ERα mRNA expression in the male rat brain
Alonso Fernández-Guasti ⁎, Paola Arteaga-López, Edwards Antonio-CabreraDepartment of Pharmacobiology, CINVESTAV, Calzada de los Tenorios 235, Col. Granjas Coapa, Del. Tlalpan, México City, D. F. 14330 México
⁎ Corresponding author. Tel.: +52 55 5483 2870; faxE-mail address: [email protected] (A. Fernánd
0031-9384/$ – see front matter © 2010 Elsevier Inc. Aldoi:10.1016/j.physbeh.2010.08.015
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 22 January 2010Received in revised form 13 August 2010Accepted 23 August 2010Keywords:Rat male sexual satietyLimbic areasAndrogen- and estrogen alpha-receptormessenger ribonucleic acidIn situ hybridization
Background/Aims: One day after male sexual behavior [one ejaculation or copulation to satiety (ad libitumcopulation during 4 h with the same female)] androgen receptor immunoreactivity (AR-ir) is decreased andestrogen receptor alpha immunoreactivity (ERα-ir) increased in various brain areas related with its control.Seven days after sexual satiety there was a limited recovery of sexual behavior accompanied by a partialrecuperation in the AR-ir. In this study we evaluated if these changes in AR-ir and ERα-ir were paralleled byvariations in their respective mRNA.Methods: Sexually experienced male rats were sacrificed at different intervals: immediately, 24 h or sevendays after sexual satiety or 24 h after one ejaculation. The changes in AR and ERα mRNA were analyzed by insitu hybridization using digoxigenine-labeled oligonucleotide probes in the MPOA, LSV and the bed nucleusof the stria terminalis, medial division, anterior (BSTMA).
Results: AR mRNA density was decreased in the MPOA and the LSV immediately and 24 h after oneejaculation or sexual satiety. Seven days after copulating to satiety, there was a recovery of AR mRNA. In theBSTMA the different behavioral conditions did not modify the AR mRNA expression. In the MPOA, LSV andBSTMA the ERα mRNA increased after a single ejaculation and at all intervals after sexual satiety.Conclusion: In some brain areas and after some intervals of sexual activity, the changes in steroid proteinreceptors expression seem to be consequence of parallel changes in the expression of the respective mRNA.© 2010 Elsevier Inc. All rights reserved.
1. Introduction
Androgens regulate a wide variety of masculine physiologicalfunctions, including copulatory behavior [1–3] and, at least partly, thiseffect is mediated via their conversion to estrogens [4]. Most androgenand estrogen actions are exerted at their intracellular receptors, whichexpression is concentrated in limbic areas involved in the regulationof male sexual behavior, such as the medial preoptic area (MPOA),lateral septum (LS), bed nucleus of the stria terminals (BST),ventromedial hypothalamic nucleus (VMH), medial amygdala(MeA) and arcuate nucleus (Arc) [5–9]. The functional relationshipbetween steroid sensitive neurons and male sexual behavior in thesebrain areas arises primarily from co-expression and direct steroidactivation results. Regarding the former, a high percentage of neuronsactivated by sexual behavior (established by c-fos) expressesandrogen receptor (AR), estrogen receptor alpha (ERα) or both [6].In relation with the later, direct implant of androgens or estrogens insome of these brain areas facilitates copulatory behavior [5,10–12],and inversely, AR blockade in MPOA and VMH effectively inhibitsexual behavior in non-castrated males [2,13]. As aforementioned,there is evidence indicating that androgens promote male sexualactivity via their aromatization to estrogens which act primarily on
: +52 55 5483 2863.ez-Guasti).
l rights reserved.
the ERα. Thus aromatase inhibition results in a loss of mating [14,15]and knock-out mice lacking either the aromatase or the ERα (but notthe ERβ)-gene present severe deficits in mating behavior [16–18].
In male rats the sexual satiety phenomenon is defined as theinhibition of sexual behavior triggered by repetitive ejaculations [19–21]. Recently we reported that in MPOA, VMH, LSV and nucleusaccumbens (Acb) the sexual behavior inhibition was paralleled by areduction in AR-immunoreactivity (AR-ir) [8,21]. However, oneejaculation or copulation to satiety did not modify the expression ofAR-ir in the BST [8,9,21]. Interestingly, the reduction in AR-ir triggeredby sexual behavior was not accompanied by changes in the levels ofserum androgens [9,21]. The functional association between thereduction in AR-ir and the inhibition of sexual behavior was studiedby analyzing whether the recovery from sexual satiety was accom-panied by an increase of the AR density. Remarkably, the inhibition ofsexual behavior coincided with a decrease in AR-ir in the MPOA 24and 48 h after sexual satiety, while seven days later, when there is apartial recovery of copulatory behavior -evidenced as the capacity ofall animals to show at least two ejaculations-, the AR density returnedto its basal levels [9]. Further functional evidence included therestoration of copulatory behavior in sexually satiated males by theadministration of a mix of anabolic androgens that induce an over-expression of AR [22]. As to the ERα, we recently reported that 24 hafter sexual satiety there was an increase in ERα density in the BST,LSV, MeA, MPOA and Acb. A single ejaculation was related to anincrease in ERα density only in the BST and MeA. ERα density in the

739A. Fernández-Guasti et al. / Physiology & Behavior 101 (2010) 738–745
Arc and VMH and serum estradiol levels remained unchanged 24 hafter one ejaculation or mating to satiety [7].
The levels of AR and ERα depend upon their synthesis, derivedfrom the translation of their mRNA, and from the activity of theproteasome that is involved in their degradation [23]. Interestingly, asfor other steroid receptors, the amount of AR and ERα also dependupon the concentration of their ligands, since for instance androgenspromote the synthesis and prevent the breakdown of AR [24,25].Accordingly, castration results in a rapid vanishing of AR in the centralnervous system, while androgens supplementation produces anincrease in the levels of AR [22,26–28]. The regulation of theexpression of the ERα in the brain seems more complex [29]. Thus,castration increases ERα mRNA in the MPOA of young male rats [26].By direct contrast, others reported that estradiol binding in the MPOAwas significantly lower in castrated compared to intact male rats [30].Treatment of male rats with hormones also affects ERα binding andimmunoreactivity. Clancy et al. [10] and Yamada et al. [31] reportedthat counts of ERα-ir cells in the MPOA, VMH and Arc weresignificantly lower in estradiol-treated male rat compared to thecontrol, and this reduction was region specific, as a difference was notdetected in the BST, LS, anteroventral periventricular nucleus orlateral mesencephalic central gray. These data indicate that in themale brain, as in the female [32], other factors interact to control ERαmRNA or protein expression in addition to simply estradiol levels [33].
The aim of the present study was to evaluate the putative relationbetween changes in AR- and ERα-mRNA levels and the expression ofsexual behavior. First we studied whether the observed decrease inAR [8,9,21] and increase in ERα [7] were paralleled by a reduction inAR mRNA and an enhancement in ERα mRNA to infer whether thedown- or up-regulation of these receptors—produced by sexualactivity—is transcriptional or translational in nature. Secondly, ifsuch parallelism is observed, would the changes in gonadal steroidreceptors reveal a putative association with the inhibition of sexualbehavior triggered by copulation? That is, if at a maximal level ofsexual behavior inhibition, for example, immediately after sexualsatiety (after 4 h of ad libitum copulation with a single sexually-receptive female), there are more pronounced changes in the levels ofAR or ERα mRNA than seven days later when a partial recovery fromsexual satiety occurs. In addition, to further explore whether theinhibition of sexual behavior could be caused by additive changes insteroid receptors levels, triggered by successive ejaculations, weadded a group of males that copulated for a single ejaculationwhich isknown to slightly modify sexual performance 24 h later [20,21]. TheMPOA, LSV and BST were selected because of the following reasons: a)their important role on rat copulatory behavior (vide supra); b) thecomparison between the putative changes in the mRNA for AR andERα with those reported for their respective proteins, and c) theinclusion of brain areas -MPOA and LSV- that show drastic changes insteroid receptors (AR- and ERα-ir) after sexual satiety [7,21], and ofanother—the BST—where the AR-ir was unmodified after satiety [21].In addition, the neocortex (anterior cingulated cortex, motor andsomatosensory areas of the frontoparietal cortices) served as a controlbrain area that does not importantly participate in the neural orneuroendocrine control of masculine sexual behavior.
2. Materials and methods
2.1. Animals
Adult male (350–400 g) albino Wistar rats were used for allexperiments. Animals were kept four to six per cage, with ad libitumaccess to rat chow and tap water, under an inverted light–dark cycle(12 h each, lights off at 10:00 AM) at 22 °C. Ratmanagement was doneaccording to the general principles of NIH publication 85-23, 1985. Allthe experiments were approved by the local committee of CINVESTAVfor ethics on animal experimentation.
2.2. Training tests
After adapting at least for one week to the inverted light–darkcycle, all males were given eight mating training tests twice weekly.All the observations were done 1 h after the onset of darkness andunder dim red light. All males were transferred, one each, tocylindrical observation cages. After a 5 min adaptation period, onereceptive female was placed with each animal. Female receptivity wasinduced by sequential s.c. injections of estradiol benzoate (8 μg/rat)followed by progesterone (2 mg/rat) 24 and 4 h before the tests,respectively. The copulatory behavior parameters recorded were:intromission latency, number of mounts and intromissions andejaculation latency. The female was removed after the male displayedone ejaculation or after a 30 min period if the animal did not show anintromission. Only the males that showed ejaculation latenciesshorter than 15 min in at least half of the tests were used for furtherexperiments.
A control group of sexually experienced males was sacrificed bydecapitation at least one week after the last training test. Theexperimental groups consisted of animals sacrificed at differentintervals: immediately, 24 h or seven days after sexual satiety. Sexualsatiety was attained after exposing a sexually experiencedmale with asingle receptive female for an ad libitum copulating period of 4 h.Previous data from our laboratory [34] showed that in 4 h all malesreach the sexual satiety criterion defined as the complete absence ofsexual behavior for at least 90 min. In addition a group of animals wassacrificed 24 h after they copulated until one ejaculation. Thesegroups and intervals were chosen since previous reports showedimportant changes in AR and ERα established by immunocytochem-istry in these brain areas, and because the inhibition of sexualbehavior triggered by repeated ejaculation is maximal immediatelyafter and 24 h following satiety [21,34], while seven days later there isa partial recovery of sexual behavior [9]. One day after a singleejaculation there is a very minor inhibition of sexual performance[20]. The n for the groups was of 4–6 animals per receptor subtype.
The rats were decapitated and the brain was obtained bydissection and washed in a cold and sterile saline solution and thenstored at−70 °C until assayed. The brains weremounted on a cryostat(−17 °C) and cut at 8 μm sections that were mounted onto poly L-lysine coated slides (75×50 mm). The brain was cut at the level of−0.40 mm anterior to bregma according to the Paxinos and Watsoncerebral atlas [35] to have in the same section the LS, the MPOA andthe BST and at a similar antero-posterior level, within these areas thesubnuclei selected were the lateral septal nucleus, ventral part (LSV)and the bed nucleus of the stria terminalis, medial division, anterior(BSTMA). These subnuclei were selected on the bases of previousreports showing changes in steroid receptor proteins associated withsexual activity [7–9,21]. In this same section the neocortex expressionof the gonadal steroid receptors was visualized.
2.3. In situ hybridization
In situ hybridization runs included sections of animals in allbehavioral conditions. Digoxigenine-labeled oligonucleotide probeswere obtained from GeneDetect.com LTD (http://www.genedetect.com; Auckland, New Zealand). For AR mRNA the 48mer antisenseoligonucleotide probe hybridizes to nucleotides 755–802 usingGenBank accession number NM_012502. For ER-α mRNA the 28merantisense oligonucleotide probe hybridizes to nucleotides 1633–1661using GenBank accession number NM_012689.1.
Antisense probes were labeled using digoxigenine (DIG). We useda commercial probe hyperlabelledwith five DIGmolecules attached tothe tail of the oligonucleotide. We followed this technique since it iseasy to detect and has a similar sensitivity than that of otherprocedures [36–38]. The use of DIG to label nucleic acids is widelyused in molecular biology experiments. The label is introduced by
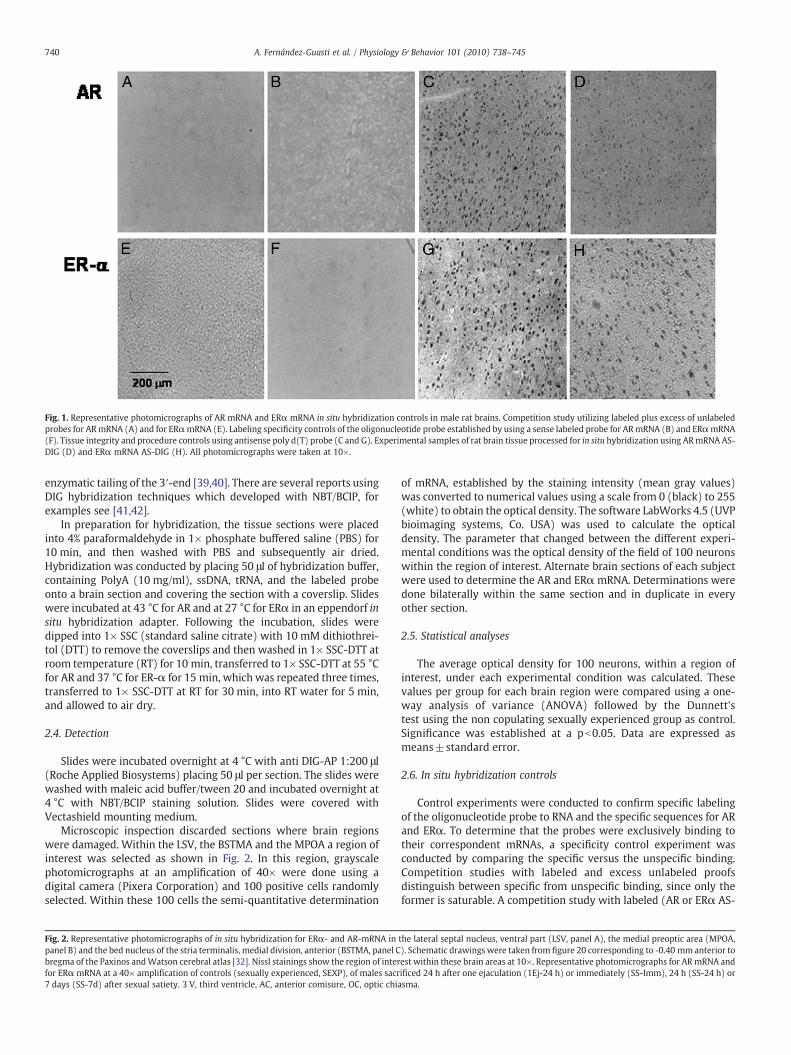
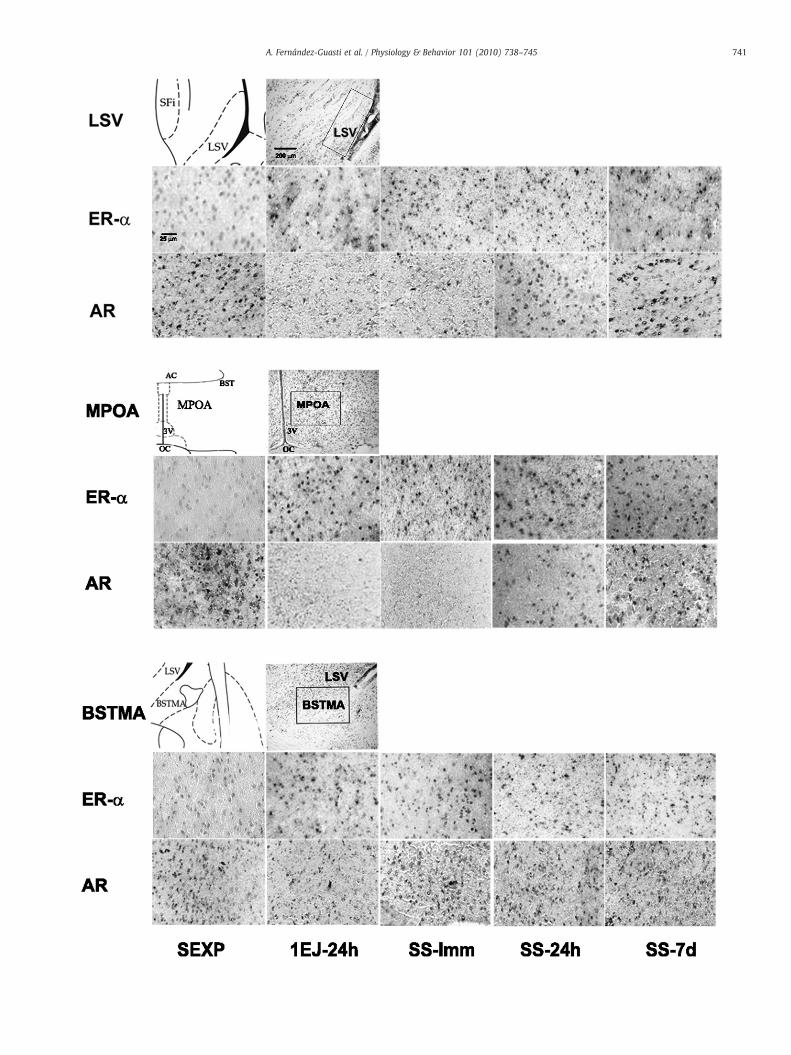
Fig. 1. Representative photomicrographs of AR mRNA and ERα mRNA in situ hybridization controls in male rat brains. Competition study utilizing labeled plus excess of unlabeledprobes for ARmRNA (A) and for ERαmRNA (E). Labeling specificity controls of the oligonucleotide probe established by using a sense labeled probe for ARmRNA (B) and ERαmRNA(F). Tissue integrity and procedure controls using antisense poly d(T) probe (C and G). Experimental samples of rat brain tissue processed for in situ hybridization using ARmRNA AS-DIG (D) and ERα mRNA AS-DIG (H). All photomicrographs were taken at 10×.
740 A. Fernández-Guasti et al. / Physiology & Behavior 101 (2010) 738–745
enzymatic tailing of the 3′-end [39,40]. There are several reports usingDIG hybridization techniques which developed with NBT/BCIP, forexamples see [41,42].
In preparation for hybridization, the tissue sections were placedinto 4% paraformaldehyde in 1× phosphate buffered saline (PBS) for10 min, and then washed with PBS and subsequently air dried.Hybridization was conducted by placing 50 μl of hybridization buffer,containing PolyA (10 mg/ml), ssDNA, tRNA, and the labeled probeonto a brain section and covering the section with a coverslip. Slideswere incubated at 43 °C for AR and at 27 °C for ERα in an eppendorf insitu hybridization adapter. Following the incubation, slides weredipped into 1× SSC (standard saline citrate) with 10 mM dithiothrei-tol (DTT) to remove the coverslips and then washed in 1× SSC-DTT atroom temperature (RT) for 10 min, transferred to 1× SSC-DTT at 55 °Cfor AR and 37 °C for ER-α for 15 min, which was repeated three times,transferred to 1× SSC-DTT at RT for 30 min, into RT water for 5 min,and allowed to air dry.
2.4. Detection
Slides were incubated overnight at 4 °C with anti DIG-AP 1:200 μl(Roche Applied Biosystems) placing 50 μl per section. The slides werewashed with maleic acid buffer/tween 20 and incubated overnight at4 °C with NBT/BCIP staining solution. Slides were covered withVectashield mounting medium.
Microscopic inspection discarded sections where brain regionswere damaged. Within the LSV, the BSTMA and the MPOA a region ofinterest was selected as shown in Fig. 2. In this region, grayscalephotomicrographs at an amplification of 40× were done using adigital camera (Pixera Corporation) and 100 positive cells randomlyselected. Within these 100 cells the semi-quantitative determination
Fig. 2. Representative photomicrographs of in situ hybridization for ERα- and AR-mRNA inpanel B) and the bed nucleus of the stria terminalis, medial division, anterior (BSTMA, panel Cbregma of the Paxinos andWatson cerebral atlas [32]. Nissl stainings show the region of interfor ERαmRNA at a 40× amplification of controls (sexually experienced, SEXP), of males sacr7 days (SS-7d) after sexual satiety. 3 V, third ventricle, AC, anterior comisure, OC, optic chia
of mRNA, established by the staining intensity (mean gray values)was converted to numerical values using a scale from 0 (black) to 255(white) to obtain the optical density. The software LabWorks 4.5 (UVPbioimaging systems, Co. USA) was used to calculate the opticaldensity. The parameter that changed between the different experi-mental conditions was the optical density of the field of 100 neuronswithin the region of interest. Alternate brain sections of each subjectwere used to determine the AR and ERα mRNA. Determinations weredone bilaterally within the same section and in duplicate in everyother section.
2.5. Statistical analyses
The average optical density for 100 neurons, within a region ofinterest, under each experimental condition was calculated. Thesevalues per group for each brain region were compared using a one-way analysis of variance (ANOVA) followed by the Dunnett'stest using the non copulating sexually experienced group as control.Significance was established at a pb0.05. Data are expressed asmeans±standard error.
2.6. In situ hybridization controls
Control experiments were conducted to confirm specific labelingof the oligonucleotide probe to RNA and the specific sequences for ARand ERα. To determine that the probes were exclusively binding totheir correspondent mRNAs, a specificity control experiment wasconducted by comparing the specific versus the unspecific binding.Competition studies with labeled and excess unlabeled proofsdistinguish between specific from unspecific binding, since only theformer is saturable. A competition study with labeled (AR or ERα AS-
the lateral septal nucleus, ventral part (LSV, panel A), the medial preoptic area (MPOA,). Schematic drawings were taken from figure 20 corresponding to -0.40 mm anterior toest within these brain areas at 10×. Representative photomicrographs for ARmRNA andificed 24 h after one ejaculation (1Ej-24 h) or immediately (SS-Imm), 24 h (SS-24 h) orsma.
0.00
0.01
0.02
0.03
0.04
0.05
0.06BSTMA
0.00
0.01
0.02
0.03
0.04
0.05
0.06
**
AR
mR
NA
mea
n O
.D. (
optic
al d
ensi
ty)
LSV
**
**
*
SEXP SS-24h SS-7d
0.00
0.01
0.02
0.03
0.04
0.05
0.06
**
MPOA
**
**
SS-Imm1EJ-24h
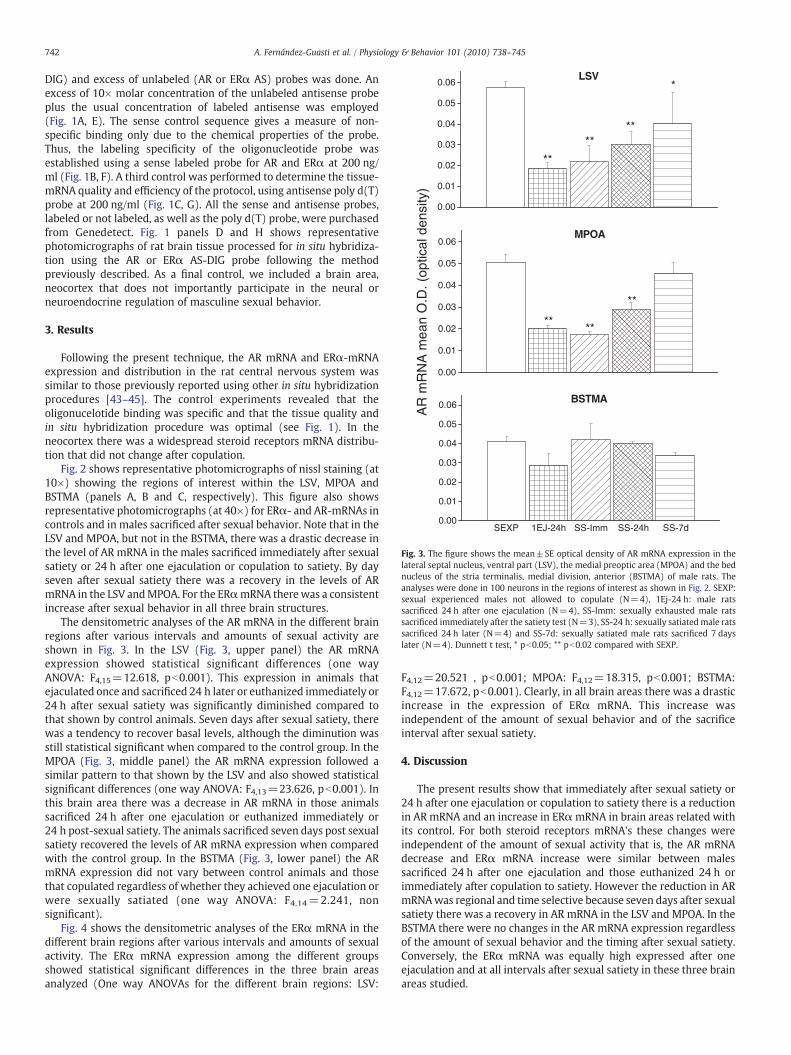
Fig. 3. The figure shows the mean±SE optical density of AR mRNA expression in thelateral septal nucleus, ventral part (LSV), the medial preoptic area (MPOA) and the bednucleus of the stria terminalis, medial division, anterior (BSTMA) of male rats. Theanalyses were done in 100 neurons in the regions of interest as shown in Fig. 2. SEXP:sexual experienced males not allowed to copulate (N=4), 1Ej-24 h: male ratssacrificed 24 h after one ejaculation (N=4), SS-Imm: sexually exhausted male ratssacrificed immediately after the satiety test (N=3), SS-24 h: sexually satiated male ratssacrificed 24 h later (N=4) and SS-7d: sexually satiated male rats sacrificed 7 dayslater (N=4). Dunnett t test, * pb0.05; ** pb0.02 compared with SEXP.
742 A. Fernández-Guasti et al. / Physiology & Behavior 101 (2010) 738–745
DIG) and excess of unlabeled (AR or ERα AS) probes was done. Anexcess of 10× molar concentration of the unlabeled antisense probeplus the usual concentration of labeled antisense was employed(Fig. 1A, E). The sense control sequence gives a measure of non-specific binding only due to the chemical properties of the probe.Thus, the labeling specificity of the oligonucleotide probe wasestablished using a sense labeled probe for AR and ERα at 200 ng/ml (Fig. 1B, F). A third control was performed to determine the tissue-mRNA quality and efficiency of the protocol, using antisense poly d(T)probe at 200 ng/ml (Fig. 1C, G). All the sense and antisense probes,labeled or not labeled, as well as the poly d(T) probe, were purchasedfrom Genedetect. Fig. 1 panels D and H shows representativephotomicrographs of rat brain tissue processed for in situ hybridiza-tion using the AR or ERα AS-DIG probe following the methodpreviously described. As a final control, we included a brain area,neocortex that does not importantly participate in the neural orneuroendocrine regulation of masculine sexual behavior.
3. Results
Following the present technique, the AR mRNA and ERα-mRNAexpression and distribution in the rat central nervous system wassimilar to those previously reported using other in situ hybridizationprocedures [43–45]. The control experiments revealed that theoligonucelotide binding was specific and that the tissue quality andin situ hybridization procedure was optimal (see Fig. 1). In theneocortex there was a widespread steroid receptors mRNA distribu-tion that did not change after copulation.
Fig. 2 shows representative photomicrographs of nissl staining (at10×) showing the regions of interest within the LSV, MPOA andBSTMA (panels A, B and C, respectively). This figure also showsrepresentative photomicrographs (at 40×) for ERα- and AR-mRNAs incontrols and in males sacrificed after sexual behavior. Note that in theLSV and MPOA, but not in the BSTMA, there was a drastic decrease inthe level of AR mRNA in the males sacrificed immediately after sexualsatiety or 24 h after one ejaculation or copulation to satiety. By dayseven after sexual satiety there was a recovery in the levels of ARmRNA in the LSV andMPOA. For the ERαmRNA therewas a consistentincrease after sexual behavior in all three brain structures.
The densitometric analyses of the AR mRNA in the different brainregions after various intervals and amounts of sexual activity areshown in Fig. 3. In the LSV (Fig. 3, upper panel) the AR mRNAexpression showed statistical significant differences (one wayANOVA: F4,15=12.618, pb0.001). This expression in animals thatejaculated once and sacrificed 24 h later or euthanized immediately or24 h after sexual satiety was significantly diminished compared tothat shown by control animals. Seven days after sexual satiety, therewas a tendency to recover basal levels, although the diminution wasstill statistical significant when compared to the control group. In theMPOA (Fig. 3, middle panel) the AR mRNA expression followed asimilar pattern to that shown by the LSV and also showed statisticalsignificant differences (one way ANOVA: F4,13=23.626, pb0.001). Inthis brain area there was a decrease in AR mRNA in those animalssacrificed 24 h after one ejaculation or euthanized immediately or24 h post-sexual satiety. The animals sacrificed seven days post sexualsatiety recovered the levels of AR mRNA expression when comparedwith the control group. In the BSTMA (Fig. 3, lower panel) the ARmRNA expression did not vary between control animals and thosethat copulated regardless of whether they achieved one ejaculation orwere sexually satiated (one way ANOVA: F4,14=2.241, nonsignificant).
Fig. 4 shows the densitometric analyses of the ERα mRNA in thedifferent brain regions after various intervals and amounts of sexualactivity. The ERα mRNA expression among the different groupsshowed statistical significant differences in the three brain areasanalyzed (One way ANOVAs for the different brain regions: LSV:
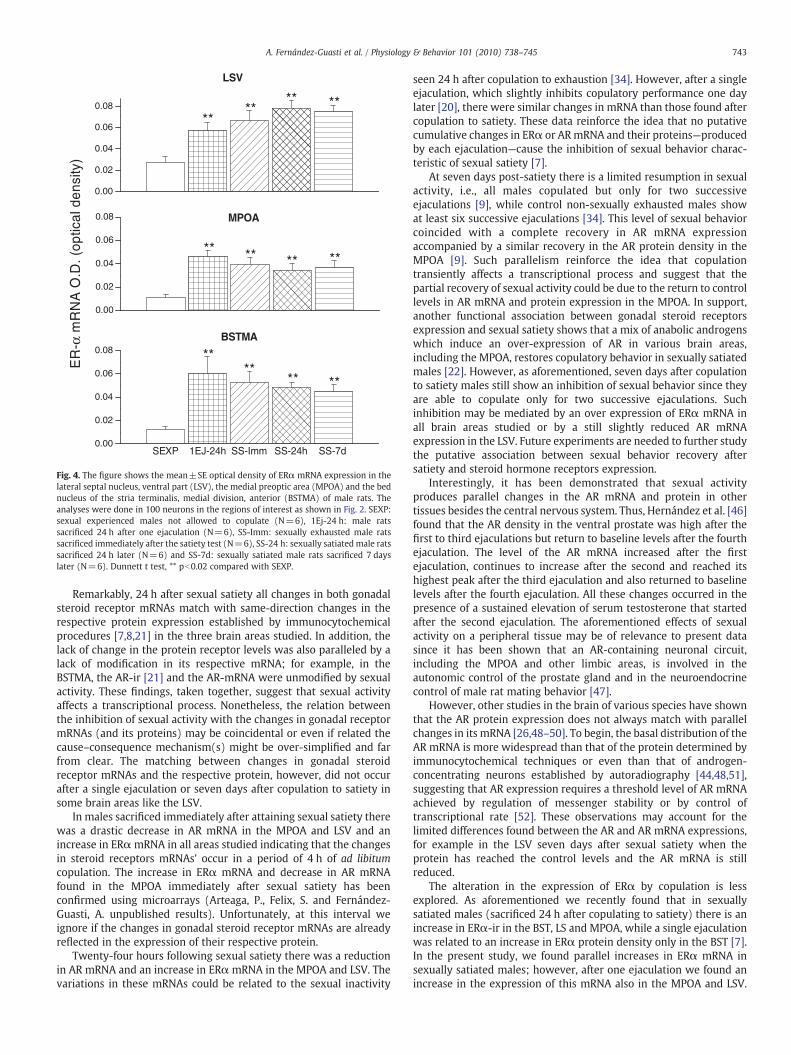
F4,12=20.521 , pb0.001; MPOA: F4,12=18.315, pb0.001; BSTMA:F4,12=17.672, pb0.001). Clearly, in all brain areas there was a drasticincrease in the expression of ERα mRNA. This increase wasindependent of the amount of sexual behavior and of the sacrificeinterval after sexual satiety.
4. Discussion
The present results show that immediately after sexual satiety or24 h after one ejaculation or copulation to satiety there is a reductionin AR mRNA and an increase in ERαmRNA in brain areas related withits control. For both steroid receptors mRNA's these changes wereindependent of the amount of sexual activity that is, the AR mRNAdecrease and ERα mRNA increase were similar between malessacrificed 24 h after one ejaculation and those euthanized 24 h orimmediately after copulation to satiety. However the reduction in ARmRNAwas regional and time selective because seven days after sexualsatiety there was a recovery in AR mRNA in the LSV and MPOA. In theBSTMA there were no changes in the AR mRNA expression regardlessof the amount of sexual behavior and the timing after sexual satiety.Conversely, the ERα mRNA was equally high expressed after oneejaculation and at all intervals after sexual satiety in these three brainareas studied.
0.00
0.02
0.04
0.06
0.08
******
BSTMA
SEXP 1EJ-24h SS-Imm SS-24h SS-7d
0.00
0.02
0.04
0.06
0.08
ER
-α m
RN
A O
.D. (
optic
al d
ensi
ty)
MPOA
0.00
0.02
0.04
0.06
0.08
**
********
******
LSV
**
Fig. 4. The figure shows the mean±SE optical density of ERα mRNA expression in thelateral septal nucleus, ventral part (LSV), the medial preoptic area (MPOA) and the bednucleus of the stria terminalis, medial division, anterior (BSTMA) of male rats. Theanalyses were done in 100 neurons in the regions of interest as shown in Fig. 2. SEXP:sexual experienced males not allowed to copulate (N=6), 1Ej-24 h: male ratssacrificed 24 h after one ejaculation (N=6), SS-Imm: sexually exhausted male ratssacrificed immediately after the satiety test (N=6), SS-24 h: sexually satiated male ratssacrificed 24 h later (N=6) and SS-7d: sexually satiated male rats sacrificed 7 dayslater (N=6). Dunnett t test, ** pb0.02 compared with SEXP.
743A. Fernández-Guasti et al. / Physiology & Behavior 101 (2010) 738–745
Remarkably, 24 h after sexual satiety all changes in both gonadalsteroid receptor mRNAs match with same-direction changes in therespective protein expression established by immunocytochemicalprocedures [7,8,21] in the three brain areas studied. In addition, thelack of change in the protein receptor levels was also paralleled by alack of modification in its respective mRNA; for example, in theBSTMA, the AR-ir [21] and the AR-mRNA were unmodified by sexualactivity. These findings, taken together, suggest that sexual activityaffects a transcriptional process. Nonetheless, the relation betweenthe inhibition of sexual activity with the changes in gonadal receptormRNAs (and its proteins) may be coincidental or even if related thecause–consequence mechanism(s) might be over-simplified and farfrom clear. The matching between changes in gonadal steroidreceptor mRNAs and the respective protein, however, did not occurafter a single ejaculation or seven days after copulation to satiety insome brain areas like the LSV.
In males sacrificed immediately after attaining sexual satiety therewas a drastic decrease in AR mRNA in the MPOA and LSV and anincrease in ERαmRNA in all areas studied indicating that the changesin steroid receptors mRNAs' occur in a period of 4 h of ad libitumcopulation. The increase in ERα mRNA and decrease in AR mRNAfound in the MPOA immediately after sexual satiety has beenconfirmed using microarrays (Arteaga, P., Felix, S. and Fernández-Guasti, A. unpublished results). Unfortunately, at this interval weignore if the changes in gonadal steroid receptor mRNAs are alreadyreflected in the expression of their respective protein.
Twenty-four hours following sexual satiety there was a reductionin AR mRNA and an increase in ERα mRNA in the MPOA and LSV. Thevariations in these mRNAs could be related to the sexual inactivity
seen 24 h after copulation to exhaustion [34]. However, after a singleejaculation, which slightly inhibits copulatory performance one daylater [20], there were similar changes in mRNA than those found aftercopulation to satiety. These data reinforce the idea that no putativecumulative changes in ERα or ARmRNA and their proteins—producedby each ejaculation—cause the inhibition of sexual behavior charac-teristic of sexual satiety [7].
At seven days post-satiety there is a limited resumption in sexualactivity, i.e., all males copulated but only for two successiveejaculations [9], while control non-sexually exhausted males showat least six successive ejaculations [34]. This level of sexual behaviorcoincided with a complete recovery in AR mRNA expressionaccompanied by a similar recovery in the AR protein density in theMPOA [9]. Such parallelism reinforce the idea that copulationtransiently affects a transcriptional process and suggest that thepartial recovery of sexual activity could be due to the return to controllevels in AR mRNA and protein expression in the MPOA. In support,another functional association between gonadal steroid receptorsexpression and sexual satiety shows that a mix of anabolic androgenswhich induce an over-expression of AR in various brain areas,including the MPOA, restores copulatory behavior in sexually satiatedmales [22]. However, as aforementioned, seven days after copulationto satiety males still show an inhibition of sexual behavior since theyare able to copulate only for two successive ejaculations. Suchinhibition may be mediated by an over expression of ERα mRNA inall brain areas studied or by a still slightly reduced AR mRNAexpression in the LSV. Future experiments are needed to further studythe putative association between sexual behavior recovery aftersatiety and steroid hormone receptors expression.
Interestingly, it has been demonstrated that sexual activityproduces parallel changes in the AR mRNA and protein in othertissues besides the central nervous system. Thus, Hernández et al. [46]found that the AR density in the ventral prostate was high after thefirst to third ejaculations but return to baseline levels after the fourthejaculation. The level of the AR mRNA increased after the firstejaculation, continues to increase after the second and reached itshighest peak after the third ejaculation and also returned to baselinelevels after the fourth ejaculation. All these changes occurred in thepresence of a sustained elevation of serum testosterone that startedafter the second ejaculation. The aforementioned effects of sexualactivity on a peripheral tissue may be of relevance to present datasince it has been shown that an AR-containing neuronal circuit,including the MPOA and other limbic areas, is involved in theautonomic control of the prostate gland and in the neuroendocrinecontrol of male rat mating behavior [47].
However, other studies in the brain of various species have shownthat the AR protein expression does not always match with parallelchanges in its mRNA [26,48–50]. To begin, the basal distribution of theAR mRNA is more widespread than that of the protein determined byimmunocytochemical techniques or even than that of androgen-concentrating neurons established by autoradiography [44,48,51],suggesting that AR expression requires a threshold level of AR mRNAachieved by regulation of messenger stability or by control oftranscriptional rate [52]. These observations may account for thelimited differences found between the AR and AR mRNA expressions,for example in the LSV seven days after sexual satiety when theprotein has reached the control levels and the AR mRNA is stillreduced.
The alteration in the expression of ERα by copulation is lessexplored. As aforementioned we recently found that in sexuallysatiated males (sacrificed 24 h after copulating to satiety) there is anincrease in ERα-ir in the BST, LS and MPOA, while a single ejaculationwas related to an increase in ERα protein density only in the BST [7].In the present study, we found parallel increases in ERα mRNA insexually satiated males; however, after one ejaculation we found anincrease in the expression of this mRNA also in the MPOA and LSV.

744 A. Fernández-Guasti et al. / Physiology & Behavior 101 (2010) 738–745
This lack of matching is puzzling andmay be related to the differentialexpression of ERα mRNA and its protein. Even in general the ERαprotein expression pattern often match with its mRNA expressionpattern [29], in certain brain structures, including the MPOA, the ERαprotein levels do not correlate with ERα mRNA expression [29,53].Thus, in the MPOA, but not in the VMH or Arc, estradiol benzoate (EB)does not suppress the expression of ER mRNA while it decreases theER-ir [54], leading to the idea that estrogen differently affects ER geneexpression depending upon the brain region.
The reduction in AR and increase in ERα observed after sexualbehavior does not correspond with changes in the systemic levels oftestosterone or estradiol [7,21] measured at the same interval; forexample, 24 h after sexual satiety there is a drastic reduction in AR-irand mRNA in the MPOA that was unaccompanied by a reduction inserum testosterone levels at this time. This observation suggests that thechanges in steroid receptors immunoreactivity, and possibly also intheir mRNA, is a ligand-independent process. Although ligand-inde-pendent regulation of gonadal steroid receptors is uncommon, growingnew evidence indicates that non-hormonal signals may influence theirexpression. As an example it has been recently demonstrated that socialcues of the environment affect progestin receptors in the rodent brain[55]. Other interpretations, however, may be proposed: for example,that the local levels of steroids may be discretely modified in specificbrain areas—like the MPOA and LS, or even in sub-nuclei within theseregions—in turn producing the specific changes in steroid receptorsmRNAs and proteins. Alternatively, the changes in AR mRNA and ERαmRNA may occur in response to variations in gonadal steroidsconcentrations produced during the execution of repeated copulation,which took place several hours before the observed variations inmRNAs. In this line, it has been reported that the plasma concentrationof testosterone increases immediately after copulation to satiety [56].Furthermore, androgens and estrogens not only regulate AR and ERα,respectively, but also may modify the heterologous receptor. Thus, forexample, estrogens increase the ARmRNA in theMPOA and the AR-ir inthe VMH, but not in the MPOA, of castrated males [26,28] and 5α-dihidrotestosterone synergizes with estradiol to decrease the ERαcontent in the BST, Arc and VMH of ovariectomized females [57]. Thesedata, taken together indicate that the changes in steroid receptors andtheirmRNA inducedby copulation are complexphenomena thatmayberegulated by a variety of processes.
In conclusion, male sexual behavior is associated with changes ingonadal steroid receptors mRNA in the mPOA, LSV and BSTMA. Formost intervals after sexual activity and brain areas studied, thesevariations paralleled with same-direction changes in the respectiveprotein expression [7,21] suggesting that sexual behavior impacts atranscriptional process.
Acknowledgements
Authorswish to thank the “ConsejoNacional de Ciencia y Tecnología,México” for the grant to A.F.-G- number 50433-M and for thefellowships to P.A.-L. and E.A.-C. numbers 050037and 204517, respec-tively. Authors also thank Mr. Víctor Flores and M. Sc. Rebeca ReyesSerrano for technical assistance and Dr. Bryan Phillips-Farfán forlanguage revision.
References
[1] Lee HJ, Chang C. Recent advances in androgen receptor action. Cell Mol Life Sci2003;60:1613–22.
[2] McGinnis MY, Montana RC, Lumia AR. Effects of hydroxyflutamide in the medialpreoptic area or lateral septum on reproductive behaviors in male rats. Brain ResBull 2002;59:227–34.
[3] Xiao L, Jordan CL. Sex differences, laterality, and hormonal regulation of androgenreceptor immunoreactivity in rat hippocampus. Horm Behav 2002;42:327–36.
[4] Baum MJ. Activational and organizational effects of estradiol on male behavioralneuroendocrine function. Scand J Psychol 2003;44:213–20.
[5] Baum MJ, Tobet SA, Starr MS, Bradshaw WG. Implantation of dihydrotestosteronepropionate into the lateral septum or medial amygdala facilitates copulation incastrated male rats given estradiol systemically. Horm Behav 1982;16:208–23.
[6] Greco B, Edwards DA, Michael RP, Clancy AN. Androgen receptors and estrogenreceptors are colocalized in male rat hypothalamic and limbic neurons that expressFos immunoreactivity induced by mating. Neuroendocrinology 1998;67:18–28.
[7] Phillips-Farfan BV, Lemus AE, Fernandez-Guasti A. Increased estrogen receptoralpha immunoreactivity in the forebrain of sexually satiated rats. Horm Behav2007;51:328–34.
[8] Phillips-Farfan BV, Fernandez-Guasti A. Endocrine, neural and pharmacologicalaspects of sexual satiety in male rats. Neurosci Biobehav Rev 2009;33:442–55.
[9] Romano-Torres M, Phillips-Farfan BV, Chavira R, Rodriguez-Manzo G, Fernandez-Guasti A. Relationship between sexual satiety and brain androgen receptors.Neuroendocrinology 2007;85:16–26.
[10] Clancy AN, Zumpe D, Michael RP. Estrogen in the medial preoptic area of male ratsfacilitates copulatory behavior. Horm Behav 2000;38:86–93.
[11] McGinnis MY, Dreifuss RM. Evidence for a role of testosterone-androgen receptorinteractions in mediating masculine sexual behavior in male rats. Endocrinology1989;124:618–26.
[12] Morali G, Hernandez G, Beyer C. Restoration of the copulatory pelvic thrustingpattern in castrated male rats by the intracerebral implantation of androgen.Physiol Behav 1986;36:495–9.
[13] Harding SM, McGinnis MY. Androgen receptor blockade in the MPOA or VMN:effects on male sociosexual behaviors. Physiol Behav 2004;81:671–80.
[14] Beyer C, Morali G, Naftolin F, Larsson K, Perez P. Effect of some antiestrogens andaromatase inhibitors on androgen induced sexual behavior in castrated male rats.Horm Behav 1976;7:353–63.
[15] Lephart ED. A review of brain aromatase cytochrome P450. Brain Res Brain Res Rev1996;22:1–26.
[16] Honda S, Harada N, Ito S, Takagi Y, Maeda S. Disruption of sexual behavior in malearomatase-deficient mice lacking exons 1 and 2 of the cyp19 gene. BiochemBiophys Res Commun 1998;252:445–9.
[17] Ogawa S, Lubahn DB, Korach KS, Pfaff DW. Behavioral effects of estrogen receptorgene disruption in male mice. Proc Natl Acad Sci U S A 1997;94:1476–81.
[18] Tetel MJ, Pfaff DW. Contributions of estrogen receptor-alpha and estrogenreceptor-beta to the regulation of behavior. Biochim Biophys Acta 2010;1800(10):1084–9.
[19] Beach FA, Jordan L. Sexual exhaustion and recovery in themale rat. Q J Exp Psychol1956;8:121–33.
[20] Larsson K. The effects of sexual satiation. In: Elgren J, editor. Conditioning and sexualbehaviour in the male albino rat. Stockholm: Academic Press; 1956. p. 101–16.
[21] Fernandez-Guasti A, Swaab D, Rodriguez-Manzo G. Sexual behavior reduceshypothalamic androgen receptor immunoreactivity. Psychoneuroendocrinology2003;28:501–12.
[22] Phillips-Farfan BV, Romano-Torres M, Fernandez-Guasti A. Anabolic androgensrestore mating after sexual satiety in male rats. Pharmacol Biochem Behav2008;89:241–6.
[23] Jaworski T. Degradation and beyond: control of androgen receptor activity by theproteasome system. Cell Mol Biol Lett 2006;11:109–31.
[24] Gobinet J, Poujol N, Sultan C. Molecular action of androgens. Mol Cell Endocrinol2002;198:15–24.
[25] Kemppainen JA, Lane MV, Sar M, Wilson EM. Androgen receptor phosphorylation,turnover, nuclear transport, and transcriptional activation. Specificity for steroidsand antihormones. J Biol Chem 1992;267:968–74.
[26] Handa RJ, Kerr JE, DonCarlos LL, McGivern RF, Hejna G. Hormonal regulation ofandrogen receptor messenger RNA in the medial preoptic area of the male rat.Brain Res Mol Brain Res 1996;39:57–67.
[27] Menard CS, Harlan RE. Up-regulation of androgen receptor immunoreactivity inthe rat brain by androgenic-anabolic steroids. Brain Res 1993;622:226–36.
[28] Lynch CS, Story AJ. Dihydrotestosterone and estrogen regulation of rat brainandrogen-receptor immunoreactivity. Physiol Behav 2000;69:445–53.
[29] Wilson ME, Westberry JM, Prewitt AK. Dynamic regulation of estrogen receptor-alpha gene expression in the brain: a role for promoter methylation? FrontNeuroendocrinol 2008;29:375–85.
[30] Roselli CE, Resko JA. Aromatase activity in the rat brain: hormonal regulation andsex differences. J Steroid Biochem Mol Biol 1993;44:499–508.
[31] Yamada S, Noguchi D, Ito H, Yamanouchi K. Sex and regional differences indecrease of estrogen receptor alpha-immunoreactive cells by estrogen in rathypothalamus and midbrain. Neurosci Lett 2009;463:135–9.
[32] Shughrue PJ, Bushnell CD, Dorsa DM. Estrogen receptor messenger ribonucleicacid in female rat brain during the estrous cycle: a comparison withovariectomized females and intact males. Endocrinology 1992;131:381–8.
[33] Wu D, Gore AC. Sexual experience changes sex hormones but not hypothalamicsteroid hormone receptor expression in young and middle-aged male rats. HormBehav 2009;56:299–308.
[34] Rodriguez-Manzo G, Fernandez-Guasti A. Reversal of sexual exhaustion byserotonergic and noradrenergic agents. Behav Brain Res 1994;62:127–34.
[35] Paxinos G, Watson C. The rat brain in stereotaxic coordinates. Fuorth ed. SanDiego: Academic Press; 1998.
[36] Iruthayanathan M, Zhou YH, Childs GV. Dehydroepiandrosterone restoration ofgrowth hormone gene expression in aging female rats, in vivo and in vitro:evidence for actions via estrogen receptors. Endocrinology 2005;146:5176–87.
[37] Akhter N, Johnson BW, Crane C, IruthayanathanM, Zhou YH, Kudo A, et al. Anteriorpituitary leptin expression changes in different reproductive states: in vitrostimulation by gonadotropin-releasing hormone. J Histochem Cytochem 2007;55:151–66.

745A. Fernández-Guasti et al. / Physiology & Behavior 101 (2010) 738–745
[38] Sanchez-Lemus E, Benicky J, Pavel J, Saavedra JM. In vivo Angiotensin II AT1receptor blockade selectively inhibits LPS-induced innate immune response andACTH release in rat pituitary gland. Brain Behav Immun 2009;23:945–57.
[39] Schmitz GG, Walter T, Seibl R, Kessler C. Nonradioactive labeling of oligonucleo-tides in vitro with the hapten digoxigenin by tailing with terminal transferase.Anal Biochem 1991;192:222–31.
[40] Holtke HJ, Sagner G, Kessler C, Schmitz G. Sensitive chemiluminescent detection ofdigoxigenin-labeled nucleic acids: a fast and simple protocol and its applications.Biotechniques 1992;12:104–13.
[41] LucasM, Frenois F, Vouillac C, Stinus L, CadorM, LeMoine C. Reactivity and plasticityin the amygdala nuclei during opiate withdrawal conditioning: differentialexpression of c-fos and arc immediate early genes. Neuroscience 2008;154:1021–33.
[42] Koji T, Brenner RM. Localization of estrogen receptor messenger ribonucleic acidin rhesus monkey uterus by nonradioactive in situ hybridization with digox-igenin-labeled oligodeoxynucleotides. Endocrinology 1993;132:382–92.
[43] Burgess LH, Handa RJ. Hormonal regulation of androgen receptor mRNA in thebrain and anterior pituitary gland of the male rat. Brain Res Mol Brain Res1993;19:31–8.
[44] Simerly RB, Chang C, Muramatsu M, Swanson LW. Distribution of androgen andestrogen receptor mRNA-containing cells in the rat brain: an in situ hybridizationstudy. J Comp Neurol 1990;294:76–95.
[45] Shughrue PJ, Lane MV, Merchenthaler I. Comparative distribution of estrogenreceptor-alpha and -beta mRNA in the rat central nervous system. J Comp Neurol1997;388:507–25.
[46] Hernandez ME, Soto-Cid A, Aranda-Abreu GE, Diaz R, Rojas F, Garcia LI, et al. Astudy of the prostate, androgens and sexual activity of male rats. Reprod BiolEndocrinol 2007;5:11.
[47] Huddleston GG, Song CK, Paisley JC, Bartness TJ, Clancy AN. Gonadal steroidreceptors colocalize with central nervous system neurons projecting to the ratprostate gland. Am J Physiol Regul Integr Comp Physiol 2007;292:R2196–205.
[48] Hamson DK, Jones BA, Watson NV. Distribution of androgen receptor immuno-reactivity in the brainstem of male rats. Neuroscience 2004;127:797–803.
[49] Metzdorf R, Gahr M, Fusani L. Distribution of aromatase, estrogen receptor, andandrogen receptor mRNA in the forebrain of songbirds and nonsongbirds. J CompNeurol 1999;407:115–29.
[50] Perez J, Kelley DB. Trophic effects of androgen: receptor expression and thesurvival of laryngeal motor neurons after axotomy. J Neurosci 1996;16:6625–33.
[51] Pfaff DW. Autoradiographic localization of radioactivity in rat brain after injectionof tritiated sex hormones. Science 1968;161:1355–6.
[52] Kelley DB, Tindall DJ. Models systems for the study of androgen-regulated geneexpression in the central nervous system. In: Pfaff DW, Arnold AP, Etgen AM,Fahrbach SE, Rubin RT, editors. Hormones, Brain and Behavior. San Diego:Academic Press; 2002. p. 527–38.
[53] Pasterkamp RJ, Yuri K, Morita N, Kawata M. Differential expression of estrogenreceptor mRNA and protein in the female rat preoptic area. Neurosci Lett 1997;239:81–4.
[54] Orikasa C, Mizuno K, Sakuma Y, Hayashi S. Exogenous estrogen acts differently onproduction of estrogen receptor in the preoptic area and the mediobasalhypothalamic nuclei in the newborn rat. Neurosci Res 1996;25:247–54.
[55] Auger AP. Steroid receptor control of reproductive behavior. Horm Behav 2004;45:168–72.
[56] Bonilla-Jaime H, Vazquez-Palacios G, Arteaga-Silva M, Retana-Marquez S.Hormonal responses to different sexually related conditions in male rats. HormBehav 2006;49:376–82.
[57] Brown TJ, Scherz B, Hochberg RB, MacLusky NJ. Regulation of estrogen receptorconcentrations in the rat brain: effects of sustained androgen and estrogenexposure. Neuroendocrinology 1996;63:53–60.




















