
Synaptotagmin 7 functions as a Ca 2+ -sensor for synaptic vesicle replenishment

Biochemical and Biophysical Research Communications 373 (2008) 509–514
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
The cytoplasmic C2A domain of synaptotagmin shows sequence specificinteraction with its own mRNA
Sunitha S. Sukumaran, Siddharth Banerjee, Salini Bhasker, Anoopkumar Thekkuveettil *
Division of Molecular Medicine, Sree Chitra Tirunal Institute for Medical Sciences and Technology, BMT Wing, Poojapura, Trivandrum, Kerala 695012, India
a r t i c l e i n f o
Article history:Received 5 June 2008Available online 26 June 2008
Keywords:RNA–protein interactionTranslation regulationC2 domainDinucleotide repeats
0006-291X/$ - see front matter � 2008 Elsevier Inc. Adoi:10.1016/j.bbrc.2008.06.063
* Corresponding author. Fax: +91 471 2341814.E-mail address: [email protected] (A. Thekkuve
a b s t r a c t
Synaptotagmin-1 (Syt1) is essential in Ca2+-dependent neurotransmitter release, but its expression reg-ulation is unknown. Here we report that the cytoplasmic Syt1 fragment forms ribonucleoprotein complexby interacting with the 30 untranslated region (3
0UTR) of its own mRNA. Two protein-binding domains,
GU15 repeat and GUCAAUG, within the Syt 30UTR and the C2 domains in Syt1, especially C2A, are essentialin this ribonucleoprotein complex formation. Furthermore, in in vitro assay the translation efficiency ofSyt1 mRNA was downregulated in presence of 30UTR. These results demonstrate for the fist time thatthe soluble fraction of Syt1 can interact with its own mRNA in a highly sequence specific manner.
� 2008 Elsevier Inc. All rights reserved.
Synaptotagmin-1 (Syt1), a presynaptic protein essential in theCa2+-dependent neurotransmitter release pathway, allows the for-mation of a ternary complex of soluble N-ethylmaleimide sensitivefactor attachment receptors (SNAREs), composed of synaptobrevin/VAMP-2, syntaxin and SNAP-25 at the active zone [1,2]. Syt1 hastwo highly conserved C2 domains, which upon binding with Ca2+
undergoes structural changes that enhance its SNARE-bindingaffinity and allows vesicles to fuse with the plasma membrane[3,4]. Both surfeit and deficit levels of Syt1 hamper the vesicle exo-cytosis [5,6]. This led to the premise that Syt1 protein synthesis oc-curs at a constant rate and is functionally moderated bydimerization with various isoforms, particularly with Syt4 havinglow Ca2+ affinity [7]. Constant level of Syt1 transcripts and upreg-ulation of other isoforms under various physiological stimuli sup-ported this notion [8]. Later studies, however, questioned theprospect of Syt1–4 dimerization because both these isoforms arenot co-localized in the neuronal terminals [9]. However, it is notclear whether an alternate molecular mechanism is involved inthe controlled expression of Syt1 in neurons.
A significant reduction in Syt1 protein has been reported in me-sial temporal lobe epilepsy patients [10]. We also observed a sim-ilar reduction in Syt1 protein level in hippocampal neurons ofrodents during status epilepticus while maintaining a constant tran-script level and synaptic vesicle density (S.S. Sukumaran and A.T.,unpublished data). All these conflicting data prompted us to exam-ine alternate molecular pathways involved in the regulation ofSyt1. Here we report that C2A domain of Syt1 has a strong binding
ll rights reserved.
ettil).
affinity for 30untranslated region (30UTR) of its own transcripts. Ourdata also suggest that the two protein-binding domains in the30UTR play a significant role in this ribonucleoprotein (RNP) com-plex formation and translation regulation.
Materials and methods
RNA and protein secondary structure analysis. The secondarystructure of Syt1 mRNA was drawn using CLC RNA Workbench ver-sion 2.1.2. The number of suboptimal structures was limited to 10and isolated base pairs were avoided in the structure prediction.The RNA-binding domains in the Syt1 were identified using BindNsoftware (http://bioinfo.ggc.org/BindN). The conserved aminoacids in Syt1 in Drosophila, rat and human were identified usingClustalW. The Syt1-C2AB structure (MMBID 62377) [11] was ana-lyzed using Cn3D (www.ncbi.nlm.nih.gov).
Recombinant Syt1 protein constructs. Syt1 gene was amplified byRT-PCR from rat brain (Table 1) and cloned (GenBank Accession No.DQ181550). From this clone the Syt1-C2, -C2A, and -C2B regionswere PCR amplified (Table 1) and cloned. GST fusion proteins wereexpressed in pGEX4T1 (Amersham Biosciences, USA) by 1 mM IPTGinduction at 37 �C for 3 h. The recombinant proteins were purifiedusing Glutathione–Sepharose 4B MicroSpin affinity columns.
Brain lysate preparation. Human brain (National Brain Bank,Bangalore) and rat brain hippocampi were homogenized in ice-cold buffer (50 mM Tris–HCl, pH 7.5, 25% glycerol, 50 mM KCl,0.1 M EDTA, and 0.5 mM DTT) containing 1 mM PMSF and 5 lg/ml aprotinin using an electronic homogenizer (Ultra Turrax T8,IKA Labortechnik, Germany) at a maximum speed of 25,000 L/min for 30 s at 4 �C. The homogenization was repeated 3–4 times

Table 1Primers and product sizes of amplified genes
Gene Primer pairs Size (bp)
Syt1 50-TGAACCAAAAATGGTGAGTGC-30 and 50-AAAGGCTTCGTTTTCCCTTTAC-30 1284SytC2 50-TGGATGACGATGCTGAAACC-30 and 50-AAAGGCTTCGTTTTCCCTTTAC-30 929SytC2A 50-TGGATGACGATGCTGAAACC-30 and 50-GGCCAAAATCCACGGTGTTC-30 406SytC2B 50-ATCTCCAGAGCGCTGAGAAAG-30 and 50-AAAGGCTTCGTTTTCCCTTTAC-30 503b-Actin 50-GCCAACCGTGAAAAGATGAC-30 and 50-AGCCACCAATCCACACAGAGTA-30 691Syt 30UTR 50-CATAGCCACAAAACAGAATAGC-30 and 50-CAAACTCTAATGTGCAAACC-30 360CA repeata 50-CAAACTCTAATGTGCAAACC-30 and 50-CATAGCCACAAAACAGAATAGC-30 360
a Reverse Syt 30UTR.
510 S.S. Sukumaran et al. / Biochemical and Biophysical Research Communications 373 (2008) 509–514
with an interval of 1 min. The homogenate was centrifuged for5 min at 1700g and the pellet was discarded. The supernatantwas centrifuged for 1 h at 100,000g at 4 �C. The supernatant (solu-ble protein fraction, SPF) and pellet were stored at �80 �C untilexperimentation. The membrane proteins may release into solu-tion because the high speed homogenization for �2 min could dis-rupt the membrane integrity of organelles.
RNA constructs. Syt 30UTR was PCR amplified from human geno-mic DNA (Table 1) and cloned in pTZ57R/T vector (GenBank Acces-sion No. DQ448032). The SytDB, SytDGT, SytDF, and SytDHepfragments were released from Syt 30UTR by restriction digestionsand subcloned in pTZ57R/T (MBI Fermentas, Canada) or pGEMT(Promega, USA) vectors. The labeled RNA probe from the aboveconstructs as well as b-actin and CA repeat RNA (Table 1) were syn-thesized by in vitro transcription using T7 promoter with[a-32P]UTP (25 lCi) in presence of transcription buffer (containing40 mM Tris–HCl, pH 7.9, 6 mM MgCl2, 10 mM DTT, 10 mM NaCl,and 2 mM Spermidine), 10 mM NTP (ATP, CTP, and GTP), 0.1 ll of20 mg/ml BSA, and 30 U RNase inhibitor by 20 U T7 RNA polymer-ase for 2 h at 37 �C. The reaction was continued for another hourafter adding 2 ll of 0.5 mM UTP. The transcribed RNA were treatedwith RNase free DNase I for 30 min at 37 �C and purified by passingthrough NucleoSpin RNA isolation column (Macherey-Nagel,Germany).
Electrophoretic mobility shift assay (EMSA). The assay was per-formed as described previously [12]. Briefly, recombinant proteinsand SPF (1 lg) were incubated with 5–7 ng [a-32P]-labeled Syt30UTR RNA for 30 min at 37 �C in binding buffer (20 mM Tris–HCl, pH 7.5, 50 mM KCl, 5 mM MgCl2, 1 mM dithiothreitol, 10%glycerol, and 100 lg/ml BSA) to allow RNP formation. The reactionmixture was resolved by 5–7% non-denaturing-PAGE and analyzedusing phosphor imager (FLA-5100, Fujifilm, Japan).
UV cross-linking assay. The assay was carried out as describedearlier [13]. Briefly, the RNP complex was UV cross-linked at254 nm for 20 min (in ice-cold conditions). DTT (100 mM) wasadded and incubated at RT for 5 min in dark. The reaction mixtureswere treated with RNase (100 ng) at 37 �C for 15 min. The sampleswere resolved by 12% SDS–PAGE.
Western blot. UV cross-linked proteins (see above) were re-solved by SDS–PAGE and transferred to a nitrocellulose membraneby wet transfer (Mini Trans-blot, Bio-Rad) at 100 V for 1 h at RT.Immunodetection was performed using ECL Advance WesternBlotting Detection Kit (Amersham Biosciences) according to manu-facturer’s instructions. Polyclonal rabbit Syt1 antibody was used ina dilution of 1:20,000. The goat anti rabbit IgG secondary antibody(HRP-conjugated) was used in a dilution of 1:200,000. The mem-branes were scanned using phosphor imager in chemilumines-cence mode.
Filter-binding assay. The assay was performed as described pre-viously [14]. Briefly, RNA–protein interactions were performedwith GST-C2 (0–160 pmol) and GST-C2A (0–230 pmol) with Syt30UTR and the mixture was passed through a sandwich of nitrocel-lulose membrane (top) and nylon membrane (bottom) using slot
blot manifold under vacuum. The wells were washed twice with1 ml binding buffer (mentioned above). Both the membranes weredried, exposed to imaging plate and scanned using phosphor ima-ger. The band intensities were calculated by densitometric analysisusing MultiGauge software (Fujifilm, Japan).
In vitro translation assay. Syt1 full-length coding region with andwithout 30UTR (Syt+UTR and SytDUTR, respectively) were con-structed in pGEMT vector (Promega, USA). For Syt+UTR, the360 bp Syt 30UTR fragment was ligated to the immediate down-stream of the full-length coding region of Syt1 gene. Circular plas-mids were used in assay for generating RNA transcripts, to avoidimmediate termination of mRNA after the stop codon. The cloneswere subjected to in vitro rabbit reticulocyte lysate transcription-translation in the presence of [35S]methionine (1000 Ci/mmol)according to the instructions mentioned in the kit (TNT Quick Cou-pled Transcription/Translation System, Promega, USA). Equivalentconcentrations of DNA with PCR enhancer were used in each assay.The reaction was performed in a 10 ll volume with 65% of TNT T7Quick Master Mix at 30 �C for 90 min. The synthesized proteinproducts were denatured by incubating at 70 �C for 10 min and re-solved by 12% SDS–PAGE. The gel was then fixed, dried, and ex-posed to imaging plate and scanned using phosphor imager.Band intensities were measured densitometrically using MultiGa-uge Software.
Results and discussion
In an attempt to ascertain regulatory sequences in the humanSyt1 mRNA, we identified two potential protein-binding domainswithin its 30UTR: a GU15 repeat domain [15] and a GUCAAUG ‘hep-tamer’ sequence analogous to the known protein-binding CDEI se-quence [A/G]TCAC[A/G]TG [16]. These regions were part of a singlestem loop structure of RNA having the heptamer sequence at theloop end and (GU)15 at the extended stem structure (Fig. 1A). Be-sides, in silico analysis of RNA-binding region in Syt1 using BindN[17] revealed that there are RNA-binding amino acid residues inC2 domains which are evolutionarily conserved (Fig. 1B). In C2Athese residues are assembled in b3, b4-strands and within the flex-ible linkers flanking b6 strand which all accumulate at the interme-diate junction between C2B (Fig. 1C). On the other hand in C2Bthese residues are distributed on the surface, mainly accumulatingin b2–b5 strands and in the second a-helix (Fig. 1C). We hypothe-sized that Syt1 may be under constant regulation to maintain a sta-ble Syt1 level and there may be a translation regulation pathway toallow this.
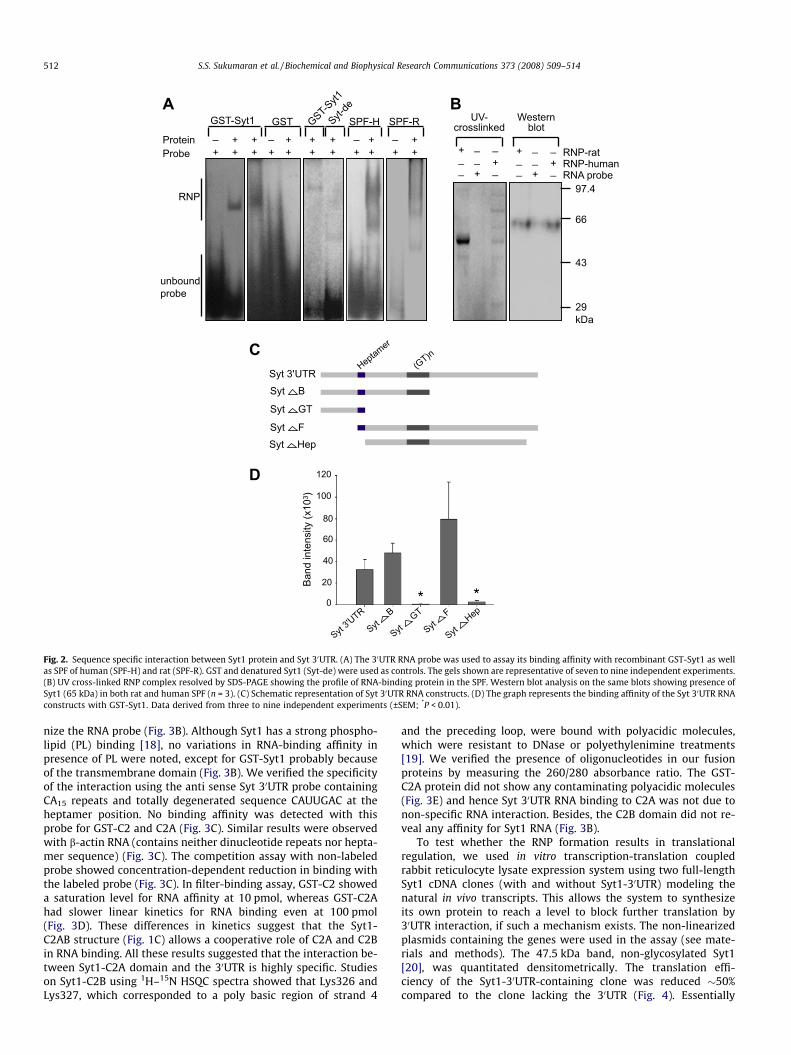
To test this, we in vitro transcribed the 360-bp region from hu-man Syt1 gene covering the GU15 repeats and heptamer sequenceto avail evidences for its discrete binding with proteins. Syt1-glu-tathione S-transferase fusion protein (GST-Syt1) was incubatedwith the RNA probe to allow RNP complex formation and was eval-uated by EMSA. The RNA probe showed distinct binding with theprotein (Fig. 2A). No RNA binding was observed for GST (Fig. 2A);while heat denatured GST-Syt1 (Syt-de) showed an altered band

Fig. 1. Secondary structures of Syt1 mRNA and C2 domains. (A) Syt1 mRNA showing heptamer (GUCAAUG) forming the loop end (shown in red box) and GU repeatsextending the stem structure (shown in green box). The secondary structure DG = �859.6 kcal/mol. (B) RNA-binding amino acid residues in C2A (ash background) and C2B(blue background) domains were identified using BindN. Those residues, which are evolutionarily conserved in Drosophila, rat and human are highlighted in yellow and inmammals alone in green. (C) The structure of Syt1-C2AB, -C2A, and -C2B showing the distribution of the highly conserved RNA-binding regions (from Drosophila to human),highlighted in yellow.
S.S. Sukumaran et al. / Biochemical and Biophysical Research Communications 373 (2008) 509–514 511
shift pattern compared to GST-Syt1 (Fig. 2A) indicating that thestructural integrity of protein may be playing a significant role inRNA interaction. The soluble protein fractions (SPF) of humanand rat hippocampal tissues also showed strong binding affinitywith Syt 30UTR (Fig. 2A); the SPF-RNA complex was UV cross-linked, to estimate the molecular weight of the proteins whichinteracts with the RNA. The results suggested that a selected num-ber of proteins in the SPF recognized the RNA (Fig. 2B); presence ofSyt1 in SPF was confirmed by Western blot analysis (Fig. 2B) (seeMaterials and methods; 100,000g pellet fraction showed a signifi-cant amount of Syt1 retained in the membrane—data not shown).
To further delineate the protein-binding sequence in the 30UTR, wetested four smaller constructs from the Syt 30UTR clone: SytDB,SytDGT, SytDF, and SytDHep (Fig. 2C). The binding affinity ofGST-Syt1 was significantly diminished in those UTR fragmentslacking heptamer or GU repeat sequences (Fig. 2D), suggesting thatboth of these sequences are imperative for the interaction.
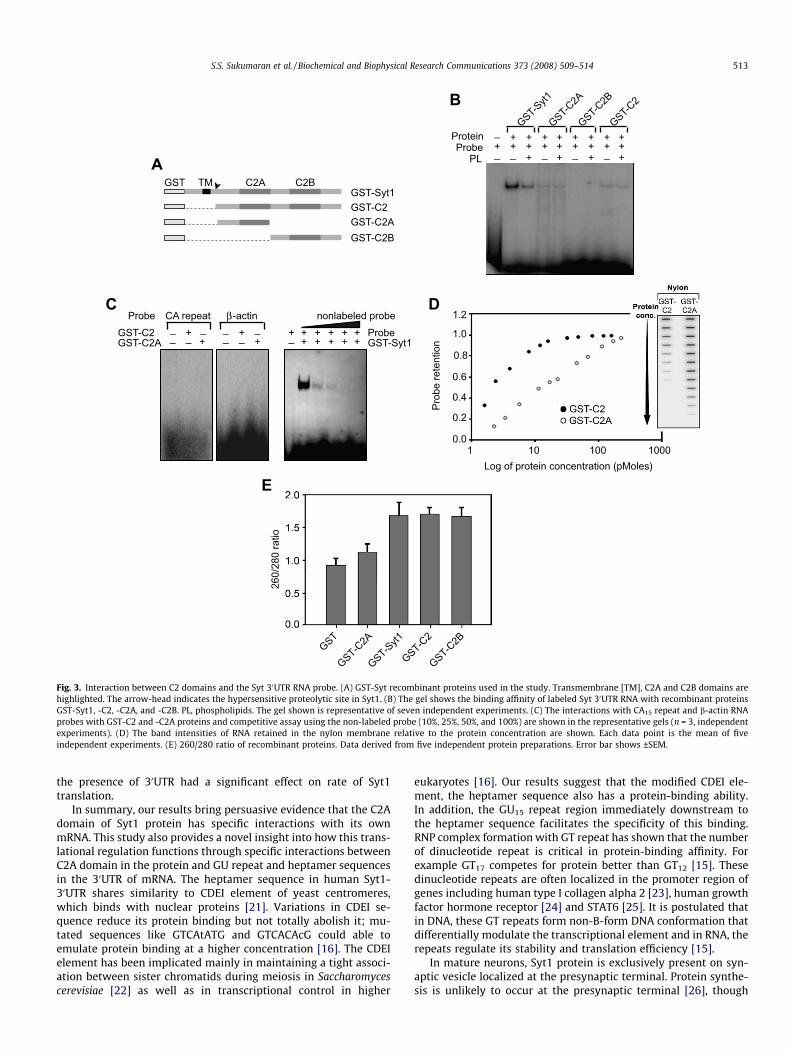
To analyze the function of the Syt1-C2A and -C2B domains inthis RNA interaction, three recombinant fusion proteins were con-structed: GST-C2 (containing both C2 domains), GST-C2A, and GST-C2B (Fig. 3A). Proteins containing the C2A domain (GST-Syt1, -C2,and -C2A) actuated RNP formation, while GST-C2B did not recog-
Syt 3'U
TR
SytB
SytGT
SytF
SytHep
Syt 3'UTR
Syt B
Syt GT
Syt F
Syt Hep
Heptamer
(GT)n
0
20
40
60
80
100
120
Ban
d in
tens
ity (
x103
)
GST-Syt1 GST Syt-de
SPF-H SPF-RGST-Syt1
+ + + + + + + + + + ++ + + + + ++_ _ _ _Protein
Probe
RNP
unboundprobe
+
++
__ _
_ _
_
+
++
__ _
_ _
_
UV-crosslinked
Westernblot
RNP-ratRNP-humanRNA probe
29kDa
43
66
97.4
A B
C
D
Fig. 2. Sequence specific interaction between Syt1 protein and Syt 30UTR. (A) The 30UTR RNA probe was used to assay its binding affinity with recombinant GST-Syt1 as wellas SPF of human (SPF-H) and rat (SPF-R). GST and denatured Syt1 (Syt-de) were used as controls. The gels shown are representative of seven to nine independent experiments.(B) UV cross-linked RNP complex resolved by SDS-PAGE showing the profile of RNA-binding protein in the SPF. Western blot analysis on the same blots showing presence ofSyt1 (65 kDa) in both rat and human SPF (n = 3). (C) Schematic representation of Syt 30UTR RNA constructs. (D) The graph represents the binding affinity of the Syt 30UTR RNAconstructs with GST-Syt1. Data derived from three to nine independent experiments (±SEM; *P < 0.01).
512 S.S. Sukumaran et al. / Biochemical and Biophysical Research Communications 373 (2008) 509–514
nize the RNA probe (Fig. 3B). Although Syt1 has a strong phospho-lipid (PL) binding [18], no variations in RNA-binding affinity inpresence of PL were noted, except for GST-Syt1 probably becauseof the transmembrane domain (Fig. 3B). We verified the specificityof the interaction using the anti sense Syt 30UTR probe containingCA15 repeats and totally degenerated sequence CAUUGAC at theheptamer position. No binding affinity was detected with thisprobe for GST-C2 and C2A (Fig. 3C). Similar results were observedwith b-actin RNA (contains neither dinucleotide repeats nor hepta-mer sequence) (Fig. 3C). The competition assay with non-labeledprobe showed concentration-dependent reduction in binding withthe labeled probe (Fig. 3C). In filter-binding assay, GST-C2 showeda saturation level for RNA affinity at 10 pmol, whereas GST-C2Ahad slower linear kinetics for RNA binding even at 100 pmol(Fig. 3D). These differences in kinetics suggest that the Syt1-C2AB structure (Fig. 1C) allows a cooperative role of C2A and C2Bin RNA binding. All these results suggested that the interaction be-tween Syt1-C2A domain and the 30UTR is highly specific. Studieson Syt1-C2B using 1H–15N HSQC spectra showed that Lys326 andLys327, which corresponded to a poly basic region of strand 4
and the preceding loop, were bound with polyacidic molecules,which were resistant to DNase or polyethylenimine treatments[19]. We verified the presence of oligonucleotides in our fusionproteins by measuring the 260/280 absorbance ratio. The GST-C2A protein did not show any contaminating polyacidic molecules(Fig. 3E) and hence Syt 30UTR RNA binding to C2A was not due tonon-specific RNA interaction. Besides, the C2B domain did not re-veal any affinity for Syt1 RNA (Fig. 3B).
To test whether the RNP formation results in translationalregulation, we used in vitro transcription-translation coupledrabbit reticulocyte lysate expression system using two full-lengthSyt1 cDNA clones (with and without Syt1-30UTR) modeling thenatural in vivo transcripts. This allows the system to synthesizeits own protein to reach a level to block further translation by30UTR interaction, if such a mechanism exists. The non-linearizedplasmids containing the genes were used in the assay (see mate-rials and methods). The 47.5 kDa band, non-glycosylated Syt1[20], was quantitated densitometrically. The translation effi-ciency of the Syt1-30UTR-containing clone was reduced �50%compared to the clone lacking the 30UTR (Fig. 4). Essentially
++ + + + + + +++++++++
+ + + +_ _ _ __
_+
GST TM C2A C2BGST-Syt1
GST-C2
GST-C2A
GST-C2B
GST-C2B
GST-Syt1
GST-C2A
GST-C2
ProteinProbe
PL
++
++__
___
___
CA repeat β-actin
GST-C2GST-C2A + + + + +
+ + + + ++_ GST-Syt1Probe
nonlabeled probe
1 10 100 10000.0
0.2
0.4
0.6
0.8
1.0
1.2
Log of protein concentration (pMoles)
Pro
be r
eten
tion
Probe
A
C D
B
GST-Syt1
GST-C2A
GST-C2B
GST-C2
GST
260/
280
ratio
E
Fig. 3. Interaction between C2 domains and the Syt 30UTR RNA probe. (A) GST-Syt recombinant proteins used in the study. Transmembrane [TM], C2A and C2B domains arehighlighted. The arrow-head indicates the hypersensitive proteolytic site in Syt1. (B) The gel shows the binding affinity of labeled Syt 30UTR RNA with recombinant proteinsGST-Syt1, -C2, -C2A, and -C2B. PL, phospholipids. The gel shown is representative of seven independent experiments. (C) The interactions with CA15 repeat and b-actin RNAprobes with GST-C2 and -C2A proteins and competitive assay using the non-labeled probe (10%, 25%, 50%, and 100%) are shown in the representative gels (n = 3, independentexperiments). (D) The band intensities of RNA retained in the nylon membrane relative to the protein concentration are shown. Each data point is the mean of fiveindependent experiments. (E) 260/280 ratio of recombinant proteins. Data derived from five independent protein preparations. Error bar shows ±SEM.
S.S. Sukumaran et al. / Biochemical and Biophysical Research Communications 373 (2008) 509–514 513
the presence of 30UTR had a significant effect on rate of Syt1translation.
In summary, our results bring persuasive evidence that the C2Adomain of Syt1 protein has specific interactions with its ownmRNA. This study also provides a novel insight into how this trans-lational regulation functions through specific interactions betweenC2A domain in the protein and GU repeat and heptamer sequencesin the 30UTR of mRNA. The heptamer sequence in human Syt1-30UTR shares similarity to CDEI element of yeast centromeres,which binds with nuclear proteins [21]. Variations in CDEI se-quence reduce its protein binding but not totally abolish it; mu-tated sequences like GTCAtATG and GTCACAcG could able toemulate protein binding at a higher concentration [16]. The CDEIelement has been implicated mainly in maintaining a tight associ-ation between sister chromatids during meiosis in Saccharomycescerevisiae [22] as well as in transcriptional control in higher
eukaryotes [16]. Our results suggest that the modified CDEI ele-ment, the heptamer sequence also has a protein-binding ability.In addition, the GU15 repeat region immediately downstream tothe heptamer sequence facilitates the specificity of this binding.RNP complex formation with GT repeat has shown that the numberof dinucleotide repeat is critical in protein-binding affinity. Forexample GT17 competes for protein better than GT12 [15]. Thesedinucleotide repeats are often localized in the promoter region ofgenes including human type I collagen alpha 2 [23], human growthfactor hormone receptor [24] and STAT6 [25]. It is postulated thatin DNA, these GT repeats form non-B-form DNA conformation thatdifferentially modulate the transcriptional element and in RNA, therepeats regulate its stability and translation efficiency [15].
In mature neurons, Syt1 protein is exclusively present on syn-aptic vesicle localized at the presynaptic terminal. Protein synthe-sis is unlikely to occur at the presynaptic terminal [26], though

Contro
l
Syt+3'U
TR
Syt3'U
TR0
500
1000
1500
2000
Contro
l
Syt+3'U
TR
Syt3'U
TRS
yt1
prot
ein
expr
essi
on
Fig. 4. Syt1 gene constructs with and without the 360-bp 30UTR were in vitrotranscribed and translated. Reactions lacking DNA served as negative control.SytDUTR, Syt1 gene construct without Syt1-30UTR; Syt+UTR, Syt1 gene constructwith the Syt1-30UTR. The graph represents values from three independentexperiments (±SEM; *P < 0.008).
514 S.S. Sukumaran et al. / Biochemical and Biophysical Research Communications 373 (2008) 509–514
there are reports suggesting possible transcription machinery inthe axons [27]. Syt1 is believed to be continuously synthesizedfrom mRNA localized at the cell body and transported directly fromthe trans-Golgi network to specialized axonal endosomes fromwhich synaptic vesicles originate [28]. A soluble p40 fragment ofSyt1, corresponding to the extravesicular portion of p65 Syt1, hasbeen detected during cellular stress [29]. This form of Syt1 is trans-lated by alternate AUG transcription initiation site at Met113 [30].Syt1 also has a protease hypersensitive site (see Fig. 3A) immedi-ately after its transmembrane domain, which on proteolysis re-leases soluble p39 Syt1 [20]. Both p39 and p40 Syts (havingsimilar structure to our GST-C2) were found to be stable [30] anddistributed both in cell body and axons similar to the splice variantof Syt6 lacking the transmembrane domain [31]. To our knowledgethis is the first report on Syt1-C2 domains forming RNP complexwith its own mRNA through a sequence specific interaction. It ishoped that this work will facilitate further understanding on themechanism of this selective interaction and its functional role.
Acknowledgments
S.S.S. and Si.B were supported by Council of Scientific andIndustrial Research and University Grants Commission, respec-tively. A.T. acknowledges financial support by Department of Sci-ence and Technology.
References
[1] R.C. Lin, R.H. Scheller, Mechanisms of synaptic vesicle exocytosis, Annu. Rev.Cell Dev. Biol. 16 (2000) 19–49.
[2] T.C. Sudhof, The synaptic vesicle cycle, Annu. Rev. Neurosci. 27 (2004) 509–547.
[3] B.A. Davletov, T.C. Sudhof, Ca(2+)-dependent conformational change insynaptotagmin I, J. Biol. Chem. 269 (1994) 28547–28550.
[4] J. Tang, A. Maximov, O.H. Shin, H. Dai, J. Rizo, T.C. Südhof, A complexin/synaptotagmin 1 switch controls fast synaptic vesicle exocytosis, Cell 6 (2006)1175–1187.
[5] M. Geppert, Y. Goda, R.E. Hammer, C. Li, T.W. Rosahl, C.F. Stevens, T.C. Sudhof,Synaptotagmin I: a major Ca2+ sensor for transmitter release at a centralsynapse, Cell 79 (1994) 717–727.
[6] K.C. Martin, Y. Hu, B.A. Armitage, S.A. Siegelbaum, E.R. Kandel, B.K. Kaang,Evidence for synaptotagmin as an inhibitory clamp on synaptic vesicle releasein aplysia neurons, Proc. Natl. Acad. Sci. USA 92 (1995) 11307–11311.
[7] J.T. Littleton, T.L. Serano, G.M. Rubin, B. Ganetzky, E.R. Chapman, Synapticfunction modulated by changes in the ratio of synaptotagmin I and IV, Nature400 (1999) 757–760.
[8] L. Vician, I.K. Lim, G. Ferguson, G. Tocco, M. Baudry, H.R. Herschman,Synaptotagmin IV is an immediate early gene induced by depolarization inPC12 cells and in brain, Proc. Natl. Acad. Sci. USA 92 (1995) 2164–2168.
[9] B. Adolfsen, S. Saraswati, M. Yoshihara, J.T. Littleton, Synaptotagmins aretrafficked to distinct subcellular domains including the postsynapticcompartment, J. Cell Biol. 2 (2004) 249–260.
[10] J.W. Yang, T. Czech, M. Felizardo, C. Baumgartner, G. Lubec, Aberrantexpression of cytoskeleton proteins in hippocampus from patients withmesial temporal lobe epilepsy, Amino Acids 4 (2006) 477–493.
[11] K.L. Fuson, M. Montes, J.J. Robert, R.B. Sutton, Structure of humansynaptotagmin 1 C2AB in the absence of Ca2+ reveals a novel domainassociation, Biochemistry 46 (2007) 13041–13048.
[12] D.R. Setzer, Measuring equilibrium and kinetic constants using gel retardationassays, Methods Mol. Biol. 118 (1999) 115–128.
[13] M.M. Hanna, L. Bentsen, M. Lucido, A. Sapre, RNA–protein cross linking withphotoreactive nucleotide analogs, Methods Mol. Biol. 118 (1999) 21–33.
[14] K.B. Hall, J.K. Kranz, Nitrocellulose filter binding for determination ofdissociation constants, Methods Mol. Biol. 118 (1999) 105–114.
[15] P.S. Gao, N.M. Heller, W. Walker, C.H. Chen, M. Moller, B. Plunkett, M.H.Roberts, R.P. Schleimer, J.M. Hopkin, S.K. Huang, Variation in dinucleotide (GT)repeat sequence in the first exon of the STAT6 gene is associated with atopicasthma and differentially regulates the promoter activity in vitro, J. Med.Genet. 41 (2004) 535–539.
[16] A. Blangy, P. Leopold, F. Vidal, M. Rassoulzadegan, F. Cuzin, Recognition of theCDEI motif GTCACATG by mouse nuclear proteins and interference with theearly development of the mouse embryo, Nucleic Acids Res. 19 (1991) 7243–7250.
[17] L. Wang, S.J. Brown, BindN: a web-based tool for efficient prediction of DNAand RNA binding sites in amino acid sequences, Nucleic Acids Res. 34 (2006)W243–W248.
[18] R.B. Sutton, B.A. Davletov, A.M. Berghuis, T.C. Sudhof, S.R. Sprang, Structure ofthe first C2 domain of synaptotagmin I: a novel Ca2+/phospholipid-bindingfold, Cell 80 (1995) 929–938.
[19] J. Ubach, Y. Lao, I. Fernandez, D. Arac, T.C. Südhof, J. Rizo, The C2B domain ofsynaptotagmin I is a Ca2+-binding module, Biochemistry 40 (2001) 5854–5860.
[20] M.S. Perin, P.A. Johnston, T. Ozcelik, R. Jahn, U. Francke, T.C. Sudhof, Structuraland functional conservation of synaptotagmin (p65) in Drosophila andhumans, J. Biol. Chem. 266 (1991) 615–622.
[21] F. Vidal, A. Blangy, M. Rassoulzadegan, F. Cuzin, A murine sequence-specificDNA binding protein shows extensive local similarities to the amyloidprecursor protein, Biochem. Biophys. Res. Commun. 189 (1992) 1336–1341.
[22] K. Bloom, A. Hill, M. Kenna, M. Saunders, The structure of a primitivekinetochore, Trends Biochem. Sci. 14 (1989) 223–227.
[23] J. Akai, A. Kimura, R.I. Hata, Transcriptional regulation of the human type Icollagen alpha2 (COL1A2) gene by the combination of two dinucleotiderepeats, Gene 239 (1999) 65–73.
[24] S. Hadjiyannakis, H. Zheng, G.N. Hendy, C.G. Goodyer, GT repeat polymorphismin the 50 flanking region of the human growth hormone receptor gene, Mol.Cell Probes 15 (2001) 239–242.
[25] K. Tamura, H. Arakawa, M. Suzuki, Y. Kobayashi, H. Mochizuki, M. Kato, K.Tokuyama, A. Morikawa, Novel dinucleotide repeat polymorphism in the firstexon of the STAT-6 gene is associated with allergic diseases, Clin. Exp. Allergy31 (2001) 1509–1514.
[26] N. Hirokawa, mRNA transport in dendrites: RNA granules, motors, and tracks,J. Neurosci. 26 (2006) 7139–7142.
[27] E. Koenig, A. Giuditta, Protein-synthesizing machinery in the axoncompartment, Neuroscience 89 (1999) 5–15.
[28] O. Mundigl, M. Matteoli, L. Daniell, A. Thomas-Reetz, A. Metcalf, R. Jahn, P. DeCamilli, Synaptic vesicle proteins and early endosomes in culturedhippocampal neurons: differential effects of Brefeldin A in axon anddendrites, J. Cell Biol. 6 (1993) 1207–1221.
[29] T.M. LaVallee, F. Tarantini, S. Gamble, C.M. Carreira, A. Jackson, T. Maciag,Synaptotagmin-1 is required for fibroblast growth factor-1 release, J. Biol.Chem. 273 (1998) 22217–22223.
[30] C. Bagalá, V. Kolev, A. Mandinova, R. Soldi, C. Mouta, I. Graziani, I. Prudovsky, T.Maciag, The alternative translation of synaptotagmin 1 mediates the non-classical release of FGF1, Biochem. Biophys. Res. Commun. 310 (2003) 1041–1047.
[31] M. Fukuda, K. Mikoshiba, A novel alternatively spliced variant ofsynaptotagmin VI lacking a transmembrane domain. Implications for distinctfunctions of the two isoforms, J. Biol. Chem. 274 (1999) 31428–31434.
Copyright © 2022 FDOKUMEN




















