
Cytotoxic Helix-Rich Oligomer Formation by Melittin and Pancreatic Polypeptide

doi:10.1016/j.jmb.2006.05.007 J. Mol. Biol. (2006) 360, 407–509
The Component Polypeptide Chains of Bovine InsulinNucleate or Inhibit Aggregation of the Parent Proteinin a Conformation-dependent Manner
Glyn L. Devlin1,2⁎, Tuomas P. J. Knowles1,3, Adam Squires1
Margaret G. McCammon2, Sally L. Gras1, Melanie R. Nilsson4
Carol V. Robinson2, Christopher M. Dobson2 and Cait E. MacPhee1
1Cavendish Laboratory,University of Cambridge,JJ Thomson Avenue,Cambridge CB3 0HE, UK2Department of Chemistry,University of Cambridge,Lensfield Road,Cambridge CB2 1EW, UK3University of Cambridge,Nanoscience Centre,11 JJ Thomson Avenue,Cambridge CB3 0FF, UK4Department of Chemistry,McDaniel College, Eaton Hall,2 College Hill, Westminster,MD 21157, USA
Present address: A. Squires, SchooUniversity of Reading, WhiteknightsUK.Abbreviations used: AFM, atomic
nano-ESI MS, nano-electrospray ionspectrometry; TEM, transmission eleE-mail address of the correspondi
0022-2836/$ - see front matter © 2006 E
Amyloid fibrils are typically rigid, unbranched structures with diameters of∼10 nm and lengths up to several micrometres, and are associated withmore than 20 diseases including Alzheimer’s disease and type II diabetes.Insulin is a small, predominantly α-helical protein consisting of 51 residuesin two disulfide-linked polypeptide chains that readily assembles intoamyloid fibrils under conditions of low pH and elevated temperature. Wedemonstrate here that both the A-chain and the B-chain of insulin arecapable of forming amyloid fibrils in isolation under similar conditions,with fibrillar morphologies that differ from those composed of intactinsulin. Both the A-chain and B-chain fibrils were found to be able to cross-seed the fibrillization of the parent protein, although these reactions weresubstantially less efficient than self-seeding with fibrils composed of full-length insulin. In both cases, the cross-seeded fibrils were morphologicallydistinct from the seeding material, but shared common characteristics withtypical insulin fibrils, including a very similar helical repeat. The broaderdistribution of heights of the cross-seeded fibrils compared to typical insulinfibrils, however, indicates that their underlying protofilament hierarchymay be subtly different. In addition, and remarkably in view of this seedingbehavior, the soluble forms of the A-chain and B-chain peptides were foundto be capable of inhibiting insulin fibril formation. Studies using massspectrometry suggest that this behavior might be attributable to complexformation between insulin and the A-chain and B-chain peptides. Thefinding that the same chemical form of a polypeptide chain in differentphysical states can either stimulate or inhibit the conversion of a protein intoamyloid fibrils sheds new light on the mechanisms underlying fibrilformation, fibril strain propagation and amyloid disease initiation andprogression.
© 2006 Elsevier Ltd. All rights reserved.
*Corresponding author
Keywords: amyloid; fibrils; insulin; protein aggregation; β-sheet breakerl of Chemistry, The, Reading, RG6 6AD,
force microscopy;ization massctron microscopy.ng author:
lsevier Ltd. All rights reserve
Introduction
Insulin is a small, predominantly α-helicalprotein consisting of 51 residues in two polypep-tide chains (see Figure 1), which readily aggregatesto form amyloid fibrils at low pH and hightemperature.1 Similar aggregates of insulin havebeen found at the site of injection in a diabeticpatient and in insulin infusion pumps.2–4 Amyloidfibrils are typically rigid, unbranched structureswith diameters ranging from 8 nm to 13 nm andwith lengths of up to several micrometres. They are
d.

Figure 1. Ribbon representation of the native bovineinsulin monomer structure. The coordinates are from thecrystal structure of the T6 hexameric crystal structure(PDB file 2A3G). The A-chain is shown in blue, the B-chainis shown in red.
498 Self-sequence Modulation of Insulin Aggregation
typified by a cross-β conformation in which thecomponent β-strands lie perpendicular to the fibrilaxis,5,6 and by binding of the dyes Congo red andthioflavin-T.7 The fibrillar assembly of polypeptidesis implicated in some 20 disease states,8 althoughthe formation of indistinguishable fibrils by non-disease-related proteins and peptides infers thatfibril formation represents a generic property ofpolypeptide chains.9 Insulin provides an interestingsystem for the study of fibrillar aggregation, as itsstructure is substantially constrained by twodisulfide bridges that crosslink the A-chain and B-chain, and a third disulfide bridge within the A-chain, all of which are retained in the fibrillarstate.10 It follows that these constraints limit theconformational space that can be accessed by theprotein and, as such, limit the number of con-formations available to fibril precursors and possi-ble protein arrangements within the fibrils.11Under zinc-free conditions, native bovine insulin
exists as dimers and tetramers,12 and the physio-logically predominant form of insulin is a zinc-coordinated hexamer formed by the association ofthree dimers.13 It is considered that the fibrilprecursor is an insulin monomer, as conditionsthat promote monomer formation, such as low pHand elevated temperature, also induce fibril ass-embly.10 The structure of the insulin monomerunder such conditions has been shown by NMR tobe partially unfolded and to resemble a proteinfolding intermediate,14 consistent with evidencefrom other polypeptide systems demonstratingthat disruption of native contacts precedes fibri-llization.8,15–17 In the insulin molecule, these disrup-tions occur predominantly in the N-terminalsegments of the A and B-chains, and the C-terminalsegment of the B-chain, all of which become
partially or fully disordered.14 The disordering ofthe C-terminal segment of the B-chain is ofparticular interest, as it is these residues that makeintermolecular contacts in the native dimer struc-ture. Furthermore, the displacement of this regionfrom its native structure is likely to expose non-polargroups normally in the helical core of the protein,and hence could provide sites to initiate aggrega-tion. Mutagenesis studies on human insulin havedemonstrated that, of these exposed residues,B10His, B17Leu and A8His in particular appear toplay vital roles in fibril nucleation and growth,18suggesting that residues of both the B-chain and theA-chain are involved in formation of the cross-βfibril core.Several studies have suggested that formation of
insulin fibrils is preceded by the assembly of thepartially folded insulin monomers into relativelysmall oligomers.19–21 Analysis of the early stages ofinsulin fibrillization by nano-electrospray ionizationmass spectrometry (nano-ESI MS) has demonstratedthe presence of oligomers containing up to 12molecules, and similar observations have beenattained by dynamic light-scattering experiments.21Such oligomers have been observed by transmissionelectron microscopy (TEM) and atomic force mi-croscopy (AFM) as small, amorphous clusters, andhave been shown to contain predominantly native-like structure.19,22 It is possible that the insulinmolecules within these oligomers undergo substan-tial rearrangement into amyloid fibrils, as has beenobserved recently for acylphosphatase,23 or that theoligomers dissociate and the fibrils form subse-quently via a separate and independent process.Whatever the nature of these structural changes, theresultant insulin fibrils are morphologically distinctfrom these early, amorphous clusters, and consist ofbetween two and six protofilaments intertwinedinto a braided fibrillar structure. A model for themolecular packing of the insulin molecule within thefibril based on cryo-electron microscopy imagereconstruction has been proposed.11 This modelsuggests that the majority of the insulin molecule,including large sequence segments of both the A andB-chains, is incorporated into the cross-β fibril core,a conclusion supported by the mutagenesis studydiscussed above that implicates residues of both Aand B-chains in the aggregation process.18In the present investigation, we have examined
the interaction of bovine insulin with its A and B-chain peptides, in both their fibrillar and solubleconformations. We demonstrate that amyloid fibrilsformed by the A-chain are capable of seeding insulinfibril formation more efficiently than B-chain fibrils.In both cases, the cross-seeded fibrils were morpho-logically distinct from the seeding material, butshared similarities to typical insulin fibrils, in thattheir helical repeats were almost identical. Thebroader distribution of heights of the cross-seededfibrils compared to typical insulin fibrils, however,indicates that their underlying protofilament hier-archy may be subtly different. Furthermore, weshow that the soluble forms of the A and B-chain
499Self-sequence Modulation of Insulin Aggregation
peptides are capable of inhibiting insulin fibrilformation, probably by forming complexes withthe insulin molecules, a result of considerableinterest in the context of the design of inhibitors ofamyloid aggregation.
Results
Characterisation of amyloid fibrils formed fromthe insulin A-chain and B-chain
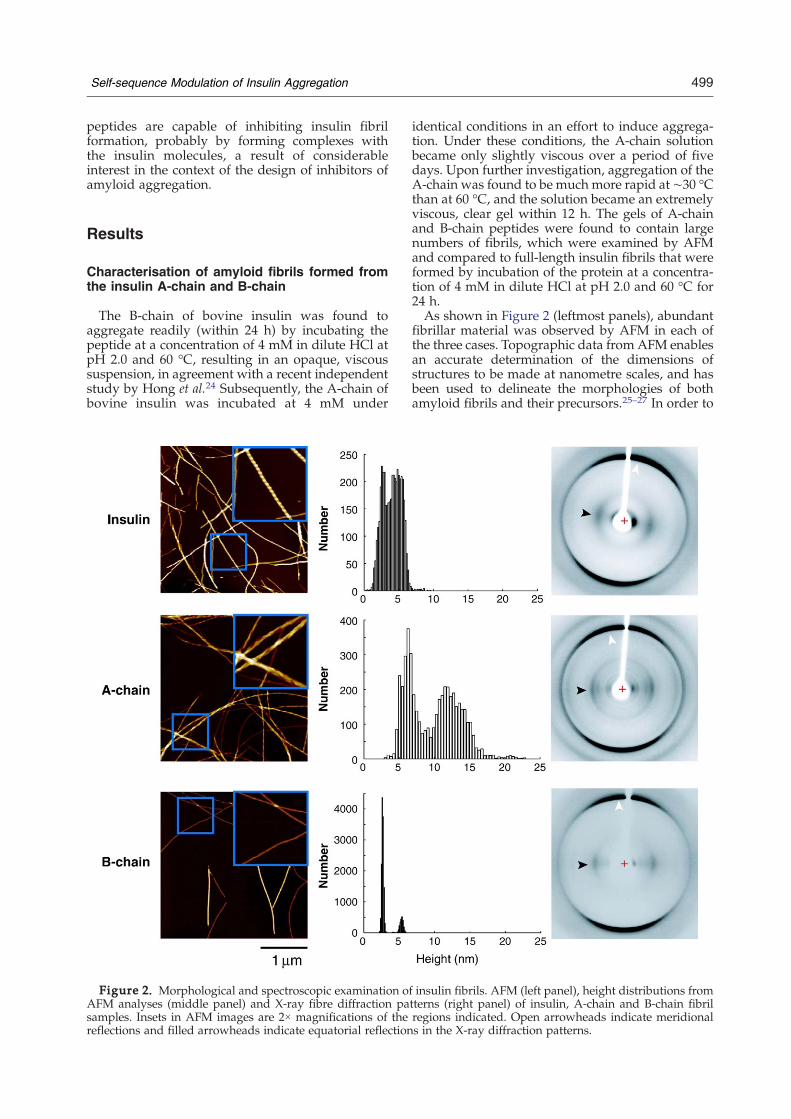
The B-chain of bovine insulin was found toaggregate readily (within 24 h) by incubating thepeptide at a concentration of 4 mM in dilute HCl atpH 2.0 and 60 °C, resulting in an opaque, viscoussuspension, in agreement with a recent independentstudy by Hong et al.24 Subsequently, the A-chain ofbovine insulin was incubated at 4 mM under
Figure 2. Morphological and spectroscopic examination oAFM analyses (middle panel) and X-ray fibre diffraction pasamples. Insets in AFM images are 2× magnifications of thereflections and filled arrowheads indicate equatorial reflection
identical conditions in an effort to induce aggrega-tion. Under these conditions, the A-chain solutionbecame only slightly viscous over a period of fivedays. Upon further investigation, aggregation of theA-chain was found to be much more rapid at ∼30 °Cthan at 60 °C, and the solution became an extremelyviscous, clear gel within 12 h. The gels of A-chainand B-chain peptides were found to contain largenumbers of fibrils, which were examined by AFMand compared to full-length insulin fibrils that wereformed by incubation of the protein at a concentra-tion of 4 mM in dilute HCl at pH 2.0 and 60 °C for24 h.As shown in Figure 2 (leftmost panels), abundant
fibrillar material was observed by AFM in each ofthe three cases. Topographic data from AFM enablesan accurate determination of the dimensions ofstructures to be made at nanometre scales, and hasbeen used to delineate the morphologies of bothamyloid fibrils and their precursors.25–27 In order to
f insulin fibrils. AFM (left panel), height distributions fromtterns (right panel) of insulin, A-chain and B-chain fibrilregions indicated. Open arrowheads indicate meridionals in the X-ray diffraction patterns.

Figure 3. Nucleation of insulin fibril formation.Insulin was incubated at a final concentration of0.44 mM at pH 2.0 and 60 °C in the absence (opensquares) and in the presence of various preformed fibrils:5% (w/w) insulin (circles), 5% (w/w) A-chain (invertedtriangles), 5% (w/w) B-chain (triangles), and 5% (w/w)lysozyme (diamonds). Aggregation was followed as theincrease in solution turbidity. The lines are only to guidethe eye.
500 Self-sequence Modulation of Insulin Aggregation
automate the data processing and to minimiseoperator-induced bias, we have developed analgorithm (the source is code available from thecorresponding author on request) that automaticallymeasures the heights at multiple locations alongindividual fibrils relative to the background (Figure2, middle panels), enabling a rigorous statisticalanalysis to be made of their dimensions. Ourmeasurements reveal interesting morphologicaldifferences between the three systems. For intactinsulin fibrils, non-twisted species with heights ofapproximately 3 nm were observed, which weascribe to being individual protofilaments. Addi-tionally, twisted fibrils with an average height ofapproximately 5.5 nm and a mean period of 45–50 nm in the direction of the fibril axis were present,suggesting that they were composed of at least twoprotofilaments assembled in a helical fashion,similar to those commonly reported for maturefibrils. We note that these dimensions are also invery good agreement with previous results fromcryo-electron microscopy image reconstructions.11For the A-chain fibrils, we observed a wide range ofmorphologies presenting with a very broad, bimod-al distribution of heights that were larger than forfibrils composed of intact insulin. The broadness ofthe distribution suggests that the protofilamenthierarchy in this system has a more complexorganization than for insulin fibrils. This is sup-ported by the irregularity of the helical repeats inthese fibrils, which varied between 65 nm and 90 nmin the direction of the fibril axis. Finally, the B-chainfibrils demonstrated some evidence of twistedmorphologies in the very early stages of theassembly process; however, mature fibrils agedfrom a few days up to a year showed little evidenceof periodicity. Furthermore, the height distributionfor B-chain fibrils was exceptionally narrow. Similarto insulin fibrils, B-chain fibril populations withheights of approximately 3 nm and 5.5 nm wereobserved; however, in the case of the B-chain, the3 nm fibrils were most abundant.X-ray fibre diffraction was used to characterize
further the insulin, A-chain and B-chain fibrils. Asshown in Figure 2 (rightmost panels), each of thefibril samples exhibited an anisotropic diffractionpattern consistent with a cross-β structure, charac-teristic of amyloid fibrils. Here, all of the fibrilsamples gave a meridional reflection at 4.77 Å,corresponding to the spacing between adjacenthydrogen bonded β-strands lying perpendicular tothe fibril axis. This reflection for the A-chain samplewas a doublet (with a second reflection at 4.66 Å), aphenomenon that has been observed in a number offibril systems.6 Equatorial reflections at 11.0 Å, 9.4 Åand 10.4 Å for insulin, A-chain and B-chain fibrils,respectively, arise from the separation between theβ-sheets that run parallel with the fibril axis. Thesedata confirm that fibrils formed by the threepolypeptides contain essentially the same funda-mental core structure. The variations observed in theequatorial reflections for the three fibril samples isprobably indicative of the incorporation of different
primary sequence into the cross-β core for eachpolypeptide resulting in varied inter-sheet packingdistances.28
Cross-seeding of insulin fibril formation byA-chain and B-chain fibrils
Fibrils composed of the insulin B-chain have beenshown in a recent independent study to be capableof nucleating the fibril formation of the full-lengthprotein.24 We next examined the capacity for insulinfibril formation to be nucleated by A-chain fibrils,and compared directly the efficiency of this processwith that elicited by B-chain and insulin fibrils.Fibril formation by intact insulin was initiated by
incubation of the protein at a concentration of0.44 mM (2.5 mg/ml) at pH 2.0 and 60 °C in thepresence and in the absence of 5% (w/w) preformedfibrils. The aggregation process was followed bymonitoring the increase in absorbance at 360 nmarising from increased sample turbidity (Figure 3).Under these conditions, insulin fibril formationfollowed sigmoidal kinetics typical of a nucleation-growth mechanism and defined by an initial lagphase during which no significant change inturbidity occurred, and a subsequent growth phasein which the turbidity reached a plateau as aggre-gation tended towards completion. As shown inFigure 3, the lag phase for insulin fibril formationlasted for approximately 500 min under theseconditions, which was followed by a rapid rise inturbidity that was complete by 700 min. Theaddition of 5% (w/w) preformed insulin fibrils to

501Self-sequence Modulation of Insulin Aggregation
an identical reaction abolished the lag phasecompletely, and the reaction was complete withinca 50 min. Addition of 5% (w/w) A-chain or B-chainfibrils did not abolish the lag phase completely,although the lag time was reduced significantly ineach case. This is consistent with a recent, indepen-dent observation by Hong and Fink that B-chainfibrils seed insulin aggregation less efficiently thanintact insulin fibrils.24 Interestingly, in the currentstudy, we found that the A-chain fibrils seededinsulin fibril formation more efficiently than B-chainfibrils, resulting in reductions in the lag time of75(±5)% and 36(±7)% (n=4), respectively. This effectwas shown to be sequence-specific, as the additionof 5% (w/w) lysozyme fibrils had no detectableeffect on the fibrillization kinetics of insulin.The morphology of the cross-seeded fibrils was
examined by AFM (Figure 4). Both the A-chain andthe B-chain seeded fibrils were morphologicallydistinct from the seed material, and exhibitedsimilarities to insulin fibrils formedwithout seeding.In each case, fibrillar species with twisted morphol-ogies were observed to have an average periodicityof ca 45–50 nm (Figure 4, left panels). The heightdistributions for these samples were broader thanfor virgin insulin fibrils (Figure 4, right panels). The
Figure 4. Morphology of cross-seeded fibrils. Insulin was i60 °C in the presence of 5% (w/w) (a) A-chain fibrils and (b) B(left panel) and height distributions were attained from tmagnifications of the regions indicated.
lower end of this distribution was approximately3 nm, suggesting that the cross-seeded fibrils werecomposed of filaments with the same diameter asthose observed for typical insulin fibrils. However,the distributions tended towards a higher upperlimit of heights (9 – 10 nm) than for insulin fibrilsformed without seeding, probably indicating thepresence of fibrils assembled from a greater numberof intertwined protofilaments.The cross-seeded fibrils were subsequently added
to fresh insulin aggregation reactions at a concentra-tion of 5% (w/w) in order to examine the formationof a second generation of cross-seeded fibrils (Figure5). In contrast to the initial cross-seeding event, theaddition of both the A-chain and B-chain cross-seeded fibrils abolished the lag phase of the reactioncompletely. The kinetic traces fit well to a mono-exponential equation, giving rates that were identi-cal, within error, with the rate attained by seedingwith virgin insulin fibrils.
Inhibition of insulin fibril formation by solubleA-chain and B-chain peptides
Initial attempts to seed insulin fibril formation bythe addition of A-chain or B-chain fibrils were
ncubated at a final concentration of 0.44 mM at pH 2.0 and-chain fibrils. The resultant fibrils were analysed by AFMhese data (right panel). Insets in AFM images are 2×

Figure 6. Inhibition of insulin fibril formation bysoluble A-and B-chain peptides. Insulin was incubated ata final concentration of 0.44 mM at pH 2.0 and 60 °C inisolation (squares) or in the presence of increasing molarratios of (a) A-chain or (b) B-chain peptide: 0.1 (triangles),0.2 (diamonds) 0.3 (inverted triangles), 0.4 (open circles)and 0.5 (filled circles). Crossed symbols represent thehighest concentrations of A-chain or B-chain peptidesused, incubated in isolation under identical conditions.Aggregation was followed as the increase in solutionturbidity. The lines are only to guide the eye.
Figure 5. Production of a second generation of cross-seeded fibrils. Insulin was again incubated at a finalconcentration of 0.44 mM at pH 2.0 and 60 °C in thepresence of 5% (w/w) fibrils previously cross-seeded byA-chain (inverted triangles) and B-chain (triangles) fibrils.Insulin seeded under identical conditions by virgin insulinfibrils is included as a control (squares). Aggregation wasfollowed as the increase in solution turbidity. The linesrepresent mono-exponential fits to the data.
502 Self-sequence Modulation of Insulin Aggregation
hindered by the presence of residual soluble A-chainor B-chain peptide in the fibrillarmaterial. The effectsof soluble A-chain and B-chain peptides on insulinfibril formation were therefore investigated.Insulin was incubated at a concentration of
0.44 mM (2.5 mg/ml) at pH 2.0 and 60 °C in thepresence and in the absence of soluble A-chain orB-chain peptide at sub-stoichiometric molar ratiosranging from 0.1 to 0.5 (i.e. 0.044 mM to 0.22 mM).At the highest concentration used, neither the A-chain nor the B-chain peptide was found toaggregate for the duration of the experiment (seeFigure 6(a) and (b), cross symbols). At a molarratio of 0.1, the soluble A-chain increased the lagtime for insulin fibrillization from 500 min to950 min, whilst ratios of 0.2 and 0.3 resulted ininhibition such that the lag times could not beresolved within the 2000 min duration of themeasurements (Figure 6(a)). Examination of thesesamples by TEM at the conclusion of the experi-ment revealed the presence of fibrillar material,albeit with fewer fibrils in those samples consistentwith greater inhibition than in the control samplethat contained no A-chain peptide (Figure 7(a) and(b)). At the highest molar ratios of 0.4 and 0.5, theaggregation behavior of insulin was markedlydifferent from that observed at lower ratios. At amolar ratio of 0.4, the lag phase was of a lengthsimilar to that of the control containing no A-chainpeptide, although the rate of growth was muchslower and the extent of solution turbidity wasgreater. At the highest molar ratio examined (0.5)the lag phase was reduced to less than 250 min and
again the rate of growth was slower relative to thecontrol. These samples were examined by TEMand were found to contain almost no fibrillarmaterial; instead, they contained large amorphousaggregates (Figure 7(c) and (d)).The generation of amorphous aggregates was not
observed when insulin was incubated in thepresence of the soluble B-chain peptide, whichresulted in a simple concentration-dependent inhi-bition at all molar ratios tested (Figure 6(b)). Here,increasing ratios of soluble B-chain extended the lagtime for insulin aggregation and resulted in lowerlevels of solution turbidity, indicating that the final

Figure 7. TEM analysis of insu-lin aggregates formed in the pres-ence of soluble A-chain and B-chainpeptides. Insulin was incubated at afinal concentration of 0.44 mM atpH 2.0 and 60 °C in the presence ofmolar ratios of (a) 0.1 A-chain, (b)0.2 A-chain, (c) 0.4 A-chain, (d) 0.5A-chain, (e) 0.1 B-chain and (f) 0.5 B-chain. Samples for TEM were pre-pared at the end of the 2000 mintime-course. The scale bars repre-sent 200 nm.
503Self-sequence Modulation of Insulin Aggregation
quantities of aggregated material formed in thesereactions was reduced relative to the control in theabsence of B-chain. Inspection of these samples byTEM at the completion of the time-course revealedthe presence of fibrillar material at all ratios ofsoluble B-chain to insulin (Figure 7(d) and (e)). Thequantity of fibrils observed on the TEM gridappeared to decrease with increasing concentrationsof added B-chain, and very little amorphous mate-rial was observed in any of these samples.The inhibitory effect elicited by the soluble A-chain
and B-chain peptides was further investigated anddemonstrated to be sequence-specific. In an experi-ment identical with that described above, 0.22 mMsoluble TTR105-115 peptide (equivalent to a molarratio of 0.5) was incubated with insulin instead ofeither the soluble A-chain or B-chain peptides.TTR105-115 was chosen, as its sequence is unrelatedto insulin and, like the insulin peptides, is essentiallydisordered in aqueous solution.29,30 Under theconditions used here, the TTR105-115 peptide did notaggregate detectably during the time-course of themeasurements (Figure 8, cross symbols). The incu-bation of TTR105-115 with insulin had no observableeffect on the aggregation of insulin, with the kinetictrace virtually overlying that of the control (Figure 8).As the incubation with the soluble A-chain and B-
chain peptides increased the lag time for insulin
aggregation dramatically, we examined whetherthis effect was due primarily to the peptidesinterfering with the insulin nucleation process byseeding insulin fibrillization in the presence ofinhibitory ratios of A and B-chain. A ratio of 0.2 ofeither A-chain or B-chain peptide was used in thisexperiment, as this ratio resulted in effectiveinhibition by both peptides, without the complica-tion of the formation of amorphous aggregatesobserved at the higher concentrations of the A-chain. The final concentration of insulin fibrils usedto seed the reactions was 5% (w/w). As shown inFigure 9(a), there is no observable lag phase in anysuch experiment, and the kinetic traces fit well to amono-exponential equation. In the absence ofexogenous peptide, the rate of aggregation was0.15(±0.004) min−1, whilst in the presence of A andB-chain, the rates of aggregation were 0.11(±0.002)min−1 and 0.12(±0.003) min−1, respectively. Thesedata suggest that the A-chain and B-chain peptidesexert their inhibition predominantly on the forma-tion of nuclei by insulin molecules, and inhibit therate of fibril growth only slightly. Interestingly, thefinal absorbance of the sample containing the A-chain peptide was almost twice as high as that forthe control or the sample with the added B-chain.The A-chain and B-chain samples were examined byTEM and, whilst both samples contained abundant

Figure 8. The effect of the TTR105-115 peptide on insulinaggregation. Insulin was incubated at a final concentrationof 0.44 mM at pH 2.0 and 60 °C in isolation (squares) or inthe presence of TTR105-115 at a molar ratio of 0.5 (triangles).Crossed symbols represent 0.22 mM TTR105-115 incubatedin isolation under identical conditions. Aggregation wasfollowed as the increase in solution turbidity. The lines areonly to guide the eye.
504 Self-sequence Modulation of Insulin Aggregation
fibrils, the sample incubated with A-chain alsocontained a significant population of amorphousaggregates (Figure 9(b)).
Complex formation between insulin and itscomponent chains
The observation that the soluble A-chain and B-chain peptides exert their inhibitory effects byinterfering with the insulin nucleation processprompted us to test whether the peptides are capableof associating with insulin under the solutionconditions used for the inhibition studies. Solutionsof insulin and its A-chain and B-chain peptides wereprepared in dilute HCl at pH 2.0 and analysed bynano-ESIMS. Them/z spectrumof 0.44mM insulin atpH 2.0 and 60 °C reveals a number of charge stateswithin them/z range 1000– 2500 (Figure 10(a)). Thesecharge states can be attributed to the presence of bothmonomers and dimers of the protein (measuredmasses of 5734(±0.12) Da and 11,469(±0.26) Da, res-pectively), each of which shows a full charge stateenvelope and unique peaks that can be assignedunambiguously. In a previous study that used nano-ESI MS to examine the oligomerization of insulinunder similar conditions, higher-order oligomerscontaining up to 12 molecules were observed.20 Thisdifference can be attributed to the higher concentra-tions of insulin used in the earlier study.Mixed polypeptide solutions were prepared to
give final concentrations of 0.44 mM insulin and0.22 mM for either the A-chain or B-chain peptide.These concentrations correspond to the highestmolar ratios examined in the inhibition studies. The
m/z spectra of insulin incubated at pH 2.0 and 60 °Cwith either the A-chain or B-chain peptide shows anumber of peaks in them/z range 1000 – 3000 (Figure10(b) and (c)). In both instances, peaks cor-responding to insulin monomer and either the A-chain or B-chain monomer can be observed, whilstother peaks can be assigned unambiguously ascomplexes of insulin with the A or B-chains. In thesample containing intact insulin and A-chain pep-tide, complexes of insulin monomer with A-chainmonomer, insulin dimer with A-chain monomer andinsulin dimer with A-chain dimer (with masses of8267(±0.08) Da, 14001(±0.17) Da and 16,534(±0.20)Da, respectively) were observed. In the samplecontaining intact insulin and B-chain peptide, com-plexes of the B-chain with insulin monomer anddimer (with masses of 9229(±0.12) and 14,962(±0.24)Da, respectively) were observed. The complexesformed between insulin and the B-chain peptidewere significantly less populated than the complexesformed between insulin and the A-chain peptide,implying a greater binding affinity for the A-chain.
Discussion
Insulin represents a very interesting protein withwhich to study the process of fibril formation, as itsstructural flexibility is heavily constrained by thepresence of three disulfide bonds within its twocomponent polypeptide chains. This degree of cross-linking results in a very stable native fold that isresistant to both chemical and thermal denatur-ation.31–33 Nevertheless, the insulin molecule popu-lates a partially unfolded conformation under fibril-forming conditions of low pH and elevated temper-ature in which the termini of its component chains inparticular become disordered and expose the helicalcore of the protein.14 This is in keeping with thenotion that the adoption of partial folds is aprerequisite to the formation of amyloid fibrils.8,9,17A large body of evidence indicates that, subsequentto the adoption of a partially folded state, amyloidfibril formation occurs via a nucleation-polymerisa-tion model, whereby the initial formation ofoligomeric nuclei is thermodynamically disfa-voured.34 Beyond a ‘‘critical nucleus’’ size, theaggregation process becomes favourable primarilyas the result of the formation of stable hydrogenbonds between the β-strand regions of polypeptidechains. Mutagenesis data imply that interactionsformed by residues within both of the componentchains of the insulin molecule are critical to thisnucleation process.18 In the current study, wedemonstrate that the nucleation of insulin fibrilformation can be modulated significantly by incu-bation of the protein with exogenous peptidessharing the amino acid sequences of its componentchains.We have observed in the current study that the A-
chain of insulin can independently form fibrilsunder conditions similar to those for the parentprotein, and we confirm the recent observation that

Figure 9. The effect of solubleA-and B-Chain peptides on self-seededinsulin aggregation. (a) Insulin wasincubated at a final concentration of0.44 mM at pH 2.0 and 60 °C in thepresence of 5% (w/w) preformedinsulin fibrils (open squares) and inthe presence of molar ratios ofsoluble A-chain (inverted triangles)and B-chain (triangles) peptides of0.2.Aggregationwas followedas theincrease in solution turbidity. Thelines represent mono-exponentialfits to the data. (b) Samples forTEMwere prepared after incubationfor 100 min. Representative imagesof seeded insulin aggregates formedin the presence of A-chain (leftpanel) and B-chain (right panel)peptides are shown. The arrow inthe left panel points to a large amor-phous aggregate. The scale bars rep-resent 200 nm.
505Self-sequence Modulation of Insulin Aggregation
the B-chain is capable of aggregating into fibrillarstructures.24 Hong and Fink have suggested thatfibrils composed solely of the B-chain are morpho-logically indistinguishable from intact insulin fibrilsas judged by TEM.24 In the current study, however,we observe by AFM varied morphologies for thefibrils composed of insulin and its two polypeptidechains (Figure 2). On the basis of our fibril heightmeasurements, the A-chain produces fibrils oflarger diameters than insulin fibrils and with alarger, less uniform helical repeat, whereas the B-chain produces exceptionally homogeneous fibrilsthat appear predominantly untwisted. Both the A-chain and B-chain fibrils were found to be able tonucleate fibril growth by insulin, although theefficiency of this process is much lower than forself-seeding with preformed fibrils of intact insulin(Figure 3). These data indicate that the accessibleregions of partially unfolded insulin molecules are,at least in part, compatible with the ends of the A-chain and B-chain fibrils. Interestingly, the A-chainfibrils consistently seeded with greater efficiency,indicating that they were more compatible withinsulin fibrillization than the B-chain fibrils. The
limited compatibility between the A-chain and B-chain fibrils and the intact insulin moleculesuggests that the primary sequence involvementand mode of polypeptide packing in the core of thefibril structure is different in each case, which issupported by the variation in equatorial reflectionsobserved in the X-ray diffraction patterns for thedifferent fibril types. The evidence implicating theinvolvement of both the A-chains and B-chains ofthe insulin molecule in the cross-β core of intactinsulin fibrils implies that the A-chain or B-chainfibrils in isolation do not contain the full comple-ment of sequence information necessary for effi-cient seeding.11,18 The nucleation of fibril growthhas been shown to be exceptionally sequence-dependent, even between homologues of the sameprotein from different species.35–38The inefficiency of the cross-seeding process
indicates that the initial binding of insulin moleculesonto the A-chain or B-chain fibril may not be asenergetically favourable as addition onto fibrilscomposed of intact insulin. Alternatively, the firstmolecules of insulin to add onto the seed fibrils mayrequire reorganization into a conformation that is

Figure 10. Interactions between insulin and itscomponent chains determined by nano-ESI MS. (a)Insulin was analysed in isolation at a concentration of0.44 mM in dilute HCl at pH 2.0 maintained at 60 °C.Insulin solutions at a concentration of 0.44 mM were alsoanalysed under identical conditions in the presence of(b) 0.22 mM A-chain or (c) B-chain. The number ofmolecules of insulin (I), A-chain (A) and B-chain (B)within each species is shown for each peak with thecharge state in parentheses.
506 Self-sequence Modulation of Insulin Aggregation
compatible with further elongation. Nonetheless,once elongation-competent insulin molecules poly-merize onto the seed fibrils, growth occurs cooper-atively, as shown by the subsequent rapid evolutionof cross-seeded fibrils, and their capacity to effi-ciently nucleate a second generation of insulin fibrilformation (Figure 5). The subtle morphologicaldifferences between virgin insulin fibrils and thecross-seeded fibrils (Figure 4), however, suggest thatthe conformation of insulin molecules may be subtlydifferent in each fibril type, as has been indicated forinsulin fibrils cross-seeded by the B-chain.24Although the A-chain and B-chain fibrils were not
observed to transmit their morphology to the cross-seeded fibrils in the current study, the transmissionof insulin fibril morphology by seeding has beenshown in experiments in which morphologicallydistinct fibrils were produced under differentsolution conditions.39 Similar observations havebeen made for fibrils composed of the β-amyloidpeptide, associated with Alzheimer’s disease.40Fibril morphology has been proposed to be thebasis of strain variability observed in mammalianprion amyloid fibrils and, as such, as the basis ofprion disease transmission.41 In the current study,the morphologies of the A-chain and B-chain fibrilsare not propagated in the cross-seeded insulinfibrils. This suggests that the molecular differencesunderlying fibril strains must be subtle; in thepresent investigation the A-chain and B-chain eachrepresent cases of substantial sequence truncationcompared to the intact protein. Elegant work by thelaboratory of Surewicz has demonstrated that thestructural differences in prion amyloid fibril strainsare controlled by just one or two residues.41The incubation of insulin with the soluble forms of
the A-chain and B-chain peptides produced effectsin stark contrast to the nucleation observed by thechains in the fibrillar form (Figures 6 and 7). In theseexperiments, both peptides inhibited the aggrega-tion of insulin in a concentration-dependent mannerat sub-stoichiometric molar ratios, although thehighest ratios of A-chain tested induced the forma-tion of amorphous aggregates. As shown in Figure10, the peptides were found to form discretecomplexes with the insulin molecules, which mayaccount for the observed inhibition of nucleationduring insulin fibril formation.The so called β-sheet breaker peptides have been
used very successfully to inhibit and reverse fibrilformation by the β-amyloid peptide and the prionprotein.42–44 These peptides are typically designedto represent an amyloidogenic region of the se-quence of the target protein with the inclusion ofseveral proline residues, which are disfavoured in β-sheet structures and decrease the aggregationpropensity of the peptides. In the current investiga-tion, however, the A-chain and B-chain peptidesequences have not been altered and can indeedthemselves assemble into fibrils. The inhibition ofaberrant conformational changes and fibril forma-tion using peptides with identical but truncatedamino acid sequences with their target protein has

507Self-sequence Modulation of Insulin Aggregation
been observed for systems other than insulin. Forexample, incubation of the hamster prion proteinwith peptide stretches from the central portion of thesequence, which includes the most amyloidogenicregion, inhibit the conversion of the protein fromnative state to the β-sheet rich, aggregation-proneconformation.45 Further, it has been reported that apeptide from the yeast protein, Sup35, inhibits theaggregation of the full-length protein, albeit to arelatively small extent.46 In these cases, the truncat-ed peptides could exert their effect by associatingwith exposed residues on the target molecule thatare critical to nucleation, effectively shielding thesesites from further associations, akin to a self-sequence chaperoning mechanism.In summary, we have demonstrated that the
associations formed between insulin and exogenouspeptides corresponding to its component polypep-tide chains are complex and critically dependent onpeptide conformation, given that the same chemicalform of the polypeptide chains in different physicalstates can either stimulate or inhibit the conversion ofinsulin into amyloid fibrils. The inhibition of insulinfibril formation by peptides representing truncatedsegments of its own primary sequence appears to bedemonstrative of a generic effect, as similar behaviorhas been observed in a variety of other systems.Short peptides are considered to represent anattractive therapeutic strategy for treatment of pro-tein misfolding disorders.44 However, since inhibi-tory peptides, by their very nature, must interactwith their target proteins, consideration needs to begiven to the fact that such peptides, under alternativeconditions, might stimulate aberrant protein aggre-gation, as demonstrated in this study.
Materials and Methods
Polypeptides
Lysozyme from chicken egg-white, insulin from bovinepancreas (FW 5733.49) and oxidized bovine insulin A-chains and B-chains (FW 2531.64 and 3495.89, respective-ly) in which all cysteine residues are modified to cysteicacid were purchased from Sigma-Aldrich Co. Ltd.(Gillingham, Dorset, UK). Insulin and B-chain peptidewere used without further manipulation; however, for theinhibition studies, the A-chain was further purified usingsolid-phase extraction on a Sep-Pak C18 cartridge (WatersCo., Elstree, Hertsfordshire, UK) and subsequently lyoph-ilized. The peptide consisting of residues 105–115 ofhuman transthyretin (TTR105-115) with the amino acidsequence, YTIAALLSPYS, was synthesized using stan-dard automated solid-phase peptide synthesis techniques,incorporating Fmoc-protected amino acids using a RaninPS3 synthesizer (Protein Technologies Inc, AZ, USA).Following de-protection and cleavage from the resin, thepeptide was lyophilized and contaminants removed bysolid-phase extraction on a Sep-Pak C18 cartridge.Characterization by matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF)mass spectrometryand HPLC confirmed the identity of the peptide andindicated that the sample purity was >95%. Therefore, nofurther purification of the peptide was performed.
Aggregation kinetics
Insulin aggregation kinetics were monitored by incu-bating the protein in a final volume of 1 ml of dilute HCl atpH 2.0 and 60 °C in Plastibrand disposable plastic UVcuvettes with Plastibrand plastic stoppers (Fisher Scien-tific Ltd., Loughborough, Leicestershire, UK) to preventevaporation. Samples were not stirred. Absorbance wasmonitored at 360 nm every 1–5 min with each time-pointaveraged for 5 s in a Varian Cary 300 UV spectrophotom-eter. The temperature was maintained at 60 °C using aPeltier temperature controller.
Seeding of fibrillisation reactions
Preformed insulin and B-chain fibrils were prepared byincubation at concentrations of 4 mM in dilute HCl at pH2.0 and 60 °C for 24 h. Preformed A-chain fibrils wereprepared by incubation at concentration of 4 mM in diluteHCl at pH 2.0 and 30 °C for 24 h. Lysozyme fibrils wereprepared at a concentration of 3.6 mM by incubation ofthe protein in dilute HCl at pH 2.0 and 65 °C until thesolution became very viscous.47 The resultant fibrillarmaterial was separated from residual soluble polypeptideby washing at least twice with 0.5 ml of dilute HCl at pH2.0 using Whatman VectaSpin Micro Centrifuge Filterswith a 0.02 μm pore size Anopore membrane (FisherScientific Ltd., Loughborough, Leicestershire, UK). Thefibrillar material was then resuspended and frozen inliquid nitrogen. Fibrils were thawed at room temperaturebefore being added to fresh polypeptide solutions forseeding.
Transmission electron microscopy
Copper transmission electron microscopy (TEM) gridswere formvar and carbon-coated. Samples were diluted to<1 mg/ml and applied to the grid. The grid was thenwashed and negatively stained with 2% (w/v) uranylacetate. TEM grids were analyzed using an FEI PhilipsTechnai 20 TEM with an acceleration voltage of 120 keV.
Atomic force microscopy
AFMmicroscopy data were acquired using a MolecularImaging Pico Plus microscope operating in AC-AM mode(tapping mode) in air. Ultrasharp Micromasch NCS36silicon cantilevers were used at resonance frequenciesbetween 150 kHz and 225 kHz. Samples for AFM analysiswere prepared by diluting the fibril solutions by a factor of30–200 in dilute HCl at pH 2.0, and 20 μl was subsequentlydeposited onto a freshly cleaved mica surface. Thesamples were then left to dry in air for 60 min whilstshielded from dust particles.
Analysis of AFM data
In light of recent evidence that individual amyloidfibrils can display several distinct morphologies alongtheir length, we chose to analyze fibril heights at multiplepoints along their lengths.27 To this end, an algorithm wasdeveloped. Two or more operator-selected points on theAFM topographs were used to specify the location of afibril such that the algorithm could automatically trace itsshape by iteratively finding the highest points along thefibril axis. Subsequently, a piecewise polynomial was

508 Self-sequence Modulation of Insulin Aggregation
fitted to these points, and every 7.8 nm a height profileperpendicular to the fibril axis was extracted. The portioncorresponding to the fibril was determined from a simplecutoff procedure, and the remaining points, which arepart of the background, are fit to a line. Finally, the heightdifference from the highest point on the fibril to thecorresponding position on the background was computedfor each of the locations along the fibrils to give the fibrilwidth. Height data were acquired from a randomselection of at least 60 different fibrils in each sample.
X-ray fibre diffraction
Amyloid fibril samples were prepared for X-raydiffraction analysis using a modification of the stretch-frame method.48 In this modification, the distancebetween capillary tubes is not changed during theproduction of the semi-aligned, dry stalk. A 10 μl dropletof fibril suspension was placed between two horizontalwax-plugged glass capillary tubes of 1 mm diameter heldapproximately 2 mm apart. Upon drying of the droplet toapproximately a quarter of its initial volume, a further 5 μlof fibril suspension was added and the process wasrepeated a further three times. Subsequently, the dropletwas left to dry completely in air. All scattering patternswere obtained on a crystallography beamline of theBiochemistry Department at Cambridge University. X-rays with a wavelength of 1.54 Å were produced by arotating copper anode, and collimated and focused byOsmic Max-flux optics. Images were acquired on a Marrimage plate. The sample–detector distance was 250 mm,and data acquisition times were typically 30 min.
Nano-electrospray ionization mass spectrometry
Mass spectrometry data were acquired on a Q-ToF 2instrument (Waters, Manchester, U.K.) modified for highmass operation.49 The following experimental parameterswere used: capillary voltage 1.4 kV, sample cone 120 V,extractor cone 4 V, collision energy 20–30 V and iontransfer stage pressure 3×10−2 mbar (1 bar=105 Pa).External instrument calibration was applied using cesiumiodide (33 mg/ml). Temperature adjustment to 60 °C atthe source was achieved using a thermocoupled nano-ESIprobe.50 Data were processed using Masslynx v3.5(Waters) and spectra are shown without backgroundsubtraction and with minimal smoothing.
Acknowledgements
We thank Dr Anna Tickler for providing theTTR105-115 peptide, Drs Ben Luisi and Dima Chir-gadze for assistance with X-ray diffraction, Profes-sor Mark Welland for valuable advice on AFM andMr Kevin Channon for useful discussions. C.E.M. isa University Research Fellow of the Royal Societyand G.L.D. is a C.J. Martin Research Fellow of theNational Health and Medical Research Council,Australia. S.L.G. is funded by a Gates CambridgeScholarship and T.P.J.K. acknowledges support fromthe IRC in nanotechnology. The research of C.M.D.is supported, in part, by ProgrammeGrants from theWellcome and Leverhulme Trusts.
References
1. Waugh, D. (1946). A fibrous modification of insulin. I.The heat precipitate of insulin. J. Am. Chem. Soc. 68,247–250.
2. Lougheed, W. D., Woulfe-Flanagan, H., Clement, J. R.& Albisser, A. M. (1980). Insulin aggregation inartificial delivery systems. Diabetologia, 19, 1–9.
3. Hutchison, K. G. (1985). Assessment of gelling ininsulin solutions for infusion pumps. J. Pharm.Pharmacol. 37, 528–531.
4. Dische, F. E., Wernstedt, C., Westermark, G. T., Wes-termark, P., Pepys, M. B., Rennie, J. A. et al. (1988).Insulin as an amyloid-fibril protein at sites of repeatedinsulin injections in a diabetic patient. Diabetologia, 31,158–161.
5. Blake, C. & Serpell, L. (1996). Synchrotron X-raystudies suggest that the core of the transthyretinamyloid fibril is a continuous beta-sheet helix.Structure, 4, 989–998.
6. Sunde, M., Serpell, L. C., Bartlam, M., Fraser, P. E.,Pepys, M. B. & Blake, C. C. (1997). Common corestructure of amyloid fibrils by synchrotron X-raydiffraction. J. Mol. Biol. 273, 729–739.
7. Nilsson, M. R. (2004). Techniques to study amyloidfibril formation in vitro. Methods, 34, 151–160.
8. Kelly, J. W. (1998). The alternative conformations ofamyloidogenic proteins and their multi-step assemblypathways. Curr. Opin. Struct. Biol. 8, 101–106.
9. Dobson, C. M. (2003). Protein folding and misfolding.Nature, 426, 884–890.
10. Brange, J., Andersen, L., Laursen, E. D., Meyn, G. &Rasmussen, E. (1997). Toward understanding insulinfibrillation. J. Pharm. Sci. 86, 517–525.
11. Jimenez, J. L., Nettleton, E. J., Bouchard, M., Robinson,C. V., Dobson, C. M. & Saibil, H. R. (2002). Theprotofilament structure of insulin amyloid fibrils. Proc.Natl Acad. Sci. USA, 99, 9196–9201.
12. Bryant, C., Spencer, D. B., Miller, A., Bakaysa, D. L.,McCune, K. S., Maple, S. R. et al. (1993). Acidstabilization of insulin. Biochemistry, 32, 8075–8082.
13. Adams, M., Blundell, T., Dodson, E., Dodson, G., Vija-yan, M., Baker, E. et al. (1969). Structure of rhombohe-dral 2 zinc insulin crystals. Nature, 224, 491–495.
14. Hua, Q. X. & Weiss, M. A. (2004). Mechanism ofinsulin fibrillation: the structure of insulin underamyloidogenic conditions resembles a protein-foldingintermediate. J. Biol. Chem. 279, 21449–21460.
15. Booth, D. R., Sunde, M., Bellotti, V., Robinson, C. V.,Hutchinson, W. L., Fraser, P. E. et al. (1997). Instability,unfolding and aggregation of human lysozymevariants underlying amyloid fibrillogenesis. Nature,385, 787–793.
16. Guijarro, J. I., Sunde, M., Jones, J. A., Campbell, I. D. &Dobson, C. M. (1998). Amyloid fibril formation by anSH3 domain. Proc. Natl Acad. Sci. USA, 95, 4224–4228.
17. Uversky, V. N. & Fink, A. L. (2004). Conformationalconstraints for amyloid fibrillation: the importance ofbeing unfolded. Biochim. Biophys. Acta. 1698, 131–153.
18. Nielsen, L., Frokjaer, S., Brange, J., Uversky, V. N. &Fink, A. L. (2001). Probing the mechanism of insulinfibril formation with insulin mutants. Biochemistry, 40,8397–8409.
19. Bouchard, M., Zurdo, J., Nettleton, E. J., Dobson, C. M.&Robinson, C. V. (2000). Formation of insulin amyloidfibrils followed by FTIR simultaneously with CD andelectron microscopy. Protein Sci. 9, 1960–1967.
20. Nettleton, E. J., Tito, P., Sunde, M., Bouchard, M.,Dobson,C.M.&Robinson, C.V. (2000). Characterization

509Self-sequence Modulation of Insulin Aggregation
of the oligomeric states of insulin in self-assembly andamyloid fibril formation by mass spectrometry. Biophys.J. 79, 1053–1065.
21. Ahmad, A., Uversky, V. N., Hong, D. & Fink, A. L.(2005). Early events in the fibrillation of monomericinsulin. J. Biol. Chem. 280, 42669–42675.
22. Jansen, R., Dzwolak, W. & Winter, R. (2005). Amyloi-dogenic self-assembly of insulin aggregates probed byhigh resolution atomic force microscopy. Biophys. J. 88,1344–1353.
23. Plakoutsi, G., Bemporad, F., Calamai, M., Taddei, N.,Dobson, C. M. & Chiti, F. (2005). Evidence for amechanism of amyloid formation involving molecularreorganisation within native-like precursor aggre-gates. J. Mol. Biol. 351, 910–922.
24. Hong, D. P. & Fink, A. L. (2005). Independentheterologous fibrillation of insulin and its B-chainpeptide. Biochemistry, 44, 16701–16709.
25. Chamberlain, A. K., MacPhee, C. E., Zurdo, J.,Morozova-Roche, L. A., Hill, H. A., Dobson, C. M. &Davis, J. J. (2000). Ultrastructural organization ofamyloid fibrils by atomic force microscopy. Biophys. J.79, 3282–3293.
26. Malisauskas, M., Ostman, J., Darinskas, A., Zamotin,V., Liutkevicius, E., Lundgren, E. & Morozova-Roche,L. A. (2005). Does the cytotoxic effect of transientamyloid oligomers from common equine lysozyme invitro imply innate amyloid toxicity? J. Biol. Chem. 280,6269–6275.
27. Anderson, M., Bocharova, O. V., Makarava, N.,Breydo, L., Salnikov, V. V. & Baskakov, I. V. (2006).Polymorphism and ultrastructural organization ofprion protein amyloid fibrils: an insight from highresolution atomic force microscopy. J. Mol. Biol. 358,580–596.
28. Fandrich,M. &Dobson, C.M. (2002). The behaviour ofpolyamino acids reveals an inverse side chain effect inamyloid structure formation. EMBO J. 21, 5682–5690.
29. Wu, C. S. & Yang, J. T. (1981). Conformation of insulinand its fragments in surfactant solutions. Biochim.Biophys. Acta, 667, 285–293.
30. Dirix, C., Meersman, F., MacPhee, C. E., Dobson, C. M.& Heremans, K. (2005). High hydrostatic pressuredissociates early aggregates of TTR105-115, but notthe mature amyloid fibrils. J. Mol. Biol. 347, 903–909.
31. Ahmad, A., Millett, I. S., Doniach, S., Uversky, V. N. &Fink, A. L. (2003). Partially folded intermediates ininsulin fibrillation. Biochemistry, 42, 11404–11416.
32. Ahmad, A., Millett, I. S., Doniach, S., Uversky, V. N.& Fink, A. L. (2004). Stimulation of insulin fibrilla-tion by urea-induced intermediates. J. Biol. Chem.279, 14999–15013.
33. Arora, A., Ha, C. & Park, C. B. (2004). Insulin amyloidfibrillation at above 100 degrees C: new insights intoprotein folding under extreme temperatures. ProteinSci. 13, 2429–2436.
34. Harper, J. D. & Lansbury, P. T. Jr (1997). Models of ayloid seeding in Alzheimer’s disease and scrapie:mechanistic truths and physiological consequences ofthe time-dependent solubility of amyloid proteins.Annu. Rev. Biochem. 66, 385–407.
35. Krebs, M. R., Morozova-Roche, L. A., Daniel, K.,Robinson, C. V. & Dobson, C. M. (2004). Observation
of sequence specificity in the seeding of proteinamyloid fibrils. Protein Sci. 13, 1933–1938.
36. O’Nuallain, B., Williams, A. D., Westermark, P. &Wetzel, R. (2004). Seeding specificity in amyloidgrowth induced by heterologous fibrils. J. Biol. Chem.279, 17490–17499.
37. Vanik, D. L., Surewicz, K. A. & Surewicz, W. K. (2004).Molecular basis of barriers for interspecies transmis-sibility of mammalian prions. Mol. Cell 14, 139–145.
38. Wright, C. F., Teichmann, S. A., Clarke, J. & Dobson,C. M. (2005). The importance of sequence diversity inthe aggregation and evolution of proteins.Nature, 438,878–881.
39. Dzwolak, W., Smirnovas, V., Jansen, R. & Winter, R.(2004). Insulin forms amyloid in a strain-dependentmanner: an FT-IR spectroscopic study. Protein Sci. 13,1927–1932.
40. Petkova, A. T., Leapman, R. D., Guo, Z., Yau, W. M.,Mattson, M. P. & Tycko, R. (2005). Self-propagating,molecular-level polymorphism in Alzheimer’s beta-amyloid fibrils. Science, 307, 262–265.
41. Jones, E. M. & Surewicz, W. K. (2005). Fibrilconformation as the basis of species- and strain-dependent seeding specificity of mammalian prionamyloids. Cell, 121, 63–72.
42. Soto, C., Sigurdsson, E. M., Morelli, L., Kumar, R. A.,Castano, E. M. & Frangione, B. (1998). Beta-sheetbreaker peptides inhibit fibrillogenesis in a rat brainmodel of amyloidosis: implications for Alzheimer’stherapy. Nature Med. 4, 822–826.
43. Soto, C., Kascsak, R. J., Saborio, G. P., Aucouturier, P.,Wisniewski, T., Prelli, F. et al. (2000). Reversion ofprion protein conformational changes by syntheticbeta-sheet breaker peptides. Lancet, 355, 192–197.
44. Permanne, B., Adessi, C., Fraga, S., Frossard, M. J.,Saborio, G. P. & Soto, C. (2002). Are beta-sheet breakerpeptides dissolving the therapeutic problem of Alz-heimer’s disease? J. Neural. Transm. Suppl. 293–301.
45. Chabry, J., Caughey, B. & Chesebro, B. (1998). Specificinhibition of in vitro formation of protease-resistantprion protein by synthetic peptides. J. Biol. Chem. 273,13203–13207.
46. Balbirnie, M., Grothe, R. & Eisenberg, D. S. (2001). Anamyloid-forming peptide from the yeast prion Sup35reveals a dehydrated beta-sheet structure for amyloid.Proc. Natl Acad. Sci. USA, 98, 2375–2380.
47. Krebs, M. R., Wilkins, D. K., Chung, E. W., Pitkeathly,M. C., Chamberlain, A. K., Zurdo, J. et al. (2000).Formation and seeding of amyloid fibrils from wild-type hen lysozyme and a peptide fragment from thebeta-domain. J. Mol. Biol. 300, 541–549.
48. Serpell, L. C., Blake, C. C. & Fraser, P. E. (2000).Molecular structure of a fibrillar Alzheimer’s A betafragment. Biochemistry, 39, 13269–13275.
49. Sobott, F., Hernandez, H., McCammon, M. G., Tito,M. A. & Robinson, C. V. (2002). A tandem massspectrometer for improved transmission and analysisof large macromolecular assemblies. Anal. Chem. 74,1402–1407.
50. Benesch, J. L., Sobott, F. & Robinson, C. V. (2003).Thermal dissociation of multimeric protein com-plexes by using nanoelectrospray mass spectrom-etry. Anal. Chem. 75, 2208–2214.
Edited by K. Kuwajima
(Received 14 February 2006; received in revised form 30 April 2006; accepted 2 May 2006)Available online 17 May 2006
Copyright © 2022 FDOKUMEN



![Problems with a conformation assignment of aryl-substituted resorc[4]arenes](https://static.fdokumen.com/doc/165x107/6324d12685efe380f30661c8/problems-with-a-conformation-assignment-of-aryl-substituted-resorc4arenes.jpg)
















