
The adrenocorticotropic hormone (ACTH)-induced reversal of hemorrhagic shock
15
Resuscitation, 18 (1989) 253-267 Elsevier Scientific Publishers Ireland Ltd. 253 THE ADRENOCORTICOTROPIC HORMONE (ACTH)-INDUCED REVERSAL OF HEMORRHAGIC SHOCK ALFIO BERTOLINI. WILLIAM FERRARI and SALVATORE GUARINI Institute of Pharmacology, University of Modenq via G. Campi 287,411O Modena (Italy) SUMMARY Adrenocorticotropic hormone (ACTHI, while having negligible effects on cardiovascular function in the intact animal, induces a potent and sustained reversal of an otherwise invariably, rapidly fatal condition of hemorrhage- induced hypovolemic shock, in rats and dogs. The main site(s) of action are at the peripheral level; however, subsidiary site(s) of action in the CNS cannot be excluded. The studies on the mechanism of action indicate that the ACTH- induced reversal of hemorrhagic shock (a) is an extra-hormonal, adrenal- independent effect, because it is not affected by adrenalectomy and is shared by many ACTH-fragments practically devoid of corticotropic activity; (bl is antag- onized by morphine in a surmontable way; (cl needs the functional integrity of the sympathetic nervous system (it is prevented by guanethidine, reserpine, and clonidinel and the availability of peripheral alpha-adrenoceptors (it is antagonized by dibenamine, prazosin and yohimbine, but not by practolol); (dl requires the integrity of afferent vagal fibers (it is almost completely abolished by vagotomyl; (el involves central cholinergic networks (it is antagonized by atropine sulphate, but not by atropine methyl bromide: and it is prevented by the intracerebroventricular injection of hemieholinium-31; (fl is associated with a massive increase in the volume of circulating blood, likely due to a mobi- lization from peripheral pooling sites (it is largely prevented by splenectomy or by suprahepatic veins ligature, and is associated with a restoration of the ven- ous blood flow in peripheral vascular beds and with a normalization of venous PO,); (gl is associated with a restoration of heart and spleen adrenoceptors, whose number is significantly decreased during hemorrhagic shock. The survival time of hemorrhage-shocked animals, which is 26 r 3 min in controls, is greatly prolonged (44 + 18 hl by ACTH, provided that the treatment is made within 5 - 10 min after bleeding. Finally, in animals treated with ACTH within 5 - 10 min after bleeding, blood reinfusion retains its effectiveness and reverse shock even if performed 2 - 5 h later. Key words: Shock - Hemorrhage - ACTH - Melanopeptides - Opioids Printed and Published in Ireland
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The adrenocorticotropic hormone (ACTH)-induced reversal of hemorrhagic shock

Resuscitation, 18 (1989) 253-267 Elsevier Scientific Publishers Ireland Ltd.
253
THE ADRENOCORTICOTROPIC HORMONE (ACTH)-INDUCED REVERSAL OF HEMORRHAGIC SHOCK
ALFIO BERTOLINI. WILLIAM FERRARI and SALVATORE GUARINI
Institute of Pharmacology, University of Modenq via G. Campi 287,411O Modena (Italy)
SUMMARY
Adrenocorticotropic hormone (ACTHI, while having negligible effects on cardiovascular function in the intact animal, induces a potent and sustained reversal of an otherwise invariably, rapidly fatal condition of hemorrhage- induced hypovolemic shock, in rats and dogs. The main site(s) of action are at the peripheral level; however, subsidiary site(s) of action in the CNS cannot be excluded. The studies on the mechanism of action indicate that the ACTH- induced reversal of hemorrhagic shock (a) is an extra-hormonal, adrenal- independent effect, because it is not affected by adrenalectomy and is shared by many ACTH-fragments practically devoid of corticotropic activity; (bl is antag- onized by morphine in a surmontable way; (cl needs the functional integrity of the sympathetic nervous system (it is prevented by guanethidine, reserpine, and clonidinel and the availability of peripheral alpha-adrenoceptors (it is antagonized by dibenamine, prazosin and yohimbine, but not by practolol); (dl requires the integrity of afferent vagal fibers (it is almost completely abolished by vagotomyl; (el involves central cholinergic networks (it is antagonized by atropine sulphate, but not by atropine methyl bromide: and it is prevented by the intracerebroventricular injection of hemieholinium-31; (fl is associated with a massive increase in the volume of circulating blood, likely due to a mobi- lization from peripheral pooling sites (it is largely prevented by splenectomy or by suprahepatic veins ligature, and is associated with a restoration of the ven- ous blood flow in peripheral vascular beds and with a normalization of venous PO,); (gl is associated with a restoration of heart and spleen adrenoceptors, whose number is significantly decreased during hemorrhagic shock. The survival time of hemorrhage-shocked animals, which is 26 r 3 min in controls, is greatly prolonged (44 + 18 hl by ACTH, provided that the treatment is made within 5 - 10 min after bleeding. Finally, in animals treated with ACTH within
5 - 10 min after bleeding, blood reinfusion retains its effectiveness and reverse shock even if performed 2 - 5 h later.
Key words: Shock - Hemorrhage - ACTH - Melanopeptides - Opioids
Printed and Published in Ireland

INTRODUCTION
Pursuing our efforts to produce pharmacological evidence in support of the idea that ACTH-MSH peptides (melanopeptidesl may play a prominent role as physiological antagonists of opioids in a general homeostatic system [l - 31, we recently discovered that ACTH and several of its fragments have a potent anti-shock effect [4 - 61.
The idea of an opioid-antiopioid homeostatic system is shared by others [7 - 111 and has been repeatedly proposed with ample supporting evidence by our group [12-181. It is corroborated by many cogent and largely consonant ana- tomical, neurochemical and functional data indicating that melanopeptides and opioid peptides influence autonomic, behavioral, sensory, endocrine, locomotor, gastrointestinal and immune functions in an usually mutually antagonistic, or at least complementary, highly integrated and versatile fashion (for review see Ref. 31.
Thus, when it became apparent that during shock there is a massive activation of opioid systems, and that opioid antagonists reverse hypotension and extend survival in several different shock conditions [lS-271, we decided to investigate the effect of melanopeptides on some typical hemodynamic para- meters and on animal survival in an experimental model of hemorrhagic shock. Here we describe the main results so far obtained.
THE ANTI-SHOCK EFFECT OF MELANOPEPTIDES
Our experimental procedures have been repeatedly described in detail elsewhere [4-6, 28 -321. Briefly, the animals, previously anesthetized with ethylurethane (1.25 g/kg, i.p.1 and heparinized (heparin sodium, 600 I.U./kg, i.v.l, are stepwise bled to a Wiggers type of hemorrhagic shock without reinfusion (oligemic shock) [33-351 until their mean arterial blood pressure stabilizes at 18- 25 mmHg, and then are treated at various intervals.
Mean arterial pressure (MAP), pulse pressure (PP), heart rate (HR), respiratory rate (RR) and survival are continuously recorded for at least 2 h after treatment. In some cases, shocked animals have been sutured, allowed to recover from anesthesia, and then kept under observation up to the cut-off time of 15 days after treatment in order to determine the survival time.
Blood volume, complete blood count, blood gases, blood flow in selected vascular beds, influence of adrenalectomy, influence of splenectomy or of suprahepatic-vein ligature, characteristics of brain, spleen capsule and heart ventricle adrenoceptors have also been studied.
ACTH, alpha-MSH, and other ACTH-fragments have no [5] or scant [36] cardiovascular and respiratory effects in the intact animal.
On the other hand, in our extremely severe, pre-terminal condition of bleeding-induced hypovolemic shock, where all saline-treated animals die within 25 - 30 min, and where typical pressor agents (vasopressin, angiotensin
255
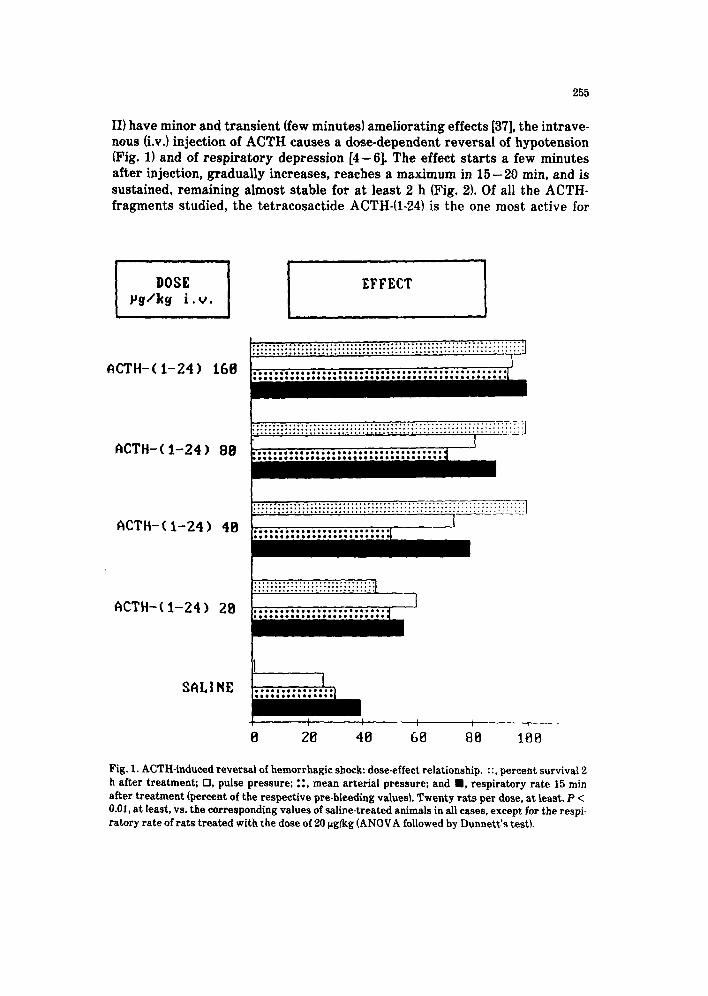
II) have minor and transient (few minutes) ameliorating effects [37], the intrave- nous Lv.) injection of ACTH causes a dose-dependent reversal of hypotension (Fig. 1) and of respiratory depression [4-61. The effect starts a few minutes after injection, gradually increases, reaches a maximum in 15-20 min, and is sustained, remaining almost stable for at least 2 h (Fig. 2). Of all the ACTH- fragments studied, the tetracosactide ACTH-(l-24) is the one most active for
DOSE Yg/kg i.u.
ACTH-( l-24) 160
.................................. ................................
...................................................................
.............................. .......... ........................... I
1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ..*.........*....*...~...............
ACTH-(l-24) 88
ACTH-( l-24) 48
ACTH-(l-24) 28
SALI NE
I 1 . . . . ..*................... .,..,...,,,...............
e 28 40 68 88 108
Fig. 1. ACTH-induced reversal of hemorrhagic shock: dose-effect relationship. ::, percent survival 2 h after treatment; ??, pulse pressure; ::, mean arterial pressure; and ??, respiratory rate 15 min after treatment (percent of the respective pre-bleeding values). Twenty rats per dose, at least. P < 0.01, at least, vs. the corresponding values of saline-treated animals in all cases, except for the respi- ratory rate of rats treated with the dose of 20 pg/kg (ANOVA followed by Dunnett’s test).
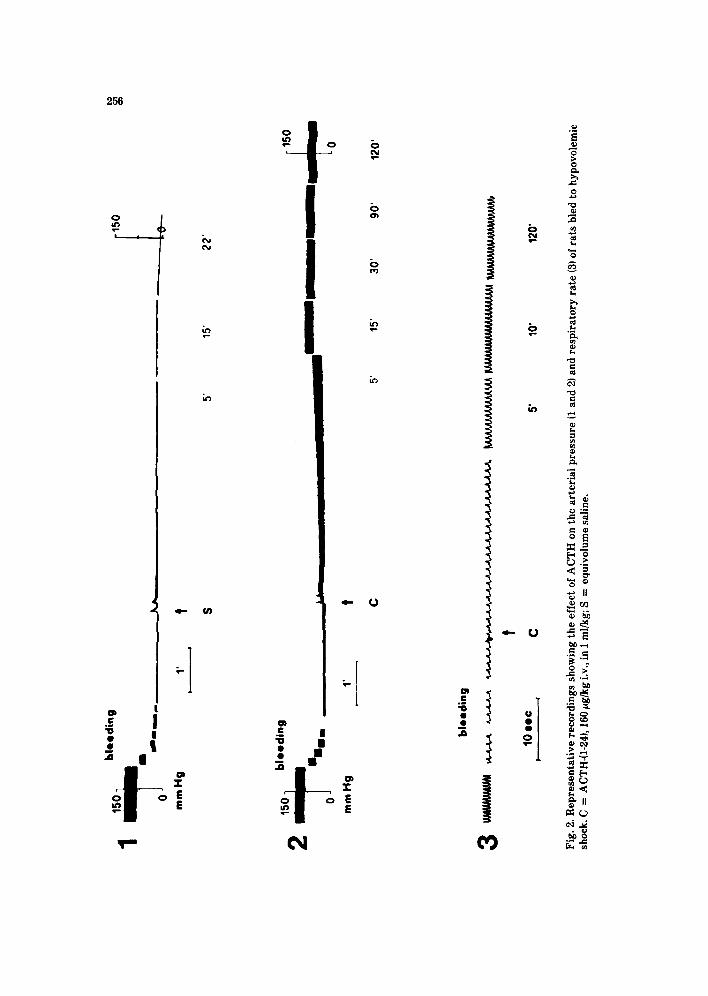
1 15
0
--(l
r AJ
h ---
I+ m
m H
g 1'
t
I I
S
5’
15’
22’
2 15
0
b 0-
n 4-
0
mm
Hg
1’
t C
5’
15’
30’
90’
120’
bloo
ding
10 SO
C
, 4
C
5 10
’ 12
0’
Fig.
2. R
epre
sent
ativ
e re
cord
ings
sho
win
g th
e ef
fect
of A
CTH
on
the
arte
rial
pre
ssur
e (1
and
2) a
nd re
spir
ator
y ra
te (3
) of r
ats
bled
to
hypo
vole
mic
sh
ock.
C =
AC
TH-(l
-24)
,160
rg/
kg i.
v., i
n 1
ml/k
g; S
= e
quiv
olum
e sa
line.
257
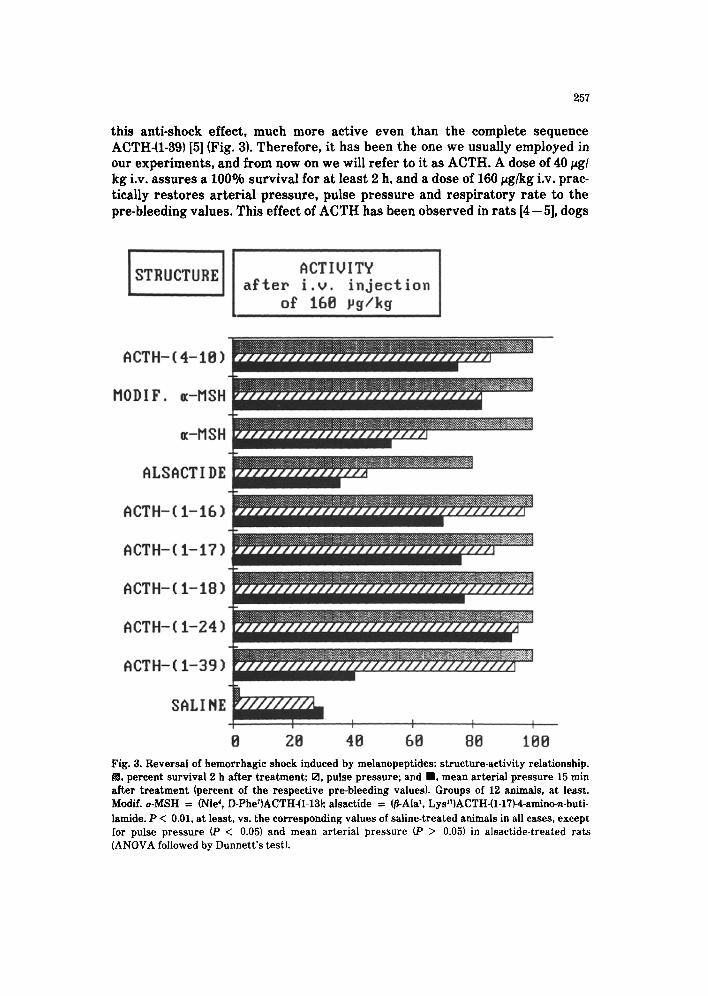
this anti-shock effect, much more active even than the complete sequence ACTH-(l-39) [5] (Fig. 3). Therefore, it has been the one we usually employed in our experiments, and from now on we will refer to it as ACTH. A dose of 40 pg/ kg i.v. assures a 100% survival for at least 2 h, and a dose of 160 rg/kg i.v. prac- tically restores arterial pressure, pulse pressure and respiratory rate to the pre-bleeding values. This effect of ACTH has been observed in rats [4 - 51, dogs
ACTH-(4-lti)
IIODIF. a-MSH
ALSACTIDE
ACTH-(l-16)
ACTH-(l-171
ACTH-(l-18)
ACTH-(l-24)
ACTH-Cl-391
SALINE
e 28 48 68 88 108 Fig. 3. Reversal of hemorrhagic shock induced by melanopeptides: structure-activity relationship. R, percent survival 2 h after treatment; 0, pulse pressure; and m, mean arterial pressure 15 min after treatment (percent of the respective pre-bleeding values). Groups of 12 animals, at least. Modif. a-MSH = (Nle’, D-PherIACTH-(l-13); alsactide = (3-Alai. Lys17)ACTH_(1-17Wamino+buti- lamide. P < 0.01, at least, vs. the corresponding values of saline-treated animals in all cases, except for pulse pressure (P < 0.05) and mean arterial pressure (P > 0.05) in alsactide-treated rats (ANOVA followed by Dunnett’s test).
258
[S] and, so far anecdotically, in humans [38,39]. It is quite remarkable that such an effect, particularly on pulse amplitude, is obtained and maintained without the need for the infusion of blood or blood substitutes.
The time-lapse between bleeding and treatment is obviously of crucial importance (Table I); when ACTH is injected 5 min after bleeding termination, animals survive for approximately 2 days (44.17 f 17.84 h), without any other treatment; when the time-lapse is 15 min, the survival time is 2.79 f 1.07 h; and when animals are treated 25 min after bleeding, i.e. almost at the point of death, ACTH no longer has any effect. A similar trend is observed with the reinfusion of the withdrawn blood. If reinfusion is performed 5 min after bleeding, shock is reversed, survival lasting 307 f 52.80 h with 75% of the animals still alive 15 days after treatment; but when blood is reinfused 15 min after bleeding, the survival time is 6.57 2 4.38 h, and when reinfusion is made 25 min after bleed- ing it no longer has any effect at all.
However, and most important, when animals have been treated with ACTH
TABLE I
IMPORTANCE OF THE TIME-LAPSE BETWEEN SHOCK AND TREATMENT Effect of saline, ACTH, blood reinfusion, and ACTH followed by blood reinfusion at different inter- vals on the survival time of rats bled to hypovolemic shock (8- 12 rats per group). ACTH, ACTH-U- 241, 160 pg&g; Blood, reinfusion of the withdrawn blood. *P < 0.01 compared with saline-treated rats (ANOVA followed by Scbeffgs test). **P < 0.01 and *+*P < 0.05 compared with saline-treated rats (Fisher’s exact probability test).
Treatment, i.v.
Saline, equivolume to the withdrawn blood, 5 min after bleeding
ACTH, 5 min after bleeding ACTH, 15 min after bleeding ACTH, 25 min after bleeding Blood, 5 min after bleeding Blood, 15 min after bleeding Blood, 25 min after bleeding ACTH, 5 min after bleeding
+ Blood, 30 min later ACTH, 6 min after bleeding
+ Blood, 60 min later ACTH, 5 min after bleeding
+ Blood, 120 min later ACTH, 5 min after bleeding
+ Blood, 300 min later
Survival time % Survival (h,m f S.E.) 15 days after treatment
0.47 zt 0.06 0 44.17 f 17.84* 0
2.79 f 1.07+ 0 0.18 f 0.03 0
> 307.20 f 52.80* 75*+ 6.57 2 4.38+ 0 0.17 f 0.03 0
> 185.75 f 100.64* 50*+
> 122.59 f 46.68* 33***
> 157.25 f 57.34+ 36***
> 92.50 * 53.87* 12

within 5- 10 min after bleeding, blood reinfusion retains its effectiveness and reverses shock even if performed 2 - 5 h later.
The anti-shock effect of ACTH can be obtained also by intra- cerebroventricular (i.c.v.1 injection, and is quantitatively and qualitatively quite similar to that observed after i.v. injection [40].
MECHANISM OF THE ANTI-SHOCK EFFECT OF MELANOPEPTIDES
Adrenal glands are not involved, because the anti-shock effect is practically the same in intact and in adrenalectomized animals, and can be obtained also with ACTH-fragments and fragment analogs almost completely devoid of corti- cotropic activity: ACTH-(l-131 (a-MSH), the a-MSH analog (Nle*, D-Phe?-ACTH- (l-131, ACTH-(4-101[5].
The restoration of arterial pressure and pulse amplitude is associated with an impressive increase in the volume of circulating blood [29]. Such increase is not due to hemodilution: the number of red cells per volume unit is in fact the same as in the controls, and the sole significant difference is leukocytosis [29]. Capillary pooling and trapping of blood (“capillary paralysis”) [33], prolonged transit time, with consequent decreased venous return, and reduced cardiac output characterize the terminal, irreversible phase of shock [41].
The increase in circulating blood volume induced by ACTH in shocked ani- mals seems to be the consequence of a mobilization of the peripherally pooled, “out of currency” blood. Indeed, the anti-shock effect of ACTH is greatly impaired in splenectomized animals [29] and in animals subjected to ligature of the suprahepatic veins [42] and is associated with a restoration of the venous blood flow in peripheral vascular beds, and with a normalization of venous PO,.
Morphine (2.5 mglkg, i.v., before starting bleeding) completely prevents the reversal of hemorrhagic shock induced by ACTH (160 pg/kg, i.v.l; however, this effect of morphine can be overcome by increasing the amount of ACTH [S]. This finding strongly supports the view that the anti-shock effect of pharmacological doses of ACTH is indeed the consequence of a functional antagonism towards endogenous opioids, which are massively released in such a condition [26,27]; the further increase of opioid tone caused by the injection of morphine requir- ing higher doses of ACTH.
As concerns the role of the autonomic nervous system, the so far available experimental data indicate that the ACTH-induced reversal of hemorrhagic shock is a complex phenomenon which involves afferent vagal fibers, a central cholinergic link and the activation of the sympathetic nervous system. Indeed, it is almost completely absent after bilateral vagotomy [28], it is prevented by atropine sulphate (hut not by atropine methylbromide) [28] and by the i.c.v. injection of hemicholinium-3, and it is completely prevented by guanethidine, reserpine and clonidine [32,43]. Moreover, it is mediated to a large extent through the activation of peripheral alpha-l- and alpha-2-adrenoceptors; it is in fact antagonized by dibenamine, prazosin and yohimbine, but not by practolol [32,43].

260
Finally, the anti-shock effect of ACTH is associated with a restoration of heart ventricle and spleen capsule adrenoceptors, the number of which is signif- icantly reduced in our experimental model of hemorrhagic shock [31].
DISCUSSION
There is increasing evidence to support the view that endogenous opioids, as well as several other neuropeptides [44,45], play a significant role in the regula- tion of the cardiovascular and respiratory functions (for review see Refs. 25 and 46) . In the CNS, chemoreceptor sensitivity at the bulbar level 1471, baroreflex activity at the level of the nucleus tractus solitarii [48], as well as somatosym- pathetic reflex at the level of the medulla oblongata [49] are influenced both by opioids and by their specific antagonists.
At the peripheral level, it has long been known that morphine and other opioids usually dilate resistance and capacitance vessels; characteristically, they produce venodilatation and move significant quantities of blood from the cen- tral to the peripheral circulation, with pooling of blood in peripheral, “storage” organs (“medical phlebotomy”) [25,50].
In shock conditions there is a massive activation of endogenous opioid systems and a sharp increase in circulating opioid peptides [26,27,51]; in particular, it has been shown that the plasma concentration of beta-endorphin rises to four times the basal level immediately after severe hemorrhage [52].
That opioid mechanisms are implicated in the pathophysiology of shock is further strongly suggested by the finding that opioid antagonists consistently raise blood pressure, increase cardiac output, and extend survival time in sep- tic, hemorrhagic, anaphylactic and neurogenic models of shock in different ani- mal species including primates [19,20,24,25,27] and, albeit not univocally [53 - 551, in humans as well [22,23,56]. However, recent animal and human data seem to question the assumption that the mechanism of action of naloxone in reversing the hemodynamic changes of shock is simply due to the blockade of opioid receptors, and make reference to a positive inotropic effect unrelated to opioid receptors. Thus, Sagy et al. have observed a positive inotropic response on iso- lated rat myocardium using ( + 1 naloxone, the non-opioid blocking isomer [57]. and the antiarrhythmic effect of naloxone [58] can similarly be obtained with the ( + ) stereoisomer as well [59]. Others have suggested that naloxone poten- tiates the effects of circulating catecholamines on the myocardium, resulting in improved contractility [60].
Finally, Roberts et al. have now reported in human cases of septic shock the unexpected finding of positive hemodynamic effects occurring only more than 4 h after the initial bolus of naloxone and the start of a continuous infusion of the drug; this is clearly not consistent with a displacement of endogenous opioids from their receptors, which should have produced a rapid effect [61].
Recent experimental data have further stressed and better defined the role of opioids in circulatory homeostasis. Schadt and coworkers [34,62] have shown, in conscious rabbits, that while rapid removal of approximately 15% of the

261
total blood volume produced a sharp increase in plasma noradrenaline with maintenance of arterial pressure at control levels (non-hypotensive, compen- sated hemorrhage), a further bleeding of another 30% of the total volume pro- duces a sudden fall in arterial pressure (hypotensive hemorrhage) and is not accompanied by any further increase in plasma noradrenaline. In the latter conditions, the injection of naloxone causes a prompt increase in plasma norad- renaline and arterial pressure, suggesting that during hemorrhagic hypoten- sion there is an opiate-mediated inhibition of sympathetic outflow and of noradrenaline release [34]. More recently, Morita et al. have shown that in con- scious rabbits hemorrhage elicits a biphasic effect on renal nerve activity, which rises initially during non-hypotensive hemorrhage and then falls during hypotensive hemorrhage, this decrease being reversed by naloxone [63].
Sandor et al. have recently shown that the bleeding volume required to induce hemorrhagic hypotension in rats is markedly reduced by the subcutaneous administration of morphine or by the i.c.v. injection of beta- endorphin, whereas it is significantly increased by the subcutaneous administration of naloxone or by the i.c.v. infusion of beta-endorphin antiserum 1641.
It is now well established that adrenergic neurons residing in a highly restricted zone (Cl area) of the rostra1 ventrolateral medulla play a critical role in the tonic and reflex regulation of arterial pressure and of respiratory func- tion [65 - 681. Abundant pharmacological evidence demonstrates that these adrenergic neurons are tonically regulated by inhibitory gabaergic and opioid- ergic inputs and by excitatory cholinergic mechanisms [69,70].
Opioids inhibit the release of several neurotransmitters [10,71,72]. They inhibit noradrenergic and dopaminergic neurotransmission in the CNS [lo] and, in particular, inhibit the firing of locus coeruleus neurons [73], while naloxone enhances it [74]. It has long been known that a number of opioid actions in the CNS and in the periphery involve the inhibition of acetylcholine release (for review see Ref. 75); in particular, in the brain, opioid administration stereospe- cifically decreases the amount of liberated acetylcholine and the basal release of acetylcholine into lateral ventricular perfusates [76 - 781, an the i.c.v. admin- istration of beta-endorphin in analgesic amounts causes a decrease in the turnover of acetylcholine in different brain areas, this effect being prevented by naltrexone [79]. So, opioids significantly depress, or abolish [80], the hyper- tensive response to a central cholinergic activation [81], most probably by inhi- biting acetylcholine release in the involved structures. Also while the injection of opioids into the rostra1 ventrolateral medulla produces cardiovascular and respiratory depression [69], the microinjection of cholinomimetic drugs in this same area reverses opioid-induced respiratory depression [82].
Conversely, melanopeptides increase noradrenaline synthesis and turnover in whole brain, as well as in more circumscribed areas (brain stem) of rats and mice (83,841 and selectively modulate restricted catecholaminergic neuronal systems [85]. In particular, ACTH increases by 97% the noradrenaline content of locus coeruleus [86], and the implantation of ACTH-(I-10) into the locus coeru-

262
leus increases noradrenaline release [87]. Moreover, melanopeptides increase acetylcholine turnover in some brain areas [10,88] and stimulate sympatho- adrenomedullary activity at the peripheral level. Indeed, ACTH increases the activity of catecholamine-synthesizing enzymes, in particular that of phenylethanolamine-N-methyltransferase in the adrenal medulla [89]; and it increases the action potential-evoked release of noradrenaline from the post- ganglionic sympathetic terminals innervating the pulmonary artery of the rab- bit, apparently by an effect at presynaptic ACTH receptors [go]. ACTH also raises the plasma noradrenaline level in vivo when injected into the right atrium of anesthetized rats [91], and potentiates the inotropic response of rat atria1 strip to noradrenaline [92].
Finally, Starke and coworkers have recently shown that the i.v. infusion of ACTH-(l-241(0.03- 1 pg/kg per mini increases the noradrenaline spillover rate and the plasma noradrenaline concentration in pithed rabbits with electrically stimulated sympathetic outflow [93].
Our present data, showing that melanopeptides are able to reverse an otherwise rapidly and invariably irreversible condition of hemorrhagic shock; that such effect involves the activation of a central choline@ pathway and occurs through an adequately restored responsivity of the sympathetic system; that it is associated with a mobilization of the peripherally pooled blood; and, finally, that it is antagonized by morphine, clearly suggest that such shock rev- ersal induced by melanopeptides is due to their functional antagonism towards endogenous opioids, with opposite effects at all levels and on all structures involved in cardiovascular homeostasis.
This further corroborates and strengthens the idea of an opioid-antiopioid homeostatic system and suggests that irreversible shock is associated with, and probably to a large extent due to, a derangement of this balanced system resulting in a massive prevalence of the opioid component, which can only be neutralized through the administration of pharmacological doses of antiopioids.
CONCLUSION
Our data lead to both conceptual and practical conclusions. From a practical point of view, also on the basis of the preliminary human
data so far available, an unforeseen, simple and safe first-aid management of the hemodynamic and respiratory consequences of massive blood losses is envisaged. In fact, ACTH and the other ACTH-fragments active in reversing shock conditions are practically devoid of acute toxicity [94], and have no other pharmacological effect, even corticotropic activity being absent in the shortest effective fragments. It therefore seems possible to restore the hemodynamic and respiratory parameters to a level compatible with survival and to postpone by hours the infusion of blood without compromising its efficacy. The outstand- ing importance of such a possibility is self-evident.
From a conceptual point of view, our data demonstrate that (i) melanopeptides, which under normal conditions are almost inactive on

263
cardiovascular function, have an extremely potent effect on blood mobilization, blood flow and arterial pressure in conditions of severe cardiovascular failure, such effect occurring through the activation of a complex mechanism that involves peripheral and central components; (ii) as a corollary, they further stress that pharmacological “activity” is a very relative concept and also depends, among many other things, on the functional condition of the consid- ered system: the case of melanopeptides is an extreme one, because they are inactive under normal conditions and extremely active and potent under path- ological conditions; (iii) our data also further stress that neuropeptides usually subserve multiple purposes in the organism, and that their sequence encodes many different messages, so that the traditional terminology is often mislead- ing or too limiting; (iv) finally, a “life-saving” mechanism may have been disclosed which, at least in mammals, is vestigial or anyhow functions only up to a limited degree and then fails, but which can be restored in a pharmacological way. Perhaps it is not out of place to remind that melanopeptides play a well known central role in the response of the organism to emergency situations, and that in some species they do in fact subserve the life-saving function of camouflage.
REFERENCES
1
2
3
4
5
10
11
12
A. Bertolini, R. Poggioli and W. Ferrari, ACTH-induced hyperalgesia in rats, Experientia. 35 (197911216- 1217. A. Bertolini and W. Ferrari, Evidence and implications of a melanocortins-endorphins homeo- static system. In: Neuropeptides and psychosomatic process. Editor: E. Endroczi, Akademiai Kiado, Budapest, 1982, pp. 245- 261. A. Bertolini, R. Poggioli, A.V. Vergoni, M. Castelli and S. Guarini, Evidence that melanocortins are physiological antagonists of opioids. In: Central actions of ACTH and related peptides. Edi- tors: D. de Wied and W. Ferrari, Liviana Press, Padova- Springer Verlag, Berlin, 1986, pp. 207 - 222. A. Bertolini, S. Guarini and W. Ferrari, Adrenal-independent anti-shock effect of ACTH-(l-24) in rats, Eur. J. Pharmacol., 122 (1986) 387 -388. A. Bertolini, S. Guarini, E. Rompianesi and W. Ferrari, a-MSH and other ACTH fragments improve cardiovascular function and survival in experimental hemorrhagic shock, Eur. J. Pharmacol.. 130 (1986119 - 26. A. Bertolini, S. Guarini, W. Ferrari and E. Rompianesi, Adrenocorticotropin reversal of experi- mental hemorrhagic shock is antagonized by morphine, Life Sci., 39 (198611271- 1280. W.H. Gispen. J.M. van Ree and D. de Wied, Lipotropin and the central nervous system, Int. Rev. Neurobiol., 20 (19771201-250. Y.F. Jacquet, Opiate effects after adrenocorticotropin or /I-endorphin injection in the peri aqueductal gray matter of rats, Science, 201(197811032- 1034. T.L. O’Donohue and D.M. Dorsa. The opiomelanotropinergic neuronal and endrocrine systems, Peptides, 3 (19821353 - 395. D. de Wied and J. Jolles, Neuropeptides derived from pro-opiocortin: behavioral, physiological and neurochemical effects, Physiol. Rev., 62 119821976- 1059. C.A. Hendrie, Opiate dependence and withdrawal-a new synthesis? Pharmacol. Biochem. Behav., 23 (19851863 - 870. A. Bertolini, R. Poggioli and W. Fratta, Withdrawal symptoms in morphine-dependent rats intracerebroventricularly injected with ACTH-(l- 24) and with 6-MSH, Life Sci., 29 (19811249 -252.

13
14
15
16
17
18
19
20
21
22 23
24
25
26
27
28
29
30
31
32
33 34
35
36
A. Bertolin& W. Fratta, M. Melis and G.L. Gessa, Possible role of ACTH-MSH peptides in mor- phine tolerance and withdrawal in rats, Adv. Biosci.. 48 (1984) 225-230. M. Castelli, G.L. Gessa and A. Bertolini, ACTH-(l-24) antagonizes the contractile effect of mor- phine on the isolated rat colon, Eur. J. Pharmacol., 108 (1985) 213-214. A.V. Vergoni. Ft. Poggioli and A. Bertolini, Corticotropin inhibits food intake in rats, Neuro- peptides, 7 (1986) 15% 158. R. Poggioli, A.V. Vergoni and A. Bertolini, ACTH-(l-24) and a-MSH antagonize feeding behav- ior stimulated by kappa opiate agonists, Peptides, 7 (1986) 843 - 848. A. Bertolini, A.V. Vergoni, R. Poggioli and G.L. Gessa, Morphine and /3-endorphin antagonize posture and locomotor disorders induced by the injection of ACTH-(l-24) in the rat locus coe- ruleus, Life Sci., 38 (1986) 373-377. R. Poggioii, R. Arletti, A.V. Vergoni, M. Castelli and A. Bertolini, ACTH-(l- 24) antagonizes the cholestatic and constipating effects of morphine, Arch. Int. Pharmacodyn., 293 (1988) 265- 272. J.W. Holaday and A.I. Faden, Naloxone reversal of endotoxic hypotension suggests role of endorphins in shock, Nature, 275 (1978) 450-451. A.I. Faden, and J.W. Holaday, Opiate antagonists: a role in the treatment of hypovolemic shock, Science, 205 (1979) 317-318. J.W. Holaday and A.I. Faden, Naloxone acts at central opiate receptors to reverse hypoten- sion, hypotermia and hypoventilation in spinal shock, Brain Res., 189 (1980) 295- 299. M. Tiengo, Naloxone in irreversible shock, Lancet, i (1980) 690. W.P. Peters, P.A. Friedman, M.W. Johnson and W.E. Mitch, Pressor effect of naloxone in sep- tic shock, Lancet, i (1981) 529-532. N.J. Guril, D.G. Reynolds, T. Vargish and R. Lechner, Naloxone without transfusion prolongs survival and enhances cardiovascular function in hypovoiemic shock, J. Pharmacol. Exp. Ther., 220 (1982) 621- 624. J.W. Holaday, Cardiovascular effects of endogenous opiate systems, Annu. Rev. Pharmacol. Toxieol., 23 (1983) 541- 594. R. Elam, F. Bergmann and G. Feuerstein, Simultaneous changes of catecholamines and Leu- enkephaline-like immunoreactivity in plasma and cerebrospinal fluid of cats undergoing acute hemorrhage, Brain Res., 303 (1984) 313-317. E.W. Bernton, J.B. Long and J.W. Holaday, Opioids and neuropeptides: mechanisms in circula- tory shock, Fed. Proc., 44 (1985) 290- 299. S. Guarini. E. Rompianesi, W. Ferrari and A. Bertoiini, Influence of vagotomy and of atropine on the anti-shock effect of adrenocorticotropin, Neuropeptides, 8 (1986) 19-24. S. Guarini, W. Ferrari, G. Mottillo and A. Bertolini, Anti-shock effect of ACTH: haematological changes and influence of splenectomy, Arch. Int. Pharmacodyn., 289 (1987) 311-318. S. Guarini, A. Bertolini, N. Lancellotti, E. Rompianesi and W. Ferrari, Different cholinergic pathways are involved in the improvement induced by CCK8 and by ACTH-(l-24) in massive acute haemorrhage, in rats, Pharmacol. Res. Commun., 19 (1987) 511- 516. M. Sandrini, S. Guarini and A. Bertolini, Influence of the intravenous administration of ACTH- (l-24) on the characteristics of brain, heart and spleen adrenoceptors of haemorrhage-shocked rats, Pharmacol. Res. Commun., 20 (1988) 739 - 749. S. Guarini, W. Ferrari and A. Bertolini, Involvement of the sympathetic nervous system in the cardiovascular effects of ACTH-(l-24) during hemorrhagic shock in rats, Naunyn-Schmiede- berg’s Arch. Pharmacol., 337 (1988) 556-560. C.J. Wiggers. Physiology of shock, Commonwealth Fund, New York, 1950, p. 459. J.C. Schadt and R.R. Gaddis, Endogenous opiate peptides may limit norepinephrine release during hemorrhage, J. Pharmacol. Exp. Ther., 232 (1985) 656-660. J. Barroso-Aranda, G.W. Schmid-Schonbein, B.W. Zweifach and R.L. Engler, Granulocytes and no-reflow phenomenon in irreversible hemorrhagic shock, Circ. Res., 63 (1988) 437 - 447. I. Iyoda, H. Takahashi, L.C. Lee, S. Kambara, H. Okajima, S. Sasaki, K. Takeda, M. Yoshimura and H. Ijichi. Centrally induced cardiovascular and sympathetic responses to adrenocorti- cotrophic hormone, Cardiovasc. Res., 20 (1986) 778- 782.

265
37
38
39
40
41 42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
68
59
60
S. Guarini, S. Tagliavini. W. Ferrari and A. Bertolini, Reversal of hemorrhagic shock in rats by cholinomimetic drugs, Br. J. Pharmacol. (19891 in press. A. Bert&d, Nuove prospettive per il trattamento dello shock: gli antagonisti endogeni degli oppioidi, Clin. Ter., 119 (19861365-367. A. Bert&r& S. Gu&ni, W. Ferrari, G. Noera, C. Massini and S. Di Tizio, ACTH-G-241 restores blood pressure in acute hypovolaemia and haemorrhagic shock in humans, Eur. J. Clin. Phar- macol., 32 (19871637 - 638. S. Guarb& A.V. Vergoni and A. Bertolini, Anti-shock effect of ACTH-(l-241: comparison between intracerebroventricular and intravenous route of administration, Pharmacol. Res. Commun., 19 (19871256-260. H.D. Green, Physiology of peripheral circulation in shock, Fed. Proc. 20. suppl. 9 (1961) 61- 68. S. Guarini, W. Ferrari and A. Bertolini, Anti-shock effect of ACTH-U-241: influence of subtotal hepatectomy, Pharmacol. Res. Commun., 20 (19881395- 403. A. Bertolini, S. Guarini, M. Sandrini and W. Ferrari, ACTHeatecholamines interactions in the haemorrhagic shock. In: New Perspectives in Pharmacological Sciences. Editors: F. Drago and J.M. Van Ree, It. Pharmacol. Sot., 1987, pp. 59-61. D. Ganten, M. Printz. M.I. Phillips and B.A. Sch61 Kens, The Renin-Angiotensin System in the Brain. Experimental Brain Research, Springer-Verlag, Berlin, 1982, suppl. 4. M. Palkovits, Neuropeptides and biogenic amines in central cardiovascular control mecha- nisms. In: Perspectives in Cardiovascular Research. Editors: J.P. Buckley and C.M. Ferrario, Vol. 6, Raven Press. New York, 1981, pp. 73 - 87. J.L. Reid and P.C. Rubin, Peptides and central neural regulation of the circulation, Physiol. Rev., 67 (19871725- 749. M. Denavit-Saubie, J. Champagnat and W. Zielgansberger, Effects of opiates and methionine- enkephalin on pontine and bulbar respiratory neurones of cat, Brain Res., 155 (1978156 - 67. M.A. Petty and W. de Jong, Cardiovascular effects of /I-endorphin after microinjection into the nucleus tractus solitarii of the anesthetized rat, Eur. J. Pharmacol., 81(1982) 449-457. D. Sandor. W. de Jong and D. de Wied, Endorphinergic mechanisms in cerebral blood flow autoregulation, Brain Res., 386 (19861122 - 129. R.P. Henney. J.S. Vasko. R.K. Brawley, H.N. Oldham and A.G. Morrow, The effect of morphine on the resistance and capacitance vessels of the peripheral circulation, Am. Heart J., 72 (1966) 242-246. L.D. Farrell, T.S. Harrison and L.M. Demers, Immunoreactive met-enkephalin in the canine adrenal; response to acute hypovolemic stress, Proc. Sot. Exp. Biol. Med., 173 (1983) 515-518. R.E. Lang, U.B. Bruckner, K. Hermann, B. Kempf, W. Rascher, V. Sturm, Th. Unger and D. Ganten, Effect of hemorrhagic shock on the concomitant release of endorphin and enkephalin- like peptides from the pituitary and adrenal gland in the dog, Adv. Biochem. Psychopharma- col., 33 (19821363 - 368. J.J. Montaatruc, J.P. Richard and B. Cathala, Naloxone fails to reverse blood pressure in shock: a double-blind study in man, J. Pharmacol., 16 (1985) 313-314. P. Rock, H. Silverman and D. Plump, Efficacy and safety of naloxone in septic shock, Crit. Care Med., 13 (1985128-33. A. De Maria, JJ. Heffeman, G.A. Grindlinger. D.E. Craven, T.K. McIntosh and W.R. McCabe, Naloxone versus placebo in treatment of septic shock, Lancet, i (1985) 1363- 1365. J.S. Groeger. G.W. Carlon and W.S. Howland, Naloxone in septic shock, Crit. Care Med., 11 (19831650 - 654. M. Sagy, G. Shavit, Y. Oron, B.A. Vidne, S. Gitter and Y. Sarne, Nonopiate effect of naloxone on cardiac muscle contractility, J. Cardiovasc. Pharmacol., 9 (19871682-685. 0. Fagbemi, I. Lepran, J.R. Parroti and L. Szekeres, Naloxone inhibits early arrhythmias resulting from acute coronary ligation, Br. J. Pharmacol., 76 (19821504 - 506. Y. Same, I. Hochman, M. Eshed and E. Oppenheimer, Antiarrhythmic action of naloxone: direct, nonopiate effect on the rat heart, Life Sci., 43 (1988) 859- 864. R.B. Lechner, N.J. Gurll and D.G. Reynolds, Naloxone potentiates the cardiovascular effects of catecholamines in canine hemorrhagic shock, Circ. Shock, 16 (1985) 347- 361.

266
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
D.E. Roberts, K.W. Hall, K.E. Dobson and R.B. Light, Effects of prolonged naloxone infusion in septic shock, Lancet, ii (19881699-702. J.C. Schadt, M.D. McKnown, D.P. McKnown and D. Franklin, Hemodynamic effects of hemor- rhage and subsequent naloxone treatment in conscious rabbits, Am. J. Physiol., 247 (1984) R497 - R508. H. Morita, Y. Nishida, H. Motochigawa, N. Uemura, H. Hosomi and S.F. Vatner, Opiate recep- tor-mediated decrease in renal nerve activity during hypotensive hemorrhage in conscious rabbits, Circ. Res., 63 (19881165- 172. P. Sandor, W. de Jong, V. Wiegant and D. de Wied, Central opioid mechanisms and cardiovas- cular control in hemorrhagic hypotension, Am. J. Physiol., 253 (1987) H507-H511. R.A.L. Dampney, Brain stem mechanisms in the control of arterial pressure, Clin. Exp. Hyper- tension, 3 (19811379 - 391. M. Kalia, Anatomical organization of central respiratory neurons, Annu. Rev. Physiol.. 43 (19811105- 120. D.J. Reis, The brain and hypertension: reflections on 35 years of inquiry into the neurobiology of the circulation, Circulation, 70, suppl. III (1984131- 45. C.A. Ross, D.A. Ruggiero, T.H. Joh, D.H. Park and D.J. Reis, Rostra1 ventrolateral medulla: selective projections to the thoracic autonomic cell column from the region containing Cl adrenaline neurons. J. Comp. Neurol., 228 (19841168 - 184. S. Punnen, R.N. Willette, A.J. Krieger and H.N. Sapru, Cardiovascular response to injections of enkephalin in the pressor area of the ventrolateral medulla, Neuropharmacology, 23 (19841 939 - 946. D.J. Reis, Neurotransmitters acting in the Cl area in the tonic and reflex control of blood pres- sure, J. Cardiovasc. Pharmacol. 10, suppl. 12 (19871 S22 - 525. H.D. Taube, E. Borowski, T. Endo and K. Starke, Enkephalin: a potential modulator of norad- renaline release in rat brain, Eur. J. Pharmacol., 38 (19761377-380. D.H.G. Versteeg, Interaction of peptides related to ACTH, MSH and gLPH with neurotrans- mitters in the brain, Pharmacol. Ther., 11(19801535-557. J. Korf, B.S. Bunney and G.K. Aghajanian, Noradrenergic neurons: morphine inhibition of spontaneous activity, Eur. J. Pharmacol., 25 (19741165- 169. H.K. Strahlendorf, J.C. Strahlendorf and CD. Barnes, Endorphin-mediated inhibition of locus coeruleus neurons. Brain Res., 19lG9801284 - 288. M. Weinstock, Acetylcholine and cholinesterase. In: Narcotic Drugs: Biochemical Pharmacol- ogy. Editor: D.H. Clouet, Plenum Press, New York, 1971, pp. 254-260. D. Beleslin and R.L. Polak, Depression by morphine and chloralose of acetylcholine release from the cat’s brain, J. Physiol., 177 (19651411-419. K. Jhamandas, J. Sawynok and M. Sutak, Enkephalin effects on release of brain acetylcholine, Nature, 269 (19771433- 434. J. Crossland and K.Z. Ahmed, Brain acetylcholine during morphine withdrawal, Neurochem. Res., 9 (19841351-366. F. Moroni, D.L. Cheney and E. Costa, fl-endorphin inhibits Ach turnover in nuclei of rat brain, Nature, 267 (19771267-268. V.M. Varagic and V. Stojanovic, The effect of enkephalins and of /3-endorphin on the hyperten- sive response to physostigmine in the rat, Br. J. Pharmacol., 92 (19871197-202. H.E. Brezenoff and R. Giuliano, Cardiovascular control by cholinergic mechanism in the central nervous system, Annu. Rev. Pharmacol. Toxicol., 22 (19821341-381. R.N. Willette, B.M. Doorley and H.N. Sapru, Activation of cholinergic mechanisms in the med- ulla oblongata reverse intravenous opioid-induced respiratory depression, J. Pharmacol. Exp. Ther., 240 (19871352-358. D.H.G. Versteeg. Effect of two ACTH-analogs on noradrenaline metabolism in rat brain, Brain Res., 49 (19731483-485. A.J. Dunn and N.R. Kramarcy. Neurochemical responses in stress: relationship between the hypothalamic-pituitary-adrenal and catecholamine system. In: Drugs Neurotransmitters and

267
85
86
87
88
89
90
91
92
93
94
Behavior. Editors: L.L. Iversen, SD. Iversen and S.H. Snyder, Handb. Psychopharmacol., Vol. 18, Plenum Press, New York and London, 1984. pp. 455- 515. D.H.G. Versteeg, ACTHMSH-related neuropeptides as modulators of central neurotransmis- sion. In: New Perspectives in Pharmacological Sciences. Editors: F. Drago and J.M. van Ree, The Italian Pharmacol. See.. 198’7, pp. 56-58. M. Fekete, J. Herman, M. Palkovits and E. Stark, ACTH-induced changes in the transmitter- amine concentration of individual brain nuclei of the rat. In: Catecholamines and Stress. Edi- tors: E. Usdin, R. Kvetnansky and I.J. Kopin, Pergamon Press, New York, 19’76, pp. 69-75. E. Endrijczi, Effect of ACTH,,, and ACTH,_,, on the distribution of BH-noradrenaline in the brain of adrenalectomized rats, Acta Physiol. Acad. Sci. Hung., 48 (1976159-64. P.L. Wood, D. Malthe-SGrenssen, D.L. Cheney and E. Costa, Increase of hippocampal acetyl- choline turnover rate and the stretching-yawning syndrome elicited by alpha-MSH and ACTH, Life Sci., 22 (19781673 - 678. J. Axelrod, Catecholamines: effects of ACTH and adrenal corticoids, Ann. N.Y. Acad. Sci., 297 (19771275 - 279. M. Giithert and F. Hentrich, Role of CAMP for regulation of impulse-evoked noradrenaline release from the rabbit pulmonary artery and its possible relationship to presynaptic ACTH receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol., 328 (19841127 - 134. T. Okajima, S. Ikuyama, K. Kato and H. Ibayashi, Naloxone inhibits the plasma epinephrine response to ACTH but not to 2deoxy-D-glucose in rats, Life Sci., 35 (1984) 2177 - 2181. J.R. Bassett. F.L. Strand and K.D. Cairncross, Glucocorticoids, adrenocorticotropic hormone and related polypeptides on myocardial sensitivity to noradrenaline, Eur. J. Pharmacol., 49 (19781243- 247. B. Szabo, L. Hedler, K. Lichtwald and K. Starke, ACTH increases noradrenaline release in pithed rabbits with electrically stimulated sympathetic outflow, Eur. J. Pharmacol., 136 (1987) 391- 399. W. Mandel, M.J. Singer, H.R. Cudmunson, L. Meister and F.W.S. Modern, Intravenous use of pituitary adrenocorticotropic hormone (ACTH). A report on its administration in twenty-five patients, J. Am. Med. Assoc., 146 (1951) 546-551.




















