
The activity of antioxidant defence enzymes in the mussel Mytilus galloprovincialis from the...
9
The activity of antioxidant defence enzymes in the mussel Mytilus galloprovincialis from the Adriatic Sea Slavica S. Borkovic ´ a , Jelena S. S ˇ aponjic ´ a , Sladjan Z. Pavlovic ´ a , Duxko P. Blagojevic ´ a , Slavixa M. Miloxevic ´ a , Tijana B. Kova*evic ´ a , Ratko M. Radoji*ic ´ b , Mihajlo B. Spasic ´ a , Radoslav V. Z ˇ ikic ´ c , Zorica S. Sai*ic ´ a, * a Department of Physiology, Institute for Biological Research ‘‘Sinixa Stankovic ´’’, Bulevar despota Stefana 142, 11060 Belgrade, Serbia and Montenegro b Institute of Biochemistry and Physiology, Faculty of Biology, University of Belgrade, Studentski Trg 16, 11000 Belgrade, Serbia and Montenegro c Institute of Biology and Ecology, Faculty of Sciences, University of Kragujevac, Radoja Domanovic ´a 12, Kragujevac, Serbia and Montenegro Received 16 November 2004; received in revised form 2 August 2005; accepted 2 August 2005 Available online 15 September 2005 Abstract The activity of the antioxidant defence enzymes superoxide dismutase (SOD, EC 1.15.1.1), catalase (CAT, EC 1.11.1.6), glutathione peroxidase (GSH-Px, EC 1.11.1.9), glutathione reductase (GR, EC 1.6.4.2) and the phase II biotransformation enzyme glutathione-S- transferase (GST, EC 2.5.1.18) in whole mussels (Mytilus galloprovincialis ) were studied. The mussels were collected in winter and in spring at two localities in the Adriatic Sea: Bar Port and Tivat Bay. Our results show that the activities of SOD, GSH-Px and GST were seasonally dependent with higher activities in winter. GR activity was also higher in winter, but only in mussels from Bar Port. In mussels from Tivat Bay, GR activity was lower in winter compared to spring. In addition, a decrease in CATactivity in mussels from Bar Port compared to those from Tivat Bay was found. It can be concluded that seasonal variations should be incorporated into interpretation of biomonitoring studies in mussels. D 2005 Published by Elsevier Inc. Keywords: Adriatic Sea; Superoxide dismutase; Catalase; Glutathione peroxidase; Glutathione reductase; Glutathione-S-transferase; Mussels; Mytilus galloprovincialis 1. Introduction All aerobic organisms during their respiratory activity continuously produce reactive oxygen species (ROS) (Pe ´rez- Campo et al., 1993; Halliwell and Gutteridge, 1999). The important sites of ROS production are mitochondria (Boveris and Cadenas, 1982; Chance et al., 1979; Turrens et al., 1985), microsomes (Staats et al., 1988), peroxisomes (Dhaunsi et al., 1992) and the cytosol (Shaw and Jayatilleke, 1990). ROS induce many cellular disturbances, such as depolymerization of polysacharides and nucleic acids, oxidation of protein sulphydryl groups and peroxidation of fatty acids (Stohs et al., 2000). According to the classical concept of antioxidant defence, this system includes enzymatic (superoxide dismutase—SOD, catalase—CAT, glutathione peroxidase—GSH-Px, glutathione reductase— GR and glutathione-S -transferase—GST) and nonenzymatic components (Cadenas, 1989). This view may be modified by the notion of complex antioxidant defence (Niki, 2000) and/or the concept of gradual antioxidant defence (Van der Oost et al., 2003). According to the Environmental Risk Assessment (ERA), the components of antioxidant defence are functionally divided into biotransformation phase II components (for instance, GST and reduced/oxidized glutathione) and oxidative stress parameters (SOD, CAT, GSH-Px and GR) (Van der Oost et al., 2003). Bivalve mollusks such as Mytilus galloprovincialis are commonly used as bioindicators in ERA. These mussels 1532-0456/$ - see front matter D 2005 Published by Elsevier Inc. doi:10.1016/j.cbpc.2005.08.001 * Corresponding author. Tel.: +381 11 2078 325; fax: +381 11 2761 433. E-mail address: [email protected] (Z.S. Sai*ic ´). Comparative Biochemistry and Physiology, Part C 141 (2005) 366 – 374 www.elsevier.com/locate/cbpc
Transcript of The activity of antioxidant defence enzymes in the mussel Mytilus galloprovincialis from the...

www.elsevier.com/locate/cbpc
Comparative Biochemistry and Physiol
The activity of antioxidant defence enzymes in the mussel
Mytilus galloprovincialis from the Adriatic Sea
Slavica S. Borkovic a, Jelena S. Saponjic a, Sladjan Z. Pavlovic a, Duxko P. Blagojevic a,
Slavixa M. Miloxevic a, Tijana B. Kova*evic a, Ratko M. Radoji*ic b, Mihajlo B. Spasic a,
Radoslav V. Zikic c, Zorica S. Sai*ic a,*
a Department of Physiology, Institute for Biological Research ‘‘Sinixa Stankovic’’, Bulevar despota Stefana 142, 11060 Belgrade, Serbia and Montenegrob Institute of Biochemistry and Physiology, Faculty of Biology, University of Belgrade, Studentski Trg 16, 11000 Belgrade, Serbia and Montenegroc Institute of Biology and Ecology, Faculty of Sciences, University of Kragujevac, Radoja Domanovica 12, Kragujevac, Serbia and Montenegro
Received 16 November 2004; received in revised form 2 August 2005; accepted 2 August 2005
Available online 15 September 2005
Abstract
The activity of the antioxidant defence enzymes superoxide dismutase (SOD, EC 1.15.1.1), catalase (CAT, EC 1.11.1.6), glutathione
peroxidase (GSH-Px, EC 1.11.1.9), glutathione reductase (GR, EC 1.6.4.2) and the phase II biotransformation enzyme glutathione-S-
transferase (GST, EC 2.5.1.18) in whole mussels (Mytilus galloprovincialis) were studied. The mussels were collected in winter and in spring
at two localities in the Adriatic Sea: Bar Port and Tivat Bay. Our results show that the activities of SOD, GSH-Px and GST were seasonally
dependent with higher activities in winter. GR activity was also higher in winter, but only in mussels from Bar Port. In mussels from Tivat
Bay, GR activity was lower in winter compared to spring. In addition, a decrease in CAT activity in mussels from Bar Port compared to those
from Tivat Bay was found. It can be concluded that seasonal variations should be incorporated into interpretation of biomonitoring studies in
mussels.
D 2005 Published by Elsevier Inc.
Keywords: Adriatic Sea; Superoxide dismutase; Catalase; Glutathione peroxidase; Glutathione reductase; Glutathione-S-transferase; Mussels; Mytilus
galloprovincialis
1. Introduction
All aerobic organisms during their respiratory activity
continuously produce reactive oxygen species (ROS) (Perez-
Campo et al., 1993; Halliwell and Gutteridge, 1999). The
important sites of ROS production are mitochondria
(Boveris and Cadenas, 1982; Chance et al., 1979; Turrens
et al., 1985), microsomes (Staats et al., 1988), peroxisomes
(Dhaunsi et al., 1992) and the cytosol (Shaw and Jayatilleke,
1990). ROS induce many cellular disturbances, such as
depolymerization of polysacharides and nucleic acids,
oxidation of protein sulphydryl groups and peroxidation of
1532-0456/$ - see front matter D 2005 Published by Elsevier Inc.
doi:10.1016/j.cbpc.2005.08.001
* Corresponding author. Tel.: +381 11 2078 325; fax: +381 11 2761 433.
E-mail address: [email protected] (Z.S. Sai*ic).
fatty acids (Stohs et al., 2000). According to the classical
concept of antioxidant defence, this system includes
enzymatic (superoxide dismutase—SOD, catalase—CAT,
glutathione peroxidase—GSH-Px, glutathione reductase—
GR and glutathione-S-transferase—GST) and nonenzymatic
components (Cadenas, 1989). This view may be modified
by the notion of complex antioxidant defence (Niki, 2000)
and/or the concept of gradual antioxidant defence (Van der
Oost et al., 2003). According to the Environmental Risk
Assessment (ERA), the components of antioxidant defence
are functionally divided into biotransformation phase II
components (for instance, GST and reduced/oxidized
glutathione) and oxidative stress parameters (SOD, CAT,
GSH-Px and GR) (Van der Oost et al., 2003).
Bivalve mollusks such as Mytilus galloprovincialis are
commonly used as bioindicators in ERA. These mussels
ogy, Part C 141 (2005) 366 – 374

S.S. Borkovic et al. / Comparative Biochemistry and Physiology, Part C 141 (2005) 366–374 367
are inter-tidal filter-feeding invertebrates known to accu-
mulate high levels of trace metals and organic com-
pounds in their tissues, providing a time-integrated
indication of environmental contamination with observ-
able cellular and physiological responses (Lau and Wong,
2003). The responses in mussels make them good
bioindicators for environmental monitoring (Livingstone,
1993). They have a number of properties which make
them useful sentinels for chemical pollution: they have a
wide geographical distribution, are easy to collect and are
abundant in estuarine waters, which are submitted to high
contamination levels (Manduzio et al., 2004). Moreover,
mussels are sedentary, euryhaline and normally the
dominant species in their habitats (Sheehan and Power,
1999).
As mussels are thermocomforming organisms, they must
deal with oscillations in the environmental temperature and
their metabolic rate and consequently with oscillations in the
levels of ROS (Wilhelm Filho et al., 1993). As a
consequence, ROS generation, oxidation rates and antiox-
idant status are most likely related to the ambient temper-
ature and the metabolic activity (Wilhelm Filho et al., 2000),
as we have previously shown for fish (Pavlovic et al., 2004)
and rodents (Spasic et al., 1993).
Fig. 1. The geographical position of Bar Po
An important factor influencing musselVs biochemistry
and physiology is seasonality. Depending on the availability
of nutrients, reproductive status, growth rate related with
season and other factors, the activity of antioxidant defence
enzymes and other biomarkers fluctuate significantly
throughout the year (Sheehan and Power, 1999). Seasonal
variations in antioxidant defences were observed in tissues
of horse mussels (Modiolus modiolus) (Lesser and Kruse,
2004), blue mussels (Mytilus edulis) (Manduzio et al., 2004)
and in the digestive gland of brown mussels (Perna perna)
(Wilhelm Filho et al., 2001).
It is also known that several classes of pollutants are
capable of enhancing the formation of ROS and thereby
provoke oxidative stress. Some of these pollutants include
polychlorinated biphenyls (PCBs), polycyclic aromatic
hydrocarbons (PAHs), phenols and heavy metals (Van der
Oost et al., 2003). Many studies have shown positive
correlations between levels of antioxidant defences and the
presence of xenobiotics (Orbea et al., 2002). Oxidative
stress is also a highly seasonal phenomenon in bivalve
mollusks (Lesser and Kruse, 2004). The oxidative stress
modulation by environmental pollutants is a factor which
might complicate the interpretation of biomonitoring studies
(Sheehan and Power, 1999).
rt and Trivat Bay in the Adriatic Sea.

Table 1
Values of salinity (�), temperature (-C) and oxygen concentration (mg/L)
at 0.5 m depth in Bar Port and Tivat Bay on the day of mussel collection
during winter and spring
Bar Port Tivat Bay
winter spring winter spring
Temperature (-C) 8.6 19.9 12.0 20.1
O2 (mg/L) 9.6 7.6 8.2 7.8
Salinity (�) 32.2 36.5 33.0 34.3
S.S. Borkovic et al. / Comparative Biochemistry and Physiology, Part C 141 (2005) 366–374368
The studied areas of Bar Port and Tivat Bay were
selected because both receive extensive industrial and
urban waste water discharges. These areas have similar
climates, and the lowest mean water temperature occurs in
February and highest in August. The mean water
temperature in February is 12.5 -C and in May is 20.4
-C. Tivat Bay is characterized by a higher inflow of
120
2
4
6
8
10
12
14
******
Winter
(A)
ANOVA comparison: (S): p<0.001 (L): N.S. SxL: N.S.
Spring
Bar PortTivat Bay
450
100
200
300
400
*** #
***
ANOVA comparison: (S): p<0.001(L): p<0.01SxL: N.S.
(B)
SpringWinter
Bar Port Tivat Bay
U/g
wet
mas
sU
/mg
prot
ein
Fig. 2. The specific (A) and total (B) activities of SOD in mussels from Bar
Port (n =20) and Tivat Bay (n =13) in winter and spring. Results are
expressed as meanTSE. The statistical significance was analyzed by two-
way ANOVA test with season (S) and locality (L) as factors. N.S.=not
significant. A minimal confidence interval of 5% ( p <0.05) was considered.
freshwater than Bar Port. The mean sea depth in Bar Port
is 10 m and Tivat bay is 25 m. The bottoms of the
biotopes are covered with thick stratum of fine terri-
genous mud containing particles of detritus. The sea
currents in both localities are very irregular, in the
summer they are slight, while in winter they are very
strong (Stjep*evic, 1974).
The aim of this study was to compare the activity of
antioxidant defence enzymes in mussels between two
different localities, one with intensive industrial pollution
(Bar Port), and second with intensive anthropogenic
pollution (Tivat Bay) in two seasons. The activity of
antioxidant defence enzymes, superoxide dismutase, cata-
lase, glutathione peroxidase and glutathione reductase and
the activity of glutathione-S-transferase was measured in
musselMytilus galloprovincialis collected from the Bar Port
and Tivat Bay from the Adriatic Sea in winter and late
spring.
890
2
4
6
8
10
12
*
#
(A)
ANOVA comparison: (S): p<0.05 (L): p<0.01 SxL: N.S.
SpringWinter
Bar Port Tivat Bay
11 12
µmol
H2O
2/m
in/g
wet
mas
s
50
100
150
200
250
300
#
#
ANOVA comparison: (S): N.S.(L): p<0.01SxL: N.S.
(B)
SpringWinter
Bar Port Tivat Bay
µmol
H2O
2/m
in/m
g pr
otei
n
0
Fig. 3. The specific (A) and total (B) activities of CAT in mussels from
Bar Port (n =20) and Tivat Bay (n =13) in winter and spring. The number
of samples and type of statistical analysis was identical to that indicated in
Fig. 2.

29 300
5
10
15
20
25
30
*
*
ANOVA comparison: (S): N.S.(L): N.S.SxL: p<0.05
SpringWinter
(A)
Bar Port Tivat Bay
600
700
800
ANOVA comparison: (S): N.S.(L): N.S.SxL: p<0.05
(B)
Bar Port Tivat Bay
t mas
snm
ol N
AD
PH/m
in/m
g pr
otei
n
S.S. Borkovic et al. / Comparative Biochemistry and Physiology, Part C 141 (2005) 366–374 369
2. Materials and methods
2.1. Site description
Mussels (Mytilus galloprovincialis) were collected in
winter (February) and late spring (May) at two localities:
Bar Port (42- 05V 58� N and 19- 05V 40� E) and Tivat Bay
(42- 24V 16� N and 18- 43V 23� E) (Fig. 1).
The two sites were chosen to compare the activity of
antioxidant defence enzymes between periods of lower
metabolic activity (winter) and higher metabolic activity
(spring). At the time of sampling environmental parameters
including water temperature, oxygen concentrations and
salinity were measured (Table 1).
2.2. Sample collection and preparation
At the Bar Port, 20 (10 in winter and 10 in spring)
specimens were collected, whereas in Tivat Bay 13 (7 in
winter and 6 in spring) specimens were collected. Mussels
from Bar Port were larger (4.5–5 cm shell length) than
15 160
2
4
6
8
10
12
******
ANOVA comparison: (S): p<0.001(L): N.S.SxL: N.S.
(A)
SpringWinter
Bar Port Tivat Bay
190
100
200
300
400
******
(B)
SpringWinter
ANOVA comparison: (S): p<0.001(L): N.S.SxL: N.S.
Bar Port Tivat Bay
nmol
NA
DPH
/min
/mg
prot
ein
nmol
NA
DPH
/min
/g w
et m
ass
Fig. 4. The specific (A) and total (B) activities of GSH-Px in mussels from
Bar Port (n =20) and Tivat Bay (n =13) in winter and spring.
32 330
100
200
300
400
500*
SpringWinter
nmol
NA
DPH
/min
/g w
e
Fig. 5. The specific (A) and total (B) activities of GR in mussels from Bar
Port (n =20) and Tivat Bay (n =13) in winter and spring.
those from Tivat Bay (2.5–3 cm). The latter has a higher
inflow of freshwater that enables higher organic production
and abundant appearance of young mussels (Stjep*evic,
1974). After collecting, mussels were frozen in liquid
nitrogen and then stored at �70 -C. In each group, total
soft tissue from the whole body was dissected from all
collected mussels. The tissues were ground and homogen-
ized in 5 vol. (Lionetto et al., 2003) of 25 mmol/L sucrose
containing 10 mmol/L Tris–HCl, pH 7.5 at 4 -C with an
Ultra-Turrax homogenizer (Janke and Kunkel, IKA-Werk,
Staufen, Germany) (Rossi et al., 1983). The homogenates
were sonicated for 30s at 10 kHz on ice to release enzymes
(Takada et al., 1982) and sonicates were then centrifuged at
4 -C at 100,000 g for 90 min. The resulting supernatants
were used for biochemical analyses.
2.3. Biochemical analyses
Total protein concentration in the supernatant was
determined according to the method of Lowry et al.
(1951) and expressed in mg/g wet mass. The activity of
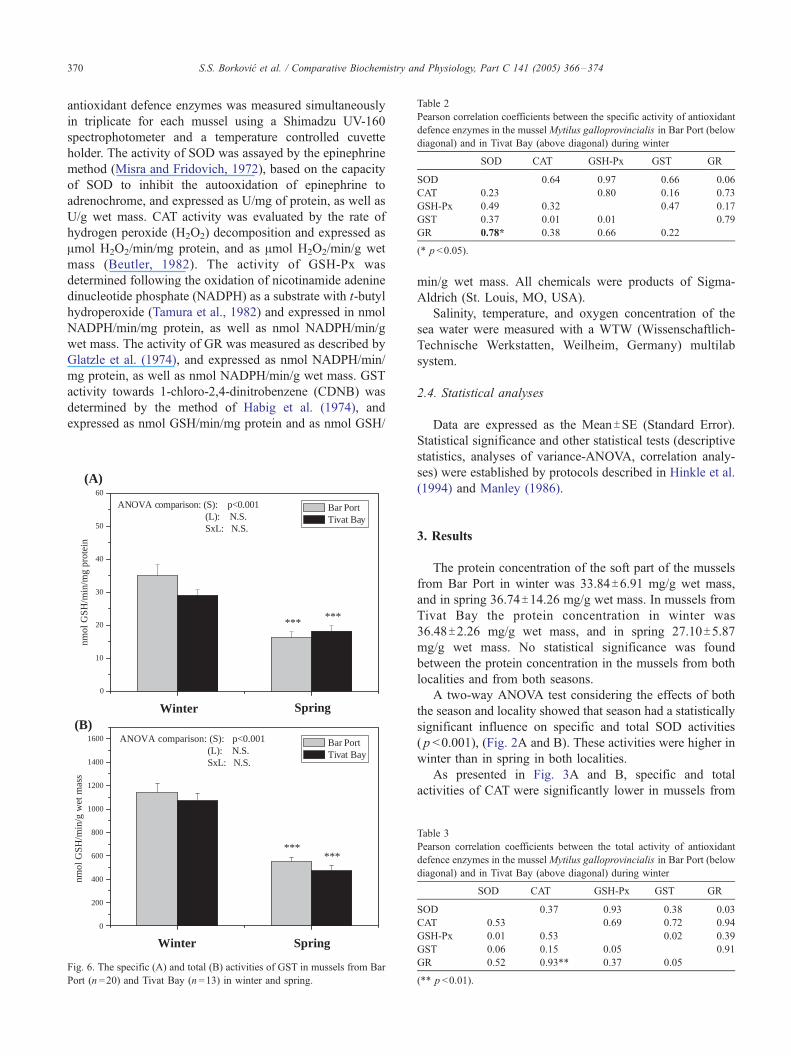
Table 2
Pearson correlation coefficients between the specific activity of antioxidant
defence enzymes in the musselMytilus galloprovincialis in Bar Port (below
diagonal) and in Tivat Bay (above diagonal) during winter
SOD CAT GSH-Px GST GR
SOD 0.64 0.97 0.66 0.06
CAT 0.23 0.80 �0.16 �0.73
GSH-Px 0.49 �0.32 0.47 �0.17
GST 0.37 0.01 �0.01 0.79
GR 0.78* �0.38 0.66 0.22
(* p <0.05).
S.S. Borkovic et al. / Comparative Biochemistry and Physiology, Part C 141 (2005) 366–374370
antioxidant defence enzymes was measured simultaneously
in triplicate for each mussel using a Shimadzu UV-160
spectrophotometer and a temperature controlled cuvette
holder. The activity of SOD was assayed by the epinephrine
method (Misra and Fridovich, 1972), based on the capacity
of SOD to inhibit the autooxidation of epinephrine to
adrenochrome, and expressed as U/mg of protein, as well as
U/g wet mass. CAT activity was evaluated by the rate of
hydrogen peroxide (H2O2) decomposition and expressed as
Amol H2O2/min/mg protein, and as Amol H2O2/min/g wet
mass (Beutler, 1982). The activity of GSH-Px was
determined following the oxidation of nicotinamide adenine
dinucleotide phosphate (NADPH) as a substrate with t-butyl
hydroperoxide (Tamura et al., 1982) and expressed in nmol
NADPH/min/mg protein, as well as nmol NADPH/min/g
wet mass. The activity of GR was measured as described by
Glatzle et al. (1974), and expressed as nmol NADPH/min/
mg protein, as well as nmol NADPH/min/g wet mass. GST
activity towards 1-chloro-2,4-dinitrobenzene (CDNB) was
determined by the method of Habig et al. (1974), and
expressed as nmol GSH/min/mg protein and as nmol GSH/
22 230
10
20
30
40
50
60
******
ANOVA comparison: (S): p<0.001(L): N.S.SxL: N.S.
(A)
SpringWinter
Bar Port Tivat Bay
25 260
200
400
600
800
1000
1200
1400
1600
******
ANOVA comparison: (S): p<0.001 (L): N.S. SxL: N.S.
SpringWinter
(B) Bar Port Tivat Bay
nmol
GSH
/min
/g w
et m
ass
nmol
GSH
/min
/mg
prot
ein
Fig. 6. The specific (A) and total (B) activities of GST in mussels from Bar
Port (n =20) and Tivat Bay (n =13) in winter and spring.
min/g wet mass. All chemicals were products of Sigma-
Aldrich (St. Louis, MO, USA).
Salinity, temperature, and oxygen concentration of the
sea water were measured with a WTW (Wissenschaftlich-
Technische Werkstatten, Weilheim, Germany) multilab
system.
2.4. Statistical analyses
Data are expressed as the MeanTSE (Standard Error).
Statistical significance and other statistical tests (descriptive
statistics, analyses of variance-ANOVA, correlation analy-
ses) were established by protocols described in Hinkle et al.
(1994) and Manley (1986).
3. Results
The protein concentration of the soft part of the mussels
from Bar Port in winter was 33.84T6.91 mg/g wet mass,
and in spring 36.74T14.26 mg/g wet mass. In mussels from
Tivat Bay the protein concentration in winter was
36.48T2.26 mg/g wet mass, and in spring 27.10T5.87mg/g wet mass. No statistical significance was found
between the protein concentration in the mussels from both
localities and from both seasons.
A two-way ANOVA test considering the effects of both
the season and locality showed that season had a statistically
significant influence on specific and total SOD activities
( p <0.001), (Fig. 2A and B). These activities were higher in
winter than in spring in both localities.
As presented in Fig. 3A and B, specific and total
activities of CAT were significantly lower in mussels from
Table 3
Pearson correlation coefficients between the total activity of antioxidant
defence enzymes in the musselMytilus galloprovincialis in Bar Port (below
diagonal) and in Tivat Bay (above diagonal) during winter
SOD CAT GSH-Px GST GR
SOD 0.37 0.93 0.38 �0.03
CAT �0.53 0.69 �0.72 �0.94
GSH-Px 0.01 �0.53 0.02 �0.39
GST �0.06 0.15 �0.05 0.91
GR 0.52 �0.93** 0.37 0.05
(** p <0.01).

Table 5
Pearson correlation coefficients between the total activity of antioxidant
defence enzymes in the musselMytilus galloprovincialis in Bar Port (below
diagonal) and in Tivat Bay (above diagonal) in spring
SOD CAT GSH-Px GST GR
SOD 0.09 0.17 �0.001 �0.04
CAT �0.30 0.45 0.59 0.15
GSH-Px 0.13 �0.01 0.80** 0.25
GST 0.32 �0.14 0.53 0.17
GR 0.06 �0.47 0.22 0.22
(** p <0.01).
S.S. Borkovic et al. / Comparative Biochemistry and Physiology, Part C 141 (2005) 366–374 371
Bar Port than from Tivat Bay in both seasons ( p <0.01). A
two-way ANOVA test considering the effects of both the
season and locality showed that the season had a statistically
significant influence only on the specific activity of CAT
( p <0.05), which was lower in winter than in spring in
mussels sampled at Tivat Bay.
The specific and total activities of GSH-Px (Fig. 4A and
B) were significantly higher in winter in both localities
( p <0.001). However, there were no significant differences
in the specific and total GSH-Px activities between Bar Port
and Tivat Bay in both seasons.
GR activity was higher in winter compared to spring in
mussels collected from Bar Port (Fig. 5A and B), while in
mussels from Tivat Bay, only the specific GR activity was
lower in winter compared to spring ( p <0.05).
As shown in Fig. 6A and B, the specific and total
activities of the phase II biotransformation enzyme GST
were significantly higher in winter than in spring in both
localities ( p <0.001), and no statistically significant differ-
ences between both localities were observed in both
seasons. Correlation analyses are statistical tools to explore
functional connections between antioxidant components on
a statistical base (Blagojevic et al., 1998; Pavlovic et al.,
2004; Jovanovic-Galovic et al., 2004; Speers-Roesch and
Ballantyne, 2005). Degree of correlation is expressed as
Pearson correlation coefficient (determines the extent to
which values of the two variables are ‘‘proportional’’ to each
other) and its statistical significance based on the assump-
tion that the distribution of the residual values (i.e., the
deviations from the regression line) for the dependent
variable follows the normal distribution, and that the
variability of the residual values is the same for all values
of the independent variable ( p <0.05 is considered as
significant). The results from such analyses from data
presented in this present paper are in Tables 2–5. We
concluded that there were no significant correlations in
winter in mussels from Tivat Bay, contrary to mussels from
Bar Port. In mussels from Bar Port collected during winter, a
positive correlation between SOD and GR and a negative
one between CAT and GST activities were detected. In
spring, several correlations were noticed in mussels from
Tivat Bay (positive between GST and GSH-Px, CAT and
GST, SOD and GR, as well as CAT and GST), and negative
between SOD and GR in mussels from Bar Port. This
Table 4
Pearson correlation coefficients between the specific activity of antioxidant
defence enzymes in the musselMytilus galloprovincialis in Bar Port (below
diagonal) and in Tivat Bay (above diagonal) in spring
SOD CAT GSH-Px GST GR
SOD 0.50 0.12 0.41 0.77**
CAT 0.38 0.51 0.68* 0.44
GSH-Px 0.12 0.47 0.71* 0.42
GST 0.61 0.73* 0.41 0.59
GR �0.71* 0.10 0.32 �0.19
(** p <0.01, * p <0.05).
suggests that basic homeostatic relationships between the
antioxidant defence enzymes in mussels from Bar Port were
different than those from Tivat Bay, depending on the period
they were collected.
4. Discussion
The seasonal pattern of antioxidant defence enzymes
found in M. galloprovincialis appears to be closely
correlated with the seasonal variations of temperature and
the reproductive cycle. The activity of antioxidant enzymes
and antioxidant compounds (vitamin E and reduced
glutathione) are known to be under extensive seasonal
control (Sheehan and Power, 1999). Lesser and Kruse
(2004) have shown that the mussel Modiolus modiolus
seasonally compensates for decreases in temperature by
increasing the concentration of rate-limiting metabolic
enzymes while maintaining the same level of antioxidant
protection in summer and winter. The increase in temper-
ature is followed by an increase in oxygen consumption
and by an increase in ROS generation. The second season-
related determining factor in the level of various antiox-
idants in mussels seems to be the metabolic particularity of
the stages of their reproductive cycle. The above results
and conclusions reinforce the importance of seasonality on
the antioxidant status of Bivalvia in relation to the
interpretation of biomonitoring data (Wilhelm Filho et
al., 2001). In our experiments both populations of mussels
expressed differences in the antioxidant defence enzymes
regarding winter and spring (data from correlation analysis,
Tables 2–5). Antioxidant defence enzymes are constitutive
in nature and susceptible to variations due to intrinsic
biological processes and cycles and ontogenic processes,
beside the extrinsic influences, such as environmental
changes and contaminants. The interpretation of these
responses, in an environmental context, is very complex to
account for all the possible causes (Sheehan and Power,
1999). When organisms are employed as monitoring
subjects, particularly for routine long-term monitoring
purposes and in areas with contaminants at sub-lethal
concentrations, the intrinsic biological variations such as
size, tissue specificity, and natural variations of the
biochemical responses, such as food availability and

S.S. Borkovic et al. / Comparative Biochemistry and Physiology, Part C 141 (2005) 366–374372
environmental changes become key factors to be consid-
ered (Lau and Wong, 2003).
It is important that besides physiological and environ-
mental factors, the contribution of ontogenetic development
is also a determining factor of antioxidant levels in mussels
(Rudneva, 1999). This is in accordance with the fact that
physiological functions in mussels are influenced by size
(Sukhotin et al., 2002). Studies regarding the effect of aging
and size on the activity of CAT in mussels have given rise to
different results, as this effect appears to be specific to
species, sex, tissue and age. Several authors found a decrease
in CAT activity in the course of ageing in the mussel Mytilus
edulis (Viarengo et al., 1991), the marine shrimp Aristeus
antennatus (Mourente and Diaz-Salvago, 1999), as well as in
cephalopods Sepia officinalis and Lolliguncula brevis
(Zielinski and Portner, 2000). In contrary, Sukhotin et al.
(2002) found a weight-specific decrease in CAT activity
within the separate age classes of M. edulis. This finding
illustrates that in those studies where age-related decrease of
CAT has been reported, it might be due to the increasing size
of the ageing mussels and not due to the effect of age per se.
Our investigations showed an increase in specific and total
activity of CAT in M. galloprovincialis collected during
May, which is probably linked to the increased metabolic
activity related to the seasonal temperature elevations and
intense reproductive activity that occurs in spring. Further-
more, in the Mediterranean Sea, the winter period is
characterized by gametogenesis, whereas spring is charac-
terized by intensive egg release. Summer is characterized by
the deposition of lipids and glucose, whereas the autumn by
the beginning of gametogenesis (Stjep*evic, 1974). This is in
accordance with the findings of other authors which
postulated that changes in the reproductive cycle are
connected with the season and the rate of metabolic activity
(Robledo et al., 1995).
The activities of SOD, GSH-Px and GST showed
elevated levels in winter. It was suggested that SOD and
GSH constituted the integral components of the cellular
antioxidant defence. Namely, reduced glutathione (GSH) is
a physiologically useful scavenger if the reaction with
superoxide is prevented (Munday and Winterbourn, 1989).
Elevated SOD activity was accompanied with elevated
GSH-Px activity and positive correlation with GR activity
suggests coordinated action toward possible hydrogen
peroxide induced oxidative attack. The study of Orbea et
al. (2002) showed a higher antioxidant defence enzyme
activities in summer. Seasonal factors might affect bio-
marker responses to a greater extent than pollution
variations. Apart from natural biological cycles, the annual
cycles of climatic conditions may also induce stress in the
organisms thereby triggering antioxidant defences. Cellular
rearrangements of metabolic components to compensate
environmental fluctuations have already been found (Lesser
and Kruse, 2004). The antioxidant defence system might
regulate its activity by rearrangements of components in
spite of changing its activity (Blagojevic et al., 1998). The
recomposition of antioxidant defence enzymes regarding
season was observed by correlation analyses in the present
work. Davenport (1983) stated that the asymmetrical pattern
of ctenidial ciliary activity and other physiological
responses depended on the water temperature and salinity.
The low temperature could reduce the metabolic activities,
and hence lower enzymatic activities in general. One
possible explanation for the higher antioxidant defence
enzyme activities in the winter samples might be the higher
sensitivity to stress induced by chemicals from the sea water
in the winter season (Power and Sheehan, 1996). Salinity is
another major physical factor affecting the physiological
responses such as inhalant and exhalant siphon closure, and
shell valve closure in mussels (Davenport, 1983). The
optimal salinity for the mussel M. galloprovincialis is in the
range of 32–37� (Renzoni, 1962). In our study, the salinity
in the seasons was within the optimal range (in winter 32–
33� and in spring 34–36.5�), which enabled mussels to
maintain normal filtration activity (Table 1). It is known that
the production of phytoplankton, which is the main energy
source for filtering organisms such as the mussel M.
galloprovincialis increases during spring. Reproductive
activity and temperature-associated changes in patterns of
food storage and utilization is also likely to cause changes in
the hormonal and nutritional status which might also affect
the levels of enzyme activity (Orbea et al., 1999).
GST, one of the phase II biotransformation enzyme
systems, has been used as a biomarker of organic industrial
effluents (Sheehan et al., 1995). In addition, GST has been
used as a biomarker of exposure to anthropogenic organics
(Fitzpatrick et al., 1997). Xenobiotic chemicals may be
biotransformed according to the simplified mechanism of
route I, which can be subdivided into phases I, II and III.
Phase I is a non-synthetic alteration (oxidation, reduction or
hydrolysis) of the original foreign molecule, which can then
be conjugated in phase II and catabolised in phase III (Van
der Oost et al., 2003). GST, a phase II biotransformation
enzyme catalyzes the initial step of mercapturic acid
synthesis and a conjugation of GSH with xenobiotics and
their metabolites such as the alkyl transferase and epoxide
transferase, detoxifying PAH epoxide produced by cyt P450
(George, 1994). This enzyme is also the most sensitive
biomarker for the influence of environmental pollution on
the organism. Our results show that the activity of GST was
significantly higher in winter which is in accordance with
higher sensitivity to oxidative stress in this period (Van der
Oost et al., 2003). At the same time, no statistically
significant differences were detected between the two
localities investigated in both seasons, indicating a similar
degree of pollution. Our finding of a positive correlation
between GST and GSH-Px activities could be explained as a
coordinated expression of total GST and its peroxidase-like
isoform. At the same time, there was significant positive
correlation between specific GST and CAT activities in both
localities, suggesting a similar pattern for hydrogen per-
oxide elimination.

S.S. Borkovic et al. / Comparative Biochemistry and Physiology, Part C 141 (2005) 366–374 373
Although many compounds possess antioxidant proper-
ties, cellular antioxidant defence is provided by functional
overlapping of antioxidant enzymes and correlation
between its activities. This means, if cellular antioxidant
defence represents a physiological system, then changes
in the activity of an individual antioxidant component
should be accompanied by subsequent changes in the
activity of others. This can be explored statistically by
correlation analysis, which determinates probability if the
activity of one component is correlated with another.
Comparative analyses of correlations between antioxidant
enzymes in different species, seasons and the environment
have been performed in our previous work (Blagojevic et
al., 1998; Pavlovic et al., 2004; Jovanovic-Galovic et al.,
2004). In the recent paper similar approach was used,
and several positive correlations between measured
antioxidant components in fish were observed (Speers-
Roesch and Ballantyne, 2005) suggesting coordinated
regulation of antioxidant enzymes. It seems that anti-
oxidant components undergo recomposition together with
the changes in its activity to respond different devel-
opmental, seasonal and environmental impact during life
cycle.
Our study represents the first comprehensive report of
antioxidant defence enzyme activities in the mussel M.
galloprovincialis collected from the Montenegrine coast-
line from the Adriatic Sea. The results obtained in this
work indicate a significant influence of seasonal factors on
specific and total activities of SOD, CAT, GSH-Px and
GST and a significant influence of location on CAT
activity. Therefore, it can be concluded that seasonal
factors should be incorporated into interpretation of
mussels-based biomonitoring studies. The relationship
between oxygen consumption and ROS generation also
seems to apply to the antioxidant status of thermocom-
formers, whose oxidative metabolism depends directly on
the environmental temperature, reproductive cycle and
seasonality (Wilhelm Filho et al., 2001). This functional
independence is important for the homeostasis of thermo-
comformers, especially regarding their oxidative metabo-
lism and should also be taken into account for the
interpretation of biomonitoring studies related to aquatic
contamination.
Acknowledgements
This work was funded by the Federal Government of
Serbia and Montenegro, Project title:‘‘Bioindicators of
contamination of the Montenegrine coastline’’, as well as
by the Ministry for Science, Environmental Protection of
Republic of Serbia. The map of the Montenegrine
coastline was kindly provided by the Institute of Marine
Biology, Kotor, Serbia and Montenegro. The authors are
thankful to Dr. David R. Jones for proofreading the
manuscript.
References
Beutler, E., 1982. Catalase. In: Beutler, E. (Ed.), Red Cell Metabo-
lism, A Manual of Biochemical Methods. Grune and Stratton, Inc.,
pp. 105–106.
Blagojevic, D., Buzad”ic, B., Korac, B., Sai*ic, Z.S., Radoji*ic, R., Spasic,
M.B., Petrovic, V.M., 1998. Seasonal changes in the antioxidative
defense in ground squirrels (Citellus citellus): possible role of GSH-Px.
J. Environ. Pathol. Toxicol. Oncol. 17, 241–250.
Boveris, A., Cadenas, E., 1982. In: Oberley, L.W. (Ed.), Production of
Superoxide Radicals and Hydrogen Peroxide in Mitochondria. CRC
Press, Boca Raton, pp. 159–168.
Cadenas, E., 1989. Biochemistry of oxygen toxicity. Ann. Rev. Biochem.
58, 79–110.
Chance, B., Sies, H., Boveris, A., 1979. Hydroperoxide metabolism in
mammalian organs. Physiol. Rev. 59, 527–605.
Davenport, J., 1983. A comparison of some aspects of the behavior and
physiology of the Indian mussel Perna (Mytilus) viridis and the
common mussel Mytilus edulis L. J. Molluscan Stud. 49, 21–26.
Dhaunsi, G.S., Gulati, S., Singh, A.K., Orak, J.K., Asayama, K.,
Singh, I., 1992. Antioxidants attenuate diabetes-induced activation
of peroxisomal functions in the rat kidney. J. Biol. Chem. 267,
6870–6873.
Fitzpatrick, P.J., OVHalloran, J., Sheehan, D., Walsh, A.R., 1997. Assess-
ment of a glutathione S-transferase and related proteins in the gill and
digestive gland of Mytilus edulis (L.) as potential organic pollution
biomarkers. Biomarkers 2, 51–56.
George, S.G., 1994. Enzymology and molecular biology of phase II
xenobiotic-conjugating enzymes. In: Malins, D.C., Ostrander, G.K.
(Eds.), Aquatic Toxicology: Molecular, Biochemical and Cellular
Perspectives. Lewis Publishers, Boca Raton, FL, pp. 37–85.
Glatzle, D., Vulliemuier, J.P., Weber, F., Decker, K., 1974. Glutathione
reductase test with whole blood a convenient procedure for the
assesment of the riboflavin status in humans. Experientia 30, 665–668.
Habig, W.H., Pubst, M.J., Jakoby, W.B., 1974. Glutathione S-transferase.
J. Biol. Chem. 249, 7130–7139.
Halliwell, B., Gutteridge, J.M.C., 1999. Free Radicals in Biology and
Medicine, 3rd edR Oxford University Press, Oxford, p. 980.
Hinkle, E.D., Wiersma, W., Jurs, G.S., 1994. Applied Statistics for
Behavioral Sciences, Third edition Houghton Mifflin Company, Boston.
Jovanovic-Galovic, A., Blagojevic, D.P., Grubor-Lajxic, G., Worland, R.,
Spasic, M.B., 2004. Role of antioxidant defense during different stages
of preadult life cycle in European corn borer (Ostrinia nubilalis,
Hubn.): diapause and metamorphosis. Arch. Insect Biochem. Physiol.
55, 79–89.
Lau, P.S., Wong, H.L., 2003. Effect of size, tissue parts and location on six
biochemical markers in the green-lipped mussel, Perna viridis. Mar.
Pollut. Bull. 46, 1563–1572.
Lesser, M.P., Kruse, V.A., 2004. Seasonal temperature compensation in
the horse mussel, Modiolus modiolus: metabolic enzymes, oxida-
tive stress and heat shock proteins. Comp. Biochem. Physiol. A 137,
495–504.
Lionetto, M.G., Caricato, R., Giordano, M.E., Pascariello, M.F., Marinosci,
L., Schettino, T., 2003. Integrated use of biomarkers (acetylcholinees-
terase and antioxidant enzyme activities) in Mytilus galloprovincialis
and Mullus barbatus in an Italian coastal marine area. Mar. Pollut. Bull.
46, 324–330.
Livingstone, D.R., 1993. Biotechnology and pollution monitoring: use of
molecular biomarkers in the aquatic environment. J. Chem. Technol.
Biotechnol. 57, 195–211.
Lowry, O.H., Rosebrough, N.L., Farr, A.L., Randall, R.I., 1951. Protein
measurement with Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Manduzio, H., Monsinjon, T., Galap, C., Leboulenger, F., Rocher, B., 2004.
Seasonal variations in antioxidant defences in blue mussels Mytilus
edulis collected from a polluted area: major contributions in gills of an
inducible isoform of Cu/Zn-superoxide dismutase and glutathione-S-
transferase. Aquat. Toxicol. 70, 83–93.

S.S. Borkovic et al. / Comparative Biochemistry and Physiology, Part C 141 (2005) 366–374374
Manley, B.F.J., 1986. In: Primer, A. (Ed.), Multivariate Statistical Methods.
Chapman and Hall, London.
Misra, H.P., Fridovich, I., 1972. The role of superoxide anion in the
autooxidation of epinephrine and simple assay for superoxide dismu-
tase. J. Biol. Chem. 247, 3170–3175.
Mourente, G., Diaz-Salvago, E., 1999. Characterization of antioxidant
systems, oxidation status and lipids in brain of wild-caught size-class
distributed Aristeus antennatus (Risso, 1816) Crustacea, Decapoda.
Comp. Biochem. Physiol. B 124, 405–416.
Munday, R., Winterbourn, C.C., 1989. Reduced glutathione in combination
with superoxide dismutase as an important biological antioxidant
defence mechanism. Biochem. Pharmacol. 15, 4349–4352.
Niki, E., 2000. Oxidative stress and ageing. Int. Med. 39, 324–326.
Orbea, A., Marigomez, I., Fernandez, C., Tarazona, J.V., Cancio, I.,
Cajaraville, M.P., 1999. Structure peroxisomes and activity of the
marker enzyme catalase in digestive epithelial cells in relation to PAH
content of mussels from two Basque estuaries (Bay of Biscay):
seasonal and site-specific variations. Arch. Environ. Contam. Toxicol.
36, 158–166.
Orbea, A., Ortiz-Zarragoitia, M., Sole, M., Porte, C., Cajaraville, M., 2002.
Antioxidant enzymes and peroxisome proliferation in relation contam-
inant body burdens of PAHs and PCBs in bivalvia molluscs, crabs and
fish from Urdiabai and Plentzia estuaries (Bay of Biscay). Aquat.
Toxicol. 58, 75–98.
Pavlovic, S.Z., Belic, D., Blagojevic, D.P., Radoji*ic, R.M., Zikic, R.V.,
Sai*ic, Z.S., Lajxic, G.G., Spasic, M.B., 2004. Seasonal variations of
cytosolic antioxidant enzyme activities in liver and white muscle of
thinlip gray mullet (Liza ramada Risso) from the Adriatic Sea. Cryo-
Lett. 25, 273–285.
Perez-Campo, R., Lopez-Torres, M., Rojas, C., Cadenas, S., Barja, G.,
1993. A comparative study of free radicals in vertebrates I. Antioxidant
enzymes. Comp. Biochem. Physiol. B 105, 749–755.
Power, A., Sheehan, D., 1996. Seasonal variation in the antioxidant defence
systems of gill and digestive gland of the blue mussel, Mytilus edulis.
Comp. Biochem. Physiol. C 114, 99–103.
Renzoni, A., 1962. Ulteriori dati sul ciclo biologico riproduttivo di Mytilus
galloprovincialis LAMK. Riv. Biol. 55, 37.
Robledo, J.A.F., Santarem, M.M., Gonzales, P., Figueras, A., 1995.
Seasonal variations in the biochemical composition of the serum of
Mytilus galloprovincialis Lmk. and its relationship to the reproductive
cycle and parasitic load. Aquaculture 133, 311–322.
Rossi, M.A., Cecchini, G., Dianzani, M.M., 1983. Glutathione peroxidase,
glutathione reductase and glutathione transferase in two different
hepatomas and in normal liver. IRCS Med. Sci. Clin. Biochem. 11, 805.
Rudneva, I.I., 1999. Antioxidant system of Black Sea animals in early
development. Comp. Biochem. Physiol. C 122, 265–271.
Shaw, S., Jayatilleke, E., 1990. Ethanol-induced iron mobilization: role of
acetaldehyde–aldehyde oxidase generated superoxide. Free Radic.
Biol. Med. 9, 11–17.
Sheehan, D., Power, A., 1999. Effects of seasonality on xenobiotic and
antioxidant defence mechanisms of bivalve molluscs. Comp. Biochem.
Physiol. C 123, 193–199.
Sheehan, D., Mc Intosh, J., Power, A., Fitzpatrick, P.J., 1995. Drug
metabolism enzymes of mussels as bioindicators of chemical pollution.
Biochem. Soc. Trans. 23, 419–422.
Spasic, M.B., Sai*ic, Z.S., Buzad”ic, B., Korac, B., Blagojevic, D., Petrovic,
V.M., 1993. Effect of long-term exposure to cold on the antioxidant
defense system in the rat. Free Radic. Biol. Med. 15, 291–299.
Speers-Roesch, B., Ballantyne, J.S., 2005. Activities of antioxidant
enzymes and citochrome c oxidase in liver of Arctic and temperate
teleosts. Comp. Biochem. Physiol. A 140, 487–494.
Staats, D.A., Lohr, D.P., Colby, H.D., 1988. Effects of tocopherol depletion
on the regional differences in adrenal microsomal lipid peroxidation and
steroid metabolism. Endocrinology 123, 975–980.
Stjep*evic, J., 1974. Ecology of mussel (Mytilus galloprovincialis LAMK.)
and oyster (Ostrea edulis L.) in cultures of Boka Kotorska bay. Stud.
Mar. 7, 3–164.
Stohs, S.J., Bagchi, D., Hassoun, E., Bagchi, M., 2000. Oxidative
mechanisms in the toxicity of chromium and cadmium ions. J. Environ.
Pathol. Toxicol. Oncol. 19, 201–213.
Sukhotin, A., Abele, D., Portner, H., 2002. Growth, metabolism and lipid
peroxidation in Mytilus edulis: age and size effects. Mar. Ecol., Prog.
Ser. 226, 223–234.
Takada, Y., Noguchit, T., Kayiyama, M., 1982. Superoxide dismutase in
various tissues from rabbits bearing the Vx-2 carcinoma in the maxillary
sinus. Cancer Res. 42, 4233–4235.
Tamura, M., Oschino, N., Chance, B., 1982. Some characteristics of
hydrogen and alkyl-hydroperoxides metabolizing systems in cardiac
tissue. J. Biochem. 92, 1019–1031.
Turrens, J.F., Alexandre, A., Lehninger, A., 1985. Ubisemiquinone is the
electron donor for superoxide formation by complex III of heart
mitochondria. Arch. Biochem. Biophys. 237, 408–414.
Van der Oost, R., Beyer, J., Vermeulen, N., 2003. Fish bioaccumulation and
biomarkers in environmental risk assessment: a review. Environ.
Toxicol. Pharmacol. 13, 57–149.
Viarengo, A., Canesi, L., Pertica, M., Livingstone, D.R., 1991. Seasonal
variation in the antioxidant defence system and lipid peroxidation of the
digestive gland of mussels. Comp. Biochem. Physiol. C 100, 187–190.
Wilhelm Filho, D., De Giulivi, C., Boveris, A., 1993. Antioxidant defences
in marine fish I. Teleosts. Comp. Biochem. Physiol. C 106, 409–413.
Wilhelm Filho, D., Torres, M.A., Marcon, J.L., Fraga, C.G., Boveris, A.,
2000. Comparative antioxidant defences in vertebrates—emphasis on
fish and mammals. Trends Comp. Biochem. Physiol. 7, 33–45.
Wilhelm Filho, D., Tribes, T., Gaspari, C., Claudio, F.D., Torres, M.A.,
Magalhaes, A.R.M., 2001. Seasonal changes in antioxidant defenses of
the digestive gland of the brown mussel (Perna perna). Aquaculture
203, 149–158.
Zielinski, S., Portner, H.O., 2000. Oxidative stress and antioxidative
defence in cephalopods: a function of metabolic rate or age? Comp.
Biochem. Physiol. B 125, 147–160.




















