
Testosterone Stimulates Intracellular Calcium Release and Mitogen-Activated Protein Kinases Via a G...
12
Testosterone Stimulates Intracellular Calcium Release and Mitogen-Activated Protein Kinases Via a G Protein- Coupled Receptor in Skeletal Muscle Cells MANUEL ESTRADA, ALEJANDRA ESPINOSA, MARIOLY MU ¨ LLER, AND ENRIQUE JAIMOVICH Centro de Estudios Moleculares de la Ce ´lula and Instituto de Ciencias Biome ´dicas, Facultad de Medicina, Universidad de Chile, Casilla 70005, Santiago 6530499, Chile Involvement of intracellular Ca 2 and ERK1/2 phosphoryla- tion in the fast nongenomic effects of androgens in myotubes was investigated. Testosterone or nandrolone produced fast (<1 min) and transient increases in intracellular Ca 2 with an oscillatory pattern. Calcium signals were slightly reduced in Ca 2 -free medium, but lack of oscillations was evident. Sig- nals were blocked by U-73122 and xestospongin B, inhibitors of inositol 1,4,5-trisphosphate (IP 3 ) pathway. Furthermore, IP 3 increased transiently 2- to 3-fold 45 sec after hormone addition. Cyproterone neither affected the fast Ca 2 signal nor the increase in IP 3 . Calcium increases could also be in- duced by the impermeant testosterone conjugated to BSA, and the effect of testosterone was abolished in cells incubated with guanosine 5-O-(2-thiodiphosphate) or pertussis toxin. Stimulation of myotubes with testosterone, nandrolone, or testosterone conjugated to BSA increased immunodetectable phosphorylation of ERK1/2 within 5 min, and this effect was not inhibited by cyproterone. Phosphorylation was blocked by the use of 1,2-bis(2-aminophenoxy)ethane-N,N,N,N- tetraacetic acid-acetoxymethylester, U-73122, and xestospon- gin B as well as by dominant negative Ras, MAPK kinase (MEK), or the MEK inhibitor PD-98059. In addition, guanosine 5-O-(2-thiodiphosphate) or pertussis toxin blocked ERK1/2 phosphorylation. These results are consistent with a fast ef- fect of testosterone, involving a G protein-linked receptor at the plasma membrane, IP 3 -mediated Ca 2 signal, and the Ras/ MEK/ERK pathway in muscle cells. (Endocrinology 144: 3586 –3597, 2003) S KELETAL MUSCLE is a target for the action of anabolic steroids (1). These hormones affect cell growth and development and are known to produce their effects by binding to intracellular androgen receptors. Upon ligand binding, the hormone-receptor complex translocates to the nucleus and binds to specific DNA sequences called hor- mone response elements. The binding of the complex to DNA results in transcription of specific genes (2, 3). However, several nongenomic actions of androgens on rat osteoblasts (4), macrophages (5), and skeletal muscle cells (6) have been reported. Common to these early effects is the fast intracel- lular calcium increase, activation of Ca 2 -dependent path- ways, and second messenger cascades (7). In skeletal muscle cells, intracellular calcium regulates contraction, but alter- native roles for calcium in muscle cells, as a regulator of gene expression, have been proposed (8, 9). These effects have been associated with functionally distinct exogenous stimuli as well as to coexistence of multiple Ca 2 release mechanisms within a single cell. In skeletal muscle cells the association between intracellular calcium increases and induction of spe- cific genomic responses is poorly understood. However, these responses could be involved in the generation of dif- ferent patterns of calcium signals (10, 11) and also in the interaction of different Ca 2 -dependent pathways (12). We have previously shown that in skeletal muscle cells, both aldosterone and testosterone produce rapid intracellular cal- cium increases (6) involving calcium release from inositol 1,4,5-trisphosphate (IP 3 )-sensitive stores. These calcium tran- sients were not modified by the use of spironolactone or cyproterone, inhibitors of intracellular mineralocorticoid and androgen receptors, respectively. Calcium signals elicited by steroid hormones, via an increase in intracellular IP 3 and associated with increased nucleoplasmic calcium, may have an important role in the regulation of unknown process in the muscle cell. An interesting route of convergence could cor- respond to ERK1/2. ERK1/2 are one class of MAPK and have been assigned an important role in the intracellular signal pathway that leads to division, growth, and/or differentia- tion in several cell types (13). This is mainly due to their capacity to phosphorylate a variety of transcription factors and other intracellular signal transduction proteins (14). Full activation of these enzymes requires dual phosphorylation of threonine and tyrosine residues (13, 14). The skeletal muscle ERK1/2 are activated by exercise (15), hormones (16, 17), growth factors (18), and diverse stress stimuli (19). Mem- brane receptors, intracellular second messengers, and local- ized changes in the intracellular calcium levels have been postulated to have a role in these effects. It is known that in skeletal muscle, androgen steroids can produce hypertrophy (20, 21); on the other hand, a role for calcium in muscle hypertrophy has been proposed (10). Thus, steroid hormones could activate rapid calcium increases as well as the MAPK cascades previous to nuclear events (22, 23). Several steroid hormones have been shown to stimulate ERK1/2 phosphor- ylation; it has been described that in breast cancer cells, estrogens can induce rapid activation of a MAPK cascade Abbreviations: BAPTA-AM, 1,2-Bis(2-aminophenoxy)ethane- N,N,N,N-tetraacetic acid-AM; CRE, cAMP response element; CREB, CRE-binding protein; GDPS, guanosine 5-O-(2-thiodiphosphate); IP 3 , inositol, 1,4,5-trisphosphate; MEK, MAPK kinase; PLC, phospholipase C; PTX, pertussis toxin; T-BSA, testosterone bound to albumin; TBST, Tris- buffered saline containing 0.1% Tween 20. 0013-7227/03/$15.00/0 Endocrinology 144(8):3586 –3597 Printed in U.S.A. Copyright © 2003 by The Endocrine Society doi: 10.1210/en.2002-0164 3586
Transcript of Testosterone Stimulates Intracellular Calcium Release and Mitogen-Activated Protein Kinases Via a G...

Testosterone Stimulates Intracellular Calcium Releaseand Mitogen-Activated Protein Kinases Via a G Protein-Coupled Receptor in Skeletal Muscle Cells
MANUEL ESTRADA, ALEJANDRA ESPINOSA, MARIOLY MULLER, AND ENRIQUE JAIMOVICH
Centro de Estudios Moleculares de la Celula and Instituto de Ciencias Biomedicas, Facultad de Medicina, Universidad deChile, Casilla 70005, Santiago 6530499, Chile
Involvement of intracellular Ca2� and ERK1/2 phosphoryla-tion in the fast nongenomic effects of androgens in myotubeswas investigated. Testosterone or nandrolone produced fast(<1 min) and transient increases in intracellular Ca2� with anoscillatory pattern. Calcium signals were slightly reduced inCa2�-free medium, but lack of oscillations was evident. Sig-nals were blocked by U-73122 and xestospongin B, inhibitorsof inositol 1,4,5-trisphosphate (IP3) pathway. Furthermore,IP3 increased transiently 2- to 3-fold 45 sec after hormoneaddition. Cyproterone neither affected the fast Ca2� signalnor the increase in IP3. Calcium increases could also be in-duced by the impermeant testosterone conjugated to BSA, andthe effect of testosterone was abolished in cells incubatedwith guanosine 5�-O-(2-thiodiphosphate) or pertussis toxin.
Stimulation of myotubes with testosterone, nandrolone, ortestosterone conjugated to BSA increased immunodetectablephosphorylation of ERK1/2 within 5 min, and this effect wasnot inhibited by cyproterone. Phosphorylation was blockedby the use of 1,2-bis(2-aminophenoxy)ethane-N,N,N�,N�-tetraacetic acid-acetoxymethylester, U-73122, and xestospon-gin B as well as by dominant negative Ras, MAPK kinase(MEK), or the MEK inhibitor PD-98059. In addition, guanosine5�-O-(2-thiodiphosphate) or pertussis toxin blocked ERK1/2phosphorylation. These results are consistent with a fast ef-fect of testosterone, involving a G protein-linked receptor atthe plasma membrane, IP3-mediated Ca2� signal, and the Ras/MEK/ERK pathway in muscle cells. (Endocrinology 144:3586–3597, 2003)
SKELETAL MUSCLE is a target for the action of anabolicsteroids (1). These hormones affect cell growth and
development and are known to produce their effects bybinding to intracellular androgen receptors. Upon ligandbinding, the hormone-receptor complex translocates to thenucleus and binds to specific DNA sequences called hor-mone response elements. The binding of the complex to DNAresults in transcription of specific genes (2, 3). However,several nongenomic actions of androgens on rat osteoblasts(4), macrophages (5), and skeletal muscle cells (6) have beenreported. Common to these early effects is the fast intracel-lular calcium increase, activation of Ca2�-dependent path-ways, and second messenger cascades (7). In skeletal musclecells, intracellular calcium regulates contraction, but alter-native roles for calcium in muscle cells, as a regulator of geneexpression, have been proposed (8, 9). These effects havebeen associated with functionally distinct exogenous stimulias well as to coexistence of multiple Ca2� release mechanismswithin a single cell. In skeletal muscle cells the associationbetween intracellular calcium increases and induction of spe-cific genomic responses is poorly understood. However,these responses could be involved in the generation of dif-ferent patterns of calcium signals (10, 11) and also in theinteraction of different Ca2�-dependent pathways (12). Wehave previously shown that in skeletal muscle cells, both
aldosterone and testosterone produce rapid intracellular cal-cium increases (6) involving calcium release from inositol1,4,5-trisphosphate (IP3)-sensitive stores. These calcium tran-sients were not modified by the use of spironolactone orcyproterone, inhibitors of intracellular mineralocorticoid andandrogen receptors, respectively. Calcium signals elicited bysteroid hormones, via an increase in intracellular IP3 andassociated with increased nucleoplasmic calcium, may havean important role in the regulation of unknown process in themuscle cell. An interesting route of convergence could cor-respond to ERK1/2. ERK1/2 are one class of MAPK and havebeen assigned an important role in the intracellular signalpathway that leads to division, growth, and/or differentia-tion in several cell types (13). This is mainly due to theircapacity to phosphorylate a variety of transcription factorsand other intracellular signal transduction proteins (14). Fullactivation of these enzymes requires dual phosphorylation ofthreonine and tyrosine residues (13, 14). The skeletal muscleERK1/2 are activated by exercise (15), hormones (16, 17),growth factors (18), and diverse stress stimuli (19). Mem-brane receptors, intracellular second messengers, and local-ized changes in the intracellular calcium levels have beenpostulated to have a role in these effects. It is known that inskeletal muscle, androgen steroids can produce hypertrophy(20, 21); on the other hand, a role for calcium in musclehypertrophy has been proposed (10). Thus, steroid hormonescould activate rapid calcium increases as well as the MAPKcascades previous to nuclear events (22, 23). Several steroidhormones have been shown to stimulate ERK1/2 phosphor-ylation; it has been described that in breast cancer cells,estrogens can induce rapid activation of a MAPK cascade
Abbreviations: BAPTA-AM, 1,2-Bis(2-aminophenoxy)ethane-N,N,N�,N�-tetraacetic acid-AM; CRE, cAMP response element; CREB,CRE-binding protein; GDP�S, guanosine 5�-O-(2-thiodiphosphate); IP3,inositol, 1,4,5-trisphosphate; MEK, MAPK kinase; PLC, phospholipase C;PTX, pertussis toxin; T-BSA, testosterone bound to albumin; TBST, Tris-buffered saline containing 0.1% Tween 20.
0013-7227/03/$15.00/0 Endocrinology 144(8):3586–3597Printed in U.S.A. Copyright © 2003 by The Endocrine Society
doi: 10.1210/en.2002-0164
3586

independent of both transcription and protein synthesis, butrequire mobilization of intracellular calcium (24). Moreover,it has been reported that estrogens can indirectly enhance thekinase activity of an estrogen receptor, which then activatesMAPK via the Ras pathway (25). In prostate cancer cells,dihydrotestosterone leads to a rapid and reversible activa-tion of ERK1/2 via the androgen receptor (26). In this workwe describe a pathway of intracellular signaling that includesa G protein-linked membrane receptor, Ca2� released fromintracellular stores through IP3 receptors, and activation ofthe ERK phosphorylation cascade. This mechanism would beeither parallel or previous to the traditional mode of actionproposed for steroid hormones.
Materials and MethodsChemical reagents
Testosterone, testosterone-BSA, cyproterone acetate (6-chloro-1�,2�-dihydro-17-hydroxy-3�H-cycloropropa[1,2]-pregna-1,4,6-triene-3,20-dioneacetate), nandrolone (4-estren-17�-ol-3-one), 17�-estradiol, pro-gesterone, dexamethasone, U-73122, and luciferin were purchased fromSigma-Aldrich Corp. (St. Louis, MO). Fluo-3 acetoxymethylester (Fluo-3/AM) and 1,2-bis(2-aminophenoxy)ethane-N,N,N�,N�-tetraacetic acid-acetoxymethylester (BAPTA-AM) were purchased from MolecularProbes (Eugene, OR). PD-98059, Bordetella pertussis toxin (PTX),guanosine 5�-O-(2-thiodiphosphate) (GDP�S), and genistein were ob-tained from Calbiochem (La Jolla, CA). Xestospongin B was a gift fromDr. Jordi Molgo (Centre National de la Recherche Scientifique, Gif-sur-Yvette, France). Antiphospho-ERK1/2 antibody was obtained from CellSignaling Technology, Inc. (Beverly, MA). Total ERK1/2 and horserad-ish peroxidase-linked antirabbit IgG were purchased from New EnglandBiolabs, Inc. (Beverly, MA). Enhanced chemiluminescence reagents andfluorescein isothiocyanate-conjugated goat antirabbit IgG were obtainedfrom Pierce Chemical Co. (Rockford, IL). Antiandrogen receptor anti-body (C-19) was purchased from Santa Cruz Biotechnology (Santa Cruz,CA). The plasmids encoding dominant negative Ha-RasN17, dominantnegative MAPK kinase (MEK), pON249 in which �-galactosidase isexpressed from a constitutive cytomegalovirus promoter, and the re-porter construct p-cAMP response element (CRE)-Luc were provided byDr. Sergio Lavandero (Facultad de Ciencias Quımicas y Farmaceuticas,Universidad de Chile). Other reagents were of the analytical grade.
Cell cultures
Rat myotubes were cultured as reported previously (27). Briefly,myoblasts were obtained from rat neonatal hind limbs, and the tissuewas mechanically dispersed and then treated with 10% (wt/vol) colla-genase for 15 min at 37 C under mild agitation. The suspension wasfiltered through a Nytex (Sartorius, Goettingen, Germany) membraneand spun down at low speed, preplating was used to partially eliminatefibroblasts, and finally, cells were plated onto dishes (60 mm) at a densityof 1.2 � 106/dish. The culture medium was DMEM/F-12 without phenolred, 10% bovine serum, 2.5% fetal calf serum, 100 mg/liter penicillin,50 mg/liter streptomycin, and 2.5 mg/liter amphotericin B. To eliminateremaining fibroblasts, 10 �m cytosine arabinoside was added on thethird day of culture for 24 h. The medium was then replaced withserum-free medium. Myotubes with an estimated purity of more than90% were visible after the fifth day of culture. Unless indicated, we used6- to 8-d-old cultures exhibiting a fairly homogeneous population ofmyotubes with central nuclei and measuring 200–300 �m long and20–40 �m wide, corresponding to young, not fully differentiated cells.
Intracellular calcium measurement
Cytosolic calcium images were obtained from single noncontractingmyotubes preloaded with Fluo-3-AM using an inverted confocal mi-croscope (Axiovert 135M, LSM Microsystems, Carl Zeiss, New York,NY) or a fluorescence microscope (T041, Olympus Corp., New HydePark, NY) equipped with a cooled CCD camera and image acquisitionsystem (MCD 600, Spectra Source Instruments, Westlake Village, CA).
Myotubes were washed three times with Krebs buffer [145 mm NaCl, 5mm KCl, 2.6 mm CaCl2, 1 mm MgCl2, 10 mm HEPES-sodium, and 5.6 mmglucose (pH 7.4)] to remove serum and loaded with 5.4 �m Fluo-3-AM(coming from a stock in 20% pluronic acid-dimethylsulfoxide) for 30 minat room temperature. After loading, myotubes were washed with thesame buffer and used within 2 h. The cell-containing coverslips weremounted in a 1-ml capacity plastic chamber and placed in the micro-scope for fluorescence measurements after excitation with a 488-nmwavelength argon laser beam or filter system. Hormones were addeddirectly to the medium, or the solution was fast changed in the chamber(1 sec). The fluorescent images were collected every 0.4–2.0 sec for fastsignals and were analyzed frame by frame with the image data acqui-sition program (MCD 600, Spectra-Source) of the equipment. An objec-tive lens PlanApo �40 (numerical aperture, 1.4) was generally used. Inmost of the acquisitions, the image dimension was 512 � 120 pixels. Theinhibitors were added during the dye incubation; times and concentra-tions are indicated in Results. To assess the role of G proteins, myotubeswere incubated either with 1 �g/ml PTX for 5 h or in a permeabilizationsolution [100 mm KCl, 20 mm NaCl, 5 mm MgSO4, 1 mm NaH2PO4, 25mm NaHCO3, 3 mm EGTA, 1 mm CaCl2, 20 mm Tris-HCl (pH 7.4), 0.1%BSA, 1 mm ATP, 0.1% glucose, and 40 mg/ml saponin] for 5 min in thepresence or absence of 100 nm GDP�S, a nonhydrolyzable analog ofGDP, before hormone stimulation. Intracellular calcium was expressedas a percentage of fluorescence intensity relative to basal fluorescence,which was stable for at least 5 min during resting conditions. Theincrease in fluorescence intensity is proportional to the rise in intracel-lular calcium level (28).
Digital image processing
Elimination of out of focus fluorescence was performed by software.Both “no-neighbors” deconvolution algorithm and Castleman’s pointspread function theoretical model were used. Complementary to res-toration methods, a procedure was created to segment the images. Tosection an image, an initial contour can be entered manually, and arecursive algorithm that adapts automatically to the region of interest(adaptable contour) can be applied (6). To quantify fluorescence, thesummed pixel intensity was calculated for the section delimited by acontour. As a way of increasing the efficiency of these data manipula-tions, action sequences were generated. To avoid possible interferencein the fluorescence by changes in volume after exposure to steroids, thearea of fluorescent cell was determined by image analysis using adaptivecontour and then creating a binary mask, which was compared with abrightfield image.
Determination of IP3 levels
IP3 levels were determined as described previously (27). Briefly, acrude rat cerebellum membrane preparation was obtained after homog-enization in 50 mm Tris-HCl (pH 7.7), 1 mm EDTA, 2 mm �-mercapto-ethanol, and centrifugation at 20,000 � g for 15 min. This procedure wasrepeated three times, resuspending the final pellet in the same solutionplus 0.3 m sucrose and freezing it at �80 C until use. The membranepreparation was calibrated for IP3 binding with 1.6 nm [3H]IP3 and 1–120nm cold IP3, carrying out the sample analysis in a similar way, butadding an aliquot of the neutralized supernatant instead of cold IP3. Theremaining membrane-bound [3H]IP3 radioactivity was measured byliquid scintillation.
Immunocytochemistry
Intracellular androgen receptor was localized using indirect immu-nofluorescence. Myotubes (control and testosterone stimulated) werewashed and then fixed with 100% methanol at �20 C for 20 min andtreated with a blocking solution of 1% BSA in PBS for 30 min. Cells wereincubated with the primary polyclonal antiintracellular androgen re-ceptor antibody (1:100) overnight at 4 C. Later, myotubes were washedin PBS and incubated with fluorescein isothiocyanate-conjugated goatantirabbit IgG diluted 1:200 for 2 h at room temperature. Cells werewashed, and Vectashield (Vector Laboratories, Inc., Burlingame, CA)was added to prevent bleaching. Myotubes were examined with a con-focal microscope (135-M LSM Microsystems, Carl Zeiss). Controls were
Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes Endocrinology, August 2003, 144(8):3586–3597 3587
performed both without the primary antibody and by displacement withthe antigenic peptide as a test for specificity.
ERK1/2 phosphorylation by steroids and Western blot
Myotubes were grown on 60-mm plates and serum-starved for 24 hbefore being exposed to steroids for various times, as indicated in Resultsand the figure legends. Cells were washed with ice-cold PBS and scrapedoff plates in 70 �l harvesting lysis buffer [150 mm NaCl, 1 mm EGTA,1% Nonidet P-40, 50 mm Tris-HCl (pH 7.4), 5 mm sodium orthovanadate,20 mm NaF, 1 �g/ml aprotinin, 1 �m pepstatin, 20 �m leupeptin, 1 mmbenzamidine, and 0.2 mm 4-(2-aminoethyl)benzene sulfonyl fluoride].They were then sonicated and subjected to centrifugation at 15,000 � gfor 20 min. Supernatants were removed and divided into portions forERK1/2 determination and protein assay. Equal amounts of proteins(20 �g) were denatured at 100 C in 30% glycerol, 8% sodium dodecylsulfate, 10% 2-mercaptoethanol, 25 mm Tris-HCl (pH 6.8), and 0.1%bromophenol blue; resolved by 10% SDS-PAGE; and transferred to0.22-�m pore nitrocellulose filters. Nonspecific staining was blockedwith Tris-buffered saline containing 0.1% Tween 20 (TBST) and 5%nonfat dry milk at room temperature for 1 h. Nitrocellulose membraneswere incubated overnight at 4 C with primary antibody raised againstphosphorylated ERK1/2 (1:1000). Next, the membranes were washedthree times in TBST, incubated with peroxidase-conjugated secondaryantibody (1:2000) for 2 h at room temperature, and washed three ad-ditional times in TBST. Enhanced chemiluminescence techniques wereused to visualize the immunoblots according to the manufacturer’sprotocol. The membranes were stripped according to conventionalmethods and immunoblotted with pan-ERK (phosphorylated and non-phosphorylated ERK1/2) for total ERK1/2 protein. Digitized imagesof the immunoblots were used for densitometric measurements withScionImage software (NIH, Bethesda, MD). Relative enzyme activationwas determined by normalization of densities image to that of the totalERK1/2 from the same membrane.
Transient transfection of primary myotubes
Cells were grown in 60-mm dishes to 60% confluence. Before trans-fection, the cells were washed to remove serum and were transientlytransfected with either empty vectors or dominant negative Ras, MEK,or pON249 (2–4 �g) using Lipofectamine (Invitrogen Life Technologies,Inc., Gaithersburg, MD) following the manufacturer’s instructions. Asingle plate of transfected cells was then used to set-up the experimentalcultures required for each assay, ensuring equal transfection efficienciesbetween different treatments, and cells were cultured for an additional24 h before treatment with androgens. For reporter gene assays, myo-tubes were cotransfected with pCRE-Luc (2 �g/ml) and pON249 (2�g/ml) plasmids, and then luciferase and �-galactosidase activities weremeasured. The ratio between luciferase values and �-galactosidase val-ues was used to correct for differences in transfection efficiency.
Statistics
Data are expressed as the mean � sem. Differences between basal andpoststimulated points were determined using a paired t test. P � 0.05was considered statistically significant.
ResultsEffects of testosterone on intracellular calcium in myotubes
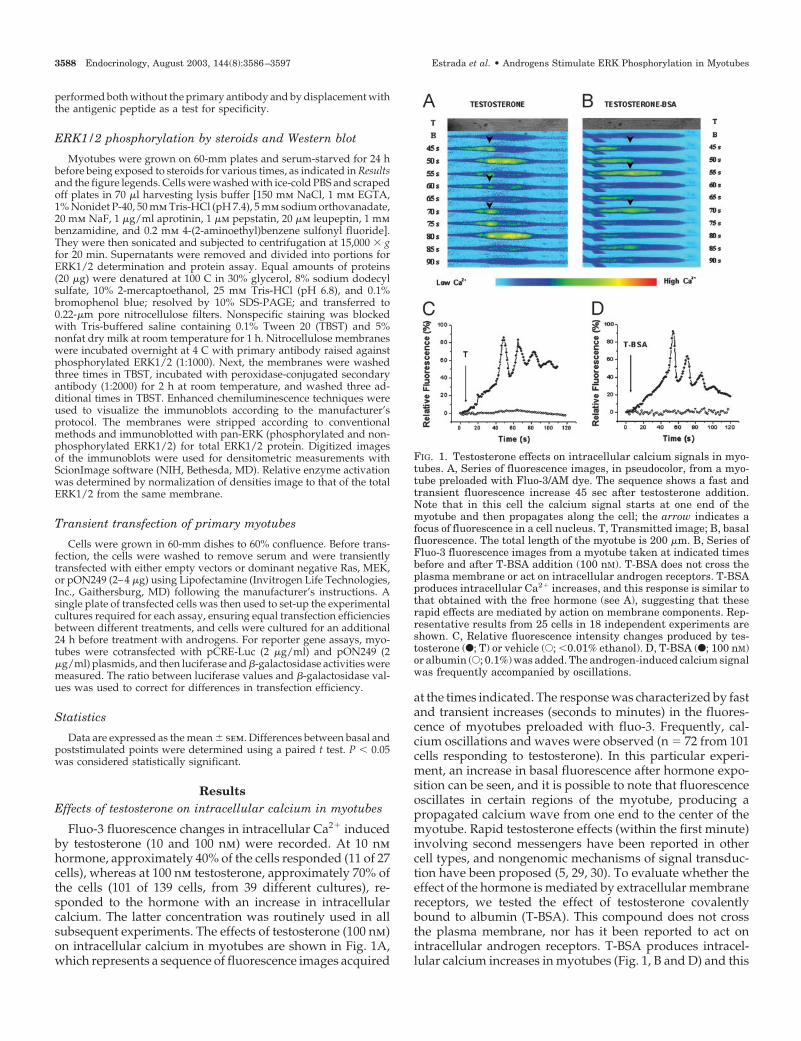
Fluo-3 fluorescence changes in intracellular Ca2� inducedby testosterone (10 and 100 nm) were recorded. At 10 nmhormone, approximately 40% of the cells responded (11 of 27cells), whereas at 100 nm testosterone, approximately 70% ofthe cells (101 of 139 cells, from 39 different cultures), re-sponded to the hormone with an increase in intracellularcalcium. The latter concentration was routinely used in allsubsequent experiments. The effects of testosterone (100 nm)on intracellular calcium in myotubes are shown in Fig. 1A,which represents a sequence of fluorescence images acquired
at the times indicated. The response was characterized by fastand transient increases (seconds to minutes) in the fluores-cence of myotubes preloaded with fluo-3. Frequently, cal-cium oscillations and waves were observed (n � 72 from 101cells responding to testosterone). In this particular experi-ment, an increase in basal fluorescence after hormone expo-sition can be seen, and it is possible to note that fluorescenceoscillates in certain regions of the myotube, producing apropagated calcium wave from one end to the center of themyotube. Rapid testosterone effects (within the first minute)involving second messengers have been reported in othercell types, and nongenomic mechanisms of signal transduc-tion have been proposed (5, 29, 30). To evaluate whether theeffect of the hormone is mediated by extracellular membranereceptors, we tested the effect of testosterone covalentlybound to albumin (T-BSA). This compound does not crossthe plasma membrane, nor has it been reported to act onintracellular androgen receptors. T-BSA produces intracel-lular calcium increases in myotubes (Fig. 1, B and D) and this
FIG. 1. Testosterone effects on intracellular calcium signals in myo-tubes. A, Series of fluorescence images, in pseudocolor, from a myo-tube preloaded with Fluo-3/AM dye. The sequence shows a fast andtransient fluorescence increase 45 sec after testosterone addition.Note that in this cell the calcium signal starts at one end of themyotube and then propagates along the cell; the arrow indicates afocus of fluorescence in a cell nucleus. T, Transmitted image; B, basalfluorescence. The total length of the myotube is 200 �m. B, Series ofFluo-3 fluorescence images from a myotube taken at indicated timesbefore and after T-BSA addition (100 nM). T-BSA does not cross theplasma membrane or act on intracellular androgen receptors. T-BSAproduces intracellular Ca2� increases, and this response is similar tothat obtained with the free hormone (see A), suggesting that theserapid effects are mediated by action on membrane components. Rep-resentative results from 25 cells in 18 independent experiments areshown. C, Relative fluorescence intensity changes produced by tes-tosterone (F; T) or vehicle (E; �0.01% ethanol). D, T-BSA (F; 100 nM)or albumin (E; 0.1%) was added. The androgen-induced calcium signalwas frequently accompanied by oscillations.
3588 Endocrinology, August 2003, 144(8):3586–3597 Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes
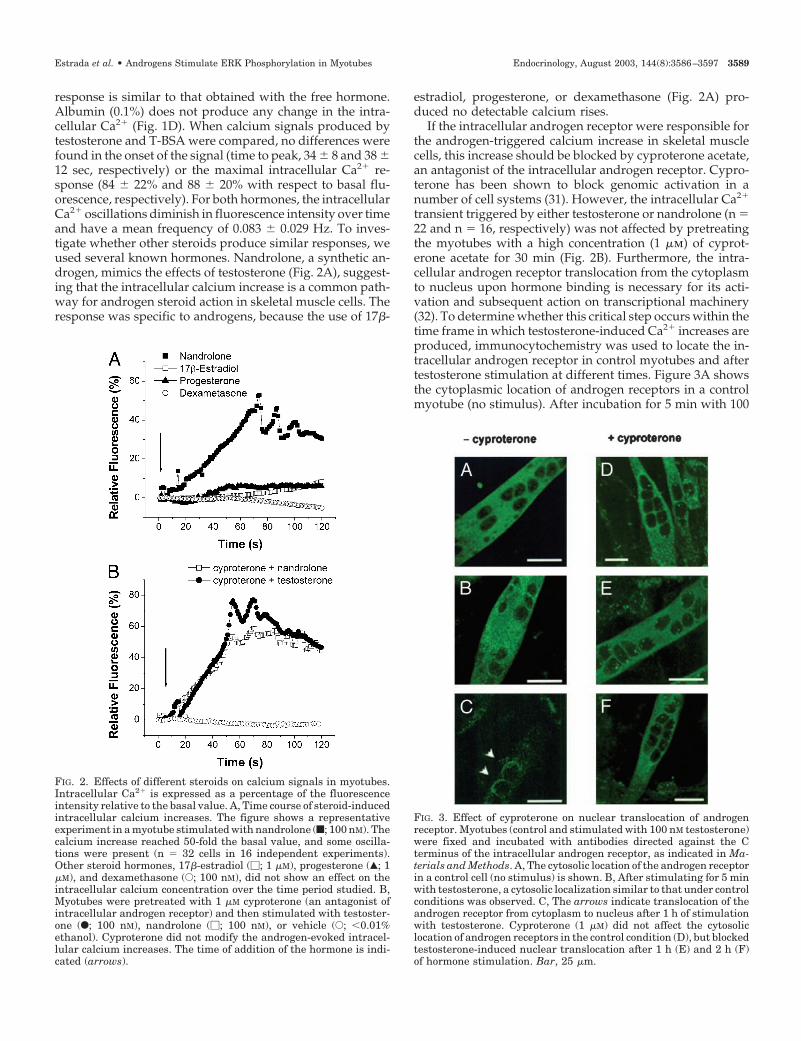
response is similar to that obtained with the free hormone.Albumin (0.1%) does not produce any change in the intra-cellular Ca2� (Fig. 1D). When calcium signals produced bytestosterone and T-BSA were compared, no differences werefound in the onset of the signal (time to peak, 34 � 8 and 38 �12 sec, respectively) or the maximal intracellular Ca2� re-sponse (84 � 22% and 88 � 20% with respect to basal flu-orescence, respectively). For both hormones, the intracellularCa2� oscillations diminish in fluorescence intensity over timeand have a mean frequency of 0.083 � 0.029 Hz. To inves-tigate whether other steroids produce similar responses, weused several known hormones. Nandrolone, a synthetic an-drogen, mimics the effects of testosterone (Fig. 2A), suggest-ing that the intracellular calcium increase is a common path-way for androgen steroid action in skeletal muscle cells. Theresponse was specific to androgens, because the use of 17�-
estradiol, progesterone, or dexamethasone (Fig. 2A) pro-duced no detectable calcium rises.
If the intracellular androgen receptor were responsible forthe androgen-triggered calcium increase in skeletal musclecells, this increase should be blocked by cyproterone acetate,an antagonist of the intracellular androgen receptor. Cypro-terone has been shown to block genomic activation in anumber of cell systems (31). However, the intracellular Ca2�
transient triggered by either testosterone or nandrolone (n �22 and n � 16, respectively) was not affected by pretreatingthe myotubes with a high concentration (1 �m) of cyprot-erone acetate for 30 min (Fig. 2B). Furthermore, the intra-cellular androgen receptor translocation from the cytoplasmto nucleus upon hormone binding is necessary for its acti-vation and subsequent action on transcriptional machinery(32). To determine whether this critical step occurs within thetime frame in which testosterone-induced Ca2� increases areproduced, immunocytochemistry was used to locate the in-tracellular androgen receptor in control myotubes and aftertestosterone stimulation at different times. Figure 3A showsthe cytoplasmic location of androgen receptors in a controlmyotube (no stimulus). After incubation for 5 min with 100
FIG. 2. Effects of different steroids on calcium signals in myotubes.Intracellular Ca2� is expressed as a percentage of the fluorescenceintensity relative to the basal value. A, Time course of steroid-inducedintracellular calcium increases. The figure shows a representativeexperiment in a myotube stimulated with nandrolone (f; 100 nM). Thecalcium increase reached 50-fold the basal value, and some oscilla-tions were present (n � 32 cells in 16 independent experiments).Other steroid hormones, 17�-estradiol (�; 1 �M), progesterone (Œ; 1�M), and dexamethasone (E; 100 nM), did not show an effect on theintracellular calcium concentration over the time period studied. B,Myotubes were pretreated with 1 �M cyproterone (an antagonist ofintracellular androgen receptor) and then stimulated with testoster-one (F; 100 nM), nandrolone (�; 100 nM), or vehicle (E; �0.01%ethanol). Cyproterone did not modify the androgen-evoked intracel-lular calcium increases. The time of addition of the hormone is indi-cated (arrows).
FIG. 3. Effect of cyproterone on nuclear translocation of androgenreceptor. Myotubes (control and stimulated with 100 nM testosterone)were fixed and incubated with antibodies directed against the Cterminus of the intracellular androgen receptor, as indicated in Ma-terials and Methods. A, The cytosolic location of the androgen receptorin a control cell (no stimulus) is shown. B, After stimulating for 5 minwith testosterone, a cytosolic localization similar to that under controlconditions was observed. C, The arrows indicate translocation of theandrogen receptor from cytoplasm to nucleus after 1 h of stimulationwith testosterone. Cyproterone (1 �M) did not affect the cytosoliclocation of androgen receptors in the control condition (D), but blockedtestosterone-induced nuclear translocation after 1 h (E) and 2 h (F)of hormone stimulation. Bar, 25 �m.
Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes Endocrinology, August 2003, 144(8):3586–3597 3589
nm testosterone, the intracellular distribution was similar tothat under control conditions (Fig. 3B), and only after 60 minof testosterone addition was receptor translocation to thenuclear compartment observed (Fig. 3C). This time is farlarger than that required to produce effects on intracellularCa2�. To confirm that the dose of cyproterone used was ableto inhibit androgen receptor translocation, myotubes wereincubated with 1 �m of the antagonist for 30 min beforetestosterone stimulation. Figure 3D shows that cyproteronealone did not affect the cytosolic localization of androgenreceptors and largely inhibited the testosterone-induced nu-clear translocation normally seen after 1 and 2 h (Fig. 3, E andF, respectively), indicating that this concentration of cypro-terone was effective in inhibiting the genomic effects of thehormone. Collectively, these results indicate that intracellu-lar androgen receptors are not involved in the testosterone-induced intracellular Ca2� increases in myotubes.
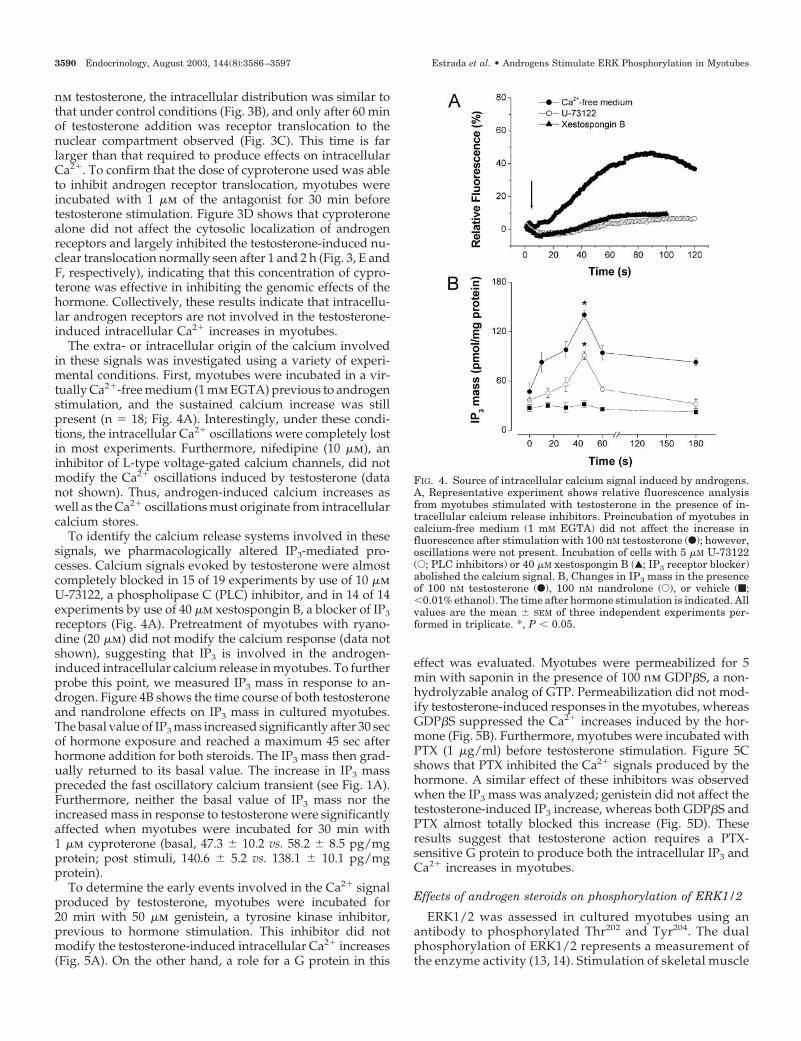
The extra- or intracellular origin of the calcium involvedin these signals was investigated using a variety of experi-mental conditions. First, myotubes were incubated in a vir-tually Ca2�-free medium (1 mm EGTA) previous to androgenstimulation, and the sustained calcium increase was stillpresent (n � 18; Fig. 4A). Interestingly, under these condi-tions, the intracellular Ca2� oscillations were completely lostin most experiments. Furthermore, nifedipine (10 �m), aninhibitor of L-type voltage-gated calcium channels, did notmodify the Ca2� oscillations induced by testosterone (datanot shown). Thus, androgen-induced calcium increases aswell as the Ca2� oscillations must originate from intracellularcalcium stores.
To identify the calcium release systems involved in thesesignals, we pharmacologically altered IP3-mediated pro-cesses. Calcium signals evoked by testosterone were almostcompletely blocked in 15 of 19 experiments by use of 10 �mU-73122, a phospholipase C (PLC) inhibitor, and in 14 of 14experiments by use of 40 �m xestospongin B, a blocker of IP3receptors (Fig. 4A). Pretreatment of myotubes with ryano-dine (20 �m) did not modify the calcium response (data notshown), suggesting that IP3 is involved in the androgen-induced intracellular calcium release in myotubes. To furtherprobe this point, we measured IP3 mass in response to an-drogen. Figure 4B shows the time course of both testosteroneand nandrolone effects on IP3 mass in cultured myotubes.The basal value of IP3 mass increased significantly after 30 secof hormone exposure and reached a maximum 45 sec afterhormone addition for both steroids. The IP3 mass then grad-ually returned to its basal value. The increase in IP3 masspreceded the fast oscillatory calcium transient (see Fig. 1A).Furthermore, neither the basal value of IP3 mass nor theincreased mass in response to testosterone were significantlyaffected when myotubes were incubated for 30 min with1 �m cyproterone (basal, 47.3 � 10.2 vs. 58.2 � 8.5 pg/mgprotein; post stimuli, 140.6 � 5.2 vs. 138.1 � 10.1 pg/mgprotein).
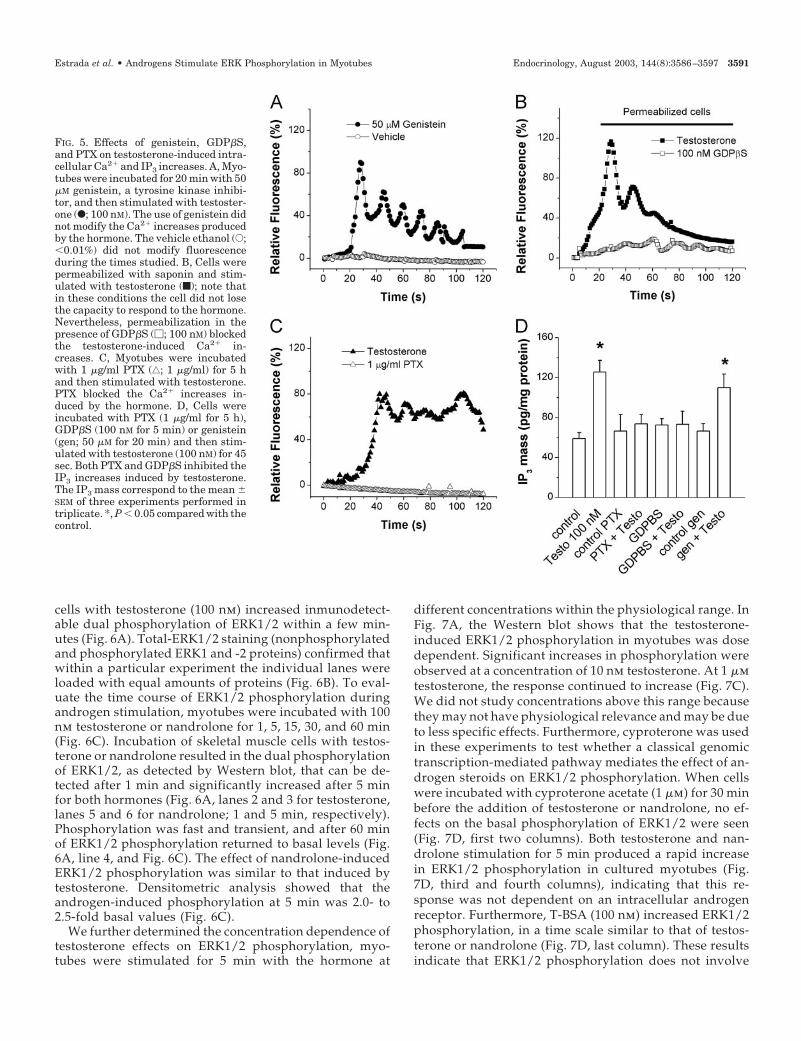
To determine the early events involved in the Ca2� signalproduced by testosterone, myotubes were incubated for20 min with 50 �m genistein, a tyrosine kinase inhibitor,previous to hormone stimulation. This inhibitor did notmodify the testosterone-induced intracellular Ca2� increases(Fig. 5A). On the other hand, a role for a G protein in this
effect was evaluated. Myotubes were permeabilized for 5min with saponin in the presence of 100 nm GDP�S, a non-hydrolyzable analog of GTP. Permeabilization did not mod-ify testosterone-induced responses in the myotubes, whereasGDP�S suppressed the Ca2� increases induced by the hor-mone (Fig. 5B). Furthermore, myotubes were incubated withPTX (1 �g/ml) before testosterone stimulation. Figure 5Cshows that PTX inhibited the Ca2� signals produced by thehormone. A similar effect of these inhibitors was observedwhen the IP3 mass was analyzed; genistein did not affect thetestosterone-induced IP3 increase, whereas both GDP�S andPTX almost totally blocked this increase (Fig. 5D). Theseresults suggest that testosterone action requires a PTX-sensitive G protein to produce both the intracellular IP3 andCa2� increases in myotubes.
Effects of androgen steroids on phosphorylation of ERK1/2
ERK1/2 was assessed in cultured myotubes using anantibody to phosphorylated Thr202 and Tyr204. The dualphosphorylation of ERK1/2 represents a measurement ofthe enzyme activity (13, 14). Stimulation of skeletal muscle
FIG. 4. Source of intracellular calcium signal induced by androgens.A, Representative experiment shows relative fluorescence analysisfrom myotubes stimulated with testosterone in the presence of in-tracellular calcium release inhibitors. Preincubation of myotubes incalcium-free medium (1 mM EGTA) did not affect the increase influorescence after stimulation with 100 nM testosterone (F); however,oscillations were not present. Incubation of cells with 5 �M U-73122(E; PLC inhibitors) or 40 �M xestospongin B (Œ; IP3 receptor blocker)abolished the calcium signal. B, Changes in IP3 mass in the presenceof 100 nM testosterone (F), 100 nM nandrolone (E), or vehicle (f;�0.01% ethanol). The time after hormone stimulation is indicated. Allvalues are the mean � SEM of three independent experiments per-formed in triplicate. *, P � 0.05.
3590 Endocrinology, August 2003, 144(8):3586–3597 Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes
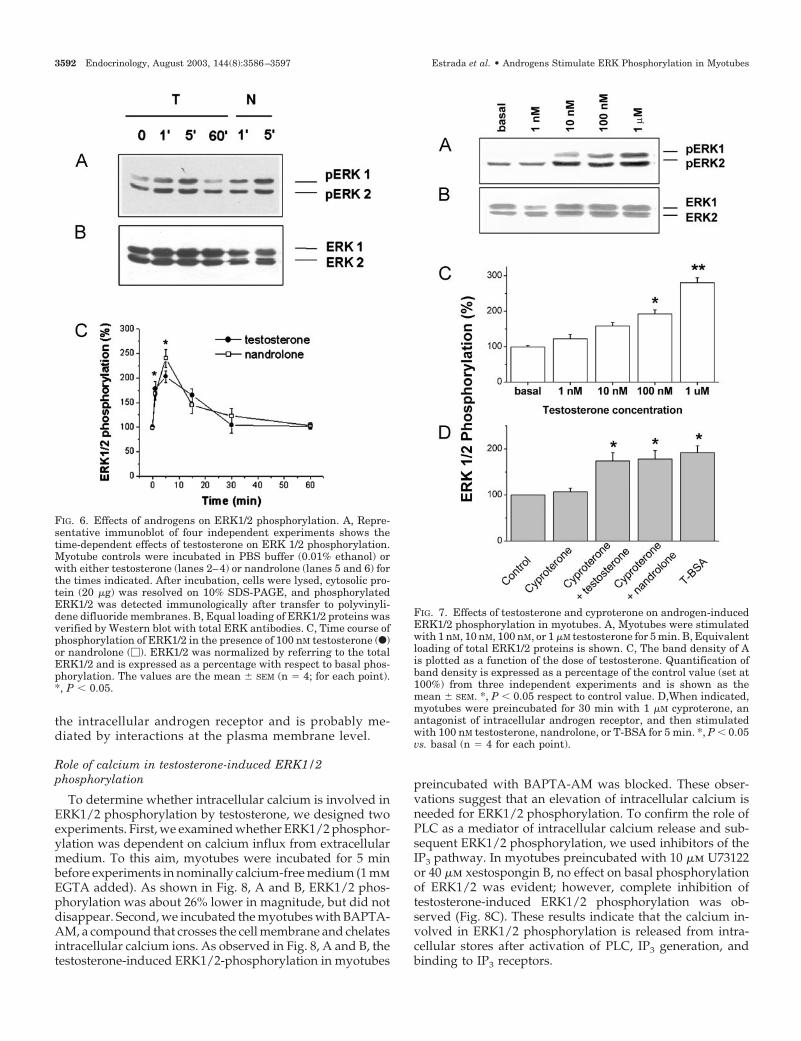
cells with testosterone (100 nm) increased inmunodetect-able dual phosphorylation of ERK1/2 within a few min-utes (Fig. 6A). Total-ERK1/2 staining (nonphosphorylatedand phosphorylated ERK1 and -2 proteins) confirmed thatwithin a particular experiment the individual lanes wereloaded with equal amounts of proteins (Fig. 6B). To eval-uate the time course of ERK1/2 phosphorylation duringandrogen stimulation, myotubes were incubated with 100nm testosterone or nandrolone for 1, 5, 15, 30, and 60 min(Fig. 6C). Incubation of skeletal muscle cells with testos-terone or nandrolone resulted in the dual phosphorylationof ERK1/2, as detected by Western blot, that can be de-tected after 1 min and significantly increased after 5 minfor both hormones (Fig. 6A, lanes 2 and 3 for testosterone,lanes 5 and 6 for nandrolone; 1 and 5 min, respectively).Phosphorylation was fast and transient, and after 60 minof ERK1/2 phosphorylation returned to basal levels (Fig.6A, line 4, and Fig. 6C). The effect of nandrolone-inducedERK1/2 phosphorylation was similar to that induced bytestosterone. Densitometric analysis showed that theandrogen-induced phosphorylation at 5 min was 2.0- to2.5-fold basal values (Fig. 6C).
We further determined the concentration dependence oftestosterone effects on ERK1/2 phosphorylation, myo-tubes were stimulated for 5 min with the hormone at
different concentrations within the physiological range. InFig. 7A, the Western blot shows that the testosterone-induced ERK1/2 phosphorylation in myotubes was dosedependent. Significant increases in phosphorylation wereobserved at a concentration of 10 nm testosterone. At 1 �mtestosterone, the response continued to increase (Fig. 7C).We did not study concentrations above this range becausethey may not have physiological relevance and may be dueto less specific effects. Furthermore, cyproterone was usedin these experiments to test whether a classical genomictranscription-mediated pathway mediates the effect of an-drogen steroids on ERK1/2 phosphorylation. When cellswere incubated with cyproterone acetate (1 �m) for 30 minbefore the addition of testosterone or nandrolone, no ef-fects on the basal phosphorylation of ERK1/2 were seen(Fig. 7D, first two columns). Both testosterone and nan-drolone stimulation for 5 min produced a rapid increasein ERK1/2 phosphorylation in cultured myotubes (Fig.7D, third and fourth columns), indicating that this re-sponse was not dependent on an intracellular androgenreceptor. Furthermore, T-BSA (100 nm) increased ERK1/2phosphorylation, in a time scale similar to that of testos-terone or nandrolone (Fig. 7D, last column). These resultsindicate that ERK1/2 phosphorylation does not involve
FIG. 5. Effects of genistein, GDP�S,and PTX on testosterone-induced intra-cellular Ca2� and IP3 increases. A, Myo-tubes were incubated for 20 min with 50�M genistein, a tyrosine kinase inhibi-tor, and then stimulated with testoster-one (F; 100 nM). The use of genistein didnot modify the Ca2� increases producedby the hormone. The vehicle ethanol (E;�0.01%) did not modify fluorescenceduring the times studied. B, Cells werepermeabilized with saponin and stim-ulated with testosterone (f); note thatin these conditions the cell did not losethe capacity to respond to the hormone.Nevertheless, permeabilization in thepresence of GDP�S (�; 100 nM) blockedthe testosterone-induced Ca2� in-creases. C, Myotubes were incubatedwith 1 �g/ml PTX (‚; 1 �g/ml) for 5 hand then stimulated with testosterone.PTX blocked the Ca2� increases in-duced by the hormone. D, Cells wereincubated with PTX (1 �g/ml for 5 h),GDP�S (100 nM for 5 min) or genistein(gen; 50 �M for 20 min) and then stim-ulated with testosterone (100 nM) for 45sec. Both PTX and GDP�S inhibited theIP3 increases induced by testosterone.The IP3 mass correspond to the mean �SEM of three experiments performed intriplicate. *, P � 0.05 compared with thecontrol.
Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes Endocrinology, August 2003, 144(8):3586–3597 3591
the intracellular androgen receptor and is probably me-diated by interactions at the plasma membrane level.
Role of calcium in testosterone-induced ERK1/2phosphorylation
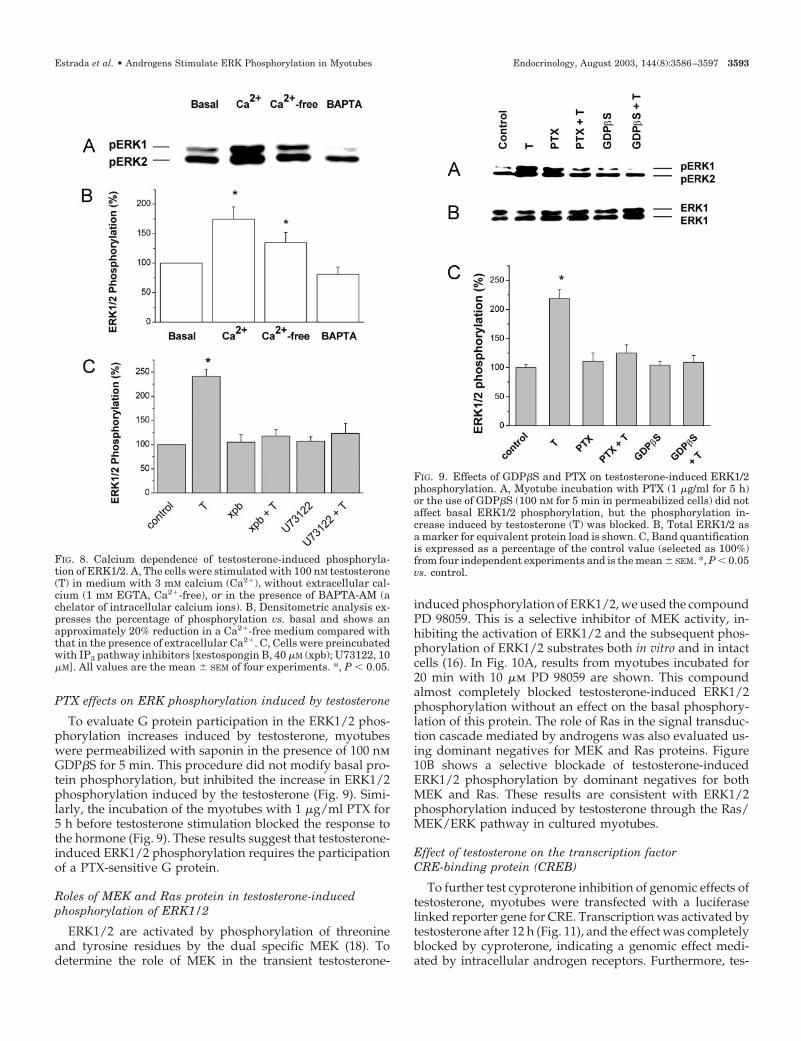
To determine whether intracellular calcium is involved inERK1/2 phosphorylation by testosterone, we designed twoexperiments. First, we examined whether ERK1/2 phosphor-ylation was dependent on calcium influx from extracellularmedium. To this aim, myotubes were incubated for 5 minbefore experiments in nominally calcium-free medium (1 mmEGTA added). As shown in Fig. 8, A and B, ERK1/2 phos-phorylation was about 26% lower in magnitude, but did notdisappear. Second, we incubated the myotubes with BAPTA-AM, a compound that crosses the cell membrane and chelatesintracellular calcium ions. As observed in Fig. 8, A and B, thetestosterone-induced ERK1/2-phosphorylation in myotubes
preincubated with BAPTA-AM was blocked. These obser-vations suggest that an elevation of intracellular calcium isneeded for ERK1/2 phosphorylation. To confirm the role ofPLC as a mediator of intracellular calcium release and sub-sequent ERK1/2 phosphorylation, we used inhibitors of theIP3 pathway. In myotubes preincubated with 10 �m U73122or 40 �m xestospongin B, no effect on basal phosphorylationof ERK1/2 was evident; however, complete inhibition oftestosterone-induced ERK1/2 phosphorylation was ob-served (Fig. 8C). These results indicate that the calcium in-volved in ERK1/2 phosphorylation is released from intra-cellular stores after activation of PLC, IP3 generation, andbinding to IP3 receptors.
FIG. 6. Effects of androgens on ERK1/2 phosphorylation. A, Repre-sentative immunoblot of four independent experiments shows thetime-dependent effects of testosterone on ERK 1/2 phosphorylation.Myotube controls were incubated in PBS buffer (0.01% ethanol) orwith either testosterone (lanes 2–4) or nandrolone (lanes 5 and 6) forthe times indicated. After incubation, cells were lysed, cytosolic pro-tein (20 �g) was resolved on 10% SDS-PAGE, and phosphorylatedERK1/2 was detected immunologically after transfer to polyvinyli-dene difluoride membranes. B, Equal loading of ERK1/2 proteins wasverified by Western blot with total ERK antibodies. C, Time course ofphosphorylation of ERK1/2 in the presence of 100 nM testosterone (F)or nandrolone (�). ERK1/2 was normalized by referring to the totalERK1/2 and is expressed as a percentage with respect to basal phos-phorylation. The values are the mean � SEM (n � 4; for each point).*, P � 0.05.
FIG. 7. Effects of testosterone and cyproterone on androgen-inducedERK1/2 phosphorylation in myotubes. A, Myotubes were stimulatedwith 1 nM, 10 nM, 100 nM, or 1 �M testosterone for 5 min. B, Equivalentloading of total ERK1/2 proteins is shown. C, The band density of Ais plotted as a function of the dose of testosterone. Quantification ofband density is expressed as a percentage of the control value (set at100%) from three independent experiments and is shown as themean � SEM. *, P � 0.05 respect to control value. D,When indicated,myotubes were preincubated for 30 min with 1 �M cyproterone, anantagonist of intracellular androgen receptor, and then stimulatedwith 100 nM testosterone, nandrolone, or T-BSA for 5 min. *, P � 0.05vs. basal (n � 4 for each point).
3592 Endocrinology, August 2003, 144(8):3586–3597 Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes
PTX effects on ERK phosphorylation induced by testosterone
To evaluate G protein participation in the ERK1/2 phos-phorylation increases induced by testosterone, myotubeswere permeabilized with saponin in the presence of 100 nmGDP�S for 5 min. This procedure did not modify basal pro-tein phosphorylation, but inhibited the increase in ERK1/2phosphorylation induced by the testosterone (Fig. 9). Simi-larly, the incubation of the myotubes with 1 �g/ml PTX for5 h before testosterone stimulation blocked the response tothe hormone (Fig. 9). These results suggest that testosterone-induced ERK1/2 phosphorylation requires the participationof a PTX-sensitive G protein.
Roles of MEK and Ras protein in testosterone-inducedphosphorylation of ERK1/2
ERK1/2 are activated by phosphorylation of threonineand tyrosine residues by the dual specific MEK (18). Todetermine the role of MEK in the transient testosterone-
induced phosphorylation of ERK1/2, we used the compoundPD 98059. This is a selective inhibitor of MEK activity, in-hibiting the activation of ERK1/2 and the subsequent phos-phorylation of ERK1/2 substrates both in vitro and in intactcells (16). In Fig. 10A, results from myotubes incubated for20 min with 10 �m PD 98059 are shown. This compoundalmost completely blocked testosterone-induced ERK1/2phosphorylation without an effect on the basal phosphory-lation of this protein. The role of Ras in the signal transduc-tion cascade mediated by androgens was also evaluated us-ing dominant negatives for MEK and Ras proteins. Figure10B shows a selective blockade of testosterone-inducedERK1/2 phosphorylation by dominant negatives for bothMEK and Ras. These results are consistent with ERK1/2phosphorylation induced by testosterone through the Ras/MEK/ERK pathway in cultured myotubes.
Effect of testosterone on the transcription factorCRE-binding protein (CREB)
To further test cyproterone inhibition of genomic effects oftestosterone, myotubes were transfected with a luciferaselinked reporter gene for CRE. Transcription was activated bytestosterone after 12 h (Fig. 11), and the effect was completelyblocked by cyproterone, indicating a genomic effect medi-ated by intracellular androgen receptors. Furthermore, tes-
FIG. 8. Calcium dependence of testosterone-induced phosphoryla-tion of ERK1/2. A, The cells were stimulated with 100 nM testosterone(T) in medium with 3 mM calcium (Ca2�), without extracellular cal-cium (1 mM EGTA, Ca2�-free), or in the presence of BAPTA-AM (achelator of intracellular calcium ions). B, Densitometric analysis ex-presses the percentage of phosphorylation vs. basal and shows anapproximately 20% reduction in a Ca2�-free medium compared withthat in the presence of extracellular Ca2�. C, Cells were preincubatedwith IP3 pathway inhibitors [xestospongin B, 40 �M (xpb); U73122, 10�M]. All values are the mean � SEM of four experiments. *, P � 0.05.
FIG. 9. Effects of GDP�S and PTX on testosterone-induced ERK1/2phosphorylation. A, Myotube incubation with PTX (1 �g/ml for 5 h)or the use of GDP�S (100 nM for 5 min in permeabilized cells) did notaffect basal ERK1/2 phosphorylation, but the phosphorylation in-crease induced by testosterone (T) was blocked. B, Total ERK1/2 asa marker for equivalent protein load is shown. C, Band quantificationis expressed as a percentage of the control value (selected as 100%)from four independent experiments and is the mean � SEM. *, P � 0.05vs. control.
Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes Endocrinology, August 2003, 144(8):3586–3597 3593
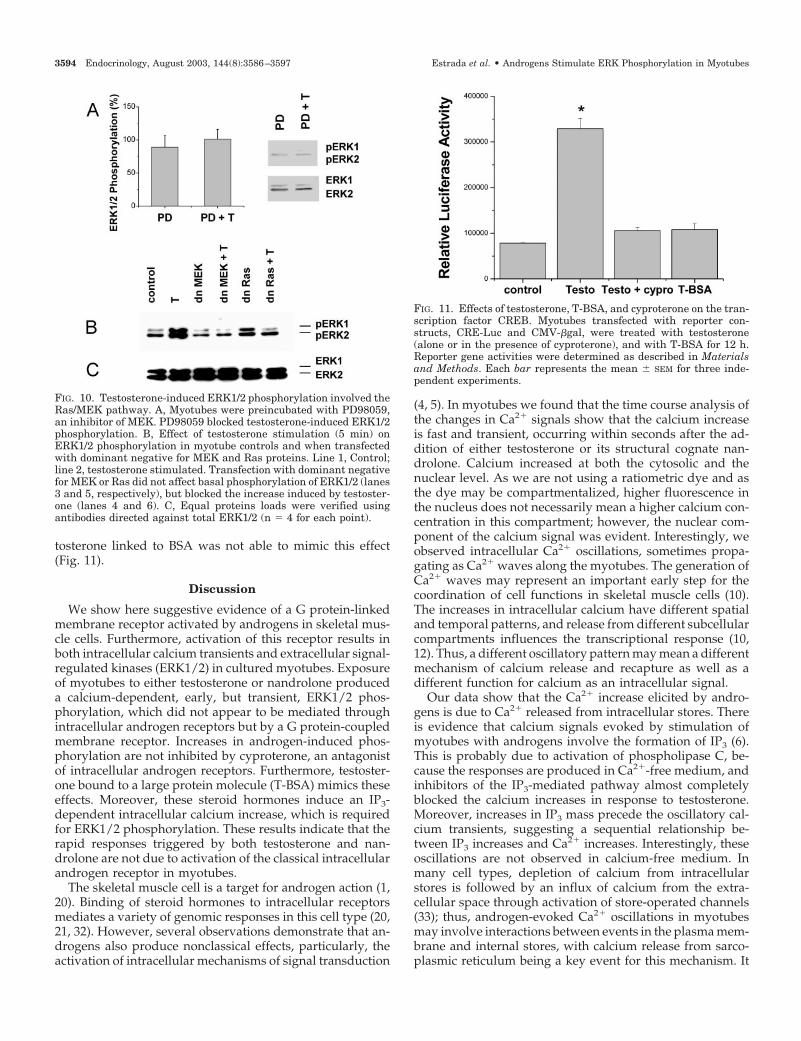
tosterone linked to BSA was not able to mimic this effect(Fig. 11).
Discussion
We show here suggestive evidence of a G protein-linkedmembrane receptor activated by androgens in skeletal mus-cle cells. Furthermore, activation of this receptor results inboth intracellular calcium transients and extracellular signal-regulated kinases (ERK1/2) in cultured myotubes. Exposureof myotubes to either testosterone or nandrolone produceda calcium-dependent, early, but transient, ERK1/2 phos-phorylation, which did not appear to be mediated throughintracellular androgen receptors but by a G protein-coupledmembrane receptor. Increases in androgen-induced phos-phorylation are not inhibited by cyproterone, an antagonistof intracellular androgen receptors. Furthermore, testoster-one bound to a large protein molecule (T-BSA) mimics theseeffects. Moreover, these steroid hormones induce an IP3-dependent intracellular calcium increase, which is requiredfor ERK1/2 phosphorylation. These results indicate that therapid responses triggered by both testosterone and nan-drolone are not due to activation of the classical intracellularandrogen receptor in myotubes.
The skeletal muscle cell is a target for androgen action (1,20). Binding of steroid hormones to intracellular receptorsmediates a variety of genomic responses in this cell type (20,21, 32). However, several observations demonstrate that an-drogens also produce nonclassical effects, particularly, theactivation of intracellular mechanisms of signal transduction
(4, 5). In myotubes we found that the time course analysis ofthe changes in Ca2� signals show that the calcium increaseis fast and transient, occurring within seconds after the ad-dition of either testosterone or its structural cognate nan-drolone. Calcium increased at both the cytosolic and thenuclear level. As we are not using a ratiometric dye and asthe dye may be compartmentalized, higher fluorescence inthe nucleus does not necessarily mean a higher calcium con-centration in this compartment; however, the nuclear com-ponent of the calcium signal was evident. Interestingly, weobserved intracellular Ca2� oscillations, sometimes propa-gating as Ca2� waves along the myotubes. The generation ofCa2� waves may represent an important early step for thecoordination of cell functions in skeletal muscle cells (10).The increases in intracellular calcium have different spatialand temporal patterns, and release from different subcellularcompartments influences the transcriptional response (10,12). Thus, a different oscillatory pattern may mean a differentmechanism of calcium release and recapture as well as adifferent function for calcium as an intracellular signal.
Our data show that the Ca2� increase elicited by andro-gens is due to Ca2� released from intracellular stores. Thereis evidence that calcium signals evoked by stimulation ofmyotubes with androgens involve the formation of IP3 (6).This is probably due to activation of phospholipase C, be-cause the responses are produced in Ca2�-free medium, andinhibitors of the IP3-mediated pathway almost completelyblocked the calcium increases in response to testosterone.Moreover, increases in IP3 mass precede the oscillatory cal-cium transients, suggesting a sequential relationship be-tween IP3 increases and Ca2� increases. Interestingly, theseoscillations are not observed in calcium-free medium. Inmany cell types, depletion of calcium from intracellularstores is followed by an influx of calcium from the extra-cellular space through activation of store-operated channels(33); thus, androgen-evoked Ca2� oscillations in myotubesmay involve interactions between events in the plasma mem-brane and internal stores, with calcium release from sarco-plasmic reticulum being a key event for this mechanism. It
FIG. 11. Effects of testosterone, T-BSA, and cyproterone on the tran-scription factor CREB. Myotubes transfected with reporter con-structs, CRE-Luc and CMV-�gal, were treated with testosterone(alone or in the presence of cyproterone), and with T-BSA for 12 h.Reporter gene activities were determined as described in Materialsand Methods. Each bar represents the mean � SEM for three inde-pendent experiments.
FIG. 10. Testosterone-induced ERK1/2 phosphorylation involved theRas/MEK pathway. A, Myotubes were preincubated with PD98059,an inhibitor of MEK. PD98059 blocked testosterone-induced ERK1/2phosphorylation. B, Effect of testosterone stimulation (5 min) onERK1/2 phosphorylation in myotube controls and when transfectedwith dominant negative for MEK and Ras proteins. Line 1, Control;line 2, testosterone stimulated. Transfection with dominant negativefor MEK or Ras did not affect basal phosphorylation of ERK1/2 (lanes3 and 5, respectively), but blocked the increase induced by testoster-one (lanes 4 and 6). C, Equal proteins loads were verified usingantibodies directed against total ERK1/2 (n � 4 for each point).
3594 Endocrinology, August 2003, 144(8):3586–3597 Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes

has been proposed that the generation and propagation ofintracellular Ca2� oscillations may reduce the thresholds foraction of this second messenger, and different Ca2� oscil-latory patterns can preferentially activate or inactivatecalcium-dependent processes (34).
Testosterone exerts its effects through binding to and ac-tivation of cytosolic androgen receptor, which translocates tothe nucleus and functions as a transcription factor (3, 30). Inour system cyproterone, an antagonist of the intracellularandrogen receptor was able to block both the receptor trans-location and CRE-mediated transcription, events probablymediated by the intracellular androgen receptor. Neverthe-less, androgen-induced second messenger generation wasnot blunted by cyproterone, and the membrane-impermeanttestosterone conjugate (T-BSA) induced similar effects oncalcium compared with the free hormone, suggesting thatthese rapid effects are mediated by the actions of androgenson membrane components. The IP3-mediated calcium re-lease from internal stores can be due to activation of tyrosinekinase as well as G protein-coupled membrane receptors.The treatment of myotubes with genistein, a tyrosine kinaseinhibitor, did not modify the intracellular Ca2� or IP3 in-creases induced by testosterone. On the other hand, severallines of evidence demonstrate that androgens can activatePTX-sensitive G proteins (5, 35); to determine whether tes-tosterone may interact with a G proteins in myotubes, theCa2� increase was evaluated in myotubes treated withGDP�S and PTX. The fact that G protein inhibitors blockedfast effects of testosterone reinforces the idea of a membranereceptor for this hormone. The presence of membrane bind-ing sites for androgens has been previously suggested (4, 5,29). According to our results in myotubes, testosterone-induced intracellular calcium increases have also been de-scribed in other cells, including rat osteoblasts (4), micesplenic T cells (29), and macrophages (5). Interestingly, mac-rophages, lacking classical intracellular androgen receptors,respond to testosterone with intracellular Ca2� increases.Thus, a transient increase in calcium should affect certaincellular processes directly (nongenomic pathway), or alter-natively, they could act through Ca2�-binding proteins toregulate slower processes (genomic pathway) via proteinsynthesis.
Steroid hormones have been associated with the growthand differentiation of muscle cells, but to date little evidencehas accumulated on the role of intracellular signaling path-ways in these processes. Upon activation by phosphoryla-tion, ERK1/2 translocate from the cytoplasm into the nucleus(14), and this results in the phosphorylation or induction oftranscription factors, leading to the expression of genes in-volved in the control of cellular growth. The results reportedhere show that testosterone, nandrolone, and T-BSA caninduce dual phosphorylation of ERK1/2 within a few min-utes. In skeletal muscle an increase in the ERK1/2 phos-phorylation correlates with its activity (15, 19). The rapidphosphorylation of ERK1/2 by androgens was transient anddose dependent, demonstrating that myotubes respond totestosterone in the physiological range of concentrations. Therelationship among intracellular calcium increases, the IP3pathway, and ERK1/2 phosphorylation was illustrated bypharmacologically manipulating the calcium sources. Thus,
testosterone-induced ERK1/2 phosphorylation was slightlyinhibited in a nominally Ca2�-free medium, a possible ex-planation being that the effect of intracellular Ca2� increasesdepends on the time course, i.e. ERK1/2 phosphorylationwill be different depending on whether the calcium increaseoccurs with an oscillatory pattern or not. A similar effect hasbeen suggested for the effects of ouabain, a steroid deriva-tive, on the transcription factor nuclear factor-�B, whichreaches maximal activation during calcium oscillations, butnot during maintained Ca2� increase (36). On the other hand,steroid-induced ERK1/2 phosphorylation was completelyblocked by chelation of intracellular calcium withBAPTA-AM or the use of either U-73122, a PLC inhibitor, orxestospongin B, an inhibitor of IP3 receptors. Moreover, Gprotein-coupled receptors can induce ERK1/2 activation (13,14). Testosterone-induced ERK1/2 phosphorylation in themyotubes was inhibited by treatment with GDP�S and PTX,suggesting the participation of a PTX-sensitive G protein inthis response. These results are in accordance with the in-hibition of intracellular Ca2� and IP3 increases by the Gprotein inhibitors and the Ca2� requirement for the ERK1/2phosphorylation induced by testosterone in myotubes.
Some researchers have shown that antagonists for intra-cellular receptors can block steroid-induced ERK1/2 phos-phorylation (37). However, after incubation of myotubeswith 1 �m cyproterone in the present study, neither basalERK1/2 phosphorylation nor testosterone-dependent phos-phorylation was modified. Even though these effects werenot blocked by cyproterone in skeletal muscle cells, whichsuggests that activation of the transcription machinery wasnot involved, nongenomic effects may play a physiologicalrole in events previous to the androgen receptor activation.A likely possibility is phosphorylation of coactivators thatamplify the androgen action, as has been proposed by Zhuet al. (38), or a biphasic system with both high affinity nuclearas well as low affinity membrane androgen receptors, asdescribed in rat Sertoli and human prostatic cell lines (39).Several mitogens can produce trans-activation of steroid re-ceptors through phosphorylation (40, 41). An interesting hy-pothesis to be explored is that intracellular calcium increasesand ERK1/2 activation are events that elicit androgen re-ceptor activation in skeletal muscle cells. Moreover, theMAPK cascade triggered by the binding of hormones orgrowth factors to surface receptors may produce the phos-phorylation events that have been implicated in cell growthand differentiation (18). Gredinger et al. (16) have shown thatthe MAPK pathway plays a positive role in myogenesis andthat ERK activity is substantially increased during terminalmuscle differentiation in muscle cells and cooperates withMyoD to activate muscle-specific transcription. In accor-dance with the rapid effects of steroid hormones, Morelli etal. (23) have recently shown that 1,25-dihydroxyvitamin D3activates ERK1/2 in skeletal muscle cells and has implicatedthe MAPK cascade in hormone control of myoblast prolif-eration. ERK1/2 are activated by phosphorylation of boththreonine and tyrosine residues; these reactions are catalyzedby MEK. MEK itself is activated by at least two types ofkinases, Raf-1 and MEK kinase (13, 14, 18). Raf-1, which isalso activated by phosphorylation, appears to be a centralpoint in receiving signals from upstream activated kinases,
Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes Endocrinology, August 2003, 144(8):3586–3597 3595

located at the plasma membrane. Most of the signaling path-ways that have been proposed to be involved in the activa-tion of ERK1/2 after increasing intracellular Ca2� convergeon Ras, thus indicating a key role for this protein in thisprocess (14, 18). The observations that overexpression ofdominant negative mutants of Ras and MEK inhibits testos-terone-mediated ERK1/2 phosphorylation reflects that thishormone activates the Ras/MEK/ERK pathway in myo-tubes. The mechanism of Ca2�-mediated Ras activation re-mains elusive; however, in other cell types, a Ras-dependentactivation of ERK1/2 by either PKC (18) or Ca2�-regulatedprotein kinase such as calmodulin (12) has been described.Buitrago et al. (42) recently demonstrated in avian myotubesthat 1�,25-dihydroxyvitamin D3-induced MAPK activationoccurs through both a Raf-1 via Ras pathway as well as aPKC�-dependent pathway. The intracellular targets for thesteroid-stimulated ERK1/2 cascade have not been estab-lished. It has been reported that androgens can activate thetranscription factor CREB (43). However, in myotubes thetranscriptional CREB activity induced by testosterone, butnot by T-BSA, indicates that a hormone-bound intracellularandrogen receptor is necessary for this effect. There is evi-dence that activation of several signaling pathways involv-ing protein kinase A and C leads to stimulation of androgen-regulated trans-activation through interaction of componentsof these pathways with the androgen receptor (41). It ispossible, then, that the interaction of activated androgenreceptor with some of the signaling pathways will eventuallylead to activation of the element-binding protein by phos-phorylation (43). A possible role for the ERK pathway wouldbe to phosphorylate hormone-bound androgen receptors(44) and modulate transcriptional activity (22, 30). Anotherpossibility could be that ERK-activated pathways could in-fluence the genomic response through a synergistic mecha-nism by phosphorylation of coactivators of the intracellularreceptor at the nuclear level, as reported for both thyroidreceptor and estrogen receptor (45, 46).
A scheme of the proposed G protein-dependent action forandrogens in skeletal muscle cells is depicted (Fig. 12). Itincludes a putative membrane androgen receptor coupled toPTX-sensitive G protein, PLC activation, and calcium release;ERK1/2 phosphorylation appears to be triggered by calcium.Taken together, our results indicate that increases in intra-cellular calcium as well as activation of the Ras/MEK/ERK1/2 pathway represent an intermediate step in the in-tracellular signaling toward as yet undefined downstreameffects of androgen steroids on skeletal muscle cells.
Acknowledgments
Received December 18, 2002. Accepted May 1, 2003.Address all correspondence and requests for reprints to: Dr. Enrique
Jaimovich, Instituto de Ciencias Biomedicas, Facultad de Medicina, Uni-versidad de Chile, Independencia 1027, Casilla 70005, Santiago 6530499,Chile. E-mail: [email protected].
This work was supported by Fondo Nacional de Ciencia y TecnologıaGrants 15010006 (to E.J.) and 2000-055 (to M.E.). M.E. is the recipient ofa graduate student fellowship from the Comision Nacional de Investi-gacion Cientıfica y Tecnologica and Instituto de Ciencias Biomedicas.
References
1. Powers ML, Florini JR 1975 A direct effect of testosterone on muscle cells intissue culture. Endocrinology 97:1043–1047
2. Michel G, Baulieu E 1980 Androgen receptor in rat skeletal muscle: charac-terization and physiological variations. Endocrinology 107:2088–2098
3. Simental JA, Sar M, Lane MV, French FS, Wilson EM 1991 Transcriptionalactivation and nuclear targeting signals of the human androgen receptor. J BiolChem 266:510–518
4. Lieberherr M, Grosse B 1994 Androgens increase intracellular calcium con-centration and inositol 1,4,5-trisphosphate and diacylglycerol formation via apertussis toxin-sensitive G-protein. J Biol Chem 269:7217–7223
5. Benten WP, Lieberherr M, Stamm O, Wrehlke C, Guo Z, Wunderlich F 1999Testosterone signaling through internalizable surface receptors in androgenreceptor-free macrophages. Mol Biol Cell 10:3113–3123
6. Estrada M, Liberona JL, Miranda M, Jaimovich E 2000 Aldosterone- andtestosterone-mediated intracellular calcium response in skeletal muscle cellcultures. Am J Physiol 279:E132–E139
7. Wehling M 1997 Specific, nongenomic actions of steroid hormones. Annu RevPhysiol 59:365–393
8. Powell JA, Carrasco MA, Adams D, Drouet B, Rios J, Muller M, Estrada M,Jaimovich E 2001 IP3 receptor function and localization in myotubes: anunexplored Ca2� signaling pathway in skeletal muscle. J Cell Sci 114:3673–3683
9. Araya R, Liberona JL, Cardenas JC, Riveros N, Estrada M, Powell J, CarrascoMA, Jaimovich E 2003 Dihydropyridine receptors as voltage sensors for adepolarization-evoked, IP3R-mediated, slow calcium signal in skeletal musclecells. J Gen Physiol 121:3–16
10. Semsarian C, Wu M, Ju Y, Marciniec T, Yeoh T, Allen D, Harvey R, GrahamRM 1999 Skeletal muscle hypertrophy is mediated by a Ca2�-dependent cal-cineurin signalling pathway. Nature 400:576–580
11. Estrada M, Cardenas C, Liberona JL, Carrasco MA, Mignery GA, Allen PD,Jaimovich E 2001 Calcium transients in 1B5 myotubes lacking ryanodinereceptors are related to inositol trisphosphate receptors. J Biol Chem 276:22868–22874
12. Cruzalegui FH, Bading H 2000 Calcium-regulated protein kinase cascades andtheir transcription factor targets. Cell Mol Life Sci 57:402–410
13. Blenis J 1993 Signal transduction via the MAP kinase: proceed at your ownRSK. Proc Natl Acad Sci USA 90:5889–5892
14. Seger R, Krebs EG 1995 The MAPK signaling cascade. FASEB J 9:726–73515. Aronson D, Violan MA, Dufresne SD, Zangen D, Fielding RA, Goodyear LJ
1997 Exercise stimulates the mitogen-activated protein kinase pathway inhuman skeletal muscle. J Clin Invest 99:1251–1257
16. Gredinger E, Gerber AN, Tamir Y, Tapscott SJ, Bengal E 1998 Mitogen-activated protein kinase pathway is involved in the differentiation of musclecells. J Biol Chem 273:10436–10444
17. Napoli R, Gibson L, Hirshman MF, Boppart MD, Dufresne SD, Horton ES,Goodyear LJ 1999 Epinephrine and insulin stimulate different mitogen-acti-vated protein kinase signaling pathways in rat skeletal muscle. Diabetes 47:1549–1554
18. Garrington TP, Johnson GL 1999 Organization and regulation of mitogen-activated protein kinase signaling pathways. Curr Opin Cell Biol 11:211–218
19. Aronson D, Wojtaszewski JF, Thorell A, Nygren J, Zangen D, Richter EA,Ljungqvist O, Fielding RA, Goodyear LJ 1998 Extracellular-regulated protein
FIG. 12. Scheme for the fast effects of testosterone in myotubes. PLCactivation, through a putative membrane androgen receptor (AR)coupled to a PTX-sensitive G protein, increases IP3 levels, which, inturn, produces intracellular Ca2� increases. This increase in cytosoliccalcium may be through a Ca2�-dependent protein kinase; it canregulate Ras activity at the plasma membrane level and will initiatea signaling cascade leading to ERK1/2 phosphorylation.
3596 Endocrinology, August 2003, 144(8):3586–3597 Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes

kinase cascades are activated in response to injury in human skeletal muscle.Am J Physiol 275:C555–C561
20. Mooradian AD, Morley JE, Korenman SG 1987 Biological actions of andro-gens. Endocr Rev 8:1–28
21. Sheffield-Moore M, Urban R, Wolf S, Jiang J, Catlin D, Herndon D, WolfeR, Ferrando A 1999 Short-term oxandrolone administration stimulates netmuscle protein synthesis in young men. J Clin Endocrinol Metab 84:2705–2711
22. Watters JJ, Campbell JS, Conningham MJ, Krebs EG, Dorsa DM 1997 Rapidmembrane effects of steroids in neuroblastoma cells: effects of estrogen onmitogen activated protein kinase signaling cascade and c-fos immediate earlygene transcription. Endocrinology 138:4030–4033
23. Morelli S, Buitrago C, Vasquez G, de Boland A, Boland R 2000 Involvementof tyrosine kinase activity in 1�,25(OH)2-vitamin D3 signal transduction inskeletal muscle cells. J Biol Chem 275:36021–36028
24. Improta-Brears T, Whorton AR, Codazzi F, York JD, Meyer T, McDonnell DP1999 Estrogen-induced activation of mitogen-activated protein kinase requiresmobilization of intracellular calcium. Proc Natl Acad Sci USA 96:4686–4691
25. Castoria G, Barone MV, Di Domenico M, Bilancio A, Ametrano D, Migli-accio A, Auricchio F 1999 Non-transcriptional action of oestradiol and pro-gestin triggers DNA synthesis. EMBO J 18:2500–2510
26. Peterziel H, Mink S, Shonert A, Becker M, Klocker H, Cato ACB 1999 Rapidsignalling by androgen receptor in prostate cancer cells. Oncogene 18:6322–6329
27. Jaimovich E, Liberona JL, Reyes R, Powell JA 2000 Nuclear IP3 receptors, IP3transients and nucleus-associated calcium signals in cultured skeletal muscle.Am J Physiol 278:C998–C1010
28. Minta A, Kao JPY, Tsien RY 1986 Fluorescent indicators for cytosolic calciumbased on rhodamine and fluorescent chromophores. J Biol Chem 264:8171–8178
29. Benten WP, Lieberherr M, Giese G, Wrehlke C, Stamm O, Sekeris CE,Mossmann H, Wunderlich F 1999 Functional testosterone receptors in plasmamembranes of T cells. FASEB J 13:123–133
30. Cato AC, Peterziel H 1998 The androgen receptor as mediator of gene ex-pression and signal transduction pathways. Trends Endrocrinol Metab 4:150–154
31. Kuil CW, Mulder E 1996 Deoxyribonucleic acid-binding ability of androgenreceptors in whole cells: implications for the actions of androgens and anti-androgens. Endocrinology 137:1870–1877
32. Lucas PC, Granner DK 1992 Hormone response domains in gene transcription.Annu Rev Biochem 61:1131–1173
33. Putney Jr JW, Broad LM, Braun FJ, Lievremont JP, Bird GS 2001 Mechanismsof capacitative calcium entry. J Cell Sci 114:2223–2229
34. Dolmetsch RE, Xu K, Lewis RS 1998 Calcium oscillations increase the effi-ciency and specificity of gene expression. Nature 392:933–936
35. Liu D, Dillon JS 2002 Dehydroepiandrosterone activates endothelial cell ni-tric-oxide synthase by a specific plasma membrane receptor coupled to G�i2,3. J Biol Chem 277:21379–21388.
36. Aizman O, Uhlen P, Lal M, Brismar H, Aperia A 2001 Ouabain, a steroidhormone that signals with slow calcium oscillations. Proc Natl Acad Sci USA98:13420–13424
37. Migliaccio A, Castoria G, Di Domenico M, de Falco A, Bilancio A, LombardiM, Barone MV, Ametrano D, Zannini MS, Abbondanza C, Auricchio F 2000Steroid-induced androgen receptor-oestradiol receptor �-Src complex triggersprostate cancer cell proliferation. EMBO J 19:5406–5417
38. Zhu X, Liu JP 1997 Steroid-independent activation of androgen receptor inandrogen-independent prostate cancer. A possible role for the MAP kinasesignal transduction pathway? Mol Cell Endocrinol 134:9–14
39. Lyng FM, Jones GR, Rommerts FFG 2000 Rapid androgen actions on calciumsignaling in rat sertoli cells and two human prostatic cell lines: similar biphasicresponses between 1 picomolar and 100 nanomolar concentrations. Biol Re-prod 63:736–747
40. Nordeen SK, Moyer ML, Bona BJ 1994 The coupling of multiple signaltransduction pathways with steroid response mechanisms. Endocrinology134:1723–1732
41. Reinikainen P, Palvimo J, Janne OA 1996 Effects of mitogens on androgenreceptor-mediated transactivation. Endocrinology 137:4351–4357
42. Buitrago CG, Gonzalez V, de Boland AR, Boland R 2003 Activation of raf-1through ras and PKC� mediates 1�,25(OH)2-vitamin D3 regulation of themitogen-activated protein kinase pathway in muscle cells. J Biol Chem 278:2199–2205
43. Kim J-G, Amano O, Wakayama T, Takahagi H, Iseki S 2001 The role of cyclicAMP response element-binding protein in testosterone-induced differentia-tion of granular convoluted tubule cells in the rat submandibular gland. ArchOral Biol 46:495–507
44. Ueda T, Bruchovsky N, Sadar MD 2002 Activation of the androgen receptorN-terminal domain by interleukin-6 via MAPK and STAT3 signal transductionpathways. J Biol Chem 277:7076–7085
45. Davis PJ, Shih A, Lin H, Martino LJ, Davis FB 2000 Thyroxine promotesassociation of mitogen-activated protein kinase and nuclear thyroid hormonereceptor (TR) and causes serine phosphorylation of TR. J Biol Chem 275:38032–38039
46. Feng W, Webb P, Nguyen P, Liu X, Li J, Karin M, Kushner PJ 2001 Poten-tiation of estrogen receptor activation function 1 (AF-1) by Src/JNK througha serine 118-independent pathway. Mol Endocrinol 15:32–45
Estrada et al. • Androgens Stimulate ERK Phosphorylation in Myotubes Endocrinology, August 2003, 144(8):3586–3597 3597




















