
Late-glacial recolonization and phylogeography of European red deer ( Cervus elaphus L.)
Upload
independentCategory
view
1download
0
Bone 44 (2009) 691–698
Contents lists available at ScienceDirect
Bone
j ourna l homepage: www.e lsev ie r.com/ locate /bone
Testosterone, but not IGF-1, LH, prolactin or cortisol, may serve as antler-stimulatinghormone in red deer stags (Cervus elaphus)☆
Luděk Bartoš a,⁎, Dieter Schams b, George A. Bubenik c
a Department of Ethology, Institute of Animal Science, Přátelství 815, 106 00 Praha 10-Uhříněves, Czech Republicb Institute of Physiology, University of Munich-Weihenstephan, Freising-Weihenstephan, Germanyc University of Guelph, Guelph, Ontario, Canada
☆ Sources of support: Funded by grants from the Cznumber 523/08/0808) andMinistry of Agriculture of theMZe 0002701402 and MZe 0002701404). All authors ha⁎ Corresponding author. Fax: +420 267 710 779.
E-mail addresses: [email protected] (L. Bartoš), [email protected] (G.A. Bubenik).
8756-3282/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.bone.2008.12.004
a b s t r a c t
a r t i c l e i n f oArticle history:
The role of androgens and Received 1 October 2008Revised 12 November 2008Accepted 5 December 2008Available online 16 December 2008Edited by: T. Matsumoto
Keywords:Antler growthTestosteroneIGF-1Red deerCervus elaphus
insulin-like growth factor 1 (IGF-1) in antler growth has been disputed. Wepredicted that the secretory of IGF-1 may be associated with an acceleration of body growth rather than withantler growth. Furthermore we anticipated a relationship between the increase of testosterone and theprogress of antler growth. If IGF-1 is involved in the stimulation of antler growth, this should be moreobvious in young than in mature stags. Eight two-year-old red deer stags (Cervus elaphus), and twelve adultred deer stags were blood sampled and the length of their velvet antlers was measured in one-week intervalsduring the period of antler growth. Concentrations of testosterone, cortisol, IGF-1, luteinizing hormone (LH),and prolactin were determined in plasma by enzyme immunoassay or radioimmunoassay. Antler growth perday was primarily dependent on changes in testosterone concentration per day in both groups of stags. Asexpected, only in two-year-old stags we detected a possible role of IGF-1 in the antler growth regulation, butthat was not in agreement with previously published studies. Nevertheless, this effect was still utilized ininteraction with testosterone. In addition to total antler length, only concentrations of testosterone and LHwere significantly higher in adult males in comparison to two-year-old males. Our present results lead us toconclude that it is not IGF-1 but testosterone which is responsible for the intensity of antler growth insubadult and adult red deer stags.
© 2008 Elsevier Inc. All rights reserved.
Introduction
The regeneration and growth of deer antlers has been proposed asa model for biomedical research in several publications [1,2]. Asshown previously [3,4], understanding of the processes in antlers maybe helpful in human osteological research. Role of steroids, testoster-one and estrogen in particular, in bone biology seems to be of specialimportance in elderly people. Generally, testosterone promotessecondary sexual characters across many taxa [5]. The observedevolutionary conservation in the regulation of traits induced bytestosterone in male vertebrates suggests that this system evolves as acomplex, with the linkage between testosterone and male traits beinginseparable [6]. In this respect, some authors suggest, that antlers, astypical male secondary sexual characters, should be controlledotherwise. When Suttie et al. [7, 8] compared seasonal variations ofhormones with the progress of antler growth in red deer, Cervus
ech Science Foundation (grantCzech Republic (grant numbersve no conflicts of interest.
@wzw.tum.de (D. Schams),
l rights reserved.
elaphus, they concluded that insulin-like growth factor 1 (IGF-1) is anantler-stimulating hormone. This hypothesis replaced an earliernotion which suggests that the antler-stimulating hormones areandrogens, particularly testosterone or its derivatives [9–12]. Tofurther support the role of IGF-1 as the antler-stimulating hormone, anumber of in vivo and in vitro studies have been performed [13–17]. Asimilar increase of IGF-1 concentrations corresponding to the progressof antler growthwas also reported in roe deer, Capreolus capreolus [18]and pudu, Pudu puda [19]. Nevertheless, in a later study we havedemonstrated that in pudu the increase of IGF-1 is found only in thedominant males but not in the subordinate ones [20]. Data on maleand female reindeer, Rangifer tarandus [21,22] and on castrated fallowdeer, Dama dama [23] did not confirm Suttie et al.'s [7] conclusionseither. In another study with yearling red deer stags, Suttie et al. [24]reported that IGF-1 correlated positively with antler growth rate, butthe pattern of the hormone shown in their Fig. 1 raises somequestions. Themost pronounced discrepancywas apparent during thelast two months of the antler growth period presented in the samefigure in that study [24], when IGF-1 concentrations were sharplydecreasing, while antlers were still growing. Similar discrepancy maybe found also in a study on reindeer calves [21]. Thus, the roleof androgens and IGF-1 in antler growth remains disputed. IGF-1is generally an important component of the growth process and a

Fig. 1. Total antler length (means±S.E.) of two-year-old and adult red deer males.
692 L. Bartoš et al. / Bone 44 (2009) 691–698
certain role in antler growth seems very plausible [25]. However, therole of IGF-1 as “the” principle hormone responsible for controllingantler growth is controversial. Otherwise all reports examining IGF-1time course during the period of antler growth should be consistent.One of the reasons for the inconsistency in the reports may be the ageof the subjects. In their first experiment Suttie et al. [7] used yearlingstags. Also all other studies providing information on function andregulation of IGF-1 in red deer have been made on calves [24,26–30].Yearling red deer are not yet mature and their antler growth occurs atthe time of the accelerated body growth [31,32]. IGF-1 correlatesstrongly with body growth [33]. Hence, we can hypothesize that theIGF-1 pattern may be basically associated with the accelerated bodygrowth rather than with the antler growth. As such, an effect of thehormones on the antler growth should be tested in principle inmaturemales with finished body growth and not in still growing yearlings. Inaddition increasing IGF-1 concentrations associated with body growthspurt in yearlings could mimic association with antler enlargementspurting at the same time. Therefore, we expected to find differencesin the IGF-1 time courses of young and mature stags as they relate toantler growth. Furthermorewe anticipated a relationship between theincrease in testosterone and the progress of antler growth. If IGF-1 isinvolved in the stimulation of antler growth, this should be moreobvious in young, than in mature stags. Finally, taking into account anincrease in the antler length with age [34], an increase in antler sizeshould be associated with a comparable increase in concentrations ofhormones responsible for antler growth.
Material and methods
Animals and procedures
Eight two-year-old red deer stags (Cervus elaphus) and twelve stags,age 4 to 9 years (adults), were kept in a paddock at the animal facility ofthe Institute of Animal Science (former Research Institute of AnimalProduction). The deer were kept in a group outdoors and were fed ongrass supplementedwith barley (0.75 kg/animal/day) and hay. Alwaysat the same time of day they were handled in a crush and bloodsampled and the length of their velvet antlers was measured using aflexible measuring tape. Deer were sampled in one week intervalsranging from the time prior the antler casting to the end of antlergrowth and velvet shedding. Left and right antlers weremeasured andmean values of the antler lengths from both sides entered the analysis.
Blood samples were collected using VENOSAFE™ blood collectionsystem (TERUMO Europe N. V.).
Hormone analyses
Concentrations of testosterone, cortisol, IGF-1, and luteinizinghormone (LH) were determined in plasma by enzyme immunoassay(EIA) or radioimmunoassay (RIA). For each hormone, all samples wereassayed at the same time.
Testosterone was measured by EIA [35] with a double antibodytechnique after extraction of 1 ml plasma with 3 ml tertiarybutylmethylether/petrolether 30/70 (v/v). After freezing, the solventfraction was decanted and evaporated in a water bath at 60 °C. Theextract was dissolved in 100 μl assay buffer, and 20 μl/tube wereanalyzed. The assay used a polyclonal antibody raised in a rabbitagainst testosterone-11-hemisuccinate-BSA, and the label was testos-terone-3-carboxy methyl-oxime-horseradish peroxidase. Cross reac-tivities to other steroids were as follows: testosterone 100%, 5a-dihydrotestosterone 10%, androstendione 2%, oestradiol b0.1% andprogesterone b0.1%. The standard curve ranged from 0.2–25 pg/tube,corresponding to a sensitivity of 10 pg/ml plasma or 1 pg/ml plasmaafter concentration (dissolving in 100 μl buffer only). The ED50 of thestandard curve was 2 pg/tube. The intra- and inter-assay coefficientsof variation were an average 6.7% and 10.5% respectively.
IGF-1 was determined by RIA after acid–ethanol extraction of 50 μlof blood plasma [18]. Recombinant human IGF-1 (Lilly Company, SaintLouis, USA) was used for iodination according to the lactoperoxidasemethod and as standard. Separation of bound and free hormone wasaccomplished by a combination of second-antibody techniquewith 4%polyethylene glycol. The assay was validated for red deer plasma byrecovery studies (IGF-1 added to plasma samples before extraction)and was an average 89% and complete parallel displacement wasobserved between extracted samples and the standard curve. Intra-and inter-assay CVs were b10% and b16%, respectively.
Cortisol (4-pregnene-11β-17α-21-triol-3, 20-dione, Steraloids,Wilton,N. H., USA) concentrations were measured with a competitive EIA [36].The antibody was raised in a rabbit against cortisol-21-hemisuccinate-BSA. Cross reactivities were: cortisol 100%, cortisone 8%, corticosterone9.5%, prednisolone18%. Bloodplasmawas extractedwith a 10 fold volumeof tert-butylmethyl ether.After freezingat–60 °C, extractsweredecanted,evaporated and reconstituted. Interassay CV was between 7 and 14%.
LH was determined by a homologous bovine assay with no crossreactivity to other pituitary hormones [37,38]. The reference prepara-tion was a pure bovine pituitary extract prepared in our laboratory(LH-DSA with a biological activity of 1.0 times NIH-LH-S1). The assaywas validated for fallow deer plasma by recovery studies of 4 differentconcentrations added to plasma. Average recovery was 96.4±6.4%.

Fig. 2. Concentrations of testosterone, IGF-1, LH, cortisol, and prolactin (means±S.E.) in the period from the day of antler casting to the end of antler growth.
693L. Bartoš et al. / Bone 44 (2009) 691–698
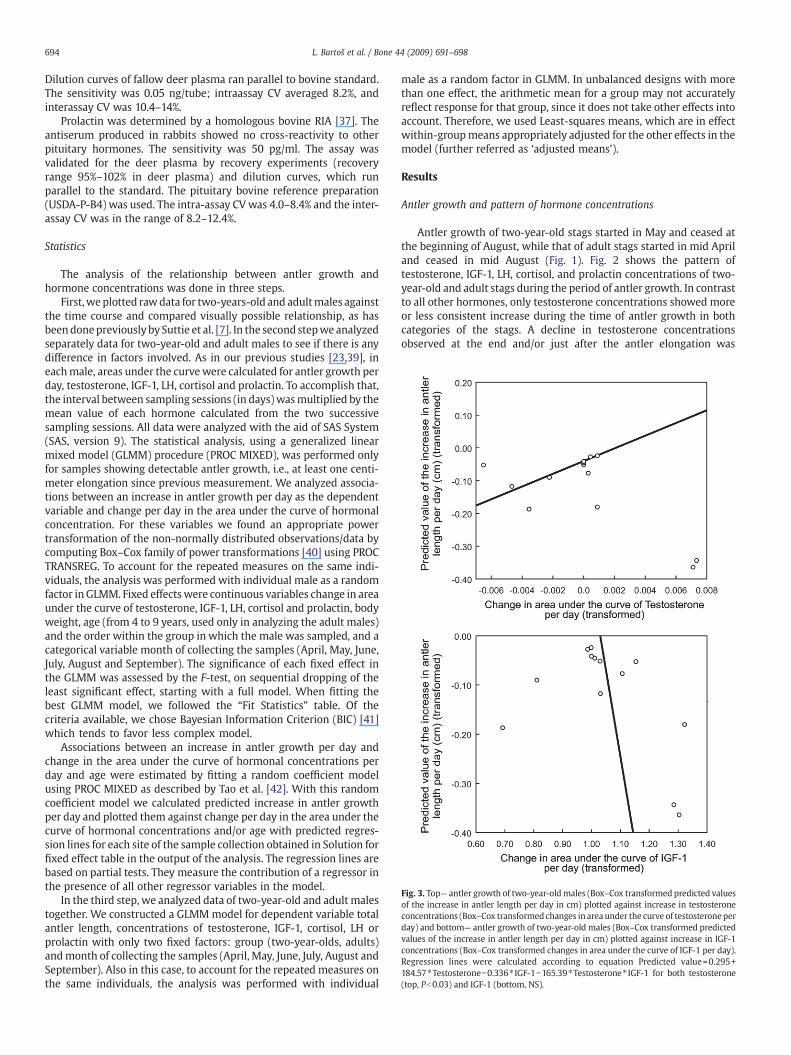
Fig. 3. Top— antler growth of two-year-oldmales (Box–Cox transformed predicted valuesof the increase in antler length per day in cm) plotted against increase in testosteroneconcentrations (Box–Cox transformedchanges in area under the curve of testosteroneperday) and bottom— antler growth of two-year-old males (Box–Cox transformed predictedvalues of the increase in antler length per day in cm) plotted against increase in IGF-1concentrations (Box–Cox transformed changes in area under the curve of IGF-1 per day).Regression lines were calculated according to equation Predicted value=0.295+184.57⁎Testosterone−0.336⁎ IGF-1−165.39⁎Testosterone⁎ IGF-1 for both testosterone(top, Pb0.03) and IGF-1 (bottom, NS).
694 L. Bartoš et al. / Bone 44 (2009) 691–698
Dilution curves of fallow deer plasma ran parallel to bovine standard.The sensitivity was 0.05 ng/tube; intraassay CV averaged 8.2%, andinterassay CV was 10.4–14%.
Prolactin was determined by a homologous bovine RIA [37]. Theantiserum produced in rabbits showed no cross-reactivity to otherpituitary hormones. The sensitivity was 50 pg/ml. The assay wasvalidated for the deer plasma by recovery experiments (recoveryrange 95%–102% in deer plasma) and dilution curves, which runparallel to the standard. The pituitary bovine reference preparation(USDA-P-B4) was used. The intra-assay CVwas 4.0–8.4% and the inter-assay CV was in the range of 8.2–12.4%.
Statistics
The analysis of the relationship between antler growth andhormone concentrations was done in three steps.
First,weplotted rawdata for two-years-old and adultmales againstthe time course and compared visually possible relationship, as hasbeendonepreviously by Suttie et al. [7]. In the second stepwe analyzedseparately data for two-year-old and adult males to see if there is anydifference in factors involved. As in our previous studies [23,39], ineachmale, areas under the curvewere calculated for antler growth perday, testosterone, IGF-1, LH, cortisol and prolactin. To accomplish that,the interval between sampling sessions (in days)wasmultiplied by themean value of each hormone calculated from the two successivesampling sessions. All data were analyzed with the aid of SAS System(SAS, version 9). The statistical analysis, using a generalized linearmixed model (GLMM) procedure (PROC MIXED), was performed onlyfor samples showing detectable antler growth, i.e., at least one centi-meter elongation since previous measurement. We analyzed associa-tions between an increase in antler growth per day as the dependentvariable and change per day in the area under the curve of hormonalconcentration. For these variables we found an appropriate powertransformation of the non-normally distributed observations/data bycomputing Box–Cox family of power transformations [40] using PROCTRANSREG. To account for the repeated measures on the same indi-viduals, the analysis was performed with individual male as a randomfactor in GLMM. Fixed effectswere continuous variables change in areaunder the curve of testosterone, IGF-1, LH, cortisol and prolactin, bodyweight, age (from 4 to 9 years, used only in analyzing the adult males)and the order within the group in which the male was sampled, and acategorical variable month of collecting the samples (April, May, June,July, August and September). The significance of each fixed effect inthe GLMM was assessed by the F-test, on sequential dropping of theleast significant effect, starting with a full model. When fitting thebest GLMM model, we followed the “Fit Statistics” table. Of thecriteria available, we chose Bayesian Information Criterion (BIC) [41]which tends to favor less complex model.
Associations between an increase in antler growth per day andchange in the area under the curve of hormonal concentrations perday and age were estimated by fitting a random coefficient modelusing PROC MIXED as described by Tao et al. [42]. With this randomcoefficient model we calculated predicted increase in antler growthper day and plotted them against change per day in the area under thecurve of hormonal concentrations and/or age with predicted regres-sion lines for each site of the sample collection obtained in Solution forfixed effect table in the output of the analysis. The regression lines arebased on partial tests. They measure the contribution of a regressor inthe presence of all other regressor variables in the model.
In the third step, we analyzed data of two-year-old and adult malestogether. We constructed a GLMM model for dependent variable totalantler length, concentrations of testosterone, IGF-1, cortisol, LH orprolactin with only two fixed factors: group (two-year-olds, adults)andmonth of collecting the samples (April, May, June, July, August andSeptember). Also in this case, to account for the repeated measures onthe same individuals, the analysis was performed with individual
male as a random factor in GLMM. In unbalanced designs with morethan one effect, the arithmetic mean for a group may not accuratelyreflect response for that group, since it does not take other effects intoaccount. Therefore, we used Least-squares means, which are in effectwithin-groupmeans appropriately adjusted for the other effects in themodel (further referred as ‘adjusted means’).
Results
Antler growth and pattern of hormone concentrations
Antler growth of two-year-old stags started in May and ceased atthe beginning of August, while that of adult stags started in mid Apriland ceased in mid August (Fig. 1). Fig. 2 shows the pattern oftestosterone, IGF-1, LH, cortisol, and prolactin concentrations of two-year-old and adult stags during the period of antler growth. In contrastto all other hormones, only testosterone concentrations showed moreor less consistent increase during the time of antler growth in bothcategories of the stags. A decline in testosterone concentrationsobserved at the end and/or just after the antler elongation was

Fig. 4. Antler growth of two-year-old males (Box–Cox transformed predicted values of the increase in antler length per day in cm) plotted against interaction between increase intestosterone concentrations (Box–Cox transformed changes in area under the curve of testosterone per day) and increase in IGF-1 concentrations (Box–Cox transformed changes inarea under the curve of IGF-1 per day).
695L. Bartoš et al. / Bone 44 (2009) 691–698
completed, was followed by another steady increase of testosteroneconcentrations (not shown here).
Relationship between antler growth and hormones in two-year-olds
The GLMM model showed a significant effect of changes intestosterone concentration per day (GLMM, F(1, 18)=5.57, Pb0.03),non-significant changes in IGF-1 concentration per day (GLMM, F(1, 18)=1.84, NS) and again, a significant effect of the interaction between thechanges in testosterone and IGF-1 concentrations per day (GLMM, F(3,42)=6.64, Pb0.02). Antler growth (an increase of antler length per day)was positively dependent on changes in testosterone concentrationper day (Fig. 3 top). The potentially negative dependency of antlergrowth on increase in IGF-1 shown in Fig. 3 (bottom) was notsignificant. Fig. 4 displays the relationship between antler growth and
Fig. 5. Antler growth of adult males (Box–Cox transformed predicted values of theincrease in antler length per day in cm) plotted against increase in testosteroneconcentrations (Box–Cox transformed changes in area under the curve of testosteroneper day).
the interaction between testosterone and IGF-1 concentrationsdemonstrating that antler growth was slowest with the highestconcentrations of the two hormones when effect of testosterone as asingle factor was statistically significant. There was no othersignificant GLMM model which would contain testosterone and anyother single hormone.
Relationship between antler growth and hormones in adults
In adults, we fitted the final GLMM model containing fixed factorschanges in testosterone concentrations, age and the month whensamples were taken. Antler growth per day was dependent onchanges in testosterone concentration per day (Fig. 5, F(1, 42)=15.94,Pb0.001), the month when samples were taken (F(4, 42)=11.06,Pb0.001) and the interaction between age of the male and changesin testosterone concentration per day (Fig. 6, F(3, 42)=9.56, Pb0.001).Antler growth was not dependent on any other hormone involved,either alone or in interaction with other factors.
Comparison between two-year-old and adult males in antlerlength and hormonal concentrations Fig. 7 shows adjusted means(±S.E.) of total antler length (F(1, 20)=22.78, Pb0.001) and overallconcentrations of testosterone (F(1, 20)=15.58, Pb0.001), IGF-1 (F(1, 20)=0.10, NS), LH (F(1, 20)=11.19, Pb0.001), cortisol (F(1, 20)=0.29, NS) andprolactin (F(1, 20)=0.76, NS). Apart from the total antler length,only concentrations of testosterone and LH, but not of IGF-1 orprolactin, were significantly higher in adult males than in thetwo-year-old males.
Discussion
In both age categories it was basically the change in testosteroneconcentrations which was positively associated with antler growth.This is in full agreement with our previous results on castrated fallowdeer [23,39]. In Iberian red deer the most precocious males in antlergrowth also showed higher levels of testosterone during the first andsecond year of age [43]. Moreover, it is also coherent with studies onmice and humans. Androgen receptor activation dominated normaltrabecular bone development and cortical bone modeling in male
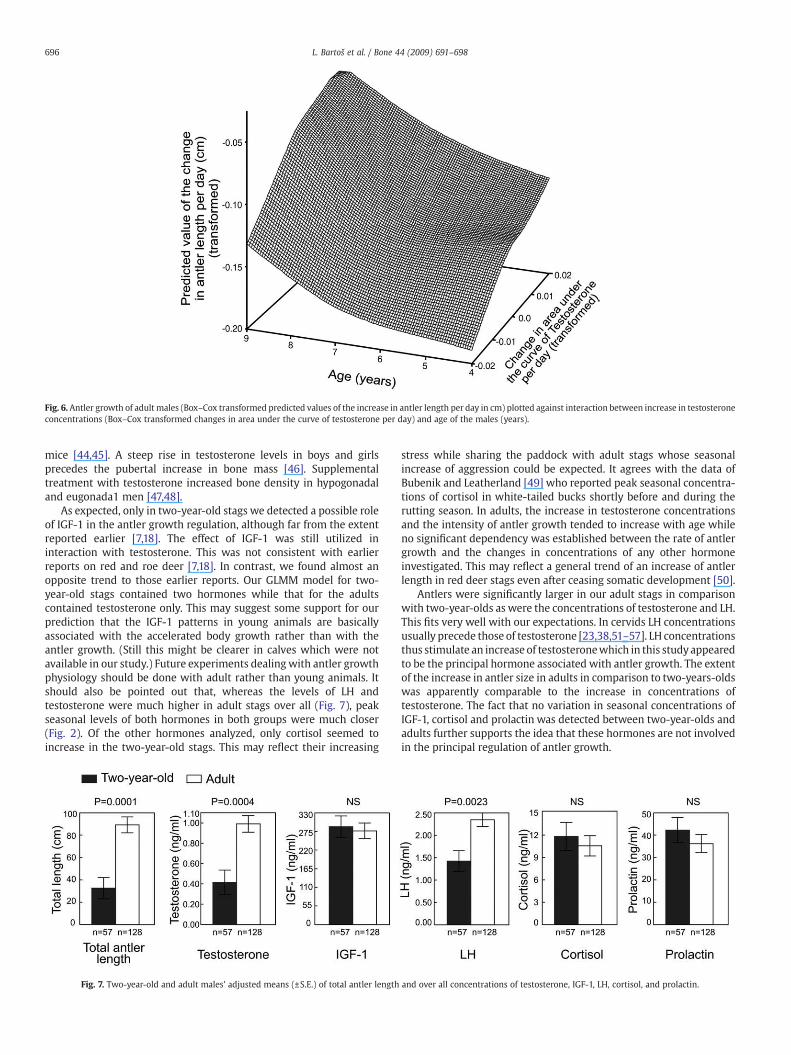
Fig. 6. Antler growth of adult males (Box–Cox transformed predicted values of the increase in antler length per day in cm) plotted against interaction between increase in testosteroneconcentrations (Box–Cox transformed changes in area under the curve of testosterone per day) and age of the males (years).
696 L. Bartoš et al. / Bone 44 (2009) 691–698
mice [44,45]. A steep rise in testosterone levels in boys and girlsprecedes the pubertal increase in bone mass [46]. Supplementaltreatment with testosterone increased bone density in hypogonadaland eugonada1 men [47,48].
As expected, only in two-year-old stags we detected a possible roleof IGF-1 in the antler growth regulation, although far from the extentreported earlier [7,18]. The effect of IGF-1 was still utilized ininteraction with testosterone. This was not consistent with earlierreports on red and roe deer [7,18]. In contrast, we found almost anopposite trend to those earlier reports. Our GLMM model for two-year-old stags contained two hormones while that for the adultscontained testosterone only. This may suggest some support for ourprediction that the IGF-1 patterns in young animals are basicallyassociated with the accelerated body growth rather than with theantler growth. (Still this might be clearer in calves which were notavailable in our study.) Future experiments dealing with antler growthphysiology should be done with adult rather than young animals. Itshould also be pointed out that, whereas the levels of LH andtestosterone were much higher in adult stags over all (Fig. 7), peakseasonal levels of both hormones in both groups were much closer(Fig. 2). Of the other hormones analyzed, only cortisol seemed toincrease in the two-year-old stags. This may reflect their increasing
Fig. 7. Two-year-old and adult males' adjusted means (±S.E.) of total antler length
stress while sharing the paddock with adult stags whose seasonalincrease of aggression could be expected. It agrees with the data ofBubenik and Leatherland [49] who reported peak seasonal concentra-tions of cortisol in white-tailed bucks shortly before and during therutting season. In adults, the increase in testosterone concentrationsand the intensity of antler growth tended to increase with age whileno significant dependency was established between the rate of antlergrowth and the changes in concentrations of any other hormoneinvestigated. This may reflect a general trend of an increase of antlerlength in red deer stags even after ceasing somatic development [50].
Antlers were significantly larger in our adult stags in comparisonwith two-year-olds as were the concentrations of testosterone and LH.This fits very well with our expectations. In cervids LH concentrationsusually precede those of testosterone [23,38,51–57]. LH concentrationsthus stimulate an increase of testosteronewhich in this studyappearedto be the principal hormone associated with antler growth. The extentof the increase in antler size in adults in comparison to two-years-oldswas apparently comparable to the increase in concentrations oftestosterone. The fact that no variation in seasonal concentrations ofIGF-1, cortisol and prolactin was detected between two-year-olds andadults further supports the idea that these hormones are not involvedin the principal regulation of antler growth.
and over all concentrations of testosterone, IGF-1, LH, cortisol, and prolactin.

697L. Bartoš et al. / Bone 44 (2009) 691–698
As mentioned earlier, IGF-1 was associated with antler growth insome studies [7,18,19] but not in others including the present results[20,22,23]. In contrary to these data, antler growth was invariablyassociated with an increase of testosterone concentrations in allstudies across many cervid species, such as red deer [7], roe deer[58–60], fallow deer [23,39,61,62], white-tailed deer, Odocoileusvirginianus [63–66], Columbian black-tailed deer, Odocoileus hemi-onus columbianus [67], axis deer, Axis axis [55,68], pudu [19], Eld'sdeer, Cervus eldi thamin [69], reindeer [21], and rusa deer, Cervus(Rusa) timorensis [70]. It is obvious that the sharp increase intestosterone concentration which causes the mineralization ofantlers and the shedding of velvet occurs only after the cessationof antler growth and the completion of antler bone development. Theconclusion that a low concentration of testosterone and not IGF-1serves as a permissive antler-stimulating hormone however does notchallenge the fact that higher levels of testosterone stop all bonegrowth and induce antler bone mineralization, as observed acrossvarious deer taxa [12]. This fits very well with the earlier study ofBrown et al. [10] who found positive correlations between increasingserum androgen concentrations and increasing antler mass in white-tailed bucks. They concluded that low concentrations of testosteronecan stimulate bone growth, while a higher level will cause inhibition[10]. Recent in vitro studies support the notion that androgens (suchas testosterone, DHT or DHEA) excert dose-dependent mitogeniceffect on antler cells derived from the proliferation zone of the antlertip. However, the androgen effect may also depend on other serumfactors such as anabolic hormones and serum-containing culturemedium [71]. Testosterone has also a direct stimulatory effect on thegrowth of the mouse male-derived mandibular condyle. Overtreat-ment with testosterone (regarding both dosage and duration) causesenhanced calcification at the expense of chondrogenic activity,resulting in reduced overall condylar growth [72].
Recently, low but significant levels of testosterone and estradiolwere detected in growing antler bone of white-tailed deer [73] thusindicating a role of sexual steroids in antler development. A growthpromoting effect of steroids can be brought about by increasing theproduction of IGF-1, which then acts as an autocrine, paracrine orendocrine growth factor [23,74]. In a number of studies and varioustaxa it has been reported, that testosterone stimulates growth andlocal production of IGF-I [72,73,75,76].
A lack of statistical difference between over all levels of IGF-1 insubadult and adult stags in the present study contrasts sharply withthe significant differences found for LH, testosterone and the rate ofantler elongation (Fig. 7). Furthermore the time course of IGF-1 in bothgroups is rather flat, unlike a rising trend of testosterone concentra-tions. Our present results thus lead us to conclude that it istestosterone (possibly in an interaction with other steroids) and notIGF-1 which is responsible primarily for the intensity of antler growthin subadult and adult red deer stags.
Acknowledgments
The authors gratefully acknowledge the assistance of Jiří Šiler,Soběslav Losos and Petr Janovský in managing the deer as well as theconsultations on statistic with Petr Šimeček. We wish to thank twoanonymous referees for their helpful comments. This study wassupported by the Czech Science Foundation (grant number 523/08/0808) and Ministry of Agriculture of the Czech Republic (grantnumbers MZe 0002701402 and MZe 0002701404).
References
[1] Bubenik GA. The antlers as a model in biochemical research. In: Bubenik GA,Bubenik AB, editors. Horns, pronghorns, and antlers: evolution, morphology,physiology, and social significance. New York, Berlin, Heidelberg, London, Paris,Tokyo, Hong Kong: Springer-Verlag; 1990. p. 474–87.
[2] Miller SC, Bowman BM, Jee WSS. Available animal-models of osteopenia— smalland large. Bone 1995;17:S117–23.
[3] Landete-Castillejos T, Currey JD, Estevez JA, Gaspar-Lopez E, Garcia A, Gallego L.Influence of physiological effort of growth and chemical composition on antlerbone mechanical properties. Bone 2007;41:794–803.
[4] Landete-Castillejos T, Estevez JA, Martinez A, Ceacero F, Garcia A, Gallego L. Doeschemical composition of antler bone reflect the physiological effort made to growit? Bone 2007;40:1095–102.
[5] Hau M. Regulation of male traits by testosterone: implications for the evolution ofvertebrate life histories. BioEssays 2007;29:133–44.
[6] Ketterson ED, Nolan V. Adaptation, exaptation, and constraint: a hormonalperspective. Am Natur 1999;154:S4–S25.
[7] Suttie JM, Gluckman PD, Butler JH, Fennessy PF, Corson ID, Laas FJ. Insulin-likegrowth factor1 (IGF-1) antler-stimulatinghormone?Endocrinology1985;116:846–8.
[8] Suttie JM, Fennessy PF, Gluckman PD, Corson ID. Elevated plasma IGF-1 levels stagsprevented from growing antlers. Endocrinology 1988;122:3005–7.
[9] Bubenik GA, Brown GM, Bubenik AB, Grota LJ. Immunohistological localization oftestosterone in the growing antler of the white-tailed deer (Odocolieusvirginianus). Calcif Tissue Res 1974;14:121–30.
[10] Brown RD, Cowan RL, Griel LC. Correlation between antler and long bone relativebone mass and circulating androgens inwhite-tailed deer (Odocolieus virginianus).Am J Vet Res 1978;39:1053–6.
[11] Bartoš L. The date of antler casting, age and social hierarchy relationships in the reddeer stag. Behav Process 1980;5:293–301.
[12] Bubenik GA. Endocrine regulation of the antler cycle. In: Brown RD, editor. Antlerdevelopment in Cervidae. Kingsville, Texas: Caesar Kleberg Wildl. Res. Inst.; 1982.p. 73–107.
[13] Elliott JL, Oldham JM, Ambler GR, Bass JJ, Spencer GSG, Hodkinson SC, et al.Presence of insulin-like growth factor-I receptors and absence of growth hormonereceptors in the antler tip. Endocrinology 1992;130:2513–20.
[14] Elliott JL, Oldham JM, Ambler GR, Molan PC, Spencer GSG, Hodkinson SC, et al.Receptors for insulin-like growth factor-II in the growing tip of the deer antler.J Endocrinol 1993;138:233–41.
[15] Suttie JM, Li CY, Sadighi M, Gunn J, Fleming JS. Physiological control of antlergrowth. In: Milne JA, editor. Recent developments in deer biology. Macaulay LandUse Research Institute, Craigiebuckler, Aberdeen, UK. Edinburgh, UK: MoredunResearch Institute; 1994. p. 189–96.
[16] Li CY, Littlejohn RP, Suttie JM. Effects of insulin-like growth factor 1 and testos-terone on the proliferation of antlerogenic cells in vitro. J Exp Zool 1999;284:82–90.
[17] Sadighi M, Li C, Littlejohn RP, Suttie JM. Effects of testosterone either alone or withIGF-I on growth of cells derived from the proliferation zone of regenerating antlersin vitro. Growth Horm IGF Res 2001;11:240–6.
[18] Schams D, Barth D, Heinze-Mutz EM, Pflaum G, Karg H. Antler growth in roe deer:effects of casting and correlations with growth hormone and insulin like growthfactor I. In: Brown RD, editor. The biology of deer. New York, Berlin, Heidelberg,London, Paris, Tokyo, Hong Kong, Barcelona, Budapest: Springer-Verlag; 1992.p. 505–10.
[19] Reyes E, Bubenik GA, Lobos A, Schams D, Bartoš L. Seasonal levels of cortisol, IGF-1and triiodothyronine in adult male pudu (Pudu puda). Folia Zool 1997;46:109–16.
[20] Bartoš L, Reyes E, Schams D, Bubenik GA, Lobos A. Rank dependent seasonal levelsof IGF-1, cortisol and reproductive hormones in male pudu (Pudu puda). CompBiochem Physiol, A 1998;120:373–8.
[21] Blake JE, Rowell JE, Suttie JM. Characteristics of first-antler growth in reindeer andtheir association with seasonal fluctuations in steroid and insulin-like growthfactor 1 levels. Can J Zool 1998;76:2096–102.
[22] Bubenik GA, Schams D, White RG, Rowell J, Blake J, Bartoš L. Seasonal levels ofmetabolic hormones and substrates in male and female reindeer (Rangifertarandus). Comp Biochem Physiol, C 1998;120:307–15.
[23] Bartoš L, Schams D, Kierdorf U, Fischer K, Bubenik GA, Šiler J, et al. Cyproteroneacetate reduced antler growth in surgically castrated fallow deer. J Endocrinol2000;164:87–95.
[24] Suttie JM, Corson ID, Gluckman PD, Fennessy PF. Insulin-like growth factor-1,growth and body composition in red deer stags. Anim Prod 1991;53:237–42.
[25] Ditchkoff SS, Spicer LJ, Masters RE, Lochmiller RL. Concentrations of insulin-likegrowth factor-I in adult male white-tailed deer (Odocoileus virginianus): associa-tions with serum testosterone, morphometrics and age during and after thebreeding season. Comp Biochem Physiol, A 2001;129:887–95.
[26] Adam CL, Kyle CE, Young P, Atkinson T. Effect of nutritional growth restriction ontiming of reproductive development and plasma concentrations of insulin-likegrowth factor-1 and growth hormone in male red deer (Cervus elaphus) reared inconstant photoperiod. Anim Sci 1995;61:155–60.
[27] Adam CL, Kyle CE, Young P. Seasonal patterns of growth, voluntary food intake andplasma concentrations of prolactin, insulin like growth factor 1, LH and gonadalsteroids in male and female prepubertal red deer (Cervus elaphus) reared in eithernatural or constant daylength. Anim Sci 1996;62:605–13.
[28] Webster JR, Corson ID, Littlejohn RP, Stuart SK, Suttie JM. Effects of season andnutrition on growth hormone and insulin-like growth factor-I in male red deer.Endocrinology 1996;137:698–704.
[29] Webster JR, Corson ID, Littlejohn RP, Stuart SK, Suttie JM. Photoperiodicrequirements for rapid growth in young male red deer. Anim Sci 1998;67:363–70.
[30] Webster JR, Corson ID, Littlejohn RP, Martin SK, Suttie JM. The roles of photoperiodand nutrition in the seasonal increases in growth and insulin-like growth factor-1secretion in male red deer. Anim Sci 2001;73:305–11.
[31] HamiltonWJ, Blaxter KL. Growth of red deer stags under farm conditions. J Agr Sci1981;97:329–34.

698 L. Bartoš et al. / Bone 44 (2009) 691–698
[32] Gaspar-López E, García AJ, Landete-Castillejos T, Carrión D, Estevez JA, Gallego L.Growth of the first antler in Iberian red deer (Cervus elaphus hispanicus). Eur JWildl Res 2008;54:1–5.
[33] Suttie JM, Fennessy PF, Corson ID, Laas FJ, Crosbie SF, Butler JH, et al. Pulsatilegrowth hormone, insulin-like growth factors and antler development in red deer(Cervus elaphus scoticus) stags. J Endocrinol 1989;121:351–60.
[34] Huxley JS. The annual increment of the antlers of the red deer (Cervus elaphus).Proc R Soc Lond 1926;67:1021–36.
[35] Blottner S, Hingst O, Meyer HHD. Seasonal spermatogenesis and testosteroneproduction in roe deer (Capreolus capreolus). J Reprod Fertil 1996;108:299–305.
[36] Sauerwein H, Dürsch I, Meyer HHD. Quantitation of glucocorticoid receptors inbovine skeletal muscle: topographical distribution, sex effect and breedcomparisons. J Steroid Biochem Mol Biol 1991;39:941–5.
[37] Schams D, Karg H. Radioimmunologische Bestimmung von Prolactin in Blutserumvom Rind. Milchwissenschaft 1969;24:262–5.
[38] Schams D, Barth D, Karg H. LH, FSH and progesterone concentrations in peripheralplasma of the female roe deer (Capreolus capreolus) during the rutting season.J Reprod Fertil 1980;60:109–14.
[39] Bartoš L, Bubenik GA, Tománek M. The role of adrenal androgens in antler growthof castrated fallow deer (Dama dama): possible stimulation of adrenal secretion oftestosterone by stress. In: Suttie JM, Haines SR, Li C, editors. Antler science andproduct technology. Mosgiel, New Zealand: AgResearch, Invermay AgriculturalCentre; 2004. p. 45–53.
[40] Timm NH, Mieczkowski TA. Univariate and multivariate general linear models:theory and application using SAS software. Cary, NC, USA: SAS Institute Inc.; 1997.
[41] Schwarz G. Estimating the dimension of a model. Ann Stat 1978;6:461–4.[42] Tao J, Littel R, Patetta M, Truxillo C, Wolfinger R. Mixed model analyses using the
SAS system course notes. Carry, NC, USA: SAS Institute Inc.; 2002.[43] Goméz JA, García AJ, Landete-Castillejos T, Gallego L. Effect of advancing births on
testosterone until 2.5 years of age and puberty in Iberian red deer (Cervus elaphushispanicus). Anim Reprod Sci 2006;96:79–88.
[44] Venken K, De Gendt K, Boonen S, Ophoff J, Bouillon R, Swinnen JV, et al. Relativeimpact of androgen and estrogen receptor activation in the effects of androgens ontrabecular and cortical bone in growing male mice: a study in the androgenreceptor knockout mouse model. J Bone Miner Res 2006;21:576–85.
[45] Venken K,Movérare-Skrtic S, Kopchick JJ, Coschigano KT, Ohlsson C, Boonen S, et al.Impact of androgens, growth hormone, and IGF-I on bone andmuscle inmalemiceduring puberty. J Bone Miner Res 2007;22:72–82.
[46] Yilmaz D, Ersoy B, Bilgin E, Gumuser G, Onur E, Pinar ED. Bone mineral density ingirls and boys at different pubertal stages: relation with gonadal steroids, boneformation markers, and growth parameters. J Bone Miner Metab 2005;23:476–82.
[47] Amory JK, Watts NB, Easley KA, Sutton PR, Anawalt BD, Matsumoto AM, et al.Exogenous testosterone or testosterone with finasteride increases bone mineraldensity in older men with low serum testosterone. J Clin Endocrinol Metab 2004;89:503–10.
[48] Anderson FH, Francis RM, Faulkner K. Androgen supplementation in eugonadalmenwith osteoporosis— effects of 6months of treatment on bonemineral densityand cardiovascular risk factors. Bone 1996;18:171–7.
[49] Bubenik GA, Leatherland JF. Seasonal levels of cortison and thyroid hormones inintact and castrated mature male white-tailed deer. Can J Zool 1984;62:783–7.
[50] Kruuk LEB, Slate J, Pemberton JM, Brotherstone S, Guinness F, Clutton-Brock T.Antler size in red deer: heritability and selection but no evolution. Evolution2002;56:1683–95.
[51] Lincoln GA, Kay RNB. Effects of season on the secretion of LH and testosterone inintact and castrated red deer stags (Cervus elaphus). J Reprod Fert 1979;55:75–80.
[52] Mirarchi RE, Howland RE, Scanlon PF, Kirkpatrick RL, Sanford LM. Seasonalvariation in plasma LH, FSH, prolactin, and testosterone concentrattions in adultwhite-tailed deer. Can J Zool 1978;56:121–72.
[53] Bubenik GA, Morris JM, Schams D, Claus A. Photoperiodicity and circannual levelsof LH, FSH, and testosterone in normal and castrated male, white-tailed deer. Can JPhysiol Pharmacol 1982;60:788–93.
[54] Bubenik GA, Bubenik AB, Schams D, Leatherland JF. Circadian and circannualrhythms of LH, FSH, testosterone (T), prolactin, cortisol, T3 and T4 in plasma ofmature, male white-tailed deer. Comp Biochem Physiol 1983;76A:37–45.
[55] Bubenik GA, Brown RD, Schams D. Antler cycle and endocrine parameters inmale Axis deer (Axis-axis)— seasonal levels of LH, FSH, testosterone, and
prolactin and results of GnRH and ACTH challenge tests. Comp Biochem Physiol1991;99:645–50.
[56] Bubenik GA, Reyes E, Schams D, Lobos A, Bartoš L. Seasonal levels of LH, FSH,testosterone and prolactin in adult male pudu (Pudu puda). Comp Biochem Physiol1996;115B:417–20.
[57] Kameyama Y, Miyamoto A, Kobayashi S, Kuwayama T, Ishijima Y. Annual changesin serum LH and testosterone concentrations in male sika deer (Cervus nippon).J Reprod Dev 2002;48:613–7.
[58] Sempéré AJ, Boissin J. Relationship between antler development and plasmaandrogen concentrations in adult roe deer (Capreolus capreolus). J Reprod Fertil1981;62:49–53.
[59] Sempéré AJ, Lacroix A. Temporal and seasonal relationships between LH,testosterone and antlers in fawn and adult male roe deer (Capreolus capreolus L.)—a longitudinal study from birth to 4 years age. Acta Endocrinol 1982;99:295–301.
[60] Sempéré AJ, Mauget R, Bubenik GA. Influence of photoperiod on the seasonalpattern of secretion of luteinizing hormone and testosterone and on the antlercycle in roe deer (Capreolus-capreolus). J Reprod Fertil 1992;95:693–700.
[61] Rolf HJ, Fischer K. Serum testosterone (T) and 5-alpha-dihydrotestosterone (DHT)in male fallow deer (Dama-dama L)— seasonality and age dependence. CompBiochem Physiol 1990;95A:445–52.
[62] Rolf HJ, Fischer K. Serum testosterone, 5-alpha-dihydrotestosterone and differentsex characteristics in male fallow deer (Cervus dama): a long-term experimentwith accelerated photoperiods. Comp Biochem Physiol 1996;115:207–21.
[63] Mirarchi RE, Scanlon PF, Kirkpatrick RL, Schreck CB. Androgen levels and antlerdevelopment in captive and wild white-tailed deer. J Wildl Manage 1977;41:178–83.
[64] Brown RD, Chao CC, Faulkner LW. The endocrine control of the initiation andgrowth of antlers in white-tailed deer. Acta Endocrinol 1983;103:138–44.
[65] Bubenik GA, Pomerantz DK, Schams D, Smith PS. The role of androstenedione andtestosterone in the reproduction and antler growth of a male white-tailed deer.Acta Endocrinol 1987;114:147.
[66] Bubenik GA, Schams D, Coenen G. The effect of artificial photoperiodicity andantiandrogen treatment on the antler growth and plasma levels of LH, FSH,testosterone, prolactin and alcaline phosphatase in the male white-tailed deer.Comp Biochem Physiol 1987;87A:551–9.
[67] West NO, Nordan HC. Hormonal regulation of reproduction and the antler cycle inthe male Columbian black-tailed deer (Odocoileus hemionus columbianus). Part I.Seasonal changes in the histology of the reproductive organs, serum testosterone,sperm production, and the antler cycle. Can J Zool 1976;54:1617–36.
[68] Loudon ASI, Curlewis JD. Cycles of antler and testicular growth in an aseasonaltropical deer (Axis axis). J Reprod Fertil 1988;83:729–38.
[69] Monfort SL, Brown JL, Bush M, Wood TC, Wemmer C, Varga A, et al. Circannualinter-relationship among reproductive hormones, gross morphometry, behavior,ejaculate characteristics and testicular histology in Eld's deer (Cervus eldi thamin).J Reprod Fertil 1993;98:471–80.
[70] van Mourik S, Stelmasiak T. Endocrine mechanisms and antler cycles in rusa deerCervus (Rusa) timorensis. In: Bubenik GA, Bubenik AB, editors. Horns, pronghorns,and antlers: evolution, morphology, physiology, and social significance. New York,Berlin, Heidelberg, London, Paris, Tokyo, Hong Kong: Springer-Verlag; 1990.p. 416–25.
[71] Rolf HJ, Wiese KG, Siggelkow H, Schliephake H, Bubenik GA. In vitro-studies withantler bone cells: structure forming capacity, osteocalcin production and influenceof sex steroids. Osteology 2006;15:245–57.
[72] Maor G, Segev Y, Phillip M. Testosterone stimulates insulin-like growth factor-I andinsulin-like growth factor-I-receptor gene expression in the mandibular condyle—a model of endochondral ossification. Endocrinology 1999;140:1901–10.
[73] Bubenik GA, Miller KV, Lister AL, Osborn DA, Bartoš L, Van Der Kraak GJ.Testosterone and estradiol concentrations in serum, velvet skin and growingantler bone of male white-tailed deer. J Exp Zool 2005;303A:186–92.
[74] Westley BR, May FEB. Role of insulin-like growth factors in steroid modulatedproliferation. J Steroid Biochem Mol Biol 1994;51:1–9.
[75] Craft WH, Underwood LE. Effects of androgens on plasma somatomedin-C insulin-like growth factor 1 responses to growth hormone. Clin Endocrinol 1984;20:549–703.
[76] Žofková I. Hormonal aspects of the muscle-bone unit. Physiol Res 2008;57:S159–69.
Copyright © 2022 FDOKUMEN




















