
Late-glacial recolonization and phylogeography of European red deer ( Cervus elaphus L.)
10
See discussions, stats, and author profiles for this publication at: http://www.researchgate.net/publication/235668686 Rabbits and hominin survival in Iberia ARTICLE in JOURNAL OF HUMAN EVOLUTION · APRIL 2013 Impact Factor: 3.87 · DOI: 10.1016/j.jhevol.2013.01.002 · Source: PubMed CITATIONS 16 DOWNLOADS 396 VIEWS 349 4 AUTHORS: John Fa Manchester Metropolitan University 152 PUBLICATIONS 2,373 CITATIONS SEE PROFILE John R Stewart Bournemouth University 71 PUBLICATIONS 1,318 CITATIONS SEE PROFILE Lluís Lloveras University of Barcelona 24 PUBLICATIONS 219 CITATIONS SEE PROFILE J. Mario Vargas University of Malaga 129 PUBLICATIONS 1,295 CITATIONS SEE PROFILE Available from: John R Stewart Retrieved on: 19 September 2015
-
Upload
bournemouth -
Category
Documents
-
view
0 -
download
0
Transcript of Late-glacial recolonization and phylogeography of European red deer ( Cervus elaphus L.)

Seediscussions,stats,andauthorprofilesforthispublicationat:http://www.researchgate.net/publication/235668686
RabbitsandhomininsurvivalinIberia
ARTICLEinJOURNALOFHUMANEVOLUTION·APRIL2013
ImpactFactor:3.87·DOI:10.1016/j.jhevol.2013.01.002·Source:PubMed
CITATIONS
16
DOWNLOADS
396
VIEWS
349
4AUTHORS:
JohnFa
ManchesterMetropolitanUniversity
152PUBLICATIONS2,373CITATIONS
SEEPROFILE
JohnRStewart
BournemouthUniversity
71PUBLICATIONS1,318CITATIONS
SEEPROFILE
LluísLloveras
UniversityofBarcelona
24PUBLICATIONS219CITATIONS
SEEPROFILE
J.MarioVargas
UniversityofMalaga
129PUBLICATIONS1,295CITATIONS
SEEPROFILE
Availablefrom:JohnRStewart
Retrievedon:19September2015

at SciVerse ScienceDirect
Journal of Human Evolution xxx (2013) 1e9
Contents lists available
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
Rabbits and hominin survival in Iberia
John E. Fa a,b,*, John R. Stewart c, Lluís Lloveras d, J. Mario Vargas a,e
aDurrell Wildlife Conservation Trust, Les Augrès Manor, Trinity, Jersey JE3 5BP, UKb ICCS, Division of Biology, Imperial College London, Silwood Park Campus, Buckhurst Road, Ascot SL5 7PY, UKc School of Applied Sciences, Bournemouth University, Talbot Campus, Fern Barrow Poole, Dorset BH12 5BB, UKd SERP, Department de Prehistòria, Història Antiga i Arqueología, Universitat de Barcelona, Montalegre 6-8, 08001 Barcelona, SpaineDepartamento de Biología Animal, Facultad de Ciencias, Universidad de Málaga, Campus de Teatinos s/n, Málaga 29071, Spain
a r t i c l e i n f o
Article history:Received 4 April 2012Accepted 7 January 2013Available online xxx
Keywords:Oryctolagus cuniculusAnatomically modern humansNeanderthalsSpecies assemblagesMammalian standing biomassMass hunting
* Corresponding author.E-mail addresses: [email protected] (J.E. Fa),
(J.R. Stewart), [email protected] (L. Lloveras), jmvy
0047-2484/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.jhevol.2013.01.002
Please cite this article in press as: Fa, J.E., et10.1016/j.jhevol.2013.01.002
a b s t r a c t
High dependence on the hunting and consumption of large mammals by some hominins may havelimited their survival once their preferred quarry became scarce or disappeared. Adaptation to smallerresidual prey would have been essential after the many large-bodied species decreased in numbers. Wefocus on the use of a superabundant species, the rabbit, to demonstrate the importance of this taxon inIberia as fundamental to predators. We show that the use of the rabbit over time has increased, and thatthere could have been differential consumption by Neanderthals and Anatomically Modern Humans(AMH). Analysis of bone remains from excavations throughout Iberia show that this lagomorph wasa crucial part of the diet of AMH but was relatively unutilised during the Mousterian, when Neanderthalswere present. We first present changes in mammalian biomass and mean body mass of mammals over50,000 years, to illustrate the dramatic loss of large mammalian fauna and to show how the rabbit mayhave contributed a consistently high proportion of the available game biomass throughout that period.Unlike the Italian Peninsula and other parts of Europe, in Iberia the rabbit has provided a food resource ofgreat importance for predators including hominins. We suggest that hunters that could shift focus torabbits and other smaller residual fauna, once larger-bodied species decreased in numbers, would havebeen able to persist. From the evidence presented here, we postulate that Neanderthals may have beenless capable of prey-shifting and hence use the high-biomass prey resource provided by the rabbit, to theextent AMH did.
� 2013 Elsevier Ltd. All rights reserved.
Introduction
There is considerable evidence from present-day studies ofhuman predation that hunters will shift to smaller-bodied (moresustainable) prey species once the larger animals have decreased innumbers (e.g., Cannon, 2000; Jerozolimski and Peres, 2003). Such‘extinction filters’ (Balmford, 1996) suggest that faunas can beimpoverished as a result of overexploitation to include only thosespecies able to survive anthropogenic impacts. Nonetheless, thereare other circumstances where hunters may focus on the smaller,abundant prey as the mainstay of their diets, though they mayoccasionally hunt larger animals. How past human hunter/gathererpopulations responded to changes in availability of prey size classesis still largely unexplored.
[email protected]@uma.es (J.M. Vargas).
All rights reserved.
al., Rabbits and hominin sur
Archaeological analysis of faunal remains and of lithic tools hassuggested differences in the prey pursued by Neanderthals incontrast to Anatomically Modern Humans (AMH). Hunting of me-dium to large mammals may have been a major element in Nean-derthal subsistence, with smaller prey being of minor importance.Neanderthals may have behaved as large game specialists (Stewart,2004), obtaining almost all of their dietary protein from these an-imal sources (Richards et al., 2000). Recent studies indicate thatNeanderthals also consumed small game such as birds, fish, mol-luscs and some plants (Blasco and Fernández Peris, 2009; Brownet al., 2011; Hardy and Moncel, 2011; Henry et al., 2011). In spiteof this, archaeological evidence shows that hunting of smaller, fastmoving prey species occured more frequently in the Upper Palae-olithic (Villaverde et al., 1996; Martínez Valle, 2001; Pérez Ripolland Martínez Valle, 2001; Hockett and Haws, 2002; Cochard andBrugal, 2004; Stiner et al., 2008). This period, and certainly theAurignacian, Gravettian and later technocomplexes, is generallybelieved to be the product of AMH since Neanderthals were notpresent for much of the time involved. Thus, Neanderthals may
vival in Iberia, Journal of Human Evolution (2013), http://dx.doi.org/

J.E. Fa et al. / Journal of Human Evolution xxx (2013) 1e92
have disappeared as the large prey items they may have specialisedupon were severely depleted.
Prey body size has been used as a proxy for understanding preyrank and return rate relationships in zooarchaeology (Broughtonet al., 2011). Body size is one of the most fundamental propertiesof any organism. The study of correlates and consequences of bodysize offer a valuable perspective on how scaling of structure andfunction affects processes at the individual as well as the biologicalcommunity level (Brown, 1995). Because size is associated withlifespan, home range size and other aspects of life history andecology, the relationship between body size and abundance is anessential link between the individual- and population-level traits ofspecies and the structure and dynamics of ecological communities(Peters, 1983). Issues related to prehistoric foraging efficiency anddiet breadth have been enabled through proxies such as body size ofhunted species to estimate prey encounter rates (Waguespack andSurovell, 2003). The body size proxy, supported by available eth-nographic and sometimes experimentally derived return-rate data,can further inform past hunting patterns (Broughton et al., 2011).
In this paper, we summarise data on prey size and abundance ofmammals to determine probable encounter rates through time. Wefocus on mammalian assemblages in Iberia and southern Francewithin the prehistoric distribution of the European rabbit (Orycto-lagus cuniculus). The rabbit originated in the Iberian Peninsula(Branco et al., 2000) and remained restricted to the Iberian Pen-insula and southern France until the Middle Ages, when throughhuman mediated colonisation it was spread across central andnorthern Europe and the British Isles (Corbet, 1994; Callou, 1995).Today, rabbits can be found throughout the world (Flux, 1994). Theimportance of this lagomorph as a keystone resource for manypredators is well documented (Delibes-Mateos et al., 2008).Moreover, rabbits are a ubiquitous feature of faunal assemblagesfrom almost every Spanish and Portuguese archaeological site andare therefore accepted as a fundamental resource for hominins(Aura et al., 2002; Sanchís Serra and Fernández Peris, 2008).However, the importance of the rabbit to Neanderthal versus AMHhas not been documented.
We analyse published data on mammal species and volume oftheir remains excavated from caves and rock shelters in Iberia andsouthern France across 50 kyr BP (thousands of years before pre-sent). We use prey size relationships to estimate faunal transitionsand mammalian community biomass changes through time. Wethen focus on the contribution made to the community biomass bythe rabbit relative to other mammals, as well as the potentialconsumption of this prey item by Neanderthals and modernhumans across the same time period. We speculate on the impor-tance of such smaller prey items on the survival of hominins inIberia, and discuss the evidence on probable use of this species byNeanderthals and AMH.
Materials and methods
Study species and time periods
We concentrated on macromammals (defined as species withaverage adult body mass equal to or greater than 1 kg). We con-sidered only macromammals because: 1) these species have beenactively recorded by archaeologists; micromammals (<1 kg) haveoften been ignored or not systematically recovered or recorded, 2)the taxa are less prone to serious bias in the representation ofspecies and their relative abundance (Van Valen, 1964); and 3)there is evidence that all of these species were hunted either inprehistoric or present times.
Because carbon dating for many sites were not available, weused seven cultural time periods divisible into three main
Please cite this article in press as: Fa, J.E., et al., Rabbits and hominin sur10.1016/j.jhevol.2013.01.002
chronological phases to determine temporal changes in faunas: 1)Middle Palaeolithic e Mousterian (c. 300e30 kyr BP), warm andcold phases; 2) Upper Palaeolithic (c. 45e10 kyr BP) e Aurignacian(c. 32e26 kyr BP), Gravettian (c. 28e22 kyr BP), Solutrean (c. 21e17 kyr BP), Magdalenian (c. 18e10 kyr BP) and 3) Mesolithic(<c. 9 kyr BP). These time periods are progressively youngeralthough some overlap in age may exist. However, the age overlapswill only cause observed changes in faunas to bemore conservative.We use the term ‘Mesolithic’ here to include also the ‘Epi-palaeolithic’. This period refers to the final Upper Palaeolithic in-dustries occurring at the end of the terminal Pleistocene, whichappear tomerge technologically into theMesolithic. We consideredthose deposits dated later than the Greenland Interstadial 1 (GI-1)as Epipalaeolithic or Mesolithic, according to the nomenclatureestablished by García-Argüelles et al. (2009).
Inferring species encounter frequencies
The strong global empirical relationships between body size andpopulation density of all forms of life (Peters, 1983) can be used toestimate relative encounter frequencies for prey species in anyecosystem, present or past (Waguespack and Surovell, 2003).Relating prey selectivity to the natural abundance of available preyspecies creates a framework for examining the types of prey uti-lised by hunteregatherers (Gamble et al., 2004). Because encounterrate is primarily a function of prey population density, estimatedpopulation densities based on body size provide a reasonableapproximation of prey encounter rates for specific environments(Surovell and Waguespack, 2009). Generally speaking, there isa negative power relationship between body size and populationdensity, meaning that, all things being equal, small bodied animalsare more common than large ones.
We obtained the average body mass (in kg) of extant (Joneset al., 2009) and extinct species (Smith et al., 2003) detected dur-ing each time period. Crude population densities of extant taxawere available from Jones et al. (2009). However, densities forextinct mammals were predicted from the inverse relationshipbetween body size and abundance using the data in Jones et al.(2009) for all extant mammals.
Faunal transitions
We compiled macromammal occurrence in each time periodfrom published faunal lists from 104 excavation units in 30 sites inSpain, Portugal and southern France (Table S1 and S2).We also usedinformation contained in the Stage 3 Project Faunal Database for254 Mousterian and Aurignacian units (Stewart et al., 2003). Weselected only those sites where the rabbit was present throughoutthe seven time periods used in this study. We excluded thoseprehistoric sites in northern Spain (e.g., in the Cantabrian Moun-tains, see Álvarez-Lao and García, 2011) where the rabbit does notoccur naturally (Villafuerte, 2007) and did not appear in excava-tions. The reason for this choice was our interest in investigatingthe importance of the species relative to others.
Standing biomass changes
To evaluate the ‘supply’ of mammalian prey across time, wecalculated crude standing biomass (kg/km�2) and mean body massper assemblage in each time period. Community biomass (orstanding crop), which represents the sum of the live weights ofindividuals occupying a given area was computed by multiplyingthe known or predicted density of each macromammal present, byits average live weight and then summing their biomasses(Robinson and Bennett, 2004). We also calculated the percentage
vival in Iberia, Journal of Human Evolution (2013), http://dx.doi.org/
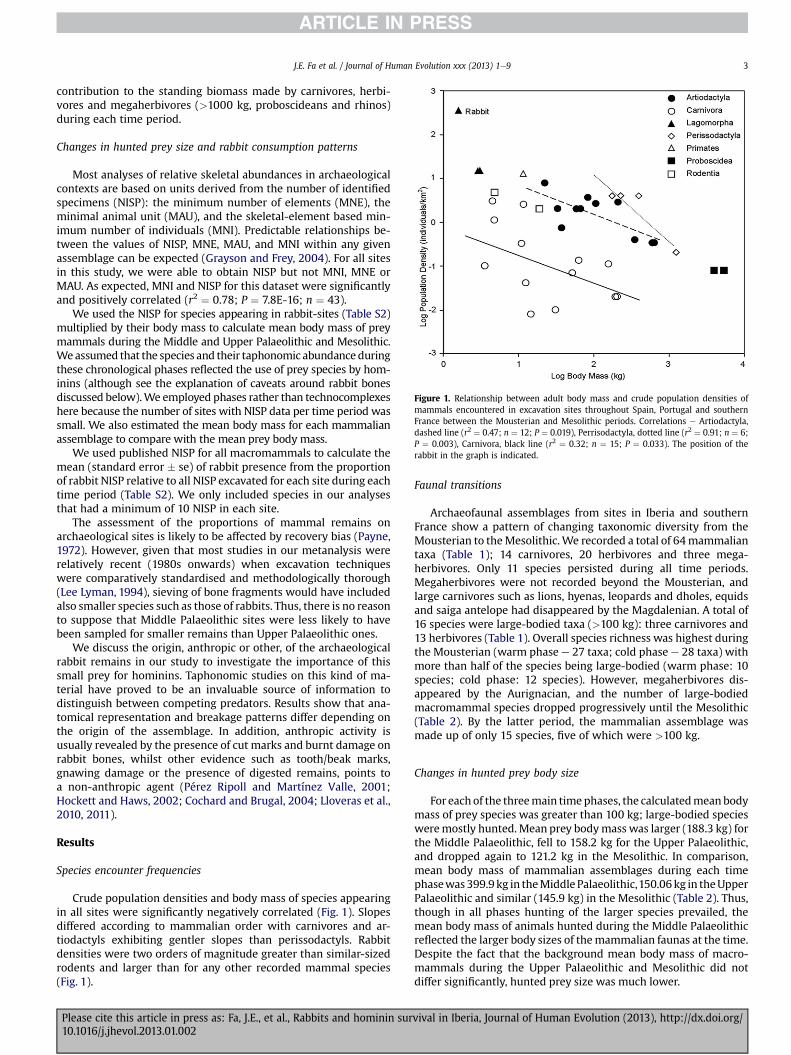
Figure 1. Relationship between adult body mass and crude population densities ofmammals encountered in excavation sites throughout Spain, Portugal and southernFrance between the Mousterian and Mesolithic periods. Correlations e Artiodactyla,dashed line (r2 ¼ 0.47; n ¼ 12; P ¼ 0.019), Perrisodactyla, dotted line (r2 ¼ 0.91; n ¼ 6;P ¼ 0.003), Carnivora, black line (r2 ¼ 0.32; n ¼ 15; P ¼ 0.033). The position of therabbit in the graph is indicated.
J.E. Fa et al. / Journal of Human Evolution xxx (2013) 1e9 3
contribution to the standing biomass made by carnivores, herbi-vores and megaherbivores (>1000 kg, proboscideans and rhinos)during each time period.
Changes in hunted prey size and rabbit consumption patterns
Most analyses of relative skeletal abundances in archaeologicalcontexts are based on units derived from the number of identifiedspecimens (NISP): the minimum number of elements (MNE), theminimal animal unit (MAU), and the skeletal-element based min-imum number of individuals (MNI). Predictable relationships be-tween the values of NISP, MNE, MAU, and MNI within any givenassemblage can be expected (Grayson and Frey, 2004). For all sitesin this study, we were able to obtain NISP but not MNI, MNE orMAU. As expected, MNI and NISP for this dataset were significantlyand positively correlated (r2 ¼ 0.78; P ¼ 7.8E-16; n ¼ 43).
We used the NISP for species appearing in rabbit-sites (Table S2)multiplied by their body mass to calculate mean body mass of preymammals during the Middle and Upper Palaeolithic and Mesolithic.Weassumed that the species and their taphonomic abundanceduringthese chronological phases reflected the use of prey species by hom-inins (although see the explanation of caveats around rabbit bonesdiscussed below).We employed phases rather than technocomplexeshere because the number of sites with NISP data per time period wassmall. We also estimated the mean body mass for each mammalianassemblage to compare with the mean prey body mass.
We used published NISP for all macromammals to calculate themean (standard error � se) of rabbit presence from the proportionof rabbit NISP relative to all NISP excavated for each site during eachtime period (Table S2). We only included species in our analysesthat had a minimum of 10 NISP in each site.
The assessment of the proportions of mammal remains onarchaeological sites is likely to be affected by recovery bias (Payne,1972). However, given that most studies in our metanalysis wererelatively recent (1980s onwards) when excavation techniqueswere comparatively standardised and methodologically thorough(Lee Lyman, 1994), sieving of bone fragments would have includedalso smaller species such as those of rabbits. Thus, there is no reasonto suppose that Middle Palaeolithic sites were less likely to havebeen sampled for smaller remains than Upper Palaeolithic ones.
We discuss the origin, anthropic or other, of the archaeologicalrabbit remains in our study to investigate the importance of thissmall prey for hominins. Taphonomic studies on this kind of ma-terial have proved to be an invaluable source of information todistinguish between competing predators. Results show that ana-tomical representation and breakage patterns differ depending onthe origin of the assemblage. In addition, anthropic activity isusually revealed by the presence of cut marks and burnt damage onrabbit bones, whilst other evidence such as tooth/beak marks,gnawing damage or the presence of digested remains, points toa non-anthropic agent (Pérez Ripoll and Martínez Valle, 2001;Hockett and Haws, 2002; Cochard and Brugal, 2004; Lloveras et al.,2010, 2011).
Results
Species encounter frequencies
Crude population densities and body mass of species appearingin all sites were significantly negatively correlated (Fig. 1). Slopesdiffered according to mammalian order with carnivores and ar-tiodactyls exhibiting gentler slopes than perissodactyls. Rabbitdensities were two orders of magnitude greater than similar-sizedrodents and larger than for any other recorded mammal species(Fig. 1).
Please cite this article in press as: Fa, J.E., et al., Rabbits and hominin sur10.1016/j.jhevol.2013.01.002
Faunal transitions
Archaeofaunal assemblages from sites in Iberia and southernFrance show a pattern of changing taxonomic diversity from theMousterian to theMesolithic.We recorded a total of 64mammaliantaxa (Table 1); 14 carnivores, 20 herbivores and three mega-herbivores. Only 11 species persisted during all time periods.Megaherbivores were not recorded beyond the Mousterian, andlarge carnivores such as lions, hyenas, leopards and dholes, equidsand saiga antelope had disappeared by the Magdalenian. A total of16 species were large-bodied taxa (>100 kg): three carnivores and13 herbivores (Table 1). Overall species richness was highest duringthe Mousterian (warm phase e 27 taxa; cold phase e 28 taxa) withmore than half of the species being large-bodied (warm phase: 10species; cold phase: 12 species). However, megaherbivores dis-appeared by the Aurignacian, and the number of large-bodiedmacromammal species dropped progressively until the Mesolithic(Table 2). By the latter period, the mammalian assemblage wasmade up of only 15 species, five of which were >100 kg.
Changes in hunted prey body size
For each of the threemain timephases, the calculatedmeanbodymass of prey species was greater than 100 kg; large-bodied speciesweremostly hunted. Mean prey bodymass was larger (188.3 kg) forthe Middle Palaeolithic, fell to 158.2 kg for the Upper Palaeolithic,and dropped again to 121.2 kg in the Mesolithic. In comparison,mean body mass of mammalian assemblages during each timephasewas399.9 kg in theMiddle Palaeolithic,150.06kg in theUpperPalaeolithic and similar (145.9 kg) in the Mesolithic (Table 2). Thus,though in all phases hunting of the larger species prevailed, themean body mass of animals hunted during the Middle Palaeolithicreflected the larger body sizes of themammalian faunas at the time.Despite the fact that the background mean body mass of macro-mammals during the Upper Palaeolithic and Mesolithic did notdiffer significantly, hunted prey size was much lower.
vival in Iberia, Journal of Human Evolution (2013), http://dx.doi.org/

Table 1Mammalian species assemblages for time periods from the Mousterian to the Mesolithic in Iberia and southern France.
Order/species Body mass(kg)
Population density(individuals/km�2)
Biomass(kg/km�2)
Middle Palaeolithic Upper Palaeolithic Mesolithic
Mousterian(warm)
Mousterian(cold)
Aurignacian Gravettian Solutrean Magdalenian Mesolithic
ProboscideaStraight-tusked elephant Elephas (Palaeoloxodon)
antiquusa3940.03 0.08 315.20 X
Woolly mammoth Mammuthus primigeniusa 5500.00 0.08 440.00 XPrimatesBarbary macaque Macaca sylvanusb 11.50 12.50 143.75 XRodentiaEuropean beaver Castor fiber 19.00 2.00 38.00 X XAlpine marmot Marmota marmota 4.76 4.84 23.04 XLagomorphaBrown hare Lepus europaeus/capensis 2.93 14.75 43.22 X X X X X X XArctic hare Lepus timidus 3.12 14.75 46.02 XEuropean rabbit Oryctolagus cuniculus 1.59 357.90 569.06 X X X X X X XCarnivoraEuropean badger Meles meles 11.88 2.52 29.94 X X X X X X XWolverine Gulo gulo 14.53 0.0079 0.12 XBrown bear Ursus arctos 196.29 0.02 3.93 X X X X X X XCave bear Ursus spelaeaa 206.00 0.02 4.12 X X XIberian lynx Lynx pardina 11.05 0.32 3.54 X X X X X X XLeopard Panthera pardusb 52.40 0.07 3.67 X X X X XLion Panthera leob 158.62 0.11 17.45 X X X XWild cat Felis sylvestris 4.57 2.95 13.48 X X X X X X XArctic fox Alopex lagopus 3.50 0.10 0.35 X XArctic fox/red fox Alopex lagopus/Vulpes vulpes 3.60 0.10 0.36 X XDhole Cuon alpinusb 12.76 0.04 0.51 X XRed fox Vulpes vulpes 4.82 1.10 5.30 X X X X X X XWolf Canis lupus 31.76 0.01 0.32 X X X X X X XSpotted hyena/Cave hyena Crocuta crocuta/Hyaena spelaeab 63.37 0.13 8.24 X XPerissodactylaHorse Equus caballusa 400.00 4.05 1620.00 X X X X X X XEuropean ass Equus hydruntinusa 230.00 4.05 931.50 X X X X X XEuropean soft nose rhinoceros Dicerorhinus hemitoechusa 1266.67 0.21 266.00 XMerck’s rhinoceros Dicerorhinus (Stephanorhinus)
mercki/D. kirckbergensi/Woolly rhinoceros Coelodontaantiquitatisa
2000.00 0.21 420.00 X
ArtiodactylaEurasian elk Alces alces 355.00 0.40 142.00 XAuroch Bos sp./B. primigeniusa 613.00 0.34 208.42 X X X X X X XAuroch/Bison Bos sp./Bison sp. 643.00 0.33 212.19 X X X X X X XPyrenean ibex Capra ibex 69.30 2.00 138.60 X X X XIberian wild goat Capra pyrenaica 60.20 2.00 120.40 X X X X X X XRoe deer Capreolus capreolus 22.50 7.61 171.23 X X X X X XRed deer Cervus elaphus 217.75 2.90 631.48 X X X X X XMuskox Ovibos moschatus 313.00 0.32 100.16 XReindeer Rangifer tarandus 109.09 2.59 282.54 X XChamois Rupicapra rupicapra 32.80 2.00 65.60 X X X X X X XSaiga antelope Saiga tataricab 37.40 0.75 28.05 XWild boar Sus scrofa 84.50 3.57 301.67 X X X X X X
Species lists were taken from 104 sites listed in Supplementary Table S1 and from 254 Mousterian and Aurignacian sites in Stewart et al. (2003). Population density and body mass data for extant species from Jones et al. (2009).Body mass for extinct taxa from Smith et al. (2003).
a Globally extinct.b Extinct in Western Europe.
J.E.Faet
al./Journal
ofHum
anEvolution
xxx(2013)
1e9
4Pleasecite
thisarticle
inpress
as:Fa,J.E.,et
al.,Rabbitsand
hominin
survivalinIberia,Journalof
Hum
anEvolution
(2013),http://dx.doi.org/10.1016/j.jhevol.2013.01.002

Table
2Stan
dingbiom
ass,sp
eciesnumbe
rsan
dmea
nbo
dymassof
macromam
mals,separated
into
carn
ivores,h
erbivo
resan
dmeg
aherbivo
res,in
Iberia
andsouthernFran
ceacco
rdingto
themaincu
lturaltimeperiods.
Carnivores
Herbivo
res
Meg
aherbivo
res2
Bod
ymass(kg)
Totals
Biomass
(kg/km
�2)
Percen
tof
totalbiom
ass
Total
species
Large-bo
died
speciesa
Biomass
(kg/km
�2)
Percen
tof
totalb
iomass
Total
species
Large-bo
died
species
Biomass
(kg/km
�2)
Percen
tof
totalb
iomass
Total
species
Large-bo
died
species
Mea
nStan
dard
error
Med
ian
Total
species
Total
biom
ass
(kg/km
�2)
Middle
Palae
olith
icMou
sterian(w
arm)
90.5
1.5
113
5195
.188
.614
558
1.2
9.9
22
309.9
149.6
60.2
2758
66.8
Mou
sterian(cold)
61.5
1.1
102
4721
.183
.716
886
0.0
15.2
22
400.7
203.2
46.5
2856
42.6
Upper
Palae
olith
icAurign
acian
91.2
1.7
133
5295
.998
.313
6e
ee
124.9
34.8
56.3
2653
87.1
Grave
ttian
77.6
1.9
82
4043
.998
.112
5e
ee
142.4
43.8
56.3
2041
21.5
Solutrea
n77
.62.0
82
3905
.398
.111
5e
ee
146.3
46.0
52.4
1939
82.9
Mag
dalen
ian
60.2
1.6
71
3603
.698
.411
5e
ee
145.6
48.6
42.6
1836
63.8
Mesolith
icMesolithic
56.5
1.4
61
3900
.198
.69
4e
ee
155.7
57.1
32.8
1539
56.6
aSp
ecies>10
0kg
.bSp
ecies>10
00kg
.
J.E. Fa et al. / Journal of Human Evolution xxx (2013) 1e9 5
Please cite this article in press as: Fa, J.E., et al., Rabbits and hominin sur10.1016/j.jhevol.2013.01.002
Standing biomass changes and contribution of the rabbit
Overall mammalian standing biomass in Iberia/southern France(Fig. 2) varied from a maximum of 5866.9 kg/km�2 during theMousterian (cold) to a minimum of 3956.6 kg/km�2 by the Meso-lithic (Table 2). This drop in standing biomass throughout time wassignificant (r2 ¼ 0.75; P ¼ 0.012). As expected, terrestrial biomassdecreased markedly at each higher trophic level. Thus, for all timeperiods pooled, carnivores contributed a mean of 1.60 � 0.12% ofthe standing biomass whereas herbivores made up 94.83 � 2.30%(Table 2). During the Mousterian, megaherbivores supplied12.55 � 2.65% of the total biomass.
Percentage of the total terrestrial standing biomass contributedby the rabbit (Fig. 2) rose significantly from around 10% in theMousterian to about 15% by the Mesolithic (r2 ¼ 0.71; P ¼ 0.018).This should be considered from the perspective that rabbit standingbiomass would have been among the five highest for all speciesrecorded during theMousterian in the study, just lower than equidsEquus spp. and the red deer Cervus elaphus (see Table 1).
Rabbit consumption patterns
The average number of NISP recorded per taxon was449.21 � 56.17 (Median 232; range 11e3554; total NISP 45,370).Presence of the rabbit through the different time periods increasedfrom 37.8 � 9.2% per site in the Mousterian, to 75.1 � 2.9% during
Figure 2. Changes in the overall terrestrial mammalian standing biomass (kg/km�2)(grey bars), calculated for faunal assemblages from the Mousterian to the Mesolithic.The black line denotes the percentage contribution to the overall standing biomassmade by the rabbit for each time period. MP ¼ Middle Palaeolithic; UP ¼ UpperPalaeolithic; M ¼ Mesolithic. In this study, data for the Mousterian are from the latterstages of this period, between 70,000 and 40,000 BP, where warm and cold climaticregions were present. Here we represent faunal assemblages for these two types ofenvironments. The Iberian Peninsula constituted one of the southernmost limits of thesteppe-tundra expanse and was occupied by these faunas during the coldest periods(Álvarez-Lao and García, 2011). In the Iberian fossil record, the presence of cold-adapted mammal species is occasional and restricted to specific time spans and, inmost cases, to limited geographical areas. The sites that have yielded cold-adaptedspecies are distributed irregularly along the Iberian Peninsula, most of them locatedin the north, where the climatology and landscapes are more similar to central Eu-ropean localities. It is likely that the Pyrenees and Cantabrian Mountain Range func-tioned as geographic barriers for some of these species. This faunal community isclosely related with the steppe-tundra characteristic of the Pleistocene glacial periods,disappeared at the end of the Pleistocene alongside the mammoth fauna. After the endof the Pleistocene. Some of these species (reindeer, musk-ox and arctic fox) movednorthward to arctic areas, a few others (as saiga antelope) migrated to the central Asiansteppe and some other species (as woolly mammoth and woolly rhinoceros) becameextinct. The loss of these megaherbivores is clearly observed in the drop in biomassshown in the graph. Although climatic differences in the north and south of the IberianPeninsula also occurred within the other time periods, we have only differentiate thesefor the Mousterian because of the distinctiveness of the cold-adapted faunas thatmigrated from Eurasia.
vival in Iberia, Journal of Human Evolution (2013), http://dx.doi.org/
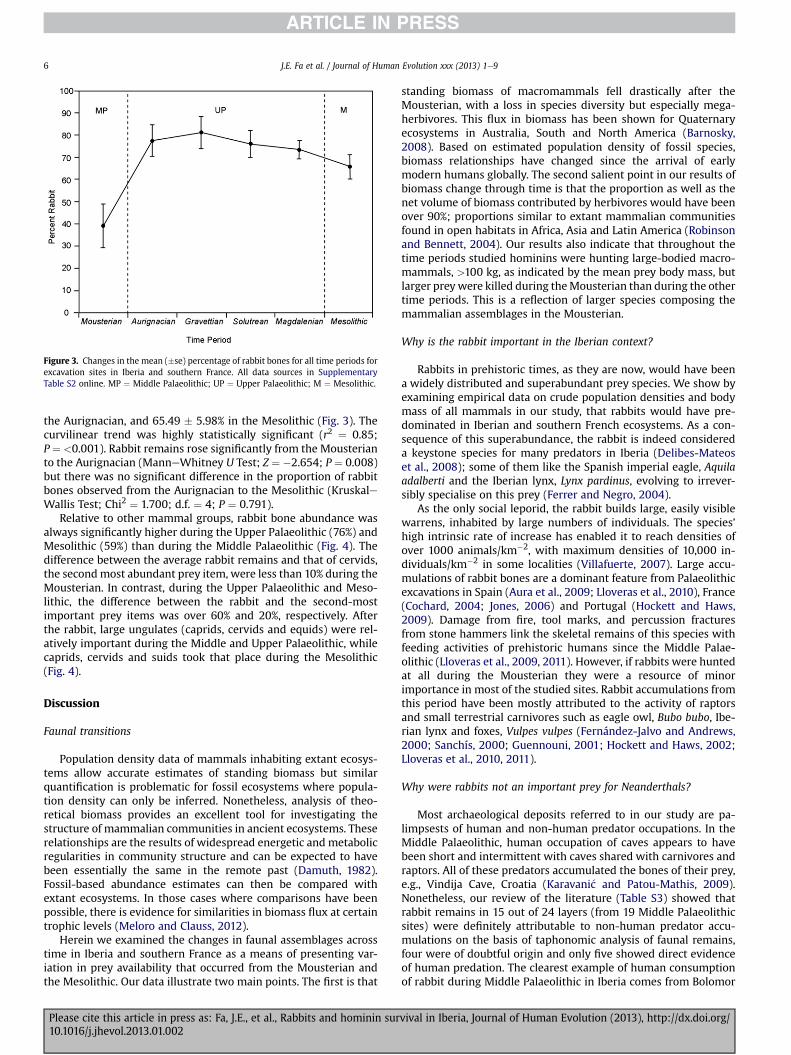
Figure 3. Changes in the mean (�se) percentage of rabbit bones for all time periods forexcavation sites in Iberia and southern France. All data sources in SupplementaryTable S2 online. MP ¼ Middle Palaeolithic; UP ¼ Upper Palaeolithic; M ¼ Mesolithic.
J.E. Fa et al. / Journal of Human Evolution xxx (2013) 1e96
the Aurignacian, and 65.49 � 5.98% in the Mesolithic (Fig. 3). Thecurvilinear trend was highly statistically significant (r2 ¼ 0.85;P ¼ <0.001). Rabbit remains rose significantly from the Mousterianto the Aurignacian (ManneWhitney U Test; Z ¼ �2.654; P ¼ 0.008)but there was no significant difference in the proportion of rabbitbones observed from the Aurignacian to the Mesolithic (KruskaleWallis Test; Chi2 ¼ 1.700; d.f. ¼ 4; P ¼ 0.791).
Relative to other mammal groups, rabbit bone abundance wasalways significantly higher during the Upper Palaeolithic (76%) andMesolithic (59%) than during the Middle Palaeolithic (Fig. 4). Thedifference between the average rabbit remains and that of cervids,the secondmost abundant prey item, were less than 10% during theMousterian. In contrast, during the Upper Palaeolithic and Meso-lithic, the difference between the rabbit and the second-mostimportant prey items was over 60% and 20%, respectively. Afterthe rabbit, large ungulates (caprids, cervids and equids) were rel-atively important during the Middle and Upper Palaeolithic, whilecaprids, cervids and suids took that place during the Mesolithic(Fig. 4).
Discussion
Faunal transitions
Population density data of mammals inhabiting extant ecosys-tems allow accurate estimates of standing biomass but similarquantification is problematic for fossil ecosystems where popula-tion density can only be inferred. Nonetheless, analysis of theo-retical biomass provides an excellent tool for investigating thestructure of mammalian communities in ancient ecosystems. Theserelationships are the results of widespread energetic and metabolicregularities in community structure and can be expected to havebeen essentially the same in the remote past (Damuth, 1982).Fossil-based abundance estimates can then be compared withextant ecosystems. In those cases where comparisons have beenpossible, there is evidence for similarities in biomass flux at certaintrophic levels (Meloro and Clauss, 2012).
Herein we examined the changes in faunal assemblages acrosstime in Iberia and southern France as a means of presenting var-iation in prey availability that occurred from the Mousterian andthe Mesolithic. Our data illustrate two main points. The first is that
Please cite this article in press as: Fa, J.E., et al., Rabbits and hominin sur10.1016/j.jhevol.2013.01.002
standing biomass of macromammals fell drastically after theMousterian, with a loss in species diversity but especially mega-herbivores. This flux in biomass has been shown for Quaternaryecosystems in Australia, South and North America (Barnosky,2008). Based on estimated population density of fossil species,biomass relationships have changed since the arrival of earlymodern humans globally. The second salient point in our results ofbiomass change through time is that the proportion as well as thenet volume of biomass contributed by herbivores would have beenover 90%; proportions similar to extant mammalian communitiesfound in open habitats in Africa, Asia and Latin America (Robinsonand Bennett, 2004). Our results also indicate that throughout thetime periods studied hominins were hunting large-bodied macro-mammals, >100 kg, as indicated by the mean prey body mass, butlarger preywere killed during theMousterian than during the othertime periods. This is a reflection of larger species composing themammalian assemblages in the Mousterian.
Why is the rabbit important in the Iberian context?
Rabbits in prehistoric times, as they are now, would have beena widely distributed and superabundant prey species. We show byexamining empirical data on crude population densities and bodymass of all mammals in our study, that rabbits would have pre-dominated in Iberian and southern French ecosystems. As a con-sequence of this superabundance, the rabbit is indeed considereda keystone species for many predators in Iberia (Delibes-Mateoset al., 2008); some of them like the Spanish imperial eagle, Aquilaadalberti and the Iberian lynx, Lynx pardinus, evolving to irrever-sibly specialise on this prey (Ferrer and Negro, 2004).
As the only social leporid, the rabbit builds large, easily visiblewarrens, inhabited by large numbers of individuals. The species’high intrinsic rate of increase has enabled it to reach densities ofover 1000 animals/km�2, with maximum densities of 10,000 in-dividuals/km�2 in some localities (Villafuerte, 2007). Large accu-mulations of rabbit bones are a dominant feature from Palaeolithicexcavations in Spain (Aura et al., 2009; Lloveras et al., 2010), France(Cochard, 2004; Jones, 2006) and Portugal (Hockett and Haws,2009). Damage from fire, tool marks, and percussion fracturesfrom stone hammers link the skeletal remains of this species withfeeding activities of prehistoric humans since the Middle Palae-olithic (Lloveras et al., 2009, 2011). However, if rabbits were huntedat all during the Mousterian they were a resource of minorimportance in most of the studied sites. Rabbit accumulations fromthis period have been mostly attributed to the activity of raptorsand small terrestrial carnivores such as eagle owl, Bubo bubo, Ibe-rian lynx and foxes, Vulpes vulpes (Fernández-Jalvo and Andrews,2000; Sanchís, 2000; Guennouni, 2001; Hockett and Haws, 2002;Lloveras et al., 2010, 2011).
Why were rabbits not an important prey for Neanderthals?
Most archaeological deposits referred to in our study are pa-limpsests of human and non-human predator occupations. In theMiddle Palaeolithic, human occupation of caves appears to havebeen short and intermittent with caves shared with carnivores andraptors. All of these predators accumulated the bones of their prey,e.g., Vindija Cave, Croatia (Karavani�c and Patou-Mathis, 2009).Nonetheless, our review of the literature (Table S3) showed thatrabbit remains in 15 out of 24 layers (from 19 Middle Palaeolithicsites) were definitely attributable to non-human predator accu-mulations on the basis of taphonomic analysis of faunal remains,four were of doubtful origin and only five showed direct evidenceof human predation. The clearest example of human consumptionof rabbit during Middle Palaeolithic in Iberia comes from Bolomor
vival in Iberia, Journal of Human Evolution (2013), http://dx.doi.org/

Figure 4. Mean percentage (�se) contribution of rabbit and other main mammalian groups derived from number of identified specimens recovered from excavation sites in Spain,Portugal and southern France. All NISP data and sources in Supplemental Tables S1eS3 online.
J.E. Fa et al. / Journal of Human Evolution xxx (2013) 1e9 7
Cave in Valencia, 350 kyr BP (Martínez Valle, 2001; Sanchís Serraand Fernández Peris, 2008). In contrast, all rabbit accumulationsfrom 27 layers in four Aurignacian, five Gravettian, six Solutrean,five Magdalenian, and four Mesolithic sites were associated withmodern human archaeology (Aura et al., 2009). Although it seemsunlikely that non-human predators were never involved in rabbitaccumulation in Upper Palaeolithic sites, the involvement ofhumans is very much clearer than in Middle Palaeolithic sites.
The lower rabbit bone proportion appearing for the MiddlePalaeolithic, which we argue is animal predator and not homininrelated, contrasts against the abundance of other larger body sizedquarry. This is also reflected in the overall higher species numbersand mammalian standing biomass during the Middle Palaeolithic.But, even though ungulates are consumed during all periods therelative proportions of rabbit for the Upper Palaeolithic and Mes-olithic periods were higher than during the Middle Palaeolithic.Our evidence indicates that hominin diets may have changed fromone dependent on largemammals during theMiddle Palaeolithic todiets dominated by smaller species in the Upper Palaeolithic, inparticular by the wild rabbit. The explanation for this transition iseither that climate change negatively impacted large mammalpopulations, forcing prehistoric people to incorporate smaller ver-tebrate species into their diets, or that people themselves neg-atively impacted large mammal populations, which in turn forcedforagers to add smaller species to their diets.
Whatever the reasons for the loss of megaherbivores, our dataon potential productivity and biomass of the rabbit points to theIberian Peninsula holding abundant prey of easy access availablefor human consumption. Given this, it is possible that the latesurvival of Neanderthals in Iberia may have been related to morefavourable climatic conditions here (Grayson and Cannon, 1999).However, if low levels of rabbit consumption by Neanderthalscontinued after the loss of the herbivorous megafauna, was thishominin’s survival handicapped? Residual faunas made up of highdensity fast-reproducing species are very often the chief supportof modern hunter communities (Cowlishaw et al., 2005). The factthat the rabbit was extensively exploited by AMH can be seen asthis hominin taking full advantage of a steady and abundantsource of meat. An alternative explanation for the difference indeposition of rabbit bones between the Middle Palaeolithic andother periods could be that Neanderthals processed rabbit else-where and their remains were not deposited in Mousterian caves.Yet, this would be a departure from what is understood to be thenorm in prehistoric hunteregatherer societies, including Nean-derthals, where prey would be taken back to shelter for processingand consumption. The inescapable fact from our data is that the
Please cite this article in press as: Fa, J.E., et al., Rabbits and hominin sur10.1016/j.jhevol.2013.01.002
mean percentage of rabbit bones approximately doubles betweenthe Mousterian and the Aurignacian and remains consistently highup until the Mesolithic. Even if we assumed that all of the rabbitremains found in the Mousterian sites were of anthropogenicorigin, the trend from low use to substantial consumption is highlysignificant.
Was mass collection of rabbits easier for AMH?
The volume of rabbit remains found in some excavation sitesfrom the Upper Palaeolithic and Mesolithic has been explained interms of mass collecting of the species (Jones, 2006). Evidence ofmass collecting of rabbits in south-western France at thePleistoceneeHolocene transition is available (Jones, 2006) andemerging data from theMolí del Salt in north-eastern Spain (Ibáñezand Saladié, 2004), and other sites in Iberia also support this(Hockett and Bicho, 2000). Rabbit mass collecting is of significancebecause this form of prey acquisition alters the relationship be-tween prey body size and energetic return (Madsen and Schmitt,1998; Jones, 2004; Ugan, 2005). Current hypotheses about chang-ing diet breadth at the PleistoceneeHolocene transition rest on theassumption that rabbits would have been a neglected prey itembecause it is lower in energetic value than large ungulates. How-ever, mass collecting events may invalidate this assumption despiteincreased handling and start-up costs associated with this techni-que. This is because warren-based harvests would have providedhigh numbers of animals in a relatively short space of time (Hockettand Haws, 2003). And, even though small prey taken in mass mayappear lower ranked in energetic gain per unit of effort than largerpackage items, the increase in harvest efficiency would have ren-dered rabbits collected directly from the warren as higher-rankedresources (Jones, 2006). This is so because individual rabbits(adults typically ranging from 1 to 1.5 kg) on average provide1.98 kcal/g. If up to 60 rabbits are present in a warren, an overalltake from a full warren could provide over 50,000 total kilocalories(Jones, 2006). We suggest that this prey item would have beena significant source of meat for AMH societies. Gender-based divi-sion of labour may have been possible, in which women may haveconcentrated on warren harvesting and men on the pursuit oflarger prey. Additionally, the role of dogs in hunting by AMH hasbeen recently highlighted as affording an adaptive advantage forthis hominin over Neanderthals by being able to hunt smaller preymore efficiently (Germonpré et al., 2012; Shipman, 2012). Thehigher level of hunting of rabbits observed in this study during theUpper Palaeolithic and Mesolithic, and by AMH since Neanderthals
vival in Iberia, Journal of Human Evolution (2013), http://dx.doi.org/

J.E. Fa et al. / Journal of Human Evolution xxx (2013) 1e98
had already disappeared could have been facilitated by huntingdogs, as is common practice in present times (Lupo, 2011).
More refined analyses on how climatic changes over time mayhave affected faunal distribution and abundance in Iberia arerequired. Though changes in absolute climate values and variabilitywould have affected faunal assemblages and individual speciesranges, it is unlikely that the biomass relationships we present herewould have differed, given the strong empirical rule associatedbetween body mass and abundance. It is likely, however, that theloss of megaherbivores, associated with fluctuating climates orhuman overkill created an enormous gap in standing biomass anda loss of large animals which would have shifted hominin survivalpatterns. We contend that if Neanderthals selected large prey tohunt, they would have had two basic options once the megafaunabecame scarcer. One optionwould have been to track these animalsas their ranges contracted. A second option, certainly not supportedby our data, would have been for Neanderthals to become moresedentary and to shift hunting emphasis onto the remainingsmaller prey. If for whatever reason Neanderthals did not adapt toresidual faunas, it is possible that this hominin would have beenseverely limited.
As suggested by Hockett and Haws (2003), although many fac-tors affect mortality in human populations, nutrition in the form ofdiverse essential nutrient intake may greatly influence maternaland foetal-to-infant mortality. This may have occurred in the de-mographic revolution in Europe when all Neanderthal populationsdisappeared. Alongside evidence from stable isotopes, faunal re-mains, and the palaeopathology of human skeletons we add thesuggestion that the specialised diet of Neanderthals consisting oflarge and medium-sized terrestrial herbivores may have madethemmore vulnerable at a timewhen these animals disappeared orbecame scarce. There is little doubt that Neanderthals were largegame specialists and lived this lifestyle for around 300,000 years.Nothing like this is seen in the archaeological record of modernhumans with the exception of people living in environments whereprey-switching really is not an option, like in grasslands and tundra.Neanderthals may have been particularly invariant in terms of theuse of smaller prey, as shown by the rather static technology andzooarchaeological evidence for the Mousterian.
Neanderthals may have been less capable of prey-switching (forwhatever reason), at least to the extent that AMH were. It is alsoplausible that they could have been less able to gather faster,smaller prey at a rate that could satisfy their daily nutritional needsbecause of anatomical constraints as suggested by Steudel-Numbers and Wall-Scheffler (2009). Differences in division of la-bour (Kuhn and Stiner, 2006) or even feeding preferences (gen-eralist versus specialist) could also explain the contrasting use ofthe rabbit by Neanderthals versus AMH. But whatever the reason,there is sufficient evidence to argue that AMH took maximumadvantage of the wild rabbit as a unique and abundant foodresource in the Iberian Peninsula whilst Neanderthals did not. Thelatter hominin may have resorted to foraging on coastal molluscsrather than pursuing smaller motile prey (Stiner et al., 1999; Cortés-Sánchez et al., 2011).
The nature and causes of the disappearance of Neanderthals andtheir apparent replacement by modern humans will still be thesubject of considerable debate. Many researchers have proposedbiologically or technologically mediated dietary differences be-tween the two groups as one of the fundamental causes of Nean-derthal disappearance. Our intention here was to open another lineof discussion by systematically examining the question, what ifNeanderthals were ‘large game specialists’ and were unable toexploit residual prey after megafaunal collapse. The Iberian Pen-insula, where Neanderthals survived longer than anywhere inEurope, was unique in containing a prey species of extraordinary
Please cite this article in press as: Fa, J.E., et al., Rabbits and hominin sur10.1016/j.jhevol.2013.01.002
abundance and wide distribution. The rabbit would have provideda steady volume of meat to hunters as we indicate from our datasetof faunal remains from late Pleistocene archaeological sitesthroughout Iberia and SW France. By examining the relationshipbetween body size and archaeological abundance, we found thatlarge mammals were the most abundant and regularly occurringanimals in Middle Palaeolithic faunal assemblages with rabbit re-mains more generally attributable to non-human predation.Because there is a strong inverse correlation between body size andpopulation density among animals, we argued that this faunal re-cord could only have been produced by hunteregatherers who didnot take up opportunities to take small game in favour of concen-trating on larger prey.
Acknowledgements
We thank C. Finlayson, B. Hockett, E.L. Jones, E.J. Milner-Gulland,J. Nadal, T. Surovell and three anonymous referees made usefulcomments on the paper. Amy Hall drew the figures and JesusDuarte helped with the statistics. J. Mario Vargas was supported bythe Programa Nacional deMovilidad de Recursos Humanos del PlanNacional I þ D þ i 2008e11 of the Ministerio de Educación, Spain.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.jhevol.2013.01.002.
References
Álvarez-Lao, D.J., García, W.D., 2011. Geographical distribution of Pleistocene coldadapted large mammal faunas in the Iberian Peninsula. Quatern. Int. 233,159e170.
Aura, J.E., Jordá, J.F., Morales, J.V., Pérez, M., Villalba, M.-P., Alcover, J.A., 2009. Eco-nomic transitions in finis terra: the western Mediterranean of Iberia, 15e7 ka BP. Before Farming 2, 1e17.
Aura, J.E., Villaverde, V., Pérez Ripoll, M., Martínez, R., Guillem, P., 2002. Big gameand small prey: Paleolithic and Epipaleolithic economy from Valencia (Spain).J. Archaeol. Method. Th. 9, 209e262.
Barnosky, A., 2008. Megafauna biomass tradeoff as a driver of Quaternary andfuture extinctions. Proc. Natl. Acad. Sci. 105, 11543e11548.
Blasco, R., Fernández Peris, J., 2009. Middle Pleistocene bird consumption at level XIof Bolomor cave (Valencia, Spain). J. Archaeol. Sci. 36, 2213e2223.
Balmford, A., 1996. Extinction filters and current resilience: the significance of pastselection pressures for conservation biology. Trends Ecol. Evol. 11, 193e196.
Branco, M., Ferrand, N., Monnerot, M., 2000. Phylogeography of the European rabbit(Oryctolagus cuniculus) in the Iberian Peninsula inferred from RFLP analysis ofthe cytochrome b gene. Heredity 85, 307e317.
Brown, J.H., 1995. Macroecology. University of Chicago Press, Chicago.Broughton, J.M., Cannon, M.D., Bayham, F.E., Byers, D.A., 2011. Prey body size and
ranking in zooarchaeology: theory, empirical evidence, and applications fromthe northern Great Basin. Am. Antiq. 76, 403e428.
Brown, K., Fa, D.A., Finlayson, G., Finlayson, C., 2011. Small game and marineresource exploitation by Neanderthals: the evidence from Gibraltar. In:Bicho, N.F. (Ed.), Trekking the Shores: Changing Coastlines and the Antiquity ofCoastal Settlement Interdisciplinary Contributions to Archaeology. SpringerScience. http://dx.doi.org/10.1007/9 469 78-1-4419-8219-3_10.
Callou, C., 1995. Modifications de l’aire de répartition du Lapin (Oryctolagus cuni-culus) en France et en Espagne, du Pléistocène à l’époque actuelle. État de laquestion. Anthropozoologica 21, 95e114.
Cannon, M.D., 2000. Large mammal relative abundance in Pithouse and Puebloperiod archaeofaunas from southwestern New Mexico: resource depressionamong the Mimbres-Mogollon? J. Anthropol. Archaeol. 19, 317e347.
Cochard, D., 2004. Les léporidés dans la subsistance Paléolithique du sud de laFrance. Ph.D. Dissertation, Université Bordeaux I, Bordeaux.
Cochard, D., Brugal, J.P., 2004. Importance des fonctions de sites dans les accumu-lations paléolithiques de Léporidés. In: Brugal, J.P., Desse, J. (Eds.), Petits Ani-maux et Sociétés Humaines. Du Complément Alimentaire Aux RessourcesUtilitaires, pp. 283e296 (Actes des XXIVe Rencontres Internationales d’Arch-éologie et d’Histoire d’Antibes: Editions APDCA).
Cortés-Sánchez, M., Morales-Muñiz, A., Simón-Vallejo, M.D., Lozano-Francisco, M.C.,Vera-Peláez, J.L., et al., 2011. Earliest known use of marine resources by Nean-derthals. PLoS ONE 6 (9), e24026.
vival in Iberia, Journal of Human Evolution (2013), http://dx.doi.org/

J.E. Fa et al. / Journal of Human Evolution xxx (2013) 1e9 9
Corbet, G.B., 1994. Taxonomy and origins. In: Thompson, K., King, C.M. (Eds.), TheEuropean Rabbit. The History and Biology of a Successful Colonizer. OxfordUniversity Press, Oxford, pp. 1e7.
Cowlishaw, G., Mendelson, S., Rowcliffe, J.M., 2005. Evidence for post-depletionsustainability in a mature bushmeat market. J. Appl. Ecol. 42, 460e468.
Damuth, J., 1982. Analysis of the preservation of community structure in assem-blages of fossil mammals. Paleobiology 8, 434e446.
Delibes-Mateos, M., Delibes, M., Ferreras, P., Villafuerte, R., 2008. Key role of Eu-ropean rabbits in the conservation of the western Mediterranean basin hotspot.Conserv. Biol. 22, 1106e1117.
Fernández-Jalvo, Y., Andrews, P., 2000. The taphonomy of Pleistocene caves, withparticular reference to Gibraltar. In: Stringer, C.B. (Ed.), Neanderthals on theEdge. Oxbow Books, Oxford, pp. 171e182.
Ferrer, M., Negro, J.J., 2004. The near extinction of two large European predators:super specialists pay a price. Conserv. Biol. 18, 344e349.
Flux, J.E.C., 1994. World distribution. In: Thompson, H.V., King, C.M. (Eds.), TheEuropean Rabbit. Oxford Science Publications, Oxford, pp. 8e21.
Gamble, C., Davies, W., Pettitt, P., Richards, M., 2004. Climate change and evolvinghuman diversity in Europe during the last glacial. Proc. R. Soc. Lond. B Bio. 359,243e254.
García-Argüelles, P., Nadal, J., Fullola, J.M., 2009. From Magdalenian to Early Neo-lithic: hunter-gatherers in transition in north-eastern Iberia. In: McCartan, S.(Ed.), Mesolithic Horizons. Oxbow Books, Oxford, pp. 500e506.
Germonpré, M., Láznicková-Galetová, M., Sablin, M., 2012. Palaeolithic dog skulls atthe Gravettian Predmostí site, the Czech Republic. J. Archaeol. Sci. 39, 84e202.
Grayson, D., Frey, C.J., 2004. Measuring skeletal part representation in archaeo-logical faunas. J. Taphon. 2, 27e42.
Grayson, D.K., Cannon, M.D., 1999. Human paleoecology and foraging theory in theGreat Basin. In: Beck, C. (Ed.), Human Paleoecology and Foraging Theory in theGreat Basin, Models for the Millennium, Great Basin Anthropology Today.University of Utah Press, Salt Lake City, pp. 141e151.
Guennouni, K.E., 2001. Les Lapins du Pleistocene Moyen et Superieur de QuelquesSites Préhistoriques de L’Europe Mediterranéenne: Terra-Amata, Orgnac 3,Baume Bonne, Grotte du Lazaret, Grotte du Boquete de Zafarraya, Arma delleManie. Etude Paleóntologique, Taphonomique et Archéozoologique. Ph.D. Dis-sertation, Museum National d’Histoire Naturelle de Paris.
Hardy, B.L., Moncel, M.-H., 2011. Neanderthal use of fish, mammals, birds, starchyplants and wood 125e250000 years ago. PLoS ONE 6 (8), e23768.
Henry, A.G., Brooks, A.S., Piperno, D.R., 2011. Microfossils in calculus demonstrateconsumption of plants and cooked foods in Neanderthal diets (Shanidar III,Iraq; Spy I and II, Belgium). Proc. Natl. Acad. Sci. 108, 486e491.
Hockett, B.S., Bicho, N.F., 2000. The rabbits of Picareiro Cave: small mammalhunting during the Late Upper Palaeolithic in the Portuguese Estremadura.J. Archaeol. Sci. 27, 715e723.
Hockett, B.S., Haws, J., 2009. Continuity in animal resource diversity in the LatePleistocene human diet of Central Portugal. Before Farming. 2/2.
Hockett, B.S., Haws, J.A., 2002. Taphonomic and methodological perspectives ofleporid hunting during the Upper Palaeolithic of the western Mediterraneanbasin. J. Archaeol. Method. Th. 9, 269e302.
Hockett, B.S., Haws, J.A., 2003. Nutritional ecology and diachronic trends in Paleo-lithic diet and health. Evol. Anthropol. 12, 211e216.
Ibáñez, N., Saladié, P., 2004. Acquisition anthropique d’Oryctolagus cuniculus dans lesite du Molí del Salt (Catalogne, Espagne). In: Brugal, J.P., Desse, J. (Eds.), PetitsAnimaux et Sociétés Humaines. Du Complément Alimentaire aux RessourcesUtilitaires. XXIVe Rencontres Internationales d’Archéologie et d’Histoire d’An-tibes, pp. 255e260.
Jones, E.L., 2004. Dietary evenness, prey choice, and humaneenvironment in-teractions. J. Archaeol. Sci. 31, 307e317.
Jerozolimski, A., Peres, C.A., 2003. Bringing home the biggest bacon: a cross-siteanalysis of the structure of hunter-kill profiles in neotropical forests. Biol.Conserv. 111, 415e425.
Jones, E.L., 2006. Prey choice, mass collecting, and the wild European rabbit(Oryctolagus cuniculus). J. Anthropol. Archaeol. 25, 275e289.
Jones, K.E., Bielby, J., Cardillo, M., Fritz, S.A., O’Dell, J., Orme, C.D.L., Safi, K.,Sechrest, W., Boaks, E.H., Carbone, C., Connolly, C., Cutts, M.J., Foster, J.K.,Grenyer, R., Habib, M., Plaster, C.A., Price, S.A., Rigby, E.A., Rist, J., Teacher, A.,Bininda-Emonds, O.R.P., Gittleman, J.L., Mace, G.M., Purvis, A., 2009. PanTHERIA:a species-level database of life history, ecology, and geography of extant andrecently extinct mammals. Ecology 90, 2648.
Karavani�c, I., Patou-Mathis, M., 2009. Middle/Upper Paleolithic interface in VindijaCave (Croatia): new results and interpretations. Sourceb. Paleolithic Transit. 4,397e405.
Kuhn, S.L., Stiner, M.C., 2006. What’s a mother to do? The division of labor amongNeandertals and modern humans in Eurasia. Curr. Anthropol. 47, 953e981.
Lee Lyman, R., 1994. Vertebrate Taphonomy. Cambridge University Press,Cambridge.
Lloveras, Ll., Moreno-García, M., Nadal, J., Maroto, J., Soler, J., Soler, N., 2010. Theapplication of actualistic studies to assess the taphonomic origin of Mousterian
Please cite this article in press as: Fa, J.E., et al., Rabbits and hominin sur10.1016/j.jhevol.2013.01.002
rabbit accumulations from Arbreda Cave (North-East Iberia). Archaeofauna 19,99e119.
Lloveras, Ll., Moreno-García, M., Nadal, J., 2009. Butchery, cooking and humanconsumption marks on rabbit (Oryctolagus cuniculus) bones: an experimentalstudy. J. Taphon. 7, 179e201.
Lloveras, Ll., Moreno-García, M., Nadal, J., Zilhão, J., 2011. Who brought in the rab-bits? Taphonomical analysis of Mousterian and Solutrean leporid accumulationsfrom Gruta do Caldeirão (Tomar, Portugal). J. Archaeol. Sci. 38, 2434e2449.
Lupo, K., 2011. A dog is for hunting. In: Albarella, U., Trentacoste, A. (Eds.), Ethno-zooarchaeology. Oxbow Press, Oxford, pp. 4e12.
Madsen, D.B., Schmitt, D.N., 1998. Mass collecting and the diet breadth model:a Great Basin example. J. Archaeol. Sci. 25, 445e455.
Martínez Valle, R., 2001. Cazadores de pequeña presas. In: Villaverde, V. (Ed.), DeNeandertales a Cromañones. El Inicio del Poblamiento Humano en TierrasValèncianas. Universitat de València, València, pp. 129e130.
Meloro, C., Clauss, M., 2012. Predator-prey biomass fluctuations in the Plio-Pleis-tocene. Palaios 27, 90e96.
Payne, S., 1972. Partial recovery and sample bias: the result of some sieving ex-periments. In: Higgs, E.S. (Ed.), Papers in Economic Prehistory. CambridgeUniversity Press, Cambridge, pp. 49e64.
Pérez Ripoll,M.,MartínezValle, R., 2001. La caza, el aprovechamientode las presas yelcomportamiento de las comunidades cazadores prehistóricos. In: Villaverde, V.(Ed.), De Neandertales a Cromañones: El Inicio del Poblamiento Humano en lasTierras Valencianas. Universitat de Valencia, Valencia, pp. 73e98.
Peters, R.H., 1983. The Ecological Importance of Body Size. Cambridge UniversityPress, Cambridge.
Richards, M.P., Pettitt, P.B., Trinkaus, E., Smith, F.H., Paunovi�c, M., Karavani�c, I., 2000.Neanderthal diet at Vindija and Neanderthal predation: the evidence fromstable isotopes. Proc. Natl. Acad. Sci. 97, 7663e7666.
Robinson, J.G., Bennett, E.L., 2004. Having your wildlife and eating it too: an analysisof hunting sustainability across tropical ecosystems. Anim. Conserv. 7, 397e408.
Sanchís Serra, A., Fernández Peris, J., 2008. Procesado y consumo antrópico deconejo en la Cova del Bolomor (Tavernes de la Valldigna, Valencia). El nivelXVIIc (ca 350 ka). Complutum 19, 25e46.
Sanchís, A., 2000. Los restos de Oryctolagus cuniculus en las tafocenosis de Bubobubo y Vulpes vulpes y su aplicación a la caracterización de registro faunísticoarqueológico. Saguntum PLAV 32, 31e50. València.
Shipman, P., 2012. Do the eyes have it? Dog domestication may have helpedhumans thrive while Neandertals declined. Am. Sci. 100, 198e205.
Smith, F.A., Lyons, S.K., Ernest, S.K.M., Jones, K.E., Kaufman, D.M., Dayan, T.,Marquet, P.A., Brown, J.H., Haskell, J.P., 2003. Body mass of late Quaternarymammals. Ecology 84, 3403.
Steudel-Numbers, K., Wall-Scheffler, C., 2009. Optimal running speed and theevolution of hominin hunting strategies. J. Hum. Evol. 56, 355e360.
Stewart, J.R., 2004. Neanderthal-modern human competition? A comparison be-tween the mammals associated with Middle and Upper Palaeolithic industriesin Europe during OIS 3. Int. J. Osteoarchaeol. 14, 178e189.
Stewart, J.R., van Kolfschoten, M., Markova, A., Musil, R., 2003. In: VanAndel, T.H.,Davies, W.D. (Eds.), Neanderthals and Modern Humans in the European Land-scape of the Last Glaciation e Archaeological Results of the Stage 3 Project. TheMcDonald Institute for Archaeological Research, Cambridge, pp. 103e129.Accessed from: http://www.esc.cam.ac.uk/research/researchgroups/oistage3/stage-three-projectdatabase-downloads.
Stiner, M.C., Beaver, J.E., Munro, N.D., Surovell, T.A., 2008. In: Bocquet-Appel, J.P.(Ed.), Modeling Paleolithic Predator-prey Dynamics and the Effects of HuntingPressure on Prey ‘Choice’. Recent Advances in Palaeodemography. Springer,Netherlands, pp. 143e178.
Stiner, M.C., Munro, N.D., Surovell, T.A., Tchernov, E., Bar Yosef, O., 1999. Paleolithicpopulation growth pulses evidenced by small animal exploitation. Science 283,190e194.
Surovell, T.A., Waguespack, N.M., 2009. Human prey choice in the late Pleistoceneand its relation to megafaunal extinctions. In: Haynes, G. (Ed.), AmericanMegafaunal Extinctions at the End of the Pleistocene. Springer Science, NewYork, pp. 77e105.
Ugan, A., 2005. Does size matter? Body size, mass collecting, and their implicationsfor understanding prehistoric foraging behavior. Am. Antiq. 70, 75e89.
Van Valen, L., 1964. Relative abundance of species in some fossil mammal faunas.Am. Nat. 98, 109e116.
Villafuerte, R., 2007. El conejo. In: Palomo, L.J. (Ed.), Atlas y Libro Rojo de losMamíferos Terrestres de España. Dirección General para la Biodiversidad-SECEM-SECEMU, Madrid, pp. 487e489.
Villaverde, V., Martinez-Valle, R., Guillem, P., Fumanal, M., 1996. Mobility and therole of small game in the Middle Palaeolithic of the central region of theSpanish Mediterranean: a comparison of Cova Negra with other Palaeolithicdeposits. In: Carbonell, E. (Ed.), The Last Neandertals, the First AnatomicallyModern Humans. Cambridge University Press, Cambridge, pp. 267e288.
Waguespack, N.M., Surovell, T., 2003. Clovis hunting strategies or how to make outon plentiful resources. Am. Antiq. 68, 333e352.
vival in Iberia, Journal of Human Evolution (2013), http://dx.doi.org/












