
Monitoring presence, abundance and survival probability of the stag beetle, Lucanus cervus, using...
11
ORIGINAL PAPER Monitoring presence, abundance and survival probability of the stag beetle, Lucanus cervus, using visual and odour-based capture methods: implications for conservation Stefano Chiari • Agnese Zauli • Paolo Audisio • Alessandro Campanaro • Pier Francesco Donzelli • Federico Romiti • Glenn P. Svensson • Massimiliano Tini • Giuseppe M. Carpaneto Received: 31 July 2013 / Accepted: 3 February 2014 Ó Springer International Publishing Switzerland 2014 Abstract A capture-mark-recapture study on the threa- tened saproxylic beetle Lucanus cervus was carried out in a chestnut (Castanea sativa) woodland located in northern Italy, using visual encounter surveys (VES) as well as aerial flight interception traps and pitfall traps (PT), both of which were baited with different odour lures. In total, 111 males and 25 females were captured, and VES was by far the most efficient method, accounting for 93 % of first captures, and 95 % of all captures. Stag beetles were not significantly attracted to any tested odour, and many PT were damaged by wild boars (Sus scrofa). Flying males were the most frequent adults observed during the season. The use of a net is necessary to capture the stag beetles, in order to evaluate the population parameters and to assess the local status of threat for the species. Capture data revealed that body size (i.e. body weight and elytron length) influences the survival probability of stag beetles, showing a lower survival probability for larger males. Felt- tip pen marking on the ventral sclerites of head and pronotum is a reliable and long lasting method for marking beetles, as proved by the use of an independent marking procedure with a small drill. Assessing the presence and threat status of L. cervus across Europe is urgently needed, and with no efficient odour-based strategy available, col- lection of adult beetles, dead or alive, by VES is the most reliable way to monitor this emblematic species. Keywords Capture-mark-recapture Á Dead wood Á Italy Á Lucanidae Á Population size estimates Á Saproxylic Introduction Methods for counting animals have evolved over the last 70 years (Andrewartha and Birch 1954) and the study of demographic parameters, such as population abundance S. Chiari Á P. Audisio Á A. Campanaro Department of Biology and Biotechnologies ‘Charles Darwin’, Sapienza University of Rome, Via A. Borelli 50, 00161 Rome, RM, Italy e-mail: [email protected] A. Campanaro e-mail: [email protected] S. Chiari (&) Á A. Campanaro Centro Nazionale per lo Studio e la Conservazione della Biodiversita ` Forestale ‘Bosco Fontana’, Corpo Forestale dello Stato, Strada Mantova 29, 46045 Marmirolo, MN, Italy e-mail: [email protected] A. Zauli Á P. F. Donzelli Á F. Romiti Á M. Tini Á G. M. Carpaneto Department of Sciences, University Roma Tre, Viale G. Marconi 446, 00146 Rome, RM, Italy e-mail: [email protected] P. F. Donzelli e-mail: [email protected] F. Romiti e-mail: [email protected] M. Tini e-mail: [email protected] G. M. Carpaneto e-mail: [email protected] P. F. Donzelli Coordinamento Territoriale per l’Ambiente di Verbania per il Parco Nazionale della Val Grande, Corpo Forestale dello Stato, Piazza Pretorio 1, 28805 Vogogna, VB, Italy G. P. Svensson Department of Biology, Lund University, Ecology Building, Solvegatan 37, 223 62 Lund, Sweden e-mail: [email protected] 123 J Insect Conserv DOI 10.1007/s10841-014-9618-8
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Monitoring presence, abundance and survival probability of the stag beetle, Lucanus cervus, using...

ORIGINAL PAPER
Monitoring presence, abundance and survival probabilityof the stag beetle, Lucanus cervus, using visual and odour-basedcapture methods: implications for conservation
Stefano Chiari • Agnese Zauli • Paolo Audisio • Alessandro Campanaro •
Pier Francesco Donzelli • Federico Romiti • Glenn P. Svensson •
Massimiliano Tini • Giuseppe M. Carpaneto
Received: 31 July 2013 / Accepted: 3 February 2014
� Springer International Publishing Switzerland 2014
Abstract A capture-mark-recapture study on the threa-
tened saproxylic beetle Lucanus cervus was carried out in a
chestnut (Castanea sativa) woodland located in northern
Italy, using visual encounter surveys (VES) as well as
aerial flight interception traps and pitfall traps (PT), both of
which were baited with different odour lures. In total, 111
males and 25 females were captured, and VES was by far
the most efficient method, accounting for 93 % of first
captures, and 95 % of all captures. Stag beetles were not
significantly attracted to any tested odour, and many PT
were damaged by wild boars (Sus scrofa). Flying males
were the most frequent adults observed during the season.
The use of a net is necessary to capture the stag beetles, in
order to evaluate the population parameters and to assess
the local status of threat for the species. Capture data
revealed that body size (i.e. body weight and elytron
length) influences the survival probability of stag beetles,
showing a lower survival probability for larger males. Felt-
tip pen marking on the ventral sclerites of head and
pronotum is a reliable and long lasting method for marking
beetles, as proved by the use of an independent marking
procedure with a small drill. Assessing the presence and
threat status of L. cervus across Europe is urgently needed,
and with no efficient odour-based strategy available, col-
lection of adult beetles, dead or alive, by VES is the most
reliable way to monitor this emblematic species.
Keywords Capture-mark-recapture � Dead wood � Italy �Lucanidae � Population size estimates � Saproxylic
Introduction
Methods for counting animals have evolved over the last
70 years (Andrewartha and Birch 1954) and the study of
demographic parameters, such as population abundance
S. Chiari � P. Audisio � A. Campanaro
Department of Biology and Biotechnologies ‘Charles Darwin’,
Sapienza University of Rome, Via A. Borelli 50,
00161 Rome, RM, Italy
e-mail: [email protected]
A. Campanaro
e-mail: [email protected]
S. Chiari (&) � A. Campanaro
Centro Nazionale per lo Studio e la Conservazione della
Biodiversita Forestale ‘Bosco Fontana’, Corpo Forestale dello
Stato, Strada Mantova 29, 46045 Marmirolo, MN, Italy
e-mail: [email protected]
A. Zauli � P. F. Donzelli � F. Romiti � M. Tini �G. M. Carpaneto
Department of Sciences, University Roma Tre, Viale G. Marconi
446, 00146 Rome, RM, Italy
e-mail: [email protected]
P. F. Donzelli
e-mail: [email protected]
F. Romiti
e-mail: [email protected]
M. Tini
e-mail: [email protected]
G. M. Carpaneto
e-mail: [email protected]
P. F. Donzelli
Coordinamento Territoriale per l’Ambiente di Verbania per il
Parco Nazionale della Val Grande, Corpo Forestale dello Stato,
Piazza Pretorio 1, 28805 Vogogna, VB, Italy
G. P. Svensson
Department of Biology, Lund University, Ecology Building,
Solvegatan 37, 223 62 Lund, Sweden
e-mail: [email protected]
123
J Insect Conserv
DOI 10.1007/s10841-014-9618-8

and survival probability, has become one of the most
important issues in conservation biology and landscape
management (Clobert et al. 2001). Biologists have long
known that the smaller the population, the more susceptible
it is to extinction (Shaffer 1981). Consequently, one of the
primary roles of a wildlife monitoring program should be
to track the current status of populations with high accu-
racy so that appropriate management actions can be taken
when substantial changes in population size are detected
(MacKenzie and Nichols 2004).
If population abundance is defined as the number of all
individuals of the same species occupying a particular space
at a particular time (Krebs 2001), including all stages of the
life cycle, then it is very difficult to calculate this parameter
for many holometabolous insects, because adults and larvae
can live in different habitats, with different ecological and
food requirements for periods of time often longer than the
adult stage. Therefore, Tikkamaki and Komonen (2011)
suggested studying only adults when monitoring the status
and extinction risk of insect populations. Moreover, in
species where adults can be easily captured and marked, the
capture-mark-recapture (CMR) approach is suitable for
quantifying population characteristics (Amstrup et al.
2005). During the last decade, several CMR studies have
been performed on large threatened beetles to estimate
population size using various methods for capturing adults,
including odour-baited traps and visual surveys (Ranius
2001; Larsson and Svensson 2009; Tikkamaki and Komo-
nen 2011; Drag et al. 2011; Chiari et al. 2013).
Saproxylic invertebrates, i.e. organisms dependent on
fungal decay of wood in living or dead trees, or on other
saproxylic organisms during at least some part of their life
cycle (Speight 1989; Alexander 2008), have been identified
as one of the most threatened guilds of the European fauna
(Speight 1989; Berg et al. 1994; Nieto and Alexander
2010). Their decline is mostly due to intensive commercial
forestry and changed agricultural management practices,
which have decreased the amount and quality of dead wood
(Davies et al. 2008). The European stag beetle, Lucanus
cervus (Linnaeus, 1758) (Coleoptera: Lucanidae), is a
saproxylic beetle whose larva develops underground and
feeds on decaying wood of stumps and roots from a wide
range of broad-leaved trees and shrubs (Percy et al. 2000).
This species, listed in the IUCN Red List of Threatened
Species as ‘‘near threatened’’, in the EU Habitats Directive
as a priority species of community interest (Appendix II)
(Nieto and Alexander 2010) and in the Bern Convention
(Appendix III), is indubitably the most charismatic and
popular saproxylic insect in Europe. The stag beetle was
also mentioned as one of the most emblematic flagship
species for biological conservation in Europe and a focal
species for the conservation of suitable habitats for sapr-
oxylic beetles (Thomaes et al. 2008). Despite that, it is
thought to be declining across much of its European range
(Harvey et al. 2011a) and no CMR study has been per-
formed previously to analyse population parameters of this
species.
The topic of how to capture stag beetles to gain infor-
mation about population parameters has been discussed for
decades. Many investigators have studied the attraction of
this beetle to various odour lures, either in the laboratory or
under natural and semi-natural conditions (woodlands and
urban gardens, respectively) obtaining various levels of
behavioural response (Brustel and Clary 2000; Chapman
et al. 2002; Krenn et al. 2002; Fremlin and Hendriks 2011;
Jansson 2011; Harvey et al. 2011b; Vrezec and Kapla 2007;
Vrezec et al. 2006, 2007). For example, promising data on
short-range attraction to several odour sources under labo-
ratory conditions did not translate into large captures of
adults in the field when such odours were used in aerial
flight interception traps (AFIT) or pitfall traps (PT) (Harvey
et al. 2011b). Such contradictory results could be an effect
of low density of beetles in the study area, or low level of
long-range attraction to the odour sources applied. Ideally,
the use of feeding resources and attractive compounds as
lures should be compared with other methods for monitor-
ing beetles as well with as available elements present in
nature in a certain area, in order to reach valid conclusions
concerning the reliability of the potential attractants, and to
assess an effective method for monitoring adults of L.
cervus across its entire distributional range.
In this study, a CMR approach on L. cervus was per-
formed in northern Italy, testing different capture methods
and odour lures to increase the current knowledge of the
threat status of the species and survival probability of
individual beetles in that region, and to improve conser-
vation programmes through the optimization of field effort
and costs for population monitoring. In particular, four
specific questions were addressed: (1) What is the best
method for capturing stag beetles in nature and monitoring
populations among the ones tested in this study? (2) Is
CMR an applicable method to evaluate stag beetle popu-
lation abundance? (3) Does the body size (as a function of
sex, body weight and elytron length) influence the survival
probability of the individuals? (4) Is felt-tip pen marking a
long lasting method for coding individual beetles to ensure
proper identification in case of subsequent recapture?
Materials and methods
Study area and field work
The study area was an old-growth chestnut (Castanea sati-
va) woodland situated in Monterosso (UTM Zone 32 T,
WGS84, 463232 5088073, between 329 and 622 m a.s.l.), a
J Insect Conserv
123

hill located near Verbania, Piedmont (northern Italy).
Monterosso is a woodland area that includes some human
settlements, i.e. a small village (Cavandone) and scattered
country houses. The woodland is managed as a stand of high
chestnut trees of different ages, regrown over an ancient
coppice; tree density is variable with an open canopy near
the tracks and in clearings. Deadwood is abundant and
represented by stumps, fallen logs and small wood debris.
Adults of L. cervus were sampled using both traps and
visual inspection of dirt tracks and clearings (visual
encounter surveys, hereby VES like in Chiari et al. 2013).
The trap types were two: AFIT and PT, the same as in
Harvey et al. (2011b). The AFIT were suspended from tree
branches at 2–4 m height, and the PT were buried with the
opening at soil level, covered with a wooden roof to pre-
vent predator attack (Fig. 1). In 2012, 35 trap replicates
were set along three 3 m wide dirt tracks (Fig. 2), and from
28th May to 27th July the replicates were checked every
second day, from 15:00 to 22:30. Each replicate contained
one AFIT and one PT, both baited with the same potential
attractant, and one unbaited trap of each type, serving as
control. Therefore 70 baited traps were set (7 AFIT and 7
PT, each type with 5 potential attractants) and 70 control
traps (one for each baited trap), for a total of 140 traps. The
traps were set 2–10 m apart (Harvey et al. 2011b; Svensson
and Larsson 2008) (Fig. 1). Ginger root, cherry juice,
maple syrup, mango juice and red wine were tested as
potential attractants, chosen among those used by Harvey
et al. (2011b). For solid and liquid substances, 20 g or
20 ml samples were used respectively (Harvey et al.
2011b), and the lures were replaced every two checks (i.e.
5 days). To avoid position effects, the relative positions of
AFITs within replicates were exchanged every two checks,
simultaneously with the replacing of lures, whereas for PTs
only the roofs, with the annexed lure holders, were
exchanged (Svensson et al. 2011; Harvey et al. 2011b).
Each replicate was separated by at least 100 m to guarantee
the spatial independence between presumed different lure
effects (Svensson et al. 2011). The maximum distance
among replicates was about 2500 m, which is shorter than
the maximum dispersal distance expected for males (Rink
and Sinsch 2007). The sequence of potential attractants in
each set of five replicates was randomly chosen (without
replacement), repeating the first extraction if the potential
attractant was the same of the last of the previous set,
avoiding two consecutive replicates with the same attrac-
tant in order to make the sampling more heterogeneous
across the landscape (Fig. 2). Each trap was marked and
geo-referenced using a Garmin GPS (MAP 60 CSX). The
VES were carried out by two operators who walked along
both sides of the dirt tracks, during trap checking, and
crossing two clearings (Fig. 2). On the whole, the pathway
measured 8.5 km. Stag beetles observed during the VES
were captured by hand when possible, i.e. when the beetles
were on the ground, trunks or rocks. An entomological net
(http://www.insectnet.eu/), consisting of a circular frame
(50 cm diameter) and a telescopic handle (up to 199 cm),
was used to catch stag beetles in flight. In addition, all stag
beetle body fragments, mostly remains of predation
(Campanaro et al. 2010), were collected. The order in
which operators surveyed the three dirt tracks (e.g. A, B
and C) was systematically changed at each survey,
repeating the permutation of the possible combinations of
the dirt tracks (e.g. ABC, ACB, BCA, BAC, CAB, CBA,
and so on) until the end of the study period.
Upon first capture, each stag beetle was weighed, mea-
sured and then marked with an individual identification
code, following a double procedure: (1) by an indelible
felt-tip pen (Uni Paint Marker, medium-fine paint marker
2.2–2.8 mm line ø, in red and white colours) on the ventral
sclerites of head and pronotum (Campanaro et al. 2011);
(2) by a small drill (Dremel Lithium Cordless 8000JE) on
both elytra (Chiari et al. 2013). The double marking pro-
cedure of each beetle increased the probability to identify
the fragments of previously captured individuals, and was
also carried out to test the durability of the felt-tip pen
mark. If a recaptured beetle showed only the mark on the
elytra it was assumed that the felt-tip pen mark on the head
and/or pronotum had been lost. Finally, the estimate of
marking loss can be used to correct population estimates
(Sutherland 2006). The body mass was measured using a
portable balance (PESOLA AG, Switzerland) (precision
10 mg). The length of the elytral suture (elytron length)
was measured from the base of the scutellum to the apex of
the elytra, in the midline, with a calliper rule (precision
0.05 mm). The elytron length was preferred to the most
common body length since total body size (from clypeus to
pygidium) is not a very precise measure of size due to the
mobility of segments (Juliano 1986). The sex of beetles
was determined by the shape of the mandibles, which
shows a strong sexual dimorphism in this species (Fran-
ciscolo 1997). After the marking procedure, and successive
captures, beetles were released at the place of capture. No
beetle appeared to be injured by the marking procedure and
no discernible leak of haemolymph was observed.
Data analysis
Capture data, efficiency of monitoring methods and body
measurements
Differences between male and female captures including
all three monitoring methods were tested by the Yates
corrected v2 test, and differences in number of captures
(both sexes) between odour-baited and unbaited traps
(AFIT and PT combined) were tested by the Fisher’s exact
J Insect Conserv
123

test. In addition, the Kruskal–Wallis ANOVA was per-
formed to compare the total number of captures between
monitoring methods (male and female data pooled). The
Shapiro test was used to test whether body measurements
data were normally distributed. The assumption of equal
variances for data on body mass and elytron length in both
males and females was verified by the Brown-Forsythe test,
and subsequent analysis of sex-related body size values
was performed using the Mann–Whitney U test. The
Pearson correlation coefficient was used to analyse the
relation between body mass and elytron length for both
sexes of beetles. All analyses were performed using
STATISTICA 7.0 (StatSoft Inc.), with a significance level
of 0.05 to reject the null hypothesis.
Population size and survival estimates
The mark-recapture data were analysed in order to inves-
tigate some aspects of the demography, size and survival
probability of the studied stag beetle population. Estimates
were calculated using information only from VES captures.
Overall population size and survival probability esti-
mates were performed for both sexes separately. The Jolly-
Seber method (POPAN parameterization) was used to
estimate the total population size, and the Cormack-Jolly-
Seber method (Jolly 1965; Southwood 1978) was used to
estimate and model the survival probability. Both methods
are suitable for analyses of open populations, and they
allow estimations of three parameters: (1) the survival
Fig. 1 Detail of aerial flight
interception trap in place, with
lure in the upper chamber, and
diagram representing one of the
35 trap replicates. Flowerpots
were used as pitfall traps, buried
with the opening at soil level
with lure chamber attached in
the middle of the wooden roof
Fig. 2 Overview of the 35 trap replicates located along three dirt
tracks (black lines), which cross the woodlands where two operators
carried out the visual encounter surveys of Lucanus cervus. Black
symbols correspond to the use of different potential attractants (circle
cherry juice, square ginger, triangle mango juice, pentagon maple
syrup, hexagon red wine). Open circles around the black symbols
indicate the position of traps in two clearings
J Insect Conserv
123
probability (/), (2) the capture probability (p), (3) the total
population size (NT). The first two parameters can be
constant (.), or can respond over time in a linear manner (t).
The survival probability was modelled in two ways: (1)
assuming that the parameter is a function of body mass
(BM) or of elytron length (EL) of a beetle at the time it was
first marked and released; (2) assuming that the parameter
is maximized for beetles having a mean body mass
(BM,BM^2) or a mean elytron length (EL,EL^2). These
analyses were performed using the program MARK (White
and Burnham 1999). For each model, differing in param-
eterization, MARK computed the AICc, a measure of
goodness of fit of the model combining likelihood and
complexity (Burnham and Anderson 2002). The model
with the highest support was assumed to be the one with
the lowest value of AICc (Burnham and Anderson 2002). A
model is usually considered plausible if DAICc \ 2
(Burnham and Anderson 2002). Finally, the coefficient of
variation (CV) was calculated as the standard error divided
by the number of individuals estimated and indicates the
precision of the population size estimate.
Results
Capture data
In total, 153 captures of 136 individuals (111 males, 25
females) of L. cervus were observed (Table 1). Of 17
recaptures, 14 were carried out with the same method and
lure and in the same replicate (or during VES), whereas
two took place with a different method and lure, and only
one with the same method but with a different lure, all in
the same replicate. The male captures outnumbered female
captures (v12 = 62.771, p \ 0.01). All recaptured beetles
were recaptured only once, with a recapture rate of 13 %
(15/111) for males and 8 % (2/25) for females. The mean
time between recaptures was 5.4 days for males and
6.0 days for females, with a maximum time period of
14 days for males and 8 days for females between cap-
tures. A low proportion 3 % (4/136) of the marked beetles
(all males) were found dead, all during VES. Most of the
captures were performed between the middle of June and
the middle of July, with a peak around 25th June (Fig. 3).
Monitoring methods
There was a highly significant difference in the number of
L. cervus captured with the different monitoring methods
(Kruskal–Wallis ANOVA, df = 12, N = 175, H =
27.707, p = 0.006). The highest rank sum (best perfor-
mance) was reached by VES (Sum of Ranks = 3278.500),
discriminating this method as by far the best in capturing
this species (Fig. 3). In contrast, there was no difference in
captures between odour-baited and unbaited traps for any of
the two trapping methods (Fisher’s exact test, p = 1.000).
Body measurements
The elytron was significantly longer in males (range:
17.16–29.25 mm) than females (range: 17.89–24.16 mm)
(median: males, 22.48 mm; females, 21.25 mm; U = 905.0,
p = 0.007). In contrast, there was no significant difference
in the median body mass between males (range:
0.80–4.50 g) and females (range: 1.30–3.20 g) (median:
males, 2.20 g; females, 2.10 g; U = 1241.0, p = 0.410). In
both sexes there was a highly significant correlation between
elytron length and body mass (Pearson Correlation Coeffi-
cient: females, r2 = 0.617, rp = 0.785, p = 0.000; males,
r2 = 0.876, rp = 0.936; p = 0.000) (Fig. 4).
Population size and survival estimates
The Jolly-Seber (POPAN parameterization) estimates of
population size for males and females were 2207 (NT:
confidence interval 1283–3132; CV = 0.21) and 747 indi-
viduals (NT: confidence interval 208–1284; CV = 0.37),
respectively.
The best-fitting Cormak-Jolly-Seber models for male
survival probability in relation to body mass resulted in two
plausible models (i.e. DAICc \ 2) (Table 2). In the first
model the male survival probability was constant in time (w/
(.)p(.) = 0.49), while in the second one it was dependent on
body mass (w/(BM)p(.) = 0.26). Heavier males had a lower
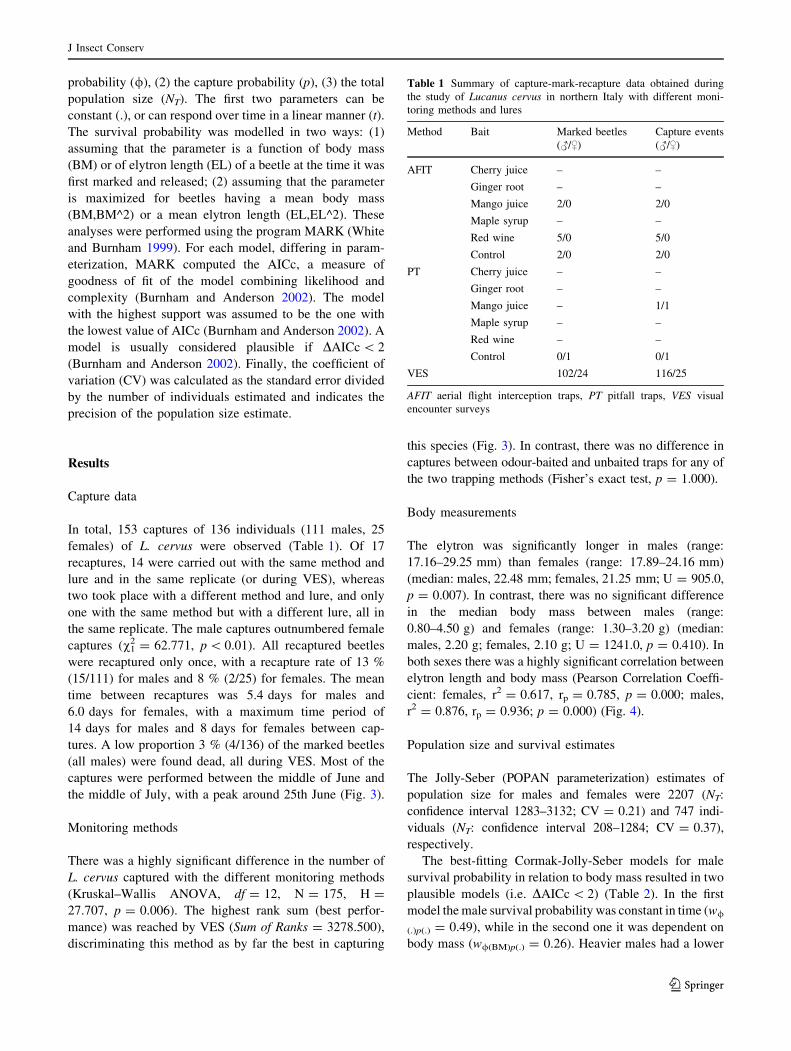
Table 1 Summary of capture-mark-recapture data obtained during
the study of Lucanus cervus in northern Italy with different moni-
toring methods and lures
Method Bait Marked beetles
(#/$)
Capture events
(#/$)
AFIT Cherry juice – –
Ginger root – –
Mango juice 2/0 2/0
Maple syrup – –
Red wine 5/0 5/0
Control 2/0 2/0
PT Cherry juice – –
Ginger root – –
Mango juice – 1/1
Maple syrup – –
Red wine – –
Control 0/1 0/1
VES 102/24 116/25
AFIT aerial flight interception traps, PT pitfall traps, VES visual
encounter surveys
J Insect Conserv
123

survival probability (Fig. 5a). For females three models
were equally good (Table 2). In the first model the female
survival probability was dependent on body mass in a linear
way (w/(BM)p(.) = 0.38) (Fig. 5b), the second model sup-
ported the normalizing selection (w/(BM,BM^2)p(.) = 0.36),
indicating that females with a body mass near the mean value
were favoured, and the third model suggested that female
survival probability was constant over time (w/
(.)p(.) = 0.26).
The best-fitting Cormak-Jolly-Seber models for the sur-
vival probability of males in relation to elytron length
resulted in two plausible models (Table 3). In the first model
the male survival probability was constant in time (w/
(.)p(.) = 0.51), while in the second one it was dependent on
elytron length (w/(EL)p(.) = 0.23). Males with longer elytron
length had a lower survival probability. For females, there
was only one fitting model (Table 3) which strongly sup-
ported the normalizing selection (w/(EL,EL^2)p(.) = 0.65),
indicating that females with the elytron length near the mean
value were favoured.
Methodological issues
During the first four surveys, on average 30 % of the PTs
per day were found broken by wild boar (Sus scrofa), with
no difference among baited and unbaited traps (v12 = 0.02,
p [ 0.05). Therefore, after the fifth survey, PTs were
removed from 26 of the 35 replicates and left only in the
more anthropized zone not reachable by wild boars.
The 95 % (134/141) of VES captures, including dead
individuals, were performed from 20:00 to 22:00, mostly in
two large clearings (132 out of 141) (Fig. 2). The majority
74 % (101/136) of VES captures of living adults were
performed using a net, and captures of living females by
hand or net were significantly lower than those of males
(v12 = 6.140, p = 0.01). All recaptured beetles (17 out of
136) showed the felt-tip pen mark on the ventral sclerites of
head and pronotum.
Discussion
Capture data, monitoring methods and body
measurements
The threatened saproxylic beetle L. cervus was caught more
abundantly than expected for a single sampling season. In a
study conducted in England over 4–5 years, using a similar
set of methods, 55 individuals were captured with odour-
baited AFIT and PT, and 153 individuals were recorded
during road transect surveys (Harvey et al. 2011b), indi-
cating smaller populations in that region. This is probably
due to substantial differences between the habitat investi-
gated in our study and in Harvey et al. (2011b): chestnut
woodland in Italy versus urban gardens in England.
Significantly more males than females were captured
and marked, but the recapture rate was low, independently
Fig. 3 Daily captures events of Lucanus cervus performed by different monitoring methods and lures (AFIT aerial flight interception traps, PT
pitfall traps, VES visual encounter surveys)
Fig. 4 Correlations between elytron length and body mass of adult
males and females stag beetles measured during the capture-mark-
recapture study
J Insect Conserv
123

of sex, as all recaptured beetles were recaptured only once.
This suggests that is not easy to recapture adult stag beetles
and the low recapture rate could make estimates of popu-
lation parameters less precise. This is a major problem, but
estimates of population parameters are absolutely neces-
sary in red-list assessment and population monitoring of
threatened species (Tikkamaki and Komonen 2011). The
biased capture-recapture sex ratio observed for L. cervus in
this study, and in previous ones (Table 4), seems to be an
artefact mostly due to the sampling method used and does
not necessarily reflect a true difference in the number of
males and females present, as documented also in other
beetles and insects since about 30 years (Shibata 1986;
Kuussaari et al. 1996; Stoks 2001; Ranius 2001; Larsson
and Svensson 2009; Chiari et al. 2013). It is still not clear
whether the effective sex ratio of L. cervus is even, and
further studies are needed to clarify the male/female pro-
portion because this parameter could affect the reproduc-
tive rate and the level of inbreeding (Fitz-Earle and Barclay
1989; Werren 1993; Heimpel and Lundgren 2000).
In this study, VES was by far the best method to capture
both males and females of L. cervus, accounting for 93 %
of first captures, and 95 % of all captures. In contrast, just
few catches were observed in odour-baited traps and adults
were not attracted at significant numbers to any of the
odour sources tested. These results are thus in agreement
with those by Harvey et al. (2011b), and confirm that the
few captures in odour-baited traps in England was not due
to low population densities at the study site, but instead a
true lack of attraction of beetles to the test odours applied.
Thus, there is still no efficient odour lure available for
monitoring stag beetle populations. Instead, visual
encounters of living adults, collection of dead individuals
and body fragments (Harvey and Gange 2006; Campanaro
et al. 2010; Campanaro and Bardiani 2012), and road
transect surveys (Harvey et al. 2011b) are currently the
most reliable methods to study population trends in this
species. Moreover, these methods are inexpensive and
easily applicable by volunteers to screen for presence and
abundance of L. cervus.
Fig. 5 Survival probability (/) of the stag beetle, males (a) and
females (b), modeled in relation to body mass (BM) using the
Cormack-Jolly-Seber method in the program MARK. Used model /(BM) p(.). The black line indicates the predicted survival probability,
the grey dotted lines indicate the 95 % confidence intervals
Table 2 Summary of model selection statistics for the Cormak-Jolly-Seber models used to estimate survival probability (/) and capture
probability (p), in realtion to the body mass (BM) and to the mean body mass (BM,BM^2)
Males Females
Model K -2Log (L) DAICc w Model K -2Log (L) DAICc w
/(.) p(.) 2 115.37 0.00 0.49 /(BM) p(.) 2 7.51 0.00 0.38
/(BM) p(.) 3 91.69 1.27 0.26 /(BM,BM^2) p(.) 3 5.01 0.13 0.36
/(t) p(.) 12 95.62 3.20 0.10 /(.) p(.) 2 8.31 0.81 0.26
/(BM,BM^2) p(.) 4 87.88 3.31 0.09 /(t) p(.) 9 4.50 23.28 0.00
/(.) p(t) 14 114.42 4.42 0.05 /(t) p(t) 9 4.50 23.28 0.00
/(t) p(t) 18 114.52 11.53 0.00 /(.) p(t) 14 4.82 67.41 0.00
K represents the number of parameters in the model and -2Log (L) is twice the negative log-likelihood value. Relative AICc (Small sample
Akaike Information Criteria) values and Akaike weight, w, are reported for each model (DAICc represents the difference in AICc value relative
to the top model, Burnham and Anderson 2002). Survival probability (/) and capture probability (p) may be constant (.) or may vary between
sampling occasions (t)
J Insect Conserv
123

As in seven other European countries where stag beetle
body size was studied (Harvey et al. 2011a), also in
northern Italy males were longer than females. Surpris-
ingly, there was no significant difference in body mass
between sexes. This result could be probably due to the
largest range of body mass registered in males (Fig. 4), that
makes median values of males and females very similar
and does not reflect the documented body size dimorphism
of this species (Rink and Sinsch 2007, 2011).
Population size and survival estimates
The population size estimates (obtained from VES captures
only) differed between males and females, and also showed
a variable coefficient of variation. This information sug-
gests that L. cervus in Italy could be more abundant than
other large threatened saproxylic beetles, e.g. Osmoderma
eremita (Chiari et al. 2013). However, due to the suspected
decline across much of its European range (Harvey et al.
2011a), broad monitoring sampling programs are needed
across the distribution range of L. cervus in order to obtain
reliable information about local extinction risks for this
emblematic species.
For both sexes, body size influences the survival prob-
ability. However, while for males the increase in body size
corresponds to a decrease in survival probability, for
females the explanation of how body size may affect sur-
vival probability is not so straightforward because both the
linear and the normalizing selection are equally well sup-
ported. L. cervus exhibits a wide variation in body size,
particularly in males, which is related to mating success,
larval diet and environmental constraints (Harvey and
Gange 2006; Rowe and Ludwig 1991). The reduction in
Table 3 Summary of model selection statistics for the Cormak-Jolly-Seber models used to estimate survival probability (/) and capture
probability (p), in realtion to the elytron length (EL) and to the mean elytron length (EL,EL^2)
Males Females
Model K -2Log (L) DAICc w Model K -2Log (L) DAICc w
/(.) p(.) 2 115.37 0.00 0.51 /(EL,EL^2) p(.) 3 3.02 0.00 0.65
/(EL) p(.) 3 114.826 1.57 0.23 /(EL) p(.) 2 8.27 2.61 0.18
/(t) p(.) 12 95.62 3.20 0.10 /(.) p(.) 2 8.31 2.66 0.17
/(EL,EL^2) p(.) 4 114.48 3.37 0.09 /(t) p(.) 9 4.50 25.13 0.00
/(.) p(t) 14 91.69 4.42 0.06 /(t) p(t) 9 4.50 25.13 0.00
/(t) p(t) 18 87.88 11.53 0.00 /(.) p(t) 14 4.82 69.26 0.00
K, -2Log (L), DAICc and are presented in Table 2. Survival probability (/) and capture probability (p) may be constant (.) or may vary between
sampling occasions (t)
Table 4 Comparison of six studies on the Lucanus cervus across Europe
Harvey and
Gange (2006)
(OCC)
Rink and
Sinsch (2007)
(RAD)
Hawes (2008)
(CMR)
Campanaro
et al. (2011)
(OCC)
Harvey et al. (2011a)
(CMR)
Present study
(CMR)
Country England Germany England Italy England Italy
Study period 1998–2004 2003–2005 2006 2008 2004* or 2005–2008 2012
Study method/s Collection of dead
individuals and
body fragments
Not specified PT
Hand
Net
Collection of dead
individuals and
zbody fragments
AFIT
PT
RTS
AFIT
PT
VES
No. of beetles
Total 1255 56 138 306 55, 153* 10, 126**
Male 569 18 100 290 30, unknown* 9, 102**
Female 686 38 38 16 25, unknown* 1, 24**
Sex ratio 1:1 Female-biased Male-biased Male-biased 1:1, female-biased* Male-biased
For Harvey et al. (2011a) and for the present study it was possible to distinguish between beetles captured with different methods (* = by RTS;
** = by VES; no specifications for beetles captured by AFIT and PT
CMR capture-mark-recapture, OCC occurrence, RAD radiotelemetry; AFIT air flight interception traps, PT pit fall traps, RTS road transect
surveys, VES visual encounter surveys
J Insect Conserv
123

survival probability of larger males could be due to an
increase in predation rate, mostly inflicted by corvids,
which hunt on sight and are the most common predators of
L. cervus adults across Europe (Harvey et al. 2011a), and to
higher metabolic costs due to the larger dimensions and
related behaviours. In many species of insects that have a
marked male dimorphism, the different ‘types’ of males
have developed alternative reproductive strategies (Emlen
and Nijhout 2000). In the stag beetle the intensity of
aggressive behaviour of males is highly dependent on their
body size, and larger males are generally more aggressive
and prone to fight (Lagarde et al. 2005). Moreover, Har-
dersen et al. (2011) collected remnants of the largest males
mostly at the beginning of the season (mid May) and the
average size of collected males thereafter decreased, sug-
gesting that the threshold in body size, which determines
the morph expression of biggest males with large mandi-
bles, shifts along the season. In the female, body size may
contribute to fecundity, with larger females producing
more eggs (Harvey et al. 2011a), but Harvey and Gange
(2006) reported that the ratio between male and female size
is a limiting factor, which influences the mating success in
laboratory tests. It is not clear how the fitness regulates the
body size of L. cervus individuals with respect to a geno-
type or to a phenotype, both at larval and adult stages, and
future research should aim at gaining a quantitative
understanding of this phenomenon.
Methodological issues
The use of PT is a potentially dangerous method for cap-
turing stag beetles in woodlands because the wooden roof
is not sufficient to prevent predator attack and the wild boar
(S. scrofa) destroys these traps, irrespectively if they are
baited with odour lures or not. The wild boar is reported as
one of the major predators of L. cervus larvae (Harvey et al.
2011a), and by using PT it could became a predator also of
adult beetles and the method may thus influence the sur-
vival probability of populations. For this reason most of the
PT were removed from the experiment and left only in the
most anthropized area, where wild boars were not present.
In order to prevent an increased predation risk and mor-
tality rate of adult stag beetles, PT should not be used in
areas where there is a high density of potential predators
such as wild boar, badger (Meles meles), foxes (Vulpes
vulpes) and hedgehog (Erinaceus sp.).
Most of the captures were performed at or just after
sunset, from 20:00 to 22:00, and in two clearings, identified
as the places where beetles were more active in flight. This
result supports the statements of a stag beetle flight peak
activity at dusk (Hawes 2008) and of the ‘hot spots’ phe-
nomenon at small scale (Pratt 2000) with an aggregated
distribution of occurrence at all spatial scales (Harvey et al.
2011a). However, it still remains uncertain if the ‘hot
spots’ phenomenon is due to a restricted availability of
suitable habitat and/or to aggregation of individuals asso-
ciated with mating.
Captures of males were higher than those of females
regardless of sex-related behaviour, such as flight in the
male and seeking behaviour on the ground for the female
(Harvey and Gange 2006; Rink and Sinsch 2011). No
female was captured by AFIT but surprisingly almost the
same numbers of female captures were performed by hand
(12 out of 27) or by net (13 out of 27). In any case, for a
CMR study it is necessary to use a net in order to capture
flying beetles, which are the most frequent adults observed
during the season.
Applying individual identification codes with an indel-
ible felt-tip pen was a reliable method to mark adults of L.
cervus, because such codes were found on all recaptured
beetles, as confirmed by the independent drill marks.
Therefore, the less costly and invasive tip pens may be
considered the most favourable method for mark-recap-
turing stag beetles in monitoring programs.
Conclusions and implications for conservation
Due to the threat status of the stag beetle, monitoring
programs to assess its presence and population sizes across
Europe are urgently needed. The present paper was con-
ceived in a poor status of knowledge about the phenology
of the stag beetle in Italy, still based on anecdotal infor-
mation. For this reason, we planned an intensive fieldwork
to identify the time interval when the species is easy to
detect in nature and to prevent the loss of information that
could bias an experiment. Our research was mostly aimed
to verify the usefulness of baited traps (a costly and time-
consuming method) in comparison with simple visual
surveys. In a previous study (Harvey et al. 2011b), traps
baited with either ginger or mango had a significant
attraction effect on both sexes of L. cervus when tested in
short-range laboratory experiments, but did not attract a
significant number of adults when applied in the field. In
the present study, the attractiveness of all the substances
used was very low and not useful for the stag beetle pop-
ulation monitoring. Therefore, until now the use of traps
baited with odour attractants have not helped in sampling
efforts to gather quantitative information on L. cervus
population demography. In addition, PT may expose the
stag beetles to predation by mammals, mainly wild boars.
The road transect surveys can not be a universal method,
because outside England the stag beetle is mostly present in
woodland areas (Harvey et al. 2011a). Therefore, the col-
lection at dusk of either living adults or their remnants by
VES (e.g. walking along transects or stationing in flying
sites) seems to be the most cost-efficient and effective
J Insect Conserv
123
Standard
Evidenziato
Standard
Evidenziato
Standard
Evidenziato
Standard
Evidenziato
Standard
Evidenziato
Standard
Evidenziato

method (i.e. optimal, sensu Chiari et al. 2013) to monitor
the presence and/or abundance of this species across its
distributional range. In order to carry out a CMR study,
collectors should have an entomological net to capture
flying adults and surveys should be carry out between
20:00 and 22:00. Owing to the documented presence of
‘hot spots’, i.e. small areas where a lot of individuals are
aggregated, many walking transects should be carried out,
repeatedly during the season, in order to limit the proba-
bility of reporting false absence of the stag beetle or
inaccurate population parameters estimates, as this species
is usually detected in low numbers, and is only active
during a short period of warm nights in June and July
(Smith 2003; Harvey et al. 2011a). Due to its role as
flagship and focal species, the monitoring of L. cervus
deserves high priority for biodiversity conservation in
Europe, because if measures are taken to protect this beetle,
many other saproxylic organisms in the same habitat will
be conserved. Precautionary actions are needed to reduce
the rate of predation by corvids, such as the hooded crow
(Corvus corone cornix) whose population density is
increasing with woodland fragmentation (Andren 1992;
Sodhi and Ehrlich 2010), and to reduce the risk of local
extinctions of small stag beetles populations. Therefore,
forest management should be oriented to avoid woodland
fragmentation, control hooded crow populations and
increase the amount and quality of stumps and dead wood
where stag beetle populations are present.
Acknowledgments This study was partially supported by the pro-
ject ‘‘Studio ecologico preliminare su specie saproxiliche della
Direttiva Habitat, nel Parco Nazionale della Val Grande’’ financed by
the ‘‘Parco Nazionale della Val Grande’’. We thank the staff of the
park, in particular Tullio Bagnati (Director) and Cristina Movalli,
(Promoter of Nature Conservation and Management), who supported
the project. A special thanks is given to the agent of the State Forestry
Corps, Simone Torniai, for field assistance and precious help. The
work of S.C. is supported by the VABAS project. G.P.S. is supported
by the Swedish Research Council Formas.
References
Alexander KNA (2008) Tree biology and saproxylic Coleoptera:
issues of definitions and conservation language. Rev Ecol (Terre
Vie) 10:9–13
Amstrup SC, McDonald TL, Manly BFJ (2005) Handbook of capture-
recapture analysis. Princeton University Press, Princeton
Andren H (1992) Corvid density and nest predation in relation to
forest fragmentation: a landscape perspective. Ecology
73:794–804
Andrewartha HG, Birch LC (1954) The distribution and abundance of
animals. Univ. of Chicago Press, Chicago, Illinois
Berg A, Ehnstrom B, Gustafsson L, Hallingback T, Jonsell M,
Weslien J (1994) Threatened plant, animal, and fungus species in
Swedish forests—distribution and habitat associations. Conserv
Biol 8:718–731
Brustel H, Clary J (2000) ‘Oh, cette Gresigne!’, Acquisitions
remarquables pour cette foret et le sud-ouest de la France:
donnees faunistiques et perspectives de conservation (Coleop-
tera), (premier supplement au catalogue de Jean Rabil, 1992,
1995). Bull Soc Entomol Fr 105:357–374
Burnham KP, Anderson DR (2002) Model selection and multimodel
inference: a practical information—theoretic approach. Springer,
Berlin
Campanaro A, Bardiani M (2012) Walk transects for monitoring of
Lucanus cervus in an Italian lowland forest. Saproxylic beetles in
Europe: monitoring, biology and conservation. Studia Forestalia
Slovenica, pp 17–22
Campanaro A, Hardersen S, Toni I, Grasso DA (2010) Monitoring of
Lucanus cervus by means of Remains of Predation (Coleoptera:
Lucanidae). Entomol Gen 33:79–89
Campanaro A, Bardiani M, Spada L, Carnevali L, Montalto F,
Antonini G, Mason F, Audisio P (2011) Linee guida per il
monitoraggio e la conservazione dell’entomofauna saproxilica.
Quaderni Conservazione Habitat, 6. Cierre Grafica, Verona,
p 8. ? CD-ROM
Chapman JW, Birkett MA, Pickett JA, Woodcock CM (2002)
Chemical ecology and conservation of the stag beetle, Lucanus
cervus. Chemical communication: from hormones to semio-
chemicals. Comp Biochem Physiol A 132:63–70
Chiari S, Zauli A, Mazziotta A, Luiselli L, Audisio P, Carpaneto GM
(2013) Surveying an endangered saproxylic beetle, Osmoderma
eremita, in Mediterranean woodlands: a comparison between
different capture methods. J Insect Conserv 17:171–181
Clobert J, Danchin E, Dhondt AA, Nichols JD (2001) Dispersal.
Oxford University Press, Oxford
Davies ZG, Tyler C, Stewart GB, Pullin AS (2008) Are current
management recommendations for saproxylic invertebrates
effective? A systematic review. Biodivers Conserv 17:209–234
Drag L, Hauck D, Pokluda P, Zimmermann K, Cizek L (2011)
Demography and dispersal ability of a threatened saproxylic
beetle: a mark-recapture study of the rosalia longicorn (Rosalia
alpina). PLoSONE 6:e21345. doi:10.1371/journal.pone.0021345
Emlen DJ, Nijhout F (2000) The development and evolution of
exaggerated morphologies in insects. Rev Ecol (Terre Vie)
45:661–708
Fitz-Earle M, Barclay HJ (1989) Is there an optimal sex ratio for
insect mass rearing? Ecol Model 45:205–220
Franciscolo ME (1997) Coleoptera Lucanidae. Collana ‘‘Fauna
d’Italia’’,Volume XXXV, Calderini, Bologna
Fremlin M, Hendriks P (2011) Sugaring for stag beetles—different
feeding strategies of Lucanus cervus and Dorcus parallelipipe-
dus. Bull Amat Entomol Soc 70:57–67
Hardersen S, Macagno ALM, Sacchi R, Toni I (2011) Seasonal
constraints on the mandible allometry of Lucanus cervus
(Coleoptera: Lucanidae). Eur J Entomol 108:461–468
Harvey DJ, Gange AC (2006) Size variation and mating success in the
stag beetle, Lucanus cervus L. Physiol Entomol 31:218–226
Harvey DJ, Gange AC, Hawes CJ, Rink M (2011a) Bionomics and
distribution of the stag beetle, Lucanus cervus (L.) across
Europe. Insect Conserv Diver 4:23–38
Harvey DJ, Hawes CJ, Gange AC, Finch P, Chesmore D, Farr I
(2011b) Development of non-invasive monitoring for larvae and
adults of the stag beetle, Lucanus cervus. Insect Conserv Diver
4:4–14
Hawes CJ (2008) The stag beetle Lucanus cervus (Linnaeus, 1758)(Coleoptera: Lucanidae): a mark-release-recapture study under-
taken in one United Kingdom residential garden. Rev Ecol
(Terre Vie) 63:131–138
Heimpel GE, Lundgren JG (2000) Sex ratios of commercially reared
biological control agents. Biol Control 19:77–93
J Insect Conserv
123

Jansson N (2011) Attraction of stag beetles with artificial sap in
Sweden. Bull Amat Entomol Soc 70:51–56
Jolly GM (1965) Explicit estimates from capture-recapture data with
both death and immigration-stochastic model. Biometrika
52:225–247
Juliano SA (1986) Food limitation of reproduction and survival for
populations of Brachinus (Coleoptera: Carabidae). Ecology
67:1036–1045
Krebs CJ (2001) Ecology: the experimental analysis of distribution
and abundance, 5th edn. Benjamin Cummings, San Francisco,
California
Krenn HW, Pernstich A, Messner T, Hannappel U, Paulus HF (2002)
Kirschen als Nahrung des mannlichen Hirschkafers, Lucanus
cervus (Linnaeus 1758) (Lucanidae: Coleoptera). Entomologi-
sche Zeitschrift Stuttgart 112:165–170
Kuussaari M, Nieminen M, Hanski I (1996) An experimental study of
migration in the Glanville fritillary butterfly Melitaea cinxia.
J Anim Ecol 65:791–801
Lagarde F, Corbin J, Goujon C, Poisbleau M (2005) Polymorphisme
et performances au combat chez les males de Lucane cerf-volant
(Lucanus cervus). Rev Ecol (Terre Vie) 60:127–137
Larsson MC, Svensson GP (2009) Pheromone monitoring of rare and
threatened insects: exploiting a pheromone-kairomone system to
estimate prey and predator abundance. Conserv Biol 23:
1516–1525
MacKenzie DI, Nichols JD (2004) Occupancy as a surrogate for
abundance estimation. Anim Biodivers Conserv 27:461–467
Nieto A, Alexander KNA (2010) European red list of saproxylic
beetles. Publications Office of the European Union, Luxem-
bourg, Belgium
Percy C, Bassford G, Keeble V (2000) Findings of the 1998 national
stag beetle survey. People’s Trust for Endangered Species,
London
Pratt CR (2000) An investigation into the status history of the stag
beetle Lucanus cervus Linneus (Lucanidae) in Sussex. Coleopt
9:75–90
Ranius T (2001) Constancy and asynchrony of Osmoderma eremita
populations in tree hollows. Oecologia 126:208–215
Rink M, Sinsch U (2007) Radio-telemetric monitoring of dispersing
stag beetles: implications for conservation. J Zool (London)
272:235–243
Rink M, Sinsch U (2011) Warm summers negatively affect duration
of activity period and condition of adult stag beetles (Lucanus
cervus). Insect Conserv Diver 4:15–22
Rowe L, Ludwig D (1991) Size and timing of metamorphosis in
complex life histories, time constraints and variation. Ecology
72:413–427
Shaffer ML (1981) Minimum population size for species conserva-
tion. Bioscience 31:131–134
Shibata E (1986) Adult populations of the sugi bark borer, Semanotus
japonicus Lacordaire (Coleoptera: Cerambycidae), in Japanese
cedar stands: population parameters, dispersal, and spatial
distribution. Res Popul Ecol (Kyoto) 28:253–266
Smith MN (2003) National Stag Beetle Survey 2002. People’s Trust
for Endangered Species, London
Sodhi NS, Ehrlich PR (2010) Conservation biology for all. Oxford
University Press, New York
Southwood TRE (1978) Ecological methods with particular reference
to the study of insect populations. Chapman and Hall, London
Speight MCD (1989) Saproxylic invertebrates and their conservation.
Council of Europe, Strasbourg
Stoks R (2001) Male-biased sex ratios in mature damselfly popula-
tions: real or artefact? Ecol Entomol 26:181–187
Sutherland WJ (2006) Ecological census techniques. Cambridge
University Press, New York
Svensson GP, Larsson MC (2008) Enantiomeric specificity in a
pheromone-kairomone system of two threatened saproxylic
beetles, Osmoderma eremita and Elater ferrugineus. J Chem
Ecol 34:189–197
Svensson GP, Sahlin U, Brage B, Larsson MC (2011) Should I stay or
should I go? Modelling dispersal distances in a threatened
saproxylic beetle, Osmoderma eremita, based on pheromone
capture and radio telemetry. Biodivers Conserv 20:2883–2902
Thomaes A, Kervyn T, Maes D (2008) Applying species distribution
modelling for the conservation of the threatened saproxylic Stag
Beetle (Lucanus cervus). Biol Conserv 141:1400–1410
Tikkamaki T, Komonen T (2011) Estimating population character-
istics of two saproxylic beetles: a mark-recapture approach. J Ins
Conserv 15:401–408
Vrezec A, Kapla A (2007) Kvantitativno vzorcenje hroscev (Cole-
optera) v Sloveniji: referencna studija [Quantitative beetle
(Coleoptera) sampling in Slovenia: the reference study]. Acta
Entomol Sloven 15(2):131–160
Vrezec A, Kapla A, Grobelnik V, Govedic M (2006) Analiza
razsirjenosti in ocena velikosti populacije rogaca (Lucanus
cervus) s predlogom conacije Natura 2000 obmocja Goricko
(SI3000221). Nacionalni institut za biologijo, Ljubljana,
Slovenia
Vrezec A, Polak S, Kapla A, Pirnat A, Grobelnik V, Salamun A
(2007) Monitoring populacij izbranih ciljnih vrst hroscev –
Carabus variolosus, Leptodirus hochenwartii, Lucanus cervus in
Morinus funereus, Rosalia alpina. Nacionalni institut za biolog-
ijo, Ljubljana, Slovenia
Werren JH (1993) The Natural History of Inbreeding and Outbreed-
ing: Theoretical and Empirical Perspectives. University of
Chicago Press, Chicago, Illinois
White GC, Burnham KP (1999) Program MARK: survival estimation
from populations of marked animals. Bird Study 46:120–139
J Insect Conserv
123




















