
Odour and ammonia emissions from intensive pig units in Ireland

ISSN 0327-9383
NOTAS TÉCNICAS - TECHNICAL NOTES
Mastozoología Neotropical / J. Neotrop. Mammal.; 10(1):167-176
©SAREM, 2003
EFICIENCIA DE ATRAYENTES PARA CARNÍVOROS EN BOSQUESYUNGUEÑOS Y PRADERAS ALTOANDINAS EN BOLIVIA
Luis F. Pacheco1, 2, J. Fernando Guerra3 y Boris Ríos-Uzeda3, 4
1 Estación Biológica Tunquini, Instituto de Ecología, Casilla 10077 Correo Central, La Paz, Bolivia.Fax: 591 2 797511. 2 Centro de Estudios en Biología Teórica y Aplicada (BIOTA), Casilla 9641, LaPaz, Bolivia, <[email protected]>. 3 Conservación y Medio Ambiente (CYMA), Héctor Ormachea320, Esq. Calle 4, Obrajes, La Paz, Bolivia. <[email protected]>. 4 Dirección actual: WildlifeConservation Society – Bolivia, Casilla 3-35181 S.M., La Paz, Bolivia
RESUMEN. Comparamos la eficiencia de tres tipos de atrayentes odoríferos para carnívorosen cada uno de cuatro pisos altitudinales del Parque Nacional y Área Natural de ManejoIntegrado Cotapata, Bolivia. Los atrayentes (Canine Call, Cat Passion y una mezcla de WildCat y Canine Call) se probaron por una noche en 6-12 transectos instalados con separaciónde al menos 600 m, cada uno con 6 huelleros de 1 m2 separados 50 m entre sí. La primeranoche de revisión se instalaron los huelleros sin atrayente como control. La tasa de visitasde carnívoros a los huelleros con algún atrayente fue en general más alta que cuando nose usaron; otras especies visitaron los huelleros con atrayente, aunque algunas evidente-mente no lo hicieron atraídas por el olor y otras probablemente evitaron acercarse a loshuelleros con atrayentes para felinos. Concluimos que el atrayente más efectivo fue el CatPassion, el cual podría utilizarse para programas de monitoreo de carnívoros (especialmenteadecuado para Pseudalopex culpaeus, Oreailurus jacobita y Leopardus tigrinus) en estazona. El monitoreo de otras especies podría ser mejor si no se usan atrayentes paracarnívoros.
ABSTRACT. Efficiency of lures for carnivores in Yungas forests and high altitudeprairies in the Bolivian Andes . We compared the efficiency of three types of scent luresfor carnivores in each one of four altitudinal floors within the Cotapata National Park andNatural Area of Integrated Management, Bolivia. The lures (Canine Call, Cat Passion andWild Cat mixed with Canine Call) were tested by one night in 6-12 transects installed withseparation of at least 600 m, each one with 6 track plots of 1 m2 separated 50 m from eachother. To use as control, we did not use any lure during the first night of revision. In general,visitation rates of carnivores were higher with the use of lures than when we did not usethem; other species visited the lured track plots, though some were evidently not attractedby their scent and others probably avoided the track plots with lures for felids. We concludethat the best lure was Cat Passion, which could be used for monitoring programs of carni-vores (specially appropriate for Pseudalopex culpaeus, Oreailurus jacobita and Leopardustigrinus) in this area. Monitoring of other species could be better if lures for carnivores arenot used.
Palabras clave : atrayentes odoríferos, índice de actividad, mamíferos, Yungas, Andes,Hippocamelus, Mazama chunyi, Oreailurus, Leopardus, Pseudalopex.
Key words : scent lures, activity index, mammals, Yungas, Andes, Hippocamelus, Mazamachunyi, Oreailurus, Leopardus, Pseudalopex.
Versión on-line ISSN 1666-0536
Recibido 24 octubre 2001. Aceptación final 27 julio 2002.

168 Mastozoología Neotropical, 2003
INTRODUCCIÓN
La importancia de las estimaciones de abun-dancia de poblaciones en los aspectos teóricosy prácticos de la ecología es por demás reco-nocida (Caughley y Sinclair, 1994; Sargeant etal., 1998; Walker et al., 2000). En áreas pro-tegidas, éstas son particularmente necesariaspara diseñar los programas de protección y/omanejo pertinentes. Las estimaciones de abun-dancia de mamíferos terrestres en áreas prote-gidas de Bolivia son muy escasas (Painter etal., 1995; Cuellar y Noss, 1997; Gómez et al,2001; Ríos Uzeda et al., 2001) y casiinexistentes en áreas protegidas de montaña(Rumiz et al., 1998). En nuestro sitio de estu-dio, el Parque Nacional y Área Natural deManejo Integrado Cotapata (PN ANMICotapata), estamos aún en fase de adecuaciónde metodologías (Ríos-Uzeda, 2001).
Como parte de este esfuerzo, nos propusi-mos evaluar la eficiencia relativa de tres atra-yentes para carnívoros en cuatro pisosaltitudinales del PN ANMI Cotapata. Si bienla literatura reporta el uso de varios atrayentesodoríferos para carnívoros y ha probado sueficiencia, los resultados no son concluyentesy parecen depender de la especie, el ambientey el método de colocar los atrayentes (Morrisonet al., 1981; Harrison, 1997; Travaini et al.,2001). Además, algunos estudios argumentansobre las ventajas de no utilizar atrayentes paraevaluar abundancia de carnívoros usando par-celas de huellas (Engeman et al., 2000;Engeman y Allen, 2001) y sólo un estudio delos disponibles evalúa la eficiencia del uso deatrayentes con relación a no usarlos (Harrison1997), el cual encontró que el uso de atrayen-tes resultaba en mayores tasas de visita de car-nívoros. Sin embargo, un estudio preliminaren nuestro sitio de trabajo sugiere que el usode atrayentes no incrementa las probabilidadesde registrar especies, ni siquiera carnívoros(Ríos-Uzeda, 2001). Por ello escogimos tresatrayentes para carnívoros con base en suge-rencias de cazadores experimentados (com.pers. a A. Taber, Wildlife ConservationSociety) para evaluar su eficiencia para atraercarnívoros y evaluar a la vez el uso de algunode éstos con relación a no utilizarlos. Las pre-
guntas específicas fueron: ¿cuál de los atra-yentes utilizados es el más efectivo? y ¿es me-jor utilizar algún atrayente que ninguno? Almismo tiempo, pretendíamos obtener informa-ción base sobre la abundancia relativa de algu-nas especies de mamíferos terrestres con peso> 1kg, para su potencial uso en programas demonitoreo.
SITIO DE ESTUDIO
El Parque Nacional y Área Natural de ManejoIntegrado Cotapata (PN-ANMI Cotapata) seubica en el departamento de La Paz (Bolivia),con coordenadas referenciales: 68°02' y 16°20'SO, 68°03' y 16°05' NO, 67°43' y 16°10' NEy 67°47' y 16°18' SE. Cubre casi 60.000 hacon un rango altitudinal de 5.600 m a 1.200 msnm (Ribera-Arismendi, 1995). Este gradientealtitudinal se traduce en una gran diversidadde ambientes que fueron separados por Ribe-ra-Arismendi (1995) en cinco pisos altitudina-les, de los cuales se estudiaron cuatro:
Piso Altoandino (4.200 - 4.900 m); cubreterrenos de origen glaciar. La vegetación esdominada por herbáceas de bajo porte de losgéneros Festuca, Stipa, Poa y Calamagrostis.Son menos comunes los arbustos (Baccharis,Satureja, Brachyotum) y árboles (Polylepis) enpequeños manchones. Existen amplias zonasde bofedal, donde predominan las plantas encojín de los géneros Distichia y Plantago. Enlas zonas más rocosas es frecuente la presen-cia de especies de Azorella y Pyncophyllum.La fauna incluye vizcachas de la sierra(Lagidium viscacia), tarujas (Hippocamelusantisensis), zorros (Pseudalopex culpaeus), titio gato andino (Oreailurus jacobita y posible-mente también Oncifelis colocolo), jucumari uoso (Tremarctos ornatus) y puma (Pumaconcolor).
Páramo Yungueño (3.500 a 4.200 m); sibien no incluye el elemento típico de los pára-mos del N de Sudamérica (Espeletia), el usodel término «páramo» está justificado por suscondiciones geográficas, ecológicas y estruc-turales (López, 1998). Su vegetación es predo-minantemente herbácea, aunque los pastizalesson más altos y densos, y se mezclan más conmatorrales bajos que en el piso Altoandino.

NOTAS TÉCNICAS - TECHNICAL NOTES 169
Los géneros más comunes son Stipa, Festuca,Brachyotum, Satureja, Mutisia, Chuquiraga,Baccharis, Calceolaria y Gnaphalium. En lasquebradas son comunes los rodales de Polylepisy Buddleja, así como los matorrales arbustivosde Baccharis. Son abundantes las bromeliáceas(al parecer del género Puya), que son alimentodel jucumari. La fauna del páramo es muyparecida a la del piso Altoandino, aunque lascomunidades de micromamíferos se hacen másricas hacia las zonas más bajas.
Bosque Nublado (2.400 a 3.500 m); se tratade un bosque relativamente bajo (10 - 15 m),con árboles retorcidos y con el dosel típica-mente cubierto de abundantes epífitas. Losemergentes (Podocarpus, Miconia) alcanzanhasta 30 m de altura. Entre los géneros máscomunes están: Weinmania, Juglans,Gaultheria, Cedrela, Alnus, Clusia, Randia yMyrsine. El estrato inferior del bosque estádominado por lianas y bambúes (Chusquea).La fauna incluye al puma, jucumari, cabrilla(Mazama chunyi), tawari (Eira barabara),uchuñari (Nasua nasua), llapa con cola o jaipa(Dinomys branickii), tigresillo (Leoparduspardalis), puercoespín (Coendu bicolor) y elfélido recientemente registrado para BoliviaLeopardus tigrinus (Pacheco et al., 2001).
Bosque Montañoso de Yungas (1.200 a2.400); aquí el dosel del bosque es más alto(15 a 20 m), con emergentes que pasan los 30 m(Ficus, Aniba). Los géneros de árboles máscaracterísticos son: Juglans, Brunellia,Podocarpus, Clusia, Protium, Vismia, Virola,varias lauráceas, Cecropia, Sambucus, Urera,Erythrina, Cedrela, Allophylus, variasmelastomatáceas y rubiáceas, además de loshelechos arbóreos (Ribera-Arismendi, 1995;A.I. Roldán, datos no publicados). La fauna esparecida a la de las regiones más bajas de laAmazonía, con la presencia de especies comoel sari (Dasyprocta variegata), llapa (Agoutipaca), monos (Cebus apella, Ateles chamek,Aotus azarae), gatos (L. wiedii, Herpailurusyaguaroundi, L. tigrinus) y chanchos de mon-te (Pecari tajacu y ocasionalmente Tayassupecari).
La temperatura del área varía grandementecon la altitud, aunque no se tienen registros deuna estación climática. Se presume que la pre-
cipitación alcanza su máximo en el bosquenublado, con unos 4.000 mm anuales (Ribera-Arismendi, 1995).
MÉTODOS
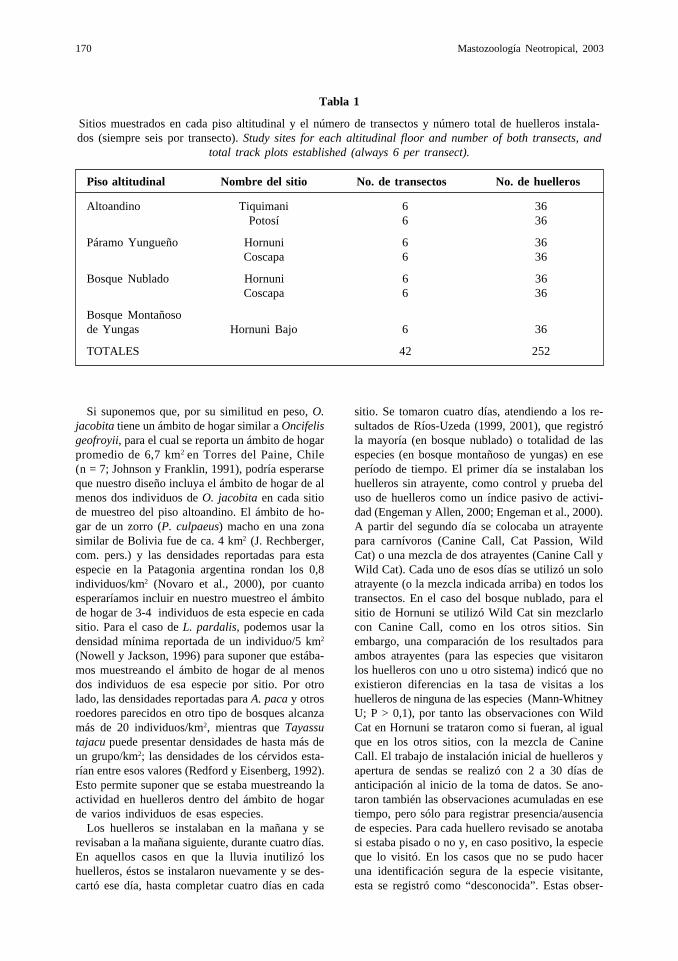
Se ubicaron dos sitios de muestreo en cada uno delos cuatro pisos altitudinales considerados dentrodel PN ANMI Cotapata, excepto en el BosqueMontañoso de Yungas, donde sólo se trabajó en unsitio (Tabla 1). El trabajo de campo se realizó enlos meses de Julio a Agosto de 1999, que corres-ponden a la época seca.
Consideramos las señales registradas en parcelaspara el registro de huellas (huelleros) como índicede actividad para aquellas especies con amplio ran-go de movimientos (carnívoros y chanchos demonte) y como índice de abundancia para aquellascon rangos de movimiento más estrechos (cérvidosy roedores). Los huelleros son el método más eficazpara detectar a los mamíferos en el sitio de estudio(Ríos-Uzeda 2001) y su aplicación es de relativobajo costo, lo cual posibilitaría su utilización enprogramas de conservación de largo plazo.
Dado que nuestro trabajo debía realizarse a pie ypor áreas de fuerte pendiente, la instalación y revi-sión de huelleros en espacios de magnitud suficien-te para evaluar abundancia de la mayoría de loscarnívoros sería prohibitiva. Nuestro diseño inclu-yó, para cada sitio de cada piso altitudinal, seistransectos de 250 m de longitud, separados 600 mentre sí, cada uno con seis huelleros de 1 m2 sepa-rados 50 m entre sí (modificado de Ríos-Uzeda,2001, ver Tabla 1). Los transectos cubrían así unespacio lineal de 3 km, atravesando un área de 9km2, si pensamos en ámbitos de hogar con forma depolígono regular. Si bien nuestros transectos esta-ban muy cercanos unos de otros para considerarlosindependientes para la mayoría de las especies(Roughton y Sweeny, 1982; Sargeant et al., 1998,Novaro et al. 2000), nuestro principal objetivo eraevaluar la eficiencia de los atrayentes y trabajamosbajo el supuesto de que no precisábamos que losindividuos bajo estudio fueran distintos entretransectos. Harrison (1997) reporta que 9 de 12individuos (distintas especies de felinos en cautive-rio) no mostraron diferencias en su respuesta a losatrayentes con el número de pruebas a las que fue-ron sometidos, por lo cual podríamos suponer quelas respuestas de los individuos bajo estudio a dis-tintos atrayentes refleja lo que sucedería de habermuestreado un área mucho mayor (y más indivi-duos por especie). De todas maneras, es útil unexamen del probable número de individuos con elque estuvimos trabajando.
170 Mastozoología Neotropical, 2003
Si suponemos que, por su similitud en peso, O.jacobita tiene un ámbito de hogar similar a Oncifelisgeofroyii, para el cual se reporta un ámbito de hogarpromedio de 6,7 km2 en Torres del Paine, Chile(n = 7; Johnson y Franklin, 1991), podría esperarseque nuestro diseño incluya el ámbito de hogar de almenos dos individuos de O. jacobita en cada sitiode muestreo del piso altoandino. El ámbito de ho-gar de un zorro (P. culpaeus) macho en una zonasimilar de Bolivia fue de ca. 4 km2 (J. Rechberger,com. pers.) y las densidades reportadas para estaespecie en la Patagonia argentina rondan los 0,8individuos/km2 (Novaro et al., 2000), por cuantoesperaríamos incluir en nuestro muestreo el ámbitode hogar de 3-4 individuos de esta especie en cadasitio. Para el caso de L. pardalis, podemos usar ladensidad mínima reportada de un individuo/5 km2
(Nowell y Jackson, 1996) para suponer que estába-mos muestreando el ámbito de hogar de al menosdos individuos de esa especie por sitio. Por otrolado, las densidades reportadas para A. paca y otrosroedores parecidos en otro tipo de bosques alcanzamás de 20 individuos/km2, mientras que Tayassutajacu puede presentar densidades de hasta más deun grupo/km2; las densidades de los cérvidos esta-rían entre esos valores (Redford y Eisenberg, 1992).Esto permite suponer que se estaba muestreando laactividad en huelleros dentro del ámbito de hogarde varios individuos de esas especies.
Los huelleros se instalaban en la mañana y serevisaban a la mañana siguiente, durante cuatro días.En aquellos casos en que la lluvia inutilizó loshuelleros, éstos se instalaron nuevamente y se des-cartó ese día, hasta completar cuatro días en cada
sitio. Se tomaron cuatro días, atendiendo a los re-sultados de Ríos-Uzeda (1999, 2001), que registróla mayoría (en bosque nublado) o totalidad de lasespecies (en bosque montañoso de yungas) en eseperíodo de tiempo. El primer día se instalaban loshuelleros sin atrayente, como control y prueba deluso de huelleros como un índice pasivo de activi-dad (Engeman y Allen, 2000; Engeman et al., 2000).A partir del segundo día se colocaba un atrayentepara carnívoros (Canine Call, Cat Passion, WildCat) o una mezcla de dos atrayentes (Canine Call yWild Cat). Cada uno de esos días se utilizó un soloatrayente (o la mezcla indicada arriba) en todos lostransectos. En el caso del bosque nublado, para elsitio de Hornuni se utilizó Wild Cat sin mezclarlocon Canine Call, como en los otros sitios. Sinembargo, una comparación de los resultados paraambos atrayentes (para las especies que visitaronlos huelleros con uno u otro sistema) indicó que noexistieron diferencias en la tasa de visitas a loshuelleros de ninguna de las especies (Mann-WhitneyU; P > 0,1), por tanto las observaciones con WildCat en Hornuni se trataron como si fueran, al igualque en los otros sitios, con la mezcla de CanineCall. El trabajo de instalación inicial de huelleros yapertura de sendas se realizó con 2 a 30 días deanticipación al inicio de la toma de datos. Se ano-taron también las observaciones acumuladas en esetiempo, pero sólo para registrar presencia/ausenciade especies. Para cada huellero revisado se anotabasi estaba pisado o no y, en caso positivo, la especieque lo visitó. En los casos que no se pudo haceruna identificación segura de la especie visitante,esta se registró como “desconocida”. Estas obser-
Tabla 1
Sitios muestrados en cada piso altitudinal y el número de transectos y número total de huelleros instala-dos (siempre seis por transecto). Study sites for each altitudinal floor and number of both transects, and
total track plots established (always 6 per transect).
Piso altitudinal Nombre del sitio No. de transectos No. de huelleros
Altoandino Tiquimani 6 36Potosí 6 36
Páramo Yungueño Hornuni 6 36Coscapa 6 36
Bosque Nublado Hornuni 6 36Coscapa 6 36
Bosque Montañosode Yungas Hornuni Bajo 6 36
TOTALES 42 252

NOTAS TÉCNICAS - TECHNICAL NOTES 171
vaciones no se analizaron. En todos los casos, conexcepción del piso Altoandino, los muestreos paralos dos sitios de cada piso se realizaron con unintervalo máximo de 10 días. Para el pisoAltoandino, la evaluación en el segundo sitio serealizó un mes después de la evaluación para elprimero. Aceptamos que los muestreos deberíanhaberse realizado simultáneamente para minimizarlos efectos temporales (distinto estado del tiempo,fase lunar o luminosidad nocturna, dependiente dela cobertura de nubes). Sin embargo, dada la dis-tancia entre los sitios de cada piso, al menos elestado del tiempo habría sido de todas maneras muydifícilmente similar en ambos. Suponemos aquí queestos factores no afectaron significativamente nues-tros resultados.
Con la proporción de huelleros con huella portransecto como variable respuesta, se evaluó la efi-ciencia relativa de los atrayentes para cada especiecon una prueba Friedman para medidas repetidas(distintos atrayentes en los mismos transectos; Zar,1999). En este caso, se consideraron 12 transectos(seis de cada sitio) para el análisis, entendiendo quela pregunta era sobre los atrayentes y no sobre lossitios. En caso que una especie sólo haya sido re-gistrada en uno de los sitios, se utilizaron solamen-te las observaciones de los seis transectos corres-pondientes al sitio con presencia de la especie encuestión Además se compararon las tasas de visitapara cada especie a los huelleros con algún cebo yaquellos sin cebo, con una prueba «U» de Mann-Whitney, con corrección por continuidad y empates(Zar, 1999). Todas las comparaciones se hicieronpor especie y se utilizó un a = 0,05. Para aquellasespecies que visitaron menos de tres huelleros nose intentó un análisis estadístico.
RESULTADOS
Se instaló un total de 42 transectos, 12 en cadapiso altitudinal (excepto el bosque Montañosode Yungas con sólo 6 transectos). Un total de252 huelleros, que en cuatro días totalizaron unesfuerzo de 1008 huelleros activos (Tabla 1).
Piso altoandino
Los transectos abarcaron alturas aproxima-das entre 4.200 y 4.600 m snm. Se registró untotal de cinco especies entre ambos sitios. Laespecie que se registró más frecuentemente enlos huelleros fue el zorro (Pseudalopexculpaeus), que llegó a visitar hasta el 67% delos huelleros de un transecto en la zona deTiquimani.
Para el caso del “titi”, no tenemos seguridadde la identidad de este felino (tal vez se tratede Oncifelis colocolo), pues es imposible ase-gurar la identidad de la especie por la huella.Sin embargo, por la gran altura del registro,creemos que debe tratarse de Oreailurusjacobita.
El tipo de atrayente utilizado sólo tuvo unefecto significativo en la tasa de visitas dezorro; la tasa de visitas difirió solamente entreCat Passion y la registrada sin atrayentes. Lastres especies de carnívoros detectados en loshuelleros visitaron éstos más frecuentementecuando se utilizó algún atrayente que sin él.No así las otras especies, cuya tasa de visita alos huelleros fue indistinta con o sin el uso deatrayentes (Tabla 2).
Páramo
Los transectos se instalaron a alturas entre3.400 y 3.800 m snm. Se registraron sólo tresespecies en este piso altitudinal y todas ellasen el sitio de Coscapa. Ningún mamífero fueregistrado por nuestros huelleros en el páramode Hornuni. El zorro fue nuevamente la espe-cie que visitó más frecuentemente los huelleros,con un máximo de 67% de huelleros visitadosen un transecto; sólo visitó los huelleros conatrayentes y de éstos el único que resultó enuna mayor tasa de visitas que los huelleros sinatrayente fue la mezcla de Wild Cat y CanineCall (Tabla 3).
Bosque Nublado
Los transectos se instalaron entre los 2.400y 3.000 m snm. Un total de ocho especies seregistraron en este piso altitudinal.
El felino más pequeño registrado era conseguridad el tilcayo (Leopardus tigrinus). Estaespecie fue recientemente registrada en el Par-que Cotapata (y por primera vez en Bolivia),donde al parecer es bastante común (Pachecoet al., 2001) y no existe otro felino con unahuella tan pequeña (3 x 3 cm, ver Becker yDalponte, 1999:99) en la zona. Suponemos queel tigresillo era L. pardalis, por el tamaño dela huella (5 x 6 cm, ver Aranda y March1987:111; Becker y Dalponte, 1999:101;Simonetti y Huareco, 1999).

172 Mastozoología Neotropical, 2003
Tabla 2
Índices de actividad (proporción de huelleros con huella por transecto) de mamíferos grandes en el pisoaltoandino. El efecto de los cebos se refiere a la eficiencia relativa de los distintos atrayentes utilizados y fuecomparado con una prueba Friedman (c2). Cebo vs sin cebo es la comparación entre la tasa de visitas a loshuelleros sin atrayente con aquella observada utilizando algún atrayente y fue realizada con una prueba deMann Whitney (Z). Las cifras para cada atrayente son mediana y rango; s/r = sin registro.
Activity indices (proportion of track plots with visits per transect) of large mammals in the high andes. Thelure effect refers to the relative efficiency of the distinct lures used and was compared with a Friedman (c2)test. With vs. without lures is the comparison of visitation rates between track plots without lure and thosewhere a lure was used, and was done with a Mann-Whitney Z test. Figures for each site are median andrange; s/r = not registered.
Especie Sin cebo Canine Call Mezcla Cat Passion Efecto de Cebos vs.los cebos Sin cebo
Taruja 0 (0-0,17) 0 (0-0,17) 0 (0-0,17) 0 (0-0,17) c2 = 1,0 Z = 0,48(Hippocamelus antisensis) P = 0,70 P = 0,63
g.l = 3 N = 12, 36
Vizcacha de la sierra 0 (0-0,17) 0 (0-0,17) s/r 0 (0-0,17)c2 = 4,38 Z = 1,28(Lagidium vizcacia) P = 0,29 P = 0,20
g.l. = 3 N = 6, 18
Zorro 0 (0-0,17) 0,08 (0-0,67) 0 (0-0,67) 0,25 (0-0,50)c2 = 9,3 Z = 2,63(Pseudalopex culpaeus) P = 0,026 P = 0,008
g.l. = 3 N = 12, 36
Titi s/r 0 (0-0,17) 0 (0-0,33) 0 (0-0,33) c2 = 4,36 Z = 2,00(Oreailurus jacobita?) P > 0,2 P = 0,045
g.l. = 3 N = 12, 36
Zorrino s/r 0 (0-0,33) 0 (0-0,17) 0 (0-0,17) c2 = 4,38 Z = 2,09(Conepatus chinga) P > 0,2 P = 0,031
g.l. = 3 N = 6, 18
Tabla 3
Índices de actividad (proporción de huelleros con huella por transecto) de mamíferos grandes en el piso delpáramo. El análisis estadístico y la leyenda son similares a los de la Tabla 2. “-.” indica que se registraronmenos de tres huelleros con huella, por tanto no se analizaron estadísticamente los datos.
Activity indices (proportion of track plots with visits per transect) of large mammals in the páramo. Statisticalanalysis and legends are similar to Table 2. “-.” Indicates that less than three track plots were visited andno statistical analysis was attempted.
Especie Sin cebo Canine Call Mezcla Cat Passion Efecto de Cebos vs.los cebos Sin cebo
Vizcacha de la sierra 0 (0-0,17) s/r s/r s/r -. -.(L. vizcacia)
Zorro s/r 0 (0-0,17) 0,17 (0-0,67) 0,08 (0-0,33) c2 = 8,82 Z = 2,25(P. culpaeus) P < 0,03 P = 0,025
g.l. = 3
Titi 0 (0-0,17) s/r s/r s/r -. -.(Oncifelis colocolo?)

NOTAS TÉCNICAS - TECHNICAL NOTES 173
La cabrilla visitó los huelleros solamentecuando se usó Canine Call o cuando no se usóatrayente, y no se registró cuando se utilizaronla mezcla de Canine Call y Wild Cat o el CatPassion. El atrayente más efectivo para eltilcayo (cuya tasa de visitas difirió de cuandono se usaron atrayentes) fue Cat Passion, mien-tras que Canine Call y Cat Passion fueron losmás efectivos para el zorro. No se hallarondiferencias en la tasa de visitas de sari y no seobtuvieron suficientes visitas de tigrecillo, llapa,chancho, venado y uchuñari (Tabla 4).
Bosque montañoso de Yungas
Los huelleros se instalaron entre 1.800 y2.000 m snm. y fueron visitados solamente porllapa (A. paca), la cual mordía los atrayentes.También se registró una huella de jucumari(Tremarctos ornatus), pero fue con seguridad
un animal en tránsito ya que la huella estabaen una esquina del huellero y no se notaba queel animal se hubiera acercado al atrayente(Tabla 5).
DISCUSIÓN
El uso de atrayentes fue útil para el estudio decarnívoros en nuestros sitios de estudio. Su usoresultó en un incremento del número de espe-cies registradas en los huelleros y los carnívo-ros siempre visitaron más los huelleros quetenían algún tipo de atrayente. Esto concuerdacon lo reportado para carnívoros centroameri-canos (Harrison, 1997), pero es contrario a loreportado por Ríos-Uzeda (2001) para algunosde los sitios que se evaluaron en nuestro estu-dio. Es posible que nuestro diseño con mayornúmero de transectos (12) con menos (seis)
Tabla 4
Índice de actividad de mamíferos grandes en el bosque nublado. Las comparaciones estadísticas y laleyenda son similares a la Tabla 2. Large mammals activity index in the cloud forest. Statistical
comparisons and legends are similar to Table 2.
Especie Sin cebo Canine Call Mezcla Cat Passion Efecto de Cebos vs.los cebos Sin cebo
Cabrilla 0 (0-0,67) 0 (0-0,33) s/r s/r c2 = 11,93 Z = 2,36(Mazama chunyi) P < 0,01 P = 0,018
g.l. = 3
Tilcayo s/r 0 (0-0,50) 0 (0-0,83) 0,17 (0-0,17) c2 = 8,82 Z = 2,52(Leopardus tigrinus) P < 0,03 P = 0,011
g.l. = 3
Tigresillo 0 (0-0,17) 0 (0-0,17) s/r 0 (0-0,17) -. -.(Leopardus pardalis?)
Uchuñari s/r s/r s/r 0 (0-0,17) -. -.(Nasua nasua)
Llapa 0 (0-0,17) s/r 0 (0-0,17) s/r -. -.(Aguti paca)
Venado 0 (0-0,17) s/r s/r s/r -. -.(Mazama americana)
Chancho s/r 0 (0-0,17) s/r s/r -. -.(Tayassu tajacu)
Zorro s/r 0 (0-0,33) s/r 0 (0-0,17) c2 = 8,14 Z = 1,04(P. culpaeus) P < 0,044 P = 0,29
g.l. = 3

174 Mastozoología Neotropical, 2003
huelleros, en lugar de tres transectos con 15huelleros, haya posibilitado la detección dediferencias.
Las diferencias entre los distintos atrayentes,que se dieron en nuestro estudio solamente parael zorro y el tilcayo, sugieren que el mejoratrayente para nuestros sitios de estudio es elCat Passion, seguido por Canine Call. El únicootro estudio disponible que evaluó el uso deCat Passion es el de Travaini et al. (2001) paraP. culpaeus en la Patagonia, pero estos autoresno hallaron diferencias entre los atrayentesusados. Creemos ahora que, en un programade monitoreo de largo plazo en nuestros sitiosde estudio, sería mejor usar uno de los dosatrayentes arriba citados, si es que sólo sepretende estudiar las tendencias poblacionalesde los carnívoros, pues las otras especies mos-traron indiferencia (el caso de la taruja y lavizcacha de la sierra) o aparente rechazo (ca-brilla) a los atrayentes usados.
Algo impactante es que, tanto en este traba-jo como en el de Ríos-Uzeda (2001), no seregistró ni una sola especie en el páramo delcerro Hornuni. Este autor argumenta que elpáramo del cerro Hornuni es muy pequeño enextensión y sería como una “isla rodeada debosque nublado” y que los animales visitaríanla zona, pero no serían residentes en ella. Ríos-Uzeda (2001) encontró heces de puma yjucumari, además de restos de bromeliáceascomidas por este último; también fue informa-do por pobladores locales de la eventual pre-sencia de taruja y cabrilla en esa zona. En todocaso, ninguna de las especies registradas en el
otro sitio de páramo (Coscapa) son exclusivasde dicho piso altitudinal; todas ellas fueronregistradas también en el piso Altoandino. Estohace pensar que el Páramo podría no ser unaformación distinta del Altoandino, desde laperspectiva de los mamíferos grandes.
Es notable que la cabrilla estuvo presente enlos huelleros sin atrayente y también cuandose usó Canine Call, pero desapareció al utili-zar los atrayentes para felinos. Ríos-Uzeda(2001) utilizó un atrayente de amplio espectro(para coyotes, linces y zorros) en este piso yobtuvo altas tasas de visita de cabrilla, tantocon el atrayente como sin él. Podría especularseque los atrayentes para felinos actúan como undisuasivo para esta especie. En general, la vi-sita de especies no carnívoras a los huelleroscon atrayentes para carnívoros no es rara(Morrison et al., 1981; Harrison, 1997), aun-que el registro de esas especies es más fre-cuente cuando no se usan atrayentes (Engemany Allen, 2000; este estudio).
El registro de L. pardalis en el bosque nu-blado indica que no es tan raro encontrarle aalturas mayores a 1.200 m, como sugierenNowell y Jackson (1996). Esta especie fuetambién registrada en bosque nublado en elParque Nacional Carrasco, en el departamentode Santa Cruz, Bolivia (Rumiz et al., 1998).
Los pobres resultados para el bosque monta-ñoso de Yungas contrastan fuertemente con loobservado por Ríos-Uzeda (2001) para estepiso altitudinal. Este autor reporta nueve espe-cies para este piso, contra dos en este trabajo.No parece que el menor número de días hubie-
Tabla 5
Índice de actividad de mamíferos grandes en el bosque montañoso de Yungas.El análisis estadístico y la leyenda son similares a la Tabla 2. Large mammals activity index in the
Yungas montane forest. Statistical analysis and legends are similar to Table 2.
Especie Sin cebo Canine Call Mezcla Cat Passion Efecto de Cebos vs.los cebos Sin cebo
Llapa s/r 0 (0-0,17) s/r 0 (0-0,17) -. -.(Aguti paca)
Jucumari s/r s/r s/r 0 (0-0,17) -. -.(Tremarctos ornatus)

NOTAS TÉCNICAS - TECHNICAL NOTES 175
ra influido en nuestros resultados, ya que Ríos-Uzeda reporta que el total de las especies fueregistrado los primeros 2-3 días; aunque Rumizy Herrera (2000) sugieren un período mayorpara registrar todas las especies presentes. Sinembargo, es más plausible que el uso de vene-no para controlar vertebrados plaga de loscultivos en la zona cercana a nuestros transectospoco antes de nuestro muestreo, haya contri-buido a que la riqueza y abundancia de mamí-feros haya resultado tan baja en nuestro sitiodel bosque montañoso de Yungas.
El total de especies registradas por Ríos-Uzeda (2001) para bosque nublado y bosquemontañoso de Yungas (no tuvo registros enpáramo) fue de diez, igual en número a loregistrado durante este trabajo. Sin embargo,tres de las especies registradas por Ríos-Uzeda(Didelphis marsupialis, Puma concolor y Eirabarbara) no lo fueron durante este estudio, eigual número de especies (L. tigrinus,Tremarctos ornatus y Pseudalopex culpaeus)fueron registradas durante este estudio y nopor Ríos-Uzeda (2001) para esos dos pisosaltitudinales. La ausencia de L. tigrinus en losregistros de Ríos-Uzeda (2001) puede ser sóloresultado de la prudencia de dicho autor enincluir todos los felinos pequeños como“Felidae sp.”, debido principalmente a que lapresencia de L. tigrinus no estaba comprobadaaún en la zona, ni en Bolivia (Pacheco et al.,2001). Tanto el puma como el jucumari tienenámbitos de hogar muy grandes (Nowell yJackson, 1996; Rechberger et al., 2001) y suregistro en ambos estudios puede haber sidocasual. No tenemos explicación para las otrasespecies no registradas en ambas campañas.
Como conclusión de este trabajo sugerimosque, un programa de monitoreo de poblacio-nes de carnívoros en el Parque Nacional y ÁreaNatural de Manejo Integrado Cotapata deberíaconsiderar el uso de huelleros cebados con elatrayente Cat Passion y el monitoreo de espe-cies no carnívoras debería realizarse sin el usode atrayentes. Falta por ajustar el número detransectos a instalarse y el número de días derevisión para cada uno. La distancia entretransectos utilizada en este trabajo no es unbuen referente para los carnívoros, pues está
por debajo de lo recomendado por la totalidadde los trabajos previos con este grupo de ma-míferos. Las distancias sugeridas varían entre1,3 a 5 km entre transectos y 0,4 – 0,8 kmentre huelleros (Roughton y Sweeny, 1982;Sargeant et al., 1998; Engeman et al., 2000;Novaro et al., 2000). Sin embargo, estas dis-tancias deben ajustarse de acuerdo a la movi-lidad de la especie de interés primario(Roughton y Sweeny, 1982). Aquí sugerimosuna distancia de 1,3 km entre transectos y de0,4 km entre huelleros (Novaro et al., 2000)como mínimo, debido a las enormes dificulta-des que implica moverse en los hábitats mon-tañosos de fuerte pendiente, especialmente debosque nublado.
AGRADECIMIENTOS
Este trabajo fue realizado con el apoyo de Wildlife Con-servation Society y John D. & Catherine T. MacArthurFoundation. Damián Rumiz y Alejandra Roldán leyeronuna versión preliminar de este trabajo. Varias personasapoyaron en el trabajo de campo y nuestro agradecimien-to va para ellos también. Andrés Novaro (especialmente)y un revisor anónimo contribuyeron a mejorar sustancial-mente este manuscrito.
LITERATURA CITADA
ARANDA, M. e I. MARCH. 1987. Guía de los mamífe-ros silvestres de Chiapas. Instituto Nacional de Inves-tigaciones sobre los Recursos Bióticos, Xalapa,Veracruz, 149 pp.
BECKER, M. y J.C. DALPONTE. 1999. Rastros demamíferos silvestres brasileros. Universidad de Bra-silia – IBAMA, Brasilia, 180 pp.
CAUGHLEY, G. y A.R.E. SINCLAIR. 1994. WildlifeEcology and Management. Blackwell Science,Cambridge, 334 pp.
ENGEMAN, R.M. y L. ALLEN. 2000. Overview of apassive tracking index for monitoring wild canidsand associated species. Integrated Pest ManagementReviews, 5:197-203.
ENGEMAN, R.M.; M.J. PIPAS, K.S.GRUVER y L.ALLEN. 2000. Monitoring coyote population changeswith a passive activity index. Wildlife Research,27:553-557.
GOMEZ, H.; R.B. WALLACE y C. VEITCH. 2001.Diversidad y abundancia de mamíferos medianos ygrandes en el noreste del área de influencia del Par-que Nacional Madidi durante la época húmeda.Ecología en Bolivia, 36:17-30.
HARRISON, R.L. 1997. Chemical attractants for CentralAmerican felids. Wildlife Society Bulletin, 25:93-97.
JOHNSON, W.E. y W.L. FRANKLIN. 1991. Feeding andspatial ecology of Felis geoffroyi in southernPatagonia. Journal of Mammalogy, 72:815-820.

176 Mastozoología Neotropical, 2003
LÓPEZ, R. 1998. ¿Páramo yungueño, pradera parámica?¿Por qué identificamos las formaciones situadas so-bre la ceja de montaña con el páramo? Ecología enBolivia, 31:93-95.
MORRISON, D.W.; R.M. EDMUNDS, G. LINSCOMBEy J.W. GOERTZ. 1981. Evaluation of specific scentstation variables in Northcentral Louisiana. Proc. Ann.Conf. S.E. Assoc. Fish & Wildl. Agencies, 35:281-291.
NOVARO, A.J.; M.C. FUNES, C. RAMBEAUD y O.MONSALVO. 2000. Calibración del índice de esta-ciones odoríferas para estimar tendencias poblacio-nales del zorro colorado (Pseudalopex culpaeus) enPatagonia. Mastozoología Neotropical, 7:81-88.
NOWELL, K. and P. JACKSON. 1996. Wild Cats. Statussurvey and conservation action plan. IUCN/SSC CatSpecialist Group, Gland, 382 pp.
PACHECO, L.F; J.F. GUERRA, S.L. DEEM y C.P.FRÍAS. 2001. Primer registro de Leopardus tigrinus(Shreber, 1775) en Bolivia. Ecología en Bolivia.
PAINTER, R.L.; R.B. WALLACE y D. PICKFORD. 1995.Relative abundances of peccaries in areas of differenthuman ressures within the Beni Biosphere Reserve,Bolivia. IBEX J.M.E., 3:49-52.
RECHBERGER, J.; R.B. WALLACE y H. TICONA.2001. Un movimiento de larga distancia de un osoandino (Tremarctos ornatus) en el norte del departa-mento de La Paz, Bolivia. Ecología en Bolivia, 36:73-74.
REDFORD, K.H. y J.F. EISENBERG. 1992. Mammalsof the Neotropics. Vol. 2. The southern cone. TheUniversity of Chicago Press, Chicago, 430 pp.
RIBERA-ARISMENDI, M.O. 1995. Aspectos ecológicos,del uso de la tierra y conservación en el ParqueNacional y Área Natural de Manejo IntegradoCotapata. Pp. 1-84. En: Caminos de Cotapata. (deMorales, C.B., ed.). Instituto de Ecología, La Paz,174 pp.
RÍOS-UZEDA, B. 1999. Evaluación de métodos indirec-tos para detectar presencia de mamíferos medianos ygrandes en hábitats de montaña. Tesis de Licenciatu-ra. Universidad Mayor de San Andrés, La Paz, Boli-via. 65 pp.
RÍOS-UZEDA, B. 2001. Presencia de mamíferos media-nos y grandes en el Parque Nacional y Área Naturalde Manejo Integrado Cotapata a través del uso demétodos indirectos. Ecología en Bolivia, 35:3-16.
RÍOS-UZEDA, B.; R.B. WALLACE, H. ARANÍBAR yC. VEITCH. 2001. Evaluación de mamíferos media-nos y grandes en el bosque semideciduo del AltoTuichi (PN y ANMI Madidi, Depto. La Paz). Ecologíaen Bolivia, 36:31-38.
ROUGHTON, R.D y M.W. SWEENY. 1982. Refinementsin scent-station methodology for assessing trends incarnivore populations. Journal of Wildlife manage-ment, 46:217-229.
RUMIZ, D.I.; C.F. EULERT y R. ARISPE. 1998. Evalua-ción de la diversidad de mamíferos medianos y gran-des en el Parque Nacional Carrasco (Cochabamba,Bolivia). Revista Boliviana de Ecología y Conserva-ción Ambiental, 4:77-90.
RUMIZ, D.I. y J.C. HERRERA. 2000. Wildlife diversityand selective mahogany logging in Bosque Chimanes,Beni, Bolivia: surveying mammals and othervertebrates by line transects, track quadrats, live-trapping and mist-netting. Pp. 235-261. En:Biodiversidad, conservación y manejo en la regiónde la Reserva de la Biosfera Estación Biológica delBeni, Bolivia (Herrera-MacBryde, O.; F. Dallmeier,B. MacBryde, James Comiskey y C. Miranda, eds.).SI/MAB Series No. 4. Smithsonian Institution, Was-hington, D.C., 423 pp.
SIMONETTI, J.A. e I. HUARECO. 1999. Uso de huellaspara estimar diversidad y abundancia relativa de losmamíferos de la Reserva de la Biósfera – EstaciónBiológica del Beni, Bolivia. Mastozoología Neotropi-cal, 6:139-144.
WALKER, R.S.; A.J. NOVARO y J.D. NICHOLS. 2000.Consideraciones para la estimación de abundancia depoblaciones de mamíferos. Mastozoología Neotropical,7:73-80.
ZAR, J.H. 1996. Biostatistical analysis. Prentice-Hall, NewJersey, 663 pp.

NOTAS TÉCNICAS - TECHNICAL NOTES 177ISSN 0327-9383Mastozoología Neotropical / J. Neotrop. Mammal.; 10(1):177-181
©SAREM, 2003 Versión on-line ISSN 1666-0536
Recibido 12 setiembre 2002. Aceptación final 23 enero 2003.
NOTAS TÉCNICAS - TECHNICAL NOTES
A SIMPLE GROUND-BASED METHOD FOR TRAPPINGSMALL MAMMALS IN THE FOREST CANOPY
Maurício E. Graipel
Depto. de Ecologia e Zoologia, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina,Campus Universitário, Trindade, 88.040-970, Florianópolis - SC - Brazil; Phone/Fax: (048) 3319626/3315156; <[email protected]>
ABSTRACT. A simple and effective ground-based method was developed for trapping smallmammals in the forest canopy. By using this method traps can be set closer to ideal gridlocations, without tree climbing, with quicker set up (3 min. after looping a cable around thechosen branch), and faster rebaiting (47 sec.) when compared to other methods applied.This method is described and its efficiency and feasibility are discussed in this paper.
RESUMEN. Un método simple con base en el suelo para el trampeo de pequeñosmamíferos en el dosel arbóreo. Se desarrolló un método simple y efectivo para el trampeode pequeños mamíferos en el dosel arbóreo desde el suelo. En este método las trampaspueden ser colocadas próximas de su posicion ideal en la cuadrícula, rápidamente y sinsubir a los árboles. Se colocan en 3 min despues de pasar una cuerda por la rama elegiday son recebadas en 47 segundos. En este trabajo se describe el método y se discuten sueficiencia y praticabilidad.
Key words: platforms, rainforest, rodents, marsupials.
Palabras clave : plataformas, floresta pluvial, roedores, marsupiales.
INTRODUCTION
Ecological aspects of arboreal small mammalsmay be investigated through several methodssuch as smoked paper tracking (Justice, 1961),direct observation (Voss and Emmons, 1996),spool and line (Miles, 1976) and radio-track-ing (Charles-Dominique et al., 1981). Livecapture of arboreal small mammals can beobtained through traps set directly on branchesin canopy (August, 1983) or alternativelyplaced on wooden platforms (Malcolm, 1991;Vieira, 1998). These procedures have beensuccessfully used to trap small mammals inseveral Neotropical forests (McClearn et al.,1994; Passamani, 1995; Leite et al., 1996;
Voltolini, 1998; Vieira and Monteiro-Filho, inpress). For trapping the bamboo rat,Kannabateomys amblyonyx, bait-less traps set-ting on artificial bridges can also be used(Kierulff et al., 1991). Nevertheless, tree climb-ing is required to place and remove platformson the canopy using the pulley method de-scribed by Malcolm (1991), traps on branches(August, 1983) or artificial bridges (Kierulff etal., 1991). For traps set on branches climbingis also required to rebait (August, 1983) and toremove the captured animals (August, 1983;Kierulff et al., 1991). Techniques to climb treeshave been described in several studies(Whitacre, 1981; Moffett and Lowman, 1995).“Free” climbing (Voltolini, 1998) or ladders

178 Mastozoología Neotropical, 2003
(Wallauer et al., 2000) are also used. How-ever, these methods are potentially dangerous(Laman, 1995) and time-consuming. Barker andSutton (1997) discussed advantages and disad-vantages of several climbing methods.
The simplest options for canopy access areoften ground-based (see Moffett and Lowman,1995; Barker and Sutton, 1997), as in Vieira(1998). The methods described by Malcolm(1991) and Vieira (1998) for trapping smallmammals demand complex platforms and re-sult in the actual location of arboreal trapsplaced at some distance from the ideal loca-tions of a grid design. Malcolm’s method re-quires tree climbing and the method describedby Vieira (1998) uses branches parallel to theground with diameter ranging from 10 to 25 cm.Vieira (1998) also describes disadvantages ofMalcolm’s method.
A simple and effective alternative ground-based method for trapping small mammals inthe canopy was developed during a smallmammal survey in the Atlantic rainforest insouthern Brazil. By using this method, trapscan be quickly set without climbing and inlocations which correspond more closely toideal grid locations, when compared to the traplocations acheived by other methods. In thisstudy this method is described and its efficiencyand feasibility are discussed.
METHODS
Study areaThe study was developed within the boundaries
of the town of Itapoá, Santa Catarina State, SouthernBrazil, at 26°04' S and 48°37' W. The study area iscovered with secondary Atlantic Forest vegetationat sea level. The study site was the ‘Reserva Parti-cular do Patrimônio Natural Volta Velha - PalmitalFarm’ (1.186 ha.). The forest was described by Klein(1984) as very uniform, with trees reaching around12-15 m in height.
Field methods and analysisSpatial arrangement of grids consisted of 100
stations in a 10 by 10 configuration. A total of 225traps were used, including 100 traps set on theground, 100 on the understory shrubs, and 25 oncanopy. Twenty five of the 100 traps set on the
ground were of medium size (45 x 15 x 15 cm) andthe rest, including understory and canopy traps, weresmall (26 x 19 x 9 cm). The distance between eachline and between each station on the same line was20m. However, all medium sized traps and canopytraps were set 40m distant from each other.
The bait used was banana mixed with peanutbutter. All traps were monitored daily for fourconsecutive days each month. The trapping periodfor canopy technique analysis was 10 months, fromNovember 2000 to August 2001, but all capturescombined ranged from September 1999 to August2001. Trees or shrubs were not chosen randomlyfor trap placement. Traps were placed in canopiesconnected to neighbouring trees or shrubs, and asnear as possible of the correspondent station.
The time required to install and rebait traps wasrecorded to compare the efficiency of the differenttrapping methods: 1) Mean time to install traps aftera cable was swung around the chosen branch; 2)Mean time to rebait traps, including the time tolower the trap from the branch and lift it back inplace. Other procedures included measuring themean height of platforms, and the distance betweenthe ideal location of the trap station, based on gridconfiguration, and the actual trap site. The meandistance, and standard deviation from the mean,between the actual trap site and the ideal gridlocation is designated scatter, and is used as astatistic to evaluate the efficiency of differentmethods in locating strap sites within a griddesign.
Platform and trap designThe platform is assembled by connecting three
pieces of wood board (2 x 2 x 30 cm) by theirextremities in order to form a “Ì” shape structure.The trap is fixed on the lower end of it, and theunique entrance of the trap faces to inside of the“Ì” structure. A thin wire with an upside down“V” shape, from where the trap hangs, is attachedto the top of the platform on its far end. After thetrap and the platform are assembled together, a cableis attached to the superior part of the “Ì” structurein such a way to maintain the balance of the structurewhen hanging from a branch (Fig. 1). Two cables3 to 5 mm in diameter were used in the operationof lifting the platform. One of the cables lifts thepulley. The other cable is inserted through thepulley and will hold the platform. When thepulley is fastened to the branch, the cable thatwas inserted through it is used to lift theplatform. Both cables are fastened shoulder-highto a nearby tree (Fig. 1).

NOTAS TÉCNICAS - TECHNICAL NOTES 179
Fig. 1. Technique to capture small mammals on the forest canopy. The pulley (in detail) and the platform are not totallysuspended. A – Cable to lift the pulley; B – Cable to lift the platform; C – Wire with “V” shape to sustain thetrap; P – Wooden platform with “Ì” shape; R – pulley. Drawing modified from Vieira (1998).
RESULTS AND DISCUSSION
The method
The method described here is “low-tech” asdefined by Barker and Sutton (1997). Thewhole trap assemblage, including cables andplatforms, were in perfect conditions after 10month of field work. The mean time of hoist-ing the trap to the canopy after the cable hasbeen thrown over a branch was (± standard
deviation) 3 min ± 50 seconds (n = 25; range2:02 - 5:44 seconds). The whole process canbe accomplished by only one person, but, inthis study, two people took part in the fieldwork. The mean time for bait reposition, in-cluding the lowering and lifting of the trap,was 47 seconds ± 12 seconds (n = 25; rangefrom 30 - 85 seconds). This result is at leastfour times faster than the method used by Vieira(1998), who managed to rebait in approximatelyfour minutes moving traps from higher levels

180 Mastozoología Neotropical, 2003
than this experiment used. The results obtainedresulted in faster time to rebaiting and trap setup than the method used by August (1983) andWallauer et al. (2000) and the methods de-scribed by Malcolm (1991) and Vieira (1998).
The mean height of the platforms was 7.25 m± 1.40 m (n = 25; range from 4.8 - 9.8 m).This height is directly related with canopy’sheight, which ranged from 12 to 15 m at theRPPN Volta Velha. Voltolini (1998) and Vieira(1998) used higher platforms (mean 9.5 m,range 7.50-12.00 m, and mean 10.1 m range6.00-15.00 m, respectively) at forests withcanopy varying from 20 to 30 m. Malcolm(1991) used platforms at mean height of 15 mnear Manaus, Brazil, in canopies reaching over30 m.
The scatter from the predetermined trap site(trapping post) based on a grid design and theactual trap site was 3.91 m ± 2.08 m (n = 25;range 0.5 - 9.3 m). Although there is no re-corded data on such variation in other trappingmethods, it is most likely that the method de-scribed here, allowing the trap set up onbranches with diameter inferior to five centi-meters, or even on vines as I have done, willresult in traps more closely placed to ideal gridloction than other methods mentioned earlier.
Trapping success in the canopy
During the capture period in the canopy Irecorded 608 captures on the ground (4,000trap-nights), 155 captures on the understory(4,000 trap-nights), and 69 captures on thecanopy (1,000 trap-nights). The overall trap-ping success (total number of captures/totalnumber of trap-nights) was 15.2% at groundlevel, 3.88% at understory and 6.9% for thecanopy. The overall trapping success at thecanopy was superior to the ones recorded byVoltolini (1998) (1.29%), Wallauer et al.(2000) (2.33%), Vieira (1998) (2.4%),McClearn et al. (1994) (3.7%), Passamani(1995) (approximately 5%) and Stallings (1989)(6.3%). The trapping success in the canopyper capture session ranged from 0 to 19%.
The number of captures per trapping effortin the canopy was superior to the number ofcaptures in the understory and inferior to thenumber at ground level. Because there is a
different fauna assemblage at different heightlevels, it is not appropriate to conclude thatthere is a difference in efficiency of capturesat different height levels. The fauna assemblageat ground level included exclusively terrestrialspecies, as well as species mainly associatedwith canopy. Besides that, Vieira (1998) notedthat a trap is more easily accessed by mam-mals when on the ground than when placed inthe canopy.
The captured fauna
One rodent and two marsupial species werecaptured in the canopy, whereas two rodentspecies (Nectomys squamipes andOligoryzomys nigripes) were captured in theunderstory and three terrestrial small mammalspecies, one marsupial and two rodent species(Metachirus nudicaudatus; Akodon montensis;Oryzomys russatus) were captured only inground level traps. All species captured in thecanopy and understory were also captured atground level (Table 1). Sciurus aestuans wasvisually observed near the study area, but itwas not trapped.
ACKNOWLEDGEMENTS
I thank Ivo R. Ghizoni Jr., Claudio R. Graipel, MarcosAzevedo, Rafael Reinicke, Jorge Cherem andundergraduate students for helping in field work. Natanoeland Lucio Machado kindly authorized the field work andprovided logistical support at RPPN Volta Velha andspecially to Luis and Chico. I also thank Marcelo Mazzollifor the English translation, Paul Richard M. Miller for theEnglish revision and Enrique Gonzalez and AlfredoLanguth for the translation of Spanish abstract, and JorgeCherem, Fernando Ávila Pires, Nilton Cáceres, EnriqueGonzales and Emerson Vieira for the improvement of themanuscript. This research project was partially funded byPICDT/CAPES.
LITERATURE CITED
AUGUST, P.V. 1983. The role of habitat complexity andheterogeneity in structuring of tropical mammalscommunities. Ecology, 64:1495-1507.
BARKER, M.G. and S.L. SUTTON. 1997. Low-techmethods for forest canopy access. Biotropica, 29:243-247.
CHARLES-DOMINIQUE, P.; M. ATRAMENTOWICZ,M. CHARLES-DOMINIQUE, H. GÉRARD, A.HLADIK, C.M. HLADIK, and M.F. PRÉVOST. 1981.Les mammifères frugivores arboricoles nocturnesd’une foret guyanaise: inter-relations plantes-animaux.Revue d’Ecologie (Terre et Vie), 35:341-435.
NOTAS TÉCNICAS - TECHNICAL NOTES 181
JUSTICE, K.E. 1961. A new method for measuring homeranges of small mammals. Journal of Mammalogy,42:462-470.
KIERULFF, M.C.; J.R. STALLINGS, and E.L. SABATO.1991. A method to capture the bamboo rat(Kannabateomys amblyonyx) in bamboo forests.Mammalia, 55:633-635.
KLEIN, R.M. 1984. Aspectos dinâmicos da vegetação dosul do Brasil. Sellowia, 36:5-54.
LAMAN, T.G. 1995. Safety recommendations for climbingrain forest trees with “single rope technique”.Biotropica, 27:406-409.
LEITE, Y.L.R.; L.P. COSTA, and J.R. STALLINGS. 1996.Diet and vertical space use of three sympatricopossums in a Brazilian Atlantic forest reserve. Journalof Tropical Ecology, 12:435-440.
MALCOLM, J.R. 1991. Comparative abundances ofNeotropical small mammals by trap height. Journalof Mammalogy, 73:188-192.
MCCLEARN, D.; J. KOHLER, K.J. MCGOWAN, E.CEDEÑO, L.G. CARBONE, and D. MILLER. 1994.Arboreal and terrestrial mammal trapping on Gigantepeninsula, Barro Colorado nature monument, Panama.Biotropica, 26:208-213.
MILES, M.A. 1976. A simple method of trackingmammals and locating triatomine vectors ofTrypanosoma cruzi in Amazonian forest. TheAmerican Journal of Tropical Medicine and Hygiene,25:671-674.
MOFFETT, M.W. and M.D. LOWMAN. 1995. Canopyaccess techniques. Pp. 3-26. In: Forest Canopies
(Lowman, M.D. and N.M. Nadkarni, eds.). AcademicPress, San Diego, California, 624 pp. + xix.
PASSAMANI, M. 1995. Vertical stratification of smallmammals in Atlantic Hill forest. Mammalia, 59:276-279.
STALLINGS, J.R. 1989. Small mammal inventories in aneastern Brazilian park. Bulletin of Florida StateMuseum of Science, 34:153-200.
VIEIRA, E.M. 1998. A technique for trapping smallmammals in the forest canopy. Mammalia, 62:306-310.
VIEIRA, E.M. and E.L.A. MONTEIRO-FILHO. (In press).Vertical stratification of small mammals in the Atlanticrain forest of south-eastern Brazil. Journal of Tropi-cal Ecology.
VOLTOLINI, J.C. 1998. Estratificação vertical demarsupiais e roedores na Floresta Atlântica do sul doBrasil. Dissertação de Mestrado. Zoologia, USP, SãoPaulo. 78 pp.
VOSS, R.S. and L.H. EMMONS. 1996. Mammaliandiversity in Neotropical lowland rainforests: apreliminary assessment. Bulletin of the AmericanMuseum of Natural History, 230:1-115.
WALLAUER, J.P.; M. BECKER, L.G. MARINS-SÁ, L.M.LIERMANN, S.H. PERRETTO, and V.SCHERMACK. 2000. Levantamento dos mamíferosda Floresta Nacional de Três Barras - Santa Catarina.Biotemas, 13:103-127.
WHITACRE, D.F. 1981. Additional techniques and safetyhints for climbing tall trees, and some equipment andinformation sources. Biotropica, 13:286-291.
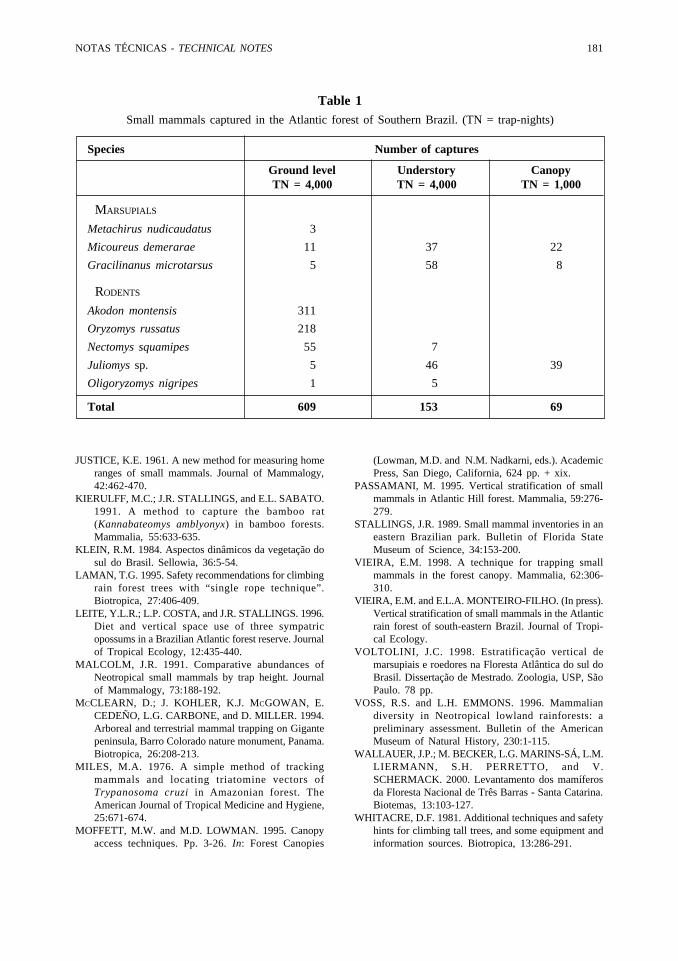
Table 1Small mammals captured in the Atlantic forest of Southern Brazil. (TN = trap-nights)
Species Number of captures
Ground level Understory CanopyTN = 4,000 TN = 4,000 TN = 1,000
MARSUPIALS
Metachirus nudicaudatus 3
Micoureus demerarae 11 37 22
Gracilinanus microtarsus 5 58 8
RODENTS
Akodon montensis 311
Oryzomys russatus 218
Nectomys squamipes 55 7
Juliomys sp. 5 46 39
Oligoryzomys nigripes 1 5
Total 609 153 69

Copyright © 2022 FDOKUMEN




















