
Why are left-right spatial codes easier to form than above-below ones
Upload
independentCategory
view
0download
0
B R I E F C O M M U N I C AT I O N
818 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 9 | SEPTEMBER 2005
terra is a left–right asymmetry gene required forleft–right synchronization of the segmentation clockLeonor Saúde1,3, Raquel Lourenço1, Alexandre Gonçalves1 and Isabel Palmeirim2
To establish the vertebrate body plan, it is fundamental to create left–right asymmetry in the lateral-plate mesoderm to correctly position the organs. However, it is also crucial to maintain symmetry between the left and the right sides of the presomitic mesoderm, ensuring the allocation of symmetrical body structures, such as the axial skeleton and skeletal muscles. Here, we show that terra is an early left-sided expressed gene that links left–right patterning with bilateral synchronization of the segmentation clock.
terra encodes a protein with a zinc-finger-like DNA-binding motif, called the DM domain1. Vertebrate genes with this domain constitute the DMRT (DM-related transcription factor) family, which is implicated in sex deter-mination2. However, mouse dmrt2 and zebrafish terra, also known as dmrt2, were also proposed to have a function in somitogenesis1.
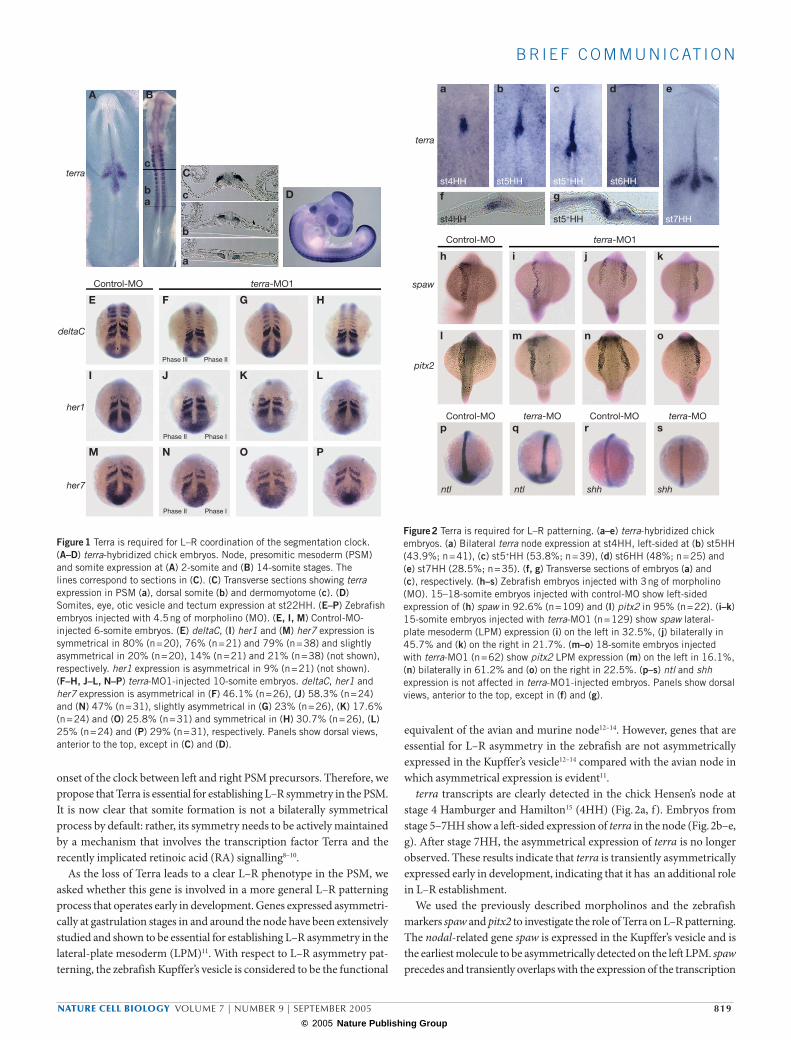
We identified the chick terra gene (see Supplementary Information, Fig. S1) and studied its expression pattern during somitogenesis. terra transcripts are detected in the anterior region of the presomitic meso-derm (PSM) and somites (Fig. 1A–D). In epithelial somites, terra is expressed in the dorsal compartment, which correlates with its later dermomyotome expression (Fig. 1C). This pattern is similar to that pre-viously reported in mouse and zebrafish1, highlighting the conserved nature of terra expression and reinforcing its role in somitogenesis.
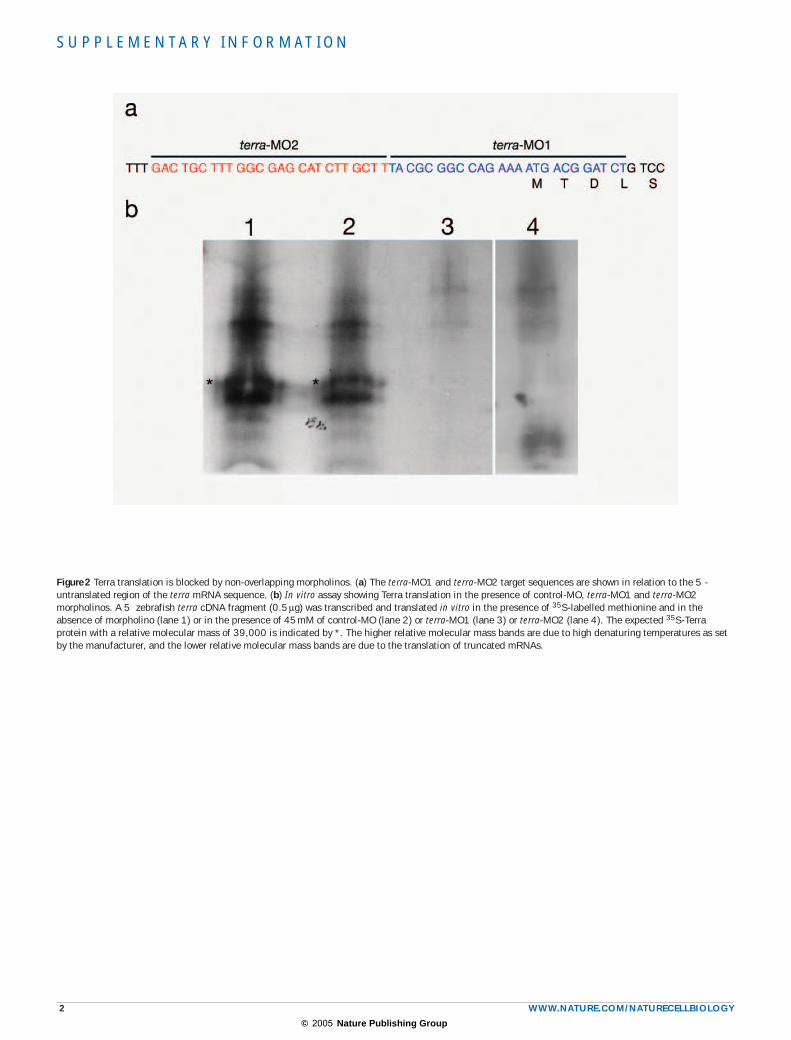
Somite formation is linked to an evolutionarily conserved molecular clock, revealed by cyclic PSM expression of components of the Notch and Wnt pathways3. To investigate the effect of reducing Terra on this clock, we used the morpholino (MO) technology in zebrafish. An anti-sense morpholino oligonucleotide against the translation initiation site of zebrafish terra (terra-MO1), a second non-overlapping morpholino (terra-MO2) and a five-mispair control oligonucleotide (control-MO) were designed and their specificities were demonstrated in an in vitro translation assay (see Supplementary Information, Fig. S2). Morpholino-injected embryos were hybridized with the zebrafish cycling genes del-taC, her1 and her7 (refs 4–6). The normal PSM expression pattern of these cyclic genes consists of 2–3 stripes in its anterior region and a dynamic expression in its posterior part. As a new somite forms, the
1Instituto Gulbenkian de Ciência, Centro de Biologia do Desenvolvimento, Rua da Quinta Grande, 6, 2780-156 Oeiras, Portugal. 2Life and Health Sciences Research Institute (ICVS), School of Health Sciences, University of Minho, 4710-057 Braga, Portugal. 3Correspondence should be addressed to L. S. (e-mail: [email protected])
Published online: 1 September 2005; DOI: 10.1038/ncb1294
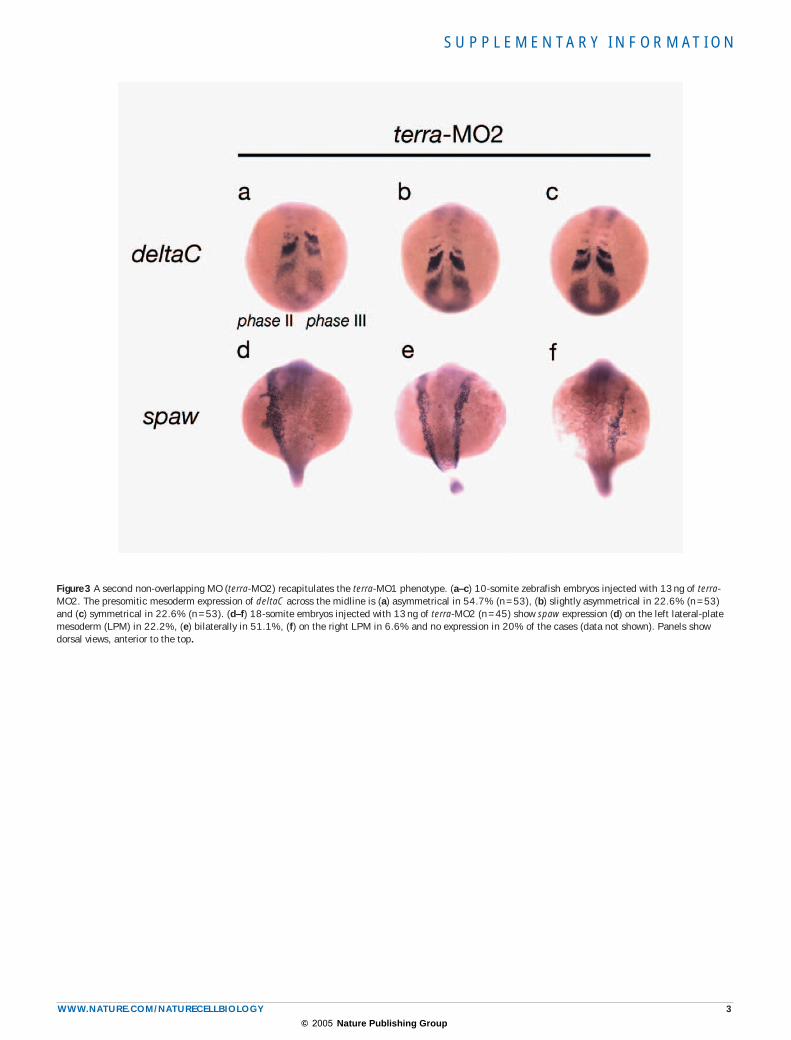
most anterior her1 and her7 stripes fade and the most anterior deltaC stripe stabilizes in the caudal somitic compartment. Concomitantly, a new wave of expression emerges from the tailbud. This dynamic pattern of expression in the PSM is bilaterally symmetrical and can be subdivided into three consecutive phases that are reiterated with the formation of a new somite pair4–6. In terra-MO1-injected embryos, the dynamic PSM expression of deltaC, her1 and her7 is maintained, and different phases of cyclic gene expression are observed (Fig. 1F–H, J–L, N–P). However, these injected embryos show striking phenotypes ranging from: cyclic gene expression on the left PSM in a different phase from the one on the right PSM (Fig. 1F, J, N); cyclic gene expression in the same phase on both left and right PSMs, although not bilaterally symmetrical (Fig. 1G, K, O); and cyclic gene expression in the same phase and bilaterally symmetrical on the left and right PSMs (Fig. 1H, L, P). The terra-MO1 phenotype was recapitulated following injection of the non-overlapping terra-MO2 (see Supplementary Information, Fig. S3).
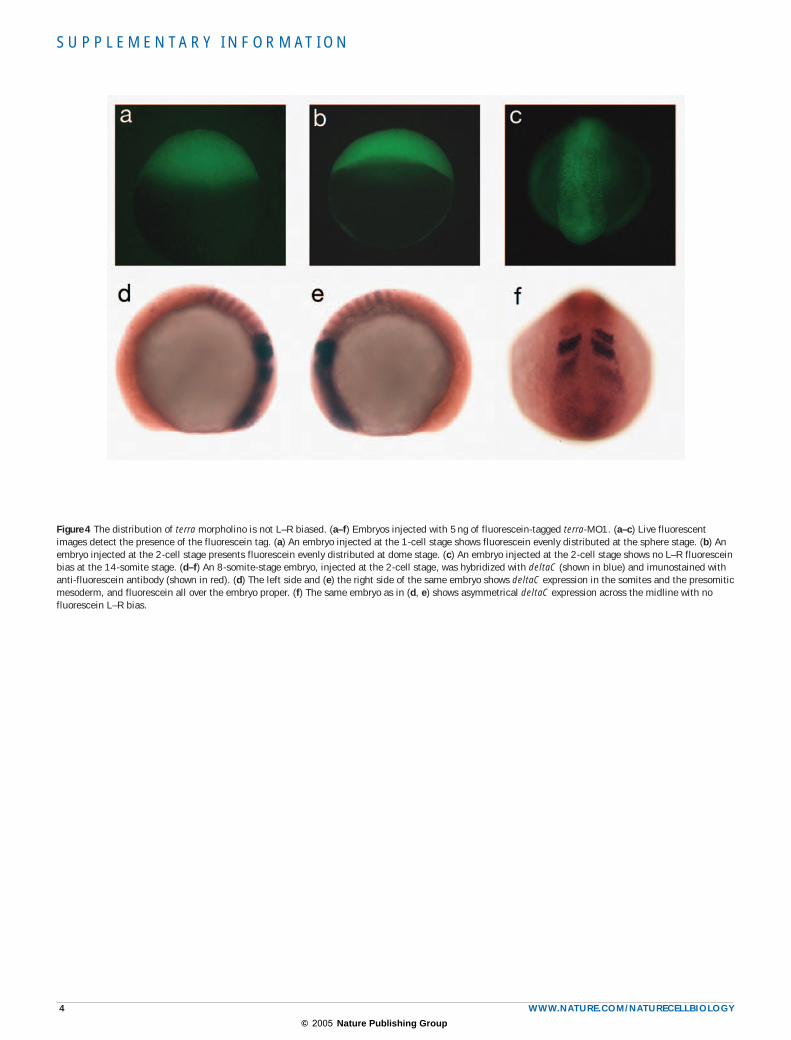
To investigate whether the observed asynchrony in gene expression between the left and the right PSMs was due to a biased distribution of the injected morpholino, we used a fluorescein-tagged version of this morpholino (terra-MO1–Fluo). Fluorescein localization was compared, at later developmental stages, between embryos injected at the 1-cell stage and embryos injected at the 2-cell stage, using fluorescent micro-scopy and imunostaining. An equal distribution of fluorescein between the left and the right sides was observed in all embryos, regardless of the injection stage. Additionally, all the embryos that showed differ-ent deltaC expression phases between the left and the right PSMs did not display any bias in left–right (L–R) fluorescein distribution (see Supplementary Information, Fig. S4).
The expression of deltaC is not affected in the somitic caudal com-partment (Fig. 1E–H), indicating that Terra is not required for anterior–posterior polarity specification. The analysis of somitic deltaC stripes revealed that intersomitic boundaries are aligned across the midline, defining somites with regular sizes (Fig. 1E–H). The preservation of a fixed somite size implies that neither the pacing of the molecular clock nor the displacement of the wavefront7 is affected in terra morphants. Therefore, this phenotype can only be explained by a desynchronized
© 2005 Nature Publishing Group
NATURE CELL BIOLOGY VOLUME 7 | NUMBER 9 | SEPTEMBER 2005 819
B R I E F C O M M U N I C AT I O N
onset of the clock between left and right PSM precursors. Therefore, we propose that Terra is essential for establishing L–R symmetry in the PSM. It is now clear that somite formation is not a bilaterally symmetrical process by default: rather, its symmetry needs to be actively maintained by a mechanism that involves the transcription factor Terra and the recently implicated retinoic acid (RA) signalling8–10.
As the loss of Terra leads to a clear L–R phenotype in the PSM, we asked whether this gene is involved in a more general L–R patterning process that operates early in development. Genes expressed asymmetri-cally at gastrulation stages in and around the node have been extensively studied and shown to be essential for establishing L–R asymmetry in the lateral-plate mesoderm (LPM)11. With respect to L–R asymmetry pat-terning, the zebrafish Kupffer’s vesicle is considered to be the functional
equivalent of the avian and murine node12–14. However, genes that are essential for L–R asymmetry in the zebrafish are not asymmetrically expressed in the Kupffer’s vesicle12–14 compared with the avian node in which asymmetrical expression is evident11.
terra transcripts are clearly detected in the chick Hensen’s node at stage 4 Hamburger and Hamilton15 (4HH) (Fig. 2a, f). Embryos from stage 5–7HH show a left-sided expression of terra in the node (Fig. 2b–e, g). After stage 7HH, the asymmetrical expression of terra is no longer observed. These results indicate that terra is transiently asymmetrically expressed early in development, indicating that it has an additional role in L–R establishment.
We used the previously described morpholinos and the zebrafish markers spaw and pitx2 to investigate the role of Terra on L–R patterning. The nodal-related gene spaw is expressed in the Kupffer’s vesicle and is the earliest molecule to be asymmetrically detected on the left LPM. spaw precedes and transiently overlaps with the expression of the transcription
A
E F G H
I J K L
M N O P
B
C
D
terra
deltaC
her1
her7
Control-MO terra-MO1
Phase III Phase II
Phase II Phase I
Phase II Phase I
c
b
a
c
ba
Figure 1 Terra is required for L–R coordination of the segmentation clock. (A–D) terra-hybridized chick embryos. Node, presomitic mesoderm (PSM) and somite expression at (A) 2-somite and (B) 14-somite stages. The lines correspond to sections in (C). (C) Transverse sections showing terra expression in PSM (a), dorsal somite (b) and dermomyotome (c). (D) Somites, eye, otic vesicle and tectum expression at st22HH. (E–P) Zebrafish embryos injected with 4.5 ng of morpholino (MO). (E, I, M) Control-MO-injected 6-somite embryos. (E) deltaC, (I) her1 and (M) her7 expression is symmetrical in 80% (n = 20), 76% (n = 21) and 79% (n = 38) and slightly asymmetrical in 20% (n = 20), 14% (n = 21) and 21% (n = 38) (not shown), respectively. her1 expression is asymmetrical in 9% (n = 21) (not shown). (F–H, J–L, N–P) terra-MO1-injected 10-somite embryos. deltaC, her1 and her7 expression is asymmetrical in (F) 46.1% (n = 26), (J) 58.3% (n = 24) and (N) 47% (n = 31), slightly asymmetrical in (G) 23% (n = 26), (K) 17.6% (n = 24) and (O) 25.8% (n = 31) and symmetrical in (H) 30.7% (n = 26), (L) 25% (n = 24) and (P) 29% (n = 31), respectively. Panels show dorsal views, anterior to the top, except in (C) and (D).
a
f
b c
g
d e
h i j k
l m n o
p q r s
terra
spaw
pitx2
st4HH
st4HH
st5HH st5+HH
st5+HH
st6HH
st7HH
Control-MO
Control-MO terra-MO terra-MOControl-MO
terra-MO1
ntl ntl shh shh
Figure 2 Terra is required for L–R patterning. (a–e) terra-hybridized chick embryos. (a) Bilateral terra node expression at st4HH, left-sided at (b) st5HH (43.9%; n = 41), (c) st5+HH (53.8%; n = 39), (d) st6HH (48%; n = 25) and (e) st7HH (28.5%; n = 35). (f, g) Transverse sections of embryos (a) and (c), respectively. (h–s) Zebrafish embryos injected with 3 ng of morpholino (MO). 15–18-somite embryos injected with control-MO show left-sided expression of (h) spaw in 92.6% (n = 109) and (l) pitx2 in 95% (n = 22). (i–k) 15-somite embryos injected with terra-MO1 (n = 129) show spaw lateral-plate mesoderm (LPM) expression (i) on the left in 32.5%, (j) bilaterally in 45.7% and (k) on the right in 21.7%. (m–o) 18-somite embryos injected with terra-MO1 (n = 62) show pitx2 LPM expression (m) on the left in 16.1%, (n) bilaterally in 61.2% and (o) on the right in 22.5%. (p–s) ntl and shh expression is not affected in terra-MO1-injected embryos. Panels show dorsal views, anterior to the top, except in (f) and (g).
© 2005 Nature Publishing Group

820 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 9 | SEPTEMBER 2005
B R I E F C O M M U N I C AT I O N
factor pitx2, thereby defining the heart field12,16. In terra-MO1- and terra-MO2-injected embryos, a randomization of the expression of these left-sided-specific genes is observed (Fig. 2i–k, m–o; and see Supplementary Information, Fig. S3), concomitant with randomization of heart position (data not shown). These results demonstrate that Terra is required to establish embryo L–R asymmetry. During development, axial midline tissues, such as the notochord and the floorplate, are required to main-tain L–R asymmetry that acts as a midline barrier11. We examined the integrity of these axial tissues in the terra-MO1-injected embryos using the markers no tail (ntl) and sonic hedgehog (shh). ntl and shh expression was not affected in terra-MO1-injected embryos (Fig. 2p–s), indicating that Terra is not necessary for midline formation. Therefore, Terra must act early in the process of L–R patterning.
This study identifies Terra as the first molecular link between two major events: L–R asymmetrical patterning and bilateral synchroniza-tion of the segmentation clock. A common feature between these two processes is the Notch signalling pathway. The segmentation clock is a Notch-based mechanism that starts well before the formation of the first somite3. Additionally, L–R determination relies on asymmetrical activation of the Notch pathway, as shown by the left-sided expression of delta-like1 and lunatic fringe around the avian node17. These data imply that prospective PSM cells on the left side of the node activate the segmentation clock earlier than those on the right side and that this would lead to L–R asymmetry of PSM cyclic gene expression. We propose that terra expression on the left side of the node has a dual function: it promotes L–R asymmetry information flow and ensures symmetry in PSM precursors.
Taking into account that, without Terra function, LPM markers are randomized (Fig. 2; and see Supplementary Information, Fig. S3), this strongly suggests that in terra-MO-injected embryos, Notch signalling activation may also be randomized. This would then imply that the seg-mentation clock is randomly activated in the absence of Terra. To test this possibility, we determined the number of somites in terra morphants, using myoD as a marker for mature somites. We found that 35% of the 12-somite-stage embryos that were analysed (n = 52) form extra somites, with no significant L–R bias (data not shown). This experiment shows that there is an unbiased desynchronization of the segmentation clock in the absence of Terra. An unbiased formation of extra somites was also
observed when the establishment of L–R information was blocked9, in contrast to the biased somite formation obtained when RA is disrupted8–
10. Taken together, these results are consistent with a model in which Terra acts early in development to promote L–R information flow and then, in combination with RA signalling, protects the PSM from the asym-metrical signals. Therefore, we conclude that Terra is the first identified coordinator of two key processes that set up the vertebrate body plan: L–R asymmetry and PSM bilateral symmetry. These are established essentially at the same time and, ultimately, share the same molecular pathways.
Note: Supplementary Information (including Methods) is available on the Nature Cell Biology website.
ACKNOWLEDGEMENTSWe thank S. Wilson and D. Stemple for the zebrafish embryos, M. Tada and C.-P. Heisenberg for reagents, I. Campos for technical support with MO injections, A. Gaspar for histological work, C. Domingues for the SDS–PAGE gels and I. Marques for sequence analysis. We are grateful to S. Thorsteinsdóttir, S. Simões, W. Wood, S. Rodrigues, R. Andrade, A. Jacinto and A. Coutinho for comments on the manuscript. This work was supported by a grant from Fundação para a Ciência e a Tecnologia (FCT) (POCTI/45914/BCI/2002). L.S. was supported by a FCT fellowship (SFRH/BPD/6755/2001). R.L. was supported by Fundação Calouste Gulbenkian (FCG)/Instituto do Emprego e Formação Profissional (IEFP). The authors are members of the FP6 European Network of Excellence ‘Cells into Organs’.
COMPETING FINANCIAL INTERESTSThe authors declare that they have no competing financial interests.
Received 14 April 2005; accepted 18 July 2005Published online at http://www.nature.com/naturecellbiology.
1. Meng, A. et al. Development 126, 1259–1268 (1999).2. Brunner, B. et al. Genomics 77, 8–17 (2001).3. Dubrulle, J. & Pourquié, O. Development 131, 5783–5793 (2004).4. Jiang, Y. J. et al. Nature 408, 475–479 (2000).5. Holley, S. A. et al. Genes Dev. 14, 1678–1690 (2000).6. Oates, A. C. & Ho, R. K. Development 129, 2929–2946 (2002).7. Cooke, J. & Zeeman, E. C. J. Theor. Biol. 58, 455–476 (1976).8. Vermot, J. et al. Science 308, 563–566 (2005).9. Kawakami, Y. et al. Nature 435, 165–171 (2005).10. Vermot, J. & Pourquié, O. Nature 435, 215–220 (2005).11. Hamada, H. et al. Nature Rev. Genet. 3, 103–113 (2002).12. Long, S. et al. Development 130, 2303–2316 (2003).13. Hashimoto, H. et al. Development 131, 1741–1753 (2004).14. Amack, J. D. & Yost, J. Curr. Biol. 14, 685–690 (2004).15. Hamburger, V. & Hamilton, H. L. J. Morphol. 88, 49–92 (1951).16. Essner, J. J. et al. Development 127, 1081–1093 (2000).17. Raya, A. et al. Nature 427, 121–128 (2004).18. Kimmel, C. B. et al. Dev. Dyn. 203, 253–310 (1995).19. Thisse, C. & Thisse, B. in Zebrafish Science Monitor 5, 8–9 (1998).20.Henrique, D. et al. Nature 375, 787–790 (1995)
© 2005 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
Figure 1 Terra protein sequence alignment. The protein sequences of human (accession number: AAF86295), chimp (accession number: XP_528529), mouse (accession number: NP_665830), zebrafish (accession number: AAD38424) and the identified chick Terra sequence (accession number: AY960292) were aligned using ClustalW. The percentage of similarity of Terra protein between chick and human, chimp, mouse and zebrafish is 82%, 82%, 81% and 72%, respectively.
© 2005 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
2 WWW.NATURE.COM/NATURECELLBIOLOGY
Figure 2 Terra translation is blocked by non-overlapping morpholinos. (a) The terra-MO1 and terra-MO2 target sequences are shown in relation to the 5 -untranslated region of the terra mRNA sequence. (b) In vitro assay showing Terra translation in the presence of control-MO, terra-MO1 and terra-MO2 morpholinos. A 5 zebrafish terra cDNA fragment (0.5 µg) was transcribed and translated in vitro in the presence of 35S-labelled methionine and in the absence of morpholino (lane 1) or in the presence of 45 mM of control-MO (lane 2) or terra-MO1 (lane 3) or terra-MO2 (lane 4). The expected 35S-Terra protein with a relative molecular mass of 39,000 is indicated by *. The higher relative molecular mass bands are due to high denaturing temperatures as set by the manufacturer, and the lower relative molecular mass bands are due to the translation of truncated mRNAs.
© 2005 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 3
Figure 3 A second non-overlapping MO (terra-MO2) recapitulates the terra-MO1 phenotype. (a–c) 10-somite zebrafish embryos injected with 13 ng of terra-MO2. The presomitic mesoderm expression of deltaC across the midline is (a) asymmetrical in 54.7% (n = 53), (b) slightly asymmetrical in 22.6% (n = 53) and (c) symmetrical in 22.6% (n = 53). (d–f) 18-somite embryos injected with 13 ng of terra-MO2 (n = 45) show spaw expression (d) on the left lateral-plate mesoderm (LPM) in 22.2%, (e) bilaterally in 51.1%, (f) on the right LPM in 6.6% and no expression in 20% of the cases (data not shown). Panels show dorsal views, anterior to the top.
© 2005 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
4 WWW.NATURE.COM/NATURECELLBIOLOGY
Figure 4 The distribution of terra morpholino is not L–R biased. (a–f) Embryos injected with 5 ng of fluorescein-tagged terra-MO1. (a–c) Live fluorescent images detect the presence of the fluorescein tag. (a) An embryo injected at the 1-cell stage shows fluorescein evenly distributed at the sphere stage. (b) An embryo injected at the 2-cell stage presents fluorescein evenly distributed at dome stage. (c) An embryo injected at the 2-cell stage shows no L–R fluorescein bias at the 14-somite stage. (d–f) An 8-somite-stage embryo, injected at the 2-cell stage, was hybridized with deltaC (shown in blue) and imunostained with anti-fluorescein antibody (shown in red). (d) The left side and (e) the right side of the same embryo shows deltaC expression in the somites and the presomitic mesoderm, and fluorescein all over the embryo proper. (f) The same embryo as in (d, e) shows asymmetrical deltaC expression across the midline with no fluorescein L–R bias.
© 2005 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
5 WWW.NATURE.COM/NATURECELLBIOLOGY
METHODS
Embryo stagingZebrafish and chick embryos were staged according to Kimmel et al.18 and Hamburger and Hamilton15, respectively.
Whole-mount in situ hybridizationChick terra expressed sequence tags were purchased from the Medical Research Council Geneservice. Zebrafish and chick mRNA in situ hybridizations were per-formed as previously described19,20. Fluorescein detection was carried out using an alkaline-phosphatase (AP)-conjugated anti-fluorescein Fab fragment diluted 1:2000, and INT (2- [4-Iodophenyl]-3-[4-nitrophenyl]-5-phenyl-tetrazolium chloride)/BCIP (5-Bromo-4-chloro- 3-indolyl phosphate, toluidine salt) as a sub-strate.
Cross-sectionsThe chick terra-labelled embryos were embedded in methacrylate (TECNOVIT 8100; Heraeus Kulzer, Wehrheim, Germany) and processed for sectioning at 20^µm using an ultramicrotome (LKB Ultrotome; Stockholm, Sweden).
Morpholino injectionsA terra-specific morpholino (terra-MO1) targeted to the ATG region (AGATCCGTCATTTTCTGGCCGCGTA), a fluorescein-tagged version of terra-MO1 (terra-MO1–Fluo), a second non-overlapping morpholino (terra-MO2) targeted to the 5 -untranslated region (AAGCAAGATGCTCGCCAAAGCAGTC) and a five-mispair control (control-MO) (AGATgCGTgATTTTgTcGCCGCcTA) were purchased from Gene Tools, LLC (Philomath, OR) and injected into the yolk cell of 1–2-cell-stage fertilized zebrafish embryos.
In vitro transcription and translationThe ‘TNT Coupled Reticulocyte Lysate System’ (Promega, Madison, WI) was used according to the manufacturer’s protocol. The reaction mix was applied in a 10% SDS–PAGE gel, and 35S-labelled proteins were detected by autoradiography.
Accession number: The partial chick terra sequence has been submitted to GenBank database (accession number: AY960292).
© 2005 Nature Publishing Group
Copyright © 2022 FDOKUMEN




















