
Contextos de producción de marfil en Valencina de la Concepción (Sevilla)

This article was published in an Elsevier journal. The attached copyis furnished to the author for non-commercial research and
education use, including for instruction at the author’s institution,sharing with colleagues and providing to institution administration.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Temporal variability of 210Pb fluxes and bioturbationin shelf sediments beneath the high primary production
area off Concepcion, central-southern Chile (36�S)
Praxedes Munoz a,b,*, Javier Sellanes a,b, Carina Lange b,c,Maritza Palma b, Marco A. Salamanca c
a Departamento de Biologıa Marina, Universidad Catolica del Norte, Casilla 116, Coquimbo, Chileb Centro de Investigacion Oceanografica en el Pacıfico Sur-Oriental (FONDAP-COPAS), Universidad de Concepcion,
Casilla 160-C, Concepcion, Chilec Departamento de Oceanografıa, Universidad de Concepcion, Casilla 160-C, Concepcion, Chile
Available online 31 August 2007
Abstract
We estimated monthly fluxes of 210Pb in shelf sediments beneath a high productivity area off central-southern Chile(36�S) during 1 year (September 2002–August 2003). Sediment cores were obtained using a multiple corer and were ana-lyzed mainly for 210Pb, total pigments, and macrofauna abundance. The 210Pb inventories and fluxes were estimated forsurface sediments (0–5 cm) and bioturbation coefficients were inferred using chlorophyll-a (reactive) profiles. In general,210Pb content was inversely correlated with phytodetritus fluxes. High photosynthetic pigment contents in surface sedi-ments were consistently associated with lower 210Pb contents. Macrofaunal activity responded to oxygen and organic mat-ter supplies at the sediment surface, generally concentrated in the first centimeters, but particularly so during months ofhigh organic matter fluxes and deficient bottom water oxygen conditions. At this study site, several processes involvedin the 210Pb surface distribution make it difficult to accurately estimate ages at the surface. We postulate that the organicfluxes promote changes in the faunal activity, which, in combination with sediment resuspension and water circulation overthe shelf, produce seasonal variations in the 210Pb inventories.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Radioisotopes; Sediment mixing; Photosynthetic pigments; Benthos; Chile; Concepcion shelf
1. Introduction
210Pb is a natural radionuclide that has been extensively used to estimate sedimentation rates and biologicalmixing in shelf and coastal environments (e.g., McCaffrey and Thomson, 1980; Levin et al., 2003; Crusius
0079-6611/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.pocean.2007.08.015
* Corresponding author. Address: Departamento de Biologıa Marina, Universidad Catolica del Norte, Casilla 116, Coquimbo, Chile.Tel.: +56 51 20 9802; fax: +56 51 20 9812.
E-mail address: [email protected] (P. Munoz).
Available online at www.sciencedirect.com
Progress in Oceanography 75 (2007) 586–602
www.elsevier.com/locate/pocean
Progress inOceanography

Author's personal copy
et al., 2004; Yeager et al., 2004). It is produced by the decay in the atmosphere of 222Rn, which emanatesmainly from rocks and soils. It is deposited onto the continents and oceans and scavenged by particles, withwashout by precipitation constituting the main source of 210Pb to the surface waters (Cochran, 1992). Othersources, such as rivers and the water column itself, are considered comparatively unimportant since shallowwater columns (<100 m water depth) do not constitute a significant source of this radionuclide from 226Radecay and 210Pb has a longer residence time in the soils (Benninger et al., 1975; Cochran, 1992).
Once the 210Pb is deposited at the ocean surface, it has a short residence time in the water column, especiallyalong ocean margins where removal by both inorganic and organic particles is efficient, particularly in produc-tive surface waters (Bruland et al., 1974; Thomson and Turekian, 1976; Bacon, 1988; Cochran, 1992; Nozakiet al., 1997). High correlations between organic particle fluxes derived from primary productivity and 210Pbfluxes to the sediments have been reported from lacustrine and oceanic settings (Nozaki et al., 1980, 1997; Bin-ford and Brenner, 1986; Moore and Dymond, 1988), constituting an important scavenging process for thisradionuclide in areas where primary productivity dominates particle deposition to the sediments.
The usual assumption when establishing a geochronology in shelf and coastal sediments is that the atmo-spheric input of 210Pb is homogeneous and constant over time. However, the seasonal and spatial variabilityof 210Pb inventories in sediments could respond to different processes, including sediment focusing (i.e., areasof enhanced deposition) and biological mixing (Radakovitch and Heussner, 1999; Smoak et al., 2000; Lewiset al., 2002). The settling particles could be enriched in 210Pb during resuspension events, or recently settled par-ticles could reach deep sections within the sediments due to biological activity, since organisms may ingest thesehigher activity particles at the sediment surface and excrete them at depth (Boudreau, 1997; Lewis et al., 2002).
The present year-long time-series studied the 210Pb content, photosynthetic pigment concentrations, andmacrofauna composition, density, and biomass at a shelf site off Concepcion (88 m water depth), reportingon their seasonal variability herein. The study area receives particulate material derived from primary produc-tion in the water column and terrigenous sources. These inputs are seasonally modulated by the occurrence ofstrong upwelling events (in spring–summer) and fluvial fluxes, which are highest during the rainy season(autumn–winter) and the ice melting period (spring) (Arcos and Wilson, 1984, www.dga.cl). We hypothesizedthat 210Pb fluxes would respond to seasonal organic matter fluxes, expecting the distribution of this element inthe surface sediments (upper 5 cm) to be appropriate for establishing a seasonal relationship with local pig-ment fluxes. Our main goals were: (i) to assess the influence of primary production export fluxes and mixingin the distribution of this radionuclide, and (ii) to evaluate the usefulness of this element for describing thehistory of sedimentation in the shelf area.
2. Materials and methods
2.1. Study area
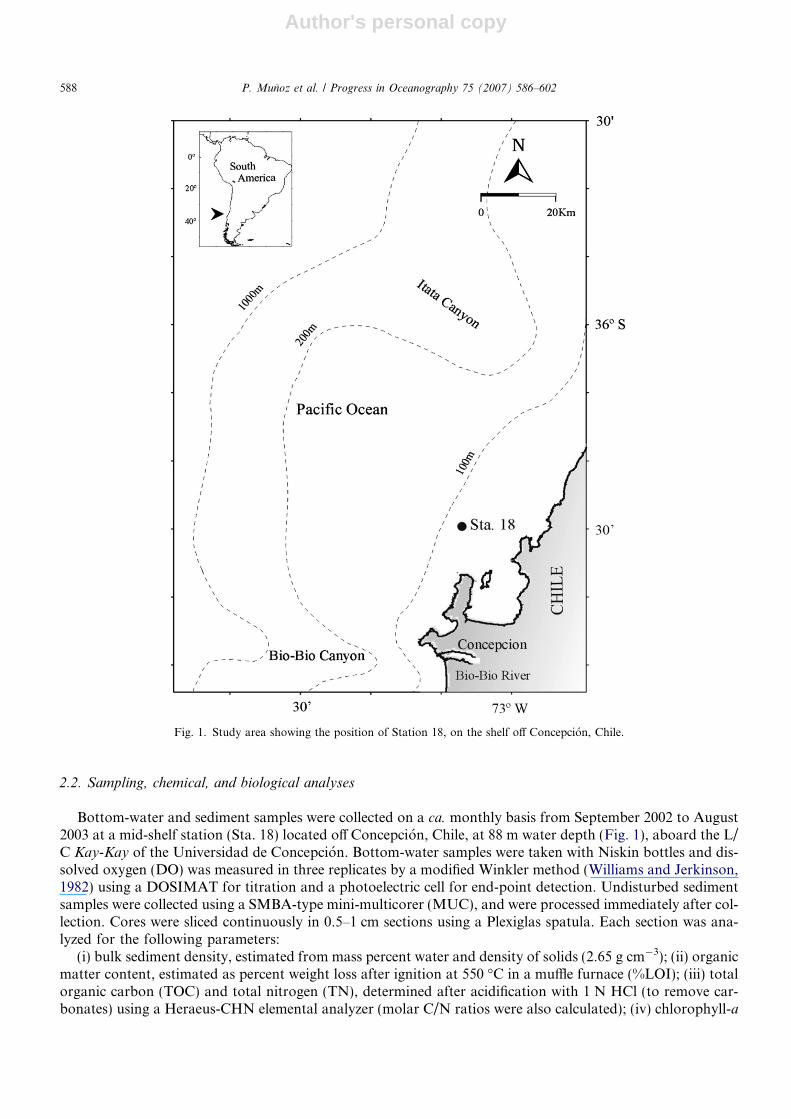
The continental shelf off central-southern Chile (�35–38�S) is wide (�65 km) and interrupted by the sub-marine canyons of the Itata River to the north and the Bio-Bio River to the south (500 and 800 m depth alongtheir respective main axes; Fig. 1). Fluvial discharges are variable during the year (1–5 km3 yr�1) (Arcos et al.,1996; Parra and Habit, 1998; Direccion General de Aguas Chile, www.dga.cl), responding to local seasonalprecipitation that fluctuates between 300 and �1000 mm yr�1 (www.meteochile.cl). The topography controlsthe presence of currents moving towards the open ocean (Sobarzo, 1999; Sobarzo et al., 2001) and the move-ment of particles discharged by the rivers, which are channeled mainly through the canyons to the slope(Scholl et al., 1970; Thornburg and Kulm, 1987a,b; Pineda, 1999).
The area off Concepcion is under the influence of an important upwelling center due to the predominance ofsouthwesterly winds from late spring through early autumn (Arcos and Wilson, 1984), resulting in high phy-toplankton biomass and primary production (highest water column integrated gross primary produc-tion = 19.9 g C m�2 d�1; Daneri et al., 2000). This promotes high fluxes of organic particles to the seafloorthat could cause anoxic conditions in the sediments or even in the bottom water (see Gutierrez et al., 2000and references therein). A flocculent layer is formed at the sediment–water interface and is composed mainlyof phytodetritus. The occurrence and thickness of this layer is variable in time; it is usually associated withbacterial mats (e.g., Beggiatoa, Thioploca), especially at shallower depths (Gutierrez, 2000; Graco et al., 2001).
P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602 587
Author's personal copy
2.2. Sampling, chemical, and biological analyses
Bottom-water and sediment samples were collected on a ca. monthly basis from September 2002 to August2003 at a mid-shelf station (Sta. 18) located off Concepcion, Chile, at 88 m water depth (Fig. 1), aboard the L/C Kay-Kay of the Universidad de Concepcion. Bottom-water samples were taken with Niskin bottles and dis-solved oxygen (DO) was measured in three replicates by a modified Winkler method (Williams and Jerkinson,1982) using a DOSIMAT for titration and a photoelectric cell for end-point detection. Undisturbed sedimentsamples were collected using a SMBA-type mini-multicorer (MUC), and were processed immediately after col-lection. Cores were sliced continuously in 0.5–1 cm sections using a Plexiglas spatula. Each section was ana-lyzed for the following parameters:
(i) bulk sediment density, estimated from mass percent water and density of solids (2.65 g cm�3); (ii) organicmatter content, estimated as percent weight loss after ignition at 550 �C in a muffle furnace (%LOI); (iii) totalorganic carbon (TOC) and total nitrogen (TN), determined after acidification with 1 N HCl (to remove car-bonates) using a Heraeus-CHN elemental analyzer (molar C/N ratios were also calculated); (iv) chlorophyll-a
Fig. 1. Study area showing the position of Station 18, on the shelf off Concepcion, Chile.
588 P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602

Author's personal copy
(Chl-a) and phaeopigments (Phaeo), analyzed fluorometrically following the methodology suggested by Gut-ierrez (2000); (v) dissolved sulfide, from sediment porewater extracted from 1- and 2-cm sediment slices (downto 15 cm) under a N2 atmosphere, using a pneumatic squeezer. Sulfide was determined colorimetrically at670 nm using the methylene blue technique (Cline, 1969); and (vi) 210Pb activities, assessed mainly by alphaspectrometry of its daughter, 210Po using 209Po as a yield tracer (Flynn, 1968). The 210Po activity was countedby a CANBERRA QUAD alpha spectrometer, model 7404, requiring 24–48 h to achieve the desired countingstatistics (4–10% 1r errors). The activity counting was performed at the Departamento de Oceanografıa, Uni-versidad de Concepcion, Chile. Six samples were counted a second time at the Universite de Montpellier 2,France, in a low background alpha counter. No differences were found between the two counters.
The activity of 210Po, assumed to be in secular equilibrium with 210Pb, was calculated using the ratiobetween the natural radionuclide and the tracer, which is multiplied by the activity of the tracer at the timeof plating. All data were corrected to the time of plating, considering the decay of 210Po (half life: 138 days)that occurred between plating and counting. Because of the short period elapsed between the collection dateand the time of the sample analyses (<6 months) compared to the half life of 210Pb (22.3 years), the calculatedactivities were not corrected for this delay.
The accuracy (5.4%) and precision (0.5%) of the radionuclide analysis were determined using IAEA 326reference material. The certified value was 53.3 Bq kg�1 (range: 48.8–57.8) and the obtained value was53.01 ± 2.85 Bq kg�1. The precision was estimated based on three replicates.
For macrofauna species composition, density, and biomass assessments, four independent replicates weretaken at each sampling (i.e., four independent MUC drops, with each replicate consisting of two pooled coresfrom the same drop; total area 0.0142 m2). In order to study the vertical distribution of the fauna, cores weresliced at 0–2, 2–5, 5–10, 10–15, and 15–20 cm. Sub-samples were then preserved in 4% buffered formalin untilfurther analysis. Once in the laboratory, macrobenthic animals (>0.5 mm) were sorted to the lowest possibletaxon under a low power stereo/microscope; biomass was evaluated in terms of wet weight with a precision of0.1 mg.
2.3. Inventories and fluxes, data analysis
210Pbxs, reactive Chl-a, total pigments, and TOC contents were calculated for the upper 15 cm of each core.In anoxic environments, a large fraction of Chl-a escapes degradation on a temporal scale from weeks tomonths (Sun et al., 1993); therefore, its concentration approaches a constant value with depth in the sediment,but never reaches zero. Thus, reactive Chl-a was estimated by subtracting the non-reactive concentration (i.e.,the lower and constant values in depth) from the total Chl-a measured. A previous study in the same areaindicates that the non-reactive Chl-a value is found between 6 and 10 cm depth (Gutierrez, 2000).
210Pbxs activities (unsupported) were determined from the total 210Pb minus the asymptotic activity at depth(1.5 ± 0.1 dpm g�1), assumed to be in equilibrium with 226Ra within the sediment (e.g., McCaffrey and Thom-son, 1980; Cochran et al., 1998). This activity is similar to 226Ra measurements obtained previously for thesame area (1.1 ± 0.2 dpm g�1; Munoz et al., 2004a). Standard deviations (SD) of 210Pb inventories were esti-mated by propagating the counting uncertainties of 210Pb and the error introduced by the estimated supported210Pb (Bevington and Robinson, 1992).
The inventories of 210Pbxs (unsupported dpm cm�3), Chl-a (reactive; lg cm�3), and total pigments (Chl-a + Phaeo; reactive fraction) were calculated integrating the concentrations in the first 5 cm of the sedimentarycolumn, where the maximum variations in the vertical distribution of the Chl-a occurred. According to pre-vious reports, this is caused by seasonal fluctuations of primary production in the column water (Gutierrez,2000), whose export to the seafloor forms a flocculent layer that could vary between 2 and 5 cm in thickness(Sellanes, unpublished observation). It is in the first 5 cm that we expect to find the largest variations in 210Pbfluxes associated with Chl-a fluxes in 1 year. In this way, the variability of the 210Pb activity by radioactivedecay (longer mean life) is greatly diminished and the changes in the fluxes can then be associated with thereactive Chl-a fluxes. TOC and total pigment inventories were also estimated. The fluxes at the sediment sur-face, sustaining the inventories, were estimated following the methodology proposed by Turekian et al. (1980):
J ¼ I � k;
P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602 589

Author's personal copy
where J is the flux at the surface (dpm 210Pbxs cm�2 yr�1; lg Chl-a cm�2 yr�1), I is the inventory (dpm210Pbxs cm�2; lg Chl-a cm�2; lg total pigments cm�2), and k is the decay constant: 0.031 yr�1 for 210Pb,0.055 d�1 for Chl-a, and 0.075 d�1 for Phaeo. Total pigment fluxes were calculated from Chl-a plus Phaeofluxes. The Chl-a and Phaeo decay constants were determined previously by Gutierrez (2000) at the same sam-pling station (Sta. 18). Organic carbon and sulfide inventories were also calculated using the total concentra-tions (lg g�1; lM) in the sediment volume (g cm�3) or porewater volume (mL cm�3).
2.4. Bioturbation (DB estimates)
The relevance of biological mixing in the inventory estimates was inferred from the bioturbation coefficient(DB). DB was estimated from the Chl-a (reactive) vertical distribution in the first 15 cm of the sediment col-umn, using the algorithms of the program Xsmixing developed by Soetaert et al. (1996). This program usesthe Levenberg–Marquardt method to adjust non-linear models. The program consists of a family of models(1, 2, 3, 4a, 4b, 5) evaluated in hierarchical order from the simplest (no mixing, Model 1) to the more complexmodels (models 2–5). Model 1 considers no mixing by organisms or other processes, explaining the verticalchanges of the tracer only by sedimentation. Model 2 includes diffusive mixing but no non-local exchange.The remaining models contain more parameters in order to explain the tracer distribution by non-localexchange; for example, Model 3 considers that part of the flux is injected at a specific depth (L) and Model4a establishes that this injection of particles is distributed in a layer with certain a width (L ± dx). From Model2 to the more complex models, the sedimentation rate is considered to be insignificant with respect to the meanlife of the tracer.
Although the increased complexity of the models implies a better visual fit between the predicted and theobserved data, after a certain point, even more complex models could not give a better fit to the tracer distri-bution. Therefore, the program uses a one-tailed F-test to choose the best model. In some cases, when only afew data were available for the F-test, the first model with a reasonable r2 was selected.
3. Results
3.1. Sediment biogeochemistry
Table 1 summarizes surface sediment values (concentration, inventories) for all parameters analyzed. Totalorganic matter content at the sediment surface fluctuated between 16% and 21%, diminishing downcore to val-ues between 14% and 18%, respectively. Maximum values were detected in January (21%) and June (19%). TheC/N ratio was highest in September (11.3) and fluctuated between 8.4 and 9.8 from November 2002 to June2003. In general, the bulk density for the first 5 cm varied between 0.27 and 0.43 g cm�3, and a lower value wasdetected at the surface in January (0.11 g cm�3). Bottom-water dissolved oxygen (DO) concentrations werevery low (<0.5 mL L�1) during most of the study period, except in September 2002 and June 2003, when val-ues were slightly higher (>0.5 and <1.2 mL L�1; Table 1).
Chl-a at the surface peaked in June 2003 (49.15 lg g�1); however, the highest Phaeo concentrations andinventories occurred in March (766.27 lg g�1 and 265.7 lg cm�2). Maximum TOC inventories were observedin January and March 2003 (72.4 and 76.5 mg C cm�2). Sulfide inventories were only calculated for 2003, andthe highest value occurred in March (10.70 mmol m�2).
Surface 210Pb activity showed slightly higher values in September 2002, April 2003, and August 2003(between 15.42 and 17.15 dpm g�1) compared to the remaining months (between 9.91 and 11.77 dpm g�1),except in June 2003, when the lowest value was observed (7.95 dpm g�1) coinciding with the highest Chl-a con-centration. Maximum 210Pbxs inventories occurred in June and August 2003 (23.8 and 29.6 dpm cm�2, respec-tively), and the lowest occurred in March 2003 (11.0 dpm cm�2), coinciding with the maximum Phaeoconcentrations. The total 210Pbxs inventories obtained from selected cores were 68.5 (January 2003) and77.1 dpm cm�2 (June 2003), sustained by fluxes of 2.1 and 2.4 dpm cm�2 yr�1, respectively. Both cores arecomparable and showed similar 210Pb inventories at the surface (0–5 cm). In general, the inventories werehigher than 25.7 dpm cm�2, except in November 2002, March, and April 2003, when values were lower,increasing the standard deviation of the mean annual inventory to about 22% (Table 1).
590 P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602
Author's personal copy
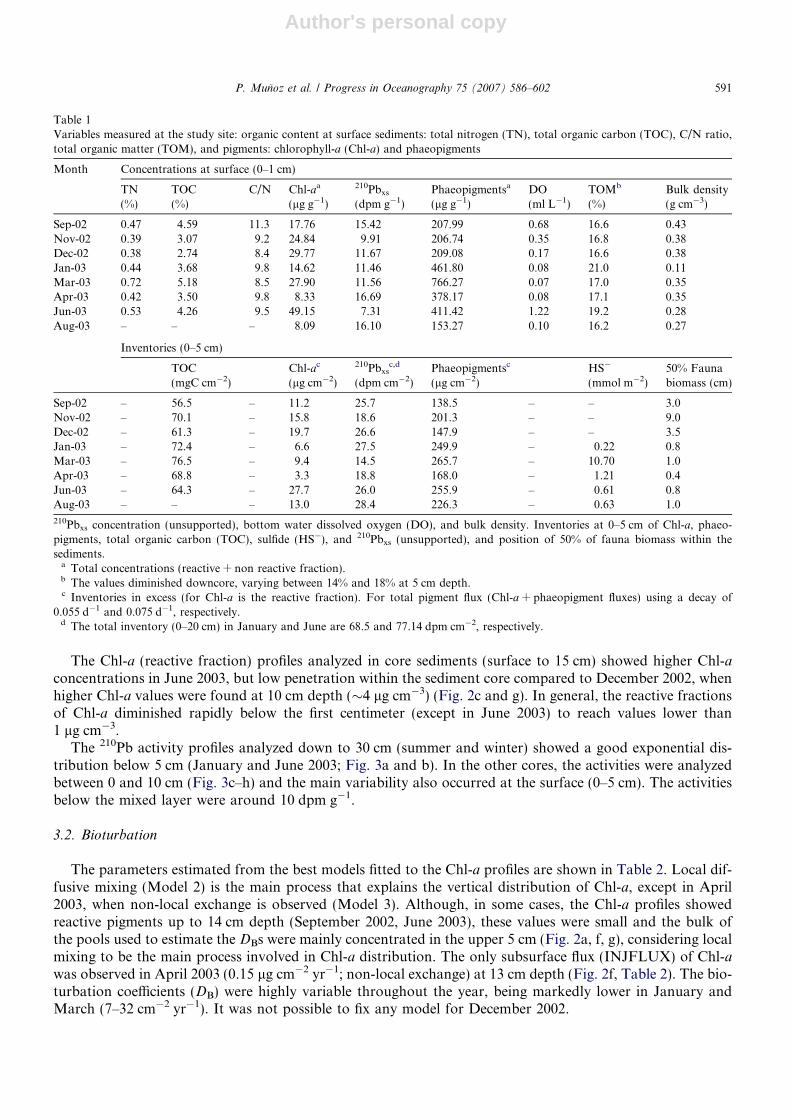
The Chl-a (reactive fraction) profiles analyzed in core sediments (surface to 15 cm) showed higher Chl-aconcentrations in June 2003, but low penetration within the sediment core compared to December 2002, whenhigher Chl-a values were found at 10 cm depth (�4 lg cm�3) (Fig. 2c and g). In general, the reactive fractionsof Chl-a diminished rapidly below the first centimeter (except in June 2003) to reach values lower than1 lg cm�3.
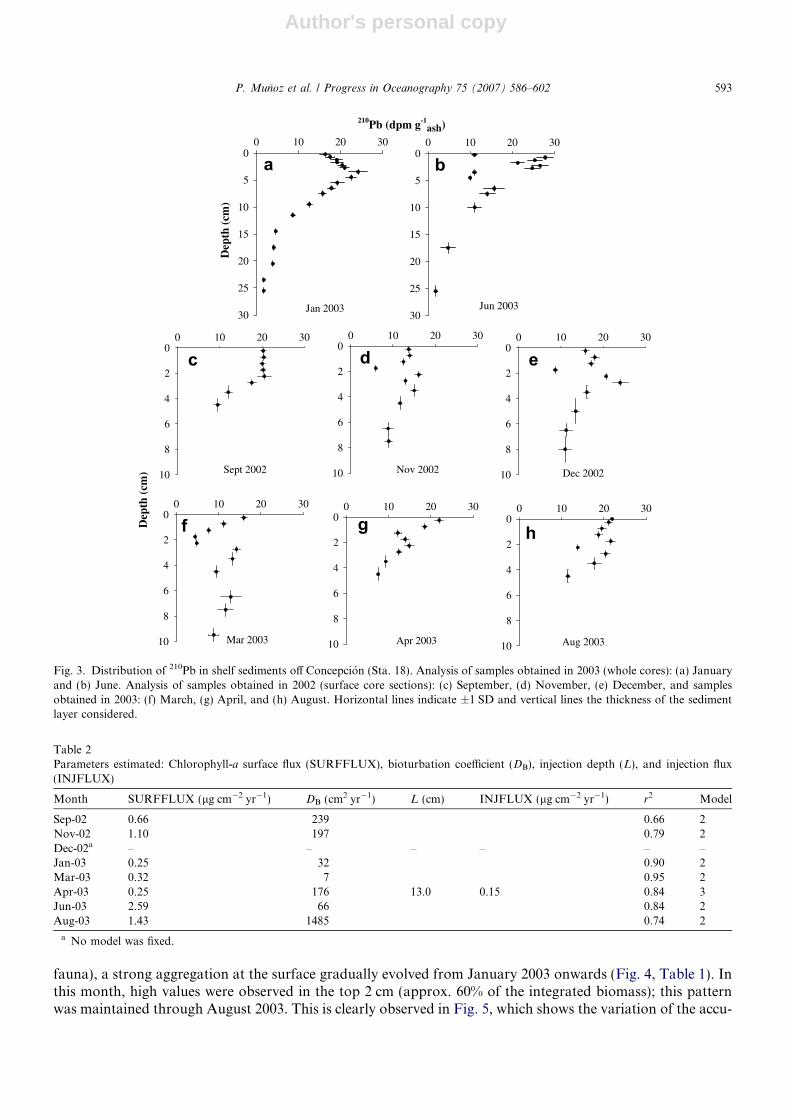
The 210Pb activity profiles analyzed down to 30 cm (summer and winter) showed a good exponential dis-tribution below 5 cm (January and June 2003; Fig. 3a and b). In the other cores, the activities were analyzedbetween 0 and 10 cm (Fig. 3c–h) and the main variability also occurred at the surface (0–5 cm). The activitiesbelow the mixed layer were around 10 dpm g�1.
3.2. Bioturbation
The parameters estimated from the best models fitted to the Chl-a profiles are shown in Table 2. Local dif-fusive mixing (Model 2) is the main process that explains the vertical distribution of Chl-a, except in April2003, when non-local exchange is observed (Model 3). Although, in some cases, the Chl-a profiles showedreactive pigments up to 14 cm depth (September 2002, June 2003), these values were small and the bulk ofthe pools used to estimate the DBs were mainly concentrated in the upper 5 cm (Fig. 2a, f, g), considering localmixing to be the main process involved in Chl-a distribution. The only subsurface flux (INJFLUX) of Chl-awas observed in April 2003 (0.15 lg cm�2 yr�1; non-local exchange) at 13 cm depth (Fig. 2f, Table 2). The bio-turbation coefficients (DB) were highly variable throughout the year, being markedly lower in January andMarch (7–32 cm�2 yr�1). It was not possible to fix any model for December 2002.
Table 1Variables measured at the study site: organic content at surface sediments: total nitrogen (TN), total organic carbon (TOC), C/N ratio,total organic matter (TOM), and pigments: chlorophyll-a (Chl-a) and phaeopigments
Month Concentrations at surface (0–1 cm)
TN(%)
TOC(%)
C/N Chl-aa
(lg g�1)
210Pbxs
(dpm g�1)Phaeopigmentsa
(lg g�1)DO(ml L�1)
TOMb
(%)Bulk density(g cm�3)
Sep-02 0.47 4.59 11.3 17.76 15.42 207.99 0.68 16.6 0.43Nov-02 0.39 3.07 9.2 24.84 9.91 206.74 0.35 16.8 0.38Dec-02 0.38 2.74 8.4 29.77 11.67 209.08 0.17 16.6 0.38Jan-03 0.44 3.68 9.8 14.62 11.46 461.80 0.08 21.0 0.11Mar-03 0.72 5.18 8.5 27.90 11.56 766.27 0.07 17.0 0.35Apr-03 0.42 3.50 9.8 8.33 16.69 378.17 0.08 17.1 0.35Jun-03 0.53 4.26 9.5 49.15 7.31 411.42 1.22 19.2 0.28Aug-03 – – – 8.09 16.10 153.27 0.10 16.2 0.27
Inventories (0–5 cm)
TOC(mgC cm�2)
Chl-ac
(lg cm�2)
210Pbxsc,d
(dpm cm�2)Phaeopigmentsc
(lg cm�2)HS�
(mmol m�2)50% Faunabiomass (cm)
Sep-02 – 56.5 – 11.2 25.7 138.5 – – 3.0Nov-02 – 70.1 – 15.8 18.6 201.3 – – 9.0Dec-02 – 61.3 – 19.7 26.6 147.9 – – 3.5Jan-03 – 72.4 – 6.6 27.5 249.9 – 0.22 0.8Mar-03 – 76.5 – 9.4 14.5 265.7 – 10.70 1.0Apr-03 – 68.8 – 3.3 18.8 168.0 – 1.21 0.4Jun-03 – 64.3 – 27.7 26.0 255.9 – 0.61 0.8Aug-03 – – – 13.0 28.4 226.3 – 0.63 1.0
210Pbxs concentration (unsupported), bottom water dissolved oxygen (DO), and bulk density. Inventories at 0–5 cm of Chl-a, phaeo-pigments, total organic carbon (TOC), sulfide (HS�), and 210Pbxs (unsupported), and position of 50% of fauna biomass within thesediments.
a Total concentrations (reactive + non reactive fraction).b The values diminished downcore, varying between 14% and 18% at 5 cm depth.c Inventories in excess (for Chl-a is the reactive fraction). For total pigment flux (Chl-a + phaeopigment fluxes) using a decay of
0.055 d�1 and 0.075 d�1, respectively.d The total inventory (0–20 cm) in January and June are 68.5 and 77.14 dpm cm�2, respectively.
P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602 591

Author's personal copy
3.3. Benthic fauna
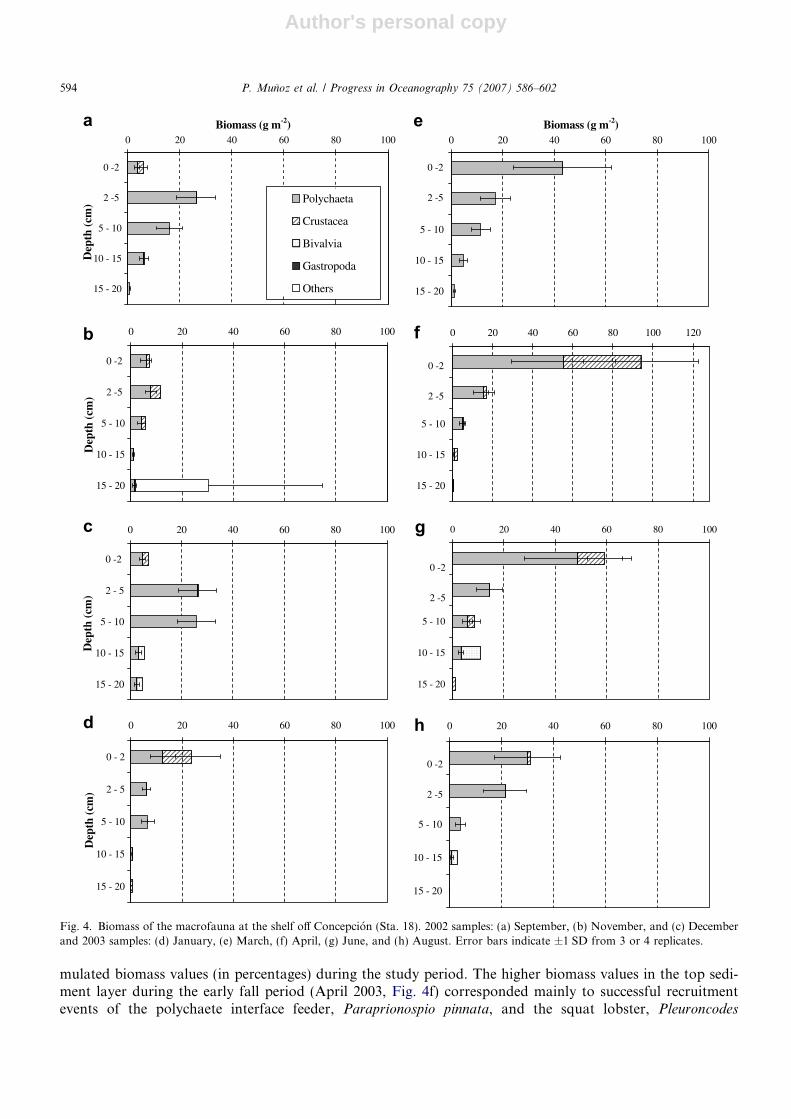
The vertical distribution of macrofaunal biomass is shown in Fig. 4. Total biomass, integrated for the upper20 cm, ranged from 37.51 ± 28.43 g m�2 (mean ± SD) in January 2003 to 94.72 ± 42.56 g m�2 in April 2003.In general, lower values were observed during the first half of the study period (September 2002–January2003), with an increasing trend towards the late summer/fall 2003.
Variations in the vertical distribution pattern showed that, in general, the months of lower biomasses coin-cided with a deeper penetration of the fauna, i.e., higher values in the 2–5 and 5–10 cm levels. Thus, whereasthe most even distribution in depth was seen from September to December 2002 (deeper penetration of the
0 2 4 6 80
2
4
6
8
10
12
14
16
Chl-a (μg cm-3) 0 2 4 6 8
0
2
4
6
8
10
12
14
16
0 2 4 6 80
2
4
6
8
10
12
14
16
0 2 4 6 80
2
4
6
8
10
12
14
16
0 2 4 6 8Dep
th (
cm)
0
2
4
6
8
10
12
14
16
0 2 4 6 80
2
4
6
8
10
12
14
16
0 5 10 15 20 250
2
4
6
8
10
12
14
16
0 2 4 6 80
2
4
6
8
10
12
14
16
a
c d
e f
g h
b
Fig. 2. Distribution of chlorophyll-a (reactive) in shelf sediments off Concepcion (Sta. 18). 2002 samples: (a) September, (b) November,and (c) December; 2003 samples: (d) January, (e) March, (f) April, (g) June, and (h) August. Dotted line indicates the predicted data fromthe best mixing model. No model was fixed in December. Horizontal lines indicate ±1 SD and vertical lines the thickness of the sedimentlayer considered.
592 P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602
Author's personal copy
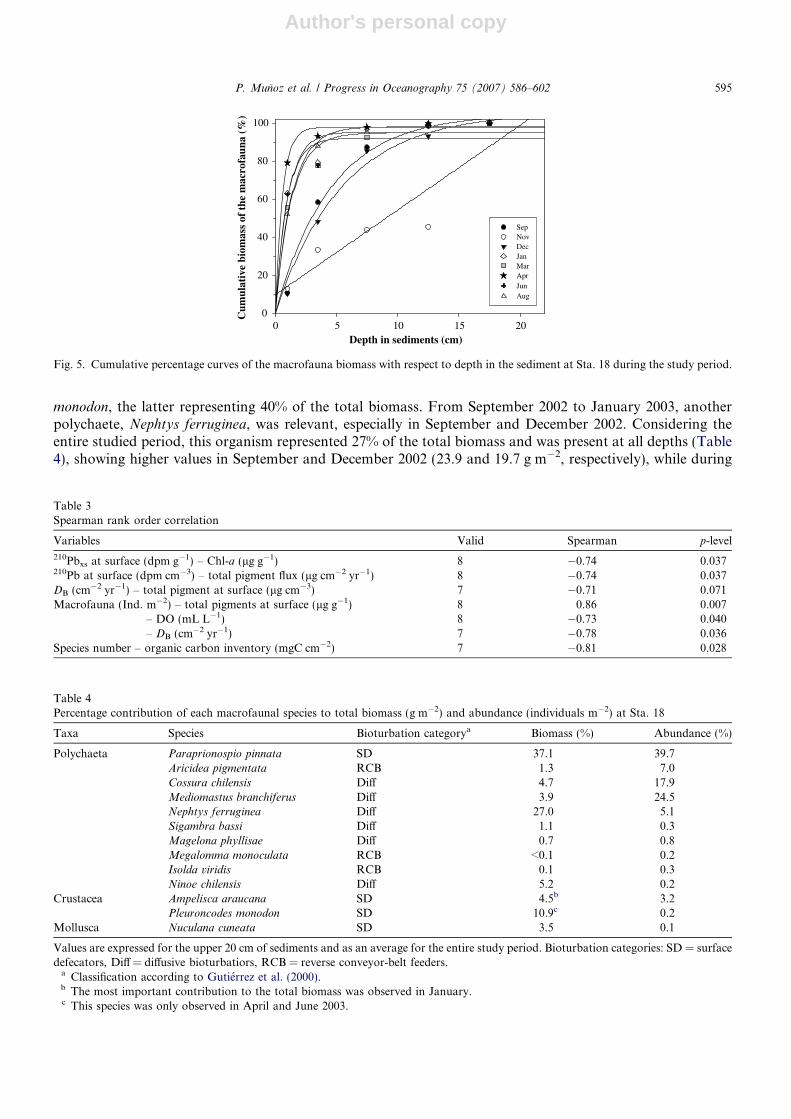
fauna), a strong aggregation at the surface gradually evolved from January 2003 onwards (Fig. 4, Table 1). Inthis month, high values were observed in the top 2 cm (approx. 60% of the integrated biomass); this patternwas maintained through August 2003. This is clearly observed in Fig. 5, which shows the variation of the accu-
0 10 20 30
Dep
th (
cm)
0
2
4
6
8
10 Sept 2002
210Pb (dpm g-1ash)
0 10 20 300
2
4
6
8
10 Nov 2002
0 10 20 300
2
4
6
8
10 Dec 2002
0 10 20 30
Dep
th (
cm)
0
5
10
15
20
25
30Jan 2003
0 10 20 300
2
4
6
8
10 Mar 2003
0 10 20 300
2
4
6
8
10 Apr 2003
0 10 20 300
5
10
15
20
25
30Jun 2003
0 10 20 300
2
4
6
8
10 Aug 2003
a b
c d e
hgf
Fig. 3. Distribution of 210Pb in shelf sediments off Concepcion (Sta. 18). Analysis of samples obtained in 2003 (whole cores): (a) Januaryand (b) June. Analysis of samples obtained in 2002 (surface core sections): (c) September, (d) November, (e) December, and samplesobtained in 2003: (f) March, (g) April, and (h) August. Horizontal lines indicate ±1 SD and vertical lines the thickness of the sedimentlayer considered.
Table 2Parameters estimated: Chlorophyll-a surface flux (SURFFLUX), bioturbation coefficient (DB), injection depth (L), and injection flux(INJFLUX)
Month SURFFLUX (lg cm�2 yr�1) DB (cm2 yr�1) L (cm) INJFLUX (lg cm�2 yr�1) r2 Model
Sep-02 0.66 239 0.66 2Nov-02 1.10 197 0.79 2Dec-02a – – – – – –Jan-03 0.25 32 0.90 2Mar-03 0.32 7 0.95 2Apr-03 0.25 176 13.0 0.15 0.84 3Jun-03 2.59 66 0.84 2Aug-03 1.43 1485 0.74 2
a No model was fixed.
P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602 593
Author's personal copy
mulated biomass values (in percentages) during the study period. The higher biomass values in the top sedi-ment layer during the early fall period (April 2003, Fig. 4f) corresponded mainly to successful recruitmentevents of the polychaete interface feeder, Paraprionospio pinnata, and the squat lobster, Pleuroncodes
0 20 40 60 80 100
0 -2
2 -5
5 - 10
10 - 15
15 - 20
Dep
th (
cm)
Biomass (g m-2) Biomass (g m-2)
Polychaeta
Crustacea
Bivalvia
Gastropoda
Others
0 20 40 60 80 100
0 -2
2 -5
5 - 10
10 - 15
15 - 20
Dep
th (
cm)
0 20 40 60 80 100
5 - 10
10 - 15
15 - 20
0 20 40 60 80 100 120
5 - 10
10 - 15
15 - 20
0 20 40 60 80 100
0 -2
2 - 5
5 - 10
10 - 15
15 - 20
Dep
th (
cm)
0 20 40 60 80 100
0 - 2
2 - 5
5 - 10
10 - 15
15 - 20
Dep
th (
cm)
0 20 40 60 80 100
5 - 10
10 - 15
15 - 20
0 20 40 60 80 100
0 -2
2 -5
5 - 10
10 - 15
15 - 20
0 -2
2 -5
0 -2
2 -5
0 -2
2 -5
a
b f
e
c g
d h
Fig. 4. Biomass of the macrofauna at the shelf off Concepcion (Sta. 18). 2002 samples: (a) September, (b) November, and (c) Decemberand 2003 samples: (d) January, (e) March, (f) April, (g) June, and (h) August. Error bars indicate ±1 SD from 3 or 4 replicates.
594 P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602
Author's personal copy
monodon, the latter representing 40% of the total biomass. From September 2002 to January 2003, anotherpolychaete, Nephtys ferruginea, was relevant, especially in September and December 2002. Considering theentire studied period, this organism represented 27% of the total biomass and was present at all depths (Table4), showing higher values in September and December 2002 (23.9 and 19.7 g m�2, respectively), while during
Depth in sediments (cm)0 5 10 15 20
Cum
ulat
ive
biom
ass
of t
he m
acro
faun
a (%
)
0
20
40
60
80
100
SepNov Dec JanMar Apr Jun Aug
Fig. 5. Cumulative percentage curves of the macrofauna biomass with respect to depth in the sediment at Sta. 18 during the study period.
Table 3Spearman rank order correlation
Variables Valid Spearman p-level
210Pbxs at surface (dpm g�1) – Chl-a (lg g�1) 8 �0.74 0.037210Pb at surface (dpm cm�3) – total pigment flux (lg cm�2 yr�1) 8 �0.74 0.037DB (cm�2 yr�1) – total pigment at surface (lg cm�3) 7 �0.71 0.071Macrofauna (Ind. m�2) – total pigments at surface (lg g�1) 8 0.86 0.007
– DO (mL L�1) 8 �0.73 0.040– DB (cm�2 yr�1) 7 �0.78 0.036
Species number – organic carbon inventory (mgC cm�2) 7 �0.81 0.028
Table 4Percentage contribution of each macrofaunal species to total biomass (g m�2) and abundance (individuals m�2) at Sta. 18
Taxa Species Bioturbation categorya Biomass (%) Abundance (%)
Polychaeta Paraprionospio pinnata SD 37.1 39.7Aricidea pigmentata RCB 1.3 7.0Cossura chilensis Diff 4.7 17.9Mediomastus branchiferus Diff 3.9 24.5Nephtys ferruginea Diff 27.0 5.1Sigambra bassi Diff 1.1 0.3Magelona phyllisae Diff 0.7 0.8Megalomma monoculata RCB <0.1 0.2Isolda viridis RCB 0.1 0.3Ninoe chilensis Diff 5.2 0.2
Crustacea Ampelisca araucana SD 4.5b 3.2Pleuroncodes monodon SD 10.9c 0.2
Mollusca Nuculana cuneata SD 3.5 0.1
Values are expressed for the upper 20 cm of sediments and as an average for the entire study period. Bioturbation categories: SD = surfacedefecators, Diff = diffusive bioturbatiors, RCB = reverse conveyor-belt feeders.
a Classification according to Gutierrez et al. (2000).b The most important contribution to the total biomass was observed in January.c This species was only observed in April and June 2003.
P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602 595

Author's personal copy
the following months values between 7.0 and 14.6 g m�2 were found. In January 2003, Ampelisca araucana
represented �50% of the biomass at the sediment surface (0–2 cm), being either not important or absent inother months (Fig. 4d).
3.4. Correlations between environmental and biological variables
Spearman rank order correlations were used to correlate all variables. No relationships were found betweenthe 210Pbxs fluxes or inventories with Chl-a or total pigment fluxes and/or inventories (reactive), even whenusing total Chl-a and pigment inventories (reactive + non reactive). The best relationships were found betweensurface content of 210Pbxs, Chl-a (reactive), and organic carbon, which showed negative correlations (Table 3).Furthermore, the macrofauna correlated negatively with DO, DB, and the organic carbon inventory except fortotal pigments at the surface. The variability observed in the 210Pb fluxes was thus explained by seasonalchanges occurring mainly at the sediment surface, but when inventories were considered, no relationships wereobserved. Therefore, the variability of the 210Pb fluxes did not reflect changes on a greater time scale and wascontrolled mainly by seasonal processes.
4. Discussion
4.1. Sediment biogeochemistry
The fluxes of organic particles to the seafloor in the study area are clearly dominated by the seasonal cycleof primary productivity, which peaks in spring–summer as a result of the upwelling of saltier, nutrient-rich,oxygen-poor Equatorial Subsurface Waters (ESSW) into the euphotic zone, injecting nutrients into the sys-tem. In the surface sediments, Chl-a content is highest at the beginning of the summer season (December2002) and phaeopigments, resulting from the Chl-a degradation, are highest at the end of summer (March,Table 1). High organic particle sedimentation toward the seafloor and high remineralization rates at the sed-iment–water interface promote oxygen-deficient conditions in the sediments and overlying bottom water(Henrichs and Farrington, 1984). Usually an oxygen minimum zone (<0.5 mL L�1) associated with the ESSWis present at depths from 50 to 250 m in Concepcion Bay and over the adjacent shelf (Ahumada et al., 1983;Fossing et al., 1995; Thamdrup and Canfield, 1996; Schulz et al., 2000; Sellanes et al., 2006, 2007). This featureappears in early spring and usually disappears during winter. During our year of observations, the lowest bot-tom water DO and the maximum concentrations of TN and TOC in the surface sediments were observed atthe end of the summer. This is explained by the settling of particles derived from primary production, whichreach maximum concentrations in summer (from January to March), simultaneous with the highest total pig-ment inventories in the sediments (Table 1) and high Chl-a content in the water column (�600 mg m�2; Pan-toja, pers. comm.).
On the other hand, maximum sulfide concentrations (in March) are generated by the anaerobic degradationof organic material, a process that has been described for the Concepcion Bay and shelf area, where sulfatereduction and denitrification are the main pathways for organic matter remineralization (see Munoz et al.,2004b and references therein). The anaerobic environment and presence of sulfides favor the formation ofinsoluble metal sulfides, trapping the 210Pb within the sediments; however, 210Pb could also be associated withMn and Fe. These elements, which are under oxidation-reduction processes, establish soluble and insolublephases that determine the adsorption or desorption of 210Pb from the solid phase, changing its distributionwithin the sediments (Todd et al., 1986; Wei and Murray, 1994). At present, the solubilization of 210Pb asso-ciated with the oxide-reduction process has only been described for lake sediments and has not been reportedin marine environments. On the contrary, Henderson and Maier-Reimer (2002) state that an irreversible scav-enging onto settling biogenic particles is the best explanation for the sustained mass balance of 210Pb in theglobal ocean.
In this study, we observed that the highest sulfide inventory (March 2003) coincided with the lowest 210Pbinventory (Table 1), but we cannot establish a clear relationship between these parameters due to a gap in theinformation as to sulfide concentrations during the first months of the time series. Therefore, this observationdeserves further attention in future studies (Table 1).
596 P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602

Author's personal copy
4.2. Bioturbation
According to the time scale considered in this study (1 year), we used Chl-a (reactive) profiles to estimatemixing in the sediments. Other elements such as 234Th and 239,240Pu have been used for evaluating mixing inthe Concepcion Bay and continental shelf sediments. DB estimates from this study were similar to thosereported earlier in the area (47–187 cm2 yr�1; Salamanca, 1989; Gutierrez, 2000). These previous values weremeasured during and after the 1997–1998 El Nino event, also showing high variability. The enhanced dis-solved oxygen content in the bottom waters during El Nino promotes the occurrence of higher bioturbationcoefficients by favoring an increase of bioturbating species as surface-subsurface deposit feeders (diffusivebioturbators; Gutierrez et al., 2000; Sellanes et al., 2007). The macrofauna abundance showed a direct cor-relation with the pigment concentrations in the surface sediments and the highest pigment concentrationswere, in turn, related to lower DB coefficients (negative correlation; Table 3). This is explained as follows:although high pigment and organic carbon inputs are related to high fauna abundance, the compositionincludes few small body-sized species that concentrate in the surface sediments (up to 2–3 cm) and thatdo not produce important mixing at deeper sections. Increased primary productivity during the spring–sum-mer period favors colonization by opportunistic polychaetes. These small worms (e.g., Paraprionospio pin-
nata) are best adapted to low oxygen concentrations, since they have a complete set of enzymes to conductanaerobic metabolism (Gonzalez and Quinones, 2000). P. pinnata has been previously recognized as thedominant species in Concepcion Bay and shelf sediments (see Carrasco and Gallardo, 1994 and referencestherein); it is present in anoxic muddy sediments, being responsible for the bioturbation of the upper 2 cm.However, since these organisms are interface feeders or tube-dwelling surface-defecators, they are consideredweak bioturbators (Gutierrez et al., 2000). Although the bioturbation coefficients (DB) estimated in thisstudy from Chl-a distribution suggest that mixing can extend beyond 5 cm (depending on fauna distribu-tion), the maximum mixing occurs in surface sediments, where �50% of the total fauna is present (0–2 cm), except in November (Table 1, Fig. 5).
Thus, the DB coefficients did not show a clear relationship with the biomass of the fauna, but correlatednegatively with its density. Negative correlations of the DB coefficients were also observed with organic carbonand pigment content at the surface (Table 3). Other studies (Boon and Duineveld, 1998; Yeager et al., 2004)indicate both positive and negative correlations between Chl-a fluxes and bioturbation. The above authorsargue that the relationship between organic matter input and biological mixing is not straightforward and thatother parameters need to be considered. These parameters are the organic matter degradation rate and thebiomass and/or abundance of macrobenthic organisms which, in turn, are also affected by the organic carbonquality and oxygen concentration (Dıaz and Rosenberg, 1995; Gutierrez et al., 2000; Sellanes et al., 2006,2007). In our case, the DBs did not show a clear relationship with oxygen concentrations, as it did with themacrofaunal abundance and total pigment concentrations in surface sediments. Although the seasonal varia-tion of dissolved oxygen during the study period is not as drastic as during El Nino conditions, a macrofaunalresponse to environmental changes is also noticeable. During El Nino conditions, bottom water dissolved oxy-gen increases, promoting higher faunal activity. After El Nino, when oxygen deficient conditions are re-estab-lished, a decreasing trend of the bioturbation potential has been reported, due mainly to the dominance ofsurface defecators (Gutierrez et al., 2000 and references therein). In our case, the faunal dominance changesfrom diffusive bioturbators to surface defecators during very low oxygen concentrations (between January toApril 2003), but the relationship of bioturbation with oxygen concentrations is not as direct. The bioturbationpotential seems to respond mainly to the organic fluxes (Table 3), which also affect oxygenation as a conse-quence of organic matter degradation. The macrofauna respond to these changes, and it seems that the thresh-old at which increases of bottom water dissolved oxygen trigger changes in the macrofaunal composition isvery low.
In summary, an increase in organic carbon (or total pigments) in the surface sediments, as is the case duringthe productive spring–summer period, results in a concentration of faunal activity at the surface and, hence, adrop in DBs. During lower organic fluxes, the fauna deepen in the sediment, and thus the DBs increase. Thesechanges result in highly variable bioturbation coefficients during the year (7–1485 cm2 yr�1; this study), of thesame order of magnitude as those measured during El Nino (286.0–59.6 cm2 yr�1) and non El Nino conditions(96.9–5.3cm2 yr�1; Gutierrez, 2000). This would suggest that the intensity of the response of the benthic fauna
P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602 597

Author's personal copy
to large-scale phenomena like El Nino is on the order of the one observed during the seasonal cycle of theorganic fluxes to the sediments.
It is known that benthic organisms mix sediment layers and thus modify the decay profiles. For example,bioturbation tends to produce upper sediment layers with similar or homogeneous concentrations, thusincreasing the inventories (Boudreau, 1997); therefore, a positive correlation of the DBs and fauna biomass(or abundance) should be observed (Yeager et al., 2004). However, in this study, no relationship was observedbetween DBs and 210Pb inventories nor between fauna biomass (or abundance) and 210Pb inventories. Theabundance and biomass of the fauna decreased gradually with depth and the 210Pb activity distribution didnot show large homogeneous layers (2–3 cm) in comparison with the Chl-a distribution (4–14 cm). We believethat this pattern could be related to a faunal preference for organic-rich particles. Although 210Pb associateswith both inorganic and organic particles, the benthic fauna is probably selectively working with particlesenriched in organic material and the mixing of these particles could be intensified. As fresh organic matter(high organic carbon quality) is deposited at the sediment surface, the fauna will work up these particles fasterthan the deeper and older ones, precluding the generation of comparable homogeneous layers of 210Pb as withthe Chl-a distribution, which seems to be altered on a different scale.
The DBs estimated from the Chl-a profiles are explained satisfactorily by local mixing as the dominant pro-cess in 75% of the cases, and a higher subsurface flux (non-local mixing) was observed only in April 2003 (sub-surface flux is observed at 13 cm depth). In this case, it must be considered that the concentration of reactiveChl-a (in excess) that reaches deeper sections within the sediments is very low, showing a weak penetration offresh pigments in the sediments (Fig. 2, Table 2). Therefore, the greatest intensity of biological activity isobserved in the first 3–5 cm, and probably is the main source for the variability of the 210Pb inventories; how-ever, no significant correlation between 210Pb inventories and DB was established (range: 14.5–28.4 dpm cm�2;Tables 1 and 3).
4.3. 210Pb fluxes
The range of 210Pbxs fluxes estimated in the first 5 cm (0.45–0.88 dpm cm�2 yr�1) could be explained, inpart, by the atmospheric inputs reported for the area (0.27–0.33 dpm cm�2 yr�1; Salamanca, 1993; Munozand Salamanca, 2003). However, higher fluxes observed during this study suggest that other sources couldbe contributing �64% of the 210Pb content in the sediments. We explored three possibilities: (i) the additionalintroduction of 210Pb during the upwelling period, (ii) variable scavenging rates, and (iii) resuspension-trans-port over the shelf.
The introduction of 210Pb by the upwelled waters associated with high primary productivity increases 210Pbfluxes from those predicted from solely atmospheric sources. Rapid scavenging of 210Pb onto sinking particleshas been reported for the study area based on the 210Pb/226Ra ratio in the water column (inner shelf: 0.5 andslope: 0.6; Salamanca, 1993). Sedimentation rates in our study area (middle shelf: 0.27 cm yr�1) seem to be asintense as in shallower waters of the inner shelf and Concepcion Bay (ca. 0.3 and 0.2 cm yr�1; Salamanca,1993; Munoz et al., 2004a). This suggests that the introduction of 210Pb by upwelled water is efficientlyremoved by the particles and, hence, its flux to the sediments should be dominated by particle deposition.However, the maximum fluxes of Chl-a (or total pigments), which occurred during the upwelling period,did not coincide with maximum 210Pb fluxes. The phytoplankton flux serves as a carrier for the depositionof 210Pb, but the fluxes of both parameters do not seem to be proportional. The seasonal increase of particleinputs (organic and inorganic) could exceed the 210Pbxs fluxes from the water column, thus diluting the 210Pbactivities in the surface sediments, at least on intra-annual scales. This supports the idea of a constant input of210Pb and its relatively homogeneous distribution over the coastal area which, in turn, is the main assumptionfor the Constant Rate of Supply model (CRS) used to estimate sedimentation rates (Appleby and Oldfield,1978; Cochran et al., 1998). A simple correlation indicates that the 210Pbxs in surface sediments is inverselyrelated with the organic fluxes. Our observations of Chl-a and total pigments, which originate in the phytode-tritus that reaches the bottom, suggest that the organic fluxes are higher than the 210Pbxs fluxes. This has alsobeen observed for 210Po–210Pb distributions in relation to the organic matter fluxes in several Florida lakes(Binford and Brenner, 1986). The authors proposed a dilution model to estimate the sedimentation rate in lakesediments considering the negative relationship between organic carbon and the 210Pb deposited in the surface
598 P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602

Author's personal copy
sediments. Although this model was controversial, since several assumptions still needed to be resolved, thedilution effect of the organic material on the radionuclide deposition was not questioned (Benoit and Hemond,1988).
Resuspension has been proposed as a mechanism to produce high 210Pb inventories (and fluxes) in the sed-iments (Lewis et al., 2002) since it increases the residence time of particles in the water column, where furtheradsorption of the radionuclide could occur. The resuspension could occur on a small scale or it could involvelarger areas, implying that the particles could settle locally or be advected by the currents (Boudreau, 1997;Radakovitch and Heussner, 1999; Dezileau et al., 2000; Dezileau et al., 2004), thereby increasing or decreasingthe radionuclide fluxes in a particular area. A recent study in the area indicates that an intensification of thebottom currents occurs during downwelling periods, which could constitute an important mechanism for sed-iment resuspension over the shelf (Sobarzo and Djurfeldt, 2004). In the study area, the development of floc-culent layers and the presence of Thioploca mats in summer suggest lower resuspension events, but theoccurrence of these events cannot be predicted in winter. Consequently, if the process of sediment resuspen-sion is seasonal or episodic during the year, this could play a role in the intra annual variability of the 210Pbinventories, and it has to be accounted for.
A general estimate of the 210Pb concentration in the suspended particles at the shelf (2.29 dpm g�1; Salam-anca, 1993) measured in springtime (upwelling) and information from a sediment trap located at the mouth ofConcepcion Bay (1.71 g cm�2 yr�1; Munoz and Salamanca, 2001) allow for preliminary calculations of theamount of sediment resuspended (Bacon et al., 1994)
D � Csusp ¼ R � Csedþ k � I
where D is the deposition rate (g cm�2 yr�1), R is the resuspension rate (g cm�2 yr�1); Csusp is the 210Pb con-tent of the suspended particles, Csed is the 210Pb content of the surface sediment (dpm g�1), k is the decayconstant, and I is the 210Pb inventory in the sediment (72.8 dpm cm�2). This allows estimates of the thicknessof the sediment (H) that needs to be resuspended in order to maintain the resuspension rate (R)
H ¼ R=q
where q is the sediment bulk density. In this way, a resuspension of up to 11 mm yr�1 was calculated, whichwould not be enough to explain by itself the variability of the 210Pb inventories, but would contribute with theseasonal variability in the 210Pb concentration at the surface layer (0–1 cm; Table 1).
On the other hand, lateral transport and particle redistribution should be facilitated by the circulation onthe shelf described by Sobarzo and Djurfeldt (2004). At the surface, water is transported offshore and north-ward during the upwelling periods, consistent with the Ekman transport, accompanied by a compensation ofmidwater (50–70 m) on-shore flows. This suggests that the surface circulation could constitute an importantsource of particles to the shelf sediments during the spring–summer season that could increase the 210Pb con-tent in the surface sediments. The bottom currents flow alongshore and southward through the year and donot constitute a compensation flow by changes in the surface water circulation. This current is intensified dur-ing downwelling periods and could transport particles offshore that could diminish the 210Pb content in thesurface sediments. However, the 210Pb activity in this sediment layer did not show a pattern comparable withenrichment during upwelling (on-shore transport) and decreased concentrations during downwelling periods(off-shore transport); on the contrary, the lowest values are observed during upwelling periods. It is likely thatthe midwater on-shore compensation flow could facilitate the recirculation of the fine particles over the shelffor a period before they settle on the sediments. Thus, reworking and redistribution increase the particles’ res-idence time in the water column and, hence, could contribute to their enrichment in 210Pb. The particles couldsettle during any upwelling relaxation or calm period during the year; therefore, the variability of the 210Pbactivity in the surface sediments is difficult to predict with this information alone.
The 210Pb activity at the surface could also be influenced by the Bio Bio river, which produces a flow thatmoves northward during upwelling periods at the southern part of the outer shelf (Sobarzo and Djurfeldt,2004). The river’s influence cannot be established at the inner shelf, but it should be episodic and dependenton the intensity of the upwelling. In spring and at the beginning of the upwelling period (September 2002),higher values of 210Pb were measured at the surface, coincident with higher values of the C/N ratio in surfacesediments (11.3; Table 1). This could be considered to be a signal of a continental source, since shelf sediments
P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602 599

Author's personal copy
usually present values around 8–9, representing a mixing of continental and marine sources. However, this wasnot observed in the next spring (August 2003). Besides, since we do not have 210Pb measurements for this river,we lack evidence to state that this input constitutes a source of 210Pb variability in the surface sediments.
5. Conclusions
1. Bioturbation affects mostly the upper 2–5 cm, showing seasonal variability associated with the organic mat-ter enrichment in the surface sediments. Since the variability of the bottom water dissolved oxygen regimewas small, a clear relationship with bioturbation was not observed. The estimated DBs are on the sameorder as previous values reported during El Nino events, suggesting that the response of benthic faunato large-scale phenomena is on the order of the one observed during the seasonal cycle of the organic fluxesto the sediments.
2. Macrofaunal activity can modify the distribution of the different parameters (e.g., 210Pb, photosyntheticpigments) in the upper sediments, but DBs and 210Pb inventories do not correlate. Therefore, bioturbationalone cannot explain the 210Pb variability in the surface sediments. It is proposed that other processes suchas organic dilution during the upwelling period could modify the 210Pb activities seasonally and, in combi-nation with the resuspension and circulation in the study area, help to redistribute the particles over theshelf.
3. For the area off Concepcion, our time-series study shows that, on short time scales, the complex processesinvolved in the distribution of 210Pb activity in surface sediments cannot be used to obtain accurate accu-mulation rates for other elements associated with carbon fluxes, but this surface variability (0–5 cm) doesnot significantly affect the annual inventories and the flux records can be resolved at least on a decadalscale.
Acknowledgements
We would like to express our gratitude to Dr. Laurent Dezileau from the Laboratoire de Dynamique de laLithosphere (LDL) of the Universite de Montpellier 2, France, who provided the facilities to count some sam-ples in the alpha counter; to Dr. Rodolfo Araya Duran, Departamento de Ingenierıa Matematica, Universi-dad de Concepcion, who helped us run the mixing model; and to all the laboratory technicians from theUniversidad de Concepcion and Universidad Catolica del Norte, who supported us during sample collectionand analysis. Our gratitude is also extended to the L/C Kay-Kay personnel for their valuable assistance duringfield activities. Three external anonymous reviewers helped to improve an early draft of this manuscript.Funding was provided by the FONDAP-COPAS Center and a DGIP Project (10301176) of the UCN grantedto P.M.
References
Ahumada, R., Rudolph, A., Martinez, V., 1983. Circulation and fertility of waters in Concepcion Bay. Estuar. Coast. Shelf Sci. 16, 95–
105.
Appleby, P.G., Oldfield, F., 1978. The calculation of lead-210 dates assuming a constant rate of supply of unsupported 210Pb to the
sediment. Catena 5, 1–8.
Arcos, D.F., Wilson, R.C., 1984. Upwelling and distribution of chlorophyll a within the bay of Concepcion, Chile. Estuar. Coast. Shelf
Sci. 18 (1), 25–35.
Arcos, D.F., Nunez, S.E., Acuna, A., 1996. Variabilidad de pequena escala en la zona nerıtica del sistema de surgencia de Talcahuano
(Chile Central): identificacion y dinamica de areas de retencion larval. Gayana Oceanol. 4 (1), 21–58.
Bacon, M.P., 1988. Tracers of chemical scavenging in the ocean: boundary effects and large scale chemical fractionation. Philos. Trans.
Roy. Acad. Lond. A 325, 147–160.
Bacon, M.P., Belastock, R.A., Bothner, M.H., 1994. 210Pb balance and implications for particle transport on the continental shelf, US
Middle Atlantic Bight. Deep-Sea Res. II 41, 511–535.
Benninger, L.K., Lewis, D.M., Turekian, K.K., 1975. The use of natural 210Pb as a heavy metal tracer in the river-estuarine system. In:
Marine Chemistry in the Coastal Environment, American Chemical Society Symposium Series, vol. 18, pp. 202–210.
Benoit, G., Hemond, H., 1988. Comment on dilution of 210Pb by organic sedimentation in lakes of different trophic states, and application
to studies of sediment–water interactions (Binford and Brenner). Limnol. Oceanogr. 33 (2), 299–304.
600 P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602

Author's personal copy
Bevington, P., Robinson, K., 1992. Error analysis. In: Bevington, P., Robinson, K. (Eds.), Data Reduction and Error Analysis for the
Physical Sciences. WCB/McGraw-Hill, USA, pp. 38–52.
Binford, M., Brenner, M., 1986. Dilution of 210Pb by organic sedimentation in lakes of different trophic states, and application to studies
of sediment–water interactions. Limnol. Oceanogr. 31, 584–595.
Boon, A.R., Duineveld, G.C.A., 1998. Chlorophyll a as a marker for bioturbation and carbon flux in southern and central North Sea
sediments. Mar. Ecol.-Prog. Ser. 162, 33–43.
Boudreau, B., 1997. Model construction. In: Boudreau, B. (Ed.), Diagenetic Models and their Implementation. Springer-Verlag, Berlin,
Germany, pp. 23–90.
Bruland, K.W., Koide, M., Goldberg, E., 1974. The comparative marine geochemistry of Lead 210 and Radium 226. J. Geophys. Res. 79
(21), 3083–3086.
Carrasco, F.D., Gallardo, V.A., 1994. Diversidad, distribucion y abundancia del macrobentos sublitoral, y observaciones sobre la
dinamica temporal de corto termino de los sedimentos de la Bahıa Concepcion, Chile. Gayana Oceanol. 2 (2), 49–68.
Cline, J.D., 1969. Spectrophotometric determination of hydrogen sulphide in natural waters. Limnol. Oceanogr. 14, 454–458.
Cochran, K., 1992. The oceanic chemistry of the U- and Th-series nuclides. In: Ivanovich, M., Harmon, R. (Eds.), Uranium Series
Disequilibrium: Applications to Earth, Marine and Environmental Sciences. Clarendon Press, Oxford, pp. 384–430.
Cochran, J.K., Frignani, M., Salamanca, M., Belluci, L.G., Guerzoni, S., 1998. Lead-210 as a tracer of atmospheric input of heavy metals
in the northern Venice Lagoon. Mar. Chem. 62 (1–2), 15–29.
Crusius, J., Bothner, M., Sommerfield, C., 2004. Bioturbation depths, rates and processes in Massachusetts Bay sediments inferred from
modeling of 210Pb and 239+240Pu profiles. Estuar. Coast. Shelf Sci. 61 (4), 643–655.
Daneri, G., Dellarossa, V., Quinones, R., Jacob, B., Montero, P., Ulloa, O., 2000. Primary production and community respiration in the
Humboldt Current System off Chile and associated oceanic areas. Mar. Ecol.-Prog. Ser. 197, 41–49.
Dezileau, L., Bareille, G., Reyss, J.L., Lemoine, F., 2000. Evidence for strong sediment redistribution by bottom currents along the
Southeast Indian Ridge. Deep-Sea Res. Pt.I. 47, 1899–1936.
Dezileau, L., Ulloa, O., Hebbeln, D., Lamy, F., Reyss, J.L., Fontugne, M., 2004. Andean iron control of past productivity in the coastal
upwelling system off the Atacama desert, Chile. Paleoceanography. 19 PA3012, doi:10.1029/2004PA001006.
Dıaz, R.J., Rosenberg, R., 1995. Marine Benthic hypoxia: a review of its ecological effects and the behavioural responses of benthic
macrofauna. Oceanogr. Mar. Biol. Ann. Rev. 33, 245–303.
Flynn, W.W., 1968. The determination of low levels of polonium-210 in environmental materials. Anal. Chim. Acta 43, 221–227.
Fossing, H., Gallardo, V.A., Jørgensen, B.B., Huttel, M., Nielsen, L.P., Schulz, H.N., Canfield, D.E., Forster, S., Glud, R.N., Gundersen,
J.K., Kuver, J., Ramsing, N.B., Teske, A., Thamdrup, B., Ulloa, O., 1995. Concentration and transport of nitrate by the mat-forming
sulfur bacterium Thioploca. Nature 374, 713–715.
Gonzalez, R., Quinones, R., 2000. Pyruvate oxidoreductases involved in glycolitic anaerobic metabolisms of polychaetes from the
continental shelf off central-south Chile. Estuar. Coast. Shelf Sci. 51, 507–519.
Graco, M., Farıas, L., Molina, V., Gutierrez, D., Nielsen, L.P., 2001. Massive developments of microbial mats following phytoplankton
blooms in a naturally eutrophhic bay: implications for nitrogen cycling. Limnol. Oceanogr. 46 (4), 821–832.
Gutierrez, D., 2000. Bioperturbacion y macrofauna en fondos sublitorales de un area de surgencias frente a Chile Central (36�30 0S):
Variacion espacial y temporal en el perıodo 1997–1999. Ph.D. thesis, Universidad de Concepcion, Casilla 160-C, Concepcion, Chile
(unpublished).
Gutierrez, D., Gallardo, V.A., Mayor, S., Neira, C., Vasquez, C., Sellanes, J., Rivas, M., Soto, A., Carrasco, F., Baltazar, M., 2000.
Effects of dissolved oxygen and fresh organic matter on the bioturbation potential of macrofauna in sublittoral bottoms off central
Chile, during the 1997–98 El Nino. Mar. Ecol.-Prog. Ser. 202, 81–99.
Henderson, G.M., Maier-Reimer, E., 2002. Advection and removal of 210Pb and stable Pb isotopes in the oceans: a general circulation
model study. Geochim. Cosmochim. Acta 66 (2), 257–272.
Henrichs, S.M., Farrington, J.W., 1984. Peru upwelling region sediments near 15�S. Remineralization and accumulation of organic
matter. Limnol. Oceanogr. 62, 1–19.
Levin, L.A., Rathburn, A.E., Gutierrez, D., Munoz, P., Shankle, A., 2003. Bioturbation by symbiont-bearing annelids in near-anoxic
sediments: implications for biofacies models and paleo-oxygen assessments. Palaeogeogr. Palaeoclimatol. Palaeoecol. 199, 129–140.
Lewis, R.C., Coale, K.H., Edwards, B.D., Marot, M., Douglas, J.N., Burton, E.J., 2002. Accumulation rate and mixing of shelf sediments
in the Monterey Bay National Marine Sanctuary. Mar. Geol. 181, 157–169.
McCaffrey, R., Thomson, J.A., 1980. Record of the accumulation of sediment and trace metals in the Connecticut salt marsh. Adv.
Geophys. 22, 165–236.
Moore, W.S., Dymond, J., 1988. Correlation of 210Pb removal with organic carbon fluxes in the Pacific Ocean. Nature 331, 339–341.
Munoz, P., Salamanca, M.A., 2001. Flujo de Pb particulado a los sedimentos marinos y su incorporacion en paraprionospio pinnata
(polychaeta: spionidae), Bahıa Concepcion, Chile. Gayana Oceanol. 65, 155–166.
Munoz, P., Salamanca, M.A., 2003. Input of atmospheric lead to polluted marine sediments of SE-Pacific (36�Lat. S). Mar. Environ. Res.
55 (4), 335–357.
Munoz, P., Lange, C.B., Gutierrez, D., Hebbeln, D., Salamanca, M.A., Dezileau, L., Reyss, J.L., Benninger, L.K., 2004a. Sedimentation
and mass accumulation rates along the Peru–Chile continental margin. Deep-Sea Res. Pt. II 51, 2523–2541.
Munoz, P., Sellanes, J., Salamanca, M., Neira, C., 2004b. Nitrogen sediment fluxes in an upwelling system off central Chile (Concepcion
Bay and adjacent shelf) during the 1997–1998 El Nino. Rev. Chil. Hist. Nat. 77, 305–318.
Nozaki, Y., Turekian, K.K., Von Damm, K., 1980. 210Pb GEOSECS water profiles from the North Pacific. Earth Planet. Sci. Lett. 49,
393–400.
P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602 601

Author's personal copy
Nozaki, Y., Zhang, J., Takeda, A., 1997. 210Pb and 210Po in the equatorial Pacific and the Bering Sea: the effects of biological productivity
and boundary scavenging. Deep-Sea Res. Pt. II 44 (9–10), 2203–2220.
Parra, O., Habit, E., 1998. Documento de sıntesis, estudio de lınea base para la evaluacion de impacto ambiental del Complejo Forestal
Industrial Itata (celulosa Arauco y Constitucion, S.A.). In: Parra, O., Habit, E. (Eds.), Chile: Centro Eula-Chile. Universidad de
Concepcion, Concepcion, Chile.
Pineda, V., 1999. El canon Submarino del Bio-Bıo: Aspectos Dinamicos y Ambientales. Ph.D. thesis. Centro EULA, Universidad de
Concepcion, Concepcion, Chile (unpublished).
Radakovitch, O., Heussner, S., 1999. Fluxes and budget of 210Pb on the continental margin of the Bay of Biscay (Northeastern Atlantic).
Deep-Sea Res. Pt. II 46, 2175–2203.
Salamanca, M.A., 1989. El efecto de la bioperturbacion en la distribucion del exceso de 210Pb y 239,240Pu en sedimentos de Bahıa
Concepcion. Cienc. Tecnol. Mar., CONA 13, 43–56.
Salamanca, M.A., 1993. Sources and sinks of 210Pb in Concepcion Bay, Chile. Ph.D. dissertation, MSRC, SUNY, USA, 253 pp.
(unpublished).
Scholl, D.W., Christensen, M.N., Von Huene, R., Marlow, M.S., 1970. Peru–Chile trench. sediments and sea-floor spreading. Geol. Soc.
Am. Bull. 81, 1339–1360.
Schulz, H.N., Strotmann, B., Gallardo, V.A., Jørgensen, B.B., 2000. Population study of the filamentous sulfur bacteria Thioploca spp. off
the bay of Concepcion. Mar. Ecol.-Prog. Ser. 200, 117–126.
Sellanes, J., Neira, E., Quiroga, E., 2006. ENSO as a natural experiment to understand environmental control of meiofaunal community
structure. PSZN Mar. Ecol. 27, 31–43.
Sellanes, J., Quiroga, E., Neira, C., 2007. Changes of macrobenthos composition under different ENSO cycle conditions on the continental
shelf off central Chile. Cont. Shelf Res. 27 (7), 1002–1016.
Smoak, J.M., Moore, W.S., Thunell, R.C., 2000. Influence of boundary scavenging and sediment focusing on 234Th, 228Th and 210Pb fluxes
in the Santa Barbara basin. Estuar., Coast. Shelf Sci. 51 (3), 373–384.
Sobarzo, M., 1999. Surgencia costera sobre una plataforma limitada por canones submarinos, Concepcion, Chile central (36�40 0S;
73�15 0W). 236 pp. Doctoral thesis, Universidad de Concepcion, Chile (unpublished).
Sobarzo, M., Djurfeldt, L., 2004. Coastal upwelling process on a continental shelf limited by submarine canyons, Concepcion, central
Chile. J. Geophys. Res. 109, CI2012. doi:10.1029/2004JC002350.
Sobarzo, M., Figueroa, D., Djurfeldt, L., 2001. Upwelling of subsurface water into the rim of the Biobıo submarine canyon as a response
to surface winds. Cont. Shelf Res. 21, 279–299.
Soetaert, K., Herman, P.M.J., Middelburg, J.J., Heip, C., de Stiger, H.S., van Weering, T.C.E., Epping, E., Helder, W., 1996. Modeling
210Pb-derived mixing activity in ocean margin sediments: diffusive versus nonlocal mixing. J. Mar. Res. 54 (6), 1207–1227.
Sun, M.Y., Lee, C., Aller, R.C., 1993. Laboratory studies of oxic and anoxic degradation of chlorophyll a in Long Island Sound
sediments. Geochim. Cosmochim. Acta 57, 147–157.
Thamdrup, B., Canfield, D., 1996. Pathways of carbon oxidation in continental margin sediments off central Chile. Limnol. Oceanogr. 41,
1629–1650.
Thomson, J., Turekian, K., 1976. 210Po and 210Pb distribution in ocean water profiles from the Eastern South Pacific. Earth Planet. Sci.
Lett. 32, 297–303.
Thornburg, T., Kulm, L.D., 1987a. Sedimentation in the Chile Trench: depositional morphologies, lithofacies and stratigraphy. Geol. Soc.
Am. Bull. 98, 33–52.
Thornburg, T., Kulm, L.D., 1987b. Sedimentation in the Chile Trench: petrofacies and provenance. J. Sediment. Petrol. 57, 55–74.
Todd, J.F., Wong, G.T.-F., Reid, D.F., 1986. The geochemistry of 210Po and 210Pb in waters overlying and within the Orca Basin, Gulf of
Mexico. Deep-Sea Res. 33, 1293–1306.
Turekian, K., Cochran, K., Benninger, L., Aller, R., 1980. The sources and sinks of nuclides in Long Island sound. Adv. Geophys. 22,
129–163.
Wei, Ch-L., Murray, J.W., 1994. The behaviour of scavenged isotopes in marine anoxic environments: 210Pb and 210Po in the water
column of the Black Sea. Geochim. Cosmochim. Acta 58 (7), 1795–1811.
Williams, P.J., Jerkinson, N.W., 1982. A transportable microprocessor controlled precise Winkler titration suitable for field station and
shipboard use. Limnol. Oceanogr. 27, 576–584.
Yeager, K.M., Santschi, P.H., Rowe, G., 2004. Sediment accumulation and radionuclide inventories (239,240Pu, 210Pb and 234Th) in the
northern Gulf of Mexico, as influenced by organic matter and macrofaunal density. Mar. Chem. 91, 1–14.
602 P. Munoz et al. / Progress in Oceanography 75 (2007) 586–602
Copyright © 2022 FDOKUMEN




















