
Temperature and development rate of Triatoma guasayana (Hemiptera: Reduviidae) eggs under laboratory...
10
Physiological Entomology (2006) 31, 361–370 DOI: 10.1111/j.1365-3032.2006.00529.x © 2006 The Authors Journal compilation © 2006 The Royal Entomological Society 361 Introduction The effect of temperature on the development rate of triatom- ine eggs has been studied for several species under a variety of environmental conditions. The first record is that of Klug (1834) with Triatoma infestans (cited in Neiva, 1913), who used different controlled temperatures between 15 and 34 °C. Subsequently, many publications have reported the develop- ment rates of triatomine eggs at controlled temperatures, particularly for the species of highest epidemiological impor- tance: Triatoma brasiliensis (Pinto Soares et al. , 2000; Guarneri et al., 2003), Triatoma dimidiata (Zeledón et al., 1970; Martínez-Ibarra et al. , 2001), Triatoma garciabesi, Triatoma guasayana, T. infestans, Triatoma patagonica, Triatoma rubrovaria, Triatoma sordida, Triatoma eratyrusi- forme, Triatoma ninoi , Triatoma delpontei and Triatoma platensis (Carcavallo & Martínez, 1972), T. guasayana (Ghilini, 1982) , T. infestans (Hack, 1955), Triatoma pallid- ipennis (Martínez-Ibarra & Katthain-Duchateau, 1999), T. patagonica (Visciarelli et al., 2001), Triatoma pseudomac- ulata (Gonçalves et al. , 1997; Pinto Soares et al. , 2000), Temperature and development rate of Triatoma guasayana (Hemiptera: Reduviidae) eggs under laboratory conditions: physiological and adaptive aspects JORGE RABINOVICH 1 , SILVIA PIETROKOVSKY 2 and CRISTINA WISNIVESKY-COLLI 2 1 Centro de Estudios Parasitológicos y de Vectores, Universidad Nacional de La Plata, La Plata and 2 Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria, Buenos Aires, Argentina. Abstract. Eggs laid in the laboratory by T. guasayana (Wygodzinsky & Abalos 1949) adults collected from wild biotopes in Santiago del Estero (Argentina) were exposed to fluctuating temperatures, and 172 egg batches, involving 1574 eggs, completed development. Their development times were fitted to the linear degree- day model and three nonlinear models (devar, Lactin, and Rueda). Parameter val- ues and 95% CL were estimated. The lower development threshold is estimated to be 15 °C (linear devar model) and 17.5 °C (Lactin model), and the temperature resulting in the shortest development time is estimated to be between 26 °C (Lactin model) and 37 °C (nonlinear devar model). The thermal death point by the Lactin model is 34.4 °C, and conforms well with other studies. Differences between the present results and those of other studies for T. guasayana are discussed in terms of the maternal effects, the influence of experimental conditions, and the rate summa- tion effect. Using the climatic conditions of the natural environment of T. guasay- ana, the results are interpreted in terms of physiological adaptation. A generalized development rate response of T. guasayana eggs to increasing temperature is pro- posed, with three ranges: between 10 and 14.8 °C development rate increases in an accelerating way; between 14.8 and 29.2 °C development rate increases more or less linearly, and between 29.2 and 34 °C development rate decreases. Key words. Development threshold, development time, degree-day model, devar model, Lactin model, parameter estimation, Rueda model, temperature response, thermal death. Correspondence: J. Rabinovich, Centro de Estudios Parasitológicos y de Vectores, Universidad Nacional de La Plata, Calle 2 no. 584, 1900 La Plata, Argentina. Tel: 54 221 423 3471; fax: 54 221 423 2327; e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Temperature and development rate of Triatoma guasayana (Hemiptera: Reduviidae) eggs under laboratory...

Physiological Entomology (2006) 31, 361–370 DOI: 10.1111/j.1365-3032.2006.00529.x
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society 361
Introduction
The effect of temperature on the development rate of triatom-ine eggs has been studied for several species under a variety of environmental conditions. The first record is that of Klug (1834) with Triatoma infestans (cited in Neiva, 1913 ), who used different controlled temperatures between 15 and 34 °C. Subsequently, many publications have reported the develop-
ment rates of triatomine eggs at controlled temperatures, particularly for the species of highest epidemiological impor-tance: Triatoma brasiliensis ( Pinto Soares et al. , 2000; Guarneri et al. , 2003 ), Triatoma dimidiata ( Zeledón et al. , 1970; Martínez-Ibarra et al. , 2001 ), Triatoma garciabesi , Triatoma guasayana , T. infestans , Triatoma patagonica , Triatoma rubrovaria , Triatoma sordida , Triatoma eratyrusi-forme , Triatoma ninoi , Triatoma delpontei and Triatoma platensis ( Carcavallo & Martínez, 1972 ), T. guasayana ( Ghilini, 1982 ) , T. infestans ( Hack, 1955 ), Triatoma pallid-ipennis ( Martínez-Ibarra & Katthain-Duchateau, 1999 ), T. patagonica ( Visciarelli et al. , 2001 ), Triatoma pseudomac-ulata ( Gonçalves et al. , 1997; Pinto Soares et al. , 2000 ),
Temperature and development rate of Triatoma guasayana (Hemiptera: Reduviidae) eggs under laboratory conditions: physiological and adaptive aspects
J O R G E R A B I N O V I C H 1 , S I L V I A P I E T R O K O V S K Y 2 and C R I S T I N A W I S N I V E S K Y - C O L L I 2 1 Centro de Estudios Parasitológicos y de Vectores, Universidad Nacional de La Plata, La Plata and
2 Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales,
Universidad de Buenos Aires, Ciudad Universitaria, Buenos Aires, Argentina.
Abstract . Eggs laid in the laboratory by T. guasayana (Wygodzinsky & Abalos 1949) adults collected from wild biotopes in Santiago del Estero (Argentina) were exposed to fluctuating temperatures, and 172 egg batches, involving 1574 eggs, completed development. Their development times were fitted to the linear degree-day model and three nonlinear models (devar, Lactin, and Rueda). Parameter val-ues and 95% CL were estimated. The lower development threshold is estimated to be 15 °C (linear devar model) and 17.5 °C (Lactin model), and the temperature resulting in the shortest development time is estimated to be between 26 °C (Lactin model) and 37 °C (nonlinear devar model). The thermal death point by the Lactin model is 34.4 °C, and conforms well with other studies. Differences between the present results and those of other studies for T. guasayana are discussed in terms of the maternal effects, the influence of experimental conditions, and the rate summa-tion effect. Using the climatic conditions of the natural environment of T. guasay-ana , the results are interpreted in terms of physiological adaptation. A generalized development rate response of T. guasayana eggs to increasing temperature is pro-posed, with three ranges: between 10 and 14.8 °C development rate increases in an accelerating way; between 14.8 and 29.2 °C development rate increases more or less linearly, and between 29.2 and 34 °C development rate decreases.
Key words. Development threshold , development time , degree-day model , devar model , Lactin model , parameter estimation , Rueda model , temperature response , thermal death .
Correspondence: J. Rabinovich, Centro de Estudios Parasitológicos y de Vectores, Universidad Nacional de La Plata, Calle 2 no. 584, 1900 La Plata, Argentina. Tel: 54 221 423 3471 ; fax: 54 221 423 2327; e-mail: [email protected]

362 J. Rabinovich et al.
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
Triatoma picturata ( Barbosa & Garcia da Silva, 2001 ), Triatoma rubrofasciata ( Braga et al. , 1998 ), T. sordida ( Mello, 1976 ), Panstrongylus geniculatus ( Cabello & Galíndez, 1998 ), Rhodnius neglectus (Da Silva Rocha et al. , 2001a; Freitas et al. , 1967; Mello, 1977 ), Rhodnius pictipes ( Da Silva Rocha et al. , 1995 ) and Rhodnius robustus (Da Silva Rocha et al. , 2001b; Jurberg & Ferreira Rangel, 1980 ).
Other publications evaluate development rates of triatom-ine eggs under cold or heat shock or variable (controlled or uncontrolled) temperature regimes: for T. infestans ( Campos et al. , 2002 ), T. picturata ( Barbosa & Garcia da Silva, 2001 ), T. pseudomaculata ( Gonçalves et al. , 1997; Pinto Soares et al. , 2000 ), T. rubrofasciata ( Braga et al. (1998) , T. sordida ( Mello, 1976) and Rhodnius prolixus ( Luz et al. , 1999 ).
There are two fundamental thermal parameters that express how the rate of development of ectotherms depends on tem-perature: the lower threshold temperature for development ( T min , temperature below which no measurable development takes place), and the thermal constant ( K , number of degree-days above temperature T min for completion of development) ( Li, 1998 ). These parameters reflect the process of heat accu-mulation ( Baskerville & Emin, 1969 ) and are useful for ana-lysing the effects of temperature on the eggs of ectothermic animals, whose development depends upon the rate of intrin-sic biological processes at a given temperature ( Honek & Kocourek, 1988 ). It has been generally accepted that the ef-fect of temperature on development rates of insects is practi-cally linear near the range of temperatures that species normally experience in the field ( Gilbert & Raworth, 1996 ).
Triatoma guasayana (Wygodzinsky & Abalos 1949) is found in the north-western Dry Chaco of Bolivia, Paraguay and Argentina, particularly in the wild, but also colonizing peridomestic biotopes and occasionally invading human dwellings. There are only two available studies reporting the effect of temperature on T. guasayana ( Carcavallo & Martínez, 1972; Ghilini, 1982 ) but they were carried out at constant temperatures. In the present study, the response of the development rate of T. guasayana eggs to a variable tem-perature regime is studied, analysing the variability of that response in physiological and adaptive terms. The results are compared with those obtained for other species, although similarities and differences should be interpreted cautiously because many species show adaptations to habitats different from the ones occupied by T. guasayana .
Materials and methods
Insect material and rearing conditions
Triatoma guasayana collected from wild biotopes near the settlement of Trinidad, Department of Moreno, Province of Santiago del Estero (27E,12 � 30 � S, 63E,02 � 30 � W), Argentina, were used. Adult females were first-generation laboratory specimens, whereas males were field-collected adults. Insects were fed weekly with bovine heparinized blood using an arti-ficial feeder ( Núñez & Lazzari, 1990 ) at room temperature. The study started with 22 females that had been kept in
separate flasks after emergence as adults. After 24 – 48 h of adult life, females were placed individually with one or two males in plastic flasks (diameter: 3.3 cm, height: 5 cm), provided with folded absorbent paper, covered with a fine nylon mesh. Flasks were kept on a shelf within a double-walled metallic box, placed in a small room with no windows, in a laboratory in Buenos Aires. The double wall of the metallic box had a fibreglass layer inbetween and, although temperature was not regulated, it effectively dampened the external (outside) tem-perature. The maximum and minimum temperatures of the interior of the box were measured daily with a bulb ther-mometer. The experiments were carried out between 30 August 1996 (winter) and 19 January 1997 (summer). Flasks with the adult couples were examined daily, and eggs laid were counted, dated and transferred to a plastic box with folded paper, and kept until hatching, in the metallic box. Each egg group was checked daily for emergence and hatch-ing dates recorded. In total, 172 egg batches, involving 1574 individual eggs, completed development and were used in the analyses.
Statistical analysis
Because the laying and hatching dates were registered for all eggs, individual eggs were assigned in a spreadsheet to different lots (batches) with a common laying and hatching date. That is, for the purpose of analysis, batches were iden-tified that had the same development time and were exposed to the same average temperature, irrespective of the calendar date that each egg was laid and of the female that had laid the eggs.
Maximum and minimum temperatures of the metallic box were converted to median temperatures by the sine wave method ( Allen, 1976; Liu et al. , 1995 ). Development times of T. guasayana eggs were converted to development rates (1/development time). After screening the literature, four models were selected to compare the effect of temperature on development rates.
The simplest model is the linear model with a threshold (known as the degree-days model) ( Allen, 1976; Higley et al. , 1986; Burden & Hart, 1989; Kim et al. , 2000 ):
R ( T ) = b ( T − T min ) if T ≥ T min
R ( T ) = 0 if T < T min
where R ( T ) is the rate of development (percentage day – 1 ), T is temperature in °C, and b is the slope of the response of the rate of development to temperature above T min , and T min is the threshold temperature below which no development takes place. The program devar was used for fitting the data to this model ( Dallwitz & Higgins, 1992 ). devar is a flexible program that allows the maximum and minimum tempera-tures to be used if temperatures are variable (as in the present study).
The second model, also obtained from Dallwitz & Higgins (1992) , will be referred to as the nonlinear devar model:
R ( T ) = b 1 10 − v 2 ( 1 − b 5 + b 5 v 2 )

Development rate of T. guasayana eggs 363
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
where:
and where R ( T ) and T are as before; b 1 is the maximum de-velopment rate (in percent day – 1 ); b 2 is approximately the temperature (< b 3 ) at which R ( T ) falls to b 1 /10; b 3 is approx-imately the temperature at which R ( T ) is a maximum; b 4 con-trols how sharply R ( T ) approaches 0 at low temperatures; and b 5 controls the asymmetry of R ( T ). The terms c 1 and c 2 allow the approximate interpretations of b 2 and b 3 given above. The program devar ( Dallwitz & Higgins, 1992 ) was also used for fitting the data to this model.
The third model is the four-parameter nonlinear function used by Lactin et al. (1995) , as an improvement of the model developed by Logan et al. (1976) , allowing for the estimate of the development threshold at low temperatures. The model by Lactin et al. (1995) is:
R ( T ) = e � T − e [ � T max − ( T max − T ) / � ] + �
where R ( T ) and T are as before, and T max is the lethal (upper) temperature at which development stops, and � , � and � are fitted parameters ( � controls the rate of increase of the devel-opment rate with temperature, � makes the curve intercept the x -axis, thus allowing the estimation of the development temperature threshold, and � is a shape parameter).
The last model tested was that of Rueda et al. (1990) , which is a modification of the original model of Sharpe & DeMichele (1977) , and has the form:
where R ( T ) is the median development rate (days – 1 ) at tem-perature K (°K = °C + 273.15). RH025 , HA , TH and HH are parameters that were estimated by the Simplex method (see below): RH025 is the development rate at 25 °C (298.15 °K) assuming no enzyme inactivation (days – 1 ); HA is the enthalpy of activation of the rate controlling enzyme reaction (cal mol – 1 ); TH is the temperature (in °K) at which the rate con-trolling enzyme is 50% active; and HH is the change in en-thalpy associated with the highest temperature inactivation of the enzyme (cal mol – 1 ). The value 1.987 is the universal gas constant (cal – 1 degree – 1 day – 1 ).
The four models were fitted to the data using three types of software: (1) a fortran program developed by one of the authors, based on the Simplex method ( Himmelblau, 1972 ), and used to obtain a first estimate of the parameters; (2) these first estimates were fed to the nonlinear estimation procedure
of statistica (Vers. 5.5A) using the Quasi-Newton subrou-tine ( StatSoft, 1999 ), to produce the coefficient of correlation and the percent of the variance explained; and (3) confidence limits on the parameters were estimated using the method of Beale (1960) for nonlinear models, and computations were performed using a fortran program developed by Lobry et al. (1991) .
To compare the performance of different models with the different number of fitted parameters the coefficient of deter-mination ( R 2 ) was adjusted as proposed by Kvalseth (1985) :
where n is the number of observations (the 172 egg batches), k is the number of parameters of the i th model, and R i
2 is cal-culated as:
where y ̄ is the observed median development rate, and y ̂ j i is the j th predicted development rate for the i th model.
Model performance was also evaluated with the Akaike Criterion (AC) ( Akaike, 1974 ) and a variation of AC called the Model Selection Criterion (MSC) ( Scientist, 1995 ). Both methods are well known goodness of fit indicators, and they are preferred over classical goodness of fit tests (such as the chi-square or the G -tests) because they ‘penalize’ models with a high number of parameters. The AC and the MSC methods represent the ‘information content’ of a given set of parameter estimates ( Burnham & Anderson, 2002 ). They re-late the coefficient of determination to the number of param-eters, and are independent of the magnitude of the data points, as well as of the number of observations. The smaller the value of the AC, the more appropriate the model is. The MSC gives the same ranking between models as the AC but is normalized so that it is independent of the scaling of the data points, and the most appropriate model is that with the largest MSC (it maximizes the ‘information content’ of the model). They are calculated as:
and:
where, Y obsi and Y calci
are the observed and predicted devel-opment rates for the i th observation, p is the number of pa-rameters, n is the number of observations, and w i is a weight to be applied to each observation; equal weights were as-signed to all observations.
In addition to the four models used for fitting the experi-mental data, the logit transformation of the median develop-ment time was also used to obtain an independent estimate of the lower development threshold temperature ( Stemp, 1988 ). For that purpose, and because all eggs had the same
364 J. Rabinovich et al.
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
development time in each egg batch, to be able to apply the logit transformation of the median development time as a function of temperature, we had to ‘divide’ the 172 egg batches into a certain number of arbitrary intervals, each with a small range of temperatures. The original data were divided into five intervals and, for each of them, the average temper-ature, and the corresponding median development times and their logit transformation were calculated. These two varia-bles were linearly regressed and its projection to y = 0 pro-vides another estimate of the lower development threshold temperature.
To compare the results with other constant temperature T. guasayana studies, the possibility was tested that a physi-ological response mechanism related to the varying tempera-ture experiment might have been present. For that purpose, the procedure of Liu et al. (1995) was applied, based upon the calculation of the development rate on a per hour basis (applying the sine wave function to the minimum and maxi-mum daily temperatures). Next, the hourly predicted devel-opment rates of the nonlinear models were accumulated until development time was complete (sum of the development rates equal one), resulting in the theoretical development time (converted to day units). This procedure was applied only to those 45 batches that had an average daily tempera-ture within the same range of the constant temperature of the literature T. guasayana studies. The theoretical (expected) development times ( Dt e ) of the three nonlinear models were compared with the observed ones ( Dt o ) and the comparison was performed using Eq. 7 from Liu et al. (1995) , which cal-culates the percentage deviation ( d ) between expected and observed development times as:
d = 100 [ ( D t e / D t o ) − 1 ]
If the development at fluctuating temperatures can be ac-counted for by the rate-summation effect, the percent devia-tions should approach a normal distribution with mean zero for a sufficiently large sample size, and the t -test can be used to test for statistical significance. Negative values of d result from development time at fluctuating temperatures ( Dt e ) be-ing shorter than those at constant temperatures ( Dt o ) with similar means. The regression between d and temperature was also carried out, with a negative slope indicating that de-velopment time at fluctuating temperatures, when compared with constant temperatures with similar means, is shorter at lower temperatures and longer at higher temperatures.
Results
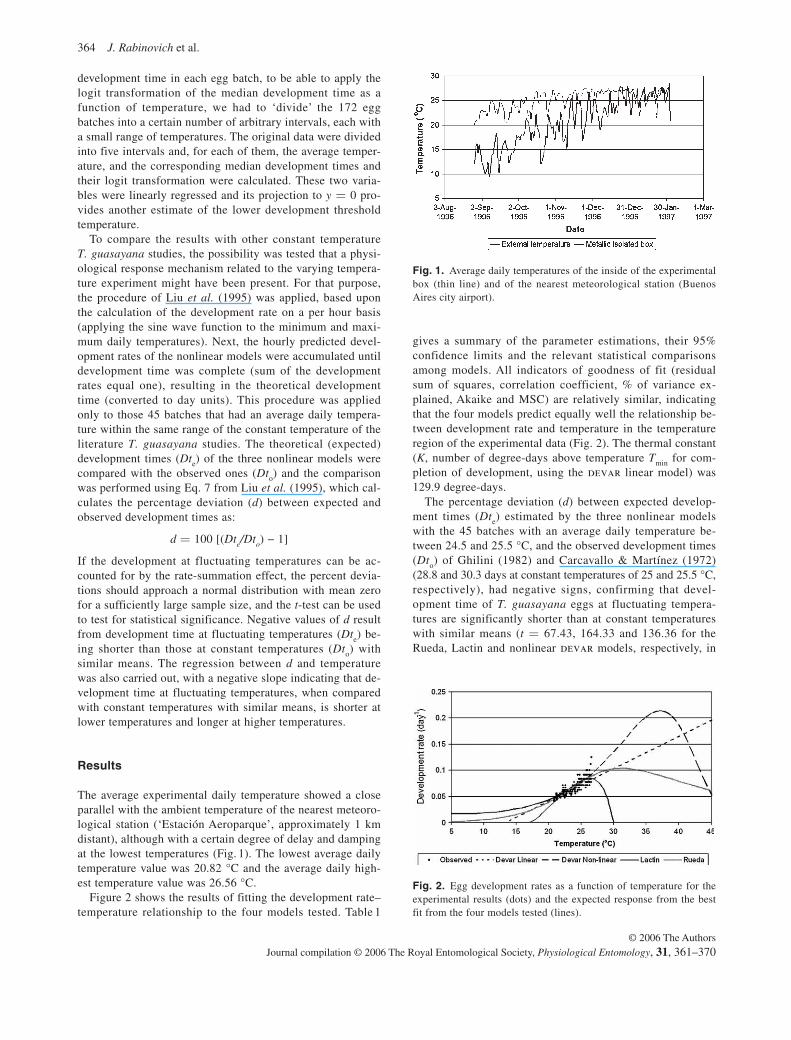
The average experimental daily temperature showed a close parallel with the ambient temperature of the nearest meteoro-logical station (‘Estación Aeroparque’, approximately 1 km distant), although with a certain degree of delay and damping at the lowest temperatures ( Fig. 1). The lowest average daily temperature value was 20.82 °C and the average daily high-est temperature value was 26.56 °C.
Figure 2 shows the results of fitting the development rate – temperature relationship to the four models tested. Table 1
gives a summary of the parameter estimations, their 95% confidence limits and the relevant statistical comparisons among models. All indicators of goodness of fit (residual sum of squares, correlation coefficient, % of variance ex-plained, Akaike and MSC) are relatively similar, indicating that the four models predict equally well the relationship be-tween development rate and temperature in the temperature region of the experimental data ( Fig. 2 ). The thermal constant ( K , number of degree-days above temperature T min for com-pletion of development, using the devar linear model) was 129.9 degree-days.
The percentage deviation ( d ) between expected develop-ment times ( Dt e ) estimated by the three nonlinear models with the 45 batches with an average daily temperature be-tween 24.5 and 25.5 °C, and the observed development times ( Dt o ) of Ghilini (1982) and Carcavallo & Martínez (1972) (28.8 and 30.3 days at constant temperatures of 25 and 25.5 °C, respectively), had negative signs, confirming that devel-opment time of T. guasayana eggs at fluctuating tempera-tures are significantly shorter than at constant temperatures with similar means ( t = 67.43, 164.33 and 136.36 for the Rueda, Lactin and nonlinear devar models, respectively, in
Fig. 1. Average daily temperatures of the inside of the experimental box (thin line) and of the nearest meteorological station (Buenos Aires city airport).
Fig. 2. Egg development rates as a function of temperature for the experimental results (dots) and the expected response from the best fit from the four models tested (lines).

Development rate of T. guasayana eggs 365
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
all cases n = 45, P = 0.0000). The slope of the regression ( b ) between the percent deviation ( d ) and temperature (within the 24.5 – 25.5 °C range) was statistically different from zero for the nonlinear devar model ( b = – 1.334, d.f. = 43, P = 0.00015) and the Rueda model ( b = – 1.963, d.f. = 43, P = 0.00075), but not for the Lactin model ( b = – 0.142, d.f. = 43, P = 0.5526). Predictions of the different models that fall outside the range of the experimental data (e.g. lower threshold temperature, or higher lethal temperatures) are justifiable because the ‘process-based’ models capture the essence of the underlying processes, and their predic-tions are accepted as a sort of hypothesis and not as extrapo-lations ( Haefner, 1996 ).
Discussion
The results of the present study confirm the generally ac-cepted knowledge that the effect of temperature on insect de-velopment rates is practically linear in the range of temperatures that a species experiences normally in the field ( Howe, 1967; Gilbert & Raworth, 1996 ). The fit to some of the three nonlinear models provides an estimate of the criti-cal upper and lower temperatures that lie outside this linear range. Additionally, all nonlinear models are able to predict
the optimal temperature of development. These three signifi-cant temperatures are discussed separately.
The optimal temperature
The linear effect of temperature on development rate falls off at average daily temperatures higher than those experi-enced normally in the field ( Howe, 1967 ), implying that there is an intermediate ‘optimum’ temperature for development. For example, Galliard (1935) shows that in three triatomine species ( T. dimidiata , T. vitticeps and R. prolixus ), under controlled temperatures between 15 and 34 °C, egg develop-ment rates are always faster at 32 °C. Guarneri et al. (2003) suggest that the optimum rate of egg development in T. bra-siliensis may be at temperatures around 25 °C, similar to the optimal temperature estimate by the Lactin model for T. gua-sayana . Alternating temperatures of 25/28 °C, 25/35 °C and 20/25 °C, and 86/75% relative humidity (RH) are found to be optimal for the development of R. prolixus ( Luz et al. , 1999 ). The structure and conceptual bases of the nonlinear models reproduce the expected ‘fall-off’ after the linear range of temperatures. The Lactin model appears to describe better the optimal temperature for egg development (see below for a discussion on field microclimate and spring temperatures).
Table 1. Fit of four development rate-temperature models to the development rate Triatoma guasayana eggs.
Model
Statistic Linear devar Nonlinear devar Lactin Rueda
Number of parameters 2 5 4 4 Residual sum of squares 106.39 117.64 116.62 108.44 Correlation coefficient 0.860 0.882 0.854 0.863 Correlation coefficient, Kvalseth (1985) adjusted
0.735 0.735 0.691 0.728
% variance explained 73.9 74.3 71.4 73.4 Akaike citerion 806.75 810.18 826.54 814.03 Model selection criterion 0.03323 0.0008 0.00503 0.00891 Parameter estimates b 1 = 0.767 (0.689 – 0.848) b 1 = 21.37 (17.50 – 25.70) T max = 34.44 (33.45 – 36.20) RH02 5 = 0.128
(0.115 – 0.139) T min = 14.96 (13.94 – 15.82) b 2 = 12.50 (10.08 – 14.84) � = 2.04 (1.84 – 2.22) HA = 35576.62
(32488.62 – 38294.13) b 3 = 37.13 (33.86 – 40.96) � = 0.010 (0.009 – 0.011) TH = 299.76
(298.77 – 300.87) b 4 = 3.10 (2.45 – 3.73) � = −1.196 [−1.207 – (−1.163)] HH = 47022.06
(43380.93 – 50999.71) b 5 = – 0.524 ( – 0.630 – 0.017) Threshold temperature 14.96 (13.94 – 15.82) – a 17.48 (12.41 – 23.00) – b Optimum temperature – 37.25 (36.09 – 40.33) 26.34 (25.82 – 28.21) 31.27 (30.39 – 33.93) Development time (days) at optimum temperature
– 4.68 11.89 9.64
Thermal constant ( K; degree-days)
129.87 – – –
Figures in parentheses after the parameter estimates are the lower and upper 95% confidence intervals. An explanation of the statistics used, the meaning of parameters and model comparison criteria is provided in the text. a It cannot be calculated for at low temperatures, development rate falls off asymptotically. However, it can be approximated by b 2 [the temperature (< b 3 ) at which the development rate falls to b 1/ 10], although this value (12.5 °C) can be considered only a very rough approximation. b It cannot be calculated for at low temperatures, development rate falls off asymptotically.

366 J. Rabinovich et al.
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
The development threshold temperature
Only two models provide the critical lower temperature: the linear devar (14.96 °C) and Lactin models (17.48 °C), whereas the nonlinear devar and Rueda models tend asymp-totically to low development rates at low temperatures. These values are very similar to the 14.8 °C development threshold temperature calculated by the logit transformation method applied to the median development times. These two esti-mates of the development threshold temperature conform well with the values found for other triatomines: in R. pro-lixus , no egg development below 20 °C and no egg hatching at 15 °C, and only different degrees of hatching success at 16 °C, depending upon the RH ( Clark, 1935 ). The development threshold temperature for the eggs of other Triatoma species is generally lower than for T. guasayana : 13 °C for T. di-midiata and 8 °C for T. vitticeps ( Galliard, 1935 ), although the eggs of T. brasiliensis do not develop below 19 °C ( Guarneri et al. , 2003 ). However, low temperatures such as 15 °C, which blocks development of R. prolixus nymphal stages during constant exposure, allow development at fluc-tuating patterns of various temperature and humidity condi-tions tested ( Luz et al. , 1999 ).
Gilbert & Raworth (1996) show that, in 36 insect species, the development threshold temperature is approximately equal to the average field temperature at first emergence from dormancy in spring. The average of the first day of spring (21 September) for three consecutive years (1995 – 97) of the Santiago del Estero station (the closest meteorological sta-tion to Trinidad, approximately 140 km distant) is 16.26 ± 2.2 °C (mean ± SD). The threshold temperature may play a critical role in the population dynamics. Gorla & Schofield (1989) show that development of T. infestans in the field is influenced more by the average minimal temperatures than by the average temperature, and mention unpublished data by Schofield & Williams indicating that the theoretical develop-ment zero for T. infestans is 16 °C, close to the 16.26 °C aver-age for the first day of spring. However, Curto de Casas & Carcavallo (1984) claim that the number of days per year with a temperature of at least 20 °C, a temperature consid-ered to permit normal biological activity of T. infestans , is more important for the distribution of this species than the minimal temperature.
The upper lethal temperature
Clark (1935) demonstrated the importance of the exposure of R. prolixus eggs to high temperatures: no eggs hatch at 35 °C above 35% RH, no eggs hatch at 33 – 34 °C at less than 35% RH, and no eggs hatch at 15 – 32 °C at less than 10% RH. Clark (1935) considered that the effects of lethal tempera-tures in the development of R. prolixus eggs is related to a hardening of the egg’s chorion that prevents hatching, and not to a physiological effect on the development of the em-bryos inside the eggs. However, Clarke (1960) suggests that the effect of temperature on development is not merely a direct one on metabolism but also one mediated by the
endocrine system. The high temperature that inhibits repro-duction is termed the sublethal temperature by Okasha (1964) and Okasha et al. (1970) .
Only the Lactin model estimates the upper temperature threshold (34.44 °C; 95% confidence interval = 33.45 – 36.20), which conforms well compared with the 35 °C ther-mal death point determined by Clark (1935) and the sublethal temperature of 34 °C determined by Okasha (1964) and Okasha et al. (1970) for R. prolixus . These comparisons have to be made with caution because, in the experiments by Clark (1935) , the eggs are subjected to various temperatures and RH in a permanent fashion. The damaging and irreversible effect of a temperature of 35 °C on the development of R. prolixus is dependent on the length of exposure ( Okasha et al. , 1970 ). Luz et al. (1999) show that an exposure for 8 h duration at 35 °C alternating with 16 h at 25 °C is not harm-ful to the development of any of the instars of R. prolixus . Eggs of T. brasiliensis do not develop above 38 °C ( Guarneri et al. , 2003 ); however, although temperatures of 35 °C or even higher can be expected in the natural habitats of tri-atomines, insects are able to avoid high temperatures by using biotopes with lower microclimatic temperatures. Conditions of extremely high temperature do not take place for extended periods of time during a given day.
The highest summer maximum temperature of the Santiago del Estero Station (years 1995 – 97) was 42.5 °C, whereas the maximum daily temperature is 33.83 ± 4.27 °C (mean ± SD). Thus, the temperature higher than the ‘optimal’ temper-ature of the Lactin model (34.44 °C) is close to the average maximum daily temperature observed in the field (33.83 °C). Gorla & Schofield (1985) determined, in an outdoor experiment also performed in the Argentinean Chaco, that climatic effects are the dominant mortality effects of T. infestans eggs.
The mean development time of egg batches exposed to the highest daily temperatures (26.3 °C) is 11.9 ± 1.1 days (Mean ± SD, n = 19 batches), which is much shorter than the values for T. guasayana eggs obtained by Carcavallo & Martínez (1972) under a constant temperature of 25 – 26 °C (28 days, n = 314), and by Ghilini (1982) under a constant temperature of 25 ± 4 °C (30.3 ± 6.59 days; three replicates, number of eggs not given). There are three possible explana-tions for these differences: (1) a maternal effect related to the age of the laying females; (2) differences in the experimental set up and environmental conditions; and (3) the different de-velopment time response between a constant temperature and a fluctuating temperature with the same mean value (the tem-perature regime in the present study has some extent of fluc-tuation between night and day, as well as between different days; Fig. 1 ). These three explanations are discussed separately.
The maternal effect
Howe (1967) suggests that there is a subtle influence on egg developmental time given by the age of the female par-ent that lays the eggs. In the present study, all females had a

Development rate of T. guasayana eggs 367
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
controlled age of 24 – 48 h after emergence as adults, whereas Carcavallo & Martínez (1972) and Ghilini (1982) do not re-port any such control
Effect of experimental and environmental conditions
Although temperature is usually a key environmental fac-tor, it interacts with humidity, food, and light. Carcavallo & Martínez (1972) and Ghilini (1982) used different humidity conditions and food sources. The experiments of Ghilini (1982) were characterized by constant temperatures of 25 ± 4 °C, 60 ± 20% RH, photoperiod of the City of La Plata, triatomines fed with live mice ‘almost always’ every week and nymphs and adults kept in 1000-cm 3 jars, with a maxi-mum of 10 large nymphs and adults per jar. The experimen-tal set up of Carcavallo & Martínez (1972) was a constant temperature of 25 – 26 °C, 60 – 65% RH, no photoperiod stated, triatomines fed with live pigeons every 2 weeks and 70 – 90 large nymphs and adults per jar (volume not stated). These conditions are very different from those employed in the present study (fluctuating temperature, no relative humidity control, nymphs and adults fed every week with bovine heparinized blood, and females and males kept in small indi-vidual vials). Additionally, there are some indications that the set up of Ghilini (1982) was not rigorously controlled be-cause the individual values of the egg development times of his three replicates are 21, 35 and 35 days. In the present study, eggs are used 0 – 24 h after being laid, whereas Ghilini (1982) used eggs that were 0 – 96 h old, and Carcavallo & Martínez (1972) do not provide the age of the eggs used. Mellanby (1935) showed that the effects of threshold tem-peratures interact with the stage of development of the em-bryos, being more resilient to threshold temperatures as the embryo is nearer development completion. Thus, egg age is another potential factor that may have affected the results of the different studies.
Rate summation effect
Development times of insects may differ, often considera-bly, between constant and variable temperature regimes with the same mean temperature ( Hagstrum & Hagstrum, 1970; Liu et al. , 1995 ). This can be attributed to two mechanisms: (1) the nonlinearity in the temperature-dependent rate func-tion and the rate-summation effect and (2) a physiological response (i.e. acceleration or deceleration of development by the temperature variation per se ) that acts in addition to the rate-summation effect ( Liu et al. , 1995 ). In the present study, the actual hourly temperatures oscillate between 18.7 and 28.9 °C, and the first mechanism should be disregarded be-cause our temperatures fluctuate within the linear range of the development time response to temperature. The test that a physiological response mechanism related to varying tem-perature might have been present proves to be statistically significant with two of the three nonlinear models. This con-clusion also conforms with the physiological results from
other triatomines: low and high temperatures, such as 15 and 35 °C, which block development of R. prolixus during con-stant exposure, allow development at the alternating patterns of temperature and humidity tested ( Luz et al. , 1999 ) in this species. Thus, this physiological response mechanism ex-plains the lower development times of the present experi-ments compared with those of Ghilini (1982) and Carcavallo & Martínez (1972) , and allow these differences to be disre-garded as a possible artifact derived from the use of a daily mean instead of the actual hourly fluctuating temperatures.
The adaptive significance of the difference in the effect of constant and fluctuating temperatures with the same mean on development times is shown by Petavy et al. (2001) with two Drosophila species: when reared under constant and alternat-ing temperature regimes, acceleration effects are observed at four alternating temperatures (in increasing order: 12 – 30, 9 – 21, 11 – 21, 16 – 26 °C), whereas retardation occurs at three other ones (in increasing order, 7 – 21, 5 – 15, 7 – 29 °C). Petavy et al. (2001) also show that very low viabilities occur under broad amplitude regimes, but the deleterious effects of some extreme temperatures, which would be lethal under constant temperatures, are recovered by daily return to a moderate temperature.
The two most common microhabitats (biotopes) used by T. guasayana for refuge, feeding and reproduction, are ‘Quimiles’ ( Opuntia quimilo ) and ‘Chaguares’ (several spe-cies of bromeliads) ( Wisnivesky-Colli et al. , 1997; Vezzani et al. , 2001 ). The temperature in these biotopes is strongly dampened, particularly in ‘Quimiles’: daily environmental maxima of the order of 28 °C are reduced to 20 °C in this biotope, and daily minima of the order of 6 °C are two-fold higher (13 °C). These actual field microclimatic conditions confirm that the development threshold temperature and the practically linear response of T. guasayana development rates with temperature is related to the temperatures that this species normally experiences in the field.
Conclusions
It is postulated that there is a generalized development rate response of T. guasayana eggs to temperature ( Fig. 3), with three ranges of response as suggested by Liu et al. (1995) : (1) between t 1 and t 2 , development rate increases in an accelerating way with increasing temperature; (2) be-tween t 2 and t 3 , development rate increases more or less lin-early with increasing temperature; and (3) between t 3 and t 4 , development rate decreases with increasing temperature. t 3 and t 4 are the low and high temperature limits permitting complete development. The value of t 1 is fixed at 13.94 °C, taken from the lower 95% confidence limit of the develop-ment threshold estimated by the linear devar model ( Table 1 ); the value of t 4 is fixed at 34 °C, taken from Okasha et al. (1970) for R. prolixus , and generally accepted as a sub-lethal temperature for triatomines; the values of t 2 and t 3 are calculated, as suggested by Liu et al. (1995) , as t 2 = t 1 + t s and t 3 = t 4 – t s , where t s = ( t 4 + t 1 )/5, resulting in t 2 = 18.0 °C and t 3 = 30.0 °C.

368 J. Rabinovich et al.
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
It is evident that, at the highest temperatures in the experi-mental metallic box (between approximately 25 and 27 °C), the development rate keeps rising, rather than beginning to slope off or starting to decrease as predicted by the theoreti-cal models. No explanation to this effect can be provided, although one possibility is the method of Liu et al. (1995) proposed for constant temperature changes, and not for fluc-tuating temperatures as is the case in the present study. Lin et al. (1954) show that the effects on development rate of vari-able temperatures that have the same daily mean as a con-stant daily temperature do not follow a consistent pattern: an acceleration of the development rate is observed in some cases but not in other cases. It is possible that, in the case of T. guasayana , the net effect would be one of an increasing development rate with higher temperatures.
In terms of physiological processes Clarke (1960) sug-gested that the effect of temperature on development is not merely a direct one on metabolism, but also an indirect one that is mediated by the endocrine system. The latter may be one of possible explanation for the effects of high tempera-ture on the reproduction (egg formation, oviposition and sterility) of the triatomine R. prolixus ( Okasha, 1964; Okasha et al. , 1970 ). Thus, it is not surprising that, as pos-tulated by Huey et al. (1991) , the relationship between de-velopment rate and temperature can be considered as the result of natural selection because this relationship changes when insects are exposed to different temperature regimes for many generations. Furthermore, Gilbert & Raworth (1996) claim that insects are selected for slow development in spring but fast development in summer and, in general, insects use temperature as an indicator of the time of sea-son. These physiological processes have adaptive and eco-logical implications. In the congeneric T. infestans , there is a cold-hardening response possibly favouring its survival in areas of South America that experience low temperatures ( Campos et al. , 2002 ). Natural selection pressures on the development time of eggs usually are related to a certain survival rate not only because the longer the development time, the higher the predation or parasitization risks, but
also because the longer the development time, the larger the eggs become, resulting in larger individuals with a higher survival and/or reproductive capacity. Additionally, the de-velopment time of the egg is also closely related to a syn-chronization process of the egg laying of the next generation of adults ( Morris, 1971 ).
Acknowledgements
We thank Patricia Gajate for help with laboratory activities and Waldo Hasperué for many aspects of programming and data management.
References
Akaike , H . ( 1974 ) A new look at statistical model identification . IEEE Transactions on Automatic Control , 19 , 716 – 23 .
Allen , J . ( 1976 ) A modified sine wave method for calculating degree days . Environmental Entomology , 5 , 328 – 396 .
Barbosa , A . & Garcia da Silva , I . ( 2001 ) Influência da temperatura na biologia de Triatomíneos. XXII. Triatoma picturata Usinger, 1939 (Hemiptera: Reduviidae) . Entomología y Vectores , 8 , 217 – 228 .
Baskerville , G . L . & Emin , P . ( 1969 ) Rapid estimation of heat accu-mulation from maximum and minimum temperatures . Ecology , 50 , 514 – 517 .
Beale , E . M . L . ( 1960 ) Confidence regions in non-linear estimation . Journal of the Royal Statistical Society B , 22 , 41 – 88 .
Braga , M . V . , Pinto , Z . T . & Lima , M . M . ( 1998 ) Life cycle and repro-ductive patterns of Triatoma rubrofasciata (De Geer, 1773) (Hemiptera: Reduviidae), under laboratory conditions . Memórias do Instituto Oswaldo Cruz , 93 , 539 – 542 .
Burden , D . J . & Hart , E . R . ( 1989 ) Degree-day model for egg eclosion of the pine needle scale (Hemiptera: Diaspididae) . Environmental Entomology , 18 , 223 – 227 .
Burnham , K . P . & Anderson , D . R . ( 2002 ) Model Selection and Multi-model Inference. A Practical Information-Theoretic Approach . Springer , New York, New York .
Cabello , D . R . & Galíndez , I . ( 1998 ) Vital statistics of Panstrongylus geniculatus (Latreille 1811) (Hemiptera: Reduviidae) under ex-perimental conditions . Memórias do Instituto Oswaldo Cruz , 93 , 257 – 262 .
Campos , S . G . P . , Rodrígues , V . L . C . C. , Wada , C . Y . et al . ( 2002 ) Effect of sequential cold shocks on survival and molting rate in Triatoma infestans Klug . Memórias do Instituto Oswaldo Cruz , 97 , 579 – 582 .
Carcavallo , R . U . & Martínez , A . ( 1972 ) Life cycles of some species of Triatoma (Hemiptera: Reduviidae) . The Canadian Entomolo-gist , 104 , 699 – 704 .
Clark , N . ( 1935 ) The effect of temperature and humidity upon the eggs of the bug, Rhodnius prolixus (Heteroptera, Reduviidae) . Journal of Animal Ecology , 4 , 82 – 87 .
Clarke , K . U . ( 1960 ) Studies on the relationships between air tem-perature and internal body temperature of Locusta migratoria L . Insect Physiology , 5 , 23 – 36 .
Curto de Casas , S . & Carcavallo , R . ( 1984 ) Límites de Triatomismo en la Argentina-Patagonia . Chagas , 9 , 35 – 40 .
Da Silva Rocha , D . , Galvão , C . , Jurberg y , J . & Lent , H . ( 1995 ) Lon-gevidad de los adultos, postura de las hembras y viabilidad de los
Fig. 3. Generalized development rate response of Triatoma guasay-ana eggs to temperature, with the three ranges of response. The dashed lined represents the approximate bounds of our experimental data in the temperature-development rate space. An interpretation and determination of the numerical values of t 1 , t 2 , t 3, and t 4 is pro-vided in the text.

Development rate of T. guasayana eggs 369
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
huevos de Rhodnius pictipes Stal, 1872 (Hemiptera, Reduviidae, Triatominae) . Entomología y Vectores , 2 , 61 – 65 .
Dallwitz , M . J . & Higgins , J . P . ( 1992 ) User’s Guide to DEVAR. A Com-puter Program for Estimating Development Rate as a Function of Temperature . Division of Entomology Report no. 2. Commonwealth Scientific and Industrial Research Organisation . (CSIRO), Australia .
Freitas , M . G . , Costa , J . O . & Costa , H . M . A . ( 1967 ) Alguns aspectos da biologia do Rhodnius neglectus Lent, 1954 (Hemiptera, Triatominae) em condicoes de Laboratorio. I. Evolucao . Arquivo Brasileiro de Medicina Veterinária e Zootecnia , 19 , 81 – 87 .
Galliard , H . ( 1935 ) Reproduction des Reduvides Hematophages . Annales de Parasitologie Humaine et Comparée , 14 , 97 – 112 .
Ghilini , J . M . ( 1982 ) Estadísticas Vitales de Triatoma guasayana Wy-godzinsky y Abalos 1949 (Hemiptera-Reduvidae) bajo Condicio-nes de Laboratorio . Revista de la Sociedad Entomólogica Argentina , 41 ( 1 – 4 ), 211 – 224 .
Gilbert , N . & Raworth , D . A . ( 1996 ) Insects and temperature – a general theory . The Canadian Entomologist , 128 , 1 – 13 .
Gonçalves , T . C . M . , Cunha , V . , de Oliveira , E . & Jurberg , J . ( 1997 ) Alguns aspectos da biologia de Triatoma pseudomaculata em condiçoes de laboratorio (Hemiptera, Reduviidae, Triatominae) . Memórias do Instituto Oswaldo Cruz , 92 , 275 – 280 .
Gorla , D . & Schofield , C . J . ( 1985 ) Analysis of egg mortality in ex-perimental populations of Triatoma infestans under natural cli-matic conditions in Argentina . Bulletin of the Society of Vector Ecologists , 10 , 107 – 117 .
Gorla , D . & Schofield , C . J . ( 1989 ) Population dynamics of Triatoma infestans under natural climatic conditions in the Argentina Chaco . Medical and Veterinary Entomology , 3 , 179 – 194 .
Guarneri , A . A . , Lazzari , C . , Xavier , A . A . P . et al . ( 2003 ) The effect of temperature on the behaviour and development of Triatoma brasiliensis . Physiological Entomology , 28 , 185 – 191 .
Hack , W . H . ( 1955 ) Estudios sobre Biología del Triatoma infestans (Klug, 1834) (Hem., Reduviidae) . Anales del Instituto de Medic-ina Regional , 4 , 125 – 147 .
Haefner , J . W . ( 1996 ) Modeling Biological Systems: Principles and Applications . Chapman & Hall , New York, New York .
Hagstrum , D . W . & Hagstrum , W . R . ( 1970 ) A simple device for pro-ducing fluctuating temperatures with an evaluation of the ecologi-cal significance of fluctuating temperatures . Annals of the Entomological Society of America , 63 , 1385 – 1389 .
Higley , L . C . , Pedigo , L . P . & Ostlie , K . R . ( 1986 ) Degday: a program for calculating degree-days, and assumptions behind the degree-day approach . Environmental Entomology , 15 , 999 – 1016 .
Himmelblau , D . M . ( 1972 ) Applied Nonlinear Programming . McGraw-Hill Book Co. , New York, New York .
Honek , A . & Kocourek , F . ( 1988 ) Thermal requirements for develop-ment of aphidophagous Coccinellidae (Coleoptera), Chrysopidae, Hemerobiidae (Neuroptera), and Syrphidae (Diptera): some gen-eral trends . Oecologia , 76 , 455 – 460 .
Howe , R . W . ( 1967 ) Temperature effects on embryonic development in insects . Annual Review of Entomology , 12 , 15 – 42 .
Huey , R . B . , Partridge , L . & Fowler , K . ( 1991 ) Thermal sensitivity of Drosophila melanogaster responds rapidly to laboratory natural selection . Evolution , 45 , 751 – 756 .
Jurberg , J . & Ferreira Rangel , E . ( 1980 ) Observaçoes sobre Rhod-nius robustus Larrousse, 1927 e Rhodnius pallescens Barber, 1932 (Hemiptera, Reduviidae, Triatominae) . Revista Brasileira de Bi-ología , 40 , 569 – 577 .
Kim , D . S . , Lee , J . H . & Yiem , M . S . ( 2000 ) Spring emergence pattern of Carposina sasakii (Lepidoptera: Carposinidae) in apple or-chards in Korea and its forecasting models based on degree-days . Environmental Entomolgy , 29 , 1188 – 1198 .
Kvalseth , T . O . ( 1985 ) Cautionary note about R 2 . The American Stat-istician , 39 , 279 – 285 .
Lactin , D . J . , Holliday , N . J . , Johnston , D . L . & Craigen , R . ( 1995 ) Improved rate model of temperature-dependent development by arthropods . Environmental Entomology , 24 , 69 – 74 .
Li , D . ( 1998 ) A linear model for description of the relationship be-tween the lower threshold temperature and thermal constant in spiders (Araneae: Arachnida) . Journal of Thermal Biology , 23 , 23 – 30 .
Lin , S . , Hodson , A . & Richards , G . ( 1954 ) An analysis of threshold temperatures for the development of Oncopeltus and Tribolium eggs . Physiological Zoology , 27 , 287 – 311 .
Liu , S . S . , Zhang , G . M . & Zhu , J . ( 1995 ) Influence of temperature variations on rate of development in insects: analysis of case stud-ies from entomological literature . Annals of the Entomological Society America , 88 , 107 – 119 .
Lobry , J . R . , Rosso , L . & Flandrois , J . P . ( 1991 ) A FORTRAN subrou-tine for the determination of parameter confidence limits in non-linear models . Binary , 3 , 86 – 93 .
Logan , J . A . , Wollkind , D . J . , Hoyt , S . C . & Tanigoshi , L . K . ( 1976 ) An analytic model for description of temperature dependent rate phe-nomena in arthropods . Environmental Entomology , 5 , 1133 – 1140 .
Luz , C . , Fargues , J . & Grunewald , J . ( 1999 ) Development of Rhod-nius prolixus (Hemiptera: Reduviidae) under constant and cyclic conditions of temperature and humidity . Memórias do Instituto Oswaldo Cruz , 94 , 403 – 409 .
Martínez-Ibarra , J . A . & Katthain-Duchateau , G . ( 1999 ) Biology of Triatoma pallidipennis Stal 1945 (Hemiptera: Reduviidae: Triato-minae) under laboratory conditions . Memórias do Instituto Oswaldo Cruz , 94 , 837 – 839 .
Martínez-Ibarra , J . A . , Miguel-Alvarez , A . , Arredondo-Jiménez , J . I . & Rodríguez-López , M . H . ( 2001 ) Update on the biology of Triat-oma dimidiata Latreille (Hemiptera: Reduviidae) under laboratory conditions . Journal of the American Mosquito Control Associa-tion , 17 , 209 – 210 .
Mellanby , H . ( 1935 ) The later embryology of Rhodnius prolixus . Quarterly Journal of Microscopic Science , 78 , 1 – 42 .
Mello , D . A . ( 1976 ) Biology of Triatominae (Reduvii, Hemiptera) from North of Formosa County (Goias – Brazil) I. Length of life cycle of Triatoma sordida (Stal 1859) . Revista de la Sociedade Brasileira de Medicina Tropical , 10 , 327 – 331 .
Mello , D . A . ( 1977 ) Biology of Triatominae (Reduviidae, Hemiptera) from North of Formosa County (Goias-Brazil) II. Length of life cycle of Rhodnius neglectus Lent, 1954 . Revista de la Sociedade Brasileira de Medicina Tropical , 11 , 63 – 66 .
Morris , R . F . ( 1971 ) Observed and simulated changes in genetic qual-ity in natural populations of Hyphantria cunea . The Canadian Entomologist , 103 , 893 – 906 .
Neiva , A . ( 1913 ) Informações sobre a Biologia da Vinchuca, Triatoma infestans Klug . Memórias do Instituto Oswaldo Cruz , 5 , 24 – 31 .
Núñez , J . A . & Lazzari , C . R . ( 1990 ) Rearing of Triatoma infestans Klug (Heteroptera: Reduviidae) in the absence of a live host. I. Some factors affecting the artificial feeder . Journal of Applied En-tomology , 109 , 87 – 92 .
Okasha , A . Y . K . ( 1964 ) Effects of high temperature in Rhodnius pro-lixus (Stal) . Nature , 204 , 1221 – 1222 .
Okasha , A . Y . K . , Hassanein , A . M . M . & Farahat , A . Z . ( 1970 ) Effects of sub-lethal high temperature on an insect, Rhodnius prolixus (Stal) . Journal of Experimental Biology , 53 , 25 – 36 .
Petavy , G . , David , J . R . , Gibert , P . & Moreteau , B . ( 2001 ) Viability and rate of development at different temperatures in Drosophila: a comparison of constant and alternating thermal regimes . Journal of Thermal Biology , 26 , 29 – 39 .

370 J. Rabinovich et al.
© 2006 The AuthorsJournal compilation © 2006 The Royal Entomological Society, Physiological Entomology, 31, 361–370
Pinto Soares , R . P . , das Graças , L . E . , Soares Laranja , L . & Diotaiuti , L . ( 2000 ) Population dynamics and feeding behavior of Triatoma brasiliensis and Triatoma pseudomaculata , main vectors of Cha-gas disease in Northeastern Brazil . Memórias do Instituto Os-waldo Cruz , 95 , 151 – 155 .
Rueda , L . M . , Patel , K . J . , Axtell , R . C . & Stinner , R . E . ( 1990 ) Temperature-dependent development and survival rates of Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae) . Journal of Medical Entomology , 27 , 892 – 898 .
Scientist ( 1995 ) Scientist Handbook (Rev. 7EEF) . MicroMath Scien-tific Software, Inc. , Salt Lake City, Utah .
Sharpe , P . J . H . & DeMichele , D . W . ( 1977 ) Reaction kinetics of poikilo-therm development . Journal of Theoretical Biology , 64 , 649 – 670 .
da Silva Rocha , D . , Jurberg , J . , Carcavallo , R . U . et al . ( 2001a ) Influên-cia da temperatura e umidade na biologia de Rhodnius neglectus Lent, 1954 em laboratório (Hemiptera, Reduviidae, Triatominae) . Re-vista da Sociedade Brasileira de Medicina Tropical , 34 , 357 – 363 .
da Silva Rocha , D . , Jurberg , J . , Carcavallo , R . U . et al . ( 2001b ) In-fluência da temperatura e umidade no desenvolvimento ninfal de Rhodnius robustus . Revista de Saúde Pública , 35 , 400 – 406 .
StatSoft ( 1999 ) Statistica for Windows (Computer Program Man-ual) . Statsoft Inc. , Tulsa, Oklahoma .
Stemp , I . C . ( 1988 ) An investigation into the temperature dependence of Triatoma infestans . Applied Project for a Diploma in Mathe-matical Statistics , pp . 1987 – 88 . Statistical Laboratory, Cambridge University , U.K.
Vezzani , D . , Schweigmann , N . J . , Pietrokovsky , S . M . & Wisnivesky-Colli , C . ( 2001 ) Characterization of Triatoma guasayana biotopes in a hardwood forest of Santiago del Estero, Argentina . Memórias do Instituto Oswaldo Cruz , 96 , 459 – 466 .
Visciarelli , E . , Ferrero , A . A . & Picollo , M . I . ( 2001 ) Caracterización morfológica del tiempo de desarrollo embrionario de Triatoma patagonica , del Ponte 1929 (Hemiptera: Reduviidae) . Ento-mología y Vectores , 8 , 449 – 461 .
Wisnivesky-Colli , C . , Schweigmann , N . J . , Pietrokovsky , S . M . et al . ( 1997 ) Spatial distribution of Triatoma guasayana (Hemiptera, Reduviidae) in hardwood forest biotopes in Santiago del Estero, Argentina . Journal of Medical Entomology , 34 , 102 – 109 .
Zeledón , R . , Guardia , V . , Zúñiga , M . et al . ( 1970 ) Biology and ethol-ogy of Triatoma dimidiata (Latreille, 1811) adults . Journal of Medical Entomology , 7 , 313 – 19 .
Accepted 4 May 2006 First published online 26 July 2006




















