
Risedronate metal complexes potentially active against Chagas disease
Upload
independentCategory
view
1download
0
iGladf
Journal of Invertebrate Pathology 76, 198–207 (2000)doi:10.1006/jipa.2000.4964, available online at http://www.idealibrary.com on
Characterization and Carbon Metabolism in Fungi Pathogenicto Triatoma infestans, a Chagas Disease Vector
M. Patricia Juarez,* Rosana Crespo,* Gustavo Calderon Fernandez,*Roberto Lecuona,† and Lazaro F. R. Cafferata‡
*Instituto de Investigaciones Bioquımicas de La Plata, Facultad de Ciencias Medicas, UNLP, calles 60 y 120, 1900 La Plata, Argentina;†Instituto de Microbiologıa y Zoologia Aplicada, Centro de Ciencias Veterinarias y Agronomicas, Instituto Nacional de Tecnologıa
Agraria, Castelar, C.C. 25, 1712 Castelar, Buenos Aires, Argentina; and ‡Ladecor, Laboratorio de Quımica Organica,Facultad de Ciencias Exactas, UNLP, calles 47 y 115, 1900 La Plata, Argentina
E-mail: [email protected]
Received March 27, 2000; accepted June 14, 2000
The pathogenicity of Metarhizium anisopliae (Ma) andBeauveria bassiana (Bb) isolates against Triatoma infes-tans, the major vector of Chagas disease in Argentina isreported. A 100% mortality was achieved with mean le-thal times varying form 5.8 (Ma6) to 7.7 (Bb5) or 11.1 days(Bb10). The fatty acid, hydrocarbon, and total lipid pat-terns were compared for glucose-grown and alkane-grown Bb10 cultures. The alkane-grown cells showed alipid pattern different from that of glucose-grown cells,with triacylglyercol as the major lipid fraction, whereassterols prevailed in the glucose-grown cells. A signifi-cant reduction in the relative amounts of linoleic aciddiminished the unsaturated/saturated fatty acid ratiofor alkane-grown cells; in addition, large amounts ofheptacosanoic and eicosanoic acids were detected in thesaturated fraction. The hydrocarbon profile of Bb10showed a saturated chain length distribution, with amarked prevalence for straight chains, ranging from n-C18 to n-C37 in the carbon skeleton, with n-C22 as themajor component. Alkane-grown cells showed no quali-tative changes in their hydrocarbon fraction, but a sim-ilar ratio for odd/even carbon chains. After 48-h incuba-tion assays, [1-14C]acetate uptake was largely dimin-shed following a period of alkane growth induction.lucose-grown cells readily incorporated 19% of the
abel into phospholipids, hydrocarbons, triacylglycerols,nd free fatty acids. In contrast, incorporation was re-uced to 5.3% for alkane-grown cells, accounting onlyor phospholipid synthesis. © 2000 Academic Press
Key Words: Beauveria bassiana; Metarhizium aniso-pliae; Triatoma infestans; hydrocarbon catabolism;fatty acids.
INTRODUCTION
Triatominae are the major vectors of Chagas disease inthe Americas, with a population of 16–18 million peoplecurrently infected (WHO, 1991); in Argentina, Triatoma
1980022-2011/00 $35.00Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
infestans is the main target of vector control programs,and massive chemical insecticide treatments have beenapplied for decades. However, biological control strate-gies are not yet well established against these insectvectors, although Romana and Fargues (1987, 1992) iso-lated several fungal strains pathogenic to Rhodnius pro-lixus, the major triatominae vector in Venezuela, Colom-bia, and Central America, and field assays were reported(Romana, 1995). Strains of Beauveria sp. and Metarhi-zium sp. (Deuteromycotina: Hyphomycetes) pathogenicto T. infestans were selected by Lecuona (1999), and someof them were used for this study. The cuticular hydrocar-bons of T. infestans comprise a complex mixture of satu-rated straight chains (mostly n-C29, n-C31, and n-C33)and branched chains of 35, 37, and 39 carbons in theirskeleton (Juarez and Blomquist, 1993). It was recentlyshown that the entomopathogenic fungi Metarhiziumanisopliae (Ma) and Beauveria bassiana (Bb) have theability to metabolize triatomine insect cuticular-like hy-drocarbons into a variety of lipid components, utilizingalkanes as their sole carbon source (Napolitano andJuarez, 1997). A high-energy yield for hydrocarbon con-sumption through complete b-oxidation mechanisms wasmeasured as volatile organic compounds (VOC) released.A period of growth induction was shown to markedlyenhance alkane-consumption activity. Evidence was alsoraised indicating that hydrocarbon degradation reactionsoccur primarily in fungal peroxisomes (Crespo et al.,2000). The ability of bacteria and yeasts to utilize medi-um-chain-length n-alkanes as their sole carbon sourcehas been frequently addressed (Makula and Finnerty,1973; Fukui and Tanaka, 1979; Gallo et al., 1973). How-ever, the performance of entomopathogenic fungi, of rel-evance in integrated pest management strategies, hasbeen scarcely studied. The biochemical mechanisms in-volved in the utilization of very-long-chain hydrocarbons,of chain lengths similar to those present at the insect

t
igs
s
N
d
199CARBON METABOLISM IN ENTOMOPATHOGENIC FUNGI
surface, and the uptake and transport of the hydrocar-bons and their metabolites into the microorganism arestill poorly understood. The outermost insect cuticularlayer is the epicuticle, covered by a thin layer of lipids,among which usually prevail long-chain hydrocarbonsconforming a highly inert coating, which provides a bar-rier against water loss and chemical or microbial inva-sion (Lockey, 1988; Juarez, 1994). Epicuticular lipid frac-ions of the lepidopteran Ostrinia mubilalis favored ger-
mination of pathogenic Beauveria sp., but they inhibitedhyphal growth of nonpathogenic strains (Lecuona et al.,1997).
The aim of this study was to further characterizelocal fungal strains of B. bassiana and a Brazilianisolate of M. anisopliae, virulent to T. infestans, and toncrease our understanding of entomopathogenic fun-al lipid metabolism to help improve current controltrategies.
MATERIALS AND METHODS
Fungal Strain Culture, Germination and GrowthConditions, and Conidia Production
B. bassiana strains were from the collection of theInstituto de Microbiologıa y Zoologıa Aplicada, IMYZA,INTA, Castelar, and M. anisopliae was obtained fromthe Centro Nacional de Pesquisa de Recursos Geneti-cos e Biotecnologia, Brasilia, Brazil. Monoconidial cul-tures were produced for reach isolate and maintainedat 26 6 0.5°C for 15 days in plates containing completeolid agar medium (CMA: 0.4 g KH2PO4, 1.4 g
Na2HPO4 z 12H2O, 0.6 g MgSO4 z 7H2O, 1.0 g KCl, 0.7 gH4NO3 z 7H20, 10 g glucose, 5 g yeast extract, 15 g
agar in 1000 ml of distilled water). Median germina-tion time (GT50 or GT90) was determined for each strainby culture of 100 ml of conidia suspension (1 3 107
conidia/ml) on petri dishes containing the nutritiveCMA medium. Germination was assessed after incu-bation at 26 6 0.5°C for 6 h in four replicates (100conidia/strain each). Conidia germination wasachieved when the germ tube was greater than orequal to conidium diameter (Beauveria sp.) or greaterthan or equal to conidium length (Metarhizium sp.)(Lecuona, 1996). Probit analysis was used for calcula-tion of the GT50 (Finney, 1964). To measure colonygrowth, 2.5 3 105 conidia/ml of each strain was cul-tured on the center of a petri dish on CMA medium andincubated at 26 3 0.5°C. After 9 days for B. bassianastrains or 15 days for M. anisopliae, the colony diam-eter was measured. Four replicates per strain wereperformed and the design was completely randomized.To determine conidia production, they were trans-ferred by an inoculating loop to test tubes containing10 ml of a sterile Tween 80 solution (0.1%). The num-ber of conidia per colony was estimated by counting ona hematocytometer.
Bioassays
To analyze the pathogenesis and to estimate themean lethal time (MLT), 4th instar T. infestansnymphs (three replicates, 30 insects each) were im-mersed in a suspension containing 1 3 108 conidia/mlfor 6 s and were then maintained in individual plasticcontainers. A control assay was similarly performedafter 6 s immersion in distilled water. Nymph mortal-ity was scored daily and MLT was estimated as inMoore et al. (1995).
Insects
T. infestans were obtained from a colony reared at30°C, 50–60% relative humidity and fed on chickens atINIBIOLP, Facultad de Ciencias Medicas, La Plata,Argentina.
Chemicals
Solvents were from Carlo Erba (Milan, Italy) andMerck (Darmstadt, FRG). Silica gel and other chemi-cals were from Aldrich (Milwaukee, WI). Hydrocarbonsand fatty acid standards were purchased from SigmaChemical Co. (St. Louis, MO). Solvents were redistilledprior to use. Ultima Gold XR scintillation cocktail waspurchased from Packard Instrument Co. Inc. (Meriden,CT). Sodium [1-14C]acetate (56 mCi/mmol) was fromNew England Nuclear (Boston, MA).
Fungal Growth on Synthetic Hydrocarbon-EnrichedMedia
Assays were performed on media agar plates withthe same composition as the CMA, deficient in glucoseand yeast extract (DMA), and containing n-octacosane(2.5 ml of a 10% hexane solution, w/v). The hydrocarbonwas layered onto the surface media and evaporated.The appropriate fungal strain already grown on CMA(FS0) was inoculated on the plates and incubated at26°C for 12–14 days. Fungi grown on DMA during thatperiod (FS1) were inoculated again under similar con-
itions for another 12- to 14-day period (FS2).
[1-14C]Acetate Utilization
Radiolabeled sodium acetate (1 ml of an ethanolicsolution containing 1 3 106 cpm) was spotted onto thecenter of either DMA or CMA agar plates. After evap-oration of the solvent, 5 3 105 conidia of Bb10 har-vested from FS0, FS1, or FS2 cultures were inoculatedon the surface of the labeled precursor and incubatedat 26°C for 48 h.
Product Analysis
After the incubation period, a 2-cm-diameter agardisk containing the growing colony was carefullyscrapped off with a metal spoon from each petri dish

sf
B
200 JUAREZ ET AL.
and transferred to a glass homogenizer. Fungal lipidswere extracted with chloroform:methanol (2:1) andpartitioned with distilled water (1:5 v/v) according tothe Folch method (Folch, 1957). The volume of eachlayer was measured, and aliquots were separated forradioactivity determination by liquid scintillationcounting. Afterward, organic layers were reduced involume under vacuum or N2 stream and stored at218°C. Agar was further extracted with n-hexane (2ml 33) to fully recover the unreacted hydrocarbon, andradioactivity was determined in aliquots as describedabove. Blanks containing [3H]hydrocarbon but noconidia were similarly extracted to correct raw data forthe extraction efficiency. The chloroform layer was an-alyzed by TLC performed on silica gel plates developedtwice in hexane, followed by petroleum ether:diethylether:acetic acid (80:20:1). Distribution of the radioac-tivity was analyzed by radio-TLC using a Bertholdscanner (Berthold, Germany). To determine labeledprecursor incorporation, radioactivity was also mea-sured by liquid scintillation counting performed with aBeckman liquid scintillation counter at 90% efficiencyfor 14C.
Lipid Analysis and Identification of FungalHydrocarbons and Fatty Acids
Fungal hydrocarbons from FS0 and FS2 cultureswere extracted by immersion in hexane (6 ml/g) andpartition with distilled water (5:1, v/v). Solvent wasconcentrated under vacuum and the extract waspassed through a minicolumn (2.5 3 0.5 cm) of Biosil(Bio-Rad Lab, Richmond, CA); hydrocarbons wereeluted with n-hexane, as previously described (Napoli-tano and Juarez, 1997). The eluent was concentratedand analyzed on a Hewlett–Packard 6890 gas chro-matograph with a flame ionization detector (FID). A30 m 3 0.32 mm I.D. 0.5-mm stationary phase DB-5 (J& W Scientific, Folson, CA) capillary column was usedin splitless mode, with an initial temperature of 50°Cfor 1 min and a temperature program from 50 to 200°Cat a rate of 50°C/min, then at 3°C/min up to 300°C, anda final holding time of 15 min at 300°C. Mass spectrom-etry analyses were carried out by interfacing a Hewlet-t–Packard 5890 gas chromatograph with FID detectionto a Hewlett–Packard 5972A quadrupole mass selec-tive detector. Helium was the carrier gas at 12 p.s.i.,and the analysis was carried out on a fused silicamethyl–phenyl silicone column (Heliflex RSL-150;bonded FSOT, 30 m length 3 0.25 mm I.D.) pro-grammed with a 5-min isothermal period at 110°Cfollowed by a temperature program of 5°C/min up to290°C and a final holding time of 10 min. Electronimpact (EI) mass spectra were obtained at 70 eV.Methyl-branched alkanes were identified by calculat-ing their equivalent chain length (ECL) by regressionanalysis derived from the straight chain hydrocarbon
components and their mass spectral fragmentationpatterns, which were interpreted as described byBlomquist et al. (1987) and Carlson et al. (1998). Forthe analysis of lipids, whole fungal cells were extractedby the Folch method as described above under ProductAnalysis, the extract was applied to high-performanceTLC plates (silica gel 60; Merck), and chromatogramswere developed using three different solvent systems.Hexane (100%) was the first solvent, followed by hex-ane:ethyl ether:acetic acid (80:20:1 v/v), both to the topof the plate. To differentiate sterols from diacylglycer-ols, the sequence was followed by a third run withbenzene:ethyl ether:water:acetic acid (40:50:2:0.2 v/v)up to 2/3 of the plate. Spots were visualized under UVlight and charred after spraying with sulfuric acid. Forphospholipid analysis, plates were run in chloroform:methanol:acetic acid:water (65:25:3:1.5 v/v). Lipidswere identified by comparison with appropriate stan-dards similarly run. For individual fatty acid analyses,fungal lipids were extracted as described above. Thechloroform extract was evaporated under N2 and re-uspended in 1 ml toluene; transesterification was per-ormed at 100°C with BF3-methanol for 60 min. The
resulting fatty acid methyl esters (FAME) were puri-fied by TLC in hexane (100%, 32) and in hexane:diethyl ether:acetic acid (80:20:1). An aliquot of theinitial chloroform extract was similarly run to analyzelipid distribution. Gas chromatographic analyses of theFAME were performed on a Hewlett–Packard 6890 gaschromatograph. Samples were analyzed on a 30 m 30.32 mm I.D. 0.5-mm stationary phase Omega Wax 250(Alltech Associates, Arlington Heights, IL) capillarycolumn. The temperature of the oven was programmedfrom 175 to 220°C at a rate of 3°C/min and then held at220°C for 15 min. Mass spectrometry analyses werecarried out with the same equipment and column asdescribed for hydrocarbon analyses. The column wasprogrammed with a 5-min isothermal period at 110°Cfollowed by a temperature program set at a rate of5°C/min up to 230°C. EI mass spectra were obtained at70 eV and the chromatographic peaks were identifiedby library search and comparison with FAME stan-dards similarly run.
RESULTS
Cultivation Conditions and Pathogenesis
Median germination time (GT50 or GT90), conidiagrowth, and conidia production of B. bassiana (Bb10,
b5) and M. anisopliae (Ma6) were estimated for FS0
cultures in CMA agar and are shown in Table 1. GT50
and GT90 were significantly larger for Ma6 than for theBb strains, and the amount of conidia produced by Ma6was markedly lower than that for Bb strains. Whenbioassays for pathogenicity on 4th instar. T. infestansnymphs were conducted as explained under Materials
toabymasftpAfcne
B
fcpd(l
tl
lidp3pMc
[
c[
dl
s
201CARBON METABOLISM IN ENTOMOPATHOGENIC FUNGI
and Methods, all three strains showed 100% mortality,with mean lethal times of 11.1 (Bb5), 7.7 (Bb10), and5.8 (Ma6) days for FS0 culture condition (Table 1).
Hydrocarbon Analysis of B. bassiana
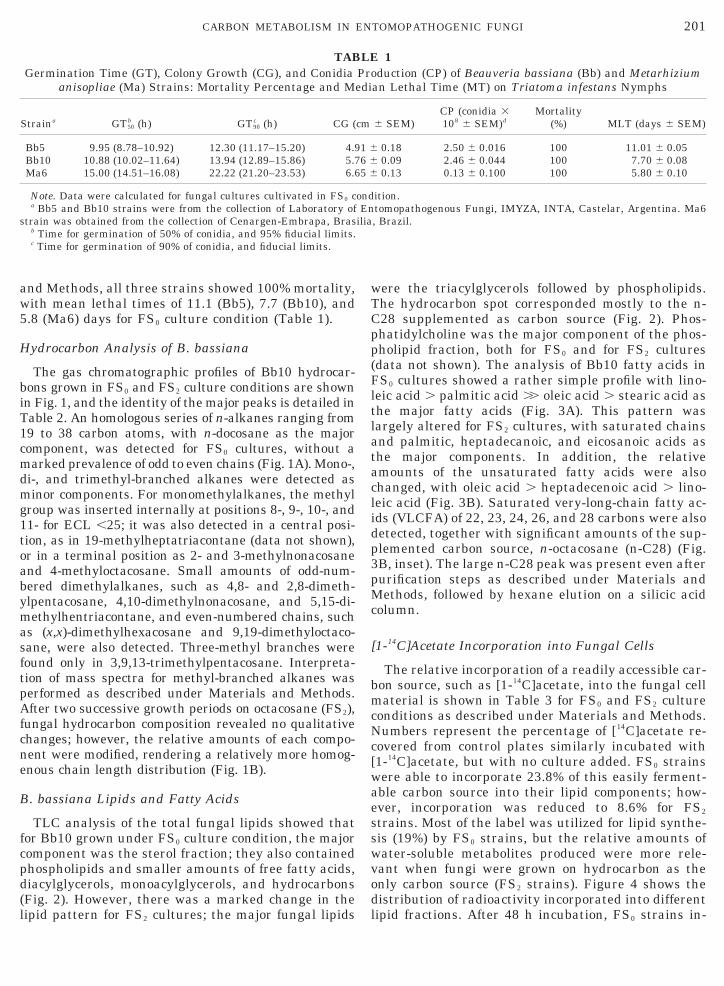
The gas chromatographic profiles of Bb10 hydrocar-bons grown in FS0 and FS2 culture conditions are shownin Fig. 1, and the identity of the major peaks is detailed inTable 2. An homologous series of n-alkanes ranging from19 to 38 carbon atoms, with n-docosane as the majorcomponent, was detected for FS0 cultures, without amarked prevalence of odd to even chains (Fig. 1A). Mono-,di-, and trimethyl-branched alkanes were detected asminor components. For monomethylalkanes, the methylgroup was inserted internally at positions 8-, 9-, 10-, and11- for ECL ,25; it was also detected in a central posi-ion, as in 19-methylheptatriacontane (data not shown),r in a terminal position as 2- and 3-methylnonacosanend 4-methyloctacosane. Small amounts of odd-num-ered dimethylalkanes, such as 4,8- and 2,8-dimeth-lpentacosane, 4,10-dimethylnonacosane, and 5,15-di-ethylhentriacontane, and even-numbered chains, such
s (x,x)-dimethylhexacosane and 9,19-dimethyloctaco-ane, were also detected. Three-methyl branches wereound only in 3,9,13-trimethylpentacosane. Interpreta-ion of mass spectra for methyl-branched alkanes waserformed as described under Materials and Methods.fter two successive growth periods on octacosane (FS2),
ungal hydrocarbon composition revealed no qualitativehanges; however, the relative amounts of each compo-ent were modified, rendering a relatively more homog-nous chain length distribution (Fig. 1B).
. bassiana Lipids and Fatty Acids
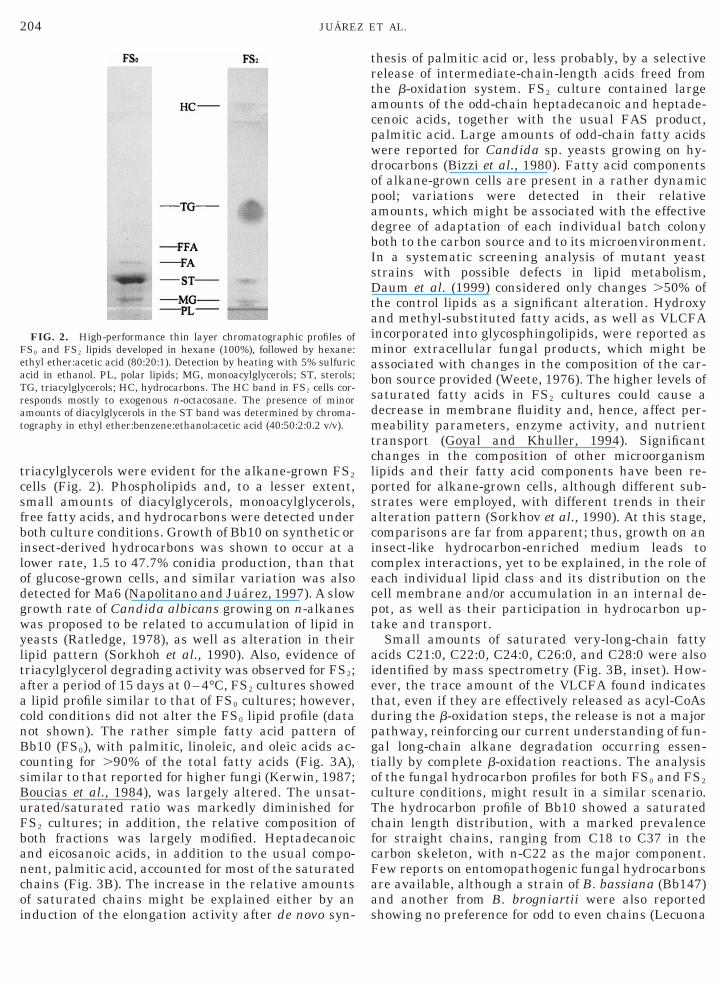
TLC analysis of the total fungal lipids showed thator Bb10 grown under FS0 culture condition, the majoromponent was the sterol fraction; they also containedhospholipids and smaller amounts of free fatty acids,iacylglycerols, monoacylglycerols, and hydrocarbonsFig. 2). However, there was a marked change in theipid pattern for FS2 cultures; the major fungal lipids
TABGermination Time (GT), Colony Growth (CG), and Conidia
anisopliae (Ma) Strains: Mortality Percentage and M
Straina GT50b (h) GT90
c (h) CG (
Bb5 9.95 (8.78–10.92) 12.30 (11.17–15.20) 4.9Bb10 10.88 (10.02–11.64) 13.94 (12.89–15.86) 5.7Ma6 15.00 (14.51–16.08) 22.22 (21.20–23.53) 6.6
Note. Data were calculated for fungal cultures cultivated in FS0 ca Bb5 and Bb10 strains were from the collection of Laboratory of
train was obtained from the collection of Cenargen-Embrapa, Brasb Time for germination of 50% of conidia, and 95% fiducial limits.c Time for germination of 90% of conidia, and fiducial limits.
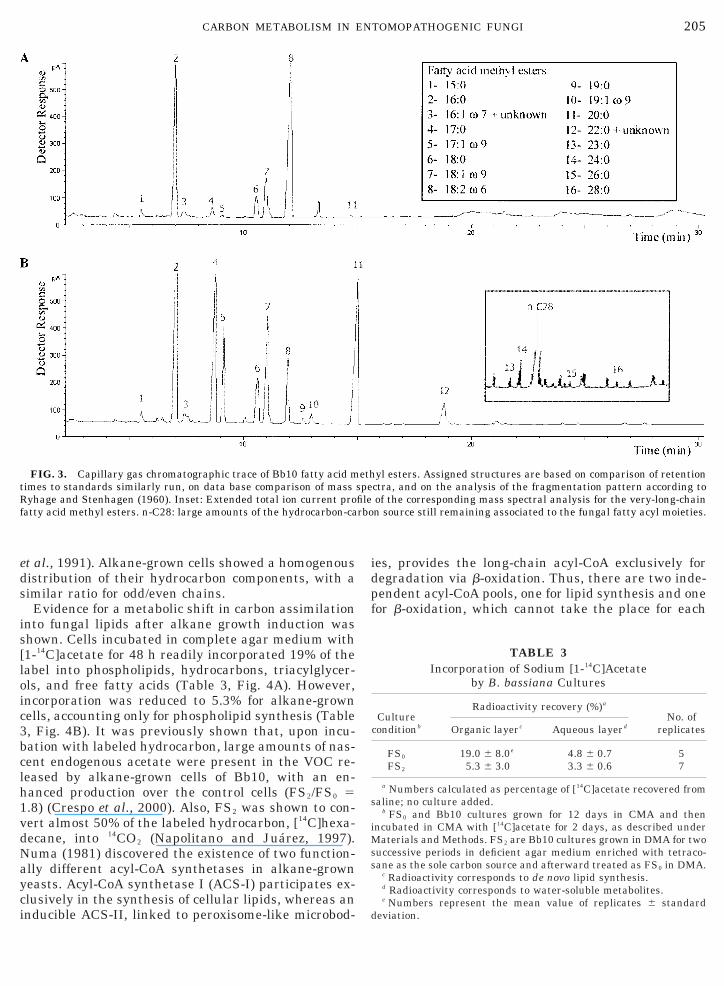
were the triacylglycerols followed by phospholipids.The hydrocarbon spot corresponded mostly to the n-C28 supplemented as carbon source (Fig. 2). Phos-phatidylcholine was the major component of the phos-pholipid fraction, both for FS0 and for FS2 cultures(data not shown). The analysis of Bb10 fatty acids inFS0 cultures showed a rather simple profile with lino-leic acid . palmitic acid .. oleic acid . stearic acid ashe major fatty acids (Fig. 3A). This pattern wasargely altered for FS2 cultures, with saturated chains
and palmitic, heptadecanoic, and eicosanoic acids asthe major components. In addition, the relativeamounts of the unsaturated fatty acids were alsochanged, with oleic acid . heptadecenoic acid . lino-eic acid (Fig. 3B). Saturated very-long-chain fatty ac-ds (VLCFA) of 22, 23, 24, 26, and 28 carbons were alsoetected, together with significant amounts of the sup-lemented carbon source, n-octacosane (n-C28) (Fig.B, inset). The large n-C28 peak was present even afterurification steps as described under Materials andethods, followed by hexane elution on a silicic acid
olumn.
1-14C]Acetate Incorporation into Fungal Cells
The relative incorporation of a readily accessible car-bon source, such as [1-14C]acetate, into the fungal cellmaterial is shown in Table 3 for FS0 and FS2 cultureconditions as described under Materials and Methods.Numbers represent the percentage of [14C]acetate re-overed from control plates similarly incubated with1-14C]acetate, but with no culture added. FS0 strains
were able to incorporate 23.8% of this easily ferment-able carbon source into their lipid components; how-ever, incorporation was reduced to 8.6% for FS2
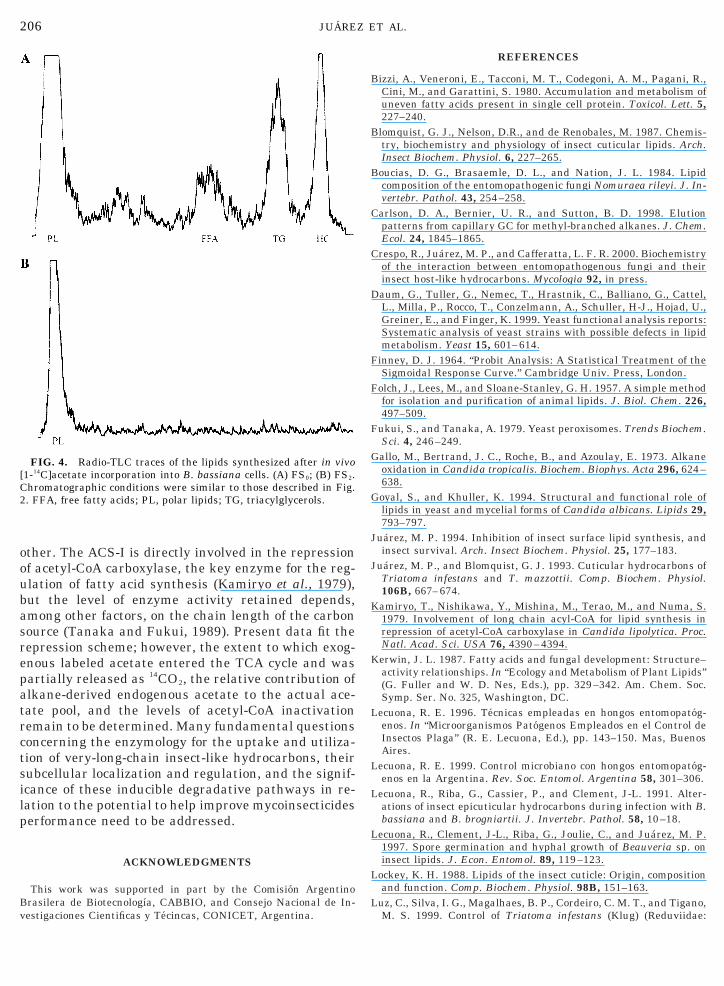
strains. Most of the label was utilized for lipid synthe-sis (19%) by FS0 strains, but the relative amounts ofwater-soluble metabolites produced were more rele-vant when fungi were grown on hydrocarbon as theonly carbon source (FS2 strains). Figure 4 shows the
istribution of radioactivity incorporated into differentipid fractions. After 48 h incubation, FS0 strains in-
1duction (CP) of Beauveria bassiana (Bb) and Metarhizium
an Lethal Time (MT) on Triatoma infestans Nymphs
6 SEM)CP (conidia 3108 6 SEM)d
Mortality(%) MLT (days 6 SEM)
0.18 2.50 6 0.016 100 11.01 6 0.050.09 2.46 6 0.044 100 7.70 6 0.080.13 0.13 6 0.100 100 5.80 6 0.10
ition.tomopathogenous Fungi, IMYZA, INTA, Castelar, Argentina. Ma6, Brazil.
LEProedi
cm
1 66 65 6
ondEn
ilia

FIG
.1.
Cap
illa
ryga
sch
rom
atog
raph
ictr
ace
ofB
b10
hyd
roca
rbon
s.N
um
bers
corr
espo
nd
tope
akn
um
bers
from
Tab
le2.
Ass
ign
edst
ruct
ure
sar
eba
sed
onm
ass
spec
trom
etri
can
alys
esac
cord
ing
toB
lom
quis
tet
al.(
1987
)an
don
data
base
and
rete
nti
onti
me
com
pari
son
tost
raig
ht
chai
nst
anda
rds.
202 JUAREZ ET AL.

a
203CARBON METABOLISM IN ENTOMOPATHOGENIC FUNGI
corporated newly synthesized fatty acids mostly intophospholipids and hydrocarbons; minor amounts weredetected in the free fatty acid and triacylglycerol frac-tions (Fig. 4A). In addition to a reduced ability to uti-lize this carbon source, FS2 strains incorporated thelabel solely into their phospholipid components (Fig.4B); similar experiments with FS1 cells showed identi-cal profiles (data not shown). The labeled spot wasscraped off the plate, solvent-extracted, and analyzedfor polar lipid distribution as detailed under Materialsand Methods; most of the label was detected in thephosphatidylethanolamine band, either for FS0 or forFS2 cultures.
DISCUSSION
The potential of entomopathogenic fungi for the con-trol of Triatominae vectors of Chagas disease has beenscarcely addressed. The high susceptibility of T. infes-tans to entomopathogenic fungi shown in this paper, ingreement with recent reports (Romana, 1995; Luz et
al., 1999), points to the need for further characteriza-tion of local strains to help the development and futureapplication of biological control tools for the erradica-tion of this vector in Argentina. Both local strains of B.
TABHydrocarbons f
Peak No. Hydrocarbon
1 9-Methyl octadecane2 n-Eicosane3 n-Heneicosane4 8-, 10-Methyl heneicosane5 n-Docosane6 11-Methyl docosane7 n-Tricosane8 9-Methyl tricosane9 n-Tetracosane
10 n-Pentacosane11 3,9,13-Trimethyl pentacosane plus 4,8- and 2,8-
dimethylpentacosane12 Hexacosane13 x,x-Dimethylhexacosane14 n-Heptacosane15 n-Octacosane16 4-Methyl plus 9,19-dimethyloctacosane17 n-Nonacosane18 2-Methyl nonacosane19 3-Methyl nonacosane plus 4,10-Dimethyl nonacosan20 3n-Triacontane21 n-Hentriacontane22 5,15-Dimethyl hentriacontane plus unknown23 n-Dotriacontane24 n-Tritriacontane25 n-Tetratriacontane26 n-Pentatriacontane27 n-Hexatriacontane28 n-Heptatriacontane29 n-Octatriacontane
a Bb10 strain.
bassiana showed similar germination times, conidiaproduction, and colony growth. However, the time re-quired to kill 100% of the insects was considerablyshorter for Bb10 (7.7 days); thus, this was the strainselected for further analyses. Although Ma6 had aMLT of 5.8 days, other parameters restricted furtheranalyses, such as slower growth and lower conidiaproduction than Bb strains and its exotic origin. For asuccessful fungal infection, various events must takeplace; adsorption and adhesion to the cuticular surface,fungal germination, and development of appresoria,prior to the penetration stage (Fargues, 1984). Thestudy of the biochemical interactions between the en-tomopathogenous fungi M. anisopliae and B. bassianaand their insect host-like hydrocarbons was previouslyaddressed at the INIBIOLP (Napolitano and Juarez,1997). Cuticular hydrocarbon degradation was shownto be a major metabolic pathway in alkane-grownfungi, related to peroxisomal proliferation (Crespo etal., 2000). Furthermore, alkane-grown B. bassiana cul-tures showed an enhanced speed of kill (M. P. Juarezand R. Crespo, unpublished). Growth on alkanescaused major changes in the lipid pattern of Bb10. Themajor lipid fractions of fungi grown in complete agarmedium (FS0) were sterols, whereas large amounts of
2B. bassianaa
ECL Diagnostic ions
18.30 140/141, 154/15520.00 28221.00 29621.36 126/127, 154/15522.00 31022.40 168/169, 182/18323.00 32423.32 140/141, 190/19124.00 33825.00 35225.95 154/155, 196/197, 224/225, 264/265, 362/363,
392/393, 140/141, 264/265, 308/30926.00 36626.9527.00 38028.00 39428.51 364/365, 408 140/ 141, 154/155, 292/29329.00 40829.56 378/379, 40829.66 364/365, 422 168/169, 296/297,364/365, 392/39330.00 42231.00 43631.77 238/239, 252/253, 406/40732.00 45033.00 46434.00 47835.00 49236.00 50637.00 52038.00 534
LErom
e
dopadbIsD
FeaTr
204 JUAREZ ET AL.
triacylglycerols were evident for the alkane-grown FS2
cells (Fig. 2). Phospholipids and, to a lesser extent,small amounts of diacylglycerols, monoacylglycerols,free fatty acids, and hydrocarbons were detected underboth culture conditions. Growth of Bb10 on synthetic orinsect-derived hydrocarbons was shown to occur at alower rate, 1.5 to 47.7% conidia production, than thatof glucose-grown cells, and similar variation was alsodetected for Ma6 (Napolitano and Juarez, 1997). A slowgrowth rate of Candida albicans growing on n-alkaneswas proposed to be related to accumulation of lipid inyeasts (Ratledge, 1978), as well as alteration in theirlipid pattern (Sorkhoh et al., 1990). Also, evidence oftriacylglycerol degrading activity was observed for FS2;after a period of 15 days at 0–4°C, FS2 cultures showeda lipid profile similar to that of FS0 cultures; however,cold conditions did not alter the FS0 lipid profile (datanot shown). The rather simple fatty acid pattern ofBb10 (FS0), with palmitic, linoleic, and oleic acids ac-counting for .90% of the total fatty acids (Fig. 3A),similar to that reported for higher fungi (Kerwin, 1987;Boucias et al., 1984), was largely altered. The unsat-urated/saturated ratio was markedly diminished forFS2 cultures; in addition, the relative composition ofboth fractions was largely modified. Heptadecanoicand eicosanoic acids, in addition to the usual compo-nent, palmitic acid, accounted for most of the saturatedchains (Fig. 3B). The increase in the relative amountsof saturated chains might be explained either by aninduction of the elongation activity after de novo syn-
FIG. 2. High-performance thin layer chromatographic profiles ofS0 and FS2 lipids developed in hexane (100%), followed by hexane:thyl ether:acetic acid (80:20:1). Detection by heating with 5% sulfuriccid in ethanol. PL, polar lipids; MG, monoacylglycerols; ST, sterols;G, triacylglycerols; HC, hydrocarbons. The HC band in FS2 cells cor-esponds mostly to exogenous n-octacosane. The presence of minor
amounts of diacylglycerols in the ST band was determined by chroma-tography in ethyl ether:benzene:ethanol:acetic acid (40:50:2:0.2 v/v).
thesis of palmitic acid or, less probably, by a selectiverelease of intermediate-chain-length acids freed fromthe b-oxidation system. FS2 culture contained largeamounts of the odd-chain heptadecanoic and heptade-cenoic acids, together with the usual FAS product,palmitic acid. Large amounts of odd-chain fatty acidswere reported for Candida sp. yeasts growing on hy-
rocarbons (Bizzi et al., 1980). Fatty acid componentsf alkane-grown cells are present in a rather dynamicool; variations were detected in their relativemounts, which might be associated with the effectiveegree of adaptation of each individual batch colonyoth to the carbon source and to its microenvironment.n a systematic screening analysis of mutant yeasttrains with possible defects in lipid metabolism,aum et al. (1999) considered only changes .50% of
the control lipids as a significant alteration. Hydroxyand methyl-substituted fatty acids, as well as VLCFAincorporated into glycosphingolipids, were reported asminor extracellular fungal products, which might beassociated with changes in the composition of the car-bon source provided (Weete, 1976). The higher levels ofsaturated fatty acids in FS2 cultures could cause adecrease in membrane fluidity and, hence, affect per-meability parameters, enzyme activity, and nutrienttransport (Goyal and Khuller, 1994). Significantchanges in the composition of other microorganismlipids and their fatty acid components have been re-ported for alkane-grown cells, although different sub-strates were employed, with different trends in theiralteration pattern (Sorkhov et al., 1990). At this stage,comparisons are far from apparent; thus, growth on aninsect-like hydrocarbon-enriched medium leads tocomplex interactions, yet to be explained, in the role ofeach individual lipid class and its distribution on thecell membrane and/or accumulation in an internal de-pot, as well as their participation in hydrocarbon up-take and transport.
Small amounts of saturated very-long-chain fattyacids C21:0, C22:0, C24:0, C26:0, and C28:0 were alsoidentified by mass spectrometry (Fig. 3B, inset). How-ever, the trace amount of the VLCFA found indicatesthat, even if they are effectively released as acyl-CoAsduring the b-oxidation steps, the release is not a majorpathway, reinforcing our current understanding of fun-gal long-chain alkane degradation occurring essen-tially by complete b-oxidation reactions. The analysisof the fungal hydrocarbon profiles for both FS0 and FS2
culture conditions, might result in a similar scenario.The hydrocarbon profile of Bb10 showed a saturatedchain length distribution, with a marked prevalencefor straight chains, ranging from C18 to C37 in thecarbon skeleton, with n-C22 as the major component.Few reports on entomopathogenic fungal hydrocarbonsare available, although a strain of B. bassiana (Bb147)and another from B. brogniartii were also reportedshowing no preference for odd to even chains (Lecuona
1v
d
205CARBON METABOLISM IN ENTOMOPATHOGENIC FUNGI
et al., 1991). Alkane-grown cells showed a homogenousdistribution of their hydrocarbon components, with asimilar ratio for odd/even chains.
Evidence for a metabolic shift in carbon assimilationinto fungal lipids after alkane growth induction wasshown. Cells incubated in complete agar medium with[1-14C]acetate for 48 h readily incorporated 19% of thelabel into phospholipids, hydrocarbons, triacylglycer-ols, and free fatty acids (Table 3, Fig. 4A). However,incorporation was reduced to 5.3% for alkane-growncells, accounting only for phospholipid synthesis (Table3, Fig. 4B). It was previously shown that, upon incu-bation with labeled hydrocarbon, large amounts of nas-cent endogenous acetate were present in the VOC re-leased by alkane-grown cells of Bb10, with an en-hanced production over the control cells (FS2/FS0 5.8) (Crespo et al., 2000). Also, FS2 was shown to con-ert almost 50% of the labeled hydrocarbon, [14C]hexa-
decane, into 14CO2 (Napolitano and Juarez, 1997).Numa (1981) discovered the existence of two function-ally different acyl-CoA synthetases in alkane-grownyeasts. Acyl-CoA synthetase I (ACS-I) participates ex-clusively in the synthesis of cellular lipids, whereas aninducible ACS-II, linked to peroxisome-like microbod-
FIG. 3. Capillary gas chromatographic trace of Bb10 fatty acid mtimes to standards similarly run, on data base comparison of massRyhage and Stenhagen (1960). Inset: Extended total ion current profatty acid methyl esters. n-C28: large amounts of the hydrocarbon-ca
ies, provides the long-chain acyl-CoA exclusively fordegradation via b-oxidation. Thus, there are two inde-pendent acyl-CoA pools, one for lipid synthesis and onefor b-oxidation, which cannot take the place for each
TABLE 3Incorporation of Sodium [1-14C]Acetate
by B. bassiana Cultures
Cultureconditionb
Radioactivity recovery (%)a
No. ofreplicatesOrganic layerc Aqueous layerd
FS0 19.0 6 8.0e 4.8 6 0.7 5FS2 5.3 6 3.0 3.3 6 0.6 7
a Numbers calculated as percentage of [14C]acetate recovered fromsaline; no culture added.
b FS0 and Bb10 cultures grown for 12 days in CMA and thenincubated in CMA with [14C]acetate for 2 days, as described underMaterials and Methods. FS2 are Bb10 cultures grown in DMA for twosuccessive periods in deficient agar medium enriched with tetraco-sane as the sole carbon source and afterward treated as FS0 in DMA.
c Radioactivity corresponds to de novo lipid synthesis.d Radioactivity corresponds to water-soluble metabolites.e Numbers represent the mean value of replicates 6 standard
eviation.
yl esters. Assigned structures are based on comparison of retentionctra, and on the analysis of the fragmentation pattern according toof the corresponding mass spectral analysis for the very-long-chainn source still remaining associated to the fungal fatty acyl moieties.
ethspefilerbo
L
L
L
L
[C2
206 JUAREZ ET AL.
other. The ACS-I is directly involved in the repressionof acetyl-CoA carboxylase, the key enzyme for the reg-ulation of fatty acid synthesis (Kamiryo et al., 1979),but the level of enzyme activity retained depends,among other factors, on the chain length of the carbonsource (Tanaka and Fukui, 1989). Present data fit therepression scheme; however, the extent to which exog-enous labeled acetate entered the TCA cycle and waspartially released as 14CO2, the relative contribution ofalkane-derived endogenous acetate to the actual ace-tate pool, and the levels of acetyl-CoA inactivationremain to be determined. Many fundamental questionsconcerning the enzymology for the uptake and utiliza-tion of very-long-chain insect-like hydrocarbons, theirsubcellular localization and regulation, and the signif-icance of these inducible degradative pathways in re-lation to the potential to help improve mycoinsecticidesperformance need to be addressed.
ACKNOWLEDGMENTS
This work was supported in part by the Comision ArgentinoBrasilera de Biotecnologıa, CABBIO, and Consejo Nacional de In-vestigaciones Cientificas y Tecincas, CONICET, Argentina.
FIG. 4. Radio-TLC traces of the lipids synthesized after in vivo1-14C]acetate incorporation into B. bassiana cells. (A) FS0; (B) FS2.hromatographic conditions were similar to those described in Fig.. FFA, free fatty acids; PL, polar lipids; TG, triacylglycerols.
REFERENCES
Bizzi, A., Veneroni, E., Tacconi, M. T., Codegoni, A. M., Pagani, R.,Cini, M., and Garattini, S. 1980. Accumulation and metabolism ofuneven fatty acids present in single cell protein. Toxicol. Lett. 5,227–240.
Blomquist, G. J., Nelson, D.R., and de Renobales, M. 1987. Chemis-try, biochemistry and physiology of insect cuticular lipids. Arch.Insect Biochem. Physiol. 6, 227–265.
Boucias, D. G., Brasaemle, D. L., and Nation, J. L. 1984. Lipidcomposition of the entomopathogenic fungi Nomuraea rileyi. J. In-vertebr. Pathol. 43, 254–258.
Carlson, D. A., Bernier, U. R., and Sutton, B. D. 1998. Elutionpatterns from capillary GC for methyl-branched alkanes. J. Chem.Ecol. 24, 1845–1865.
Crespo, R., Juarez, M. P., and Cafferatta, L. F. R. 2000. Biochemistryof the interaction between entomopathogenous fungi and theirinsect host-like hydrocarbons. Mycologia 92, in press.
Daum, G., Tuller, G., Nemec, T., Hrastnik, C., Balliano, G., Cattel,L., Milla, P., Rocco, T., Conzelmann, A., Schuller, H-J., Hojad, U.,Greiner, E., and Finger, K. 1999. Yeast functional analysis reports:Systematic analysis of yeast strains with possible defects in lipidmetabolism. Yeast 15, 601–614.
Finney, D. J. 1964. “Probit Analysis: A Statistical Treatment of theSigmoidal Response Curve.” Cambridge Univ. Press, London.
Folch, J., Lees, M., and Sloane-Stanley, G. H. 1957. A simple methodfor isolation and purification of animal lipids. J. Biol. Chem. 226,497–509.
Fukui, S., and Tanaka, A. 1979. Yeast peroxisomes. Trends Biochem.Sci. 4, 246–249.
Gallo, M., Bertrand, J. C., Roche, B., and Azoulay, E. 1973. Alkaneoxidation in Candida tropicalis. Biochem. Biophys. Acta 296, 624–638.
Goyal, S., and Khuller, K. 1994. Structural and functional role oflipids in yeast and mycelial forms of Candida albicans. Lipids 29,793–797.
Juarez, M. P. 1994. Inhibition of insect surface lipid synthesis, andinsect survival. Arch. Insect Biochem. Physiol. 25, 177–183.
Juarez, M. P., and Blomquist, G. J. 1993. Cuticular hydrocarbons ofTriatoma infestans and T. mazzottii. Comp. Biochem. Physiol.106B, 667–674.
Kamiryo, T., Nishikawa, Y., Mishina, M., Terao, M., and Numa, S.1979. Involvement of long chain acyl-CoA for lipid synthesis inrepression of acetyl-CoA carboxylase in Candida lipolytica. Proc.Natl. Acad. Sci. USA 76, 4390–4394.
Kerwin, J. L. 1987. Fatty acids and fungal development: Structure–activity relationships. In “Ecology and Metabolism of Plant Lipids”(G. Fuller and W. D. Nes, Eds.), pp. 329–342. Am. Chem. Soc.Symp. Ser. No. 325, Washington, DC.
Lecuona, R. E. 1996. Tecnicas empleadas en hongos entomopatog-enos. In “Microorganismos Patogenos Empleados en el Control deInsectos Plaga” (R. E. Lecuona, Ed.), pp. 143–150. Mas, BuenosAires.
Lecuona, R. E. 1999. Control microbiano con hongos entomopatog-enos en la Argentina. Rev. Soc. Entomol. Argentina 58, 301–306.
ecuona, R., Riba, G., Cassier, P., and Clement, J-L. 1991. Alter-ations of insect epicuticular hydrocarbons during infection with B.bassiana and B. brogniartii. J. Invertebr. Pathol. 58, 10–18.
ecuona, R., Clement, J-L., Riba, G., Joulie, C., and Juarez, M. P.1997. Spore germination and hyphal growth of Beauveria sp. oninsect lipids. J. Econ. Entomol. 89, 119–123.
ockey, K. H. 1988. Lipids of the insect cuticle: Origin, compositionand function. Comp. Biochem. Physiol. 98B, 151–163.
uz, C., Silva, I. G., Magalhaes, B. P., Cordeiro, C. M. T., and Tigano,M. S. 1999. Control of Triatoma infestans (Klug) (Reduviidae:

M
N
N
R
R
R
R
R
R
S
T
W
W
207CARBON METABOLISM IN ENTOMOPATHOGENIC FUNGI
Triatominae) with Beauveria bassiana (Bals.)Vuill.: Preliminaryassays on formulation and application in the field. Ann. Soc. En-tomol. Brazil 28, 101–110.
Makula, R., and Finnerty, W. R. 1968. Microbial assimilation ofhydrocarbons. J. Bacteriol. 95, 2102–2107.oore, D., Bateman, R. P., Carey, M., and Prior, C. 1995. Long-termstorage of Metarhizium flavoviride conidia in oil formulations forthe control of locusts and grasshoppers. Biocontrol Sci. Technol. 5,193–199.apolitano, R., and Juarez, M. P. 1997. Entomopathogenous fungidegrade epicuticular hydrocarbons of Triatoma infestans. Arch.Biochem. Biophys. 344, 208–214.uma, S. 1981. Two long chain acyl-CoA synthetases: Their differentroles in fatty acid metabolism and its regulation. Trends Biochem.Sci. 6, 113–115.atledge, C. 1978. Degradation of aliphatic hydrocarbons. In “Devel-opments in Biodegradation of Hydrocarbons” (R. J. Natkinson,Ed.), pp. 1–46, Appl. Sci., London.oberts, D. W., and Hajek, A. E. 1992. Entomopathogenic fungi asbioinsecticides. In “Frontiers in Industrial Mycology” (G. J.Leatham, Ed.), pp. 145–159. Chapman & Hall, New York.omana, C. A. 1995. Potencial del control biologico de los triatomi-
nos. In “International Workshop on Population Genetics and Con-trol of Triatominae” (C. J. Schofield, J. P. Dujardin, and J. Jurberg,Eds.), pp. 87–88. Indre, Mexico.omana, C. A., and Fargues, J. 1987. Sensibilite des larves del’hemiptere hemathophage Rhodnius prolixus aux hyphomycetesentomopathogenes. Entomophaga 32, 167–179.
omana, C. A., and Fargues, J. 1992. Relative susceptibility of dif-ferent stages of Rhodnius prolixus to the entomopathogenic hy-phomycete Beauveria bassiana. Mem. Inst. O. Cruz 87, 363–368.
yhage, R., and Stenhagen, E. 1960. Mass spectrometry in lipidresearch. J. Lipid Res. 1, 361–370.
orkhoh, N. A., Ghannoum, M. A., Ibrahim, A. S., Stretton, R. J., andRadwan, S. S. 1990. Growth of Candida albicans on hydrocarbons,lipids and sterols. Microbios 64, 159–171.
anaka, A., and Fukui, S. 1989. Metabolism of n-alkanes. In “TheYeasts” (A. Tanaka and S. Fukui, Eds.) Vol. 3, 2nd ed., pp. 261–287. Academic Press, New York.eete, J. D. 1976. Algal and fungal waxes. In “Chemistry and Bio-chemistry of Natural Waxes” (P. E. Kolattukudy, Ed.), pp. 290–334. Elsevier, Amsterdam.HO. 1991. “Control of Chagas Disease.” WHO Technical ReportSeries 811, World Health Organization, Geneva.
Copyright © 2022 FDOKUMEN




















