
Taxonomy and biogeography of the African Pygmy mice, Subgenus Nannomys (Rodentia, Murinae, Mus ) in...
16
Mammalia 72 (2008): 237–252 2008 by Walter de Gruyter • Berlin • New York. DOI 10.1515/MAMM.2008.035 Article in press - uncorrected proof 2007/35 Taxonomy and biogeography of the African Pygmy mice, Subgenus Nannomys (Rodentia, Murinae, Mus) in Ivory Coast and Guinea (West Africa) Ste ´ phane Kan Kouassi 1 , Violaine Nicolas 2 , Vladimir Aniskine 3 , Aude Lalis 2 , Corinne Cruaud 4 , Arnaud Couloux 4 , Marc Colyn 5 , Mireille Dosso 1 , Lamine Koivogui 6 , Erik Verheyen 7 , Chantal Akoua-Koffi 1 and Christiane Denys 2, * 1 Institut Pasteur de Co ˆ te d’Ivoire, 01 BP490, Abidjan, Ivory Coast 2 De ´ partement Syste ´ matique et Evolution, UMR5202CNRS – MNHN, 55, rue Buffon, 75005 Paris, France, e-mail: [email protected] 3 Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, Leninskii pr 33, 117071 Moscow, Russia 4 Genoscope, Centre National de Se ´ quenc ¸ age, 2, rue Gaston Cre ´ mieux, CP5706, 91057 Evry Cedex, France 5 Projet Fie ` vres He ´ morragiques en Guine ´ e (PFHG), CHU Donka, Conakry, Guinea 6 University of Rennes I, Station Biologique de Paimpont, 35380 Paimpont, France 7 Brussels University Royal Belgian Institute of Natural Sciences, Vautierstra ¨ at 29, 1000, Brussels, Belgium *Corresponding author Abstract Recent investigations in Guinea and Ivory Coast showed that Mus (Nannomys) are hosts of new arenaviruses. However, the taxonomy and biogeography of the western pygmy mice are poorly known, as this genus contains many cryptic species. To improve our taxonomic know- ledge on this subgenus and its distribution range in West Africa, we performed a molecular, cytogenetical and mor- phological study of all Mus (Nannomys) species in this part of Africa. This approach allowed us to detect five species in Guinea (Mus mattheyi, M. minutoides, M. setu- losus, M. baoulei, M. musculoides) and four (M. minu- toides, M. setulosus, M. baoulei, M. musculoides) in Ivory Coast. Our molecular results confirm the validity of M. baoulei and the extended distribution range of M. mattheyi. We also observed a high degree of genetic dif- ferentiation between and within these species, especially within the M. setulosus clade which may be a species complex. The canonical analyses on the craniometrical and external morphological features of the sequenced specimens are insufficient to perfectly separate the stud- ied species. Nevertheless, our results confirm that M. setulosus and M. baoulei are the largest Mus species that occur in Guinea and Ivory Coast. M. baoulei can be distinguished by its short tail and M. setulosus by an enlarged and bilobated lower molar (M3). Among the smaller species, M. musculoides is difficult to identify using morphological data alone, but differences in tail length proportions, size of tympanic bullae, the length and shape of the upper dental row allow identification of this species with some confidence. Our results allow us to provide some conclusions about the distribution rang- es of the studied Mus (Nannomys) spp. in both countries and to discuss their presence in relation to vegetation, habitat and human settlements. Keywords: cytogenetics; DNA sequencing; morphometrics; Mus; West Africa. Introduction According to Myers et al. (2000) and Kuper et al. (2004), Ivory Coast and Guinea belong to the Guineo-Congolese biodiversity hot spot. In both countries, there is a North to South gradient of aridity that results in a wide region that gradually changes from forests to savannas (White 1986). Moreover, the increasing human population den- sity in this region was accompanied by deforestation to create new areas suitable for human settlement. In these areas, murines and humans inevitably interact, hence creating the danger that pathogens can be transmitted that may cause disease in humans. For example, recent studies demonstrated that several murines in West Africa are host reservoirs for new arena- and hantaviruses (Lecompte et al. 2006, 2007, Klempa et al. 2006). Tropical Africa harbors a very rich rodent diversity that is poorly known due to the existence of numerous sibling species (Taylor 2000, Dobigny et al. 2002, Granjon and Dobigny 2003). Related to the rats and mice of the sub- family Murinae, the pygmy mice of the genus Mus (sub- genus Nannomys) are endemic to sub-Saharan Africa. These small mice are known to occur all over Africa, and currently 19 species are recognized (Musser and Carle- ton 2005). Despite recent studies that implement karyo- logical and molecular methods (Castiglia et al. 2002, Veyrunes et al. 2004, 2005), the number of species within the subgenus Nannomys remains uncertain. As a con- sequence of our poor taxonomical knowledge of this subgenus, the geographical distribution ranges of the Nannomys species remain unclear. We used a combined multidisciplinary approach to improve our knowledge on the taxonomy of the Nanno- mys species in Guinea and Ivory Coast and to use this information to infer their distribution ranges. Because the
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Taxonomy and biogeography of the African Pygmy mice, Subgenus Nannomys (Rodentia, Murinae, Mus ) in...

Mammalia 72 (2008): 237–252 � 2008 by Walter de Gruyter • Berlin • New York. DOI 10.1515/MAMM.2008.035
Article in press - uncorrected proof
2007/35
Taxonomy and biogeography of the African Pygmy mice,Subgenus Nannomys (Rodentia, Murinae, Mus) inIvory Coast and Guinea (West Africa)
Stephane Kan Kouassi1, Violaine Nicolas2,Vladimir Aniskine3, Aude Lalis2, CorinneCruaud4, Arnaud Couloux4, Marc Colyn5,Mireille Dosso1, Lamine Koivogui6, ErikVerheyen7, Chantal Akoua-Koffi1 andChristiane Denys2,*1 Institut Pasteur de Cote d’Ivoire, 01 BP490, Abidjan,Ivory Coast2 Departement Systematique et Evolution,UMR5202CNRS – MNHN, 55, rue Buffon, 75005 Paris,France, e-mail: [email protected] Severtsov Institute of Ecology and Evolution, RussianAcademy of Sciences, Leninskii pr 33, 117071Moscow, Russia4 Genoscope, Centre National de Sequencage, 2, rueGaston Cremieux, CP5706, 91057 Evry Cedex, France5 Projet Fievres Hemorragiques en Guinee (PFHG), CHUDonka, Conakry, Guinea6 University of Rennes I, Station Biologique dePaimpont, 35380 Paimpont, France7 Brussels University Royal Belgian Institute of NaturalSciences, Vautierstraat 29, 1000, Brussels, Belgium
*Corresponding author
Abstract
Recent investigations in Guinea and Ivory Coast showedthat Mus (Nannomys) are hosts of new arenaviruses.However, the taxonomy and biogeography of the westernpygmy mice are poorly known, as this genus containsmany cryptic species. To improve our taxonomic know-ledge on this subgenus and its distribution range in WestAfrica, we performed a molecular, cytogenetical and mor-phological study of all Mus (Nannomys) species in thispart of Africa. This approach allowed us to detect fivespecies in Guinea (Mus mattheyi, M. minutoides, M. setu-losus, M. baoulei, M. musculoides) and four (M. minu-toides, M. setulosus, M. baoulei, M. musculoides) in IvoryCoast. Our molecular results confirm the validity ofM. baoulei and the extended distribution range of M.mattheyi. We also observed a high degree of genetic dif-ferentiation between and within these species, especiallywithin the M. setulosus clade which may be a speciescomplex. The canonical analyses on the craniometricaland external morphological features of the sequencedspecimens are insufficient to perfectly separate the stud-ied species. Nevertheless, our results confirm thatM. setulosus and M. baoulei are the largest Mus species
that occur in Guinea and Ivory Coast. M. baoulei can bedistinguished by its short tail and M. setulosus by anenlarged and bilobated lower molar (M3). Among thesmaller species, M. musculoides is difficult to identifyusing morphological data alone, but differences in taillength proportions, size of tympanic bullae, the lengthand shape of the upper dental row allow identification ofthis species with some confidence. Our results allow usto provide some conclusions about the distribution rang-es of the studied Mus (Nannomys) spp. in both countriesand to discuss their presence in relation to vegetation,habitat and human settlements.
Keywords: cytogenetics; DNA sequencing;morphometrics; Mus; West Africa.
Introduction
According to Myers et al. (2000) and Kuper et al. (2004),Ivory Coast and Guinea belong to the Guineo-Congolesebiodiversity hot spot. In both countries, there is a Northto South gradient of aridity that results in a wide regionthat gradually changes from forests to savannas (White1986). Moreover, the increasing human population den-sity in this region was accompanied by deforestation tocreate new areas suitable for human settlement. In theseareas, murines and humans inevitably interact, hencecreating the danger that pathogens can be transmittedthat may cause disease in humans. For example, recentstudies demonstrated that several murines in West Africaare host reservoirs for new arena- and hantaviruses(Lecompte et al. 2006, 2007, Klempa et al. 2006).
Tropical Africa harbors a very rich rodent diversity thatis poorly known due to the existence of numerous siblingspecies (Taylor 2000, Dobigny et al. 2002, Granjon andDobigny 2003). Related to the rats and mice of the sub-family Murinae, the pygmy mice of the genus Mus (sub-genus Nannomys) are endemic to sub-Saharan Africa.These small mice are known to occur all over Africa, andcurrently 19 species are recognized (Musser and Carle-ton 2005). Despite recent studies that implement karyo-logical and molecular methods (Castiglia et al. 2002,Veyrunes et al. 2004, 2005), the number of species withinthe subgenus Nannomys remains uncertain. As a con-sequence of our poor taxonomical knowledge of thissubgenus, the geographical distribution ranges of theNannomys species remain unclear.
We used a combined multidisciplinary approach toimprove our knowledge on the taxonomy of the Nanno-mys species in Guinea and Ivory Coast and to use thisinformation to infer their distribution ranges. Because the

238 S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa
Article in press - uncorrected proof
number of specimens in most collections is too low toreliably distinguish whether observed differences repre-sent intraspecific variation or interspecific differences, itis often problematic to correctly identify these species.To overcome this problem, we collected new specimensand analyzed their standard karyotypes and/or cyto-chrome b (cytb) sequences. The combination of thesedata, with the morphological features of the same speci-mens, and the comparison of these data with relevanttype specimens or with topo-types, allowed us to reliablyidentify the species. The subsequent morphometric anal-yses allowed us to reveal the distinguishing morpholog-ical features of these species. This approach alsoallowed us to precisely identify the distribution ranges ofsome species, identify their habitat preferences andwhether they are commensal species to humans.
Background studies on West African Nannomys
The taxonomy of Nannomys has always been problem-atic because of the small size of these species (from 3 gto 20 g). Moreover, these mice are not easily capturedand as a result only few studies have investigated thesespecies in West and Central Africa. Rosevear (1969) pro-vided a key to distinguish between Mus setulosus,M. haussa and M. musculoides. M. musculoides andM. haussa are described as the two smaller species witha hind foot (HF) -14 mm, skull length -20 mm, uppermolar rows -3.5 mm and an upper M2 with only a t1.M. setulosus is described as a larger species of whichthe upper M2 is characterized by a very small antero-external cusp. Petter (1963, 1969) and Petter et al. (1971)combined cytogenetic data with cranial and dental char-acters to clarify the taxonomy of the West African Nan-nomys. They described M. mattheyi from Ghana Accra(Petter 1969), and provided several morphological criteriato distinguish M. mattheyi from the Nannomys species ofthe so-called sorella group (Petter 1963, Petter et al.1971). The most recently described species is M. baouleiVermeiren and Verheyen (Vermeiren and Verheyen 1980),which was described from Lamto (Ivory Coast) and wasdiagnosed on the basis of a series of morphological char-acters: a red mark close to the ear opening, a short tail,a narrow rostrum, orthodontic upper incisors, long incisorforamina, large molars and a big isolated distal t1 on theupper M1. Nevertheless, these authors indicated thatdespite the mentioned differences this taxon can beconfused with M. setulosus and in particular with M.musculoides (minutoides-musculoides complex). Thecytogenetical studies by Matthey (1966, 1967, 1970a,b),Jotterand (1972) and Jotterand-Bellomo (1984, 1986)showed numerous Robertsonian fusions that result in alarge amount of chromosomal polymorphism in the minu-toides-musculoides complex. This observation, andhence the validity of M. musculoides, was contested byseveral authors who amalgamated this species complexunder M. minutoides (see review in Veyrunes et al. 2004,2005). A general molecular phylogeny of Mus by Chevretet al. (2005) confirmed the validity of the African pygmymouse subgenus (Nannomys) and found M. setulosus tobe an early African offshoot. Recent molecular and band-ing analyses of western and southern African specimensof the minutoides-musculoides complex by Veyrunes etal. (2004, 2005) clarified the situation. These authors
showed that M. minutoides occurs in South Africa, Tan-zania, Kenya and Guinea, while M. musculoides occursin Mali and in the West African type locality (vaguelydescribed as ‘‘Gold Coast’’) that is supposed to be sit-uated in Ghana rather than Guinea. The same study indi-cates that (1) M. indutus is a valid species that onlyoccurs in South Africa, (2) M. mattheyi occurs in Mali,Senegal, Burkina Faso and Togo, (3) M. haussa is restrict-ed to the relatively dry habitats in Mali, Niger, Chad andSenegal, (4) M. setulosus occurs in Gabon, Central Afri-can Republic and Guinea, and (5) South Chad is inhab-ited by an unnamed Nannomys species. Table 1summarizes the available cytogenetic results for Mus(Nannomys) species from Guinea and Ivory Coast.
Mus minutoides from Adiopodoume (Ivory Coast) hasa 2ns32–34, FNs36 and M. musculoides has a 2ns36,FNs36 (Matthey 1970a,b). The chromosomal formula ofM. baoulei from Foro savanna (Ivory Coast) is 2ns20/21(FN unknown) (M. Tranier and J.C. Gautun unpublisheddata, cited in Musser and Carleton 2005). The karyotypeof M. mattheyi was described with 2ns36, FNs36 inKafine and Bouake, while M. minutoides with 2ns34, FNunknown was described for Bouake (Ivory Coast) (R.Matthey unpublished data). M. mattheyi was found inBobo-Diolasso (Burkina Faso) by Matthey (1970a), and inBandia, Saboya and Niokolo Koba (Senegal) by Veyruneset al. (2004). There are no published karyotype data thatconfirm the presence of M. minutoides in Guinea, but ithas been cited by V. Aniskine (C. Denys et al. unpublish-ed data) and identified through DNA cytb sequencing intwo localities of Guinea by Denys et al. (2005), Lecompteet al. (2007) and Veyrunes et al. (2005). The presence ofother species, such as M. haussa or M. mattheyi, hasnever been confirmed by chromosomes or DNAsequencing in Guinea, despite the confirmed presenceof this taxon in neighboring countries, such as Mali, Sen-egal and Burkina Faso (Matthey 1970a, Veyrunes et al.2004). Matthey (1967) determined the chromosome for-mula of the Accra type specimen of M. mattheyi as being2ns36, FNs36, with primitive sexual chromosomes. ForM. setulosus from the Central African Republic and IvoryCoast, differences in length for some chromosomes havebeen reported (Jotterand-Bellomo 1986). According tothe latter author, the differentiation of chromosomes 16and 17 observed in M. setulosus could be at the originof the species isolation from an ancestral Mus genusstock, which was later confirmed by Catzeflis and Denys(1992), Chevret et al. (2005) and Veryunes et al. (2005).Musser and Carleton (2005) recorded the presence ofM. baoulei in the old Guinea MNHN (Museum of NaturalHistory, Paris) collections based upon morphologicalgrounds. However, the validity of this species was neverconfirmed by chromosomal or molecular data, and itshould be confirmed by additional data.
Material and methods
Over a period of 3 years (2003–2006) 105 new Mus spec-imens were collected in Guinea (14 localities) and IvoryCoast (10 localities, see Figure 1). They were capturedusing Sherman traps, Pitfall traps and by hand, in bothcultured and natural habitats (Denys et al. 2005). All the

S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa 239
Article in press - uncorrected proof
Table 1 List of type localities and chromosomal data for the five West African species recognized in Musser and Carleton (2005).
Species Authors Distribution Type Chromosome Referenceslocality formula
Mus baoulei Vermeiren and Lamto 2ns20/21 Unpublished dataVerheyen 1980 Ivory Coast (Trainier M. and Gautun J.C.)
Mus haussa Thomas and Hinton Farniso 2ns28–34, Dobigny et al. 20021920 Nigeria FNs38, Veyrunes et al. 2004
X acro
Mus mattheyi Petter 1969 Accra 2ns36 FNs36 Jotterand-Bellomo 1986Ghana Veyrunes et al. 2004
Mus minutoides Smith 1834 Cape Town 2ns18–36 Veyrunes et al. 2004RSA FNs34,
X.1
Mus musculoides Temminck 1853 Gulf of Guinea 2ns18–19 Veyrunes et al. 2004, 2005Coast FNs36, X.7
Mus setulosus Peters 1876 Limbe 2ns36, Jotterand-Bellomo 1986Cameroon FNs36,
X acro, small Y
Chromosomal data (2nsnumber of chromosome pairs, FNsfundamental number) are from the synthesis by Veyrunes et al. (2004,2005). X.1sRobertsonian translocation between chromosome X and 1. X.7sRobertsonian translocation between chromosome X and7. acrosacrocentric.
Figure 1 Geographical distribution of Mus (Nannomys) sampled localities from Guinea and Ivory Coast for this study. Blackcirclessnew localities of collections searched for this study. White circlessold specimens from the collections of the MNHN.
captured animals were autopsied under P3 security pro-tocols adapted from Mills et al. (1995). External meas-urements wweight (W), head and body length (HB), taillength (TL), ear length (E) and hind foot length (HF)x weretaken during autopsies. Organs (liver, muscles) were keptin 90% ethanol for molecular studies, and the carcasseswere fixed in formalin for later preparation as skin andskull specimens.
One member of our group (V.A.) performed standardkaryotype analysis of 15 individuals from 6 localities inthe field using the colchicine method described by Leeand Elder (1980) (see Appendix A).
DNA was extracted for 81 specimens from ethanol-preserved muscles or liver by the CTAB (cetyltrimethyl-
ammoniumbromide) method (Winnepenninckx et al.1993). For all specimens, a large part of the cytochromeb gene was amplified, using PCR primers L7(59-ACCAATGACATGAAAAATCATCGTT-39), H14896 (59-TAGTTGTCGGGGTCTCCTA-39) and H15915 (59-TCTCCATTTCTGGTTTACAAGAC-39). The PCR consistedof 35 cycles: 30 s at 948C, 40 s at 51–558C and 90 s at728C. The double-stranded PCR product was purifiedand was run using an Automatic Sequencer ABI 3730(Abi, Ramsey, Minnesota, USA). Sequencing was con-ducted under BigDye� (Abi, Ramsey, Minnesota, USA)terminator cycling conditions. A 744-bp long part of thecytochrome b gene was retained for the analyses, as itwas available for all specimens. Haplotypes were iden-

240 S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa
Article in press - uncorrected proof
Figure 2 Standard karyotype of male Mus mattheyi fromNorthwest Guinea ALT47 (M) (2ns36, FNs36).
tified using TCS version 1.13 (Clement et al. 2000). Phy-logenetic relationships among haplotypes wereestimated using neighbor-joining (NJ) analyses imple-mented in PAUP 4.0b10 (Swofford 2002), and BayesianMarkov-chain Monte Carlo phylogenetic analyses(MCMC) implemented in MrBAYES version 3.1 (Huelsen-beck and Ronquist 2001, Ronquist and Huelsenbeck2003). We estimated the robustness of internal nodes by1000 bootstrapping replicates in NJ analyses. The com-puter program MODELTEST 3.04 (Posada and Crandall1998) was used to evaluate the fit of 56 nested modelsof nucleotide substitution to the data. Likelihood-ratiotests and the Akaike information criterion are commonlyused to evaluate which model best fits the data. We usedthe Akaike information criterion, as it was recently shownto have several advantages over likelihood-ratio tests(Sullivan and Joyce 2005). The Akaike information crite-rion revealed that the General Time Reversible modelwith a gamma-distributed rate variation across sites anda proportion of invariables sites (GTRqIqG) was a bestfit for these data. MCMC analyses were conducted inMrBAYES version 3.1 with the model selected byMODELTEST. The program’s default priors for parame-ters of the MCMC analyses were used for all analyses.We used three heated chains and a single cold chain inall MCMC analyses, and initiated runs with random trees.We conducted two independent MCMC runs with 2 mil-lion generations per run. We sampled trees (and para-meters) every 100 generations. The runs were evaluatedby examining the average standard deviation of split fre-quencies. As the two runs converge onto the stationarydistribution, we expect the average standard deviation ofsplit frequencies to approach zero, reflecting the fact thatthe two tree samples become increasingly similar (Ron-quist et al. 2005). Moreover, the Potential Scale Reduc-tion Factor (PSRF) should approach 1 as runs converge.For each run, the first 25% of sampled trees were dis-carded as burn-in. The mean percentage of sequencedivergence within and between clades was calculated(Jukes Cantor) with the software DNASP (Rozas et al.2003).
For classical morphometric analyses, 5 external and 11craniometrical distance measurements were taken on the105 collected specimens, 96 of which were either karyo-typed and/or sequenced (Appendix A). Due to a lot ofbroken parts on skulls and to increase the sample sizefor the morphological measurements, we added 42 spec-imens of the MNHN collections from Seredou (Guinea),Kerouane (Guinea), Adiopodoume (Ivory Coast), Faraba-na (Mali), Foro (Ivory Coast), Bouake (Ivory Coast),Damantan (Senegal) and Boukoko, La Maboke (CentralAfrican Republic), Benin, Cameroon. In all cases thesespecimens were either assigned to a species throughprevious chromosomal or molecular works or confirmedmorphologically by G. Musser in Paris MNHN (especiallyfor baoulei and setulosus specimens) (Appendix A). Weperformed Canonical analyses (CA) using XLSTAT Soft-ware version 7.2 (Addinsoft, Paris, France). The cranio-metrical measurements used were similar to those inDenys and Tranier (1992). In dorsal view: LGT: greatestlength of the skull, WZYG: bizygomatic width, CIO: inter-orbital constriction width, LNAS=WNAS: length and
width of nasal bone, WBR: braincase maximal width. Inventral view: LS13: upper molar row length, LBT: lengthof tympanic bulla. On the mandible three distances weremeasured: LMDB: length of the mandible, HMDB: heightof the mandible, LI13: length of the lower molar row.
Results
Cytogenetics
We obtained three different Mus (Nannomys) karyotypesin Guinea. The first corresponds to Mus mattheyi with2ns36, FNs36 (Figure 2). It was detected in specimenscollected in North West Guinea and South central Guinea(Altou Fonkola, Tanganya, Bantou, Khoneya localities)close to the borders of Gambia, Senegal, Liberia andSierra Leone. The second karyotype was found in spec-imens from one locality in the west, close to the borderof Guinea Bissau (Kandiafara), and in the south westernpart of Guinea (Tanganya, Bantou) (Figure 3). The stan-dard karyotypes correspond to M. minutoides with avariable formula of 2ns32–34, FNs34. The G-bandingallowed identification of the chromosomes involved in thefusions and to show that the sex chromosomes are fusedto pair 1. Males were 2ns34 (BA704, KDF30), whilefemales displayed 2ns32 (TA777, TA815) due to twoRobertsonian fusions involving pairs 2 and 17, and 3 and5, both found in heterozygous state.
In the Guinean forests, in the locality of Sangassou, wefound one male and two females (SA15, SA33, SA41)with a third karyotype 2ns36 FNs36, characterized by asmall Y chromosome which we assigned to be identifiedas Mus setulosus, according to previous works by Mat-they (1970a) and Veyrunes et al. (2004) (Figure 4).
Molecular analysis
The molecular phylogeny inferred from the cytb sequen-ces of 82 original and 18 GenBank sequences displaysfive highly supported clades, each one corresponding toone species (Figure 5). The inclusion of sequences ofspecimens from previous studies that have already beenspecifically identified by molecular/cytogenetics methodswill help to link each clade to a species name. The meangenetic distances between species varies from 12% to13%, except in the minutoides-musculoides clade wheredistances reach only 9%.
Clades I and clade II are sister-clades. This monophy-letic group is highly supported and corresponds to theso-called minutoides-musculoides species complex.Clade I is subdivided into two highly supported sub-clades: the first one groups specimens from Guinea

S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa 241
Article in press - uncorrected proof
Figure 3 Standard karyotype of male Mus minutoides fromNorthwest Guinea (KDF30) (2ns34, FNs34) (top) and standardkaryotype of female Mus minutoides from Southwest Guinea(TA777) (2ns32, FNs34) (bottom).
Figure 4 Standard karyotype of male Mus setulosus: (SA15)(2ns36, FNs36).
(Kodoko) and Ivory Coast (Gbagroube, Lamto, Touzouk-ro, Blodi, Taı, Menekre, Bakro, Aohouakro), the secondone groups specimens from South Africa and Tanzania.In each of these subclades several karyotyped speci-mens allow us to provide names to some clades. Allthese karyotypes are similar to those typical of Musminutoides. However, the relatively high percentage ofgenetic divergence between these two subclades (5.7%)suggests that M. minutoides may represent species-complex of species. As the M. minutoides type comesfrom South Africa, it is possible that the West Africanrepresentative of minutoides represents an undescribedspecies.
Based on published karyotypes and sequences ofspecimens from Mali (Veyrunes et al. 2004, 2005), cladeII corresponds to Mus musculoides. This species wasalso found in our study in Guinea (Kodoko) and IvoryCoast (Sanguiebo).
Clade III includes specimens from Mali, Senegal andNorth West Guinea (Altou Fonkola, Boke). The one karyo-typed specimen in this clade displays a chromosomalformula (Figure 2) that assigns it to Mus mattheyi fromIvory Coast and Ghana.
Clade IV includes specimens from Forest Guinea(Bhoita, Sangassou, Gbetaya, Ziama, Diecke), SouthernIvory Coast (Bakro, Kotokossou, Gbagroubet), Came-roon, Gabon and Central African Republic. Two speci-mens were also karyotyped from Sangassou andGbetaya and have a chromosome formula that is closeto the one of Mus setulosus from Central African Repub-lic. The high genetic divergence within this clade (approx-imately 8%) suggests the existence of cryptic species in
the complex. It is interesting to note that very differenthaplotypes have been detected in a single locality inIvory Coast (e.g., Gbagroubet). The amount of geneticdifferentiation that we observe between central (Came-roon, Central African Republic, Gabon) and western Afri-can populations was previously detected by cytogeneticstudies between Central African Republic and IvoryCoast (Jotterand-Bellomo 1986). Unfortunately, no karyo-type is available from the M. setulosus holotype locality(Southwest Cameroon).
Clade V contains specimens from Benin, Guinea(Kodoko) and Ivory Coast (Lamto, Sanguiebo). Unfortu-nately, we did not karyotype any specimens of clade V.Further morphological analyses may provide clues to linkit to a species name.
Morphological and morphometrical analyses ongenetically analyzed specimens: puttingnames on clades
If for clades I to IV available sequences and karyotypesallowed us to provide a preliminary taxonomic attribution,this was not the case for clade V. This clade groups quitelarge specimens with a rather short tail (Table 2), a typicalorthodontic pattern, and an isolated t1 on the M1 (Figure6). These morphological characters correspond to Musbaoulei. This species was described from Lamto (IvoryCoast), from which one specimen was sequenced here.According to Musser and Carleton (2005), this speciesalso occurs in Guinea and in another locality of IvoryCoast (Foro) and was displaying an unusual karyotype(2ns20/21) (J.C. Gautun and M. Tranier unpublisheddata). Our molecular and morphological data confirm thevalidity of M. baoulei as a species from Ivory Coast (Lam-to) and Guinea (Kodoko), and allowed us to extend itsgeographical distribution to Central Benin.
Using specimens identified through chromosomal ormolecular data, we analyzed morphological and morpho-metrical characters to search for diagnostic criteria dis-tinguishing the West African species.
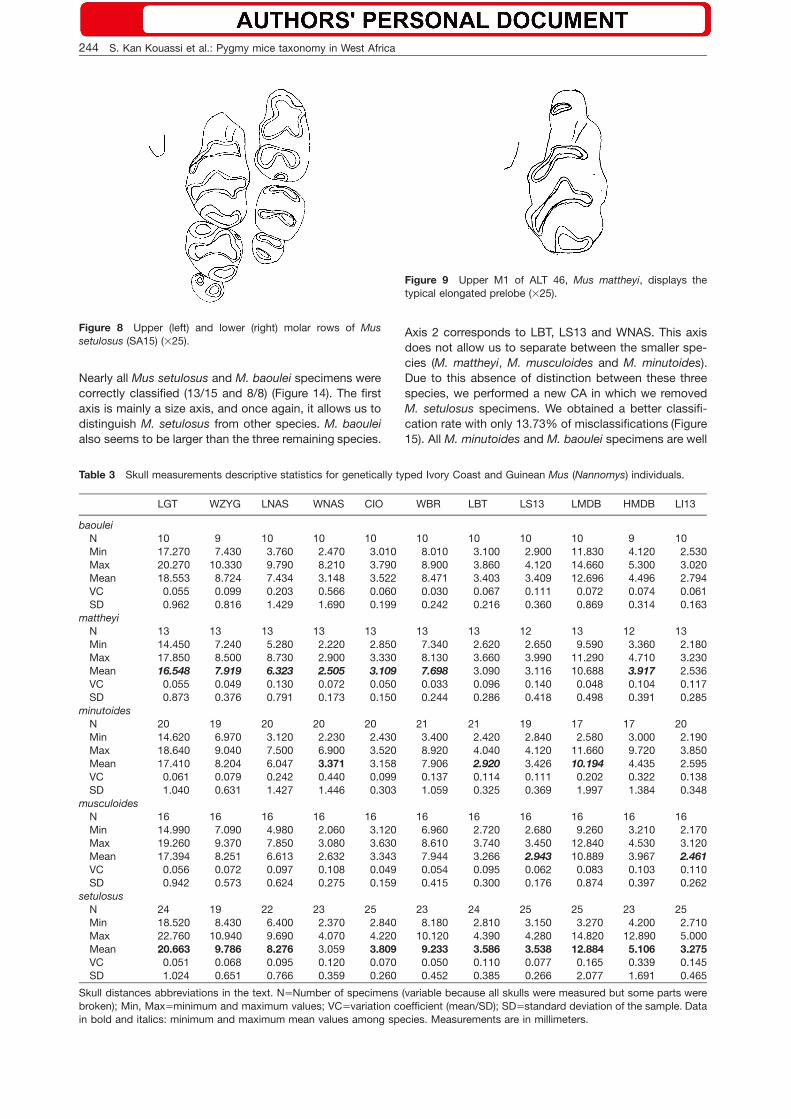
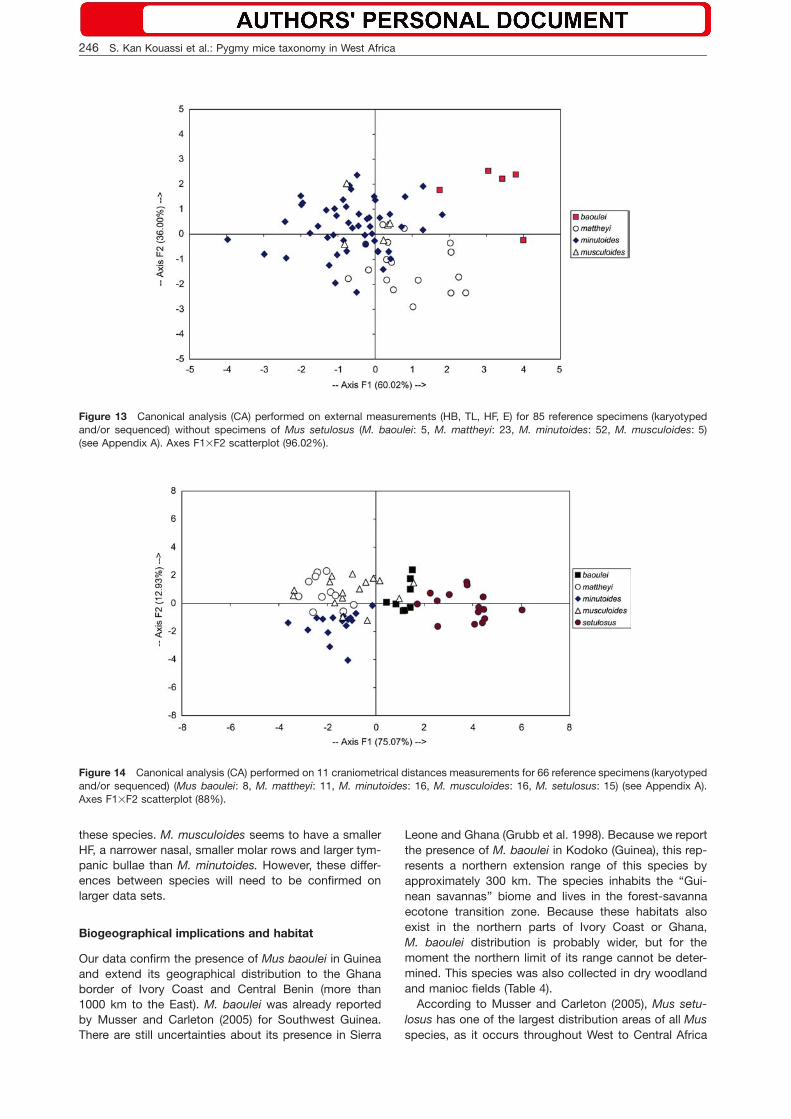
We considered molar morphology because these teethoften provide diagnostic characters for species identifi-cation within the genus Mus (Petter 1963, 1969). Forexample, M. setulosus and M. baoulei have a rather sim-ilar external morphology and both taxa are larger thanthe other West African species. However, the shape oftheir molars have a species-specific pattern: M. setulo-sus has a poorly reduced M3/3 and the lower M/3 dis-plays a two-lobe pattern which is not observed in allother West African taxa (Figures 7, 8). M. mattheyi spec-imens from Guinea have small HB and skull sizes, theyhave a long incisor foramen and display the same elon-gated prelobe of the upper M1/ as the holotype speci-men from Accra (Figure 9).
Mus minutoides and M. musculoides are very close inmolar morphology and size. They both display a muchreduced M3/3 and a relatively small size (Tables 2, 3 andFigures 10, 11). In M. minutoides, the incisor foramenstops more anteriorly than in M. musculoides where itreaches the t1 level.
To find further determination criteria, we performed acanonical analysis (CA) on four external measurementsof 99 genetically identified specimens of IC and Guinea

242 S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa
Article in press - uncorrected proof
Figure 5 Phylogeny recovered by neighbor-joining (NJ) phylogenetic analyses (GTR model). Values above branches indicate boot-strap support (NJ) and Bayesian posterior probabilities. To improve clarity, bootstrap values and posterior probabilities for the mostapical nodes are not written. Karyotyped specimens are in bold font. We included specimens already typed molecularly or cytoge-netically in order to assign clades to species. Abbreviations for Ivory Coast and Guinean localities: ahosAhouakro; altsAltou-Fonkola;baksBakro; bhsBhoita; blosBlodi; boksBoke; frsFranfina; gagsGagal; gbasGbagroubet; gbesGbetaya; kazsKazaouma; kdsKodoko; kdfsKandiafara; kotsKotokossou; lamsLamto; mensMenekre; SasSangassou; sansSanguiebo; taisTaı; tousTouzoukro;yossYossono; AFSsSouth Africa, KENsKenya; MALIsMali; TOGOsTogo; SENsSenegal; CAMsCameroon; CARsCentral AfricanRepublic; BENsBenin.
only. On the F1 axis (61.82% of variability), which isexplained by all external variables, Mus setulosus is thelargest species and M. mattheyi is the smallest one (Fig-ure 12). In this analysis, 25.25% of the specimens aremisidentified and for none of the species a 100% correctidentification was obtained. The best classified speciesis M. minutoides (48/52 well classified specimens). Thesecond axis of the analysis is partly explained by TL andseems to separate M. baoulei from the other species,due to its rather short tail in regard to its large HB (meanof HB: 66.8 mm versus mean of TL: 40 mm in M. baoulei,
against mean of HB: 74.25 versus 50.05 mm for TL in M.setulosus). When we removed M. setulosus specimensfrom the CA analysis the proportion of misidentifiedspecimens decreased to 20%, and again none of thespecies could be 100% correctly identified. On the firstaxis (Figure 13) there is an opposition between M. minu-toides and a group composed by M. mattheyi-M. baoulei.Specimens of M. musculoides are situated close to thecenter of the graph. M. minutoides has a longer tail(mean: 46 mm) compared to M. baoulei and M. mattheyi(means40 and 40.2 mm, respectively). The distinction

S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa 243
Article in press - uncorrected proof
Table 2 External measurements descriptive statistics for all genetically typed Ivory Coast and Guinean Mus (Nannomys) individualsplus specimens from Mali, Cameroon, Senegal cited in Appendix A.
Weight HB TL HF E
baouleiN 4 7 5 7 7Min 7.000 59.000 31.000 12.000 10.000Max 9.000 79.000 45.000 14.500 13.000Mean 7.750 68.286 40.000 13.286 10.929VC 0.124 0.089 0.137 0.092 0.100SD 0.829 5.599 4.899 1.129 1.015
mattheyiN 20 27 27 27 23Min 2.600 46.000 33.000 11.000 8.000Max 8.000 63.000 45.000 13.000 10.000Mean 4.500 54.444 40.370 12.056 9.261VC 0.288 0.080 0.095 0.044 0.069SD 1.262 4.298 3.763 0.515 0.623
minutoidesN 53 53 52 53 53Min 3.000 51.000 41.000 8.000 8.000Max 12.000 73.000 59.000 14.000 13.000Mean 6.840 62.660 47.269 12.423 9.708VC 0.289 0.085 0.081 0.102 0.111SD 1.957 5.291 3.773 1.261 1.070
musculoidesN 9 10 10 10 5Min 4.000 47.500 37.000 9.000 9.000Max 7.000 70.000 48.000 14.500 13.000Mean 5.644 58.850 44.700 12.450 10.800VC 0.226 0.124 0.070 0.141 0.190SD 1.202 6.928 2.968 1.665 1.833
setulosusN 24 34 30 34 21Min 6.000 60.000 40.000 11.000 10.000Max 16.000 88.000 63.000 17.500 15.000Mean 11.125 73.059 51.067 14.691 11.048VC 0.226 0.075 0.100 0.092 0.130SD 2.463 5.423 5.033 1.329 1.396
NsNumber of specimens (varying, depending on the preservation state of animals or collection labels completed); Min, Maxsmini-mum and maximum values; VCsvariation coefficient (mean/SD); SDsstandard deviation of the sample. Data in bold and italics:minimum and maximum mean values among species. All measurements are in millimeters.
Figure 6 Upper (left) and lower (right) molar rows of Musbaoulei (KD36) (=25).
Figure 7 Upper (left) and lower (right) molar rows of Mussetulosus (CIV433) (=25).
between M. musculoides and the other species is quitedifficult based on external characters, but it could be dueto our small sample sizes.
A second set of canonical analyses was performed onthe skull measurements of 66 genetically typed speci-mens. The rate of misclassification was now 19.7%.
244 S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa
Article in press - uncorrected proof
Figure 8 Upper (left) and lower (right) molar rows of Mussetulosus (SA15) (=25).
Figure 9 Upper M1 of ALT 46, Mus mattheyi, displays thetypical elongated prelobe (=25).
Table 3 Skull measurements descriptive statistics for genetically typed Ivory Coast and Guinean Mus (Nannomys) individuals.
LGT WZYG LNAS WNAS CIO WBR LBT LS13 LMDB HMDB LI13
baouleiN 10 9 10 10 10 10 10 10 10 9 10Min 17.270 7.430 3.760 2.470 3.010 8.010 3.100 2.900 11.830 4.120 2.530Max 20.270 10.330 9.790 8.210 3.790 8.900 3.860 4.120 14.660 5.300 3.020Mean 18.553 8.724 7.434 3.148 3.522 8.471 3.403 3.409 12.696 4.496 2.794VC 0.055 0.099 0.203 0.566 0.060 0.030 0.067 0.111 0.072 0.074 0.061SD 0.962 0.816 1.429 1.690 0.199 0.242 0.216 0.360 0.869 0.314 0.163
mattheyiN 13 13 13 13 13 13 13 12 13 12 13Min 14.450 7.240 5.280 2.220 2.850 7.340 2.620 2.650 9.590 3.360 2.180Max 17.850 8.500 8.730 2.900 3.330 8.130 3.660 3.990 11.290 4.710 3.230Mean 16.548 7.919 6.323 2.505 3.109 7.698 3.090 3.116 10.688 3.917 2.536VC 0.055 0.049 0.130 0.072 0.050 0.033 0.096 0.140 0.048 0.104 0.117SD 0.873 0.376 0.791 0.173 0.150 0.244 0.286 0.418 0.498 0.391 0.285
minutoidesN 20 19 20 20 20 21 21 19 17 17 20Min 14.620 6.970 3.120 2.230 2.430 3.400 2.420 2.840 2.580 3.000 2.190Max 18.640 9.040 7.500 6.900 3.520 8.920 4.040 4.120 11.660 9.720 3.850Mean 17.410 8.204 6.047 3.371 3.158 7.906 2.920 3.426 10.194 4.435 2.595VC 0.061 0.079 0.242 0.440 0.099 0.137 0.114 0.111 0.202 0.322 0.138SD 1.040 0.631 1.427 1.446 0.303 1.059 0.325 0.369 1.997 1.384 0.348
musculoidesN 16 16 16 16 16 16 16 16 16 16 16Min 14.990 7.090 4.980 2.060 3.120 6.960 2.720 2.680 9.260 3.210 2.170Max 19.260 9.370 7.850 3.080 3.630 8.610 3.740 3.450 12.840 4.530 3.120Mean 17.394 8.251 6.613 2.632 3.343 7.944 3.266 2.943 10.889 3.967 2.461VC 0.056 0.072 0.097 0.108 0.049 0.054 0.095 0.062 0.083 0.103 0.110SD 0.942 0.573 0.624 0.275 0.159 0.415 0.300 0.176 0.874 0.397 0.262
setulosusN 24 19 22 23 25 23 24 25 25 23 25Min 18.520 8.430 6.400 2.370 2.840 8.180 2.810 3.150 3.270 4.200 2.710Max 22.760 10.940 9.690 4.070 4.220 10.120 4.390 4.280 14.820 12.890 5.000Mean 20.663 9.786 8.276 3.059 3.809 9.233 3.586 3.538 12.884 5.106 3.275VC 0.051 0.068 0.095 0.120 0.070 0.050 0.110 0.077 0.165 0.339 0.145SD 1.024 0.651 0.766 0.359 0.260 0.452 0.385 0.266 2.077 1.691 0.465
Skull distances abbreviations in the text. NsNumber of specimens (variable because all skulls were measured but some parts werebroken); Min, Maxsminimum and maximum values; VCsvariation coefficient (mean/SD); SDsstandard deviation of the sample. Datain bold and italics: minimum and maximum mean values among species. Measurements are in millimeters.
Nearly all Mus setulosus and M. baoulei specimens werecorrectly classified (13/15 and 8/8) (Figure 14). The firstaxis is mainly a size axis, and once again, it allows us todistinguish M. setulosus from other species. M. baouleialso seems to be larger than the three remaining species.
Axis 2 corresponds to LBT, LS13 and WNAS. This axisdoes not allow us to separate between the smaller spe-cies (M. mattheyi, M. musculoides and M. minutoides).Due to this absence of distinction between these threespecies, we performed a new CA in which we removedM. setulosus specimens. We obtained a better classifi-cation rate with only 13.73% of misclassifications (Figure15). All M. minutoides and M. baoulei specimens are well

S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa 245
Article in press - uncorrected proof
Figure 10 Upper (left) and lower (right) molar rows of Musminutoides (CIV477) (=25).
Figure 11 Upper (left) and lower (right) molar rows of Musmusculoides (KD29) (=25).
Figure 12 Canonical analysis (CA) performed on external measurements (HB, TL, HF, E) for 99 reference specimens (karyotypedand/or sequenced) (Mus baoulei: 5, M. mattheyi: 11, M. minutoides: 52, M. musculoides: 5, M. setulosus: 26) (see Appendix A). AxesF1=F2 scatterplot (96.86%).
classified, while only 7/10 M. mattheyi and 13/16M. musculoides specimens are well classified. The graphof F1=F2 plan opposes specimens of M. minutoides onone side of axis 1 to M. baoulei at the other extremity.M. mattheyi and M. musculoides are difficult to discrim-inate both on axis 1 and 2. Axis 2 can be explained bythe upper molar length (LS13), the nasal and braincasewidth (WNAS, WBR).
Discussion
Summary of each species characteristics
In this study, we have linked species names to theobtained molecular groupings that were inferred fromDNA sequences and karyotypes. Whenever such datawere unavailable, a clade was named pending the out-come of a careful comparison of these specimens withall possible relevant type specimens. The results of ourmultivariate analyses are congruent with the morpholog-ical data of Rosevear (1969) and confirm that Mus setu-losus and M. baoulei are the largest species of pygmymice in West Africa. M. setulosus can be distinguishedfrom M. baoulei by its two-lobed lower M/3. Moreover,M. baoulei has a proportionally shorter TL and a lowermandibular height compared to M. setulosus.
It is possible to distinguish Mus mattheyi from all otherWest African pygmy mice based upon the morphologyof the upper M1, which is very elongated. Moreover,M. mattheyi can be distinguished from the M. minu-toides-musculoides species complex by its smaller bodyproportions. The distinction between M. musculoidesand M. minutoides is more difficult from a morphologicalpoint of view. On the basis of our specimens, teeth mor-phology, HB and TL are quite undistinguishable between
246 S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa
Article in press - uncorrected proof
Figure 13 Canonical analysis (CA) performed on external measurements (HB, TL, HF, E) for 85 reference specimens (karyotypedand/or sequenced) without specimens of Mus setulosus (M. baoulei: 5, M. mattheyi: 23, M. minutoides: 52, M. musculoides: 5)(see Appendix A). Axes F1=F2 scatterplot (96.02%).
Figure 14 Canonical analysis (CA) performed on 11 craniometrical distances measurements for 66 reference specimens (karyotypedand/or sequenced) (Mus baoulei: 8, M. mattheyi: 11, M. minutoides: 16, M. musculoides: 16, M. setulosus: 15) (see Appendix A).Axes F1=F2 scatterplot (88%).
these species. M. musculoides seems to have a smallerHF, a narrower nasal, smaller molar rows and larger tym-panic bullae than M. minutoides. However, these differ-ences between species will need to be confirmed onlarger data sets.
Biogeographical implications and habitat
Our data confirm the presence of Mus baoulei in Guineaand extend its geographical distribution to the Ghanaborder of Ivory Coast and Central Benin (more than1000 km to the East). M. baoulei was already reportedby Musser and Carleton (2005) for Southwest Guinea.There are still uncertainties about its presence in Sierra
Leone and Ghana (Grubb et al. 1998). Because we reportthe presence of M. baoulei in Kodoko (Guinea), this rep-resents a northern extension range of this species byapproximately 300 km. The species inhabits the ‘‘Gui-nean savannas’’ biome and lives in the forest-savannaecotone transition zone. Because these habitats alsoexist in the northern parts of Ivory Coast or Ghana,M. baoulei distribution is probably wider, but for themoment the northern limit of its range cannot be deter-mined. This species was also collected in dry woodlandand manioc fields (Table 4).
According to Musser and Carleton (2005), Mus setu-losus has one of the largest distribution areas of all Musspecies, as it occurs throughout West to Central Africa

S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa 247
Article in press - uncorrected proof
Figure 15 Canonical analysis (CA) performed on 11 craniometrical distances measurements for 51 reference specimens (karyotypedin bold font and/or sequenced) without Mus setulosus specimens (M. baoulei: 8, M. mattheyi: 10, M. minutoides: 17, M. musculoides:16) (see Appendix A). Axes F1=F2 scatterplot (84.10%).
Table 4 Habitat of the molecularly typed and karyotyped specimens for Ivory Coast and Guinea (the latter are indicated in boldfont).
Landscape setulosus minutoides musculoides baoulei matheyi
Rice fieldqrice fallowq 7q4 1q1cultivated swampBanana plantations 4q2 4Dry and open woodland 3 1 1Hevea plantation 2 2Bush inside village 5q1Houses 3Maize and egg-plant field 3Manioc field (Cassava) 2 5 1Ecotone 2 2Yam-egg-plant fields 8Primary forest 3Cocoa plantation 1Gallery forest 1 2Fallow land 1Total 14 47 3 4 4
up to Ethiopia. Our molecular data suggest that this spe-cies could represent a species complex. It is found inprimary and secondary forest, and even open fields, inSouth Ivory Coast and Southeast Guinea. Its northernlimits in Guinea and Ivory Coast seem to correspond tothe transition zone between forests and savannas.Despite a huge sampling effort it was never found in theFouta Djallon in Guinea, nor was it found in coastalGuinea in the west (C. Denys et al. unpublished data).Possibly, its western extension is limited by the Guineo-Congolese forest block. Dosso (1988) did not report thepresence of this species in the Taı forest (Ivory Coast),but it occurs in Macenta, Mount Nimba, Diecke and Zia-ma in Guinea (Heim de Balsac and Lamotte 1958, Roche1971, this study). The specimens from Macenta, Seredouand Mount Nimba attributed to M. setulosus were notkaryotyped, but an examination of skull and teeth ofspecimens preserved at MNHN confirm their presence (a
bilobated lower M/3 and a rather large body size with aHF)15 mm). Furthermore, the presence of this taxon isattested by karyotypes from Adiopodoume, while unka-ryotyped specimens (displaying the bilobated M/3) fromLamto are also attributed to M. setulosus. During our sur-vey the so-called Ivorian and Guinean M. cf. setulosushave also been recovered close to human habitats mainlyin plantations of secondary forest (Hevea, banana,cocoa, coffee fields, Table 4).
Mus minutoides seems to occur everywhere in IvoryCoast and Guinea which indicates that it cannot berestricted to a South African range previously claimed byMusser and Carleton (2005), and this is in agreement withVeyrunes et al. (2004, 2005). This species is one of thetropical African rodents with the largest distribution rang-es: from Cape Town to Central and East Guinea, whichseems to represent, for the moment, its western limit.However, the western and eastern lineages that we

248 S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa
Article in press - uncorrected proof
detected through our molecular analyses may indicatethat M. minutoides includes several cryptic species.M. minutoides is present in western Guinea (E. Lecompteet al. 2007, C. Denys et al. unpublished data, this work),in the Guinean Fouta Djallon plateau and in Eastern andCentral Guinea (Veyrunes et al. 2005). It has been foundin nearly all sampled localities from forest zones tosavannas and in open fields as well as houses (Table 4).Dosso (1988), Gautun et al. (1986) and Kadjo et al. (1996)have already recorded M. minutoides-musculoides in Tai,Adiopodoume, Lamto, and Bouake. Roche (1971) alsoindicated that M. minutoides-musculoides occurred inMacenta, Seredou and has anthropophilous affinities.Heim de Balsac and Lamotte (1958) indicated its capturein Mount Nimba and Gouecke zones. Karyotyped spec-imens of M. musculoides only occur from Senegal, Maliand Niger (Hubert et al. 1973, Granjon et al. 1992, Dobi-gny et al. 2002, Veyrunes et al. 2004, 2005).
In West Africa, Mus minutoides lives in sympatry withM. musculoides, but according to our study it seems thatM. musculoides has a more restricted distribution range:it occurs only in savanna regions of North Ivory Coastand Guinea, and in the transition zone between the Gui-nean and Sudanian zones (Kodoko, Sanguiebo) in rela-tively drier places. During our fieldwork, M. musculoideswas trapped in cultivated swamps, rice field fallows andbush (Table 4).
According to our data, Mus mattheyi occurs in North-west and Central Guinea (Altou Fonkola, Tanganya, Ban-tou) in dry Guinean savanna close to Niokolo KobaNational Park and in the transition zone between forestand savanna. Because the holotype comes from Accra(Ghana), this suggests a quite wide distribution of thisspecies which was also recorded in Senegal, Ivory Coast,Togo and Mali (Petter et al. 1971, Hubert et al. 1973,Duplantier and Granjon 1992, Grubb et al. 1998, Vey-runes et al. 2005). Our specimens fit well in the molecularphylogeny with Mali and Senegal specimens. Unfortu-nately, no specimen from the type locality wassequenced, but on karyological grounds there is no dif-ference between our Guinean samples and those fromthe Ghanean type locality. For these reasons we considerthat, contrary to Musser and Carleton (2005), M. mattheyimust not be restricted only to the type locality and is aWestern African savanna endemic species.
Diversity considerations
The present study revealed an unexpected Mus (Nan-nomys) spp. diversity with five species in Guinea and four(potentially five due to the wide genetic variabilityobserved in some localities for setulosus clade) speciesin Ivory Coast. This is rather high compared to speciescounts available for the surrounding countries. For exam-ple, only 2–3 Nannomys species have been reported inSenegal, Mali and Niger (Duplantier et al. 1997, Dobignyet al. 2002). We extend the distribution of Mus baoulei inIvory Coast close to the Ghana border, suggesting itspresence in Ghana. We add to the rodent list of Guineathe presence of M. musculoides and M. mattheyi andconfirm the one of M. baoulei in a new locality. To this,one must add the exotic M. musculus domesticus foundin the village houses of some villages (Kotokossou, Tou-
zoukro, Bakro, Menekre) that may compete with theendemic Mus species. It is also important to note thatthe indigenous Mus we recorded also occur in culturedareas and that the different species were all trapped inthe surroundings of villages. M. minutoides was some-times caught inside houses, a fact that may have sanitaryconsequences as this fact may facilitate the transmissionof viruses to humans. Only very few localities yieldedcatches consisting of only a single Mus species. In gen-eral, two species were found to coexist even in the samehabitat (e.g., M. minutoides and M. setulosus). In a singlevillage (Kodoko) (Guinea), up to three species appear tocoexist. Moreover, it is of interest to point out that it is inthe same village that a previous survey discovered a newarenavirus (LCMV) in M. minutoides (Lecompte et al.2007). This village is found at the Guinean-Sudaniantransition zone and is situated Southeast of the NigerRiver. Sampling locality Lamto station situated at theecotone between forest and savanna also yielded threedifferent species, a finding that confirms the importanceof transition zones for biodiversity.
Conclusion
Species of the subgenus Nannomys are abundant inWest Africa, and some are known to carry viruses. Thus,it is important to be able to identify these species cor-rectly. Our study demonstrates that the combination ofmolecular phylogenetic information based on mitochon-drial DNA sequences and standard karyotypes may facil-itate the correct identification of these species. We havealso been able to provide some diagnostic morphologicalcharacters for the five studied West African Mus speciesand to precisely identify their geographical distributions.The morphological and morphometrical characters thatcharacterize the studied species are useful to identifythese taxa. Nevertheless, future studies are required toconfirm that these characters alone are reliable enoughfor the identification of these species, as we did notobtain 100% correct classification in the canonicalanalyses. Similar to the case of the multimammate ratMastomys, we recommend the combined use of mor-phological, chromosomal and genetic data to reliablyidentify these species during future ecological, agricul-tural or health studies.
Acknowledgements
Ian ter Meulen, Vitaly Volobouev, Emilie Lecompte and ElisabethFichet Calvet are thanked for their collaboration in the field or inlogistics. R. Cornette, C. Houssin and A. Delapree contributedto specimen preparations at the MNHN (SPOT, service de prep-aration taxidermique et osteologique). Thanks to the PFHG (Pro-jet Fievres hemorragiques en Guinee) and Ivory Coast InstitutPasteur drivers: Peve Guilavogui, Mamadou Doumbouya, Sou-rouma Beavogui, Denis Kouassi Kaouakou, Tape Bozoua A.F.People of the CHU (Centre Hospitalier Universitaire) Donka-PFHG in Conakry helped in logistics (among them Dr. Magas-souba, the late Fidele Haba, Mme. Sow). Prof. H. Dosso, D.Coulibaly, Michel Kabi and Assi Yao Michel helped in the field-work in Ivory Coast. This work was funded by CEE-INCO-DEV-

S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa 249
Article in press - uncorrected proof
TREATCONTROL program (No. ICA4-CT2002-10050), HealthMinistry of Guinea (research permit 2003/PFHG/05/GUI) andResearch and Education Ministry of Guinea. This molecular workwas supported by the ‘‘Consortium National de Recherche enGenomique’’, and the ‘‘Service de systematique moleculaire’’ ofthe Museum National d’Histoire Naturelle (IFR 101). It is part of
the agreement no. 2005/67 between the Genoscope and theMuseum National d’Histoire Naturelle on the project ‘‘Macro-phylogeny of life’’ directed by Guillaume Lecointre. The authorsthank D. Reed and the two anonymous referees for improvingthe English and their very attentive and constructive commentson the original manuscript.
Appendix A
List of specimens used for this study.
Number Species Locality Country Karyotype cytb Morpho
BE055 baoulei Agbassa Benin EU603991BE141 baoulei Sakete Benin EU6039921920-375-3 baoulei Kerouane Guinea xKD36 baoulei Kodoko Guinea EU603995 x1973-482 baoulei Foro Ivory Coast x1980-100 baoulei Foro Ivory Coast x1980-94 baoulei Foro Ivory Coast T&G xCIV 296 baoulei Sanguiebo Ivory Coast EU603994 xCIV 697 baoulei Lamto Ivory Coast EU603998 xCIV 699 baoulei Lamto Ivory Coast EU603993 x1966-1897 mattheyi Accra Ghana M x1966-1898 mattheyi Accra Ghana M x1966-1900 mattheyi Accra Ghana M x1966-1902 mattheyi Accra Ghana M xALT100 mattheyi Altou Fonkola Guinea EU603971 xALT46 mattheyi Altou Fonkala Guinea A xALT47 mattheyi Altou Fonkola Guinea A EU603972 xBA758 mattheyi Bantou Guinea A EU603973 xBOK96 mattheyi Boke Guinea EU603970 xKO628 mattheyi Khoneya Guinea A xTA778 mattheyi Tanganya Guinea A xTA781 mattheyi Tanganya Guinea A xTA811 mattheyi Tanganya Guinea A xTA812 mattheyi Tanganya Guinea A x1990-27 mattheyi Bouake Ivory Coast M x
mattheyi Mali AJ875067mattheyi Mali AJ698876mattheyi Mali AB125781mattheyi Mali AJ875066
2004-1046 mattheyi Farabana Mali V x2004-1047 mattheyi Farabana Mali V x2004-1048 mattheyi Farabana Mali V x2004-1049 mattheyi Farabana Mali V x
mattheyi Senegal AJ8750681971-773 mattheyi Damantan Senegal M x1971-774 mattheyi Damantan Senegal M x
mattheyi Togo AJ875069R24684 minutoides Tai Cote d’ivoire EU603959241 minutoides Gagal Guinea V AJ875077 xBA704 minutoides Bantou Guinea A xBH18 minutoides Bhoita Guinea EU603960 xFR1 minutoides Franfina Guinea EU603964 xGB155 minutoides Gbetaya Guinea V AJ875076 xKD30 minutoides Kodoko Guinea EU603961 xKD42 minutoides Kodoko Guinea EU603965 xKDF30 minutoides Kandiafara Guinea A EU603936 xKDF32 minutoides Kandiafara Guinea EU603937 xTA777 minutoides Tanganya Guinea A xTA815 minutoides Tanganya Guinea A xP0486 minutoides Kazaouma Guinee EU603966CIV 1309 minutoides Gbagroubett Ivory Coast EU603963 xCIV 100 minutoides Blodi Ivory Coast EU603999 xCIV 1209 minutoides Blodi Ivory Coast EU603953 xCIV 1215 minutoides Blodi Ivory Coast EU603955 xCIV 1216 minutoides Blodi Ivory Coast EU603958 xCIV 1224 minutoides Blodi Ivory Coast EU603954 xCIV 1226 minutoides Blodi Ivory Coast EU604005 xCIV 1253 minutoides Menekre Ivory Coast EU603941 x

250 S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa
Article in press - uncorrected proof
(Appendix A Continued)
Number Species Locality Country Karyotype cytb Morpho
CIV 132 minutoides Menekre Ivory Coast EU603962 xCIV 171 minutoides Gbagroubett Ivory Coast EU603929 xCIV 172 minutoides Gbagroubett Ivory Coast EU603934 xCIV 173 minutoides Gbagroubett Ivory Coast EU603933 xCIV 187 minutoides Gbagroubett Ivory Coast EU603925 xCIV 190 minutoides Gbagroubett Ivory Coast EU603935 xCIV 379 minutoides Kotokossou Ivory Coast EU603996 xCIV 420 minutoides Touzoukro Ivory Coast EU604000 xCIV 421 minutoides Touzoukro Ivory Coast EU603942 xCIV 435 minutoides Touzoukro Ivory Coast EU603943 xCIV 436 minutoides Touzoukro Ivory Coast EU603944 xCIV 439 minutoides Touzoukro Ivory Coast EU603946 xCIV 440 minutoides Touzoukro Ivory Coast EU603947 xCIV 455 minutoides Touzoukro Ivory Coast EU603938 xCIV 473 minutoides Bakro Ivory Coast EU603931 xCIV 477 minutoides Bakro Ivory Coast EU603948 xCIV 487 minutoides Bakro Ivory Coast EU603949 xCIV 495 minutoides Blodi Ivory Coast EU603957 xCIV 524 minutoides Blodi Ivory Coast EU604001 xCIV 525 minutoides Bakro Ivory Coast EU603951 xCIV 526 minutoides Blodi Ivory Coast EU603952 xCIV 602 minutoides Gbagroubett Ivory Coast EU604003 xCIV 603 minutoides Gbagroubett Ivory Coast EU603926 xCIV 604 minutoides Gbagroubett Ivory Coast EU603927 xCIV 605 minutoides Gbagroubett Ivory Coast EU603928 xCIV 628 minutoides Gbagroubett Ivory Coast EU604002 xCIV 631 minutoides Gbagroubett Ivory Coast EU603932 xCIV 673 minutoides Gbagroubett Ivory Coast EU603930 xCIV 693 minutoides Lamto Ivory Coast EU603945 xCIV 698 minutoides Lamto Ivory Coast EU603939 xCIV 747 minutoides Ahouakro Ivory Coast EU603940 xCIV 840 minutoides Menekre Ivory Coast EU604004 xCIV 94 minutoides Blodi Ivory Coast EU603950 xCIV 96 minutoides Blodi Ivory Coast EU603956 x
minutoides Kenya AJ875084minutoides South Africa AJ875080minutoides South Africa AJ875078minutoides South Africa AJ875079minutoides South Africa AJ875081
KD01 musculoides Kodoko Guinea EU603969 xKD29 musculoides Kodoko Guinea EU603968 xCIV 298 musculoides Sanguiebo Ivory Coast EU603967 x
musculoides Mali AJ875075musculoides Mali AJ698875musculoides Mali Z96069
2004-1052 musculoides Farabana Mali V x2004-1053 musculoides Farabana Mali V x2004-1054 musculoides Samaya Mali V x2004-1055 musculoides Farabana Mali V x1971-761 musculoides Damantan Senegal M x1971-777 musculoides Damentan Senegal M x2004-1051 musculoides Farabana Senegal V xR14029 setulosus Ekom Cameroon EU603989
setulosus Central African Republic AJ8750821964-17 setulosus Boukoko Central African Republic M x1971-507 setulosus La Maboke Central African Republic M x1971-577 setulosus La Maboke Central African Republic M xR12258 setulosus Mouloukou Central African Republic EU6039901959-910 setulosus Seredou Guinea x1959-911 setulosus Seredou Guinea x1959-912 setulosus Seredou Guinea x1959-913 setulosus Seredou Guinea x1959-914 setulosus Seredou Guinea x1959-915 setulosus Seredou Guinea x1959-919 setulosus Seredou Guinea x1960-681 setulosus Seredou Guinea x1970-689 setulosus Seredou Guinea xBH19 setulosus Bhoita Guinea EU603983 xBH39 setulosus Bhoita Guinea EU603986 x

S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa 251
Article in press - uncorrected proof
(Appendix A Continued)
Number Species Locality Country Karyotype cytb Morpho
GB158 setulosus Gbetaya Guinea V AJ875083 xSA15 setulosus Sangassou Guinea A EU603982 xSA33 setulosus Sangassou Guinea A xSA41 setulosus Sangassou Guinea A xDV44 setulosus Yossono Guinea EU603976P0781 setulosus Kazaouma Guinea EU6039781998-201 setulosus Adiopodoume Ivory Coast x1998-202 setulosus Adiopodoume Ivory Coast x1998-204 setulosus Adiopodoume Ivory Coast x1998-208 setulosus Adiopodoume Ivory Coast x1998-848 setulosus Adiopodoume Ivory Coast x1998-880 setulosus Adiopodoume Ivory Coast x1998-904 setulosus Adiopodoume Ivory Coast x1998-913 setulosus Adiopodoume Ivory Coast xCIV 1288 setulosus Gbagroubett Ivory Coast EU603974 xCIV 1290 setulosus Gbagroubett Ivory Coast EU603975 xCIV 338 setulosus Kotokossou Ivory Coast EU603981 xCIV 472 setulosus Bakro Ivory Coast EU603988 xCIV 476 setulosus Bakro Ivory Coast EU603987CIV 610 setulosus Gbagroubett Ivory Coast EU603984 xCIV 640 setulosus Gbagroubett Ivory Coast EU603985CIV 644 setulosus Gbagroubett Ivory Coast EU603979 xCIV 663 setulosus Gbagroubett Ivory Coast EU603980 xCIV 664 setulosus Gbagroubett Ivory Coast EU603997 xCIV 674 setulosus Gbagroubett Ivory Coast EU603977 x
The column karyotype shows an abbreviation corresponding to the person who karyotyped or published the karyotype: T andGs M. Tranier and J.C. Gautun, unpublished data; VsVeyrunes et al. 2004; MsMatthey 1966, 1967, 1970a,b; AsV. Aniskine, thiswork. GenBank numbers are provided in column cytb. The column Morpho (morphology/morphometrics) indicates specimens usedfor the present work in the analyses, including karyotyped, molecularly typed and/or sequenced specimens, but also old collectionsof Guinean specimens previously examined, whose determination was confirmed by G. Musser, were used.
References
Castiglia, R., E. Gormung and M. Corti. 2002. Cytogenetic anal-yses of chromosomal rearrangements in Mus minutoides/musculoides from North-West Zambia through mapping ofthe telomeric sequence (TTAGGG)n and banding techniques.Chrom. Res. 10: 399–406.
Catzeflis, F.M. and C. Denys. 1992. The African Nannomys (Muri-dae): an early offshoot from the Mus lineage. Evidence forscnDNA hybridization experiments and compared morphol-ogy. Isr. J. Zool. 38: 219–231.
Chevret, P., F. Veyrunes and J. Britton-Davidian. 2005. Molecularphylogeny of the genus Mus (Rodentia: Murinae) based onmitochondrial and nuclear data. Biol. J. Linnean Soc. 84:417–427.
Clement, M., D. Posada and K. Crandall. 2000. TCS: a computerprogram to estimate gene genealogies. Mol. Ecol. 9:1657–1660.
Denys, C. and M. Tranier. 1992. Presence d’Aethomys (Mam-malia, Rodentia) au Tchad et analyse morphometrique preli-minaire du complexe A. hindei. Mammalia 56: 625–656.
Denys, C., E. Lecompte, E. Calvet, M. Camara, A. Dore, K. Kou-lemou, F. Kourouma, B. Soropogui, O. Sylla, B.K. Allali, S.Kouassi-Kan, C. Akoua-Koffi, J. ter Meulen and L. Koivogui.2005. Community analysis of Muridae (Mammalia, Rodentia)diversity in Guinea: a special emphasis on Mastomys speciesand Lassa fever distributions. In: (B.A. Huber, B.J. Sinclairand K.H. Lampe, eds) African biodiversity, molecules, organ-isms, ecosystems. Springer, New York. pp. 339–350.
Dobigny, G., A. Nomao and J.C. Gautun. 2002. A cytotaxonomicsurvey of rodents from Niger: implications for systematics,biodiversity and biogeography. Mammalia 66: 495–523.
Dosso, H. 1988. Les peuplements de rongeurs de forets et deszones defrichees et cultives de la Cote d’Ivoire. Ann. Univ.Abidjan, Ser. E (Ecol.) XX: 67–85.
Duplantier, J.M. and L. Granjon. 1992. Liste revisee des rongeursdu Senegal. Mammalia 56: 425–431.
Duplantier, J.M., L. Granjon and K. Ba. 1997. Repartition bio-geographique des petits rongeurs au Senegal. J. Afr. Zool.111: 17–26.
Gautun, J.C., I. Sankhon and M. Tranier. 1986. Nouvelle contri-bution a la connaissance des rongeurs du massif guineendes monts Nimba (Afrique occidentale). Systematique etapercu quantitatif. Mammalia 50: 205–217.¸
Granjon, L. and G. Dobigny. 2003. The importance of cytotax-onomy in understanding the biogeography of Africanrodents: Lake Chad murids as an example. Mammal Rev. 33:77–91.
Granjon, L., J.M. Duplantier, J. Catalan and J. Britton-Davidian.1992. Karyotypic data on rodents from Senegal, mainly Muri-dae and Gerbillidae. Isr. J. Zool. 38: 263–276.
Grubb, P., T.S. Jones, A.G. Davies, E. Edberg, E.D. Starin andJ.E. Hill. 1998. Mammals of Ghana, Sierra Leone and theGambia. The Tendrine Press, Cornwall. 265 pp.
Heim de Balsac, H. and M. Lamotte. 1958. XV. Mammiferesrongeurs. La reserve integrale du Mont Nimba. Mem. IFAN.53: 339–357.
Hubert, B., F. Adam and A. Poulet. 1973. Liste preliminaire desrongeurs du Senegal. Mammalia 37: 76–87.
Huelsenbeck, J.P. and F. Ronquist. 2001. MRBAYES: Bayesianinference of phylogenetic trees. Bioinformatics 17: 754–755.
Jotterand, M. 1972. Le polymorphisme chromosomique desMus (Leggadas) africains. Cytogenetique, zoogeographie,evolution. Rev. Suisse Zool. 79: 287–359.
Jotterand-Bellomo, M. 1984. L’analyse cytogenetique de deuxespeces de Muridae africains, Mus oubanguii et Mus minu-toides/musculoides: polymorphisme chromosomique etebauche d’une phylogenie. Cytogenet. Cell Genet. 38: 182–188.
Jotterand-Bellomo, M. 1986. Le genre Mus africain, un exempled’homogeneite caryotypique: etude cytogenetique de Musminutoides/musculoides (Cote d’Ivoire), de M. setulosus

252 S. Kan Kouassi et al.: Pygmy mice taxonomy in West Africa
Article in press - uncorrected proof
(Republique Centrafricaine), et de M. mattheyi (Burkina Faso).Cytogenet. Cell Genet. 42: 99–104.
Kadjo, B., H. Dosso and R. Vuattoux. 1996. Influence de la vege-tation sur le peuplement des rongeurs dans la region centrede Cote d’Ivoire. Agron. Afr. VIII (3): 153–212.
Klempa, B., E. Fichet-Calvet, E. Lecompte, B. Auste, V. Aniskin,H. Meisel, C. Denys, L. Koivogui, J. ter Meulen and D.H. Kru-ger. 2006. Hantavirus in African wood mouse, Guinea. Em.Inf. Dis. 12: 838–840.
Kuper, W., J.H. Sommer, J.C. Lovett, J. Mutke, H.P. Linder, H.J.Beentje, S. Van Rompaey, C. Chatelain, M. Sosef and W.Barthlott. 2004. Africa’s hotspots of biodiversity redefined.Ann. Missouri Bot. Gard. 91: 525–535.
Lecompte, E., E. Fichet-Calvet, S. Daffis, K. Koulemou, F. Kou-rouma, A. Dore, B. Soropogui, V. Aniskine, B.K. Allali, S.Kouassi Kan, A. Lalis, S. Gunther, L. Koivogui, C. Denys andJ. ter Meulen. 2006. Mastomys natalensis and Lassa fever,West Africa. Em. Inf. Dis. 12: 1971–1974.
Lecompte, E., J. ter Meulen, S. Emonet, S. Daffis and R.N. Char-rel. 2007. Genetic identifications of Kodoko virus, a novelarenavirus of the African pygmy mouse (Mus Nannomysminutoides) in West Africa. Virology 364: 178–183.
Lee, M.R. and F.F.B. Elder. 1980. Yeast stimulation of bone mar-row mitosis for cytogenetic investigations. Cytogenet. CellGenet. 26: 36–40.
Matthey, R. 1966. Nouvelles contributions a la cytogenetiquedes Mus africaines du sous genre Leggada. Experientia 22:1–6.
Matthey, R. 1967. Un nouveau systeme chromosomique poly-morphe chez des Leggada africaines du groupe tenellus(Rodentia – Muridae). Genetica 38: 211–226.
Matthey, R. 1970a. Nouvelles donnees sur la cytogenetique etla speciation des Leggada (Mammalia – Rodentia – Muridae).Experientia 26: 102–103.
Matthey, R. 1970b. L’eventail robertsonien chez les Mus (Leg-gada) africains du groupe minutoides-musculoides. Rev.Suisse Zool. 77: 625–629.
Mills, J.N., J.E. Childs, T.G. Ksiazek, C.J. Peters and W.M. Vel-leca. 1995. Methods for trapping and sampling small mam-mals for virologic testing. U.S. Department of Health andHuman Services, Public Health Service, Centers for DiseaseControl and Prevention, Atlanta, GA.
Musser, G.G. and M.D. Carleton. 2005. Superfamily Muroidea.In: (D.E. Wilson and D.A.M. Reeder, eds) Mammal species ofthe world: a taxonomic and geographic reference. 3rd edn.The Johns Hopkins University Press, Baltimore, MD. pp.894–1531.
Myers, N., R.A. Mittermeier, C.G. Mittermeier, G.A.B. Da Fon-seca and J. Kent. 2000. Biodiversity hotspots for conserva-tion priorities. Nature 403: 853–858.
Petter, F. 1963. Contribution a la reconnaissance des sourisafricaines. Mammalia 27: 602–607.
Petter, F. 1969. Une souris nouvelle d’Afrique occidentale Musmattheyi sp. nov. Mammalia 33: 118–123.
Petter, F., F. Adam and B. Hubert. 1971. Presence au Senegalde Mus mattheyi F. Petter 1969. Mammalia 35: 346–347.
Posada, D. and K.A. Crandall. 1998. MODELTEST: testing themodel of DNA substitution. Bioinformatics 14: 817–818.
Roche, J. 1971. Recherches mammalogiques en Guinee fores-tiere. Bull. Mus. Nat. Hist. Nat. Ser. 3, Zool. 16: 737–782.
Ronquist, F. and J.P. Huelsenbeck. 2003. MrBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinformatics19: 1572–1574.
Ronquist, F., J.P. Huelsenbeck and P. van der Mark. 2005.MrBAYES 3.1 manual. https://mrbayes.csit.fsu.edu/index.php.
Rosevear, D.R. 1969. The rodents of West Africa. British Muse-um of Natural History, London. 604 pp.
Rozas, J., J.C. Sanchez-DelBarrio, X. Messeguer and R. Rozas.2003. DnaSP, DNA polymorphism analyses by the coalescentand other methods. Bioinformatics 19: 2496–2497.
Sullivan, F. and P. Joyce. 2005. Model selection in phylogenetics.Annu. Rev. Ecol. Evol. Syst. 36: 445–466.
Swofford, D.L. 2002. PAUP*: phylogenetic analysis using parsi-mony (and other methods). Sinauer Associates, Sunderland,MA.
Taylor, P.J. 2000. Patterns of chromosomal variation in southernAfrican rodent. J. Mamm. 81: 317–331.
Vermeiren, L.J.P. and W.N. Verheyen. 1980. Notes sur les Leg-gada de Lamto, Cote d’Ivoire, avec la description de Leg-gada baoulei sp. N. (Mammalia, Muridae). Rev. Zool. Afr. 94:570–590.
Veyrunes, F., J. Catalan, B. Sicard, T.J. Robinson, J.M. Duplan-tier, L. Granjon, G. Dobigny and J. Britton-Davidian. 2004.Autosome and sex chromosome diversity among the Africanpygmy mice, subgenus Nannomys (Muridae; Mus). Chrom.Res. 12: 369–382.
Veyrunes, F., J. Britton-Davidian, T.J. Robinson, E. Calvet, C.Denys and P. Chevret. 2005. Molecular phylogeny of the Afri-can pygmy mice, subgenus Nannomys (Rodentia, Murinae,Mus): implications for chromosomal evolution. Mol. Phyl.Evol. 36: 358–369.
White, F. 1986. La vegetation de l’Afrique. Memoire accompa-gnant la carte Central African Republicte de vegetation del’Afrique UNESCO/AETFAT/UNSO. UNESCO, ORSTOM. pp.1–384.
Winnepenninckx, B., T. Backeljau and R. De Watcher. 1993.Extraction of high molecular weight DNA from molluscs.Trends Genet. 9: 407.




















