Inferring the relationship between butterfly species in the subgenus Agrodiaetus on Balkan peninsula
54
UNIVERZA V LJUBLJANI PEDAGOŠKA FAKULTETA BIOTEHNIŠKA FAKULTETA FAKULTETA ZA KEMIJO IN KEMIJSKO TEHNOLOGIJO NAROVOSLOVNOTEHNIŠKA FAKULTETA Študijski program: Kemija in Biologija UGOTAVLJANJE SORODSTVENIH ODNOSOV V PODRODU METULJEV AGRODIAETUS NA BALKANSKEM POLOTOKU DIPLOMSKO DELO INFERRING THE RELATIONSHIP BETWEEN BUTTERFLY SPECIES IN THE SUBGENUS AGRODIAETUS ON BALKAN PENINSULA GRADUATION THESIS Mentor: dr. Rudi Verovnik Kandidat: Tom Koritnik LJUBLJANA, oktober, 2014
Transcript of Inferring the relationship between butterfly species in the subgenus Agrodiaetus on Balkan peninsula

UNIVERZA V LJUBLJANI
PEDAGOŠKA FAKULTETA
BIOTEHNIŠKA FAKULTETA
FAKULTETA ZA KEMIJO IN KEMIJSKO TEHNOLOGIJO
NAROVOSLOVNOTEHNIŠKA FAKULTETA
Študijski program: Kemija in Biologija
UGOTAVLJANJE SORODSTVENIH ODNOSOV V PODRODU
METULJEV AGRODIAETUS NA BALKANSKEM POLOTOKU
DIPLOMSKO DELO
INFERRING THE RELATIONSHIP BETWEEN BUTTERFLY
SPECIES IN THE SUBGENUS AGRODIAETUS ON BALKAN
PENINSULA
GRADUATION THESIS
Mentor: dr. Rudi Verovnik
Kandidat: Tom Koritnik
LJUBLJANA, oktober, 2014

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
I
Diplomsko delo je zaključek univerzitetnega študijskega programa Kemija in Biologija.
Opravljeno je bilo na Katedri za zoologijo Oddelka za biologijo Biotehniške fakultete
Univerze v Ljubljani. Komisija za dodiplomski študij Oddelka za biologijo je za mentorja
diplomskega dela imenovala doc. dr. Rudija Verovnika. Komisija za oceno in zagovor:
Predsednik: doc. dr. Primoţ ZIDAR
Univerza v Ljubljani, Biotehniška fakulteta
Član: doc. dr. Cene FIŠER
Univerza v Ljubljani, Biotehniška fakulteta
Mentor: doc. dr. Rudi VEROVNIK
Univerza v Ljubljani, Biotehniška fakulteta
Datum zagovora:
Podpisani se strinjam z objavo svojega diplomskega dela v polnem tekstu na spletni strani
Digitalne knjiţnice Biotehniške fakultete. Izjavljam, da je diplomsko delo, ki sem ga oddal v
elektronski obliki, identično tiskani verziji.
Delo je rezultat lastnega raziskovalnega dela.
Tom Koritnik

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
II
KLJUČNA DOKUMENTACIJSKA INFORMACIJA
ŠD Dn
DK
KG metulji /Lepidoptera /modrini /Polyommatus /Agrodiaetus /Polyommatus admetus
/Polyommatus ripartii /Polyommatus eleniae /Polyommatus aroaniensis /Polyommatus
orphicus /kriptične vrste /PCR /filogenetska analiza /morfologija kril /fenotipski znaki
AV KORITNIK, Tom
SA VEROVNIK, Rudi (mentor)
KZ SI-1261Dobrunje, Pot v Podgorje 3a
ZA Univerza v Ljubljani, Pedagoška fakulteta, Biotehniška fakulteta
LI 2014
IN UGOTAVLJANJE SORODSTVENIH ODNOSOV V RODU METULJEV
AGRODIAETUS NA BALKANSKEM POLOTOKU
TD Diplomsko delo (univerzitetni študij)
OP VII, 45 str., 3 preg., 6 sl., 49 virov
IJ sl
JI sl/en
AI V diplomskem delu smo raziskovali sorodstvene odnose in vrstno pripadnost v podrodu
modrinov Polyommatus (subgenus Agrodiaetus) na Balkanskem polotoku. Metulje smo
predhodno določili na podlagi fenotipskih znakov. Vse osebke smo nato poskusili tudi
natančneje uvrstiti glede na širši nabor morfoloških znakov, ki se uporabljajo v taksonomiji
tega podroda. Vrstno pripadnost smo ugotavljali na podlagi primerjave zaporedij
mitohondrijskega gena COI, ki se uporablja kot genetska črtna koda. Filogenetske odnose smo
ugotavljali z Bayesovo analizo in jih primerjali z morfološkimi določitvami osebkov in
njihovo razširjenostjo. Glede na fenotipske znake smo lahko z gotovostjo uvrstili le osebke
vrste Polyommatus admetus, pri ostalih vrstah pa je variabilnost fenotipskih znakov prevelika
za nedvoumno določitev. Filogenetski rezultati so potrdili prisotnost vsaj petih vrst (P.
admetus, P. ripartii, P. aroaniensis, P. orphicus in P. eleniae) med našimi vzorci. Glede na
geografsko razširjenost osebkov posameznih vrst nismo potrdili alopatrije oţje sorodnih vrst.
Simpatrija med bolj sorodnimi vrstami kaţe na reproduktivne bariere tudi med morfološko
zelo podobnimi vrstami. Ugotovili smo tudi, da je območje razširjenosti vrste P. orphicus
večje, kot je navedeno v literaturi.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
III
KEY WORDS DOCUMENTATION (KWD)
DN dn
DC
CX butterflies /Lepidoptera /blues /Polyommatus /Agrodiaetus /Polyommatus admetus /
Polyommatus ripartii /Polyommatus eleniae /Polyommatus aroaniensis /Polyommatus
orphicus /cryptic species /PCR /philogenetic analisys /wing morphology /fenotype
characteristics
AU KORITNIK, Tom
AA VEROVNIK, Rudi (supervisor)
PP SI-1261 Dobrunje, Pot v Podgorje 3a
PB University of Ljubljana, Faculty of Education, Biotechnical faculty
PY 2014
TI Inferring the relationship between butterfly species in the subgenus Agrodiaetus on
Balkan Peninsula
DT Graduation Thesis (University studies)
NO VII, 45 p., 3 tab., 6 fig., 49 ref.
LA sl
AL sl/en
AI We studied the phylogenetic relationships in the subgenus of the Polyommatus (subgenus
Agrodiaetus) butterflies in the Balkan Peninsula. Preliminarily identifications of all specimens
were based on their wing morphology. In turn, all individuals were examined for a larger
number of morphological characters, that proved useful in taxonomy of the subgenus.
Phylogenetic relationships were reconstructed using the mitochondrial DNA gene COI, used
as a DNA barcodes. We reconstructed phylogeny with the Bayesian analysis and then
compared these results with the morphological characteristics and distribution ranges of
studied species. Only Polyommatus admetus can be unambigously identified using
morphological traits; the identiy of other herein studied species cannot be inferred from
morphology alone due to high intraspecific variation. The phylogentic confirmed the presence
of at least five species (P. admetus, P. ripartii, P. aroaniensis, P. orphicus in P. eleniae) in our
samples. According to the geographical distribution of the samples, we could not confirm
alopatry of closely related species. Simpatry between more closely related species suggests
that even morphologically similar species are reproductively isolated. We also found that the
distribution range of the P. orphicus species is larger than previously thought.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
IV
Kazalo vsebine
KLJUČNA DOKUMENTACIJSKA INFORMACIJA ............................................................ II
KEY WORDS DOCUMENTATION (KWD) ......................................................................... III
Kazalo vsebine ......................................................................................................................... IV
Kazalo preglednic: .................................................................................................................... VI
Kazalo slik: ............................................................................................................................... VI
Okrajšave ................................................................................................................................. VII
1 UVOD ................................................................................................................................ 1
2 PREGLED OBJAV ............................................................................................................ 2
2.1 Ţuţelke ........................................................................................................................ 2
2.2 Metulji ......................................................................................................................... 2
2.3 Druţina modrinov (Lycaenidae) .................................................................................. 4
2.4 Rod Polyommatus ........................................................................................................ 5
2.5 Podrod Agrodiaetus ..................................................................................................... 5
2.5.1 Polyommatus admetus (Esper, 1783) ................................................................... 6
2.5.2 Polyommatus ripartii (Freyer, 1830) ................................................................... 6
2.5.3 Polyommatus aroaniensis (Brown, 1976) ............................................................ 7
2.5.4 Polyommatus eleniae (Coutsis in De Prins, 2005) ............................................... 7
2.5.5 Polyommatus orphicus (Kolev, 2005) .................................................................. 7
2.6 Kriptične vrste ............................................................................................................. 8
2.7 Črtne kode DNK .......................................................................................................... 8
2.8 Reakcija PCR ............................................................................................................... 9
3 MATERIALI IN METODE ............................................................................................. 10
3.1 Vzorci ........................................................................................................................ 10

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
V
SEZNAM VZORCEV IN LOKALITET ......................................................................... 11
3.2 Morfologija kril ......................................................................................................... 13
3.3 MOLEKULARNOBIOLOŠKA ANALIZA ............................................................. 15
3.3.1 Kemikalije, potrošni material in naprave ........................................................... 15
3.3.2 Izolacija DNK .................................................................................................... 15
3.3.3 Pomnoţevanje DNK ........................................................................................... 18
3.3.4 Postopek izdelave filogenetskih dreves in Bayesova analiza ............................. 22
4 REZULTATI .................................................................................................................... 26
4.1 Morfologija barvnih vzorcev na krilih ....................................................................... 26
4.1.1 Preglednica opazovanih fenotipskih znakov ...................................................... 28
4.2 Genetska podobnost med osebki ............................................................................... 30
5 RAZPRAVA .................................................................................................................... 33
5.1 Primernost fenotipskih znakov na krilih za ločevanje vrst ........................................ 33
5.2 Določitev vzorcev s filigenetsko analizo ................................................................... 34
5.3 Alopatrija in razširjenost vrst .................................................................................... 35
6 SKLEPI ............................................................................................................................ 37
7 POVZETEK ..................................................................................................................... 38
8 VIRI .................................................................................................................................. 40
Zahvala ..................................................................................................................................... 45

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
VI
Kazalo preglednic:
Preglednica 1: Preglednica vzorcev uporabljenih v diplomski nalogi. .................................... 11
Preglednica 2: Seznam referenčnih in zunanjih vzorcev vključenih v filogenetsko drevo ...... 24
Preglednica 3: Seznam vzorcev s pripadajočimi opazovanimi fenotipskimi znaki ................. 28
Kazalo slik:
Slika 1: Zemljevid juţnega Balkana z označenimi lokacijami ulova vzorcev in unikatnimi
kodami vzorcev. ....................................................................................................................... 13
Slika 2: Shematski prikaz strukture krila metulja z označbami ............................................... 14
Slika 3: Vzorci LA38, LA53, LA60. ........................................................................................ 15
Slika 4: Vzorec LA62, na podlagi morfologije določen kot vrsta P. admetus. ........................ 15
Slika 5: Filogenetsko drevo podobnosti izračunano po metodi Bayes ..................................... 32
Slika 6: Zemljevid območja Makedonije, severne Grčije in Bolgarije z označenimi lokacijami
vzorcev LA37, LA38, LA39 in LA60 ...................................................................................... 36

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
VII
Okrajšave
DNK deoksiribonukleinska kislina
COI citokrom oksidaza I
m meter
PRC polymerase chain reaction
dNTP deoksiribonukleotidni trifosfat
TAE tris-acetatni-EDTA pufer
EDTA etilendiaminotetraična kislina
W watt
V volt
Ω-voda ultračista voda
µl mikroliter
ml mililiter
UV ultravijolično

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
1
1 UVOD
Metulji iz druţine Lycaenidae so velika skupina metuljev, ki ţivi po celem svetu. Znotraj
druţine ločimo mnogo rodov, sorodnost katerih zadnja leta proučujejo z genetskimi metodami
(Kandul in sod., 2004). Teţavna je tudi neurejena vrstna problematika. Trenutno ima status
vrste pribliţno 50 taksonov. Mnoge od teh vrst so razširjene le lokalno (Vila in sod., 2010).
Klasifikacija rodov v druţini Lycaenidae je teţavna. Največji rod v druţini je rod
Polyommatus, znotraj katerega je največ vrst uvrščenih v podrod Agrodiaetus (Kandul in sod.,
2004). Slednji je bil sprva opisan kot samostojen rod (Elliot, 1973), a so ga kasnejše analize
uvrstile kot podrod v rodu Polyommatus (Dantchenko in Lukhtanov, 1994; Vila in sod.,
2010).
Mnoge vrste iz podrodu Agrodiaetus so spolno dimorfne. Pri teh so samci modri, samice pa
različnih rjavih odtenkov. Pri številnih vrstah spolni dimorfizem ni tako izrazit. Ker je vrst
znotraj podrodu Agrodiaetus več, kot je znanih različnih vzorcev na krilih pri samcih, lahko
domnevamo, da je v podrodu večje število kriptičnih vrst. Poleg nejasnosti glede morfologije
imajo vrste znotraj podrodu tudi veliko diverziteto v kariotipih, verjetno celo največjo v
ţivalskem kraljestvu (Vila in sod., 2010). Teţave s klasifikacijo te skupine metuljev je opazil
ţe Elliot (1973), ko je zapisal: »Priznati moram popolno neuspešnost pri poskusih razdelitve
rodu Agrodiaetus v naravne skupine, zato smo vrste organizirali v 30 sekcij.« Ločevanje med
vrstami oteţujejo tudi le neznatne razlike v genitalnih strukturah, ki so sicer pomemben
določevalni znak pri metuljih (Wiemers, 2003).
Z razvojem molekulsko genetskih postopkov sekvenciranja DNK se je v zadnjih dveh
desetletjih odprla moţnost identifikacije osebkov z genetskimi markerji − s črtnimi kodami
DNK. Ta postopek omogoča identifikacijo vrst na podlagi unikatnega zaporedja nukleotidov
standardnih delov genoma. Ta postopek pogosto ni dovolj za končno identifikacijo, zato
večinoma uporabljamo še druge metode identifikacije, kot so morfološki znaki, kariotipske
raziskave, študije vedenjskih vzorcev ipd. Metulji znotraj podrodu Agrodiaetus se po
fenotipskih znakih slabo razlikujejo, prav tako imajo za določevanje neuporabne genitalije. V

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
1
praksi se uporablja kombinacija morfoloških znakov in genetskih markerjev oz. kombinacija
več različnih genetskih markerjev in kariotipizacija (Ajmal Ali in sod., 2014).
Našo raziskavo so vodili naslednji cilji:
- ugotoviti, ali se morfološki znaki, ki jih uporabljamo za ločevanje vrst, ujemajo z
molekulsko ugotovljeno filogenijo;
- ugotoviti sorodstvene odnose med vrstami Polyommatus admetus, Polyommatus
ripartii, Polyommatus aroaniensis, Polyommatus nephohiptmenos, Polyommatus
eleniae in Polyommatus orphicus na Balkanskem polotoku;
- dopolniti poznavanje razširjenosti zgornjih vrst na podlagi zbranega materiala s
klasičnimi morfološkimi in z genetskimi metodami.
V delu smo testirali dve hipotezi:
Morfološko definirane vrste iz podrodu Agrodiaetus na Balkanskem polotoku so tudi
genetsko med seboj različne.
Genetsko med seboj bolj sorodne vrste iz podrodu Agrodiaetus se na Balkanskem
polotoku pojavljajo alopatrično, torej geografsko ločeno.
V diplomskem delu smo opravili molekulske raziskave na mitohondrijskem citokromu C
oksidaznem genu podenote I (COI) gena in kvalitativno analizo morfoloških vzorcev na
krilih.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
2
2 PREGLED OBJAV
2.1 Žuželke
Ţuţelke so najštevilčnejša skupina metazojev na zemlji. Opisanih je več kot en milijon vrst
ţuţelk (Chapman, 2006), ocene številčnosti celotne skupine pa se gibljejo od 2 milijonov
(Nielsen in Mound, 2000) do skoraj 6 milijonov vrst (Novotny, 2002).
Najstarejši fosili, ki jih lahko prištevamo k ţuţelkam, so stari 250 milijonov. Te ţivali so ţe
imele kompleksno strukturo ţuţelčjega telesa, iz česar lahko sklepamo, da je skupina Insecta
mnogo starejša. Fosili, stari od 25 do 35 milijonov let, ţe kaţejo podobnost današnjim
ţuţelkam (Klots in Klots, 1970).
Ţuţelke (Insecta) spadajo v deblo členonoţcev (Arthropoda) in poddeblo šesteronoţcev
(Hexapoda). Razred ţuţelk se nato razdeli v mnogo redov, eden izmed njih je red metuljev
(Lepidoptera) (Štrus in sod., 2002).
Vse ţuţelke imajo telo iz členov. Oporo telesu daje zunanji skelet, ki je iz hitina in sklerotina.
Telo se deli v tri telesne regije: glavo, oprsje in zadek. Na glavi ločimo več struktur, kot so
tipalnice, oči in obustni aparat. Na oprsju so trije pari členjenih nog, navadno tudi dva para
kril. Zadek ima med drugim odprtine trahej in strukture za izleganje zaroda (leglica ali
preobraţena leglica npr. v ţelo) (Daccordi in sod., 1987).
2.2 Metulji
Metulji (red Lepidoptera) so se pojavili pred 100 do 130 milijoni let, kar sovpada z razvojem
kritosemenk, ki predstavljajo primaren vir hrane za večino vrst metuljev (Preston-Mafham,
1988).
Poznamo okoli 160.000 opisanih vrst metuljev, vendar je ocena končnega števila vsaj okrog
500.000 vrst. Kljub velikemu številu vrst in razširjenosti na vseh kontinentih so metulji

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
3
strukturno in ekološko homogena skupina v primerjavi z drugimi velikimi skupinami ţuţelk
(Kristensen in sod., 2007).
Metulji imajo popolno preobrazbo, kar pomeni, da imajo 4 faze razvoja: jajčece, larva, buba
in odrasel osebek. Takšen ţivljenjski cikel pomeni, da larve in odrasli osebki med seboj ne
tekmujejo za isti vir hrane (Preston-Mafham, 1988).
Metulje lahko razdelimo v dve skupini: dnevne metulje in vešče, ki veljajo za nočne metulje.
Ta delitev je umetna, saj so pri nekaterih vrstah razlike med skupinama le majhne (Preston-
Mafham, 1988).
Metulji se med ţuţelkami s popolno preobrazbo najbolj razlikujejo po luskah na svojih dveh
parih kril. Barvila, ki se kopičijo v teh luskah, dajejo metuljem barvo. Luske se ne nahajajo le
na krilih, ampak pogosto pokrivajo celotno telo (Daccordi in sod., 1988). Dolge so pribliţno
100 μm in široke okrog 50 μm. Gostota lusk se giblje med 200 do 600 lusk na mm2. Barva
metuljev je posledica mozaične sestave lusk, katerih barva in intenziteta barve se razlikujeta
(Burghardt in sod., 2000). Vzorci na krilih so najpogosteje iz treh, štirih ali petih barv. K
pestrosti vzorcev prispeva tudi oblika in zgradba luske same, ki daje vtis bleščečih barv zaradi
odboja svetlobe (Preston-Mafhan, 1988). Na glavi je par sestavljenih oči in pri nekaterih
vrstah tudi par ocelov, katerih vloga je le delno raziskana. Pomemben organ na glavi so
tipalnice, ki nosijo receptorje za voh. Receptorji za voh pa niso omejeni le na antene, ampak
jih lahko najdemo tudi drugje po telesu, predvsem na nogah. Dolţina in oblika tipalnic je
vrstno specifična, pri tem pa je treba omeniti, da se oblika in velikost anten razlikujeta tudi
med spoloma iste vrste. Antene samcev so navadno kompleksnejše, saj jih uporabljajo
primarno za iskanje samic (Daccordi in sod., 1988). Obustni aparat je večinoma v obliki
rilčka, ki ga tvorita preobraţeni maksili v obliki kanalčka za sesanje tekoče hrane. Ko rilčka
ne uporabljajo, ga lahko zvijejo s pomočjo mišic in tlaka v miksocelu. Krila so derivat
epidermisa. Oporo jima dajejo hitinaste cevke, ki jim rečemo ţile. Noge imajo tipično
ţuţelčjo strukturo, pri nekaterih vrstah pa so reducirane do te mere, da so skoraj neuporabne
za premikanje (Lewis, 1985). Zadek je relativno preprost. Sestavljen je iz desetih segmentov,
kjer sta zadnja dva segmenta spremenjena. Samci imajo strukturo, s katero primejo samico
med parjenjem, samice pa imajo leglico (Preston-Mafham, 1988).
V redu Lepidoptera je mnogo druţin. V nadaljevanju se bomo osredotočili na druţino
Lycaenidae, s katero smo se tudi ukvarjali.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
4
2.3 Družina modrinov (Lycaenidae)
Modrini so druga največja druţina dnevnih metuljev. V primerjavi z drugimi druţinami
dnevnih metuljev je večina vrst majhnih. Gosenice modrinov so različne. Najpogosteje so
gosenice kratke in čokate. Imajo kratke noge. Glava je skoraj skrita pod oprsjem in nekatere
vrste jo lahko potegnejo pod oprsje. Pri večini vrst imajo gosenice na zadku dobro
izoblikovane ţleze, ki izločajo sladke tekočine (Preston-Mafham, 1988; Bálint, 1993). Ta
izloček je priljubljena hrana nekaterih vrst mravelj, zato mravlje pogosto liţejo sladki izloček
in v zameno branijo gosenico pred plenilci. Gosenice nekaterih vrst mravlje odnesejo v
mravljišče in jih tam gojijo (Klots in Klots, 1970).
Gosenice te druţine so v načinu ţivljenja močno raznolike. Prehrana gosenic pri modrinih se
razlikuje med poddruţinami. Dve največji poddruţini, Theclinae in Polymmmatinae, sta
fitofagni in oportunistično kanibalski, odvisno od zunanjih dejavnikov. Hranilne rastline
gosenic se razlikujejo med skupinami, raziskovanje in določitev tipičnih hranilnih rastlin pa je
teţavno (Klots in Klots, 1970). Teţave povzročajo tudi velike razlike v izbiri hranilne rastline
znotraj skupin, saj se npr. Polyommatus icarus v Britaniji pojavlja v eni generaciji na leto, v
Španiji pa tudi v štirih generacijah. Slednji tako uporabljajo skozi sezono različne hranilne
rastline ţe znotraj ene populacije (Artemyeva, 2005).
V Evropi ţivijo številne vrste iz druţine modrinov, vendar je poddruţina Polyommatini vrstno
najštevilčnejša. Samci so večinoma značilne modre barve, so pa samice rjavih odtenkov
(Dinca in sod., 2013). Pri nekaterih vrstah takšne razlike v barvi ne opazimo. Mednje sodijo
tudi vrste, ki smo jih analizirali v diplomskem delu (Preglednica 3).
Poddruţino Polyommatini sestavlja več rodov, vključno rod Polyommatus (Talavera in sod.,
2012). V diplomskem delu smo se ukvarjali z rodom Polyommatus, zato se bomo osredotočili
le na opis tega rodu.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
5
2.4 Rod Polyommatus
Rod Polyommatus je vrstno najbogatejši rod modrinov in šteje pribliţno 460 vrst, četudi je
razmeroma mlad. Je splošno razširjena skupina z največ predstavniki v palearktičnih,
neotropskih in nearktičnih regijah (Talvera in sod., 2012; Leimar, 1996).
Večina modrinčkov (Polyommatus) se hrani na rastlinah iz druţine Fabaceae (metuljnice)
(Wiemersin sod., 2010). Gosenice se po morfoloških znakih med vrstami neznatno
razlikujejo.
Rod modrinčkov (Polyommatus) vključuje navadno modro obarvane metulje (samice so
rjavih odtenkov), vendar je nekaj vrst tudi takih, kjer spolnega dimorfizma v barvi skoraj ne
opazimo. Tipska vrsta znotraj rodu je Polyommatus icarus, ki je tudi najbolj geografsko
razširjena vrsta znotraj rodu (Wiemers in sod., 2010; Artemyeva, 2005). Vrste znotraj rodu
Polyommatus imajo različno število kromosomov. Poleg različnega števila se kromosomi
vrste razlikujejo tudi po velikosti (Wiemers, 2003). Zaradi nestalnih kariotipov je klasifikacija
tega rodu problematična.
Druţina Lysandra je bila v preteklosti razdeljena na mnogo rodov, novejše raziskave pa so
pokazale drugačno sorodnost. Taksonomsko se danes druţina deli na manjše število rodov v
primerjavi z delitvijo v preteklosti, ki se naprej delijo na več podrodov (Agrodiaetus,
Lysandra, Plebicula itd.) (Wiemers, 2003; Kandul in sod., 2004; Talvera in sod., 2012).
2.5 Podrod Agrodiaetus
Podrod Agrodiaetus vključuje okrog 130 vrst morfološko precej enotnih metuljev; morfološke
razlike med vrstami so neznatne (Talvera in sod., 2012). Vrste se najpogosteje pojavljajo v
dveh barvah, modri in rjavi. Samci so po navadi obarvani modro (lahko tudi vijolično ali sivo
oranţno), samice pa so rjavih barv. Pri nekaterih vrstah sta oba spola rjava. Ker je vrst znotraj
podrodu več, kot je barvnih vzorcev, lahko pričakujemo veliko morfološko kriptičnih vrst
(Vila in sod., 2010). V zadnjih desetletjih so številne morfološko identične vrste popisali s
pomočjo kariotipov (Wiemers, 2003).

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
6
Podrod Agrodiaetus je razširjen med zahodno Palearktiko in centralno Azijo. Največja
pestrost vrst je v Kavkaškem gorovju, Iranu in Sredozemlju (Dinca in sod., 2013).
Hranilne rastline vseh vrst podrodu Agrodiaetus so iz rodov Onobrychis in Hedysarum
(druţina Fabaceae). Določevanje vrste metuljev glede na hranilno rastlino je teţavno, saj je
določevanje vrste rastline znotraj rodov metuljnic teţko (Wiemers, 2003).
V diplomskem delu smo obravnavali vrste iz podrodu Agrodiaetus: P. admetus, P. ripartii, P.
eleniae, P. aroaniensis in P. orphicus, ki so podrobneje predstavljene v nadaljevanju.
2.5.1 Polyommatus admetus (Esper, 1783)
Naseljuje stepe, predvsem pobočja in pašnike. Razširjen je na Balkanskem polotoku od
Madţarske do Grčije. Gosenice prezimijo in se kot odrasli osebki pojavljajo od junija do julija
(Tolman in Lewington, 2008). Različne vrste mravelj redno obiskujejo gosenice. Hranilni
rastlini sta Onobrychis viciifolia in Onobrychis caput-galli iz druţine metuljnic (van Swaay in
sod., 2010a). Odrasli osebki imajo šibko razvito belo črto na krilih in prisotne ter dobro
razvite submarginalne pike na krilih (Tolman in Lewington, 2008).
2.5.2 Polyommatus ripartii (Freyer, 1830)
Najdemo ga na travnatih površinah, hribovitih pobočjih in med grmovnatim rastjem.
Razširjenost je precej razdrobljena, saj ga najdemo v severni Španiji, jugovzhodni Franciji, na
juţnem Balkanskem polotoku, Turčiji in na območju od juţnega Urala do Altajskega gorovja.
Gosenica prezimi in se hrani spomladi, odrasle metulje pa lahko opazimo v poletnih mesecih.
Gosenice pogosto obiskujejo mravlje različnih rodov. Hranilne rastline so Onobrychis
viciifolia, O. arenaria, O. saxatillis in O. alba (van Swaay in sod., 2010b). Odrasli osebki
imajo dobro razvito belo črto na krilih, vendar se pri nekaterih grških populacijah pojavljajo
variacije v debelini in intenziteti črte (Tolman in Lewington, 2008; Pamperis, 2009).

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
7
2.5.3 Polyommatus aroaniensis (Brown, 1976)
Pojavlja se na suhih travnatih površinah, v grmičevju ob gozdovih, na kamnitih pobočjih in na
gozdnih travnikih. Letno se izmenja ena generacija osebkov, prezimijo pa v stadiju ličinke.
Zaradi sladkega izločka gosenice pogosto obiskujejo mravlje. Hranilna rastlina je Onobrychis
arenaria iz druţine metuljnic. Vrsta je endemična za Evropo. Najdemo jo v gorah v Grčiji in
v juţni Bolgariji na nadmorski višini 400−1500 metrov (van Swaay in sod., 2010e; Melovski
in Bozhinovsk, 2014). Bele črte na krilih pri 60−70 % osebkov ni (Tolman in Lewington,
2008; Lukhtanov in sod., 2003). Vrsto P. aroaniensis nekateri avtorji obravnavajo kot
kompleks vrst, saj genetske preiskave kaţejo moţno prisotnost več vrst (Coutsis in De Prins,
2005).
2.5.4 Polyommatus eleniae (Coutsis in De Prins, 2005)
Vrsta se pojavlja na travnatih površinah. Je endemit iz gorovja Falakro v severovzhodni
Grčiji. Hranilne rastline so Onobrychis spp. Podrobnosti o habitatu in nahajališčih niso znane,
saj je vrsta redka (van Swaay in sod., 2010d). Morfološko je vrsta zelo podobna vrsti
P. aroaniensis. Črna pika v celici na sprednjih krilih je komaj vidna ali manjka. Bela črta na
krilih je slabo vidna ali manjka. Samci imajo razvito belo črto bolje kot samice, a slabše kot
vrsta P. ripartii (Coutsis in De Prins, 2005).
2.5.5 Polyommatus orphicus (Kolev, 2005)
Velja za zelo ogroţeno vrsto. Omejena je na Rodopsko gorovje na meji med Grčijo in
Bolgarijo. Glavni dejavnik, ki omejuje razširjenost te vrste, je lokalno razširjena hranilna
rastlina Onobrychis alba. Osebke najdemo na suhih in kamnitih pobočjih (van Swaay in sod.,
2010c). Po morfoloških znakih spominja na vrsti P. aroaniensis in P. dantchenkoi. Bela črta
na krilih je dobro razvita. Submarginalne pike na krilih so slabo vidne ali jih ni, ukrivljenost

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
8
pik na krilih pa je močnejša od sorodnih vrst (Kolev, 2005). Kolev (2005) vrsto P. orphicus
obravnava kot podvrsto Polyommatus dantchenkoi (P. dantchenkoi orphicus ssp.) zaradi
enakega števila kromosomov.
2.6 Kriptične vrste
V literaturi najdemo več definicij kriptičnih vrst. Poenostavljeno lahko kriptične vrste
razloţimo kot skupino vrst, ki so bile na podlagi morfoloških znakov v preteklosti prepoznane
kot ena vrsta (Trontelj in Fišer, 2009; Bickford in sod., 2007). Kriptične vrste predstavljajo
problem, saj smo v percepciji močno omejeni na uporabo vizualne informacije, ne pa tudi
drugih signalov (kemični, zvočni), ki jih uporabljajo nevretenčarji. Zato bi morali poleg
morfoloških znakov vključevati tudi molekulske (genetske) in druge znake (npr. zvočne,
vedenjske) ţe v alfa taksonomijo (opisovanje in poimenovanje novih vrst). Prav tako bi se
morali genetski vzorci novo odkritih vrst rutinsko shranjevati. Tako se v zadnjih letih
spodbuja ponovno vzorčenje ţe znanih vrst, kar je privedlo do odkritja mnogih kriptičnih vrst
(Bickfordin sod., 2007). Kriptične vrste so pogoste zlasti pri morskih organizmih, raziskave
pa kaţejo, da so kriptične vrste pogoste tudi pri ţuţelkah (Bickford in sod., 2007). Dober
primer prisotnosti kriptičnih vrst je obravnavani podrod Agrodiaetus (Vershinina in
Lukhanov, 2010).
2.7 Črtne kode DNK
Črtne kode DNK oz. »barcoding« temelji na predpostavki, da lahko na podlagi kratke
standardizirane sekvence ločimo vrste med seboj, saj so genetske razlike med vrstami večje
kot znotraj njih (Hajibabaei in sod., 2007; Wilson, 2004). Identifikacijo vrst s pomočjo
»barcodinga« omogoča uporaba kratkega odseka DNK (pribliţno 650 baznih parov) na
standardiziranem odseku genoma. Pri ţuţelkah je to najpogosteje citokrom C oksidazni gen
podenote I (COI) genu, ki ga lahko rutinsko pomnoţimo s postopkom PCR (Frézal in Leblois,
2008; Ajmal Ali in sod., 2014). Morfološko lahko prepoznavne vrste običajno določimo s
pomočjo morfoloških znakov; nasprotno pa v številnih primerih morfologija za točno

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
9
identifikacijo vrste ne zadošča, zato se moramo zateči k uporabi črtne kode (Floyd in sod.,
2002).
2.8 Reakcija PCR
Reakcija PCR (Polymerase Chain Reaction) je metoda pomnoţevanja verige DNK s pomočjo
polimeraze DNK. Metodo je razvil Kary Mullis leta 1985. Metoda pomnoţi zelo majhne
koncentracije DNK do te mere, da so uporabni za različne analize. Encim DNK polimeraza
spodbudi nastanek komplementarne verige ţe obstoječi verigi DNK, vendar se izgradnja
verige lahko začne le na ţe obstoječem 3'-OH mestu. Za to je potreben začetni oligonukleotid,
na katerega se nato veţejo nadaljnji nukleotidi. Pri reakciji PCR ločimo 3 glavne faze: fazo
denaturacije, fazo pripenjanja in fazo podaljševanja. Za uspešno reakcijo potrebujemo
vzorčno DNK, DNK polimerazo, začetne nukleotide, dNTP (posamezni nukleotidi, iz katerih
polimeraza sestavi novo komplementarno verigo DNK) in pufre, ki reakcijo stabilizirajo in
vzdrţujejo optimalne pogoje za delovanje polimeraze. V prvi fazi reakcije, fazi denaturacije,
reakcijsko zmes segrejemo na 90−97 °C. S tem dvojno vijačnico DNK razklenemo in
omogočimo nadaljnje faze. V fazi pripenjanja se začetni oligonukleotidi veţejo na
komplementarno mesto na verigi DNK, ki je enoveriţna. Ta faza poteka pri 50−60 °C. V tretji
fazi, fazi podaljševanja, polimeraza DNK podaljšuje verigo DNK naprej od mesta, ki ustreza
začetnemu oligonukleotidu. Faza poteka pri 72 °C. Pomembno je, katero DNK polimerazo
uporabljamo, saj je npr. DNK polimeraza I nestabilna pri visokih temperaturah. Večinoma se
uporablja Taq polimeraza, ki ima optimum delovanja pri okrog 70 °C. Vse tri faze so del
enega cikla. Število ciklov je odvisno od koncentracije DNK, ki jo ţelimo imeti v končni
reakcijski zmesi. Po zadnjem ciklu po navadi sledi inkubacija reakcijske zmesi na 72 °C, zato
da se konci novonastalih verig zaključita (Mohini in Deshpande, 2010).

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
10
3 MATERIALI IN METODE
3.1 Vzorci
Vzorci so bili nabrani na različnih lokacijah v juţnem delu Balkana med leti 2010 in 2013
(Preglednica 1, Slika 1). Večina primerkov, na katerih smo opravljali genetske in morfološke
preiskave, je samcev. Preliminarna določitev vzorcev je potekala ţe na terenu. Podrobnejšo
določitev smo lahko opravili le po podrobnejši analizi genetskih in morfoloških razlik med
vzorci in s primerjavo z drugimi vrstami.
Ulovljeni osebki so shranjeni v papirnatih vrečkah, ki imajo dopisano kodo vzorca, datum in
lokacijo ulova metuljev.
Za veljavno ugotavljanje sorodstvenih odnosov smo potrebovali tudi zunanje in referenčne
vzorce. Kot zunanje vzorce smo uporabili metulje iz podrodu Lysandra, ki smo jih v
filogenetskem drevesu uvrstili v rod Polyommatus (P. belargus), tako kot kaţe večina
recentnih raziskav (Kandulin sod., 2004; Wiemers in sod., 2010), in metulje iz samostojnega
rodu Cyaniris (C. semiargus) (Talavera in sod., 2012). Referenčni vzorci spadajo v podrod
Agrodiaetus in pripadajo istim vrstam kot naši vzorci (P. admetus, P. ripartii, P. orphicus,
P. eleniae in P. aroaniensis) ali pa sorodnim vrstam, ki niso bile vzorčene
(P. nephohiptamenos, P. danchenkoi, P. humedasae). S takšno zastopanostjo bolj ali manj
sorodnih vrst smo poskušali doseči čim boljšo uvrstitev naših vzorcev v filogenetsko drevo.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
11
SEZNAM VZORCEV IN LOKALITET
Preglednica 1: Preglednica vzorcev, uporabljenih v diplomski nalogi.
Koda vzorca Lokacija ulova* Drţava ulova Datum ulova Predhodna določitev
vrste
LA35 Mavrovo; Debar,
Rosoki; 910 m Makedonija 30.7.2012
Polyommatus
ripartii
LA36 Cer; Kićevo; 1100 m Makedonija 28.6.2012 P. ripartii
LA37 Cer; Kićevo; 1100 m Makedonija 28.6.2012 P. aroaniensis
LA38 Kozjak; Skopje, Nova
Breznica; 1070 m Makedonija 13.7.2013 P. aroaniensis
LA39 Kozjak; Skopje, Nova
Breznica; 1070 m Makedonija 13.7.2013 P. aroaniensis
LA50 Brajićino Bosna in
Hercegovina 9.7.2010 P. ripartii
LA51 Jelašnica Srbija 5.7.2010 P. admetus
LA52 Pletvar Makedonija 14.7.2010 P. orphicus
LA53 Pletvar Makedonija 14.7.2010 P. ripartii
LA54 Vzhodno od Kelli Grčija 13.7.2010 P. ripartii
LA55 Lavdar Albanija 21.7.2013 P. ripartii
LA56 Lavdar Albanija 21.7.2013 P. admetus
LA57 Vzhodno od Drenove Makedonija 21.7.2013 P. ripartii
LA58 Cajupi; 1000 m Albanija 27.7.2013 P. admetus
LA59 Sheper; 1290 m Albanija 27.7.2013 P. ripartii
LA60 Siatista Grčija 26.7.2013 P. ripartii
LA61 Siatista Grčija 26.7.2013 P. aroaniensis

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
12
LA62 Granitis Grčija 27.7.2013 P. admetus
LA63 Granitis Grčija 27.7.2013 P. eleniae
LA64 Gacko Bosna in
Hercegovina 27.7.2013 P. ripartii
LA65 Falakron Grčija 22.7.2013 P. neptohiptamenos
LA66 Devoll Albanija 22.7.2013 P. ripartii
LA67 Devoll Albanija 22.7.2013 P. admetus
LA68 sever Drenove Makedonija 21.7.2013 P. admetus
LA69 Gacko Bosna in
Hercegovina 19.7.2013 P. aroaniensis
* Pri nekaterih vzorcih je označena nadmorska višina v metrih: pri drugih vzorcih ta podatek ni znan.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
13
Slika 1: Zemljevid južnega Balkana z označenimi lokacijami ulova vzorcev in unikatnimi kodami vzorcev.
3.2 Morfologija kril
Kot del morfoloških razlik med vrstami metuljev iz podrodu Agrodiaetus smo preučili tudi
vzorce kril na način, ki ga predlaga Pamperis (2009).
Uporabljali smo stereomikroskop z merilcem in pinceto.
Pregledali smo 8 fenotipskih znakov, po katerih naj bi se vrste razlikovale, in sicer:

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
14
barvo resic;
prisotnost bele črte;
prisotnost submarginalne pike na zadnjih krilih;
osnovno barvo;
prisotnost črne pike;
ukrivljenost subkostalnih pik (a/b);
prisotnost modrikastega sijaja bazalno;
prisotnost pike v S1 (Slika 2).
Slika 2: Shematski prikaz strukture krila metulja z označbami. Krilo je razdeljeno na več delov, ki so poimenovani s
črkami in številkami (slika povzeta po Tolman in Lewington, 2008).

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
15
Slika 3: Vzorci LA38, LA53 in LA60. Na slikah so vidni primeri intenzitete bele črte na krilih. Osebek levo (LA38)
črte nima, pri srednjem osebku (LA53) je črta šibka, desni osebek (LA60) pa ima izrazito belo črto.
Slika 4: Vzorec LA62, ki je na podlagi morfologije določen kot vrsta P. admetus. Za to vrsto so značilne močne
submarginalne pike.
3.3 MOLEKULARNOBIOLOŠKA ANALIZA
3.3.1 Kemikalije, potrošni material in naprave
3.3.1.1 Začetni oligonukleotidi za PCR
Začetni oligonukleotidi za odsek gena COI (Folmel in sod., 1994)
LCO: 5'-GGTCAACAAATCATAAAGATTGG-3'
HCO: 5'-TAAACTTCAGGGTGACCAAAAAATCA-3'
3.3.2 Izolacija DNK
DNK smo izolirali iz noge vzorčnih metuljev (Gen EluteTM
, Mammalian genomic DNA
Miniprep Kit, Sigma-Aldricht, ZDA). Navzkriţno kontaminacijo vzorcev smo preprečili tako,
da smo po rokovanju z vzorcem secirno orodje razkuţili v 96-odstotnem etanolu in ga obrisali
s papirnato brisačko.
Nogo vsakega metulja smo dali v svojo mikrocentrifugirko, ki smo jo označili s pripadajočo
kodo vzorca.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
16
Del izolacij DNK smo delali z izolacijskim kompletom kemikalij Sigma-Aldricht, del izolacij
pa s kompletom kemikalij Smart Helix.
Protokol Sigma-Aldricht:
Nogo vsakega vzorčnega metulja poloţimo v svojo mikrocentrifugirko, ki je primerno
označena. V vsako dodamo 180 µl raztopine Lysis T.
V vsako mikrocentrifugirko dodamo 15 µl proteinaze K.
Mikrocentrifugirke inkubiramo v stresalniku pri 58 °C uro in 20 minut.
Vzorce maceriramo z macerirno palčko in pri tem pazimo, da med maceriranjem
vsakega vzorca palčko razkuţimo v 96-odstotnem etanolu in obrišemo s papirnato
brisačko.
V vsako mikrocentrifugirko ponovno dodamo 15 µl proteinaze K.
Vzorce ponovno inkubiramo v stresalniku 1−2 uri pri 58 °C.
Vzorce centrifugiramo 3 minute pri 14000 obratih.
Tekočino odpipetiramo v nove primerno označene mikrocentrifugirke in pri tem
pazimo, da koščki maceriranega vzorca ostanejo v stari mikrocentrifugirki.
V vsako mikrocentrifugirko s supernatantom dodamo 300 µl hladnega 96-odstotnega
etanola in 10 sekund vorteksiramo.
Vsebino vsake mikrocentrifugirke prepipetiramo v svojo mikrocentrifugirko s filtrom,
ki je primerno označena.
Mikrocentrifugirke s filtrom centrifugiramo 1 minuto pri 14000 obratih.
Supernatant odlijemo in v vsako mikrocentrifugirko s filtrom odpipetiramo 500 µl
Wash solutiona.
Mikrocentrifugirke ponovno centrifugiramo 1 minuto pri 6900 obratih.
Supernatant odlijemo.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
17
Ponovno odpipetiramo 500 µl Wash solutiona v vsako mikrocentrifugirko.
Centrifugiramo 3 minute pri 14000 obratih. Supernatant odlijemo in ponovno
centrifugiramo 2 minuti pri 14000 obratih.
Filtre iz mikrocentrifugirk prestavimo v nove mikrocentrifugirke, ki so primerno
označene.
V vsako mikrocentrifugirko dodamo 30 µl Elution solutiona in pustimo 5 minut, da se
filter omoči.
Centrifugiramo 1 minuto pri 6500 obratih.
Ponovno dodamo 30 µl Elution solutiona in centrifugiramo 1 minuto pri 6500 obratih.
Supernatant shranimo v zamrzovalnik za nadaljnje raziskave.
Protokol Smart Helix:
Nogo vsakega vzorčnega metulja poloţimo v svojo mikrocentrifugirko, ki je primerno
označena. V vsako dodamo 380 µl pufra D.
Vzorce maceriramo z macerirno palčko.
Dodamo po 20 µl proteinaze K in vzorce inkubiramo v stresalniku dve uri pri 58 °C,
na koncu pa še 10 minut na 99 °C.
Vzorce centrifugiramo v centrifugi 5 minut pri 10000 obratih.
S pipeto odstranimo supernant in ga prepipetiramo v sveţe mikrocentrifugirke, ki so
primerno označene.
Dodamo B-pufer (toliko, da je mikrocentrifugirka napolnjena skoraj do vrha) in
vorteksiramo, da se supernant in B-pufer dobro premešata.
700 µl mešanice odpipetiramo v mikrocentrifugirko s filtrom.
Centrifugiramo 1 minuto pri 14000 obratih.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
18
Supernant odlijemo in v isto mikrocentrifugirko s filtrom prepipetiramo še preostalo
mešanico z vzorcem.
Centrifugiramo 1 minuto na 14000 obratih in ponovno odlijemo supernant.
V mikrocentrifugirko s filtrom odpipetiramo 600 µl W-pufra.
Centrifugiramo 1 minuto na 14000 obratih in odlijemo supernant.
V mikrocentrifugirko s filtrom odpipetiramo 500 µl W-pufra.
Centrifugiramo 1 minuto pri 14000 obratih in odlijemo supernant.
Filtre iz mikrocentrifugirk s filtri prestavimo v nove mikrocentrifugirke, ki so
primerno označene.
Na filtre odpipetiramo po 30 µl E-pufra in počakamo 5 minut, da se filter omoči.
Centrifugiramo 1 minuto pri 14000 obratih.
Ponovno dodamo 30 µl E-pufra in centrifugiramo 1 minuto pri 14000 obratih.
Supernant, ki vsebuje DNK, odpipetiramo v nove primerno označene epice in jih
shranimo v zamrzovalniku za nadaljnje preiskave.
3.3.3 Pomnoževanje DNK
3.3.3.1 Reakcija PCR
Pri pripravi vzorcev za postopek PCR smo vsak vzorec pripravili z naslednjimi kemikalijami:
2,5 µl PCR-pufra;
2,5 µl dNTP;
18,3 µl Ω-vode;

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
19
0,33 µl začetnega oligonukleotida 1 (LCO);
0,33 µl začetnega oligonukleotida 2 (HCO);
0,12 µl Taq-DNK polimeraze.
To kombinacijo in volumen kemikalij smo uporabili v vseh reakcijah, ki smo jih izvajali.
Poleg tega smo pripravili 1 volumen te mešanice za slepi vzorec, ki smo ga uporabili za
kontrolo.
Pri reakcijah PCR smo zaradi neuspelih reakcij pri določenih vzorcih spreminjali 2 parametra:
volumen vzorca in protokol PCR.
a) Prvič smo pri PCR postopku uporabili protokol COI:
4 minute pri 95 °C − denaturacija DNK;
1 minuta pri 95 °C;
1 minuta pri 45 °C − hibridizacija; ta cikel se ponovi 40-krat
2,5 minute pri 72 °C − pomnoţevanje;
7 minut pri 72 °C − podaljšana sinteza.
Ta protokol smo uporabili pri vzorcih LA35, LA36, LA37, LA55, LA56, LA57, LA58, LA59
LA62, LA64, LA65, LA66, LA67, LA68 in LA69. Uporabili smo po 2 µl vzorcev. Pri vzorcu
LA54 smo uporabili 3 µl vzorca.
b) Drugič smo pri postopku PCR uporabili spremenjen protokol COI:
4 minute pri 95 °C − denaturacija DNK;
1 minuta pri 95 °C;
1 minuta pri 44 °C − hibridizacija; ta cikel se ponovi 42-krat
2,5 minute pri 72 °C − pomnoţevanje;
7 minut pri 72 °C − podaljšana sinteza.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
20
Ta protokol smo uporabili pri vzorcih LA38, LA39, LA50, LA51, LA52, LA53 in LA61.
Vseh vzorcev smo uporabili po 2 µl.
c) Tretjič smo pri postopku PCR uporabili spremenjen protokol COI:
4 minute pri 95 °C − denaturacija DNK;
1 minuta pri 95 °C;
1 minuta pri 43 °C − hibridizacija; ta cikel se ponovi 43-krat
2,5 minute pri 72 °C − pomnoţevanje;
7 minut pri 72 °C − podaljšana sinteza.
Ta protokol smo uporabili pri vzorcih LA60, kjer smo uporabili 3 µl vzorca, in LA63, kjer
smo uporabili 2 µl vzorca.
3.3.3.2 Elektroforeza
Pred izvajanjem elektroforeze smo pripravili agarozni gel.
Priprava gela:
1 gram agaroznega gela v prahu smo v erlenmajerici zmešali s 100 ml 1xTAE-pufra. V
erlenmajerico smo dali mešalni magnet in vse skupaj postavil v mikrovalovno pečico.
Mešanico smo v mikrovalovni pečici segrevali pri nastavitvi 700 W pribliţno 1 minuto oz.
dokler se ves agarozni gel v prahu ni raztopil.
Pripravili smo elektroforezno banjico, ki smo jo opremili z vsemi potrebnimi deli (plastične
pregrade), da gel ni prišel v stik z ţicami v banjici. V pipeto smo pripravili 2 µl etidijevega
bromida. Gel smo nato počasi vlivali v banjico, istočasno pa smo s pipeto dodali tudi etidijev
bromid in dobro premešali. Pazili smo, da v gelu ni bilo mehurčkov, ki bi lahko kasneje pri
elektroforezi ovirali potovanje molekul DNK. Ko je bila banjica zalita pribliţno do polovice,
smo vstavili plastični glavniček in dolili toliko gela, da so bili zobci glavnička potopljeni vanj.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
21
Elektroforezno banjico smo nato pustili vsaj 1 uro v digestoriju, da se je gel strdil in
popolnoma ohladil.
Postopek elektroforeze:
S pipeto smo odmerili 5 µl vzorca, ki je bil pomnoţen s postopkom PCR, in ga premešali s
kapljico Loading-pufra, tako da smo vzorec in Loading-pufer večkrat posrkali s pipeto.
Mešanico smo nato s pipeto prenesli v prvo luknjico v gelu. Pri tem smo pazili, da gela nismo
prebodli ali kako drugače poškodovali. Postopek smo ponovili za vse vzorce in pri tem
menjali nastavke za pipetiranje, da ni prišlo do kontaminacije. V predzadnjo luknjico smo po
enakem postopku odmerili slepi vzorec, v zadnjo luknjico pa smo dali 1,7 µl mešanice 100
baznih parov.
Banjico smo nato pokrili s pokrovom in jo preko adapterja priklopil na napetost 45 V za 5−10
minut. Po tem času smo napetost povečali na 60 V. Ko je fronta pripotovala do pribliţno
polovice gela, smo napetost izklopili.
Z banjice smo odstranili pokrov, odlil TAE-pufer in vse skupaj splaknil z malo vode.
Banjico smo nato odnesli do UV-kamere, kjer smo gel slikali in tako preverili prisotnost
zadostne količine DNK v vzorcu. DNK na sliki pod UV-svetlobo izgleda kot svetleča črtica.
Močnejši kot je sijaj, več DNK je v vzorcu.
3.3.3.3 Čiščenje DNK
Vzorce, ki so imeli dovolj veliko koncentracijo DNK po postopku PCR, smo očistili in poslali
na sekvenciranje.
Čiščenje je potekalo v napravi PCR. Po izvedbi elektroforeze je vsakega vzorca ostalo
pribliţno 20 µl.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
22
Postopek čiščenja:
V vsako epico smo dodali 0,4 µl eksonukleaze 1-EXO I in 2 µl alkalne fosfataze FAST AP.
Vzorce smo nato prestavili v napravo PRC in vklopili program čiščenja: 47 minut pri 37 °C in
15 minut pri 80 °C.
Po sekvenciranju v tujini smo nazaj dobili sekvence vzorcev v elektronski obliki.
3.3.4 Postopek izdelave filogenetskih dreves in Bayesova analiza
Sekvence, ki smo jih po pošiljanju na sekvenciranje dobili nazaj v elektronski obliki, smo
obdelali z naslednjimi programi:
ChromasPro (verzija 1.7.6);
BioEdit Sequence Alignment Editor (verzija 7.2.5);
NotePad (verzija 7.1 build 7600);
ClustalX2 (verzija 2.1);
MEGA (verzija 6.05(6140107));
Model Generator (verzija 82);
REDSECQ;
MrBayes (verzija 3.2.0);
FigTree (verzija 1.4.0);
CorelDraw X6 (verzija 16.0.0.707).

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
23
Sekvence, ki smo jih dobili, so bile v formatu .ab1. Vsako datoteko smo odprli v programu
ChromasPro in jo pregledali. Za vsak vzorec smo dobili sekvence iz obeh smeri branja. V
istem programu smo obe datoteki zdruţili v komplementarno sekvenco. Datoteko smo shranili
v formatu .gpj. Postopek smo ponovili za vse vzorce. Sekvenco vsakega vzorca smo skopirali
v program NotePad, kjer smo dobili zaporedje nukleotidov, ki se pojavljajo v vzorcih.
Poleg naših vzorcev smo na spletni bazi podatkov NCBI (http://www.ncbi.nlm.nih.gov/)
poiskali sekvence vrst, ki so filogenetsko sorodni rodu Polyommatus in vrstam iz podrodu
Agrodiaetus (Preglednica 2). Sekvence smo skopirali v program NotePad. Vzorce, ki smo jih
dobili s te spletne strani, smo uporabljali kot zunanjike in referenčne vzorce v filogenetskem
drevesu, ki je končni produkt te obdelave.
Datoteke smo nato odprli v programu ClustalX2. Vse sekvence so bile enakih dolţin in smo
jih poravnali brez presledkov. Iz poravnave so bili razvidni odstopanja in napake, ki so nastale
pri sekvenciranju. Nekatere napake smo lahko popravili ročno glede na ostala zaporedja,
nekatere pa smo pustili nepopravljene zaradi moţnih razlik v sekvencah vzorcev DNK.
Urejene, popravljene in poravnane sekvence smo nato shranili v formatu .fasta.
Preden smo začeli z Bayesovo analizo, smo v programu MEGA izrisali preliminarno
filogenetsko drevo na podlagi metode največjega verjetja »maximum likelihood«, iz katerega
smo lahko videli pribliţno porazdelitev vzorcev glede na referenčne sekvence. V programu
MEGA smo uporabili naslednje parametre: metodo Bootstrap s 100 ponovitvami, nastavitev
GTR (nastavitev general time reversible) in nastavitev G + I (gamma distributed with
invariable sites). Nukletotide, ki niso bili jasno odčitani, smo izključili iz analize (complete
deletion). Program je izrisal filogenetsko drevo, ki smo ga shranili v formatu .emf.
V naslednjem koraku smo datoteko .fasta uvozili v program Model Generator, ki za izbrani
niz podatkov preverja ustreznost različnih evolucijskih modelov in izmed 56 moţnosti izbere
najprimernejšega. Program je za moţne modela predlagal: GTR + I + G in TIM + I + G. Za
nadaljevanje analize smo izbrali model TIM + I + G (transition model + invarient sites +
gamma distrtibuted), ker se je kot predlog pojavil pri več različnih pristopih iskanja.
Datoteko .fasta smo nato v programu REDSECQ pretvorili v datoteko formata .nexus, ki se
uporablja pri Bayesovi analizi.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
24
Nadaljevali smo z Bayesovo analizo, ki se izvaja s programom MrBayes. Po uvozu datoteke
.nexus v program MrBayes smo določili parametre, po katerih je potekal izračun
filogenetskega drevesa, in sicer: NST = 6, rec = gamma (definira, po katerem modelu
evolucije naj program generira drevo), ngen = 2000000 in savebrlens = yes (2 milijona
ponovitev z moţnostjo shranjevanja dolţine vej). Po vseh ponovitvah se vsa drevesa med
seboj primerjajo. Izbrisali smo prvih 5000 dreves, ostala pa so vključena v končno
filogenetsko drevo. Pri tem so vrednosti na vejah posteriorne verjetnosti, ki povedo,
kolikokrat se posamezna veja ponovi (Ronquist in sod., 2011).
Dobljeno datoteko smo nato odprli s programom FigTree, v katerem smo lahko drevo tudi
urejali. Za urejanje smo uporabljali program CorelDraw.
Preglednica 2: Seznam referenčnih in zunanjih vzorcev, vključenih v filogenetsko drevo, s pripadajočimi unikatnimi
kodami v bazi podatkov NCBI (http://www.ncbi.nlm.nih.gov/).
VRSTA KODA VZORCA
P. admetus MW98084
P. admetus 09-V962
P. admetus AY556986
P. admetus 09-V963
P. admetus AY556867
P. admetus JC01014
P. ripartii HM210168
P. ripartii HM210167
P. ripartii 11-G195
P. ripartii EF104603
P. ripartii 11-G192
P. ripartii 11-G198
P. ripartii AY556962
P. ripartii 11-G194
P. ripartii 11-G197
P. ripartii HM210172
P. ripartii AY556858
P. ripartii 12-M016

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
25
P. ripartii J903
P. ripartii 12-M017
P. nephohiptamenos JC00045
P. nephohiptamenos AY556860
P. nephohiptamenos 09-V964
P. nephohiptamenos AY556859
P. nephohiptamenos JC00046
P. humedasae HM210169
P. dantchenkoi AY557081
P. dantchenkoi MW99276
P. dantchenkoi MW99319
P. dantchenkoi AY557073
P. dantchenkoi AY557072
P. aroaniensis JC00040
P. aroaniensis AY556856
P. icarus MW98133
P. icarus MW00412
P. bellargus RV-07-D435
P. bellargus RV-07-D149
C. semilargus MW00525
C. semilargus MW02034

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
26
4 REZULTATI
Analizirali smo 25 osebkov, ki glede na molekularne analize pripadajo petim vrstam:
Polyommatus ripartii, Polyommatus admetus, Polyommatus eleniae, Polyommatus orphicus
in Polyommatus aroaniensis. Ti rezultati niso skladni s fenotipskimi lastnostmi analiziranih
osebkov. V nadaljevanju povzemamo fenotipske razlike med osebki in rezultate filogenetske
analize.
4.1 Morfologija barvnih vzorcev na krilih
Pri morfološki diferenciaciji osebkov smo uporabljali znake na krilih, ki so v literaturi
uporabljeni za diferenciacijo vrst po zunanjih znakih (Lukhtanov in sod., 2003; Coutsis in De
Prins, 2005; Kolev, 2005; Tolman in Lewington, 2008; Pamperis, 2009). Za določanje vrst
naših vzorcev smo uporabili tudi genetske markerje, ki so vzorce razvrstili v monofiletske
skupine, ki ustrezajo zgoraj naštetim vrstam. Delitev na osnovi filogenetskih odnosov smo
uporabili za analizo morfoloških razlik med osebki.
V nadaljevanju primerjamo preliminarno uvrstitev vzorcev s tabelo opazovanih fenotipskih
znakov (Preglednica 3).
Značilni znaki za vrsto P. admetus so prisotna črna pika v celici in močno vidne (dvojne)
submarginalne pike (Pamperis, 2009) (Slika 4). Oba znaka se pojavljata pri vzorcih LA51,
LA56, LA58, LA62, LA67 in LA68. Vsi navedeni vzorci so bili na podlagi morfologije
pravilno uvrščeni.
Eden glavnih določevalnih znakov za vrsto P. ripartii je prisotna močna bela črta z
variacijami v dolţini in intenziteti pri nekaterih grških populacijah (Tolman in Lewington,
2008; Pamperis, 2009) (Slika 3). Temu kriteriju ustrezajo vzorci LA36, LA50, LA53, LA54,
LA55, LA57, LA59, LA60, LA64 in LA66. Za navedene vzorce predvidevamo, da spadajo v
vrsto P. ripartii. Iz Preglednice 3 je razvidno, da je bela črta prisotna ne le pri P. ripartii,
temveč tudi pri drugih vrstah. Diagnostična vrednost tega znaka je vprašljiva.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
27
Za vrsto P. aroaniensis so značilni naslednji znaki: bela črta šibko vidna ali manjka,
submarginalnih pik ni in črne pike v celici, ki pa lahko manjkajo (Tolman in Lewington,
2008; Pamperis, 2009). Tem kriterijem delno ustrezajo vzorci LA37, LA38, LA39, LA61 in
LA69. Slednji je edini iz klada, ki ima celico s črno piko. Četudi so bili nekateri izmed teh
vzorcev predhodno določeni pravilno, se niti en osebek v celoti ne ujema z diagnostičnimi
znaki.
P. eleniae je lokalizirana vrsta, ki se nahaja v okolici gore Falakron v Grčiji. Značilna
fenotipska znaka sta komaj vidna oz. odsotna črna pika v celici na sprednjih krilih in slabo
vidna oz. odsotna bela črta na krilih (Tolman in Lewington, 2008; Pamperis, 2009). Tem
kriterijem odgovarja vzorec LA63, ki je skladen z vsemi tipičnimi fenotipskimi znaki za to
vrsto. Tudi lokacija ulova je v območju znane razširjenosti vrste (Coutsis in De Prins, 2005).
Vrsta, ki smo jo glede na morfologijo poimenovali P. orphicus, bi bila lahko tudi podvrsta
vrste P. dantchenkoi (Pamperis, 2009; Kolev, 2005). Pri poimenovanju smo se sklicevali na
morfologijo, saj v bazi GenBank ni bilo referenčnega taksona. Kot P. orphicus smo
preliminarno določili vzorec LA52. Iz Preglednice 3 je razvidno, da se vzorec le delno ujema
z značilnimi fenotipskimi znaki za to vrsto. Submarginalne pike so odsotne, bela črta pa je
močna in dobro vidna. Preliminarna določitev je v tem primeru vprašljiva.
Vzorec LA65 je po morfologiji in lokaciji preliminarno uvrščen k vrsti P. neptohiptamenos
(tipična fenotipska znaka sta široka bela črta in črna pika v celici) (Pamperis, 2009), vendar
rezultati filogenetske analize niso potrdili te trditve. Moţna je napačna uvrstitev ali napaka v
postopku PCR. Tudi fenotipsko vzorec ni povsem skladen s to vrsto, saj manjka črna pika v
celici.
Večina preliminarnih uvrstitev je vprašljiva in nezanesljiva zaradi velike variabilnosti
fenotipskih znakov. To variabilnost dobro ponazarja primer vrste P. eleniae, kjer so lahko vsi
tipični fenotipski znaki tudi odsotni.
Za natančnejšo uvrstitev smo opravili filogenetske raziskave, ki so pokazale dejansko vrstno
pripadnost.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
28
4.1.1 Preglednica opazovanih fenotipskih znakov
Preglednica 3: Seznam vzorcev s pripadajočimi opazovanimi fenotipskimi znaki. Vrednosti v tabeli so kvalitativne. *Podani sta 2 barvi; prva barva predstavlja barvo
resic pri proksimalnem delu resice, druga barva pa predstavlja barvo resic pri distalnem delu resice. **Ukrivljenost subkostalnih pik je vrednost, ki predstavlja
razdaljo od roba krila do ocelne točke (b) in razdaljo od ocelne točke do celice (a) (Pamperis, 2009).
Koda vzorca
Barva resic* Bela črta Submarginalne pike
na zadnjih krilih Osnovna
barva Črna pika v celici
Ukrivljenost subkostalnih pik
(a/b)**
Modrikast sijaj bazalno
Bazalna pika v S1 Pravilna določitev
vzorca
LA35 rjava/ siva šibka niso prisotne svetlo rjava
prisotna šibka rahel ni prisotna ne
LA36 rjava/ sivo
rjava močna slabo vidne
svetlo rjava
ni prisotna šibka rahel ni prisotna ne
LA37 rjava/ sivo
rjava ni prisotna niso prisotne
temno rjava
ni prisotna šibka rahel ni prisotna ne
LA38 svetlo
rjava/siva ni prisotna niso prisotne rjava ni prisotna močna rahel ni prisotna ne
LA39 rjava/ belo
rjava šibka−
polovična niso prisotne
temno rjava
ni prisotna močna rahel ni prisotna ne
LA50 rjava/ sivo
rjava šibka slabo vidne rjava ni prisotna močna rahel ni prisotna ne
LA51 temno rjava/
sivo rjava ni prisotna prisotne − močne
temno rjava
prisotna močna rahel prisotna da − P. admetus
LA52 temno rjava/
sivo bela močna niso prisotne rjava prisotna močna rahel ni prisotna ne
LA53 rjava/ svetlo
rjava šibka niso prisotne rjava ni prisotna močna rahel ni prisotna ne
LA54 temno rjava/
sivo rjava šibka slabo vidne
temno rjava
ni prisotna šibka prisoten ni prisotna ne
LA55 rjava/ sivo
rjava šibka niso prisotne
svetlo rjava
ni prisotna močna prisoten ni prisotna ne
LA56 siva/ siva šibka prisotne − močne svetlo rjava
prisotna močna prisoten prisotna da − P. admetus
LA57 temno rjava/
sivo rjava močna prisotne
temno rjava
ni prisotna šibka rahel ni prisotna ne

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
29
LA58 temno rjava/
rjavo siva ni prisotna prisotne − močne
temno rjava
prisotna šibka rahel ni prisotna da − P. admetus
LA59 rjavo bela/belo
siva močna prisotne
temno rjava
prisotna šibka rahel ni prisotna ne
LA60 svetlo rjava/
sivo bela močna niso prisotne
temno rjava
ni prisotna šibka rahel ni prisotna ne
LA61 temno rjava/
sivo rjava šibka niso prisotne
temno rjava
ni prisotna močna rahel ni prisotna da − P. aroaniensis
LA62 svetlo rjava/ svetlo rjava
ni prisotna prisotne − močne temno rjava
prisotna močna rahel prisotna da − P. admetus
LA63 svetlo rjava/ svetlo rjava
šibka, široka
niso prisotne svetlo rjava
ni prisotna šibka rahel ni prisotna da − P. eleniae
LA64 temno rjava/ svetlo rjava
šibka slabo vidne temno rjava
ni prisotna šibka rahel ni prisotna ne
LA65 svetlo rjava/
siva močna slabo vidne
temno rjava
ni prisotna šibka rahel ni prisotna ne
LA66 temno rjava/ svetlo rjava
močna niso prisotne temno rjava
ni prisotna močna rahel ni prisotna ne
LA67 temno rjava/ svetlo rjava
ni prisotna prisotne − močne temno rjava
prisotna šibka rahel ni prisotna da − P. admetus
LA68 svetlo rjava/
siva ni prisotna prisotne − močne
svetlo rjava
prisotna močna rahel ni prisotna da − P. admetus
LA69 svetlo rjava/
siva ni prisotna niso prisotne
svetlo rjava
prisotna močna rahel ni prisotna ne

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
30
4.2 Genetska podobnost med osebki
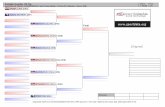
V filogenetskem drevesu se glede na referenčne vzorce in zunanjike naši vzorci uvrščajo v 5
skupin, ki ustrezajo 5 ločenim vrstam: P. ripartii, P. admetus, P. eleniae, P. orphicus in P.
aroaniensis (Slika 5). Vzorce smo glede na filogenetsko drevo razporedili v naslednje vrste:
P. ripartii, vzorci: LA36, LA50, LA52, LA53, LA54, LA55, LA57, LA59, LA64,
LA66 in LA69;
P. admetus, vzorci: LA51, LA56, LA58, LA62, LA67 in LA68;
P. aroaniensis, vzorci: LA35 in LA61;
P. eleniae, vzorci: LA63 in LA65;
P. orphicus, vzorci: LA37, LA38, LA39 in LA60.
V filogenetskem drevesu je jasno ločen kompleks P. aroaniensis z vsemi sorodnimi vrstami:
P. humedaseae, P. dantchenkoi in P. orphicus. Vrste znotraj te skupine so si med seboj
sorodne, sorodstveni odnosi med njimi pa niso povsem razjasnjeni. Globoke cepitve vej
znotraj skupine aroaniensis kaţejo, da so znotraj skupine verjetno prisotne še druge kriptične
vrste. Nekateri avtorji v to skupino uvrščajo tudi vrsto P. eleniae (Coutsis in De Prins, 2005),
ki pa je v naši analizi bliţje vrsti P. admetus. Pod skupino eleniae lahko opazimo tudi skupino
štirih vzorcev iz baze NCBI, določenih kot vrsta P. ripartii. Takšna razvrstitev lahko pomeni
moţno napačno uvrstitev teh osebkov ali moţnost prisotnosti kriptične vrste, saj so ti vzorci iz
vzhodnega dela Turčije, torej so geografsko ločeni od ostalih vzorcev P. ripartii. Podporo
kladov smo določili s posteriorno verjetnostjo, ki je bila relativno nizka (pod 95 %).
Znotrajvrstni kladi, predvsem pri vrsti P. ripartii, ima podpore večinoma celo pod 50 %.
Areali vrst se glede na lokacije ulova vzorcev delno prekrivajo. Simpatrijo opazimo pri vrstah
P. orphicus in P. ripartii, saj se obe pojavljata na Ceru v Makedoniji (vzorca LA37 in LA36).
Tudi vrsti P. aroaniensis in P. orphicus se pojavljata na isti lokaciji, in sicer v Siatisti v Grčiji
(vzorca LA60 in LA61). Vrsti P. admetus (vzorci LA56, LA67 in LA68) in P. ripartii (vzorci
LA55, LA57 in LA66) se prav tako pojavljata na istih lokacijah. Prekrivanje lahko opazimo v

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
31
Lavdarju v Albaniji (LA55 in LA56), v Drenovi v Makedoniji (LA57 in LA68) in v Devollu v
Albaniji (LA66 in LA67).

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
32
Slika 5: Filogenetsko drevo podobnosti, izračunano po metodi Bayes. Imena vrst, ki so obarvana, so naši vzorci, črno
obarvane vrste pa so referenčni vzorci ali zunanji vzorci. Oznake na razvejitvah so vrednosti posteriorne verjetnosti,
izražene v %. Z barvami so označeni naši vzorci, črno pa so obarvani zunanji in referenčni vzorci iz baze GenBank.
Znotraj skupine aroaniensis se nahajajo tudi vzorci, določeni kot P. orphicus.
- skupina ripartii
- skupina admetus
- skupina eleniae
- skupina aroaniensis

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
33
5 RAZPRAVA
5.1 Primernost fenotipskih znakov na krilih za ločevanje vrst
Od 9 opazovanih fenotipskih znakov se je za uporabne pokazalo le 5 znakov na spodnji strani
zadnjih kril, in sicer bela črta na krilih, submarginalne pike, črna pika v celici, ukrivljenost
subkostalnih pik in bazalna pika v S1. Barva resic je zaradi prevelike podobnosti med
analiziranimi osebki neuporaba za ločevanje. Osnovna barva kril se sicer uporablja za
ločevanje med vrstami, predvsem na terenu (Pamperis, 2009), vendar tudi ta fenotipski znak
ni zanesljiv, saj gre za razlike v odtenkih barv, ki so po navadi določeni subjektivno. Tako bi
npr. lahko vzorec LA38 glede na druge fenotipske znake razvrstili v vrsto P. orphicus (šibka
ali odsotna bela črta, submarginalne pike odsotne ali slabo vidne, ukrivljenost subkostalnih
pik je močna). To uvrstitev podpirajo tudi filogenetski rezultati, vendar je osnovna barva v
literaturi navedena kot značilno rumeno rjava s sivimi odtenki na bazalnem delu (Kolev,
2005), vzorec pa smo določili kot tipično rjave barve. Tudi druge vzorce, določene kot
P. orphicus, smo označili kot temno rjave barve, kar še bolj odstopa od navedb v literaturi.
Kot nezanesljiv znak pri naših vzorcih se je pokazal tudi modrikast sijaj na krilih. Ta se je
sicer pojavljal pri vseh osebkih, pri treh celo bolj izrazito (vzorci LA54, LA55 in LA56),
vendar le bazalno na krilih. Zaradi premajhne variabilnosti v modrikastem sijaju med našimi
vzorci ta znak ni primeren za določanje vrste. Trenutne raziskave kaţejo, da se je redukcija
modre obarvanosti znotraj podrodu zgodila večkrat neodvisno (Wiemers, 2003).
Določevanje vrst vzorcev glede na morfološke znake je teţavno. Po morfologiji smo lahko z
gotovostjo uvrstili le vzorce, ki smo jih določili kot P. admetus. Zanesljiva prepoznava drugih
vrst na podlagi znanih znakov ni moţna. Prav tako na vrstno pripadnost ne moremo sklepati iz
geografske razširjenosti vrst, saj se nekatere vrste pojavljajo simpatrično.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
34
5.2 Določitev vzorcev s filigenetsko analizo
Filogenetsko sorodnost smo ugotavljali na podlagi mitohondrijskega gena COI z Bayesovo
analizo. Med dobro podprtimi monofiletskimi kladi je tudi morfološko prepoznavna vrsta
P. admetus, ki je sicer z nizko podporo sestrska lokalnemu endemitu P. eleniae. Dobro
podprta je tudi tesna sorodnost P. admetus in P. eleniae s P. ripartii. To se ne ujema s podatki
iz literature, kjer vrsto P. eleniae obravnavajo kot sorodno skupini P. aroaniensis oz. jo celo
uvrščajo v kompleks aroaniensis, znotraj katerega lahko prepoznamo več vrst in podvrst zelo
sorodnih metuljev, med katerimi odnosi še niso popolnoma pojasnjeni (Coutsis in De Prins,
2005). Druga moţna razlaga sorodnosti je, da je določitev vzorcev LA63 in LA65 kot P.
eleniae napačna. Četudi se fenotipski znaki le delno ujemanja z originalnim opisom te vrste,
je ta razlaga malo verjetna, saj sta bila oba osebka ujeta znotraj znanega območja razširjenosti
te vrste in blizu tipske lokacije v okolici gore Falakron (Coutsis in De Prins, 2005). Ţal v bazi
nukleotidnih zaporedij (NCBI) ni vzorcev P. eleniae, s katerimi bi uvrstitev naših vzorcev
nedvoumno potrdili.
Vrsta P. ripartii je parafiletska, saj vključuje tudi osebke iz vzhodne Turčije. Ti tvorijo dobro
podprt klad, ki se bazalno odcepi od skupine, v kateri so vključene prej omenjene vrste.
Zaradi velike razdalje med Balkanskim polotokom in vzhodno Turčijo je moţno, da osebki iz
slednje pripadajo kriptični oz. še neopisani vrsti, ki se morfološko ne razlikuje od P. ripartii.
Za potrditev te trditve bi potrebovali vzorce P. ripartii tudi iz vmesnega območja. Vrsta
P. neptohiptamenos, ki je endemit visokih gora v vzhodni Grčiji, je sestrska kladu prej
omenjenih vrst.
Četrta velika skupina, ki je jasno ločena od prej omenjenih vrst in je podprta z visoko
posteriorno verjetnostjo, je skupina aroaniensis, znotraj katere se je razvrstilo 6 vzorcev
fenotipsko pripadajočima vrstama P. aroaniensis in P. orphicus. Poleg naših vzorcev sta
znotraj skupine tudi vrsti P. dantchenkoi in P. humedaseae. Slednja je endemit zahodnih Alp
in je tudi geografsko dobro ločena od ostalih vrst. Vrsta P. orphicus je glede na nekatere
avtorje le podvrsta P. dantchenkoi, saj imata obe enako število kromosomov (Coutsis in De
Prins, 2005; Kolev, 2005). Takšna klasifikacija pri naši raziskavi ni podprta, saj je skupina
P. orphicus jasno ločena. Ločitev sicer ni podprta z visoko posteriorno vrednostjo (54 %),
zato je razporeditev v filogenetskem drevesu nezanesljiva. Vrsta je glede na rezultate celo bolj

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
35
sorodna vrsti P. humedaseae ali P. aroaniensis, ki pa je glede na razporeditev vzorcev
parafiletska. Ţal zaradi premajhnega števila vzorcev in velikih razlik v nukleotidnih
zaporedjih ne moremo ugotoviti, ali posamezne veje predstavljajo še dodatne kriptične vrste.
Vzorci torej kaţejo v filogenetskem drevesu razdelitev v štiri glavne skupine, in sicer v
skupino ripartii, skupino admetus, skupino eleniae in skupino aroaniensis, znotraj katere so
razlike v nukleotidnih zaporedjih veliko večje kot v primeru P. ripartii in P. admetus. Na
podlagi tega lahko sklepamo, da so v skupini aoraniensis vsaj štiri ločene vrste, vendar pri
takšni delitvi dopuščamo moţnost napake zaradi majhnega števila vzorcev.
V filogenetskem drevesu se v skupini aroaniensis pojavljajo vrste, ki v to skupino spadajo
tudi glede na literaturo (Wiemers, 2003). Sorodnost znotraj skupine aroaniensis še ni
popolnoma raziskana, posebej po odkritju novih vrst ali kriptičnih vrst, ki so sorodne ţe
znanim skupinam. V našem filogenetskem drevesu je znotraj skupine aroaniensis P. orphicus
ločena vrsta, vendar je ujemanje s tipičnimi fenotipskimi znaki (Kolev, 2005) le delno. Vrsta
je bila do sedaj znana le na območju Rodopov v Bolgariji, zato se tudi lokacije ulova naših
vzorcev ne ujemajo z znano razširjenostjo te vrste (Kolev, 2005). Tudi v tem primeru v bazi
nukleotidnih zaporedij (NCBI) ni vzorcev P. orphicus, s čimer bi lahko nedvoumno potrdili
uvrstitev naših vzorcev.
5.3 Alopatrija in razširjenost vrst
Glede na zastavljene hipoteze naj bi se genetsko bolj sorodne vrste pojavljale alopatrično, saj
naj bi nastale z alopatrično speciacijo. Temu več kot očitno ni tako, saj se nekatere sorodne
vrste pojavljajo v simpatriji. Vzorec LA60 (P. orphicus) in vzorec LA61 (P. aroaniensis) sta
bila ulovljena na isti lokaciji, in sicer v Siatisti na severu Grčije. P. aroaniensis se pojavlja
tudi v Mavrovem na zahodu Makedonije, P. orphicus pa se pojavlja tudi v Kozjaku na severu
Makedonije in v Ceru na zahodu Makedonije. Do sedaj je bila vrsta P. orphicus znana le na
območju gorovja Rhodopi v Bolgariji (Kolev, 2005). Če naši vzorci dejansko pripadajo tej
vrsti, sega njen areal precej bolj proti zahodu do Makedonije in osrednjega dela severne
Grčije (Slika 6).

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
36
Slika 6: Zemljevid območja Makedonije, severne Grčije in Bolgarije z označenimi lokacijami vzorcev LA37, LA38,
LA39 in LA60, v filogenetskem drevesu določenih kot vrsta P. orphicus. S črko »R« je označeno območje gore
Rhodopi, ki velja za nahajališče vrste P. orphicus (Kolev, 2005). Rumeno je označeno hipotetično območje
razširjenosti te vrste glede na filogenetske rezultate in mesta ulova vzorcev.
Drug primer simpatrije ozko sorodnih vrst sta P. eleniae in P. admetus pri vasi Granitis
jogozahodno od gore Falakron. Glede na filogenetske raziskave sta to sestrski vrsti. Vrsti
P. admetus in P. ripartii imata večje območje razširjenosti in se pojavljata tudi izven
Balkanskega polotoka (Tolman in Lewington, 2008). Tudi ti dve sorodni vrsti se pojavljata
simpatrični v juţnem delu Albanije v Lavdarju, soteski Devollu (vzorca LA66 in LA67) in
okolici Drenove (vzorca LA57 in LA68). Na podlagi teh rezultatov lahko sklepamo, da
alopatrična distribucija pri teh vrstah ni nujno prisotna in lahko zavrnemo hipotezo o alopatriji
ozko sorodnih vrst.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
37
6 SKLEPI
Od 25 metuljev iz rodu Polyommatus smo v diplomskem delu pregledali fenotip kril vsakega
osebka. Na podlagi analize fenotipskih znakov v večini primerov nismo mogli potrditi
preliminarne določitve vrst, saj so bili znaki znotraj vrst preveč variabilni. Morfološko smo
lahko določili le osebke vrste P. admetus (LA51, LA56, LA58, LA62, LA67 in LA68). Kljub
temu lahko fenotipske znake uporabljamo kot pomoč pri identifikaciji ali vsaj za preliminarno
klasifikacijo na terenu, vendar takšna določitev ni verodostojna brez molekularnih podatkov.
Rezultati filogenetskih raziskav kaţejo na to, da so molekularno-filogenetske tehnike bolj
primerne za določevanje vrst tega kompleksa. Vseeno je treba upoštevati moţnost
nenatančnega pomnoţevanja fragmentov, ki je pogosta teţava pri tej metodi (Palumbi, 1996).
Teţava pri uporabi molekularnih podatkov so slabo raziskani sorodstveni odnosi znotraj rodu
Polyommatus in neenotnost avtorjev glede le-teh (Wiemers, 2003; Kandul in sod., 2004;
Talavera in sod., 2012).
Iz naših rezultatov je razvidno, da so vrste P. admetus, P. elleniae in P. ripartii ozko sorodne.
P. ripartii je razdeljena v dve parafiletski skupini. Ločeni klad iz vzhodne Turčije bi lahko
predstavljal kriptično vrsto.
V skupini aroaniensis so sorodstveni odnosi med vrstami nejasni, razvidno pa je, da
P. orphicus ne pripada vrsti P. dantchenkoi, čeprav je bil opisan kot njena podvrsta (Kolev,
2005).
V analizi smo ugotovili, da vrsta P. orphicus ni znana le iz Rodopov v Bolgariji (Kolev,
2005), temveč ţivi tudi v Makedoniji in Grčiji.
Ozko sorodne vrste se pogosto pojavljajo simpatrično. Simpatrijo smo ugotovili med pari vrst
P. ripartii in P. admetus, P. admetus in P. eleniae, P. aroaniensis in P. orphicus ter P.
orphicus in P. ripartii. Zaradi pojavljanja genetsko bolj podobnih vrst na istih lokacijah lahko
sklepamo, da v primeru rodu Polyommatus ne gre za alopatrično distribucijo. Simpatrija je
dokaz, da so tudi filogenetsko bolj sorodne vrste med seboj ločene z reproduktivno bariero.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
38
7 POVZETEK
V diplomskem delu smo se ukvarjali z določevanjem vzorcev metuljev iz podrodu
Agrodiaetus, ulovljenih na Balkanskem polotoku. Poleg filogenetskih raziskav smo
kvalitativno opisali tudi fenotip kril oz. 8 najpogosteje uporabnih znakov za prepoznavo vrst.
Vsi vzorci spadajo v rod metuljev Polyommatus. Vrste, določene s filogenetsko analizo, smo
primerjali z njihovo geografsko razširjenostjo in ugotavljali, ali se vrste pojavljajo
simpatrično.
V raziskavi smo imeli 25 osebkov z območja Balkanskega polotoka, in sicer iz Grčije,
Makedonije, Albanije, Bosne in Hercegovine ter Srbije. Vsak vzorec smo pregledali pod
stereomikroskopom in opazovali naslednje fenotipske znake: barvo resic, prisotnost bele črte,
prisotnost submarginalne pike na zadnjih krilih, osnovno barvo, prisotnost črne pike,
ukrivljenost subkostalnih pik (a/b), prisotnost modrikastega sijaja bazalno in prisotnost pike v
S1. Mnoge vrste kaţejo veliko znotrajvrstno variabilnost v morfologiji. Po morfologiji smo
lahko določili osebke vrste P. admetus, ki imajo edini prisotne močno vidne submarginalne
pike na krilih. Razlikovanje med drugimi vrstami na podlagi fenotipskih znakov je
nezanesljivo.
V drugem delu raziskave smo iz nog vzorcev izolirali DNK in sekvencirali dele
mitohondrijskega gena COI. Iz sekvenc smo nato z Bayesovo analizo ugotavljali sorodnost
med vzorci, ki je prikazana v obliki filogenetskega drevesa. Po tej metodi so se vzorci
razvrstili v 5 vrst znotraj rodu Polyommatus: Polyommatus ripartii, P. admetus, P. eleniae, P.
aroaniensis in P. orphicus. Poleg delitve na vrste so bili dobro podprti štirje kladi glede na
vzorec zdruţevanja v večje skupine, ki so jih tvorili skupaj z referenčnimi vzorci zaporedij
DNK iz baze GenBank. Te skupine so: skupina ripartii, skupina admetus, skupina eleniae in
skupina aroaniensis. Te skupine so deloma podprte tudi v drugih molekulskih analizah tega
podrodu (Kandul in sod., 2004; Schmitt in sod., 2005).
Bliţnja sorodnost vrst P. ripartii in P. admetus se ujema z dosedanjimi raziskavami (Vila in
sod., 2010). Znotraj skupine aroaniensis je vrsta P. orphicus ločena od P. dantchenkoi, katere
podvrsta naj bi bila (Kolev, 2005). Samostojen status te vrste je dobro podprt, vendar so
odnosi med vrstami v tej skupini nedoločeni zaradi velikih genetskih razlik med vzorci in

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
39
manjšega števila vzorcev morfološko določenih kot P. aroaniensis, zato so potrebne dodatne
raziskave in obseţnejše vzorčenje.
Hipotezo o alopatriji ozko sorodnih vrst smo ovrgli, saj se več vzorcev različnih ozko
sorodnih vrst pojavlja na istih lokacijah. Simpatrične so vrste P. ripartii in P. admetus,
P. eleniae in P. admetus ter P. orphicus in P. admetus. Glede na filogenetske raziskave lahko
sklepamo, da vrsta P. orphicu ni razširjena le v Rodopih v Bolgariji (Kolev, 2005), ampak
njen areal sega tudi v Makedonijo in severno Grčijo.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
40
8 VIRI
Ajmal Ali M., Gyulai G., Hidvégi N., Kerti B., Al Hemaid M. A. F., Pandey K. A., Lee J.
2014. The changing epitome of species identification- DNA barcoding. Saudi Journal of
Biological Sciences 21: 204-231.
Artemyeva E. A. 2005. Clinal variation in populations of the Common Blue butterfly
Polyommatus icarus Rott. (Lepidoptera, Lycaenidae). Russian Journal of Genetics 41 (8):
859-870.
Bálint Z. 1993. The threatened lycaenids of the Carpathian Basin, east-central Europe.
Conservation Biology of Lycaenidae (Butterflies). Ed. New T. R. The IUCN Species Survival
Commision. Str. 105-110.
Bickford D., Lohman J. D., Sodhi S. N., Ng K. L. P., Meier R., Winker K., Ingram K. K., Das
I. 2007. Cryptic species as a window on diversity and conservation. TRENDS in Ecology and
Evolution 22-3: 148-158.
Burghardt F., Knüttel H., Becker M., Fiedler K. 2000. Flavonoid wing pigments increase
attractiveness of female common blue(Polyommatus icarus) butterflies to mate-searching
males. Naturwissenschaften 87: 304-307.
Chapman D. A. 2009. Numbers of Living Species in Australia and the World, 2nd edition,
Report for the Australian Biological Resoueces Study, Canberra.
Coutsis G. J., De Prins J. 2005. A new brown Polyommatus (Agrodiaetus) from northern
Greece (Lepidoptera: Lycaenidae). Phegea 33(4): 129-137.
Dantchenko, A. V., & Lukhtanov, V. A., 1994. New taxa of the subgenus Agrodiaetus
HÜBNER, 1822 from Caucasus (Lepidoptera, Lycaenidae). Atalanta, 25, 207-213.
Dincă V., Runquist M., Nilsson M., Vila R. 2013. Dispersal, fragmentation, and isolation
shape the phylogeography of the European lineages of Polyommatus (Agrodiaetus) ripartii
(Lepidoptera: Lycaenidae). Biological Journal of the Linnean Society 109: 817- 829.
Elliot, J. N. 1973. The higher classification of the Lycaenidae (Lepidoptera): A tentative
arrangement. Bulletin of the British Museum (Natural Hististory) Entomology, 28:371-505.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
41
Floyd, R., Eyualem, A., Papert, A., Blaxter, M.L., 2002. Molecular barcodes for soil
nematode identification. Molecular Ecology, 11: 839–850.
Folmer O., Black M., Hoeh W., Lutz R., Vrijenhoek R. 1994. DNA primers for amplification
of mitochondrial cytochrome coxidase subunit I from diverse metazoan invertebrates.
Molecular Marine biology and Biotechnology.
Frézal, L., Leblois, R., 2008. Four years of DNA barcoding: Current advances and prospects.
Infection, genetics and evolution, doi: 10.1016/j.meegid.2008.05.005.
Hajibabaei M., Singer A. C. G., Hebert D. N. P., Hickey A. D. 2007. DNA barcoding: how it
complements taxonomy, molecular phylogenetics and population genetics. Trends in Genetics
23-4.
Kandul P. N., Lukhtanov A. V., Dantchenko V. A., Coleman W. S. J., Sekercioglu H. C.,
Haig D., Pierce E. N. 2004. Phylogeny of Agrodiaetus Hübner 1822 (Lepidoptera:
Lycaenidae) Inferred from mtDNA Sequences of COI and COII and Nuclear Sequences of
EF1-α: Karyotype Diversification and Species Radiation. Systematic Biology 53(2): 278-298.
Kandul P. N., Lukhtanov A. V., Pierce E. N. 2007. Karyotypic Diversity and Speciation in
Agrodiaetus Butterflyes. Evolution 61 (3): 546- 559.
Klots A. B, Klots E. B. 1970. Metulji- red Lepidoptera. V: Ţuţelke, Ilustrirana enciklpedija
ţivali. Šmit J. (ed.). Ljubljana, Mladinska knjiga: 184-258
Kolev Z. 2005. Polyommatus dantchenkoi (Lukhtanov & Wiemers, 2003) tentatively
identified as new to Europe, with a description of a new taxon from the Balkan Peninsula
(Lycaenidae). Nota lepidoptera 28 (1): 25- 34.
Kristensen N., Scoble J. M., Karsholt O. 2007. Lepidoptera phyogeny and systematics: the
state of inventorying moth and butterfly diversity. Zootaxa 1668: 699-747.
Leimar O. 1996. Life History Plasticity: Influence of Photoperiod on Growth and
Development in the Common Blue Butterfly. Oikos 76 (2): 228-234.
Lewis H. L. 1985. Butterflies of the World, Bracken Books, London.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
42
Lukhtanov A. V., Wiemers M., Meusemann K. 2003. Description of a new species of the
»brown« Agrodiaetus complex from South-East Turkey (Lycaenidae). Nota lepidoptera
26(1/2): 65-71.
Melovski D., Bozhinovsk E. 2014. New Records for Four Butterfly Species (Lepidoptera:
Papilionidea & Hesperioidea) in the Republic of Macedonia. Journal of Natural Sciences
Research 4 (7): 40-44.
Mohini J., Deshpande J. D. 2010. Polymerase chain reaction: methods, principles and
application. International Journal of Biomedical Research 1 (5): 81-97.
National center for biotechnology information. URL: http://www.ncbi.nlm.nih.gov/ (citirano
dne 12. 6. 2014).
Nielsen, E. S., and L. A. Mound. 2000. Global diversity of insects:the problems of estimating
numbers. Str. 212–222 v P. H. Raven and T. Williams, (ed.). Nature and human society: the
quest for a sustainable world. National Academy, Washington, DC.
Novotny, V., Basset, Y., Miller, S.E., Weiblen, G.D., Bremer, B., Cizek, L. and Drozd, P.
(2002). Low host specificity of herbivorous insects in a tropical forest. Nature 416: 841-844.
Palumbi, R. 1996. Nucleic acids II: The polymerase chain reaction. Molecular systematics,
Sinauer Associates, Inc., 2nd edition, uredniki: Hills, D. M., Moritz C., Mable, K. M. str. 655.
Pamperis N. L., 2009. The Butterflies of Greece. Editions Pamperis, Atene.
Preston-Mafham R., Preston-Mafham K. 1988. Butterflies of the World, Blandford Press,
London.
Przybylowicz L., Lukhtanov V., Lachowska- Cierlik D. 2013. Towards the understanding of
the origin of the Polish remote population of Polyommatus (Agrodiaetus) ripartii
(Lepidoptera: Lycaenidae) based on kariology and molecular phylogeny. Journal of
Zoological Systematics and Evolutionary Research 52 (1): 44-51.
Ronquist F., Huelsenbeck J., Teslenko M. 2011. Draft MrBayes version 3.2 Manual: Tutorials
and Model Summaries, str. 1-36.
Schmitt T., Varga Z., Seitz A. 2005. Are Polyommatus hispana and Polyommatus slovaticus
bivoltine Polyommatus coridon (Lepidoptera: Lycaenidae)? The discriminatory value of
genetics in taxonomy. Organisms, Diversity & Evolution 5: 297- 307.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
43
Štrus J., Drobne D., Zidar P., 2002. Navodila za vaje iz splošne zoologije, 2. del. Študentska
zaloţba, Ljubljana.
Talavera G., Lukhtanov A. V., Pierce E. N., Vila R. 2012. Establishing criteria for higher-
level classification using molecular data: the systematics of Polyommatus blue butterflies
(Lepidoptera, Lycaenidae). Cladistics 1-27.
Tolman T., Lewington R. 2008. Collins butterfly guide, The most complete guide to the
Butterflies of Britain and Europe. Fluke Art Velika Britanija (ed.). London, HarperCollins
Publishers: 15-120 str.
Triberti P., Daccordi M., Zanetti A. 1988. Butterflies and Moths, Mcdonald & Co (Publishers)
Ltd, Londonstr 10-30.
Trontelj P., Fišer C. 2008. Cryptic species diversity should not be trivialised. Systematics and
Biodiversity 7(1): 1-3.
van Swaay, C., Wynhoff, I., Verovnik, R., Wiemers, M., López Munguira, M., Maes, D.,
Sasic, M., Verstrael, T., Warren, M. & Settele, J. 2010a. Polyommatus admetus. The IUCN
Red List of Threatened Species. Version 2014.1. <www.iucnredlist.org>. [04. 07. 2014].
van Swaay, C., Wynhoff, I., Verovnik, R., Wiemers, M., López Munguira, M., Maes, D.,
Sasic, M., Verstrael, T., Warren, M. & Settele, J. 2010b. Polyommatus ripartii. The IUCN
Red List of Threatened Species. Version 2014.1. <www.iucnredlist.org>.[04. 07. 2014].
van Swaay, C., Wynhoff, I., Verovnik, R., Wiemers, M., López Munguira, M., Maes, D.,
Sasic, M., Verstrael, T., Warren, M. & Settele, J. 2010c. Polyommatus orphicus. The IUCN
Red List of Threatened Species. Version 2014.1. <www.iucnredlist.org>. [04. 07. 2014].
van Swaay, C., Wynhoff, I., Verovnik, R., Wiemers, M., López Munguira, M., Maes, D.,
Sasic, M., Verstrael, T., Warren, M. & Settele, J. 2010d. Polyommatus eleniae. The IUCN
Red List of Threatened Species. Version 2014.1. <www.iucnredlist.org>. [04. 07. 2014].
van Swaay, C., Wynhoff, I., Verovnik, R., Wiemers, M., López Munguira, M., Maes, D.,
Sasic, M., Verstrael, T., Warren, M. & Settele, J. 2010e. Polyommatus aroaniensis. The
IUCN Red List of Threatened Species. Version 2014.1. <www.iucnredlist.org>. [04 July 2014].

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
44
Vershinina A. O., Lukhanov V. A. 2010. Geographical distribution of the cryptic species
Agrodiaetus alcestis alcestis, A. Alcestis karacetinae and A. demavendi (Lepidoptera:
Lycaenidae) revealed by cytogenetic analysis. Comparative Cytogenetics. 4 (1), 1-11.
Vila R., Bell D. C., Macniven R., Goldman-Huertas B., Ree H. R., Marshall R., Bálint Z.,
Johnson K., Benyamini D., Pierce E. N. 2011. Phylogeny and palaeoecology of Polyommatus
blue butterflies show Beringa was a climate-regulated gateway to the New World.
Proceedings of the Royal Society str. 4-10.
Vila R., Lukhtanov A. V., Talavera G., Gil-T. F., Pierce E. N. 2010. How common are dot-
like distributions? Taxonomical oversplitting in western European Agrodiaetus (Lepidoptera:
Lycaenidae) revealed by chromosomal and molecular markers. Biological Journal of the
Linnean Society 101: 130-154.
Wiemers M. 2003. Chromosome differentiation and the radiation of the butterfly subgenus
Agrodiaetus (Lepidoptera: Lycaenidae: Polyommatus)- a molecular phylogenetic approach.
(Doktorska disertacija). Rheinischen Friedrich- Wilhelms- Universität Bonn, str. 5- 120.
Wiemers M., Stradomsky V. B., Vodolazhsky I. D. 2010. A molecular phylogeny of
Polyommatus s. str. and Plebicula based on mitochondrial COI and nuclear ITS2 sequences
(Lepidoptera: Lycaenidae). European Journal of Entomology 107: 325- 336.
Wilson, O.E., 2004. The encyclopedia of life. Trends in Ecology and. Evolution 18 (2): 77–
80.

Koritnik T. Ugotavljanje sorodstvenih odnosov v podrodu metuljev Agrodiaetus
na Balkanskem polotoku. Diplomsko delo. Ljubljana, Univerza v Ljubljani, Pedagoška
fakultera, Biologija in Kemija, 2014.
45
Zahvala
Zahvaljujem se mentorju doc. dr. Verovniku za pomoč, vodstvo in potrpljenje med pisanjem
diplomskega dela.
Zahvaljujem se tudi doc. dr. Fišerju za pregled diplome in vodstvo v končnih fazah pisanja ter
doc. dr. Zidarju za končni pregled in odobritev diplomskega dela.
Velika zahvala gre tudi Teu Deliću za pomoč in vodstvo pri laboratorijskem delu diplomskega
dela.
Zahvaljujem se tudi Anji, s katero sva skupaj raziskovala, pisala in si stala ob strani.
Za vse zabavne trenutke tekom študija se zahvaljujem še Joni in Nevi ter vsem ostalim, s
katerimi smo preţivljali ves ta čas.