
Significance of the social relationships of Efe Pygmy men in the Ituri forest, Zaire
13
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 78:495-507 (1989) Significance of the Social Relationships of Efe Pygmy Men in the lturi Forest, Zaire ROBERT C. BAILEY AND ROBERT AUNGER, JR. Department of Anthropology, University of Californiq Los Angeles, California 90024 KEY WORDS success, Hunter-gatherers, Pygmies, Africa Patrilocal residence, Social organization, Hunting ABSTRACT Efe Pygmies in northeast Zaire exhibit features of social or- ganization (small group size, flexible but predominantly patrilocal residence, close relations with kin, and pair bonds) that are common to many hunting and gathering societies. This study uses methods commonly employed in stud- ies of nonhuman primates to investigate the associations of Efe men with other individuals as a function of their age, sex, and kinship, and it tests the hypothesis that Efe are patrilocal because hunting efficiency is improved by hunting with male relatives. The analyses of 376 hours of focal behavior observations on 16 Efe men and observations of 71 cooperative group hunts show that the majority of associa- tions between Efe men were with other adult men. Men associated preferen- tially with kin over nonkin, and they associated with close kin more than with distant kin. Men’s close relationships (what we call “Companionships” sensu Smuts’ “Friendships” in baboons) were predominantly with other adult men, but each man who cohabited with a woman had his strongest Companionship by far with that woman. Neither the efficiency of hunts nor the hunting success of individual men were found to be related to the degree of relatedness of hunters nor the proportion of relatives on the hunt. Alternative hypotheses concerning the functional significance of patrilocality and male kin groups are considered. We conclude that strong affiliative bonds between male kin and between males and females may arise as much from the need for strong allies in the face of competitive social situations as from economic or ecological necessity. Investigations of the behavior and social organization of contemporary hunting and gathering peoples are used by students of human evolution to generate conceptual models of preagricultural life. Ideally, such models can be tested by examining the fossil and archaeological record (Binford, 1978, 1983; Gould, 1978; Isaac, 1968; Yellen, 1977). However, because direct evidence of social behavior is seldom preserved, paleoanthro- pologists have been forced to make infer- ences about the social constraints working in the past primarily from observations of ex- tant nonhuman primates and people (Good- all and Hamburg, 1975; Hill, 1982; Kinzey, 1987; Lovejoy, 1981; Tanner, 1981; Washburn and Lancaster, 1968). There are numerous pitfalls associated with attempts to use mod- ern-day peoples as analogs for a way of life that has long disappeared (Schrire, 1984; Freeman, 1968). Overriding these, however, is the likelihood that populations in the past were subject to the same fundamental evolu- tionary and ecological principles as contem- porary human populations. Also, it is likely that the same basic characteristics of social organization exhibited by modern hunters and gatherers were common to preagricul- tural humans. These include residential Received May 22,1987; revision accepted May 9,1988. 0 1989 ALAN R. LISS, INC
Transcript of Significance of the social relationships of Efe Pygmy men in the Ituri forest, Zaire

AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 78:495-507 (1989)
Significance of the Social Relationships of Efe Pygmy Men in the lturi Forest, Zaire
ROBERT C. BAILEY AND ROBERT AUNGER, JR. Department of Anthropology, University of Californiq Los Angeles, California 90024
KEY WORDS success, Hunter-gatherers, Pygmies, Africa
Patrilocal residence, Social organization, Hunting
ABSTRACT Efe Pygmies in northeast Zaire exhibit features of social or- ganization (small group size, flexible but predominantly patrilocal residence, close relations with kin, and pair bonds) that are common to many hunting and gathering societies. This study uses methods commonly employed in stud- ies of nonhuman primates to investigate the associations of Efe men with other individuals as a function of their age, sex, and kinship, and it tests the hypothesis that Efe are patrilocal because hunting efficiency is improved by hunting with male relatives.
The analyses of 376 hours of focal behavior observations on 16 Efe men and observations of 71 cooperative group hunts show that the majority of associa- tions between Efe men were with other adult men. Men associated preferen- tially with kin over nonkin, and they associated with close kin more than with distant kin. Men’s close relationships (what we call “Companionships” sensu Smuts’ “Friendships” in baboons) were predominantly with other adult men, but each man who cohabited with a woman had his strongest Companionship by far with that woman. Neither the efficiency of hunts nor the hunting success of individual men were found to be related to the degree of relatedness of hunters nor the proportion of relatives on the hunt. Alternative hypotheses concerning the functional significance of patrilocality and male kin groups are considered. We conclude that strong affiliative bonds between male kin and between males and females may arise as much from the need for strong allies in the face of competitive social situations as from economic or ecological necessity.
Investigations of the behavior and social organization of contemporary hunting and gathering peoples are used by students of human evolution to generate conceptual models of preagricultural life. Ideally, such models can be tested by examining the fossil and archaeological record (Binford, 1978, 1983; Gould, 1978; Isaac, 1968; Yellen, 1977). However, because direct evidence of social behavior is seldom preserved, paleoanthro- pologists have been forced to make infer- ences about the social constraints working in the past primarily from observations of ex- tant nonhuman primates and people (Good- all and Hamburg, 1975; Hill, 1982; Kinzey, 1987; Lovejoy, 1981; Tanner, 1981; Washburn
and Lancaster, 1968). There are numerous pitfalls associated with attempts to use mod- ern-day peoples as analogs for a way of life that has long disappeared (Schrire, 1984; Freeman, 1968). Overriding these, however, is the likelihood that populations in the past were subject to the same fundamental evolu- tionary and ecological principles as contem- porary human populations. Also, it is likely that the same basic characteristics of social organization exhibited by modern hunters and gatherers were common to preagricul- tural humans. These include residential
Received May 22,1987; revision accepted May 9,1988.
0 1989 ALAN R. LISS, INC

496 R.C. BAILEY AND R. AUNGER, JR.
groups that are small and variable in size; highly fluid and flexible residence; individu- als embedded in a social network of biologi- cal kin; close relations with a relatively small number of people over a lifetime; and bonds between individual men and women (pair bonds).
The social environment of Efe pygmies liv- ing in the Ituri Forest conforms to these fea- tures. Efe camps are small, averaging 17.9 inhabitants, but they range in size from just three to 55 individuals (Bailey and Peacock, 1988). Both the ideal and predominant pat- tern of residence is patrilocal (virilocal), but households change residence easily, com- monly moving to camps where biological kin are present. With the exception of special occasions (dances and funerals), when as many as 160 people congregate for a day in a Lese village, Efe are never exposed to large numbers of people. Since individuals range over a small, sparsely populated geographic area (approximately 3,000 km’), few Efe ever converse with more than 2,000 different peo- ple over their lifetime. Individual Efe men and women marry or cohabit for periods of months or years. They sleep in the same hut, often with others, and cooperate in some ac- tivities, including food gathering, food prep- aration, and child care (Bailey, 1985; Peacock, 1985).
The Efe, being predominantly patrilocal, have the same pattern of postmarital resi- dence as the majority of the world’s other hunting and gathering societies (Ember, 1978). The conditions that favor patrilocality among hunters and gatherers have long been under debate (e.g., Divale, 1974; Eggan, 1968; Ember, 1975, 1978; Ember and Ember, 1971; Lee, 1972; Service, 1962; Steward, 1955). One prominent hypothesis is that reliance on hunting favors patrilocal residence because hunting is most efficiently practiced, and the products of hunting are most efficiently shared, by men who are raised together (Ser- vice, 1962). Despite crosscultural studies in- dicating that high dependence on hunting does not correlate with patrilocality (Ember, 1975), it still seems likely that patrilocality should be favored in societies where hunting is important to subsistence, if hunting with male kin (versus nonkin) improves the effi- ciency of hunting. Among the ways male kin groups might have higher hunting efficiency is through greater familiarity with the hunted area and the distribution of animals within it (Service, 1962). Hunting could also
favor patrilocality through kin selection (Maynard-Smith, 1964), since individuals may increase their inclusive fitness by shar- ing meat with relatives on hunts.
The patrilocal postmarital residence pat- tern of Efe along with their emphasis on hunting by men and the general features of their social environment are common to most contemporary hunters and gatherers. The purpose of this report is to explain the func- tional significance of these aspects of social organization. We characterize the form and frequency of interactions between individual Efe, which result from these features of so- cial organization, and we investigate the ad- vantages to men of associating in the patterns we observe. After examining men’s associations in different settings with people of different age and sex classes, we test the hypothesis that men associate with close kin at greater frequencies than with distant kin or with nonkin. We then examine carefully the individuals with whom men associate at the highest frequencies. We call these high- frequency associations “special relation- ships” or “Companionships.” We ask the question: What are the advantages to men of associating with close male kin? We test the hypotheses that hunting groups with more closely related men are more successful and that men who hunt with close kin are able to procure more meat. We then discuss alterna- tive hypotheses concerning the functional significance of cooperative male kin (patrilo- cal) groups among Efe in light of similar studies of nonhuman primates and humans.
This study uses methods similar to those employed by field primatologists to discern social relationships. It does not rely upon informant reports but rather measures asso- ciation patterns between individuals using quantitative observational techniques de- rived from studies of primate behavioral ecol- ogy (e.g., Altmann, 1974). This makes possible direct comparisons with the results of studies of other primate species.
From the earliest days of primatology and human ethology researchers have used asso- ciation and spatial proximity as measures of affinity between individuals (e.g., Carpenter, 1963; Washburn and DeVore, 1963; Blurton- Jones, 1972). Numerous studies of nonhu- man primates and humans have shown that proximity is related consistently to other im- portant variables. In monkeys spatial prox- imity has been found to be correlated with grooming relationships, and both have been

SOCIAL RELATIONSHIPS OF EFE MEN 497
related to the frequency of alliance formation (e.g., Cheney, 1977; Dunbar, 1980), the like- lihood of responsiveness to another’s call (e.g., Seyfarth and Cheney, 1984), the frequency of aggressive encounters (e.g., Silk et al., 1981), the frequency of mounting and presenting (e.g., Kummer et al., 19781, the probability of future matings (e.g., Michael et al., 1978), and the formation of “special relationships” or “Friendships” between males and females (Smuts, 1985). Patterns of proximity in non- human primates have also been shown to be related to genealogical relatedness (Massey, 1977; Kurland, 19771, with individuals inter- acting more often with close kin within cer- tain age and sex classes than with distant kin.
In studies of humans, anthropologists have long focused on how variables such as age, sex, and particularly kinship can structure social behavior under a variety of circum- stances. However, there have been surpris- ingly few quantitative observational studies of how these variables impact on the pat- terns of association between people within societies. Hames (1979), studying the Ye’kwana of southern Venezuela, demon- strated that social interactions and proxim- ity increased proportionately with the degree of relatedness between interactants. He showed that the Ye’kwana kinship classifi- cations did not predict the frequency of inter- actions as well as estimates of biological kinship. These findings were consistent with the prediction derived from kin selection the- ory (Hamilton, 1964) that individuals should interact more frequently with close biologi- cal kin than with distant kin or nonrelatives (Alexander, 1974). Hames (1988) also showed that the amount of care allocated to Ye’kwana infants is positively correlated to the degree of relatedness between child and caretaker. Flinn (1983, 1984) quantified as- sociations between people in Grande Anse, a village on the northern coast of Trinidad and, like Hames, found that close kin associated more frequently than distant kin. Flinn also found sex and age differences in association patterns, and he used association frequencies between males and females as measures of mate guarding behavior by males. Betzig and Turke (1986) used associations of parents with children as an index of parental invest- ment (Trivers, 1972) on Ifaluk atoll in the southern Pacific. They found that husbands of high traditional and salaried status and their wives associated significantly more
TABLE 1. Characteristics of and hours of Observations on 16 Efe men
Hours of Marital Approx. ID No. age (years) status’ observations
530 531 532 533 534 538 630 631 632 633 634 635 636 650 656 670
53 M 32 M 22 M 58 M 24 M 40 M 57 M 35 M 53 S 21 M 24 S 31 S 22 M 32 S 36 S 62 S
32 32 24 16 16 16 24 24
8 32 24 32 24 24 24 24
‘M indicates married or cohabiting with an unrelated woman for more than 50% of days when observations were performed. S indicates single.
often with sons than did parents without such status. Their findings were consistent with the prediction, derived from Trivers and Wil- lard (19731, that parents in good condition will contribute greater parental investment to sons than daughters, while parents in poor condition will invest more in daughters than in sons.
MATERIALS AND METHODS
Results are based upon 376 hours of focal observations collected by the senior author between December 1980 and January 1982 on 16 Efe men who lived mostly in two sepa- rate but interacting bands that were associ- ated with the Lese villages near our research station. Details of the behavioral observation methodology can be found in Bailey (1985). Table I provides a profile of the 16 focal men and the number of hours of observation on each man. Using a stratified random sam- pling technique, a focal subject was prese- lected and followed for a 1-hour period. One- hour observation periods on each man were evenly distributed throughout the 12-hour day (from 6:OO AM to 6:OO PM) and across each of four seasons throughout a year. Dur- ing each hour of observation, an instanta- neous sample (Altmann, 1974) of the focal subject’s location and activity was recorded every minute on the minute. Some of the classes of behaviors recorded each minute were type and amount of food being eaten, location of the subject, his nearest nonheld neighbor, his posture or movement, his work activity, whether or not he was engaged in

498 R.C. BAILEY AND R. AUNGER. JR
child care, any overt social interaction, and any vocalizations.
Associations with other individuals were recorded in two ways. First, an instanta- neous scan sample (Altmann, 1974) of all in- dividuals within 10 m of the focal subject was recorded three times during each observa- tional hour: on the 1st minute, on the 30th minute, and on the last minute. A total of 1,128 such scans were recorded. For the pur- poses of analyses, every person appearing in a scan was recorded as an “associate,” and every record of an associate was called an “association.” We call the focal subject and any person who appears in the total sample, even if only once, a “dyad.” So, for example, a person could appear 100 times in the sam- ple as an associate of one focal man, but that person and the focal man would be recorded as a single dyad. Second, the nearest neigh- bor of the focal subject was recorded every minute on the minute. Nearest neighbors in- cluded infants unless they were held by the focal subject. The nearest neighbor data have been used in combination with the records of associates to define “Companionships.” This term is roughly equivalent to what Smuts (1985) refers to as “Friendships” among baboons.
A score combining association frequency with nearest neighbor frequency was devel- oped to define Companionship. This was nec- essary because two persons livlng in the same camp may appear as associates frequently, yet they may interact directly only rarely. However, if two people are nearest neighbors consistently, they are likely to have a pref- erential affinity for one another. Therefore we used the on-the-minute nearest-neighbor records to add a criterion to the association data to determine Companionship. To qualify as a Companion of a focal man, a person had to appear as an associate during a minimum of 20% of the scan samples of the focal subject and as nearest neighbor during at least 5% of the on-the-minute observations of the focal subject. These percentages were chosen by determining the number of Companions re- sulting from varying combinations of associ- ation and nearest-neighbor frequencies. At higher frequencies there was a sharp drop in the number of Companions; at lower frequen- cies there was a sharp rise in the number of Companions. Defining dyads as Companions emerged from these objective criteria, but the results were confirmed by the inclusion of nearly all those individuals that the senior
author considered, subjectively, to have close relationships with the focal subjects and the exclusion of those he did not. For compara- tive purposes, a companionship score was calculated by multiplying the percent of ob- servations for which the companion dyad was recorded as nearest neighbors times the per- cent of observations for which it was recorded as in association.
Additional information was recorded dur- ing hunts and foraging expeditions. A total of 71 group hunts were observed. Prey killed during each hunt were butchered and di- vided among the participants in the forest before return to camp. During each of 61 group hunts the following information was recorded: the number and identity of all par- ticipants; the number, weight, and shooter of all animals killed; the weight of all meat distributed; to whom and from whom all meat was distributed; and the total weight of all meat procured by each hunter-this included the meat retained and the meat received by each hunter after all kills were butchered, divided, and distributed.
The senior author conducted extensive in- terviews to determine each subject’s geneal- ogy and marital and reproductive history. Since such interviews were conducted with over 400 other Efe and 450 Lese in the same area, there was ample opportunity for cross- checking. Any discrepancies were carefully resolved. Special care was taken to deter- mine true biological genealogy. To calculate relatedness, we have assumed that our rec- ords of putative biological fathers are correct and that no inbreeding has occurred among the common ancestors. Relatedness (r) was defined as the expected fraction of identical genes shared by two individuals by virtue of common descent. Ages were estimated fol- lowing methods published by Howell (1979) and described in detail in Bailey (1985). In- dividuals were classified as infants if they were less than 3 years old, as juveniles if they were 3-17 years old, and as adults if they were 18 or more years old.
Data were summarized and analyzed using SAS (SAS Institute, 1985) on an IBM 4381 mainframe computer.
RESULTS Association patterns
The 16 Efe men had a total of 6,233 associ- ates over 1,128 observations, or an average of 5.53 associates each time they were ob- served. Approximately 1,050 different dyads

SOCIAL RELATIONSHIPS OF EFE MEN 499
d
c e Lr
d
c P
Lr
Id
TABLE 3. Frequencies and percentages of association with 16 focal men bv adeeree of relatedness frl'
Degree of relatedness (r) Frequency Percent
> 1/16 Males Females
Males Females
Males Females
Males F e m a 1 e s
1/16
118
114
2,841 45.4 1,797 28.8
153 2.4 42 .7
172 2.8 58 .9
191 3.1 65 1.1
1/2 Males 454 7.3 Females 236 3.8
Wives 224 3.6 Totals 6,233 99.9
'Based upon 1,128 instantaneous scan samples of all individuals within 10 m of each focal man. Scan samples were recorded during 376 hours of focal observations on the 16 men.
were observed over the 1-year period. Table 2 shows the proportion of all associations within each age-sex class observed with the 16 men during the 376 hours of focal obser- vations. The greatest proportion of associa- tions by far were with other adult men, who constitute 50.5% of all associates. Fewer than one-third of the associations were with adult women, and only 4.3% were with infants of either sex. While hunting and foraging in the forest, which constituted 32.8% of their time (Bailey, 1985; Bailey and Aunger, 19861, the Efe men were either alone or nearly ex- clusively in the company of other men. Only 4.2% of the associations while men were hunting and foraging were with females and infants; these occurred not during hunts, but during honey-extracting trips. While in camp, men associated with women and chil- dren much more often than when they were in other settings, although men and boys still constituted over 50% of the associations.
Association with kin The 16 Efe men associated much more fre-
quently with nonkin (r < 1/16) than with kin (r 2 1/16) (Table 3). The proportion of associ- ations with nonrelatives (77.8%) was greater than with all categories of relatives com- bined, and of 1,050 different dyads observed, only 116 (11%) were relatives of 1/16 degree of relatedness or greater.
500 R.C. BAILEY AND R. AUNGER, JR.
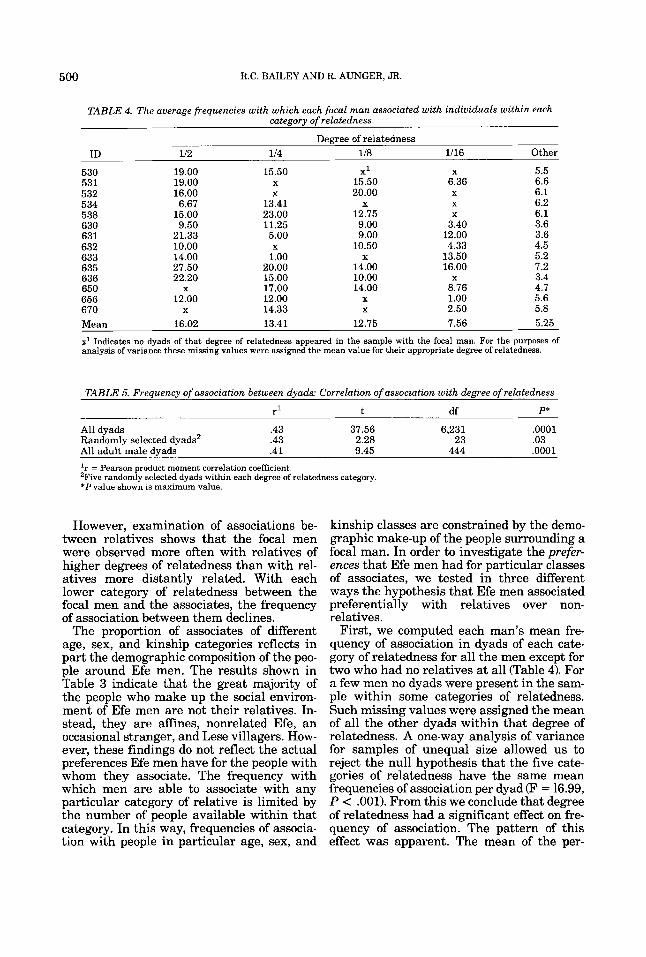
TABLE 4. The average frequencies with which each focal man associated with individuals within each category of relatedness
Degree of relatedness ID 112 114 118 1/16 Other
530 19.00 15.50 X1 X 5.5 531 19.00 X 15.50 6.36 6.6 532 16.00 X 20.00 X 6.1 534 6.67 13.41 X X 6.2 538 15.00 23.00 12.75 X 6.1 630 9.50 11.25 9.00 3.40 3.6 631 21.33 5.00 9.00 12.00 3.6 _ _ _ 632 633 635 636 650 656 670
10.00 X 14.00 1.00 27.50 20.00 22.20 15.00 X
12.00 X
17.00 12.00 14.33
10.50
14.00 10.00 14.00
X
X X
4.33 13.50 16.00
8.76 1.00 2.50
X
4.5 5.2 7.2 3.4 4.7 5.6 5.8
Mean 16.02 13.41 12.75 7.56 5.25
x1 Indicates no dyads of that degree of relatedness appeared in the sample with the focal man. For the purposes of analysis of variance these missing values were assigned the mean value for their appropriate degree of relatedness.
TABLE 5. Frequency of association between dyads: Correlation of association with degree of relatedness
rl t df P* All dyads .43 37.56 6,231 ,0001 Randomly selected dyads2 .43 2.28 23 .03 All adult male dyads .41 9.45 444 ,0001 'r = Pearson product moment correlation coefficient. 'Five randomly selected dyads within each degree of relatedness category. *P value shown is maximum value.
However, examination of associations be- tween relatives shows that the focal men were observed more often with relatives of higher degrees of relatedness than with rel- atives more distantly related. With each lower category of relatedness between the focal men and the associates, the frequency of association between them declines.
The proportion of associates of different age, sex, and kinship categories reflects in part the demographic composition of the peo- ple around Efe men. The results shown in Table 3 indicate that the great majority of the people who make up the social environ- ment of Efe men are not their relatives. In- stead, they are affines, nonrelated Efe, an occasional stranger, and Lese villagers. How- ever, these findings do not reflect the actual preferences Efe men have for the people with whom they associate. The frequency with which men are able to associate with any particular category of relative is limited by the number of people available within that category. In this way, frequencies of associa- tion with people in particular age, sex, and
kinship classes are constrained by the demo- graphic make-up of the people surrounding a focal man. In order to investigate the prefer- ences that Efe men had for particular classes of associates, we tested in three different ways the hypothesis that Efe men associated preferentially with relatives over non- relatives.
First, we computed each man's mean fre- quency of association in dyads of each cate- gory of relatedness for all the men except for two who had no relatives at all (Table 4). For a few men no dyads were present in the sam- ple within some categories of relatedness. Such missing values were assigned the mean of all the other dyads within that degree of relatedness. A one-way analysis of variance for samples of unequal size allowed us to reject the null hypothesis that the five cate- gories of relatedness have the same mean frequencies of association per dyad (F = 16.99, P < .001). From this we conclude that degree of relatedness had a significant effect on fre- quency of association. The pattern of this effect was apparent. The mean of the per-

SOCIAL RELATIONSHIPS OF EFE MEN 501
dyad associations within each category of re- latedness was higher than the mean for the next lower category, so that there was a step- wise decline in the mean frequency of asso- ciation per dyad with each degree of relatedness. Using Hochberg’s GT2-method for multiple comparison of means (Sokal and Rohlf, 1981), it was found that each mean frequency of association was significantly different from every other mean except those of 114 and 118 degrees of relatedness (m(10,67) = 1.45, P c .05, one-tailed). These results show that the focal men associated with the close relatives around them in preference to those who were more distantly related.
Next we examined the correlation between the frequency of association between all dyads and their degree of relatedness (Table 5). The relationship between degree of relat- edness and the frequency of association was highly significant (r = .43, t = 37.56, P < .0001). Since the numbers of relatives that a man had within any one category of related- ness could bias his associations with people of the same or other categories, we then ran- domly chose five out of all the possible dyads within each of the five categories of related- ness. The correlation between frequency of association and degree of relatedness for the 25 dyads was significant (r = .43; t = 2.28, P < .03).
We next investigated this relationship within one age-sex category: adult males. The findings were similar (Table 5): the focal men associated significantly more often than ex- pected with close male relatives than with distant male relatives and nonrelatives (r = .41, t = 9.45, P < .OOOl>.
Three different tests all yield similar re- sults: kinship has a significant relationship with patterns of association. The three differ- ent tests lead us to reject the null hypothesis that Efe men are associating with people randomly with respect to relatedness. Efe men associate preferentially with kin, and they prefer to associate with close kin more than with distant kin or with nonkin.
Companionship and special relationships Using our combined association and near-
est-neighbor criteria to define a Companion- ship (see “Materials and Methods”), there were 41 dyads that qualified as Companions. These constitute just 4.0% of all dyads that appear in the sample (Table 6). There was little variation in the number of Companions the males had; each man had at least one
Companion, and no man had more than four. The mean number of Companions per man was 2.56. As with associates, the majority of Companions (73%) were other men or boys. Twenty-one of the Companions were other focal men, so that each focal man appeared as a Companion of other focal men a mean of 1.3 times (range = 0-4). Just one of the com- panions was an infant, and ten (24%) of the Companions were females. Age had no effect on the number of Companions a male had. Only one (ID #670) of the five bachelor males had a woman as a Companion; he was an old widower who lived with his nephew and his nephew’s wife; it was she that qualified as his Companion. The four other unmarried males had no females as Companions.
The strongest Companionships that men had were with women, all of whom were un- related to any focal man. Ten men had com- panionships with women. No man had more than one woman as a Companion, and only one (ID #548) of the ten women who qualified as Companions appeared as a Companion of more than one man. The mean Companion- ship Score of women was significantly higher than the mean score of men’s other Compan- ions, and with just two exceptions, each fe- male Companion had a higher Companion- ship Score than any other Companion of a male. Every man but one who was defined as married (cohabiting with a woman during more than 50% of observation days) during the study (n = 10) had his wife as a Compan- ion. The one husband-wife dyad that did not appear as Companions had an ongoing dis- pute during 9 months of the study; they lived in the same camp, but did not cohabit or even talk to one another during that time.
The results show, then, that if a man had a particularly special and close relationship with any one person, it was with a woman. This is especially remarkable given the divi- sion of labor between Efe men and women, which results in men being either alone or in the nearly exclusive company of other men for approximately one-third of their time (Bailey, 1985; Bailey and Aunger, 1986).
The success of hunting with kin To test the hypothesis that group hunting
efficiency (weight of animals killed per hour) increases with the mean relatedness of hunt- ers on a hunt, it was necessary to control for hunting group size, since mean relatedness of hunters declines as hunting group size in- creases (n = 535 hunters; P < .001). Using

502 R.C. BAILEY AND R. AUNGER, JR.
TABLE 6. The companionship scores of41 individuals who qualified as companions of the focal male subjects'
ID No. of ID No. of Age-sex Nearest Companionship focal man companion class' neighbor3 Associate3 score
530 5404 AF 531 530 AM
5414 AF 532 531 AM
8604 AF 533 530 AM
5444 AF 534 532 AM
634 AM 652 AM 656 AM
538 531 AM _ ~ .
630
631
632
633
634
635
636
656 650 5484 600 632 610 6404 657 6414 630 634 630 636 6614 533 534 531 671 610 611 611
AM AM AF IM AM JM AF AM AF AM AM AM AM AF AM AM AM AM JM JM JM
8.5 10.9 13.5 11.0 7.7 5.0 11.9 6.9 7.2 25.5 8.5 7.0 5.1 7.2 23.8 6.3 7.4 10.2 11.7 6.6 8.1 18.0 9.6 20.8 5.2 14.0 5.4 6.3 12.4 5.8 8.8 11.3 6.7
25.0 34.0 32.0 39.0 29.0 25.0 31.0 20.8 29.2 35.4 41.7 37.5 37.5 41.7 50.0 20.8 22.2 23.6 27.8 20.8 20.8 34.2 25.0 34.0 20.8 26.0 25.0 26.4 36.0 24.0 37.5 54.2 30.6
212.5 370.6 432.0 429.0 223.3 125.0 368.9 143.5 210.2 902.7 354.5 262.5 191.3 300.2
1,190.0 131.0 164.3 240.7 325.3 137.3 168.5 615.6 240.0 707.2 108.2 364.0 135.0 166.3 446.4 139.2 330.0 612.5 205.0
635 AM 7.8 45.8 357.2 6434 AF 29.1 45.8 1,332.8
650 656 AM 5.3 25.0 132.5 531 AM 6.0 29.2 175.2
656 534 AM 5.2 23.6 122.7 652 AM 14.5 25.0 362.5
670 538 AM 11.7 22.2 259.7 548 AF 13.1 30.6 400.9
'Each companionship score is the product of the percent of observations that a companion appeared as nearest neighbor times the p c e n t she appeared as an associate of the focal male. AM = adult male; AF = adult female; JM = male between 4 years and 16 years; IM = infant male under 4 years. No females
between 4 years and 16 years qualified as companions. 3The values are percents of observations of the focal male. 41ndicates the wife of the focal man.
each hunt as the unit of analysis (n = 611, a multiple linear regression model of hunting efficiency on mean relatedness of hunters, number of hunters, and the interaction of the two showed that there was no significant relationship between hunting efficiency and average relatedness of hunters (df = 57; F = 5.96; P < .08). Given the low level of signifi- cance and the fact that the model accounted for only 24% of the observed variation, we conclude that mean relatedness of hunters has no significant effect on the success of the
hunt, and other factors besides relatedness and hunting group size influence hunting efficiency.
While hunting with relatives does not in- crease the total amount of meat killed per hour on a hunt, hunting with kin may still be advantageous to an individual because the few animals killed are shared unequally among the men on the hunt. While the dis- tribution of meat at the butchery site in- creases the chances of every man procuring at least some meat on each hunt (Bailey,

SOCIAL RELATIONSHIPS OF EFE MEN 503
19851, some men gain more during the shar- ing process than others (Bailey, 1985), so that some hunters procure more, and others less than they themselves kill. Hunting with male kin could be advantageous if, as the theory of kin selection (Maynard-Smith, 1964) predicts (all other things being equal), coop- eration is more likely between relatives than between nonrelatives. Thus Efe men who hunt in a group that has a higher mean re- latedness to them may receive more meat than men whose fellow hunters are less closely related to them. Similarly, if those hunters who are fortunate enough to kill an animal are sharing kills preferentially with relatives, hunting with a higher proportion of relatives (r 2 1/16) versus nonrelatives (r < 1/16) should be advantageous. A higher proportion of relatives present on the hunt would mean a greater chance of receiving at least some meat.
Using each record of a hunter on a hunt as the unit of analysis (n = 535), we tested the hypothesis that the amount of meat each hunter brought home per hour of hunting (procurement success) depended upon the mean relatedness of his fellow hunters to him. We used a multiple linear regression model with procurement success per hour as the dependent variable and the following as independent variables: mean relatedness of fellow hunters, hunting group sue, the total meat killed per hour, and the interaction of the last two variables. There was no signifi- cant relationship between a man’s procure- ment success and the mean relatedness of his fellow hunters (df = 530; F = 68.8; P > .89). Substituting the proportion of hunters re- lated to the hunter for mean relatedness of cohunters produced exactly equivalent re- sults. When we added the proportion of hunt- ers related to the focal man and the interaction of this variable with mean relat- edness of fellow hunters, the generalized lin- ear hypothesis test (Mifi and Clark, 1984:172) showed that the additional variables did not improve the regression equation (F = .001; df = 2,528; P > 9). Scatterplots of the resid- uals on both mean relatedness and propor- tion of relatives were linear and constant over all values. From this we conclude that there is no relationship between the amount of meat a man brings home from a hunt and either the mean relatedness of the men he hunts with or the proportion of his fellow hunters who are related to him.
DISCUSSION
This study shows that Efe men have strong affinities for one another. Men associate with other males more frequently than any other age-sex class. Not only do they associate al- most exclusively with other males while they are hunting and foraging in the forest, they have a predominance of male associates at all other times as well. Furthermore, the great majority of close relationships or Com- panionships that Efe men have are with other men.
Among both their associates and their Com- panions, Efe men show clear preference for kin over nonkin. These findings are consis- tent with their expressed preference for pa- trilocal residence, which we should expect to result in males frequently associating with close male kin.
However, the closest relationships that men have-those with the highest frequencies of association and closest proximity-are not with other men, but are reserved for women. Miliative bonds between husbands and wives appear to be very strong. Married men are found in close proximity to their wives more than with any other individual, and most men do not have Companionships with any woman other than their wife (for the one exception see the “Results” section above). Similarly, women are not Companions of any man if they are not married or cohabiting with him. Husbands spend more time in close proximity with their wives than with anyone else, despite spending approximately one- third of their day engaged in subsistence ac- tivities that nearly exclude female participation.
The picture that emerges of Efe social or- ganization based on the observed association patterns of these 16 men is of males forming cooperative kin groups whereby men live, hunt, and associate preferentially with men who are relatives. Within these cooperative male kin groups, men have close affXiative relationships with particular females who join or are recruited from other groups. This picture is consistent with patrilocal resi- dence and exogamous marriage patterns.
Our data are not consistent with the hy- pothesis that Efe are patrilocal because hunt- ing with kin increases hunting success. We have found no advantage to men of cooperat- ing with their kin while hunting. Hunts are no more efficient if hunters are highly re-

504 R.C. BAILEY AND R. AUNGER, JR.
lated to one another than if they are dis- tantly related or unrelated to one another. Those that hunted with a higher proportion of kinsmen did not procure any more meat than other men. These results strongly indi- cate that factors other than increased hunt- ing, eMiciency are favoring patrilocal residence by Efe men. Thus, they are incon- sistent with earlier views of hunting as the driving force for patrilocality among hunters and gatherers (Steward, 1955; Service, 1962) but consistent with a more recent crosscul- tural study finding that greater dependence on hunting does not predict patrilocality (Ember, 1975).
For what reasons other than the procure- ment of meat might Efe men form close ties with their kinsmen? During the rest of this discussion we consider several alternative hypotheses. Underlying each hypothesis is the assumption that related males have greater coincidence of interest than males chosen from the population at random by virtue of their greater genetic relatedness.
One plausible hypothesis is that males form alliances with related males in order to de- fend areas against other cooperative kin groups. Efe are not strictly territorial, but each patriclan is identified with and has ac- knowledged rights to specific areas of forest. Neighboring clans may forage in a patri- clan’s acknowledged home range, but they generally do not do so without recognition from resident clan members. If a resource is disputed (a beehive, for example), the intrud- ers generally defer to the residents.
Perhaps more important than areas of for- est are the trading rights of specific Efe clans with specific Lese villages. Since approxi- mately 65% of their calories come from vil- lage gardens (Bailey and Peacock, 1988) and most of the material wealth of Efe comes from their Lese villager trading partners (Bailey, 1985), rights of access to a village with its gardens and purveyors of material wealth are important sources of resources for the members of an Efe patriclan. Further- more, since it is women who rely most on the villages as sources of the food they garner, the patriclans that can maintain close and exclusive ties with productive villages are more attractive to Efe women. The ability of men to persuade women to take up residence with them within the patriclan home range depends in part upon the reliability of the food and material resources available to them. By maintaining constant access to a
Lese village, cooperative groups of related males can assure females of access to valu- able resources.
Despite the importance of access to the Lese villages, direct confrontations between Efe patriclans over trading and working rights with specific villages are infrequent, proba- bly because the Efe compete with one an- other indirectly through villagers. However, disputes between patriclans over rights to villages do occur, and one patriclan may be displaced from association with a village by another. Two such conflicts between patri- clans were observed by the senior author in 1981, and informants told of other incidents occurring in the past.
Another possible reason why Efe males form cooperative kin groups is to recruit fe- males more effectively and to defend females and young from harassment or capture by other males (Irons, 1981). This hypothesis is compelling given the constant high level of disputes between patriclans over women and children. Also, because of a pattern of hyper- gyny whereby Efe women many villagers but Efe men never marry villager women, approximately 14% of Efe women are lost as potential marriage partners. This increases competition among Efe men for access to women (Bailey, 1988).
The prescribed marriage system entailing “sister exchange” between clans makes it advantageous to an individual Efe man to remain in his natal group with his male and unmarried female relatives in order to im- prove his chances of recruiting a wife through exchange. Females are exchanged between patriclans, but such exchanges are not al- ways immediately reciprocated (often be- cause a woman from one clan wishes to move to the clan of her lover, but her lover’s clan either refuses or is unable to give up a woman in return), so that inequalities between patri- clans are common. This results in disputes so numerous that every clan is involved at all times in at least one, but usually several, ongoing arguments over women and their children.
Armed conflict between patriclans over women is not uncommon. Two armed inter- clan confrontations involving five or more men on each side were observed during the study, one resulting in the recapture of a clan woman, and the other in the failure to cap- ture the wife of a clan member. Two other such conflicts were recorded but not observed during the study, and numerous others were

SOCIAL RELATIONSHIPS OF EFE MEN 505
reported to have occurred in the recent past. To this day, women are strongly discouraged from and obviously fearful of traveling alone, and men often escort women between camps or villages with bow and arrows in hand to protect them from other men.
Prescriptions also exist for children’s mem- berships to patriclans, and serious conflicts arise between patriclans over possession of children. Often elaborate and costly negotia- tions take place between patriclans for rights over children. Although serious child injury or infanticide have never been observed, mild physical abuse was observed several times during intense arguments over children, with the child tugged and pulled by one side and the other, and infanticide was threatened. In all the observed disputes over women and children, alliances among male relatives against members of other patriclans were ap- parent, and often individual males sought and were given prompt support from fellow patriclan members. Similarly, when wives or female relatives were threatened with attack or kidnapping (or seduction) by males outside the clan, patriclan members quickly rallied to retrieve or protect them.
Most authors considering the emergence of unique human social relationships place em- phasis on the division of labor and the con- sequent need for economic exchange between males and females (Galdikas and Teleki, 1981; Tanner, 1981; Lovejoy, 1981; Washburn and Lancaster, 1968; Hill, 1982). Most scena- rios of human evolution depict early homi- nids with relatively undifferentiated social relationships until food sharing and the di- vision of labor brought males and females together into practical economic arrange- ments that benefited both sexes. Yet recent studies of nonhuman primates show that highly differentiated, long-term relation- ships between males and females exist in the absence of any economic exchange (de Waal, 1982; Smuts, 1985; Wrangham, 1986). It seems likely that hominid males were able to offer important social benefits to females and their young long before they were devot- ing much time to hunting. As Smuts and others have so convincingly shown, the non- human primate data strongly suggest “that protection from aggression by other homi- nids, and especially by other males, was probably among the most important benefits that (hominid males) could provide (fe- males)” (Smuts, 1985260). The strong bonds between males and females that have been
detected in many species of monkey as well as the bonds observed between members of the same sex in monkeys and apes (Wran- gham, 1975,1980,1986) exist in the absence of significant food exchange or exchange of material goods.
While the economic benefits of cooperation among Efe cannot be ignored (e.g., Bailey, 1985), the patrilocal residence and male as- sociation patterns showing strong affiliative bonds between male kin and between partic- ular males and particular females may arise as much from the need for strong allies in the face of a complex web of competitive so- cial situations as from any economic or eco- logical necessity.
The results of this specific study examining the significance of Efe association patterns should never be used to make straightfor- ward analogies concerning the form and fre- quency of social interactions among hunters and gatherers in the past. However, our find- ings suggest that strictly economic and eco- logical approaches to the study of the evolution of human social behavior ignore the significance of relationships based upon the reciprocal exchange of social benefits. Greater emphasis on social relationships by field researchers seeking to apply studies of contemporary hunting and gathering peo- ples to inquiries into human evolution may eventually stimulate paleoanthropologists and others to make better-informed infer- ences about the distant past.
ACKNOWLEDGMENTS
This work was supported by grants to RCB from The National Geographic Society, The L.S.B. Leakey Foundation, and the Academic Senate of UCLA, as well as grant BNS-79- 26640 to h e n DeVore. We are grateful to Dr. Kabamba Nkamany and his staff at CE- PLANUT in Kinshasa, Zaire, for their gen- erous assistance. We thank Jeff Gornbein for his expert guidance with the analyses and the following for their helpful comments: Richard Wrangham, Monique Borgerhoff Mulder, Lynn Fairbanks, Raymond Hames, Marc Hauser, Nadine Peacock, Ross Sackett, Joan Silk, Barbara Smuts, and two anony- mous reviewers. Special thanks go to the 16 Efe men and their families for allowing us into their lives.
LITERATURE CITED
Afifi AA, and Clark V (1984) Computer-Aided Multivar- iate Analysis. Belmont, CA: Wadsworth, Inc.

Alexander RD (1974) The evolution of social behavior. Annu. Rev. Ecol. Systematics 5325-383.
Altmann J (1974) Observational study of behaviour: Sampling methods. Behaviour 48:141.
Bailey RC (1985) The Socioecology of Efe Pygmy Men in the Ituri Forest, Zaire. Ph.D. dissertation, Harvard University.
Bailey RC (1988) The significance of hypergyny for un- derstanding subsistence behavior among contempo- rary hunters and gatherers. In BV Kennedy and GM LeMoine (eds.): Diet and Subsistence: Current Archae- ological Perspectives. Calgary: University of Calgary, pp. 57-65.
Bailey RC, and Aunger R (1986) Association patterns among Efe men. Paper presented at the Fifty-Fifth Annual Meeting of the American Association of Phys- ical Anthropologists, Albuquerque, New Mexico.
Bailey RC, and Peacock NR (1988) Efe Pygmies of north- east Zaire: Subsistence strategies in the Ituri Forest. In I de Garine and GA Harrisson (eds.): Coping With
Uncertainty in Food Supply. Oxford Oxford University Press, pp. 88-117.
Betzig LL, and Turke PW (1986) Parental investment by sex on Ifaluk. Ethol. Sociobiol. 7:29-37.
Binford LR (1978) Dimensional analysis of behavior and site structure: Learning from an Eskimo hunting stand. Am. Antiquity 43:330-361.
Binford LR (1983) In Pursuit of the Past: Decoding the Archaeological Record. New York: Thames and Hudson.
Blurton-Jones NG (1972) Ethnological Studies of Child Behaviour. London: Cambridge University Press.
Carpenter CR (1963) Societies of monkeys and apes. In CH Southwick (ed.): Primate Social Behavior. New York: Van Nostrand Reinhold, pp. 24-51.
Cheney DL (1977) The acquisition of rank and the devel- opment of reciprocal alliances among free-ranging im- mature baboons. Behav. Ecol. Sociobiol. 2:303-318.
de Waal FBM (1982) Chimpanzee Politics: Power and Sex Among Apes. New York Harper and Row.
Divale WT (1974) Migration, external warfare, and ma- trilocal residence. Behav. Sci. Res. 9:75-133.
Dunbar RIM (1980) Determinants and evolutionary con- sequences of dominance among female gelada baboons. Behav. Ecol. Sociobiol. 7:253-265.
Eggan F (1968) Comments on “typology and reconstruc- tion.” In RB Lee and I DeVore (eds.): Man the Hunter. Chicago: Aldine, pp. 161-162.
Ember CR (1975) Residential variation among hunter- gatherers. Behav. Sci. Res. 10:199-227.
Ember CR (1978) Myths about hunter-gatherers. Ethnol- ogy 17:439-448.
Ember M, and Ember CR (1971) The conditions favoring matrilocal and patrilocal residence. Am. Anthropol. 73571-594.
Flinn MV (1983) Resources, Mating, and Kinship: The Behavioural Ecology of a Trinidadian Village. Ph.D. Thesis, Northwestern University.
Flinn MV (1984) Mate guarding in a Trinidadian village. Paper presented at the Eighty-Second Annual Meeting of the American Anthropological Association, Denver.
Freeman LG (1968) A theoretical framework for inter- preting archaeological materials. In RB Lee and I DeVore (eds.): Man the Hunter. Chicago: Aldine, pp.
Galdikas BMF, and Teleki G (1981) Variations in subsis- tence activities of female and male pongids: New per- wectives on the oridns of hominid labor division. Curr.
262-267.
Goodall J, and Hamburg DA (1975) Chimpanzee behav- ior as a model for the behavior of early man: New evidence on possible origins of human behavior. In DA Hamburg and HKH Brodie (eds.): American Handbook of Psychiatry: New Psychiatric Frontiers, Vol. 6. New York: Basic Books, pp. 14-43.
Gould R (1978) Explorations in Ethnoarchaeology. Al- berquerque: University of New Mexico Press.
Hames RB (1979) Relatedness and interaction among the Ye’kwana: A preliminary analysis. In NA Chagnon and W Irons (eds.): Evolutionary Biology and Human Behavior: An Anthropological Perspective. North Sci- tuate, M A Duxbury Press, pp. 239-249.
Hames RB (1988) The allocation of parental care among the Ye’kwana. In L Betzig, M Borgerhoff Mulder, and P
Turke (eds.): Human Reproductive Behavior: A Dar- winian Perspective. Cambridge: Cambridge Univer- sity Press, pp. 237-251.
Hamilton WD (1964) The genetical evolution of social behavior (Parts I and 10. J. Theor. Biol. 7:l-52.
Hill K (1982) Hunting and human evolution. J. Hum. Evol. 11:521-544.
Howell N (1979) Demography of the Dobe !Kung. New York Academic Press.
Irons W (1981) Why lineage exogamy? In RD Alexander and DW Tinkle (eds.): Natural Selection and Social Behavior: Recent Research and New Theory. New York: Chiron Press, pp. 476-489.
Isaac GL (1968) Traces of Pleistocene hunters: An east African example. In RB Lee and I DeVore (eds.): Man the Hunter. Chicago: Aldine, pp. 253-261.
Kinzey WG (edJ(1987) The Evolution of Human Behav- ior: Primate Models. Albany: Suny Press.
Kummer H, Abegglen JJ , Bachmann C , Falett J, and Sigg H (1978) Grooming relationships and object com- petition among hamadryas baboons. In DJ Chivers and J Herbert (eds.): Recent Advances in Primatology, Volume 1: Behavior. London: Academic Press, pp. 31- 38.
Kurland JA (1977) Kin selection in the Japanese mon- key. Contrib. Primatol. 12:l-145.
Lee RB (1972) Work effort, group structure, and land use in contemporary hunter-gatherers. In P J Ucko, R Tringham, and GW Dimbleby (eds.): Man, Settlement, and Urbanism. London: Duckworth, pp. 177-185.
Lovejoy CO (1981) The origin of man. Science 211:341- 350.
Massey A (1977) Agonistic aids and kinship in a group of pigtail macaques. Behav. Ecol. Sociobiol. 2:31-40.
Maynard-Smith J (1964) Group selection and kin selec- tion. Nature 20(4924):1145-1147.
Michael RP, Bonsall RW, and Zurnpe D (1978) Consort bonding and operant behavior by female rhesus mon- keys. J. Comp. Physiol. Psychol. 92:837-845.
Peacock NR (1985) Time Allocation, Work and Fertility Among Efe Pygmy Women in the Ituri Forest of North- east Zaire. Ph.D. dissertation, Harvard University.
SAS Institute Inc. (1985) SAS User’s Guide: Statistics, Version 5 Edition. Cary, NC: SAS Institute Inc.
Service ER (1962) Primitive Social Organization: An Ev-
Seyfarth RM, and Cheney DL (1984) Grooming alliances olutionary Perspective. New York: Random House.
and reciprocal altruism in vervet monkeys. Nature 308:541-543.
Schrire C (1984) Wild surmises on savage thoughts. In C Schrire (ed.): Past and Present in Hunter Gatherer Studies. Orlando: Academic Press, pp. 1-25.
Silk JB, Samuels A. and Rodman PS (1981) The influ-
506 R.C. BAILEY AND R. AUNGER, JR.
Anthropol. 22:241-256. ence of kinship, rank, and sex on affiliation and

SOCIAL RELATIONSHIPS OF EFE MEN 507
aggression between adult females and immature bon- net macaques (Macaca radiata). Behaviour 78:lll-137.
Smuts BB (1985) Sex and Friendship in Baboons. New York Aldine.
Sokal FJ, and Rohlf FJ (1981) Biometry: The Principle and Practice of Statistics in Biological Research. New York W.H. Freeman.
Steward JH (1955) Theory of Culture Change. Urbana: University of Illinois Press.
Tanner NM (1981) On Becoming Human. Cambridge: Cambridge University Press.
Trivers RL (1972) Parental investment and sexual selec- tion. In BH Campbell (ed.): Sexual Selection and the Descent of Man, 1871-1971. Chicago: Aldine, pp. 136- 179.
Trivi.rs RL, and Willard DE (1973) Natural selection of parental ability to vary the sex ratio of offspring. Sci- ence 179:90-92.
Washburn SL, and DeVore I (1963) The social life of
baboons. In CH Southwick (ed.): Primate Social Behav- ior. New York: Van Nostrand Reinhold, pp. 98-113.
Washburn SL, and Lancaster CS (1968) The evolution of hunting. In RB Lee and I DeVore (eds.): Man the Hunter. Chicago: Aldine, pp. 293-303.
Wrangham RW (1975) Behavioral Ecology of Chimpan- zees in Gombe National Park, Tanzania. Ph.D. disser- tation, University of Cambridge.
Wrangham RW (1980) An ecological model of female- bonded primate groups. Behaviour 75:262-300.
Wrangham RW (1986) Ecology and social relationships in two species of chimpanzee. In DI Rubenstein and RW Wrangham (eds.): Ecological Aspects of Social Evo- lution: Birds and Mammals. Princeton: Princeton Uni- versity Press, pp. 352-378.
Yellen JE (1977) Archaeological Approaches to the Pres- ent: Models for Reconstructing the Past. New York: Academic Press.




















