
Tagungsband - 8. BOKU-Symposium TIERERNÄHRUNG
346
Tagungsband 8. BOKU-Symposium TIERERNÄHRUNG Mineralstoffe in der Tierernährung Leistung – Gesundheit – Umweltschutz 01. Oktober 2009, Wien Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Department für Lebensmittel- wissenschaften und -technologie BOKU – University of Natural Resources and Applied Life Sciences, Vienna Universität für Bodenkultur Wien
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Tagungsband - 8. BOKU-Symposium TIERERNÄHRUNG

Tagungsband
8. BOKU-Symposium TIERERNÄHRUNG
Mineralstoffe in der Tierernährung
Leistung – Gesundheit – Umweltschutz
01. Oktober 2009, Wien
Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie
Department für Lebensmittel- wissenschaften und -technologie
BOKU – University of Natural Resources and Applied Life Sciences, Vienna Universität für Bodenkultur Wien

Seite II 8. BOKU-Symposium Tierernährung 2009
Tagungsband:
8. BOKU-Symposium TIERERNÄHRUNG
Mineralstoffe in der Tierernährung
Leistung – Gesundheit – Umweltschutz
01. Oktober 2009, Wien
Herausgeber:
Mag. Christiane Mair Margit Kraft Univ.-Prof. Dr. Wilhelm Matthias Windisch
Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie
Department für Lebensmittelwissenschaften und -technologie
Universität für Bodenkultur Wien
Gregor Mendel-Straße 33, A-1180 Wien
www.dlwt.boku.ac.at/tte.html
Eigenverlag:
Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie
Department für Lebensmittelwissenschaften und -technologie der BOKU Wien
ISBN 978-3-900962-81-4
Für den Inhalt der Beiträge sind allein die Autoren verantwortlich

Inhaltsverzeichnis
8. BOKU-Symposium Tierernährung 2009 Seite III
Inhaltsverzeichnis
Übersichtsvorträge
Erben, R.G. Knochenstoffwechsel und Mengenelemente 1
Rodehutscord, M., Haese E. Wechselwirkungen zwischen der Phosphorversorgung und der mikrobiellen Gemeinschaft des Verdauungstraktes 5
Martens, H., Stumpff, F. Magnesium Stoffwechsel: Regulation und Risiken 11
Kienzle, E. Kationen-Anionen-Bilanz beim Schwein – Möglichkeiten, Risiken und Nebenwirkungen 24
Flachowsky, G., Franke, K. Röttger, A., Meyer, U. Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft 28
Windisch, W., Kickinger, T., Würzner, H. Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung? 37
Würzner, H. (Ent)warnung für Schwermetalle in Futtermitteln 45
Poster: Mineralstoffe und Spurenelemente / Minerals and trace elements
Baulez, M. Improved selenium content in high value pork product 51
Boulianne, M., Roch, G., Baulez, M. Effect of organic selenium on growth performances, feathering, skin strength and cellulitis prevalence of challenged broiler chickens 54
Bourdonnais, A. Intestinal absorption of zinc from zinc sulfate and zinc amino acid chelates in growing pigs 57
Chrenková, M., Čerešňáková, Z., Fľak, P., Poláčiková, M. The effect of heat treatment on phosphorus releasing from soya bean and pea 62
Doležal, P., Skládanka J., Poštulka R., Zeman, L. The Effect of Stage Maturity of Alfalfa (Medicago sativa L.) on the Content of Major Minerals 66
Fry, R.S., Spears, J.W., Schlegel, P., Durosoy, S. Effects of dietary zinc source and level on immune responses and health of cattle 70
Hammerl, S., Windisch, W., Li, Q., Schedle, K. Experimentelle Untersuchungen einer hohen Jodversorgung auf die zootechnischen Leistungen sowie die Schlachtkörperqualität bei Mastschweinen 76
Kozłowski, K., Jankowski, J., Jeroch, H. Efficacy of different phytase preparations in broiler rations 82
Kulpys, J., Stankevičius, R., Urbonas, R. Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen 86

Inhaltsverzeichnis
Seite IV 8. BOKU-Symposium Tierernährung 2009
Nitrayová, S., Brestenský, M., Patráš, P., Brož, J., Heger, J. Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs 94
Rapp, C., Flocke, F., Gövert, J., Heseker, A., Hopp, J. Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen 102
Sager, M., Laguna Paredes, C. Selected nutrient- and trace element contents in hens´eggs – Nähr- und Spurenelemente in Hühnereiern 110
Sager, M., Werteker, M. Distribution of Elements in Flour Fractions – Elementverteilung in Mahlfraktionen von Weizen 114
Schlegel, P., der Kinderen, L., Mul, A., Ubbink-Blanksma, M., Durosoy, S., Bruininx, E. Effects of supplemental trace mineral (Fe, Cu, Mn, Zn) source and level on post-weaning piglet trace mineral excretion and status 116
Seresinhe, T., Manawadu, A., Iben, C. Effect of concentrate–mineral supplements on the overall nutritional status of cross bred milking cows in two small farms in Sri Lanka 121
Vacchina, V., Oguey, S., Lobinski, R., Bravo, D. Determination of Zn-glycinate complexes in diluted media 129
Večerek, M., Jancikova, P., Zeman, L., Mareš, P. Influence of Addition of Organic Manganese on its Digestibility in Sport Horses 137
Wetscherek, W., Wetscherek-Seipelt, G. Zweck und Durchführung einer Toleranzstudie bei tragenden und säugenden Zuchtsauen am Beispiel einer Phytase 142
Poster: Leistungsfördernde Futtermittelzusatzstoffe / Growth promoting feed additives
Bravo, D., Utterback, P., Parsons, C.M., Vikari, A. Evaluation of a mixture of carvacrol, cinnamaldehyde and capsicum oleoresin for improving growth performance and metabolizable energy for broiler chicks fed corn-soybean meal 148
Wendler, K.R., Mader, A., Asamer, A. Influence of precaecal apparent digestibility in young broilers by a phytogenic feed additive 153
Cypriano, L., Piccini, I., Filho, J.B.P., Pastore, N.S., Mader, A., Asamer, A. Performance and tibia characteristics of broilers fed a phytogenic feed additive 157
Gärtner, S., Zentek, J. Untersuchungen zur Wirkung eines pflanzlichen Futterzusatzes auf einen enteropathogenen Escherichia coli- Stamm 160
Kang, C.W., Ahn, B.K., Kim, J.S., Tschirner, K. Performance parameters and meat quality of broiler fed diets supplemented with a mixture of Quaternary Benzophenanthrinde and Protopine alkaloids (Sangrovit®) 165
Mareš, P., Zeman, L., Večerek, M. Spice Industry By-products in Pig Nutrition 171
Reisinger, N., Steiner, T., Nitsch, S., Schatzmayr, G., Applegate, T.J. Effects of essential oils on broiler performance and intestinal physiology during a coccidial vaccine challenge 174

Inhaltsverzeichnis
8. BOKU-Symposium Tierernährung 2009 Seite V
Steiner, T., Hong, J.-C., Lien, T.-F. Effects of Phytogenics on intestinal parameters and growth performance of broilers 179
Jerešiūnas, A., Kulpys, J., Sudikas, G., Jerešiūnienė, O. Comparable assessment of the alternative zinc oxide growth promoters, organic acids and probiotics in the rations of weaned piglets 184
Roth, N., Germ, T., Klimitsch, A., Urbaityte, R., Nitsch, S. Antibacterial activity of formic and propionic acids against Salmonella and E. coli and their influence on broiler performance 188
Wetscherek, W., Rohrer, E., Wetscherek-Seipelt, G. Einsatz von organischen Säuren in der Ferkelaufzucht 194
Urbaityte, R., Padoan, D., Roth, N. Effect of urinary acidifier on reproduction performance in sows 203
Matusevicius, P., Asmenskaite, L. Zum Einfluss von Zichoriewurzelmehl auf Leistungsparameter von Mastkaninchen 208
Liu, J., Trautwein, J., Pietsch, M., Dusel, G. Einfluss von Lignocellulose in der Broilerfütterung 212
Kroismayr, A., Braunsberger, F., Neufeld, K. Lignocellulose als Nahrungsfaserquelle in der Zuchtsauenfütterung 219
Wetscherek-Seipelt, G., Wetscherek, W. Einsatz von Weizenkleie bzw. Trockenschnitzel bei Ferkel 226
Maevis, G., Koch, C., Romberg, F.-J., Landfried, K., Kühl, J., Steingaß, H., Südekum, K.-H. Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen 232
Vielhaber, B., Zitterl-Eglseer, K., Gallnböck, M., Tichy, A., Hagmüller, W. Einsatz von Oligogalakturoniden zur Reduzierung des Absetzdurchfalles 239
Wehr, U., von Rosenberg, S. Lanthan-Verbindungen wirken einem erhöhten Knochenverlust im Osteoporosemodell der ovariektomierten Ratte entgegen 246
Poster: Weitere Themen zur Tierernährung / Further aspets in animal nutrition
Kluge, H., Bartelt, J., Stangl, G. Eine Valinsupplementierung ergibt neue Möglichkeiten bei der Proteinreduzierung im Ferkelfutter 250
Rutzmoser, K., Ettle, T., Obermaier, A. Vorschlag eines Systems von nutzbaren Aminosäuren bei Mastbullen 255
Ettle, T., Rutzmoser, K., Obermaier, A. Anwendung des Systems von nutzbaren Aminosäuren bei einem Fütterungsversuch mit Mastbullen 259
Hošková, Š., Vašátková, A., Balabánová, M., Hošek, M., Zeman, L. The effect of feeding DDGS on broiler meat composition 264
Brunauer, C., Mair, C., Schreiner, M., Windisch, W. Qualität des intramuskulären Fettes von Grauviehalmochsen im Vergleich zum durch- schnittlichen österreichischen Rindfleisch mit besonderem Augenmerk auf essentielle n6- und n3-Fettsäuren 268

Inhaltsverzeichnis
Seite VI 8. BOKU-Symposium Tierernährung 2009
Wetscherek-Seipelt, G., Stückler, R., Mandl, J., Wetscherek, W. Stichprobenuntersuchung von österreichischen Schweinefleischproben zum Fettgehalt und zur Fettzusammensetzung 274
Wetscherek, W., Kocsis, L., Liebhard, P. Nährstoffgehalt verschiedener getrockneter Topinamburkrautsorten 281
Mayrhofer, S., Domig, K.J., van Hoek, A.H.A.M., Aigner, S., Kneifel, W. Evaluierung von Laktobazillen-Stämmen bezüglich erworbener Antibiotikaresistenzgene 287
Petersson, A., Domig, K.J., Dombazyan, V., Nagel, P., Zollitsch, W., Hagmüller, W., Kneifel, W. Einsatz der Denaturierenden Gradientengelelektrophorese (DGGE) zur Untersuchung der Darmmikrobiota des Schweins 292
Müller, A., Kern, C., Sattler, V.A., Wegl, G., Schatzmayr, G., Klose, V. Gut Microbiota – Studying bacterial DGGE profiles 297
Wegl, G., Applegate, T.J., Schatzmayr, G., Klose, V. Evaluation and selection of a stable endogenous reference gene for studying immunological host response with quantitative reverse transcription PCR (qRT- PCR) in coccidiosis challenged chickens 303
Balabánová, M., Hošková, Š., Vašátková, A., Večerek, M., Zeman, L. Effect of sample conservation on blood parameters analyses 308
Finck, E., Dusel, G., Klaßen, M., Landfried, K. Untersuchung der Energie- und Nährstoffbilanzen in der Vorbereitungs- und Säugephase von Sauen 314
Novák, L., Zeman, L., Mares, P. Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period 321
Trautwein, J., Landfried, K., Hlawitschka, B., Dusel, G. Einsatz einer Trocken-TMR in der Mastrinderfütterung 328
Autorenverzeichnis 335
Sponsoren 338

Erben: Knochenstoffwechsel und Mengenelemente
8. BOKU-Symposium Tierernährung 2009 Seite 1
Knochenstoffwechsel und Mengenelemente
Reinhold G. Erben
Institut für Physiologie, Pathophysiologie und Medizinische Physik, Vetmeduni Vienna
Einleitung Die wesentlichen Funktionen von Knochengewebe bestehen 1) in einer mechanischen Stützfunktion für die Muskelaktivität, wie z.B. für die Fortbewegung notwendig, 2) in einer Schutzfunktion für innere Organe oder Weichgewebe, wie z.B. das Gehirn, und 3) in einer Funktion als Speicherorgan für den Mineralhaushalt. 99% des gesamten Körperbestandes an Calcium befinden sich im Knochen und Kno-chen ist das einzige Speicherorgan für Calcium. Da die Mineralkomponente des Knochens, die ca. 2 Drittel seiner Trockenmasse ausmacht, aus Hydroxylapatit [Ca10(PO4)6(OH)2] besteht, sind Calcium und Phosphor die für den Knochenaufbau wesentlichsten Mengenelemente. Daneben enthält Knochen auch in geringen Mengen noch Magnesium. Es hat sich jedoch in den letzten Jahren zunehmend ge-zeigt, dass die anderen Mengenelemente, also Natrium, Kalium, Chlor und Schwefel, über die Beeinf-lussung der Kationen/Anionen-Bilanz und damit des Säure-Basenhaushalts ebenfalls eine wichtige Rolle im Knochenstoffwechsel spielen. Deshalb sind Knochenstoffwechsel und Mineralhomöostase untrennbar verbunden.
Genetische und Umwelteinflüsse auf den Knochenstoffwechsel Studien bei Mäusen und am Menschen haben gezeigt, dass ca. 60 – 80% der Variabilität in der Kno-chenmineraldichte (bone mineral density, BMD) durch genetische Einflussfaktoren erklärt werden kön-nen (Beamer et al 1999, Beamer et al 2001, Eisman 1999). Die genetische Prädisposition spielt des-halb eine sehr große Rolle für die Entwicklung der Skelettmasse. Obwohl bei weitem noch nicht alle Gene, die an der Ausbildung der Skelettmasse und am Knochenstoffwechsel beteiligt sind, einschließ-lich ihrer Interaktion charakterisiert sind, sind in den letzten Jahren doch erhebliche Fortschritte in der Identifikation der stärksten Einflussfaktoren gemacht worden. Genomweite Assoziationsstudien beim Menschen haben gezeigt, dass die genetische Variabilität im Bereich des Osteoprotegerin (OPG)-Gens und des LRP5 (lipoprotein-receptor-related protein)-Gens erheblichen Einfluss auf die BMD und das Frakturrisiko haben (Richards et al 2008). Beide Gene sind Teil von größeren Zytokin- bzw. Signalt-ransduktions-Netzwerken, die für den Knochen von sehr großer Bedeutung sind. OPG ist Teil des RANK (Receptor activator of nuclear factor-κB)/RANKL (RANK ligand)/OPG-Systems (Kostenuik 2005). Dabei ist RANK ein membranständigen Rezeptor, der von osteoklastären Zellen exprimiert wir und für die Osteoklasten-Entwicklung essentiell ist. RANKL ist der extrazelluläre Ligand, der RANK aktiviert. In der Abwesenheit von RANKL oder RANK können in vivo keine Osteoklasten mehr gebildet werden. RANKL wird von osteoblastären Zellen und aktivierten T-Zellen als membranständiges Protein oder als löslicher Faktor (soluble RANKL, sRANKL) produziert. OPG ist ein lösliches Rezeptorprotein, das unter anderem von osteoblastären Zellen produziert wird und RANKL bindet. Damit wird die Bindung von RANKL an RANK inhibiert. OPG ist deshalb ein stark antiresorptives Protein. Das RANKL/OPG-Verhältnis ist von entscheidender Bedeutung für die Steuerung der Knochenresorption, denn es spie-gelt wider, wie viel RANKL bioverfügbar ist. In der Abwesenheit von OPG kommt es zu einer massiven Steigerung der Knochenresorption und Spontanfrakturen bei Mäusen. Durch genetische Variabilität bedingte Unterschiede in der OPG-Expression bzw. in der biologischen Aktivität von OPG können des-halb einen großen Einfluss auf den Knochenstoffwechsel haben. LRP5 ist Teil des Wnt (Wingless)-Signaltransduktionswegs und agiert als Wnt-Corezeptor (Glass and Karsenty 2007, Liu et al 2008).

Erben: Knochenstoffwechsel und Mengenelemente
Seite 2 8. BOKU-Symposium Tierernährung 2009
Aktivierende Mutationen in LRP5 sowie inaktivierende Mutationen der Inhibitoren Sclerostin und Dick-kopf führen zu einer starken Erhöhung der Skelettmasse. In den letzten Jahren hat sich gezeigt, dass der Wnt-Signaltransduktionsweg, der ursprünglich in Drosophila melanogaster charakterisiert wurde, entscheidende Funktionen in der Osteo- und Chondrogenese sowie in der Knochenregeneration beim Säugetier hat. Es macht deshalb biologischen Sinn, dass die genetische Variabilität in diesen Loci ei-nen starken Einfluss auf die Skelettmasse hat.
Obwohl die Heritabilität der Skelettmasse und der BMD hoch ist, gibt es doch auch bedeutende Um-weltfaktoren, die den Knochen beeinflussen. Im Vordergrund stehen dabei physische Aktivität, Ernäh-rung und Sexualsteroide.
Es ist seit langem bekannt, dass Knochen als Organ auf biomechanische Belastung reagiert. Vermin-derte Aktivität induziert Knochenabbau, während verstärkte biomechanische Belastung zu einem ge-steigerten osteoblastären Knochenaufbau führt. Der Schlagarm bei Profi-Tennisspielern zeigt z.B. eine ca. 30%ige Erhöhung der BMD gegenüber dem kontralateralen Arm.
Da die Zusammensetzung von Knochenmineral chemisch vorgegeben ist, wird für eine adäquate Mine-ralisierung des Knochens Calcium und Phosphor benötigt. Steht eines oder beide dieser Elemente nicht in ausreichendem Maße zur Verfügung, kommt es zu gestörter Knochenmineralisation und ver-minderter Knochenmasse. Die Ernährung und damit auch die Zufuhr von Calcium und Phosphor für den Knochenaufbau und die Knochenmineralisierung können im besten Fall jedoch nur dazu führen, dass das genetische Potenzial für die peak bone mass optimal ausgenutzt wird. Es ist sehr wichtig, sich klar zu machen, dass eine vermehrte Zufuhr von Calcium und Phosphor über die Diät nicht auto-matisch zu einer höheren Knochenmasse und einer verbesserten biomechanischen Belastbarkeit führt. Die Menge an Mineral, die in die Knochenmatrix eingelagert werden kann, ist physikalisch-chemisch limitiert. Eine Erhöhung der Skelettmasse und der biomechanischen Belastbarkeit bei einer gegebenen genetischen Konstitution und gegebenem Sexualhormon-Status ist nur durch physische Aktivität er-reichbar. Die Funktion der Ernährung für den Knochen besteht letztlich also lediglich darin, das geneti-sche Potenzial optimal auszunutzen.
Endokrine Kontrolle des Knochenstoffwechsels Die Aufrechterhaltung einer konstanten Konzentration von ionisiertem Calcium im Extrazellulärraum ist von essentieller Bedeutung für höhere Organismen, denn Calcium spielt eine sehr wichtige Rolle in der Enzymaktivierung, Blutgerinnung, Permeabilität von Zellen und in der Signaltransduktion. Die Konzent-ration von ionisiertem Calcium im Extrazellulärraum wird deshalb sehr fein endokrin reguliert, wobei Parathormon (PTH), das Vitamin D-Hormon [1,25(OH)2D] und Sexualhormone die wichtigste Rolle spielen. Der praktisch alleinige Kurzzeitregulator des ionisierten Calciums im Extrazellulärraum und der endokrine Hauptregulator von Knochenresorption und Knochenremodeling ist PTH aus der Neben-schilddrüse. PTH steuert damit unter anderem auch den Fluss von Calcium und Phosphor aus dem Knochen in den Extrazellulärraum. Die biologisch wichtigste Wirkung von 1,25(OH)2D liegt in der Sti-mulation der intestinalen Calcium- und Phosphorresorption. Daneben fördert 1,25(OH)2D die renal-tubuläre Reabsorption von Calcium. 1,25(OH)2D ist für ein normales Wachstum und eine normale Mineralisierung von neugebildetem, unmineralisiertem Knochengewebe notwendig. Diese Wirkung von 1,25(OH)2D beruht jedoch nicht auf einem direkten Einfluss am Knochen, sondern auf der Bereitstel-lung von Calcium und Phosphat für den Mineralisierungsprozess durch eine gesteigerte intestinale Resorption. 1,25(OH)2D hat in physiologischen Konzentrationen wahrscheinlich keine direkte Wirkung am Knochen (Weber et al 2009). Sexualhormone haben eine komplexe direkte Wirkung am Knochen. Während bei höheren Säugern Estrogene vor allem eine antiresorptive Wirkung zeigen, stimulieren Estrogene beim Huhn und bei der Maus die endocorticale Knochenapposition. Androgene induzieren die Ausbildung des männlichen Skelettphänotyps und stimulieren bei Säugern das Längenwachstum sowie die radiäre periostale Knochenapposition, wodurch insgesamt größere Knochen entstehen.

Erben: Knochenstoffwechsel und Mengenelemente
8. BOKU-Symposium Tierernährung 2009 Seite 3
Knochen als endokrines Organ für die Mineralhomöostase In den letzten Jahren hat sich gezeigt, dass Knochen nicht nur als Zielorgan für endokrine Signale dient, sondern selbst auch endokrine Signalstoffe produziert, die im Wesentlichen an der Niere ihre Wirkung entfalten. Genetische und Tumor-Erkrankungen beim Menschen, so genannte „Phosphate-wasting disorders“, haben zur Entdeckung von Fibroblast growth factor-23 (FGF-23) geführt (Shimada et al 2001, The ADHR Consortium 2000). FGF-23 gehört zu den endokrin wirkenden Fibroblasten-Wachstumsfaktoren und ist ein phosphaturisches Hormon, das von Osteocyten sezerniert wird. Phos-phat und 1,25(OH)2D3 stimulieren die FGF-23-Sekretion. FGF-23 supprimiert am proximalen Nierentu-bulus die Expression der renalen 1α-Hydroxylase und inhibiert die renal-tubuläre Phosphat-Reabsorption. Damit sinkt indirekt – durch die verminderte 1,25(OH)2D3–Synthese - die intestinale Phosphataufnahme und die Niere scheidet mehr Phosphat aus. Dieser endokrine Faktor stellt somit eine negative Feed-back-Achse zwischen Knochen und Niere dar.
Säure-Basen-Metabolismus und Knochenstoffwechsel Seit langem war bekannt, dass die Resorptionsaktivität von Osteoklasten in vitro sehr stark durch den pH im Zellkulturmedium beeinflusst wird (Arnett et al 1994, Arnett and Spowage 1996). Ein sinkender pH verstärkt die Resorptionsaktivität, während eine Alkalisierung die Aktivität von Osteoklasten hemmt. Die Aufklärung des Zusammenhangs zwischen einer alkalischen Stoffwechsellage und dem Auftreten von Gebärparese bei der Kuh hat wesentlich zu der Erkenntnis beigetragen, dass dem Säu-re-Basenmetabolismus eine wichtige Rolle in der Steuerung der Knochenresorption zukommt (Goff 2006). Die Hemmung der Knochenresorption durch eine alkalische Stoffwechsellage in der peripartu-rienten Phase ist einer der wichtigsten Risikofaktoren bei der Gebärparese. Deshalb besteht eine der effektivsten Gebärparese-Präventionsmaßnahmen in der Ansäuerung der Diät durch bessere Rations-gestaltung unter Berücksichtigung der Natrium-, Kalium-, Chlorid-Gehalte oder durch externe Zugabe von sauren Salzen. Inzwischen haben große epidemiologische Studien beim Menschen ebenfalls nahe gelegt, dass es einen Zusammenhang zwischen osteoporotischem Frakturrisiko und der diätetischen Säurebelastung geben könnte (Dargent-Molina et al 2008). Falls sich der Zusammenhang von diäteti-scher Säurebelastung und Osteoporose beim Menschen erhärtet, könnte dies von erheblicher Bedeu-tung für die Prophylaxe dieser Erkrankung sein.
Zusammenfassung Die Forschung der letzten Jahre hat zu einem bedeutend besseren Verständnis der genetischen Steue-rung der Skelettmasse und des Knochenstoffwechsels, des Knochens als Organ sowie seiner Einbin-dung in endokrine Signal-Netzwerke und in den Gesamt-Stoffwechsel geführt. Es ist davon auszuge-hen, dass ein Teil dieser neuen Erkenntnisse auch einen bedeutenden Einfluss auf die Tier- und Hu-manernährung haben wird.
Literatur
Arnett T.R., Boyde A., Jones S.J., and Taylor M.L. (1994): Effects of medium acidification by alteration of carbon dioxide or bicarbonate concentrations on the resorptive activity of rat osteoclasts. J.Bone Miner.Res. 9, 375-379.
Arnett T.R., Spowage M. (1996): Modulation of the resorptive activity of rat osteoclasts by small changes in extracellular pH near the physiological range. Bone 18, 277-279.
Beamer W.G., Shultz K.L., Churchill G.A., Frankel W.N., Baylink D.J., Rosen C.J., and Donahue L.R. (1999): Quantitative trait loci for bone density in C57BL/6J and CAST/EiJ inbred mice. Mamm.Genome 10, 1043-1049.

Erben: Knochenstoffwechsel und Mengenelemente
Seite 4 8. BOKU-Symposium Tierernährung 2009
Beamer W.G., Shultz K.L., Donahue L.R., Churchill G.A., Sen S., Wergedal J.R., Baylink D.J., and Rosen C.J. (2001): Quantitative trait loci for femoral and lumbar vertebral bone mineral density in C57BL/6J and C3H/HeJ inbred strains of mice. J.Bone Miner.Res. 16, 1195-1206.
Dargent-Molina P., Sabia S., Touvier M., Kesse E., Breart G., Clavel-Chapelon F., and Boutron-Ruault M.C. (2008): Proteins, dietary acid load, and calcium and risk of postmenopausal fractures in the E3N French women prospective study. J.Bone Miner.Res. 23, 1915-1922.
Eisman J.A. (1999): Genetics of osteoporosis. Endocr.Rev. 20, 788-804.
Glass D.A., Karsenty G. (2007): In vivo analysis of Wnt signaling in bone. Endocrinology 148, 2630-2634.
Goff J.P. (2006): Major advances in our understanding of nutritional influences on bovine health. J.Dairy Sci. 89, 1292-1301.
Kostenuik P.J. (2005): Osteoprotegerin and RANKL regulate bone resorption, density, geometry and strength. Curr.Opin.Pharmacol. 5, 618-625.
Liu F., Kohlmeier S., and Wang C.Y. (2008): Wnt signaling and skeletal development. Cell Signal. 20, 999-1009.
Richards J.B., Rivadeneira F., Inouye M., Pastinen T.M., Soranzo N., Wilson S.G., Andrew T., Falchi M., Gwilliam R., Ahmadi K.R., Valdes A.M., Arp P., Whittaker P., Verlaan D.J., Jhamai M., Kumanduri V., Moorhouse M., van Meurs J.B., Hofman A., Pols H.A., Hart D., Zhai G., Kato B.S., Mullin B.H., Zhang F., Deloukas P., Uitterlinden A.G., and Spector T.D. (2008): Bone mineral density, osteoporosis, and osteoporotic fractures: a genome-wide association study. Lancet 371, 1505-1512.
Shimada T., Mizutani S., Muto T., Yoneya T., Hino R., Takeda S., Takeuchi Y., Fujita T., Fukumoto S., and Yamashita T. (2001): Cloning and characterization of FGF23 as a causative factor of tumor-induced osteomalacia. Proc.Natl.Acad.Sci.U.S.A 98, 6500-6505.
The ADHR Consortium (2000): Autosomal dominant hypophosphataemic rickets is associated with mutations in FGF23. Nat.Genet. 26, 345-348.
Weber K., Bergow C., Hirmer S., Schuler C., and Erben R.G. (2009): Vitamin D-independent therapeutic effects of extracellular calcium in a mouse model of adult-onset secondary hyperparathyroidism. J.Bone Miner.Res. 24, 22-32.
Autorenanschrift
Prof. Dr. Dr. Reinhold G. Erben Vetmeduni Vienna Institut für Physiologie, Pathophysiologie und Medizinische Physik Dept. für Biomedizinische Wissenschaften Veterinärplatz 1, A-1210 Wien, Österreich E-Mail: [email protected]

Rodehutscord und Haese: Wechselwirkungen zwischen der Phosphorversorgung und der mikrobiellen Gemeinschaft des Verdauungstraktes
8. BOKU-Symposium Tierernährung 2009 Seite 5
Wechselwirkungen zwischen der Phosphorversorgung und der mikrobiellen Gemeinschaft des Verdauungstraktes
Markus Rodehutscord und Eva Haese
Institut für Tierernährung, Universität Hohenheim
Einleitung Die Lagerstätten für Rohphosphate sind die wichtigsten Ressourcen, aus denen der Kreislauf Phos-phor(P)-haltiger Verbindungen kontinuierlich gespeist wird. Dass diese Lagerstätten endlich sind, ist lange bekannt (Mengel 1997). Europa ist weitgehend frei von solchen Lagerstätten und daher auf Importe angewiesen. Für die langfristige Sicherung der Phosphatversorgung gibt es daher keine Alter-native zu dem Anspruch, P-Kreisläufe möglichst geschlossen zu halten und hierdurch den weiteren Abbau der Lagerstätten von Rohphosphaten zu minimieren.
Seit etwa zwei Jahrzehnten ist die Optimierung der P-Versorgung landwirtschaftlicher Nutztiere Ge-genstand der Tierernährungsforschung. Für verschiedene Tierarten wurden die Empfehlungen zur Versorgung vom Ausschuss für Bedarfsnormen (AfBN) der Gesellschaft für Ernährungsphysiologie (GfE) überarbeitet. Zudem wurden die Methoden zur Ermittlung der Verwertbarkeit verschiedener P-Quellen weiterentwickelt. Anlass für diese Arbeiten gab zunächst die wachsende Erkenntnis, dass landwirtschaftliche Produktion in Regionen mit sehr hohen Phosphatgehalten der Böden für einen überhöhten Eintrag von Phosphaten in die Umwelt, insbesondere die Oberflächengewässer, mit ver-antwortlich ist. Zunehmend kam das Bewusstsein für die Begrenzung der Lagerstätten für Rohphos-phate und die Einbindung der Tierproduktion in die P-Kreisläufe hinzu. Während es bei den Empfeh-lungen zur Versorgung zunächst hauptsächlich darum gegangen war, einen Mangel in der Versorgung der Nutztiere sicher auszuschließen, wurde bei der Überarbeitung von Versorgungsempfehlungen in den zurückliegenden ca. 15 Jahren zunehmend auch der Anspruch berücksichtigt, eine hohe Verwer-tung des P durch die Tiere zu erreichen und die Ausscheidungen von P zu minimieren. Neue For-schungsergebnisse ermöglichten es, die Empfehlungen zur P-Versorgung für verschiedene Tierarten zu reduzieren.
Die Klärschlammverordnung und später das Verbot der Verfütterung von Tier- und Fleisch-knochenmehlen haben die Möglichkeiten für den Erhalt der Kreisläufe eingeschränkt, und je nach Verwendungsalternative gehen Phosphate dem Kreislauf unwiederbringlich verloren. Aus quantitativer Sicht ist die Vermeidung der Verluste, die mit der gegenwärtigen Praxis der Verwendung von Tier-mehlen und kommunalen Abwässern verbunden sind, eine gesamtgesellschaftlich viel größere Heraus-forderung als die weitere Optimierung der Fütterung unserer Nutztiere (Rodehutscord 2008).
Die Mikroorganismen des Verdauungstraktes spielen im Zusammenhang mit der P-Versorgung des Tieres eine eigene Rolle. Einerseits haben sie für ihren Stoffwechsel und ihre Vermehrung einen eige-nen Bedarf an P, insbesondere, aber nicht ausschließlich, für die Bildung von Nukleinsäuren (Van Ne-vel und Demeyer 1977) und den Energieumsatz. Andererseits sind sie zur Bildung von Enzymen befä-higt, die zur partiellen Hydrolyse von Phytin (Phytinsäure (IP6) plus Phytate) in der Lage sind. In pflanzlichen Samen liegen etwa zwei Drittel des Phosphors in Form von Phytin vor. Je nach Lokalisati-on im Verdauungstrakt wird durch die mikrobielle Gemeinschaft die Verwertung von Phytin-P unter-stützt. In diesem Beitrag sollen einige dieser Wechselwirkungen betrachtet werden, und zwar differen-ziert nach Wiederkäuern und Schweinen. Dabei soll auch der Frage nachgegangen werden, ob es bei einer reduzierten, aber noch bedarfsdeckenden P-Versorgung des Wirtstieres zu einer Einschränkung der mikrobiellen Aktivität kommen kann.

Rodehutscord und Haese: Wechselwirkungen zwischen der Phosphorversorgung und der mikrobiellen Gemeinschaft des Verdauungstraktes
Seite 6 8. BOKU-Symposium Tierernährung 2009
Wiederkäuer Bislang wird davon ausgegangen, dass die mikrobiell gebildeten Phytasen im Pansen eine weitgehend vollständige Hydrolyse des Phytins und eine entsprechend hohe Verwertung des P ermöglichen. Dies stützt sich auf Ergebnisse aus Versuchen mit kleinen Wiederkäuern, in denen eine Zulage von Futter-mitteln mit hohen Phytingehalten nicht zu einer Veränderung der P-Ausscheidung mit dem Kot geführt hat (Kincaid und Rodehutscord 2005; Koddebusch und Pfeffer 1988). In den Versorgungs-empfehlungen für Wiederkäuer wird daher im Hinblick auf die Verwertbarkeit des P nicht zwischen verschiedenen P-Quellen differenziert und ein einheitlicher Wert von 70 % unterstellt (GfE 1993; GfE 1995; GfE 2001). Aus Einzeluntersuchungen neueren Datums ist jedoch bekannt, dass es auch bei Wiederkäuern zur Ausscheidung von Phytin mit dem Kot kommen kann (Kincaid et al. 2005; Park et al. 2002) und der ruminale Phytinabbau nicht immer vollständig ist. Hierzu passen Befunde aus in situ-Studien, nach denen zwischen verschiedenen Einzelfuttermitteln erhebliche Unterschiede sowohl im ruminal löslichen als auch im ruminal potenziell abbaubaren Anteil des Phosphors bestehen (Bravo et al., 2000).
Durch eine Vorbehandlung von Futtermitteln, wie sie zum Beispiel zur Beeinflussung des Abbaus von Rohprotein vorgenommen wird, kann auch der Abbau des Phytins im Pansen beeinflusst sein. Eine Behandlung von Futtermitteln mit Formaldehyd führte zu einem Rückgang im Abbau von IP6, ähnlich wie im Abbau des Rohproteins (Martín-Tereso et al. 2009). (Bravo et al. 2002) untersuchten mit lak-tierenden Ziegen und trockenstehenden Kühen in situ die Auswirkungen einer Formaldehyd-behandlung von Raps- und Sojaextraktionsschrot. Die ruminale Löslichkeit des Phosphors ging bei beiden Tierarten nach Behandlung mit Formaldehyd signifikant zurück. Nach Formaldehydbehandlung war in in situ-Studien auch der ruminale Abbau von Trockenmasse aus Weizen und verschiedenen Ölschroten (Soja-, Raps- und Sonnenblumenextraktionsschrot) vermindert (Bravo et al. 2000). Gleich-zeitig ging die ruminale P-Freisetzung aus den Nylonbeuteln im Vergleich zu den unbehandelten Fut-termitteln deutlich zurück. Die Autoren vermuten, dass dies hauptsächlich auf einen verminderten Abbau von Phytin infolge einer durch Formaldehydbehandlung erschwerten Zugänglichkeit des Sub-strates für mikrobielle Phytasen zurückzuführen sei. Auch nach einer Hitzebehandlung ging der effek-tive Abbau des Phytins, ermittelt in in situ-Studien mit Schafen, signifikant zurück (Konishi et al. 1999).
In Untersuchungen mit duodenal-fistulierten Schafen war nach einer Hitzebehandlung von Raps-extraktionsschrot der Fluss von Inositolphosphaten (IP6, IP5, IP4 und IP3) am Duodenum signifikant erhöht (Park et al. 2000). Der überwiegende Anteil der Inositolphosphate entfiel auf IP6. Bis zu 55% des in Inositolphosphaten gebundenen Phosphors wurden nach Hitzebehandlung im Duodenum wie-dergefunden, während es beim unbehandelten Schrot 22% waren.
Yanke et al. (1998) bestimmten in vitro die Phytaseaktivität anaerober ruminaler Bakterien. Zur Ge-winnung der ruminalen Proben wurden fistulierte Ochsen bei einem unterschiedlichen Verhältnis von Heu und Gerste in der Ration gefüttert. Je höher der Anteil der Gerste in der Ration war, desto höher war die Phytaseaktivität in der mittels Zentrifugation gewonnenen Mikrobenmasse. Da Gerste einen deutlich höheren Phytingehalt als Heu aufweist, liegt die Vermutung nahe, dass die ruminalen Bakteri-en mit entsprechend erhöhter Phytaseaktivität reagiert haben.
In einem Pansensimulationssystem (Rusitec) ermittelten Godoy und Meschy (2001) die Phytase-aktivität ruminaler Mikroorganismen bei Verwendung von Rationen mit hohem bzw. niedrigem Grob-futteranteil (80 bzw. 60% der TM) und hohem Phytingehalt. Es wurde mit zwei verschiedenen Puffern gearbeitet (75% des P im Puffer entweder in anorganischer oder organischer Form (Na-Phytat)). Der Abbau des Phytins lag bei hohem Grobfutteranteil und organischem P im Puffer bei 65%, bei niedri-gem Grobfutteranteil bei 70%. Die unvollständige Hydrolyse des Phytins kann möglicherweise auf eine Sättigung der Enzymaktivität zurückgeführt werden, da der Phytingehalt in diesem Versuch sehr hoch war. Bei anorganischem P im Puffer betrug der Abbau des Phytins 51% bei hohem und 48% bei nied-rigem Grobfutteranteil, und war damit signifikant geringer als bei Verwendung von organischem P im

Rodehutscord und Haese: Wechselwirkungen zwischen der Phosphorversorgung und der mikrobiellen Gemeinschaft des Verdauungstraktes
8. BOKU-Symposium Tierernährung 2009 Seite 7
Puffer. Hier ist es möglicherweise zu einer Hemmung der Phytasebildung der Mikroben durch anorga-nisches Phosphat gekommen.
In diesem Zusammenhang ist von Interesse, wie hoch der „Bedarf“ der Pansenmikroorganismen an P ist. Es ist wiederholt gezeigt worden, dass durch einen induzierten starken Mangel an P in der Ration Futteraufnahme, Nährstoffverdaulichkeit, Fermentation und Effizienz der mikrobiellen Proteinsynthese deutlich vermindert sind (Breves und Höller 1987; Durand et al. 1982; Müschen et al. 1988; Petri et al. 1988). Diese Effekte sind bei laktierenden Wiederkäuern eher zu beobachten als bei nicht-laktierenden, weil bei letzteren der Bedarf des Tieres an P geringer ist und die endogene Phosphatsek-retion über den Speichel eher zu einem Ausgleich von Defiziten in der Zufuhr mit dem Futter führen kann. Die Phosphorkonzentration in der Pansenflüssigkeit kann je nach P-Gehalt der Ration sehr un-terschiedlich sein und korreliert eng mit den Gehalten an anorganischem Phosphat im Blut der Tiere (Rodehutscord et al. 1994). Die Ableitung eines Mindestgehaltes an P in der Pansenflüssigkeit zur Gewährleistung einer uneingeschränkten mikrobiellen Aktivität ist allerdings schwierig und unter ande-rem von dem gewählten Wirkungskriterium abhängig. Untersuchungen hierzu wurden häufig mit in vitro-Ansätzen durchgeführt. Es darf aber davon ausgegangen werden, dass die P-Versorgung des Tieres, solange sie die Versorgungsempfehlungen nicht unterschreitet, immer auch den spezifischen P-Bedarf der Mikroorganismen des Pansens deckt.
Schweine Beim Schwein haben die Wechselwirkungen zwischen der P-Versorgung und den Bakterien in Dünn- und Dickdarm eine unterschiedliche Bedeutung. Die Zusammensetzung der Ration, insbesondere die Fraktion der Kohlenhydrate, wirkt sich unterschiedlich aus. In einer Kooperation zwischen den Univer-sitäten Halle und Hohenheim wurden diese Zusammenhänge mit wachsenden Schweinen in zwei Dok-torarbeiten näher untersucht (Baumgärtel 2009; Metzler-Zebeli 2007) und einige Ergebnisse sollen nachfolgend präsentiert werden. Die Schweine waren mit T-Kanülen am terminalen Ileum versehen, so dass Messungen zu den praecaecalen und postilealen Umsetzungen gemacht werden konnten.
Bei Einsatz einer P-armen Grundmischung auf der Basis von Körnermais und Sojaextraktionsschrot betrug die Verdaulichkeit des P im gesamten Verdauungstrakt 28% und praecaecal 26% (Tab. 1). Dies bestätigt die Größenordnung der Werte vergleichbarer früherer Versuche und zudem die Beo-bachtung, dass postileal keine nennenswerte Nettoabsorption von P erfolgt. Eine Zulage von Lignocel-lulose zur Grundmischung führte nicht zu Veränderungen in den ermittelten Verdaulichkeitswerten. Zulagen von Maisstärke und Apfelpektin wirkten sich hingegen aus, und zwar unterschiedlich. Die Zulage von Maisstärke bewirkte tendenziell eine Verminderung der praecaecalen P-Verdaulichkeit, die sich auch in einem verminderten Abbau von IP6 widerspiegelte. Die Zulage von Pektin hingegen führ-te zu einer verminderten Verdaulichkeit des P am Ende des Verdauungstraktes. Die postileale Netto-absorption des P war folglich zwischen den Varianten mit Stärke- und Pektinzulage signifikant ver-schieden. Während die Messwerte für die Stärkezulage auf eine geringfügige Nettoabsorption von P im Dickdarm hindeuten, hat es bei Pektinzulage eine Nettosekretion von P gegeben. Diese Unterschie-de dürften mit der mikrobiellen Aktivität zusammenhängen und die Mikrobengemeinschaft beeinflusst haben, weil zumindest im Dickdarm der Kohlenhydratabbau auf mikrobielle Aktivität zurückzuführen ist. Zieht man die Konzentrationen der flüchtigen Fettsäuren (VFA) im Chymus und Kot als Indikatoren heran, so deuten sie deutliche Unterschiede im Umfang der mikrobiellen Umsetzung an, die sich auch im ermittelten Abbau der organischen Masse widerspiegeln. So war im Chymus die VFA-Konzentration bei Zulage von Stärke höher als bei Zulage der beiden anderen Quellen, während sie im Kot bei Zula-ge von Pektin höher war als bei den anderen beiden Quellen. In isolierten Bakterien des Kotes (über Zentrifugation gewonnene Mischfraktion) waren der P-Gehalt signifikant vermindert und das N:P-Verhältnis signifikant erhöht, wenn die Schweine die Zulage von Pektin erhielten. Nach Verabreichung von Pektin per fistulam war die Aktivität von Cellulase im Kot der Schweine signifikant vermindert und die der Polygalakturonase signifikant erhöht (Metzler et al. 2008). All dies deutet auf Veränderungen

Rodehutscord und Haese: Wechselwirkungen zwischen der Phosphorversorgung und der mikrobiellen Gemeinschaft des Verdauungstraktes
Seite 8 8. BOKU-Symposium Tierernährung 2009
sowohl in der Größe als auch in der Zusammensetzung der Mikrobengemeinschaft des Verdauungs-traktes hin. In der Tat wurde in ergänzenden Studien festgestellt, dass interessanterweise die Zulage von Maisstärke zu einer deutlichen Erhöhung der Konzentration von Bakterien, insbesondere der Lac-tobacillen, im Ileumchymus führte (Metzler et al. 2009b). Ob hierüber auch der bei Stärkezulage beo-bachtete Rückgang im Abbau des IP6 erklärt werden kann, weil eventuell die bakterielle Phytase-synthese beeinflusst war, oder der bei veränderter Milchsäuresynthese beeinflusste pH-Wert die Akti-vität der (geringen) pflanzlichen Phytaseaktivität beeinflusst hat, ist noch nicht geklärt.
Tabelle 1: Einfluss von Zulagen verschiedener Kohlenhydratquellen zu einer P-armen Grundmischung (GM) bei Schweinen (Baumgärtel et al. 2008; Metzler et al. 2009a)
GM 75% GM plus 25% 100% Cellulose Stärke Pektin Praecaecal
P-Verdaulichkeit (%) 26 25 15* 17 IP6-Abbau (%) 60 60 18 64 VFA-Konzentration im Chymus (mmol/kg TM)
665ac 429b 830a 414bc
Postileale P-Verdaulichkeit (%) 3ab -5b 9a -4b Gesamttrakt bzw. Kot
P-Verdaulichkeit (%) 28a 24ab 23ab 15b P in Mikrobenmasse (g/kg TM) 23a 20a 19ab 13b N:P-Verh. in Mikrobenmasse 2,6a 3,3ab 3,2ab 4,3b VFA-Konzentration im Kot (mmol/kg TM)
349ac 195b 244bc 451a
a,b,c Ungleiche Hochbuchstaben kennzeichnen signifikante Unterschiede (P≤0,05) * im Vergleich zur Grundmischung, P<0,10
Wenn Schweine auf einem niedrigen Niveau der P-Versorgung nur mit pflanzlichem P versorgt wer-den, scheiden sie im Kot kaum IP6 aus, auch wenn die Futterration im Wesentlichen auf Mais und Sojaextraktionsschrot basiert (Baumgärtel 2009). Der IP6-Abbau erfolgt überwiegend im Dickdarm, allerdings ist der praecaecale Abbau von IP6 bei Zusatz von Phytase zum Futter erhöht. Eine Erhö-hung der P-Versorgung durch Zulage von Monocalciumphosphat (MCP) führte nicht nur zu Verände-rungen in der Verdaulichkeit des P insgesamt, sondern auch zu einem deutlich verminderten postilea-len Abbau von IP6 und somit zur Ausscheidung von IP6 mit dem Kot. Dies kann als Bestätigung für in vitro-Befunde angesehen werden, in denen eine hemmende Wirkung einer erhöhten Phosphatkon-zentration auf die Aktivität bakterieller Phytase nachgewiesen wurde (Konietzny und Greiner 2002).
Häufig wurde gezeigt, dass bei Zusatz von mikrobieller Phytase zu einer Futtermischung auf Basis von pflanzlichen P-Quellen die praecaecale Hydrolyse von Inositolphosphaten und die Verdaulichkeit des P gesteigert werden. Baumgärtel (2009) zeigte, dass dies bei knapper P-Versorgung auch mit einer Ver-änderung der postilealen P-Umsetzung gekoppelt ist: eine Phytasezulage führte zur signifikanten Ver-minderung der postilealen Nettoabsorption des P von 0 auf -10%. Vermutlich hat die Verminderung in der Phytatanflutung im Dickdarm, die mit der Steigerung der praecaecalen Verdaulichkeit verbunden war, zu einer Sekretion von P geführt, womit die Minderversorgung der Mikroben zumindest teilweise kompensiert werden konnte. Die Ergebnisse zeigten aber auch, dass die postileale Fermentation des per fistulam verabreichten Pektins durchgehend hoch war. Weder durch den Zusatz von MCP noch durch den Zusatz von Phytase wurde sie gesteigert, so dass auch die niedrige P-Versorgung des Tie-res für die Fermentation des Pektins keine Begrenzung darstellte.
Man kann in der Gesamtschau die folgenden Schlussfolgerungen ziehen. Verschiedene Kohlenhydrat-quellen beeinflussen je nach Verdaulichkeit und Fermentierbarkeit die mikrobielle Gemeinschaft des

Rodehutscord und Haese: Wechselwirkungen zwischen der Phosphorversorgung und der mikrobiellen Gemeinschaft des Verdauungstraktes
8. BOKU-Symposium Tierernährung 2009 Seite 9
Verdauungstraktes von Schweinen quantitativ und qualitativ. Dies kann mit einem Einfluss auf die praecaecale Hydrolyse von Phytat und somit auf die Versorgung des Schweins mit verdaulichem P verbunden sein. Im Dickdarm beeinflusst die anflutende fermentierbare Energie den Umfang der mikrobiellen Aktivität sowie die Zusammensetzung der mikrobiellen Gemeinschaft und damit auch den P-Bedarf der Mikroben. Ist die Anflutung praecaecal unverdaulichen Phosphors gering, kann es zu einer Sekretion von P in den Dickdarm kommen, der den Mikroben zur Bedarfsdeckung zur Verfügung steht und mit dem Kot ausgeschieden wird. Auch wenn P nicht in einer für das Schwein bedarfs-deckenden Konzentration im Futter enthalten ist, wird die Fermentation im Dickdarm uneingeschränkt verlaufen.
Literatur
Baumgärtel, T. (2009): Einfluss einer Zulage unterschiedlicher Kohlenhydrate auf die praecaecale und postileale Umsetzung und die Ausscheidung von Phosphor beim Schwein. Diss. agr., Universität Halle-Wittenberg
Baumgärtel, T., Metzler, B.U., Mosenthin, R., Greiner, R. und Rodehutscord, M. (2008): Precaecal and postileal metabolism of P, Ca and N in pigs as affected by different carbohydrate sources fed at low level of P intake. Arch. Anim. Nutr. 62, 169 - 181.
Bravo, D., Meschy, F., Bogaert, C. und Sauvant, D. (2000): Ruminal phosphorus availability from several feedstuffs measured by the nylon bag technique. Reprod. Nutr. Dev. 40, 149-162.
Bravo, D., Meschy, F., Bogaert, C. und Sauvant, D. (2002): Effects of fungal phytase addition, formaldehyde treatment and dietary concentrate content on ruminal phosphorus availability. Anim. Feed Sci. Techn. 99, 73-95.
Breves, G. und Höller, H. (1987): Effects of dietary phosphorus depletion on dry matter and organic matter digestibility. J. Anim. Physiol. Anim. Nutr. 58, 281-286.
Durand, M., Bertier, B., Hannequart, G. und Guéguen, L. (1982): Influence d'une subcarence in phosphore et d'un excès de calcium alimentaire sur la phoshatémie et les teneurs en phosphore et calcium des contenus de rumen du mouton. Reprod. Nutr. Dévelop. 22, 865-879.
Gfe (1993): Überarbeitete Empfehlungen zur Versorgung von Milchkühen mit Calcium und Phosphor. Proc. Soc. Nutr. Physiol. 1, 108-113.
Gfe (1995): Energie- und Nährstoffbedarf landwirtschaftlicher Nutztiere. Nr. 6: Empfehlungen zur Energie- und Nährstoffversorgung der Mastrinder. DLG-Verlag, Frankfurt a. M.
Gfe (2001): Empfehlungen zur Energie- und Nährstoffversorgung der Milchkühe und Aufzuchtrinder. DLG-Verlag, Frankfurt am Main.
Godoy, S. und Meschy, F. (2001): Utilisation of phytate phosphorus by rumen bacteria in a semi-continuous culture system (Rusitec) in lactating goats fed on different forage to concentrate ratios. Reprod. Nutr. Dev. 41, 259-265.
Kincaid, R. und Rodehutscord, M. (2005): In: E. Pfeffer und A.N. Hristov, Eds. Nitrogen and phosphorus nutrition of cattle: Phosphorus metabolism in the rumen. CABI Publishing: Wallingford, 2005; pp 187-193.
Kincaid, R.L., Garikipati, D.K., Nennich, T.D. und Harrison, J.H. (2005): Effect of grain source and exogenous phytase on phos-phorus digestibility in dairy cows. J. Dairy Sci. 88, 2893-2902.
Koddebusch, L. und Pfeffer, E. (1988): Untersuchungen zur Verwertbarkeit von Phosphor verschiedener Herkünfte an laktierenden Ziegen. J. Anim. Physiol. Anim. Nutr. 60, 269-275.
Konietzny, U. und Greiner, R. (2002): Molecular and catalytic properties of phytate-degrading enzymes (phytases). Intern. J. Food Sci. Technol. 37, 791-812.
Konishi, C., Matsui, T., Park, W., Yano, H. und Yano, F. (1999): Heat treatment of soybean meal and rapeseed meal suppresses rumen degradation of phytate phosphorus in sheep. Anim. Feed Sci. Techn. 80, 115-122.
Martín-Tereso, J., Gonzalez, A., Van Laar, H., Burbano, C., Pedrosa, M.M., Mulder, K., Den Hartog, L.A. und Verstegen, M.W.A. (2009): In situ ruminal degradation of phytic acid in formaldehyde-treated rice bran. Anim. Feed Sci. Techn. In Press, Corrected Proof,
Mengel, K. (1997): Agronomic measures for better utilization of soil and fertilizer phosphates. Eur. J. Agron. 7, 221-233.
Metzler-Zebeli, B.U. (2007): Effects of fermentable carbohydrates and dietary P supply on bacterial P incorporation, activity and composition in the gastrointestinal tract of pigs. Diss. agr., Universität Hohenheim, Cuvillier Verlag, Göttingen

Rodehutscord und Haese: Wechselwirkungen zwischen der Phosphorversorgung und der mikrobiellen Gemeinschaft des Verdauungstraktes
Seite 10 8. BOKU-Symposium Tierernährung 2009
Metzler, B.U., Mosenthin, R., Baumgartel, T. und Rodehutscord, M. (2008): The effect of dietary phosphorus and calcium level, phytase supplementation, and ileal infusion of pectin on the chemical composition and carbohydrase activity of fecal bacteria and the level of microbial metabolites in the gastrointestinal tract of pigs. J. Anim. Sci. 86, 1544-1555.
Metzler, B.U., Mosenthin, R., Baumgärtel, T. und Rodehutscord, M. (2009a): Effects of fermentable carbohydrates and low dietary phosphorus supply on the chemical composition of faecal bacteria and microbial metabolites in the gastrointestinal tract of pigs. J. Anim. Physiol. Anim. Nutr. 93, 130-139.
Metzler, B.U., Vahjen, W., Baumgärtel, T., Rodehutscord, M. und Mosenthin, R. (2009b): Changes in bacterial populations in the ileum of pigs fed low-phosphorus diets supplemented with different sources of fermentable carbohydrates. Anim. Feed Sci. Techn. 148, 68-89.
Müschen, H., Petri, A., Breves, G. und Pfeffer, E. (1988): Response of lactating goats to low phosphorus intake. 1. Milk yield and faecal excretion of P and Ca. J. Agric. Sci., Camb. 111, 255-263.
Park, W.-Y., Matsui, T., Yano, F. und Yano, H. (2000): Heat treatment of rapeseed meal increases phytate flow into the duode-num of sheep. Anim. Feed Sci. Techn. 88, 31-37.
Park, W.-Y., Matsui, T. und Yano, H. (2002): Post-ruminal phytate degradation in sheep. Anim. Feed Sci. Techn. 101, 55-60.
Petri, A., Müschen, H., Breves, G., Richter, O. und Pfeffer, E. (1988): Response of lactating goats to low phosphorus intake. 2. Nitrogen transfer from rumen ammonia to rumen microbes and proportion of milk protein derived from microbial amino acids. J. Agric. Sci., Camb. 111, 265-271.
Rodehutscord, M. (2008): Ansatzpunkte zur Schonung der begrenzten Phosphorressourcen. Arch. Tierz. 51, 39-48.
Rodehutscord, M., Pauen, A., Windhausen, P., Brintrup, R. und Pfeffer, E. (1994): Effects of drastic changes in P intake on P concentrations in blood and rumen fluid of lactating ruminants. J. Vet. Med. A 41, 611-619.
Van Nevel, B.J. und Demeyer, D.I. (1977): Determination of rumen microbial growth in vitro from 32P-labelled phosphate incor-poration. Br. J. Nutr. 38, 101-114.
Yanke, L.J., Bae, H.D., Selinger, L.B. und Cheng, K.-J. (1998): Phytase activity of anaerobic ruminal bacteria. Microbiology 144, 1565-1573.
Autorenanschrift
Prof. Dr. M. Rodehutscord und Eva Haese Institut für Tierernährung Universität Hohenheim Emil-Wolff-Str. 10 70599 Stuttgart E-Mail: [email protected]

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
8. BOKU-Symposium Tierernährung 2009 Seite 11
Magnesium Stoffwechsel: Regulation und Risiken
Holger Martens und Friederike Stumpff
Institut für Veterinär-Physiologie, Freie Universität Berlin, Oertzenweg 19b, 14163 Berlin
Einleitung Magnesium ist ein essentieller Mineralstoff, der nicht durch Hormone reguliert wird, obwohl eine Be-einflussung der Blutmagnesiumkonzentration durch Hormone wie Insulin (Perrson and Luhmann 1974) und Katecholamine möglich ist (Rayssiguier 1977). Die zahlreichen Funktionen des Magnesiums bein-halten die Aktivierung von Enzymen (Günther 1977), die Regulation von Ionenkanälen (Stumpff und Martens 2006) und die Beeinflussung der Synapsenfunktion (Lit. siehe Meyer und Scholz 1972). Fer-ner dient Magnesium als Strukturkomponente des Knochens. Entsprechend den Funktionen des Mag-nesiums ergibt sich die Verteilung im Körper: 2- 3 g im extrazellulären Raum (EZR), 150 g intrazellulär (IZR) und 270 – 280 g im Knochen (Kuh 650 kg KGW).
Störungen des Mg-Haushalts werden mit Ausnahme der Wiederkäuer selten beobachtet und verursa-chen bei geringen Hypomagnesämien relativ unspezifische, bei gravierenden Hypomagnesämien die typischen motorischen Störungen.
Kenntnisse über die Prinzipien der Regulation des Mg-Haushalts erleichtern einerseits das Verständnis der Pathogenese der Hypomagnsämie einschließlich der klinischen Symptome und zeigen andererseits auch mögliche Gefährdungen des Mg-Haushalts auf. In der vorliegenden Arbeit werden die hierzu notwendigen Erkenntnisse und Zusammenhänge aufgezeigt.
Regulation des Magnesiumstoffwechsels Die physiologische Konzentration von Magnesium im Blut (0.75 – 1.10 mmol/l) wird einerseits be-stimmt durch die Absorption aus dem Verdauungskanal (a) bzw. dem Transfer des Mg in das Blut oder EZR. Die Deckung des Magnesiumbedarfs erfolgt andererseits aus dem Mg-Pool des Blutes bzw. des EZR und umfasst den Mg-Bedarf für das Wachstum (b: Gewebe und Knochen), für das foetale Wachs-tum im Falle einer Trächtigkeit (c), die Milchsekretion (d) und die nicht vermeidbaren Verluste in den Magen-Darm-Kanal (e). Die Blutmagnesiumkonzentration wird konstant gehalten, wenn
a – [(b) + (c) + (d) + (e)] > 0 (Gleichung 1)
ist. Die über den Bedarf aus dem Magen-Darm-Kanal absorbierte Mg-Menge wird renal ausgeschie-den. Da wegen der fehlenden hormonellen Regulation die intrazellulär und im Knochen vorhandenen Mg-Mengen nicht mobilisiert werden können, ergibt sich zwangsläufig eine Hypomagnesämie (Mg-Pool im EZR↓), wenn (a) infolge unzureichender Mg-Aufnahmen und/oder gestörter Magnesiumabsorption geringer ist als der Bedarf [(b) + (c) + (d) + (e)].
Hypomagnesämie: a < [(b) + (c) + (d) + (e)] (Gleichung 2)
Unter diesen Umständen verringert sich die renale Mg-Ausscheidung und die Mg-Konzentration im Urin sinkt unter 1 mmol/l (siehe unten).
Da aufgrund vieler Untersuchungen bei landwirtschaftlichen Nutzieren der Mg-Bedarf und die zur Be-darfsdeckung notwendigen oralen Aufnahmen bekannt sind und somit (a) (Gleichung 1) definiert wer-

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
Seite 12 8. BOKU-Symposium Tierernährung 2009
den kann, werden Hypomagnesämien aufgrund unzureichender Mg-Aufnahmen mit dem Futter selten beobachtet, die jedoch dennoch auftreten können, wenn trotz bedarfsdeckender Mg-Aufnahmen die absorbierte Mg-Menge (a) infolge der verringerter Mg-Absorption aus dem Verdauungskanal den ak-tuelle Mg-Bedarf [(b) + (c) + (d) + (e)] nicht decken kann. Eine Verringerung der Mg-Absorption durch Futterinhaltsstoffe ist im ausgeprägten Maßstab nur von Wiederkäuern bekannt, wobei bei Milchkühen eine gestörte Absorption und ein erhöhter Mg-Bedarf durch die Milchproduktion gleichzei-tig auftreten können und dadurch das Risiko von Hypomagnesämien/Tetanien erhöht wird. Aus die-sem Grunde beschränken sich die weiteren Ausführungen auf die Problematik der möglichen Störung des Mg-Stoffwechsels bei Kühen.
Risken des Magnesiumstoffwechsels Hypomagnesämie des Rindes Klinische Symptome und Inzidenz: Eine Hypomagnesämie verursacht aufgrund der vielfältigen Funktionen des Mg eine Reihe relativ unspezifischer Symptome wie verminderte Futteraufnahme und Milchproduktion, unsicherer Gang, Zähneknirschen und schließlich Festliegen und Tetanien, die ohne Behandlung häufig zum Tode führen können. Die klinischen Symptome Festliegen und Tetanien wer-den ausgelöst durch ein Abfallen der Mg-Konzentration in der cerebrospinalen Flüssigkeit (< 0.7 mmol/l), weil die hemmende Wirkung des Mg bei der synaptischen Übertragung abnimmt und da-durch unkontrollierte Erregungsübertragungen erleichtert werden (Lit. siehe Meyer und Scholz 1972). Blutmagnesiumkonzentration < 0.7 mmol/l sind als Hypomagnesämie anzusehen und Konzentrationen < 0.5 mmol/l Mg im Blut bedeuten ein erhöhtes Tetanierisiko, weil bei dieser Blutkonzentration (< 0.5 mmol/l) die Mg-Konzentration in der cerebrospinalen Flüssigkeit nicht länger > 0.7 mmol/l stabilisiert werden kann (Meyer und Scholz 1972).
Obwohl Berichte über das Vorkommen von Tetanien aus vielen Ländern vorliegen, gibt es keine epi-demiologisch verlässlichen Daten und aktuell dürfte das Risiko << 1 % sein. Dennoch können auch heute Hypomagnesämien und Todesfälle gehäuft vorkommen, die sich in der Regel dann jedoch im-mer nur auf einen Betrieb beziehen (Donovan et al., 2004). Subklinische Hypomagnesämien kommen wahrscheinlich wesentlich häufiger vor (McCoy et al. 1996), weil mögliche Symptome wie verminderte Futteraufnahmen oder Milchproduktion sehr unspezifisch sind. Zur Diagnose sollte immer die Bestim-mung der Konzentration von Magnesium im Blut (Verdacht ab < 0.8 – 0.9 mmol/l) und im Harn (< 1 mmol/l; siehe unten) erfolgen.
Ätiologie: Das mögliche Risiko einer Hypomagnesämie wurde abstrakt mit der Gleichung (2) be-schrieben. Da bei erwachsenen Tieren der Mg-Bedarf für das Gewebe/Knochen (b), für das foetale Wachstum (c) und die endogenen Verluste (e) entweder gering (b + c) oder als konstant (e) anzuse-hen sind, wird die Blutmagesiumkonzentration maßgeblich durch die Absorption und den Bedarf für die Milch bestimmt. Die mögliche Labilität der Blutmagnesiumkonzentration wird erkennbar, wenn man sich den Mg-Pool im EZR in Erinnerung ruft, der nur 2 - 3 g Mg beträgt. Da die Konzentration von Mg in der Milch relativ konstant gehalten wird und 120 – 150 mg/l beträgt, werden bei einer Milchleis-tung von 40 l/d 4.8 – 6.0 g Mg oder ungefähr die zweifache Menge des Mg-Pools im EZR benötigt. Wegen der fehlenden hormonellen Regulation können die Mg-Mengen im IZR oder im Knochen nicht zur akuten Bedarfsdeckung mobilisiert werden (Abb. 1).
Aufgrund des raschen Turnovers des Mg im EZR (Abb. 1) kann die Konzentration von Mg im Blut nur konstant gehalten werden, wenn der Mg-Bedarf für die Milch kontinuierlich durch eine entsprechende Absorption aus dem Verdauungskanal ersetzt wird. Ist das bei gestörter Absorption nicht der Fall, ergibt sich das Risiko einer Hypomagnesämie, da die Mg-Konzentration in der Milch unabhängig vom Mg-Status der Kuh konstant gehalten wird.

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
8. BOKU-Symposium Tierernährung 2009 Seite 13
Abbildung 1: Schematische Darstellung des Mg-Stoffwechsels einer Milchkuh mit einer Milchproduk-tion von 40 l/d (Martens and Schweigel 2000, with permission). Absorptiont: Wahre Verdaulichkeit
Von maßgeblicher Bedeutung für die physiologische Mg-Konzentration im Blut ist daher die ausrei-chende und ungestörte Absorption von Mg aus dem Verdauungskanal. Genaue Kenntnisse über die Mg-Absorption und deren mögliche Beeinflussung sind somit als wesentliche Voraussetzung zum Ver-ständnis der Pathogenese der Hypomagnesämie und Tetanie infolge gestörter Absorption beim Rind anzusehen.
Magnesiumabsorption im Magen-Darm-Kanal: Die scheinbare Verdaulichkeit von Magnesium bei Kühen nimmt im Bereich der üblichen oralen Mg-Aufnahme von 40 – 80 g/d linear zu und beträgt etwa 20 %, kann jedoch auf Werte < 10 % sinken. Diese niedrige scheinbare Verdaulichkeit ist nicht auf eine erhöhte Sekretion von Mg in den Magen-Darm-Kanal zurückzuführen, die mit 3 – 4 mg/k KGW relativ konstant ist, sondern ist bedingt durch Futterinhaltsstoffe (siehe unten), deren Wirkung nur verstanden werden kann, wenn a) die Lokalisation und b) der Mechanismus der Mg-Absorption bekannt ist.
Lokalisation der Mg-Absorption: Die Absorption von Mg erfolgt nur in den Vormägen mit Hilfe eines aktiven Transportmechanismus und ist essentiell für die Aufrechterhaltung der physiologischen Mg-Konzentration im Blut (Literatur siehe Martens und Schweigel 2000). Im Dünn- und Dickdarm findet keine nennenswerte Nettoresorption von Mg statt und eine gestörte Mg-Absorption in den Vormägen wird nicht im Dünn- und Dickdarm kompensiert.
Mechanismus der Mg-Absorption: Zwei luminale Aufnahmemechanismen sind im Pansenepithel für Mg beschrieben worden. Ergebnisse von in vivo und in vitro Versuchen unterstützen die Annahme, dass die Aufnahme des Mg2+ Ions aus der Pansenflüssigkeit durch die luminale (apikale) Membran des Pansenepithels (wahrscheinlich) durch einen Kanal erfolgt (Abb. 2).

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
Seite 14 8. BOKU-Symposium Tierernährung 2009
Abbildung 2: Schematische Darstellung des ruminalen Mg-Transportes. Die Mg-Aufnahme aus der Pansenflüssigkeit durch die luminale Membran in die Epithelzelle erfolgt einerseits als Mg2+ Ion (wahr-scheinlich) durch einen Kanal (1) und andererseits im Cotransport zusammen mit 2 A- Ionen (wahr-scheinlich Cl-; 2). Der basolaterale Heraustransport aus der Zelle wird durch einen Na+/Mg2+ Austau-scher vermittelt (3). Das dabei in die Zelle aufgenommene Na wird mit Hilfe der Na+/K+-ATPase (4) aktiv heraustransportiert. PDa, PDb: Potenzialdifferenz der apikalen und basolateralen Membran. PDt: transepitheliale Potenzialdifferenz: PDt = PDa – PDb. Modifiziert nach Martens und Schweigel, 2000.
Diese luminale Mg-Aufnahme ist als passiv anzusehen und wird daher maßgeblich durch die Potenzial-differenz der apikalen Membran, PDa, bestimmt. Diese Abhängigkeit vom elektrischen Gradienten als Triebkraft hat zu der Bezeichnung potenzialabhängiger Mg-Transport geführt. Ferner existiert ein wei-terer, potenzialunabhängiger Aufnahmemechanismus. Es handelt sich sehr wahrscheinlich um einen Cotransport des zweiwertigen Mg2+ Ions mit 2 Cl- Ionen, der elektroneutral und daher potenzialunab-hängig ist.
Mg2+
Mg2+
2 A -
Mg2+
Na+ Mg2+
- + - +
Na+
K+
K+
K+ K+
Mg2+
Mucosal Serosal
Na/K-ATPase Carrier
(PDa) (PDb)
- + PDt = PDa - PDb
(1)
(2)
(3)
(4)
Kanal
2 A -
2 A -
K+

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
8. BOKU-Symposium Tierernährung 2009 Seite 15
Die Abgabe durch die basolaterale Membran wird mit Hilfe eines Na+/Mg2+-Austauschers ermöglicht. Das dabei aufgenommene Na+ wird mit Hilfe der Na+/K+-ATPase wieder aus der Zelle entfernt, d.h. der transepitheliale Mg2+-Transport ist als sekundär aktiv zu bezeichnen (Abb. 2; Literatur zum epithe-lialen Mg-Transport: Martens and Blume 1987; Martens et al. 1987a; Leonhard-Marek and Martens 1996; Schweigel et al. 1999; Schweigel et al. 2000; Schweigel and Martens 2003; Leonhard-Marek et al. 2005).
Die Existenz zweier luminaler Aufnahmemechanismen für Mg2+ führt zu der Frage der physiologischen Bedeutung. Da die Mg-Absorption aus den Vormägen zur Sicherstellung einer normalen Blutmagnesi-umkonzentration essentiell ist, müssen die ruminalen Mg-Transportmechanismen insbesondere auch bei sehr niedrigen Mg2+-Konzentrationen (1 mmol/l oder weniger) eine ausreichende Mg-Absorption ermöglichen. Diese Aufgabe kann durch den potenzialabhängigen Mg-Transport unter Nutzung des elektrischen Gradienten der PDa erfolgen. Mit Hilfe dieses Mechanismus ist eine Mg-Aufnahme sogar dann noch möglich, wenn die luminale Mg-Konzentration gleich oder geringfügig kleiner ist als in der Epithelzelle, in der die Mg-Konzentration 0.5–1.0 mmol/l beträgt. Die Mg-Aufnahme mit Hilfe des elektroneutralen Cotransportes wird allein durch die chemischen Gradienten der beteiligten Ionen Mg2+ und Cl– bestimmt. Hohe Mg-Konzentrationen im Pansen erleichtern daher die Mg-Absorption mit Hilfe dieses Transportmechanismus.
Beeinflussung der ruminalen Mg-Absorption: Kalium. Es ist seit vielen Jahrzehnten bekannt, dass Kalium die Mg-Verdaulichkeit durch Verringerung der Mg-Absorption aus dem Pansen herabsetzt (Literatur siehe Martens et al. 1987a). Dieser negative Effekt des Kaliums ist auf elektrophysiologische Veränderungen des Pansenepithels zurückzuführen, die mit Hilfe der Abb. 2 erklärt werden können. Die Erhöhung der K-Aufnahme verursacht eine Zunahme der ruminalen K-Konzentration, die wieder-um die Diffusion von K aus der Epithelzelle (siehe Abb. 2) verringert mit der Konsequenz, dass PDa depolarisiert wird und damit die Triebkraft für apikale Mg-Aufnahme mit Hilfe des potenzialabhängigen Mg-Transports abnimmt. Diese durch in vivo und in vitro Versuche bestätigte Abnahme des Mg-Transportes durch Erhöhung der ruminalen K-Konzentration beschränkt sich auf den PD-abhängigen oder K-sensitiven Mg-Transport (siehe oben). Das bedeutet, dass mit hohen K-Aufnahmen eine maxi-male (negative) Wirkung auf den Mg-Transport bewirkt wird, die durch weitere Erhöhungen der K-Konzentration nicht mehr gesteigert werden kann, weil einerseits der PD-abhängige Mg-Transport weitgehend wegen der fehlenden Triebkraft gehemmt wird und andererseits der PD-unabhängige Mg-Transport (K-insensitiv) weiterhin eine Mg-Absorption ermöglicht. Dieser Zusammenhang wurde schon sehr früh von Fontenot und Mitarbeitern erkannt, die feststellten, dass beim Schaf die Mg-Absorption abnimmt, wenn der K-Gehalt im Futter von 1 – 3 % in der Trockenmasse erhöht wird. Höhere K-Gehalte verstärken die negativen Wirkungen auf die Mg-Absorption nicht (siehe Rahnema und Fonte-not 1986; Martens et al. 1988). Die verbleibende Mg-Absorption erfolgt über den PD-unabhängigen oder K-insensitiven Mg-Transport, der mit steigenden Mg-Aufnahmen zunimmt. Die Zusammenhänge zwischen der oralen K-Aufnahme und der Mg-Verdaulichkeit bei unterschiedlichen Mg-Aufnahmen haben Ram et al. (1997) an Schafen untersucht (Tab. 1).
Tabelle 1: Effekt von Kalium (1 % versus 3.6 % in der Trockenmasse) auf die Mg-Verdaulichkeit von Schafen bei steigenden Mg-Aufnahmen. In Klammern: Scheinbare Verdaulichkeit in %. Versuchergeb-nisse Ram et al. 1998.
Mg-Aufnahme
(g/d)
Mg-Absorption (g/d)
K 1 % K 3.6 %
Abnahme
Mg-Abs. (g/d)
Relative
Veränderung
1.64 0.58 (35) 0.27 (16) 0.31 54
3.14 1.17 (37) 0.81 (26) 0.36 31
4.66 1.56 (33) 1.14 (24) 0.42 27

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
Seite 16 8. BOKU-Symposium Tierernährung 2009
Es ist erkennbar, dass die Abnahme der Mg-Absorption durch den erhöhten K-Gehalt (3.6 %) bei allen Mg-Aufnahmen etwa die gleiche Größenordnung von 0.31 – 0.42 g/d hat. Hierbei handelt es sich um die Verringerung des PD-abhängigen oder K-sensitiven Mg-Transports. Es ist ferner zu betonen, dass bei den geringen Mg-Aufnahmen die Mg-Absorption um 54 % verringert wird, während bei den hohen Mg-Aufnahmen die relative Abnahme nur 27 % beträgt. Diese Befunde bestätigen das vorgeschlagene Transportmodell für Mg (siehe Abb. 2), das annimmt, dass bei niedrigen Mg-Aufnahmen und dann niedrigen Mg-Konzentrationen in der Pansenflüssigkeit Mg primär über den PD-abhängigen Mg-Transport absorbiert wird, der K-sensitiv ist. Infolgedessen ergeben sich erhebliche Abnahmen der Mg-Verdaulichkeit bei einer Kombination von niedrigen Mg- mit hohen K-Aufnahmen. Entsprechende Ergebnisse wurden auch in Versuchen mit nicht laktierenden Kühen erzielt. Jittakhot et al. (2004) beobachteten bei Kühen mit einer niedrigen Mg-Aufnahme (40 – 41 g/d) eine Abnahme der scheinba-ren Verdaulichkeit von 12.6 % (K-Gehalt 2.1 %) auf nur 2 % (K-Gehalt 7.5 %). Bei hohen Mg-Aufnahmen (69 g/d) ergab sich eine scheinbare Verdaulichkeit von 18.6 % (K-Gehalt 2.1 %) und 7 % (K-Gehalt 7.5 %). Widersprüchliche Ergebnisse liegen jedoch auch vor. So konnten Holtenius et al. (2008) die scheinbare Verdaulichkeit von Mg bei laktierenden Kühen durch steigende K-Aufnahmen nicht negativ beeinflussen.
Als Quintessenz lässt sich festhalten, dass der negative Effekt von K auf die Mg-Verdaulichkeit nicht konstant ist und bei einer Abschätzung der Mg-Verdaulichkeit daher die Mg- und die K-Aufnahme Be-rücksichtigung finden müssen. In einer Metaanalyse über die Mg-Verdaulichkeit bei Kühen hat Weiss (2004) diese Voraussetzungen einbezogen und ist zu folgenden Schlussfolgerungen gekommen. Für die Verdaulichkeit von Mg ergab sich aufgrund dieser Metaanalyse folgende Gleichung (3)
Mg Absorption = 4.5 (± 4.0) + 0.24 (±0.07) x Mg-Aufnahme – 4.4 (± 2.2) x K (3) Mg Absorption und Mg-Aufnahme in g/d und K-Gehalt in % of TM
Diese Gleichung ergibt eine Mg-Verdaulichkeit von etwa 24 % bei einem K-Gehalt von 1 % in der TM im Erhaltungsbedarf und auch bei steigender Milchproduktion und folglich höheren Mg-Aufnahmen (siehe Tabelle 2; Angaben in Klammern = scheinbare Verdaulichkeit in %). Wie in dem Beispiel der Tab. 1 zeigt sich auch hier, dass die negativen Effekte erhöhter K-Aufnahmen bei niedrigen Mg-Aufnahmen wie z. B. im Erhaltungsbedarf ganz ausgeprägt sind. Hier ergibt sich eine Abnahme der scheinbaren Mg-Verdaulichkeit von 24 % bei einem K-Gehalt von 1 % auf 5.8 %, wenn der K-Gehalt 3 % beträgt (Tab. 2). Vice versa, die negative Wirkung von K fällt bei höheren Mg-Aufnahmen geringer aus (siehe Tab. 2).
Tabelle 2: K Gehalt der Diät und dessen Effekt auf die notwendige Mg-Aufnahme für eine Kuh mit einem Körpergewicht von 700 kg mit steigender Milchproduktion. Es wurde ein Mg-Bedarf von 2.8 g/d für die Erhaltung und von 0.15 g Mg/l Milch angenommen. Die Berechnung erfolgte mit Hilfe der Glei-chung (3) (Weiss 2004)*. Angaben in Klammern: Scheinbare Verdaulichkeit in Prozent.
Milchmenge kg/d
Mg Milch g/d
Mg-Bedarf** g/d
Notwendige Mg-Aufnahme (g/d) 1 % K 2 % K 3 % K
0 0 2.8 11.3 (24) 29.6 (9.5) 47.9 (5.8) 20 3.0 5.8 23.8 (24) 42.1 (14) 60.4 (9.6) 40 6.0 8.8 36.3 (24) 54.6 (16) 72.9 (12) 60 9.0 11.8 48.8 (24) 67.1 (18) 85.4 (14)
* Es muss betont werden, dass die Gleichung (3) abgeleitet wurde mit Hilfe von Versuchsdaten aus Fütterungsversuchen, in denen der K-Gehalt zwischen 1 – 3 % variierte. Die Gleichung (3) sollte daher nicht für höhere K-Gehalte verwendet werden. ** Mg-Erhaltungsbedarf. Der Gesamtbedarf ergibt sich aus dem Erhaltungsbedarf (2.8 g/d) plus der Mg-Menge in der Milch.

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
8. BOKU-Symposium Tierernährung 2009 Seite 17
Aus diesen Befunden lässt sich die Schlussfolgerung ableiten, dass das Risiko einer Hypomagnesämie immer dann groß ist, wenn bei geringen Mg-Aufnahmen der K-Gehalt des Futters und die Milchpro-duktion hoch sind: Kombination von reduzierter Absorption mit erhöhtem Bedarf.
Mit der Problematik der negativen Beeinflussung der Mg-Absorption bei Milchkühen hat sich in Europa die Arbeitsgruppe Schonewille in Utrecht intensiv befasst. Aufgrund der Untersuchungen dieser Auto-ren wird in den Niederlanden folgende Gleichung für die Ermittlung des Mg-Bedarfs genutzt (Literatur siehe Schonewille et al. 2008).
Mg-Absorption (%) = 34.9 – 0.45xK (g/kg) (4)
Diese Gleichung besagt, dass bei einem K-Gehalt von 1 % in der TM (10 g K/kg) die Mg-Absorption etwa 30 % beträgt. Dieser Wert ist etwas höher als die mit Hilfe der Gleichung (3) ermittelte Mg-Absorption (siehe auch Tab. 3). Berechnet man nun mit Hilfe der Gleichung (4) die notwendigen Mg-Aufnahmen mit den Rahmendaten der Tab. 2 ergeben sich folgende Werte.
Tabelle 3: K Gehalt der Diät und dessen Effekt auf die notwendige Mg-Aufnahme für eine Kuh mit einem Körpergewicht von 700 kg mit steigender Milchproduktion. Es wurde ein Mg-Bedarf 2.8 g/d für die Erhaltung und von 0.15 g Mg/l Milch angenommen. Die Berechnung erfolgte mit Hilfe der Glei-chung (4). Angaben in Klammern: Scheinbare Verdaulichkeit in Prozent.
Milchmenge kg/d
Mg Milch g/d
Mg-Bedarf* g/d
Notwendige Mg-Aufnahme (g/d) 1 % K 2 % K 3 % K
0 0 2.8 9.32 (30) 10.8 (26) 13.1 (21) 20 3.0 5.8 19.3 (30) 22.4 (26) 27.1 (21) 40 6.0 8.8 29.3 (30) 34.0 (26) 41.1 (21) 60 9.0 11.8 39.3 (30) 45.6 (26) 55.1 (21)
* Mg-Erhaltungsbedarf. Der Gesamtbedarf ergibt sich aus dem Erhaltungsbedarf (2.8 g/d) plus der Mg-Menge in der Milch.
Da die Versuchsdaten, die der Gleichung 4 zugrunde liegen, große Schwankungen bezüglich der Mg-Verdaulichkeit der Kühe aufwiesen, wird empfohlen die Werte der Tabelle 4 mit einem Sicherheitszu-schlag von 1.6 zu versehen (Schonewille, persönliche Mitteilung), d. h. im Erhaltungsbedarf (1 % K) müsste die Aufnahme z. B. nicht 9.32 sondern 14.9 g/d betragen. Entsprechend nähern sich mit die-sem Sicherheitszuschlag die Werte der Tab. 2 und 3 an.
Beeinflussung der ruminalen Mg-Absorption: Na-Mangel. Es ist seit vielen Jahren bekannt, dass bei einem Na-Mangel der Na-Gehalt des Speichels abnimmt und in gleichem Ausmaß (mmol/mmol) die Kaliumkonzentration ansteigt (Denton 1956). Die hohen Speichelsekretionsraten verursachen dann infolge der veränderten Speichelzusammensetzung einen Anstieg der ruminalen K-Konzentration mit der Konsequenz der erläuterten elektrophysiologischen Veränderungen des Panse-nepithels und des PD-abhängigen (K-sensitiven) Mg-Transportes (Martens et al. 1987a). In der Tabel-le 4 sind die entsprechenden Versuchsergebnisse von Schafen zusammengestellt worden. Es ist er-kennbar, dass die Veränderungen infolge des Na Mangels zu der erwarteten Beeinflussung der rele-vanten Parameter geführt hat (Tab. 4). Ein Na-Mangel und damit das Risiko der Hypomagnesämie sind häufig gegeben bei ausschließlicher Weidehaltung ohne Zufütterung von Kraftfutter oder ohne Angebot von Lecksteinen. Infolgedessen ist ein Na-Mangel bei Weidehaltung der weiterhin am häu-figsten übersehene Risikofaktor.

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
Seite 18 8. BOKU-Symposium Tierernährung 2009
Tabelle 4: Effekt eines Na-Mangels auf Parameter des Mg-Stoffwechsels bei Schafen. Modifiziert von Martens et al. 1987b.
Parameter Kontrolle Na-Mangel Mg-Aufnahme (g/d) 2.24 2.24 Absorption (g/d) Ges. Verdauungstrakt Vormägen
0.78 1.00
0.56 0.70
Ruminale K-Konzentr. (mmol/l) 54.3 112.2 PDt (mV) 38.0 55.0 Mg Urin (g/d) 0.44 0.23 Scheinb. Verdaulichkeit (%) 34.6 24.9
Beeinflussung der ruminalen Mg-Absorption: N-Aufnahme. Untersuchungen über mögliche Wechselwirkungen zwischen der N-Aufnahme und dem Auftreten von Hypomagnesämien/Tetanien zeichnen sich durch Widersprüchlichkeit aus. Feldbeobachtungen haben immer wieder zu der Vermu-tung geführt, dass z. B. der hohe Rohproteingehalt des Grases das Auftreten von Hypomagnesämien begünstigt. Entsprechende Bilanzversuche der Arbeitsgruppe Fontenot et al. (1973) haben aber nie eine negative Wirkung der N-Aufnahme auf die Mg-Absorption aufzeigen können. Im Gegensatz hierzu wurde im akuten Versuch mit dem Modell des vorübergehend isolierten Pansens sowohl beim Schaf (Martens und Rayssiguier, 1980) als auch beim jungen Rind (Martens et al. 1988) gezeigt, dass eine Erhöhung der ruminalen NH4
+ -Konzentration eine Verringerung der Mg-Absorption aus dem Pansen verursacht. Dieser scheinbare Widerspruch wurde aufgelöst durch einen Versuchsansatz, der als Hypothese annahm, dass die negative Wirkung von NH4
+ auf die Mg-Absorption ein akuter Effekt ist, der infolge von Adaptationsvorgängen im Pansenepithel aufgehoben wird. Gäbel und Martens (1986) haben in einem entsprechenden Versuch mit Schafen diese Hypothese geprüft, in dem die ruminale NH4
+ Konzentration durch eine intraruminale Harnstoffinfusion akut erhöht wurde. Die renale Mg-Ausscheidung als möglicher Parameter der ruminalen Mg-Absorption wurde vor und nach der Erhö-hung der Ammoniakkonzentration bestimmt. Es zeigte sich, dass die renale Mg-Ausscheidung nur 2- 3 Tage nach Beginn der Erhöhung der ruminalen Ammoniakkonzentration verringert war und dann wie-der der Ausscheidungsmenge entsprach, die vor der Erhöhung der ruminalen Ammoniakkonzentration beobachtet wurde. Die Arbeitshypothese einer akuten Ammoniakwirkung wurde also bestätigt. Über die Mechanismen der Adaptation des Pansenepithels an hohe Ammoniakkonzentrationen, die zur Normalisierung der Mg-Absorption führen, kann keine Aussage getroffen werden.
Renale Ausscheidung von Mg: Der renalen Ausscheidung von Mg kommt eine überragende Bedeu-tung im Hinblick auf die Regulation des Mg-Haushalts zu (siehe auch Abb. 3 und Gleichung 1 und 2) und ist als einzige Regelgröße des Mg-Haushalts anzusehen. Wie die Abb. 3 zeigt, verläuft die Aus-scheidung von Mg mit dem Urin kurvilinear und wird außerordentlich gering bei einer Blutmagnesium-konzentration unterhalb der physiologischen Schwankungsbreite von < 0.75 mmol/l. Eine ausreichen-de Mg-Versorgung wird angenommen, wenn die renale Mg-Ausscheidung > 2.5 g/d bei Kühen beträgt (Kemp and Geurink 1978; Mayland 1988).

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
8. BOKU-Symposium Tierernährung 2009 Seite 19
Abbildung 3: Beziehung zwischen der Mg-Konzentration im Blut und der renalen Mg-Ausscheidung von Kühen. Die Mg-Ausscheidung nähert sich asymptotisch fast Null bei Mg-Konzentration im Blut, die als hypomagnesämisch (< 0.75 mmol/l) anzusehen sind. (von Schonewille et al. 2000 mit Erlaubnis).
Der Parameter „tägliche Mg-Ausscheidung im Urin“ ist in der Praxis nicht bestimmbar. Die Mg-Konzentration im Harn kann jedoch weiterhelfen. Diese Erkenntnis hat dazu geführt, dass schon vor vielen Jahren die Mg-Konzentration im Harn als sicherer Indikator der Mg-Versorgung angesehen wur-de (Kemp 1983; Tab. 5): Eine ausreichende Mg-Versorgung liegt vor bei Mg-Konzentrationen > 4 mmol/l, die Mg-Aufnahme wird als nicht ausreichend angesehen bei Mg-Konzentrationen von 0.9 – 4.0 mmol/l und eine gravierende Unterversorgung mit dem Risiko von Tetanien ist anzunehmen bei Mg-Konzentrationen < 0.9 mmol/l.
Tabelle 5: Mg-Konzentration im Harn als Indikator der Mg-Versorgung (Kemp 1983).
Mg-Konzentration mg/l
Mg-Konzentration Mmol/l
Bewertung
> 100 > 4.40 Ausreichende Mg Versorgung 20 - 100 0.87 – 4.40 Marginal; u. U. nicht ausreichend
< 20 0.87 Gravierende Unterversorgung; Tetanierisiko
Die Abb. 3 zeigt, dass die Niere das zuviel absorbierte Mg schnell und regulativ ausscheidet. Spekula-tiv kann man daher annehmen, dass in der Evolution offensichtlich immer reichlich Mg verfügbar war, sodass primär die renale Ausscheidung von zuviel absorbiertem Mg zur Regulation erfolgte. Die Regu-lation einer Unterversorgung war offensichtlich nicht erforderlich, die dann aber zum Risiko wird,

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
Seite 20 8. BOKU-Symposium Tierernährung 2009
wenn die ursprünglichen Bedingungen nicht mehr gegeben sind. Hierbei handelt es sich um die Ver-änderung der Fütterung (hohe K-Aufnahmen) und vor allem um den erhöhten Bedarf infolge der Milchproduktion.
Diagnose, Prophylaxe und Behandlung: Eine Diagnose wird immer dann erleichtert, wenn klini-sche Symptome wie Zähneknirschen, Muskeltremor oder unsicherer Gang auftreten. Eine Objektivie-rung kann dann durch die Bestimmung der Blutmagnesiumkonzentration (< 0.7 – 0.8 mmol/l) erfol-gen. Unsicherheiten sind jedoch immer dann gegeben, wenn niedrige Blutmagnesiumkonzentrationen (0.75 – 0.9 mmol/l) ohne eindeutige klinische Symptome auftreten und der Verdacht einer unzurei-chenden Mg-Versorgung besteht. In diesen Fällen sollte die Bestimmung der Mg-Konzentration im Harn (siehe oben) zusätzlich erfolgen.
Eine erfolgreiche Prophylaxe der Hypomagnesämie des Rindes ist aufgrund der erweiterten Kenntnisse über mögliche Störungen der Mg-Absorption und aufgrund der gut gesicherten Bedarfsnormen leicht und wirksam zu verwirklichen. In der Regel lässt sich der K-Gehalt der Diät nicht beeinflussen. Aus diesem Grunde sind entsprechend erhöhte Mg-Aufnahmen notwendig, die sich leicht realisieren las-sen, wenn zur Sicherung der Milchproduktion Kraftfutter mit den erforderlichen Mg-Gehalten verfüttert werden. Im Hinblick auf eine mögliche Na-Unterversorgung ist in gleicher Weise zu verfahren. Die transiente negative Wirkung hoher Stickstoffaufnahmen wird nur wirksam bei plötzlicher Futterumstel-lung, die durch einen langsamen Futterwechsel vermieden werden kann. Somit lässt sich feststellen, dass bei Milchkühen eine mögliche Gefährdung durch eine Hypomagnesämie/Tetanie bei den heute üblichen Fütterungsbedingungen (Kombination aus Grundfutter plus leistungsgerechte Kraftfutterzula-gen) nachhaltig vermieden werden kann.
Wenn jedoch diese Bedingungen nicht gegeben sind, kann es zu vermehrten Fällen von Hypomagne-sämie/Tetanien kommen, wie es z. B. in England der Fall war, als infolge der Einführung der Milch-quote die Kraftfutterzulagen reduziert wurden und damit die „Prophylaxe“ durch die Konzentratfütte-rung ausblieb (Whitaker et al. 1985). Ferner ist eine Gefährdung bei überwiegender Weidehaltung und Grasfütterung gegeben. Eine Supplementation von Mg mit Hilfe von Lecksteinen ist nur bedingt wirk-sam, weil Kühe auch im Mg-Mangel entsprechende Angebote nur unzureichend wahrnehmen.
Die Behandlung klinisch erkrankter Tiere erfolgt durch intravenöse Infusion von Mg-haltigen Lösun-gen. Eine „Erste Hilfe“ kann auch durch Laien erfolgen, in dem rektal 60 g MgCl2∙6H2O in 200 ml Wasser appliziert wird. Unter diesen Bedingungen kann die Absorptionsfähigkeit des Colons/Rektums für Mg genutzt werden, die unter physiologischen Bedingungen infolge der geringen Mg-Löslichkeit im Lumen keine Bedeutung hat.
Magnesium und Milchfieber Eine Hypomagnesämie beeinträchtigt die Regulation der Ca-Konzentration im Blut, weil die Freiset-zung und Synthese von Parathormon (PTH) sowie die Bindung von PTH an seinen Rezeptor Mg erfor-dert (Goff 2008). Infolgedessen wird die Signalkaskade der Regulation des Ca-Haushalts wiederholt gestört. Diese zunächst nur bei Versuchstieren erarbeiteten Zusammenhänge treffen offensichtlich auch für die Kuh zu, so dass eine unzureichende Mg-Versorgung die Ca-Mobilisation aus dem Knochen erschwert und das Risiko der Gebärparese erhöht. Entsprechende Berichte liegen in der Literatur vor (Sampson et al. 1983). Eine unzureichende Mg-Versorgung ist jedoch häufig schwer zu objektiveren, weil die Mg-Konzentration im Blut auch dann noch im unteren „physiologischen“ Bereich gehalten wird, wenn die Mg-Versorgung schon unzureichend ist (siehe oben). Ein wertvoller Indikator für die Mg-Versorgung ist der Verlauf der Mg-Konzentration im Blut während der Geburt. Der Abfall der Ca-Konzentration im Blut erhöht die PTH Konzentration im Blut. Da PTH in der Niere die Ausscheidung von Mg verringert, ergibt sich bei ausreichender Mg-Versorgung ein Anstieg der Mg-Konzentration bei gleichzeitiger Verringerung der Ca-Konzentration im Blut. Wenn dieser typische physiologische Anstieg der Mg-Konzentration nicht beobachtet wird, kann von einer Unterversorgung ausgegangen werden.

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
8. BOKU-Symposium Tierernährung 2009 Seite 21
Die Bedeutung einer ausreichenden Mg-Versorgung zur Prophylaxe der Gebärparese geht aus einer Metaanalyse von Lean et al. (2006) hervor, die zu der Schlussfolgerung führte, dass Mg-Gehalte von 0.3 – 0.4 % in der TM prophylaktisch wirksam sind. Diese Mg-Gehalte liegen erheblich über den zur Bedarfsdeckung empfohlenen Wert von 0.2 %. Eine mögliche Erklärung könnte die Reduzierung der Futteraufnahme vor der Geburt sein, so dass die hohen Mg-Gehalte eine ausreichende Mg-Aufnahme gewährleisten.
Zusammenfassung und Perspektive Magnesium ist ein essentieller Mineralstoff ohne hormonales Regulationssystem mit vielfältigen Funk-tionen als Cofaktor zahlreicher Enzyme, als Regulator von Ionenkanälen und Modulator bei der synap-tischen Erregungsübertragung. Es überrascht daher nicht, dass ein Mg-Mangel mit vielen und zu-nächst unspezifischen Symptomen wie Appetitlosigkeit, unsicherer Gang und Absonderung aus dem Herden/Gruppenverband einhergeht. Ausgeprägte Hypomagnesämien (< 0.7 mmol/l) führen letztend-lich zum Verlust des Stehvermögens und zu Tetanien, die wiederum tödlich enden können. Hypo-magnesämien werden immer dann beobachtet, wenn die Regulation der normalen Blutmagnesium-konzentration nicht aufrechterhalten wird. Da ein hormonelles Regulationssystem nicht existiert, ba-siert die Aufrechterhaltung der Blutmagnesiumkonzentration auf der aus dem Magen-Darm-Kanal absorbierten (a) Magnesiummenge einerseits und der Bedarfsdeckung für (b) Wachstum, (c) Milchleis-tung, (d) die Versorgung des Fetus und (e) endogene Verluste in den Verdauungskanal andererseits. Die Blutmagnesiumkonzentration wird im physiologischen Bereich gehalten, wenn (a) > (b) + (c) + (d) + (e) und somit die Differenz zwischen (a) – [(b) + (c) + (d) + (e)] > 0 ist. Diese über dem Be-darf zur Verfügung stehende Magnesiummenge wird renal ausgeschieden. Umgekehrt ergeben sich Hypomagnesämien, wenn die orale Magnesiumaufnahme nicht bedarfsdeckend ist, die Magnesiumab-sorption aus dem Verdauungskanal gestört wird oder wenn der Bedarf (b, c, d oder e) erhöht ist. Un-ter diesen Bedingungen sinkt die renale Magnesiumausscheidung und die Mg-Konzentration im Harn ist < 1mmol/l.
Bei landwirtschaftlichen Nutztieren werden Hypomagnesämien und Tetanien fast ausschließlich bei Wiederkäuern beobachtet, die in der Regel auf eine Kombination von gestörter Mg-Absorption mit einem erhöhten Mg-Bedarf für die Milchsekretion zurückzuführen sind. Die in den letzten Jahrzehnten erarbeiteten Erkenntnisse über die Mg-Absorption aus den Vormägen und dessen Beeinflussung durch Futterinhaltsstoffe erlauben eine bedarfsgerechte Abschätzung der notwendigen Mg-Aufnahme, so dass eine effektive Prophylaxe der Hypomagnesämie der Wiederkäuer möglich ist bzw. Risiken aufge-zeigt werden können.
Die mögliche Bedeutung einer ausreichenden peripartalen Mg-Versorgung zur Prophylaxe der Gebär-parese wird zur Diskussion gestellt und erfordert weitere Untersuchungen.
Danksagung
Die Untersuchungen der Autoren sind viele Jahre von der Deutschen Forschungsgemeinschaft unterstützt worden. Darüber hinaus erfolgte eine Unterstützung durch Stipendien des DAAD, der Alexander von Humboldt Stiftung, der H.-W. Schaumann Stiftung und der Margaret-Markus-Charity.
Literatur
Denton D.A. (1956): The effect of Na+-depletion on the Na+:K+ ratio of the parotid saliva of the sheep. J. Physiol. 131, 516-525
Donovan G. A., Steenholdt C., McGehee K., and Lundquist R. (2004): Hypomagnesemia among cows in a confinement-house dairy system. J. Am. Vet. Med. Ass. 224, 96-99.

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
Seite 22 8. BOKU-Symposium Tierernährung 2009
Fontenot J.P., Wise M. B., and Webb, K. E. (1973): Interrelationships of potassium, nitrogen, and magnesium in ruminants. Fed. Proc. 32, 1925-1928.
Gäbel, G., and Martens H. (1986): The effect of ammonia on magnesium metabolism in sheep. J. Animal. Phys. Animal. Nutr. 55, 278-287
Goff J. (2008): The monitoring, prevention, and treatment of milk fever and subclinical hypocalcemia in dairy cows. Vet. J. 176, 50-57.
Günther T. (1977): Stoffwechsel und Wirkungen des intrazellulären Magnesiums. J. Clin. Chem. a. Clin. Biochem. 15, 433-438
Holtenius K., Kronquist C., Briland E., and Spörndly R. (2008): Magnesium absorption by lactating cows on grass silage-based diet supplied with different potassium and magnesium levels. J. Dairy Sci. 91, 743-748
Jittakhot S., Schonewille J.T., Wouterse H., Yuangklang C., and Beynen A.C. (2004): Apparent magnesium absorption in dry cows fed at 3 levels of potassium and 2 levels of magnesium intake. J. Dairy Sci. 87, 379-385.
Kemp A. (1983): The effect of fertilizer treatment of grassland on the biological availability of magnesium to ruminants. In: J.P. Fontenot, G.E. Bunce, K.E. Webb, V. Allen (eds), Proc. of John Lee Pratt international symposium on the role of magnesium in animal nutrition. Virginia Polytechnic Institute and State University Blacksburg, Virginia, 143-157.
Kemp A., and Geurink, JH. (1978): Granssland farming and minerals in cattle. Neth. J. Agric. Sci. 26, 161- 175
Lean I.J., DeGaris P.J., McNeil D.M., and Block E. (2006): Hypocalcemia in dairy cows: Meta analysis and dietary cation anion difference theory revisited. J. Dairy Sci. 89, 669-684.
Leonhard-Marek , and Martens H. (1996): Effects of potassium on magnesium transport across rumen epithelium. Am. J. Physiol. 271, G1034-1036.
Leonhard-Marek S., Stumpff F., Brinkmann I., Breves G., and Martens H. (2005): Basolateral Mg2+/Na+ exchange regulates apical nonselective cation channel in sheep rumen epithelium via cytosolic Mg2+. Am. J. Physiol. 288, 630-645.
H. Martens H., and Rayssiguier Y. (1980) Magnesium metabolism and hypomagnesaemia .MTP Press. In: Digestive Physiology and Metabolism in Ruminants Clermont-Ferrand. P. 447-466
Martens H., and Schweigel M. (2000): Grass Tetany and other Hypomagnesaemias. In: Herdt, T. (eds) Veterinary Clinics of North America. Food Animal Practice: Metabolic Disorders of Ruminants. 16, 339-368. W. B. Saunders - Philadelphia
Martens H., Heggemann G., and Regier K. (1988): Studies about the absorption of magnesium from the temporarily isolated rumen of heifers. Effect of K, Na, NH4, VFA and CO2. Zbl. Vet. Med. A 35, 73-80.
Martens H., Gäbel G., Strozyk H. (1987a): The effect of potassium and the transmural potenzial difference on the magnesium transport across an isolated preparation of sheep rumen epithelium. Q. J. Exp. Physiol. 72, 181-188
Martens H., Kubel O.W., Gäbel G., and Honig H. (1987b): Effects of low sodium intake on magnesium metabolism in sheep. J. agric. Sci. 108, 237-243
Mayland HF. (1988) Gras Tetany. In: Church DC (ed): The ruminant animal. Digestive Physiology and Nutrition. New Jersey, Resten Book. P. 511-523
McCoy A. A., Goodall E. A., and Kennedy D. G. (1996): Incidence of bovine hypomagnesaemia in Northern Ireland and methods of magnesium supplementation. Vet. Rec. 13, 41-43.
Meyer H., and Scholz H. (1972): Untersuchungen über die Pathogenese der hypomagnesaemischen Tetanie: Beziehungen zischen dem Mg-Gehalt im Blut und im Liquor cerebrospinalis beim Schaf. Dtsch. Tierärztl. Wochenschr. 79: 55-61
Persson J., and Luhman J. (1974): The effect of insulin, glucose and catecholamines on some blood minerals in sheep. Acta Vet. Scand. 15: 519-564.
Rahnema S.H., and J. Fontenot (1986): Effect of potassium on association of minerals with various fractions of digesta and feces of sheep fed hay. J. Anim. Sci. 63, 1491-1501
Ram L., Schoneville, J. Th., Van‘t Klooster, A. Th., Martens, H., and Beynen, A. C. (1998): The decrease in the amount of magnesium absorbed in weathers fed potassium bicarbonate is independent of the dietary magnesium concentration. J. Dairy Sci. 81, 2485-2492.
Rayssiguier Y.(1977): Hypomagnesemia resulting from adrenaline infusion in ewes: Ist relation to lipolysis. Horm. Metab. Res. 9: 309-316.
Sampson B.F., Manston R., and Vagg M.J. (1983): Magnesium and Milk fever. Vet. Rec. 112, 447-449.
Schonewille J., Everts H., Jittakhot S., and Beynen A.C. (2008): Quantitative prediction of magnesium absorption in dairy cows. J. Dairy Sci. 91, 271-278

Martens und Stumpff: Magnesium Stoffwechsel: Regulation und Risiken
8. BOKU-Symposium Tierernährung 2009 Seite 23
Schonewille J.Th., van’t Klooster, A.Th., Woutersee, H., and Beynen, A.C. (2000): Time cours of plasma magnesium concentrations and urinary magnesium excretion in cows subjected to acute changes of potassium intake. The Vet. Quart. 22, 136-140.
Schweigel M. and Martens H. (2003): Anion dependent Mg2+ influx and a role for a vacuolar H+-ATPase. Am. J. Physiol. 285, G45-G53.
Schweigel M. Lang I., and Martens H. (1999): Mg2+ transport in sheep rumen epithelium: evidence for an electrodiffusive up-take mechanism. Am. J. Physiol. 277, G976-G982.
Schweigel M., Vormann M., and Martens H. (2000): Mechanisms of Mg2+ transport in cultured epithelial cells. Am. J. Physiol. 278, G400-G408.
Stumpff, F. Martens, H.: A role for magnesium in the regulation of ruminal sodium transport. In: Focus on signal transduction research / Gavin McAlpine (Hrsg.) - Hauppauge NY, USA: Nova Science Publishers, 2006. - Chapter 2; ISBN: 1-60021-376-6) S. 37-66
Weiss W. P. (2004): Macromineral digestion by lactating cows: Factors affecting digestibility of magnesium. J. Dairy Sci. 24, 2167-2171.
Whitaker D. A., Kelly J. M., and Smith E. J. (1985): Hypomagnesaemia increases. Vet. Rec. 116, 451-452.
Autorenanschrift
Prof. Dr. Holger Martens Inst. f. Veterinär-Physiologie Freie Universität Berlin Oertzenweg 19b, 14163 Berlin E-Mail: [email protected]

Kienzle: Kationen-Anionen-Bilanz beim Schwein – Möglichkeiten, Risiken und Nebenwirkungen
Seite 24 8. BOKU-Symposium Tierernährung 2009
Kationen-Anionen-Bilanz beim Schwein – Möglichkeiten, Risi-ken und Nebenwirkungen
Ellen Kienzle
Lehrstuhl für Tierernährung und Diätetik, LMU München
Säuernde Rationen werden bei Sauen um den Zeitpunkt der Geburt zur nutritiven Prophylaxe des MMA-Syndroms eingesetzt. Als Wirkungsmechanismus werden eine Absenkung des Keimgehaltes im Harn bei Harn-pH-Werten unter 6 sowie indirekte Effekte wie eine vermehrte Wasseraufnahme auf-grund der Zulagen von Harnsäuerern und/oder indirekte Wirkungen auf die Responsivität der Pa-rathormonrezeptoren diskutiert (Jürgens 1991, Finkensiep 1993, Roux 2005). Roux (2005) fütterten eine acidierende Ration während der gesamten Laktation und berichteten, dass die so gefütterten Sauen während der auf diese Laktation folgenden Reproduktionsperiode mehr lebende Ferkel hatten. DeRouchey et al. (2003) beschreiben eine höhere Überlebensrate bei Ferkeln, deren Mütter in der aktuellen Laktation acidierend gefüttert wurden. Dove and Haydon (1994) fanden dagegen keinen derartigen Effekt, wobei ihre Rationen allerdings deutlich höhere Kationen-Anionen-Bilanzen aufwiesen und damit weniger stark säuernd waren als die Rationen in den anderen genannten Experimenten. Bei wachsenden Schweinen führten säuernde Rationen dagegen in der Mehrzahl der Untersuchungen zu einer Leistungsreduktion (Yen et al. 1981, Patience et al. 1987). Ein depressiver Effekt acidierender Rationen auf das Wachstum ist schon lange bekannt Langendorf (1963). Ein möglicher Grund ist der Aminosäurenverbrauch zur Bildung von puffernden Carbonationen (Foley et al. 1995)
Säuernde Rationen machen daher vor allem bei Sauen Sinn. Bei der praktischen Durchführung der Harnsäuerung bei Sauen steht die Frage der Dosierung der harnsäuernden Substanzen (Ammoni-umchlorid, Ca-Chlorid, Methionin, Phosphorsäure) im Mittelpunkt des Interesses. Zum einen sind diese Substanzen teuer, zum anderen ist der Geschmack meist unangenehm. Starke Überdosierung führt i.d.R. zu einer metabolischen Acidose und zur Einstellung der Futteraufnahme. Für den Harn-pH-Wert ist jedoch nicht nur die Menge der zusätzlich verabreichten Harnsäuerer entscheidend, sondern die Bilanz der insgesamt aufgenommenen säuernden und alkalisierenden Equivalente. Diese Bilanz wird als Kationen-Anionen-Bilanz (KAB) im Futter bezeichnet und ermöglicht eine näherungsweise Abschät-zung der Wirkung des Futters auf den Harn-pH-Wert (Krohn 1993, Beeker 1999). In der folgenden Formel sind das Atomgewicht und die Wertigkeit bereits berücksichtigt, die Gehalte im Futter werden in g/kg TS eingesetzt:
Kationen-Anionen-Bilanz (KAB, mmol/kg TS)=50*Ca+83*Mg+26*K+44*Na-59*P-62*S-28*Cl.
Statt Schwefel kann näherungsweise auch der Gehalt an Methionin und Cystin (mit Faktor -13) einge-setzt werden, wenn nicht größere Mengen an Sulfaten enthalten sind.
Der Begriff der KAB wird im englischen Schrifttum auch, sehr treffend, als "dietary undetermined ani-ons" bezeichnet, da mit dieser Berechnung im wesentlichen alkalisierende Anionen wie Carbonate und organische Säureanionen, die nicht durch acidierende Substanzen neutralisiert sind, erfasst werden. Die KAB ist nicht als physiologische Gesetzmäßigkeit zu betrachten, da einige Aspekte des Effektes des Futters auf den Säuren-Basen-Haushalt (u.a. unterschiedliche Absorptionsraten von An- und Kationen z.B. bei Ca-Chlorid) nicht erfasst werden. Als Orientierungshilfe um in der Praxis eine acidierende Ra-tion zusammenzustellen ist sie jedoch sehr gut geeignet. Der Harn-pH-Wert lässt sich anhand der folgenden quadratischen Gleichung aus der KAB abschätzen: pH=3*10-6KAB2 +0,003KAB + 6,19 (Krohn 1993, Beeker 1999).

Kienzle: Kationen-Anionen-Bilanz beim Schwein – Möglichkeiten, Risiken und Nebenwirkungen
8. BOKU-Symposium Tierernährung 2009 Seite 25
Eine mäßig acidierende Wirkung ist bei einer schwach negativen KAB zu erwarten. In der Praxis berei-tet die Berechnung der KAB und die Schätzung des Harn-pH-Wertes oft Schwierigkeiten. Deshalb wer-den einige Faustzahlen dargestellt. Eine nicht mineralisierte Ration aus Getreide und Sojaschrot weist i.d.R. eine KAB von -100 bis +100 mmol/kg TS auf, und liegt damit beinahe im gewünschten Bereich. Durch Zusatz von 3 % eines handelsüblichen Mineralfutters, das i.d.R. auch Ca-Karbonat enthält, steigt die KAB auf ca. +300 mmol/kg TS. Wird statt dessen der Ca-Bedarf der Sauen ausschließlich durch Ca-Carbonat (kohlensaurer Futterkalk) abgedeckt, so werden fast +500 mmol/kg TS erreicht. Der Verzicht auf Ca-Carbonat entweder direkt oder durch Mineralfutter zugesetzt reduziert die KAB daher erheblich. Nach anekdotischen Berichten aus der Praxis entsteht dadurch nicht das Risiko einer Hypokalzämie, eine Absicherung durch kontrollierte Untersuchungen besteht aber nicht. Statt Ca-Karbonat kann Ca-Chlorid eingesetzt werden. Zusatz von 1 % Ca-Chlorid deckt etwa die Hälfte des Ca-Bedarfs. Ca-Chlorid verringert die Akzeptanz, daher können größere Mengen problematisch sein. Besonders stark reduziert wird die Akzeptanz, wenn Ca-Chlorid in Verbindung mit Ammoniumchlorid und/oder Kochsalz eingesetzt wird, da die verschiedenen Chloride dann einen salzig-bitteren Ge-schmack erzeugen. Ammoniumchlorid, Methionin oder Phosphorsäure sind stark azidierend. 1 % Am-moniumchlorid oder 1 % Methionin senkt die KAB einer nicht mineralisierten Getreide-Soja-Ration um 200 mmol/kg TS. Damit ist bereits eine ausreichende Harnsäuerung gewährleistet. Bei Verzicht auf Mineralfutter genügen daher 1 % Ammoniumchlorid oder Methionin um einen Harn-pH-Wert um 6 zu erreichen. Ca-Chlorid besitzt einen leicht säurenden Effekt, welcher mit der obigen Formel nicht er-fasst wird. Daher muss beim Einsatz größerer Mengen an Ca-Chlorid eine Korrektur vorgenommen werden: Pro Gramm Ca-Chlorid im kg müssen nach Praxiserfahrungen 3,4 mmol von der KAB abge-zogen werden, i.a. der Zusatz von 1 % Ca-Chlorid senkt die KAB um 34 mmol.
Soll die säuernde Ration zusätzlich faserreich gestaltet werden, so muss darauf geachtet werden, dass über die faserreichen Futtermittel nicht alkalisiert wird. Vom Standpunkt des Säuren-Basen-Haushaltes sind Kleie und Biertreber gut geeignet, Trockenschnitzel und Grünmehl dagegen weniger.
Organische Säuren wie z.B. Zitronensäure wirken nicht zuverlässig harnsäuernd, da sie vollständig verstoffwechselt werden können, ohne dass Protonen frei werden. Eine säuernde Wirkung im Inter-mediärstoffwechsel und damit ein Effekt auf den Harn-pH-Wert kommt in diesem Fall nicht zustande. Die Alkali- oder Erdalkalisalze organischer Säuren wirken sogar alkalisierend, da bei ihrer Verstoff-wechselung Protonen aufgebraucht werden (Kienzle 1991). Phenolsäurederivate wie z.B. Benzoesäure werden dagegen unter Bildung organischer Säuren, vor allem Hippursäure, verstoffwechselt, die über die Niere ausgeschieden werden und dadurch zu einer Senkung des Harn-pH-Wertes führen können (Foley et al. 1995). Dies wurde für Benzoesäure auch beim Schwein gezeigt (Kluge et al. 2006). Hier wurden auch leistungsfördernde Effekte in der Mast beobachtet.
Der pH-Wert und der Ca-Gehalt im Trinkwasser haben keinen wesentlichen Einfluss auf den Harn-pH-Wert, da der mengenmäßige Gehalt an säuernden und alkalisierenden Substanzen im Verhältnis zum Futter sehr gering ist.
Eine langfristige acidierende Ernährung birgt erhebliche Risiken (Langendorf 1963), da sie zu Verände-rungen im Mineralstoffhaushalt führt. Bei Katzen und Pferden wurde gezeigt, dass der Blut-pH-Wert nicht streng mit der KAB korreliert, sondern vor allem von der Menge stark säuernder Substanzen, insbesondere des Ammoniumchlorids abhängt (Kienzle und Wilms-Eilers 1993, Lisa Berchtold##). Hinsichtlich der Veränderungen am Mineralstoffhaushalt infolge acidierender Fütterung ist beim Schwein insbesondere die reduzierte Verfügbarkeit von Phosphor zu nennen. Jambor und Prochazka (1977) zeigten, dass die Verfügbarkeit des Phosphors bei Mastschweinen von 92 auf 50 % zurück ging, wenn die Ration acidiert wurde. Dabei stieg sowohl die faecale als auch die renale Exkretion an. Derartige Effekte werden bei der Ableitung von Bedarfszahlen nicht berücksichtigt. Aus Gründen des Umweltschutzes müssen die Sicherheitsspannen beim Phosphorbedarf niedrig gehalten werden. Bei säuernden Rationen, die längerfristig eingesetzt werden sollen, sollte daher die Phosphorversorgung oberhalb der neueren Bedarfszahlen gehalten werden. Ein entsprechend höherer Eintrag in die Gülle

Kienzle: Kationen-Anionen-Bilanz beim Schwein – Möglichkeiten, Risiken und Nebenwirkungen
Seite 26 8. BOKU-Symposium Tierernährung 2009
ist dann allerdings nicht zu vermeiden. Phosphormangelsymptome beim Schwein im Kontext mit aci-dierender Ernährung wurden bereits 1932 von Marek und Wellmann beschrieben. Es ist eine Bein-schwäche mit auffälliger Säbelbeinigkeit zu erwarten. Bei anderen Species (Hund, Katze) wurde auffäl-lige Durchtrittigkeit beschrieben (Kienzle et al. 1998; Dobenecker und Kienzle 2008), die sich inner-halb weniger Stunden oder Tage entwickelte und nicht schmerzhaft war. Auch reduzierte Futterauf-nahme, Schwäche oder Anämie können Folgen eines P-Mangels sein. Plasmaphosphorgehalte zeigen einen Mangel leider nicht zuverlässig an. Bei Katzen wurde eine interstitielle Nephritis beschrieben, wenn bei chronischer acidierender Ernährung die Kaliumzufuhr knapp war (DiBartola et al. 1993).
Literatur
Beeker, S. (1999): Einstellung des Harn-pH-Wertes bei Sauen. LMU München, Vet. Diss.
Berchtold, L. (2009): Untersuchungen zum Einfluss der Anionen-Kationen-Bilanz auf den Mineralstoff- und Säure-Basen-Haushalt bei Ponys. LMU München, Vet. Diss
DeRouchey, J. M., J. D. Hancock, R. H. Hines, K. R. Cummings, D. J. Lee, C. A. Maloney, D.W. Dean, J. S. Parks, Cao, H. (2003): Effects of dietary electrolyte balance on the chemistry of blood and urine in lactating sows and sow litter performance. J. Anim. Sci. 81:3067-3074.
DiBartola, S. P., Buffington, C. A., Chew, D. J., McLoughlin, M. A., Sparks, R. A. (1993): Development of chronic renal disease in cats fed a commercial diet. J. Am. Vet. Med. Assoc. 202: 744-751.
Dobenecker, B., Kienzle, E. (2008): Normal calcium and low phosphorus intake in puppies leads to extremely bowed legs which are reversible by phosphorus repletion. ESVCN Conference Wien, Abstract, p. 26.
Dove, C. R., Haydon, K.D. (1994): The effect of various diet nutrient densities and electrolyte balances on sow and litter performance during two seasons of the year. J. Anim. Sci. 72:1101-1106.
Finkensiep, A. (1993): Untersuchungen über den Einfluß des Harnstatus auf das Puerperalsyndrom der Sau unter Berücksichtigung der Fütterung und des peripartalen Trinkwasserverbrauchs. Hannover, Tierärztliche Hochschule, Diss.
Foley, W.J., McLean, S., Cork, S.J. (1995): Consequences of biotransformation of plant secondary metabolites on acid–base metabolism in mammals. A final common pathway. J. Chem. Ecol. 21, 721–743.
Jambor, V., Procházka, Z. (1977): Effect of various anion and cation concentrations in the diet on the utilization of calcium and phosphorus in swine. Arch Tierernährg. 27: 701-709.
Jürgens, C. (1991): Einfluß von Futterzusatzstoffen auf den pH-Wert und die Inhaltsstoffe des Harns bei Zuchtsauen. Hannover, Tierärztliche Hochschule, Diss.
Kienzle, E. (1991): Ernährung und Urolithiasis bei Haussäugetieren. Übers. Tierernährg. 19: 157-200.
Kienzle, E., Wilms-Eilers, S. (1993): Untersuchungen zur Struvitsteindiätetik: 2. Einfluß von Ammoniumchlorid und Carbonaten auf den Säuren-Basen- und Mineralstoffhaushalt der Katze. Deutsche Tierärztliche Wochenschrift 100: 399-405.
Kienzle, E., Pessinger, C., Thielen, C. (1998): Phosphorous requirements of adult cats. Journal of Animal Physiology and Animal Nutrition 80: 90-100
Kluge, H., Broz, J., Eder, K. (2006): Effect of benzoic acid on growth performance, nutrient digestibility, nitrogen balance, gastrointestinal microflora and parameters of microbial metabolism in piglets. Journal of Animal Physiology and Animal Nutrition 90: 316–324
Krohn, U. (1993): Beeinflussung des Säure-Basen-Haushalts von Zuchtsauen durch Futterzusätze. Hannover, Tierärztliche Hochschule, Diss.
Langendorf, H. (1963): Säurebasengleichgewicht und chronische acidogene und alkalogene Ernährung. Zeitschrift für Ernährungswissenschaften. Supplementum 2.
Marek, J., Wellmann, O. (1932): Die Rachitis. Biochemischer Teil. Gustav Fischer Verlag, Jena.
Patience, J. F., R. E. Austic, Boyd, R.D. (1987): Effect of dietary electrolyte balance on growth and acid-base status in swine. J. Anim. Sci. 64: 457-466.
Roux, M. (2005): The effect of diets varying in dietary cation-anion difference fed in late gestation and in lactation on sow productivity. Louisiana State University Master thesis.

Kienzle: Kationen-Anionen-Bilanz beim Schwein – Möglichkeiten, Risiken und Nebenwirkungen
8. BOKU-Symposium Tierernährung 2009 Seite 27
Yen, J. T., G. Pond, Prior, R.L. (1981): Calcium chloride as a regulator of feed intake and weight gain in pigs. J. Anim. Sci. 52: 778-782.
Autorenanschrift
Prof. Dr. Ellen Kienzle Veterinärwissenschaftliches Department Lehrstuhl für Tierernährung und Diätetik Schönleutnerstraße 8 85764 Oberschleißheim E-Mail: [email protected]

Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
Seite 28 8. BOKU-Symposium Tierernährung 2009
Jod in der Tierernährung und Jodtransfer in Lebensmittel tieri-scher Herkunft
G. Flachowsky, K. Franke, A. Röttger und U. Meyer
Institut für Tierernährung, Friedrich-Loeffler-Institut (FLI), Bundesforschungsinstitut für Tiergesundheit, Bundesallee 50, D-38116 Braunschweig
Einleitung Jod ist ein essentielles Spurenelement für Mensch und Tier. Als Bestandteil der Schilddrüsenhormone, vor allem von Trijodthyronin (T3) und Thyroxin (T4), hat Jod eine herausragende Bedeutung für wich-tige Umsetzungen im Körper (Zus. u.a. bei Mc Dowell et al. 2003, Preedy et al. 2009, Underwood and Suttle et al. 1999). Der Jodbedarf des Menschen wird in Abhängigkeit vom Alter, Körpergewicht, phy-siologischem Stadium und Geschlecht zwischen 40 und 280µg je Tag angegeben (Tabelle 1), jener der Tiere schwankt zwischen 0,15 und 0,5mg je kg Futtertrockensubstanz (TS; Tabelle 1). Unterschiede in den Bedarfsangaben bestehen auch zwischen den verschiedenen wissenschaftlichen Gremien (z.B. Mensch: WHO, FAO, DACH; Tier: NRC, GfE).
Tabelle 1: Empfehlungen zum Jodbedarf und Höchstmengen der Jodaufnahme (Upper Levels, UL)
Mensch Tier Versorgungsempfehlungen (DACH, 2008)
40 – 280 µg/Tag Erwachsene ≈ 200 µg/Tag
(GfE, 1999; 2001; 2006) 0,15 – 0,5 mg/kg TS
Höchstmengen (UL) (DACH, 2008) < 500 µg/Tag (SCF, 2002) 200 – 600 µg/Tag Erwachsene: 600 µg/Tag)
(EU, 2005) 10 mg/kg TS außer: Milchkühe, Legehennen: 5 mg/kg TS Pferde: 4 mg/kg TS Fische: 20 mg/kg TS
Die Angaben zum Jodbedarf von Mensch und Tier sind durchaus vergleichbar, wenn der Bezug auf die Trockenmasseaufnahme erfolgt (≈ 0,3-0,5 mg/kg TS).
In verschiedenen Regionen der Erde leiden Menschen und Tiere an Jod-Defiziten. Nach Einschätzung der WHO (2001) werden gegenwärtig etwa 800 Mio. Menschen mit Jod unterversorgt. Infolge dieser Situation gehört Jod in die Versorgungskategorie 1, denn das Risiko eines Defizits ist relativ hoch (Ta-belle 2).
Daraus resultieren umfangreiche Bemühungen, die Jodversorgung zu verbessern, wie z.B. durch Jod-supplementation des Speisesalzes, den Einsatz von jodiertem Salz in der Lebensmittelindustrie und im Haushalt sowie über die Jodsupplementation von Futtermitteln eine Erhöhung des Jodgehaltes in Le-bensmitteln tierischer Herkunft zu erreichen. Andererseits ist die Spanne zwischen Bedarf und Höchstmenge für Jod in der Humanernährung nicht sehr groß (Bedarf des Erwachsenen: ≈ 200µg; UL: 500-600µg/Tag, s. Tabelle 1), so dass sich nur ein Verhältnis von ≈ 1:3 ergibt.

Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
8. BOKU-Symposium Tierernährung 2009 Seite 29
Tabelle 2: Versorgungs- und Risikokategorien von Spurenelementen und Vitaminen beim Menschen unter Berücksichtigung von Aufnahme und Bedarf (nach BfR, 2004; EFSA, 2006; Gaßmann, 2006)
UL ist nicht definiert oder Faktor > 100Gering
Mittlere Differenz (Faktor 5 – 100)Mittel
Geringe Differenz zwischen Versorgungsempfehlungen und maximal zulässiger Menge (UL; Faktor < 5)
Hoch
Risikokategorie
Aufnahme über Empfehlungen4
Ausreichende Aufnahme3
Mögliches Risiko eines Defizits2
Hohes Risiko eines Defizits1KriteriumVersorgungskategorie
UL ist nicht definiert oder Faktor > 100Gering
Mittlere Differenz (Faktor 5 – 100)Mittel
Geringe Differenz zwischen Versorgungsempfehlungen und maximal zulässiger Menge (UL; Faktor < 5)
Hoch
Risikokategorie
Aufnahme über Empfehlungen4
Ausreichende Aufnahme3
Mögliches Risiko eines Defizits2
Hohes Risiko eines Defizits1KriteriumVersorgungskategorie
Demnach existiert unter bestimmten Bedingungen auch die Gefahr einer Überdosierung und Jod wur-de deshalb in die Risikokategorie „Hoch“ eingeordnet (Tabelle 2).
Diese Situation schlägt sich auch in der EFSA (2005)-Stellungnahme nieder, in der die EFSA die Bei-träge von Lebensmitteln tierischer Herkunft in Abhängigkeit von der Jodversorgung der Tiere zur Jod-versorgung der Menschen wertet. In der Stellungnahme wird geschlussfolgert:
1. Es sind mehr Dosis-Wirkungsstudien mit Lebensmittel-liefernden Tieren notwendig, um den Beitrag dieser Lebensmittel an der Jodversorgung der Menschen besser bewerten zu können.
2. Die Kenntnisse über weitere Einträge in die Lebensmittel tierischer Herkunft sind zu verbes-sern.
3. Der Jodbedarf von Nutztieren mit hohen Leistungen ist zu überprüfen.
In den zurückliegenden Jahren wurden am Institut für Tierernährung des FLI in Kooperation mit ver-schiedenen Partnern (u.a. TLL Jena, MRI mit Standort Kulmbach) zu den Komplexen 1 und 2 ver-schiedenen Versuche mit Lebensmittel-liefernden Tieren durch geführt, über die nachfolgend unter Berücksichtigung der internationalen Literatur berichtet werden soll.
Versuchsansätze Zur Bearbeitung der Fragestellung wurden umfangreiche Versuchsserien am Institut für Tierernährung des FLI durchgeführt (Tabelle 3). Weitere wichtige Einflussgrößen waren unterschiedliche Jodspezies (KI und Ca(IO3)2; (EU 2005) sowie die Gestaltung der Futtermischungen ohne bzw. mit einem glucosi-nolathaltigen Futtermittel (Rapsextraktionsschrot; 3,5 µmol Gesamtglucosinolate/g).
Nachfolgend werden einige wesentliche Versuchsergebnisse bezüglich des Überganges von Jod in Lebensmittel tierischer Herkunft zusammengestellt. Details der Versuchsdurchführung und Ergebnisse können den in Tabelle 3 angeführten Literaturquellen entnommen werden.

Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
Seite 30 8. BOKU-Symposium Tierernährung 2009
Tabelle 3: Dosis-Wirkungs-Versuchsansätze am ITE des FLI
Tierart/-kategorie Versuchstage Literaturquelle Milchkühe Vorversuch (5 Tiere),
0,2; 1,2; 5,5; 10,0 mg I/kg TS; Hauptversuch (32 Tiere), 0,2; 0,6; 1,2; 2,0; 3,2; 5,0 mg I/kg TS; KI und Ca(IO3)2; ohne und mit Rapsextraktionsschrot (3,5 µmol Gesamtglucosinulate/g TS)
Schöne et al. (2009) Franke (2009) Flachowsky et al. (2007)
Mastrinder 0,5; 4 und 10 mg I/kg TS Meyer et al. (2008) Mastschweine 0,2; 0,5; 1, 2 und 5 mg I/kg Futter;
2 bzw. 7 mg I/kg Futter und unterschiedliche Glucosinolatanteile
Berk et al. (2008) Franke et al. (2008) Franke (2009)
Masthähnchen 0,2; 1,0; 5,0; 10,0 mg I/kg Futter; KI und Ca(IO3)2
Röttger (2009)
Pekingenten1) 0,2; 1,0; 5,0; 10,0 mg I/kg Futter Röttger (2009) Legehennen 0,4; 1,0; 2,0; 4,0; 7,0 mg I/kg Futter, KI und
Ca(IO3)2, mit und ohne Rapsextraktionsschrot Röttger (2009)
1) Für den Versuch mit Pekingenten liegen noch keine Ergebnisse zur Jodkonzentrationen in Lebens-mitteln vor
Ergebnisse von Dosis-Wirkungs-Versuchen Versuche mit Milchkühen
Die Jodergänzung des Milchkuhfutters bewirkte einen dramatischen Anstieg der Jodkonzentration in der Milch. Im Vorversuch verursachte die Zulage von 10 mg I/kg TS eine Milchjodkonzentration von > 2mg I/kg Milch (Abb. 1). Diese Ergebnisse waren u.a. eine Grundlage für die Reduzierung der ma-ximal zulässigen Joddosierung im Milchviehfutter von 10 auf 5 mg/kg in der EU (2005).
Daraufhin wurde in den folgenden Versuchen die maximale Joddosierung auf 5 mg/kg TS reduziert. Als Jodspezies kamen KI und Ca(IO3)2 in Rationen ohne und mit Rapsextraktionsschrot zum Einsatz (s. Tabelle 3). Während die Jodspezies keinen signifikanten Einfluss auf die Milchjodkonzentration ausüb-ten, bewirkte das Rapsextraktionsschrot eine dramatische Reduzierung des Milchjodgehaltes (auf etwa 30 – 40 % im Vergleich zur rapsfreien Ration; Abbildung 2). Neben dem Futter existieren auch weitere Eintragsmöglichkeiten von Jod in das Lebensmittel, vor allem in die Milch. Häufig sind Reinigungs- und Euterpflegemittel (Dippmittel) jodhaltig, wobei Reinigungsmittel grundsätzlich nicht in die Milch gelan-gen sollen. Der Jodeintrag in die Milch kann beispielsweise in Abhängigkeit vom Jodgehalt des Dipp-mittels, der Höhe der Milchleistung (Verdünnungseffekt) und anderen Faktoren zwischen 11 und 150 µg/l betragen. In eigenen Versuchen (Flachowsky et al. 2007) wurden 54µg Jod/kg Milch bei Verwendung eines Euterpflegemittels mit 5 g Jod/l ermittelt.

Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
8. BOKU-Symposium Tierernährung 2009 Seite 31
0500
1000150020002500300035004000
0.2 1.2 5.5 10
Jodkonzentration (mg/kg T)
Jod
(µg
kg-1
)
101d ± 32343c ± 109
1215b ± 222
2762a ± 852
0500
1000150020002500300035004000
0.2 1.2 5.5 10
Jodkonzentration (mg/kg T)
Jod
(µg
kg-1
)
101d ± 32343c ± 109
1215b ± 222
2762a ± 852
1) Transfer des supplementierten Jods in die Milch: 31 – 38 %
Abbildung 1: Jodkonzentration der Milch (µg/kg) in Abhängigkeit von der Jodsupplementation) (n = 5, Schöne et al. 2009)
Abbildung 2: Abhängigkeit der Milchjodkonzentration vom Jodgehalt des Futters sowie der eingesetz-ten Proteinquelle und Jodspezies (Franke 2009)
Versuch mit Mastrindern
Im Vergleich zur Milch stieg in den Gewebeproben (außer der Schilddrüse) vom Mastrind die Jodkon-zentration nach Jodzulage zum Futter nur unwesentlich an. Im Muskel bzw. im Fleisch wurden dabei geringere Werte als in der Leber bzw. im Serum ermittelt (Tabelle 4).
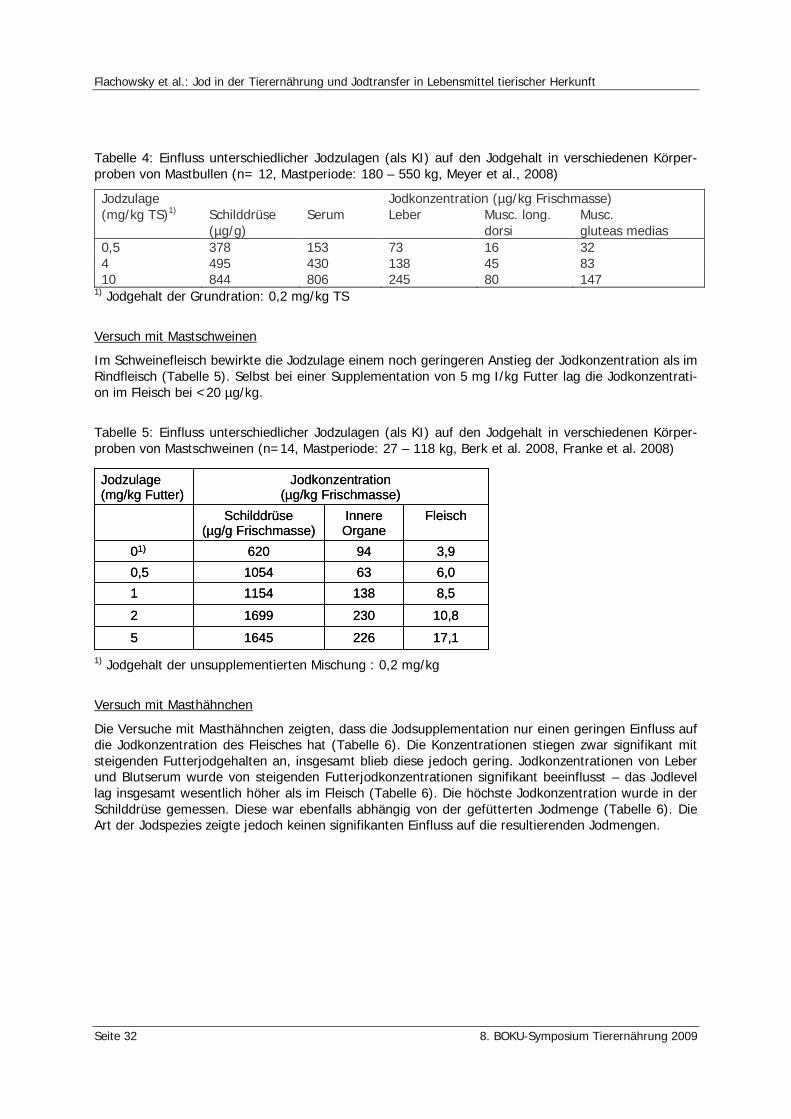
Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
Seite 32 8. BOKU-Symposium Tierernährung 2009
Tabelle 4: Einfluss unterschiedlicher Jodzulagen (als KI) auf den Jodgehalt in verschiedenen Körper-proben von Mastbullen (n= 12, Mastperiode: 180 – 550 kg, Meyer et al., 2008)
Jodzulage Jodkonzentration (µg/kg Frischmasse) (mg/kg TS)1) Schilddrüse
(µg/g) Serum Leber Musc. long.
dorsi Musc. gluteas medias
0,5 378 153 73 16 32 4 495 430 138 45 83 10 844 806 245 80 147
1) Jodgehalt der Grundration: 0,2 mg/kg TS
Versuch mit Mastschweinen
Im Schweinefleisch bewirkte die Jodzulage einem noch geringeren Anstieg der Jodkonzentration als im Rindfleisch (Tabelle 5). Selbst bei einer Supplementation von 5 mg I/kg Futter lag die Jodkonzentrati-on im Fleisch bei <20 µg/kg.
Tabelle 5: Einfluss unterschiedlicher Jodzulagen (als KI) auf den Jodgehalt in verschiedenen Körper-proben von Mastschweinen (n=14, Mastperiode: 27 – 118 kg, Berk et al. 2008, Franke et al. 2008)
1) Jodgehalt der unsupplementierten Mischung : 0,2 mg/kg
Versuch mit Masthähnchen
Die Versuche mit Masthähnchen zeigten, dass die Jodsupplementation nur einen geringen Einfluss auf die Jodkonzentration des Fleisches hat (Tabelle 6). Die Konzentrationen stiegen zwar signifikant mit steigenden Futterjodgehalten an, insgesamt blieb diese jedoch gering. Jodkonzentrationen von Leber und Blutserum wurde von steigenden Futterjodkonzentrationen signifikant beeinflusst – das Jodlevel lag insgesamt wesentlich höher als im Fleisch (Tabelle 6). Die höchste Jodkonzentration wurde in der Schilddrüse gemessen. Diese war ebenfalls abhängig von der gefütterten Jodmenge (Tabelle 6). Die Art der Jodspezies zeigte jedoch keinen signifikanten Einfluss auf die resultierenden Jodmengen.
17,122616455
10,823016992
8,5138115416,06310540,53,99462001)
Innere Organe
FleischSchilddrüse(µg/g Frischmasse)
Jodkonzentration(µg/kg Frischmasse)
Jodzulage(mg/kg Futter)
17,122616455
10,823016992
8,5138115416,06310540,53,99462001)
Innere Organe
FleischSchilddrüse(µg/g Frischmasse)
Jodkonzentration(µg/kg Frischmasse)
Jodzulage(mg/kg Futter)

Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
8. BOKU-Symposium Tierernährung 2009 Seite 33
Tabelle 6: Einfluss unterschiedlicher Jodzulagen und Jodspezies auf den Jodgehalt verschiedener Kör-perproben von Masthähnchen (Median; n=6, Röttger 2009)
Jodzulage
(mg/kg)1)
Jod-
spezies
Jodkonzentration (µg/kg Frischmasse) Schilddrüse [µg/g]
Serum Leber Brustfleisch Schenkelfleisch
01) KI 2317 29 22 5,8 5,9 Ca(IO3)2 2323 28 28 7,0 12,2
1 KI 3315 50 31 10,3 6,9 Ca(IO3)2 3170 50 44 11,5 22,9
2,5 KI 4599 226 103 39,3 38,6 Ca(IO3)2 3117 179 104 27,9 37,7
5 KI 5053 382 173 58,0 63,1 Ca(IO3)2 3652 350 181 52,1 67,8
1) Jodgehalt der unsupplementierten Mischung : 0,5 mg/kg
Versuch mit Legehennen
Ähnlich wie bei der Milch (s. Abb.1 und 2) erfolgt auch im Hühnerei eine erhebliche Anreicherung des supplementierten Jods (Tabelle 7). Dabei ist die Jodkonzentration im Eidotter mehr als 10 x so hoch wie die im Eiklar.
Tabelle 7: Einfluss der Futterjodkonzentration auf den Jodgehalt im Hühnerei (µg/kg Frischmasse; Mittelwerte, n=6; Röttger 2009)
Jodspezies
Jodsupplementation [mg/kg] 0 1 2 4 7
Dotter KI 426 814 1430 3773 5250 Ca(IO3)2 419 657 1811 2913 5522
Eiklar KI 14 36 58 125 224 Ca(IO3)2 33 33 75 131 281
Gesamtei KI 144 281 505 1304 1888 Ca(IO3)2 157 230 582 989 1866
Jodmenge pro Ei [µg/Ei] KI 7.1 13.7 25.2 64.0 87.8 Ca(IO3)2 8.2 12.6 30.7 50.0 96.9
1) Jodgehalt der unsupplementierten Mischung : 0,4 mg/kg
Ein sechsmonatiger Versuch, in dem Legehennen zusätzlich zu den zwei Jodquellen und den verschie-denen Konzentrationsstufen ohne und vergleichend mit 10 % Rapskuchen (Glucosinolatgehalt: 13,8 µmol/g) gefüttert wurden, zeigte, dass der Glucosinolatgehalt die Jodmenge im Ei signifikant verminderte. Die Jodkonzentration der Eier wurde durch den Rapszusatz um bis zu 50 % vermindert.
Vergleichende Wertung
Unter Berücksichtigung der Ergebnisse der am ITE durchgeführten Versuche und der verfügbaren Literatur erfolgt in Tabelle 8 eine Zusammenstellung der Jodkonzentrationen in wichtigen Lebensmit-teln tierischer Herkunft.

Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
Seite 34 8. BOKU-Symposium Tierernährung 2009
Tabelle 8: Einfluss der Höhe der Jodsupplementation im Futter und von Antagonisten auf den Jodge-halt in ausgewählten Lebensmitteln tierischer Herkunft (µg/kg; nach Studien am ITE)
Lebensmittel tierischer Herkunft
Jodsupplementation zum Futter (mg/kg) 0
(0,1-0,4) 0,5 1 2 5 10²)
Milch ohne Raps
50-100
100-20
200-300
600-800
1000-1500
nicht
erlaubt mit Raps3) 20-40 30-80 60-120 200-300 300-600 Rindfleisch 6-10 12-20 15-25 20-40 40-60 60-100 Schweinefleisch 3-5 4-8 6-10 9-12 15-20 20-40 Broilerfleisch Schenkel
3-6
5-8
9-14
25-40
40-70
70-100
Brust 8-10 8-12 12-18 30-50 50-80 80-120 Eier ohne Raps
80-120
100-200
300-400
600-800
1200-1500
nicht
mit Raps3) 50-80 60-120 150-200 300-400 500-800 erlaubt 1) Grundgehalt (nativ) des Futters 2) nur bei wachsenden Tieren 3) 10-20% Rapsextraktionsschrot im Futter
Dabei wird offensichtlich, dass Fleisch nur unwesentlich zur Jodversorgung des Menschen beiträgt, während Milch und Eier wesentliche Jodquellen sind. Aus Gründen des vorbeugenden Verbraucher-schutzes (Jod wird der höchsten Risikokategorie zugeordnet, s. Tabelle 2) wurde deshalb die Ober-grenze für Jod in der Milchvieh- und Legehennenfütterung von 10 auf 5mg/kg Futter reduziert (EU 2005, s. Tabelle 1). Die vorgelegten Ergebnisse sind Anlass, bei glucosinolatfreien Milchviehrationen über eine weitere Reduzierung des maximal möglichen Jodzusatzes nachzudenken.
Transfer von Jod ins Lebensmittel
Unter Transfer wird die Menge des supplementierten Nährstoffs verstanden, die in der essbaren Frak-tion (Milch, Fleisch, Eier) wiedergefunden wird:
Menge des Nährstoffs Menge des Nährstoffs in essbaren Teilen nach _ in essbaren Teilen ohne Zusatz (mg) Zusatz (mg)
Transfer (%) = ________________________________________________________ x 100 Menge des aufgenommenen zugesetzten Nährstoffes (mg)
Derartige Kalkulationen sind für Milch und Eier relativ einfach durchführbar. Für vom Tierkörper stammende Lebensmittel, wie Fleisch oder innere Organe, sind Transferberechnungen deutlich schwieriger, da häufig Ausgangswerte („Starttiere“) fehlen, die Zuordnung zur essbaren Fraktion in verschiedenen Regionen unterschiedlich ist und häufig auch analytische Daten fehlen. Besonders problematisch ist der Umgang mit spezifischen Speicherorganen, wie im Falle von Jod mit der Schild-drüse (>70% der Körperreserven). Bei Transferberechnungen bleiben diese Organe meist unberück-sichtigt, wenn sie keine (z. B. Schilddrüse) oder kaum Bedeutung (z.B. Blut) für die Humanernährung haben.
In Tabelle 9 werden zu erwartende Transferwerte für Jod zusammengestellt. Dabei zeigt sich, dass für Milch und Eier deutlich höhere Werte ermittelt wurden als für Fleisch. Durch Einsatz von Rapsproduk-ten als Proteinquelle vermindern sich die Transferwerte etwa auf die Hälfte.

Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
8. BOKU-Symposium Tierernährung 2009 Seite 35
Tabelle 9: Transfer vom supplementiertem Jod vom Futtermittel in Lebensmittel tierischer Herkunft (% der Supplementation in essbare Fraktion; nach Versuchen des ITE im FLI)
Zulage (mg/kg TS) 0,51) 1 2 5 10 Milch ohne Antagonsiten2) mit Antagonisten3)
50 - 60 25 - 30
40 - 50 20 - 25
35 - 45 15 - 20
30 - 40 10 - 20
(30 - 40)
k.D. Fleisch
Rindfleisch
0,5 - 1
0,4-0,8
0,3-0,6
0,2 - 0,5
0,1 - 0,2 Schweinefleisch 0,3 - 0,5 0,3-0,4 0,2 - 0,3 0,1 - 0,2 0,05-0,1 Broilerfleisch 0,1-0,2 0,2-0,4 0,2-0,4 0,2-0,4 0,1-0,3
Eier ohne Antagonisten mit Antagonisten
10-20 5-10
10-20 5-10
10-20 5-10
10 - 20 5 - 10
k.D. k.D.
1) Basaltgehalt: 0,1 - 0,2 mg/kg 2) Glucosinolatfreie Mischung 3) mit 10-20% Rapsprodukten im Futter k.D.: keine Daten
Zusammenfassung Für Jod als essentielles Spurenelement besteht ein hohes Risiko für Defizite in der Human- und Tier-ernährung. Andererseits ist die Spanne zwischen Versorgungs-empfehlung und maximal zulässiger Menge in der Humanernährung relativ gering (1 : 3), so dass der möglichen Jodanreicherung in Le-bensmitteln tierischer Herkunft sowohl aus Gründen der Bedarfsabdeckung beim Menschen als auch der Vermeidung von Überschüssen erhöhte Aufmerksamkeit zu widmen ist.
In Dosis-Wirkungs-Studien mit Lebensmittel liefernden Tieren wurde der Übergang von Jod unter Berücksichtigung verschiedener Einflussfaktoren (Jodspezies, Jodantagonisten) in Milch, Fleisch und Eier untersucht. Dabei zeigte sich, dass der Jodtransfer ins Fleisch deutlich geringer ist (meist <1%) als der in Milch und Eier (meist 10 bis 40%) und das Fleisch keine wesentliche Jodquelle für den Men-schen darstellt.
Unter Berücksichtigung des hohen Jodtransfers in Milch und Eier war aus Gründen des vorbeugenden Verbraucherschutzes die Reduzierung der Jodobergrenze von 10 auf 5 mg/kg TS gerechtfertigt.
Raps (glucosinolat-)haltige Futtermischungen führen etwa zu einer Halbierung der Jodkonzentration in Milch und Eiern.
Literatur
Berk, A., Wagner, H., Flachowsky, G. (2008): Zum Einfluss unterschiedlicher Glucosinolatgehalte im Schweinefutter auf den Jodgehalt von Schilddrüse und Schlachtkörper von Mastschweinen. Forum angewandte Forschung, Fulda, 09./10.04.2008, 183-185
BfR (2004): Teil I: Verwendung von Vitaminen in Lebensmitteln. Teil II: Verwendung von Mineralstoffen in Lebensmitteln. Toxikologische und ernährungsphysiologische Aspekte. BfR-Pressestelle, Berlin, BfR-Wissenschaft 03 und 04/2004
DACH (2008): Referenzwerte für die Nährstoffzufuhr, 1. Aufl., 3. korr. Nachdruck, Umschau Braus GmbH Verlagsges., Frankfurt/M. 2008, p. 179-184
EFSA (2005): Opinion of the Scientific Panel on Additives and Products or Substances used in Animal Feed on the request from the Commission on the use of Iodine in feedingstuffs. The EFSA J. 168, 1-42
EFSA (2006): Tolerable upper intake levels for vitamins and minerals. http://ec.europa.eu/comm./food/fs/scf/outcome
EU (2005): Regulation (EC) No. 1459/2005 of the European Parliament and of the Council of 8. Sept. 2005 amending the condi-tions for authorisation of a number of feed additives belonging to the group of trace elements. OJEU 2005, 48. L233/8 – L 233/10

Flachowsky et al.: Jod in der Tierernährung und Jodtransfer in Lebensmittel tierischer Herkunft
Seite 36 8. BOKU-Symposium Tierernährung 2009
Flachowsky, G., Schöne, F., Leiterer, M., Bemmann, D., Spolders, M., Lebzien, P. (2007): Influence of an iodine depletion period and teat dipping on the iodine concentration in serum and milk of cows. J. Anim. Feed Sci. 15, 18-25
Franke, K. (2009): Effect of various iodine supplementations and species on the iodine transfer into milk and on serum, urinary and faecal iodine of dairy cows fed rations varying in the glucosinolate content. Diss. Univ. Halle/S. 119 p.
Franke, K., Schöne, F., Berk, A., Leiterer, M., Flachowsky, G. (2008): Influence of dietary iodine on the distribution of the trace element in the body. European J. Nutr. 47, 40-46
Gassmann, B. (2006): Zum Festlegen von Höchst- und Mindestmengen an Vitaminen und Mineralstoffen in Lebensmitteln. Ernähr.-Umsch. 53, 336-343
McDowell, L.R. (2003): Minerals in animal and human nutrition. Elsevier, 2nd ed., 644 p.
Meyer, U., Weigel, K., Schöne, F., Leiterer, M., Flachowsky, G. (2008): Effect of dietary iodine on growth and iodine status of growing-fattening bulls. Livestock Sci. 115, 219-225
Preedy, V.R., Burrow, G.N., Watson, R. (2009): Comprehensive Handbook of Iodine. Nutritional, Biochemical, Pathological and Therapeutic aspects. Oxford: Academic Press,
Röttger, A. (2009): Einfluss unterschiedlicher Jodmengen und –quellen auf den Jodtransfer in Eier und Geflügelfleisch. Diss.TiHo Hannover, in Vorbereitung
SCF (Scientific Committee on Food, 2002): Opinion of the Scientific Committee on Food on the tolerable upper intake level of Iodine. Expressed on 26. Sept. 2002
Schöne, F., Leiterer, M., Lebzien, P., Bemmann, D., Spolders, M., Flachowsky, G. (2009): Iodine concentration of milk in a dose-respinse study with dairy cows and implications for consumer iodine intake. J. Trace Elements. Med. Biol. (2009), doi: 10.1016/j.jtemb. 2009.02.004
Schöne, F., Rajenchram, R. (2009): Iodine in Farm Animals. In: V.R. Preedy, G.N. Burrow, V.R. Watson (Eds.) Comprehensive Handbook of Iodine Oxford, Acad. Press, 151-170
Underwood, L.J., Suttle, N.F. (1999): The mineral nutrition of livestock. 3rd ed. CABI-Publ., UK
WHO, UNICEF, ICCIDD (2001): Assessment of the iodine deficiency disorders and monitoring their elimination. Geneva: WHO publ. WHO/NHD/01.1.; p. 1-107
Autorenanschrift
Prof. Dr. Gerhard Flachowsky Institut für Tierernährung Friedrich-Loeffler-Institut Bundesforschungsinstitut für Tiergesundheit Bundesallee 50 D-38116 Braunschweig E-Mail: [email protected]

Windisch et al.: Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung?
8. BOKU-Symposium Tierernährung 2009 Seite 37
Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierer-nährung?
Wilhelm Windisch1, Thomas Kickinger2, Herbert Würzner2 1 Universität für Bodenkultur Wien, Österreich 2 AGES Wien, Österreich
Zink und Kupfer in der „guten fachlichen Fütterungspraxis“ Zink (Zn) und Kupfer (Cu) sind essentielle Spurenelemente und müssen über das Futter aufgenom-men werden. Der Bedarf heutiger landwirtschaftlicher Nutztiere ist in der Regel so hoch, dass die nati-ven Gehalte des Futters normalerweise nicht mehr ausreichen und Zink und Kupfer über das Mineral-futter supplementiert werden müssen. Im Falle von Zink besteht die Notwendigkeit der Ergänzung praktisch bei allen monogastrischen Nutztieren. Bei Wiederkäuern steht dagegen hauptsächlich Kupfer im Vordergrund, das aufgrund chemischer Umsetzungen in den Vormägen in der Regel weniger bio-verfügbar ist als bei Monogastriern. Um eine ausreichende Versorgung der Nutztiere mit diesen beiden Spurenelementen zu gewährleisten, sollte die Gesamtration je nach Spezies und Kategorie etwa 40 bis 100 mg Zn bzw. 4 bis 10 mg Cu/kg Trockenmasse enthalten (GfE 1999, 2001, 2006). In diesen Emp-fehlungen sind Sicherheitszuschläge bereits enthalten.
Aus der Sicht der Körperzellen stellen Zink und Kupfer, wie auch die anderen metallischen Spuren-elemente, hochtoxische Gefahrstoffe dar, deren Umgang im Stoffwechsel präzise kontrolliert werden muss. Ein wichtiger Steuermechanismus ist die homöostatische Regulation der Absorption und Exkre-tion. Sie sorgt dafür, dass der Organismus nur so viel Zink und Kupfer aus der Nahrung aufnimmt, wie er tatsächlich benötigt. Der Überschuss wird über die Exkremente wieder abgegeben und landet un-weigerlich im Wirtschaftsdünger.
Unter normalen Fütterungsbedingungen ist nur ein Teil des über die Nahrung aufgenommenen Zinks und Kupfers überhaupt absorbierbar. Der nicht absorbierte Teil gelangt unmittelbar in die Wirtschafts-dünger. Darüber hinaus ist es ein Grundprinzip der fachgerechten Fütterung, den Tieren absorbierba-res Zink und Kupfer in einer Menge vorzulegen, die den tatsächlichen Bedarf leicht überschreitet. Da-mit werden einerseits vorübergehende Phasen einer Unterversorgung sicher ausgeschlossen, anderer-seits kann der Organismus über die homöostatische Regulation die aus dem Futter tatsächlich absor-bierten Mengen an Zink und Kupfer selbst steuern. Diese Sicherheitszuschläge an Zink und Kupfer gelangen ebenfalls in die Wirtschaftsdünger.
Die Höhe des Austrags an Zink und Kupfer in die Wirtschaftsdünger hängt somit entscheidend vom Grad der Überversorgung der Tiere an diesen Spurenelementen ab. In diesem Zusammenhang sind die Zn- und Cu-Gehalte im Futter durch das Futtermittelrecht nach oben hin begrenzt (bezogen auf 88 % Trockenmasse: 150 mg Zn/kg für alle Nutztierspezies; 35 mg Cu/kg für Rinder, 25 mg Cu/kg für Schweine und Geflügel mit Ausnahme von 170 mg Cu/kg für Ferkel bis zu einem Lebensalter von 12 Wochen; Commission Regulation (EC) No 1334/2003). Diese Obergrenzen liegen deutlich oberhalb der Empfehlungen für eine adäquate Versorgung der Nutztiere mit diesen essentiellen Spurenelementen. Sie beinhalten somit einen Überschuss an Zink und Kupfer, der vom Organismus nicht verwertet wird und deshalb quantitativ vollständig über die Exkremente in die Wirtschaftsdünger gelangt.
Die Konzentrationen an Zink und Kupfer im Wirtschaftsdünger, die aus der vollen Ausnutzung der futtermittelrechtlich zulässigen Grenzen resultieren, lassen sich zumindest hinsichtlich ihrer Obergren-zen relativ einfach kalkulieren. Unter der Annahme, dass beim Schwein 25 % der Futtertrockenmasse

Windisch et al.: Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung?
Seite 38 8. BOKU-Symposium Tierernährung 2009
über Kot und Harn ausgeschieden werden, beim Geflügel 20 % und beim Rind 30 %, beträgt der An-reicherungsfaktor vom Futter in den Exkrementen etwa 4 (Schwein), 5 (Geflügel), bzw. 3,3 (Rind). Unter der übertriebenen Annahme, dass das gesamte Zink und Kupfer des Futters wieder ausgeschie-den wird und neben den Exkrementen sonst keine organische Substanz in die Wirtschaftsdünger ge-langt (z.B. Einstreu), sind für Schweine, Geflügel und Rindern höchstens 600, 750 und 500 mg Zn bzw. 100, 130 und 120 mg Cu je kg Wirtschaftsdünger-Trockenmasse zu erwarten, vorausgesetzt, man verzichtet auch beim Ferkelfutter auf exzessive Cu-Dosierungen und verfüttert maximal 25 mg Cu/kg.
Exzessive Dosierungen von Zink und Kupfer Zink und Kupfer üben ihre Funktionen als essentielle Spurenelemente nur im ernährungsphysiologisch relevanten Konzentrationsbereich des Futters aus, wie er durch die Versorgungsempfehlungen defi-niert wird. Überschüssig aufgenommene Mengen werden im Zuge der homöostatischen Regulation ausgeschieden. Diese Kompensationsfähigkeit ist bei Zink und Kupfer zwar sehr leistungsfähig, stößt jedoch bei einer Überdosierung in der Größenordnung von etwa Faktor 50 über dem physiologischen Bedarf an ihre Grenzen (dieser Wert kann je nach Bioverfügbarkeit der zugesetzten Verbindung auch deutlich niedriger liegen!). Oberhalb dieser Schwelle strömen größere Mengen an Zink und Kupfer aus dem Futter in den Organismus, die dann auch für die toxischen Wirkungen verantwortlich sind. Zwar kommt es schon vorher zu Akkumulationen an Zink und Kupfer vor allem in der Leber, aber die ent-sprechenden Mengen sind im Vergleich zu den Gehalten im Futter vernachlässigbar klein. Dies bedeu-tet, dass bis zum Erreichen der akuten Toxizität praktisch die gesamten Mengen an Zink und Kupfer des Futters, soweit sie den physiologischen Bedarf überschreiten, vollständig in die Wirtschaftsdünger überführt werden.
Exzessive Dosierungen an Zink und Kupfer im Futter sind für ihre leistungsfördernden Effekte und pharmakologischen Wirkungen zur Prävention bzw. Behandlung von Durchfällen seit Jahrzehnten be-kannt und gut beschrieben (Cromwell et al., 1989; Hahn and Baker, 1993; Coffey et al., 1994; Apgar et al., 1995; Apgar and Kornegay, 1996; Poulsen, 1995; Smith et al, 1995; Carlson et al, 1999; Win-disch et al., 1999; Windisch et al., 2001). Sie sind vor allem im Bereich der Ferkelfütterung weit ver-breitet, scheinen aber auch noch in der Schweinemast Anwendung zu finden (Kickinger et al. 2008). Die erforderlichen Mindestdosierungen im Futter liegen oberhalb von etwa 2000 mg Zn/kg bzw. 100 mg Cu/kg. Futtermittelrechtlich ist dies bei Kupfer nur im Falle des Ferkels abgedeckt (max. 170 mg Cu/kg bis zu einem Alter von 12 Wochen). Bei Zink liegt die Obergrenze im Futter für alle Spezies bei einheitlich 150 mg/kg, so dass exzessive Dosierungen futtermittelrechtlich in keinem Fall zulässig sind. Allerdings gestatten einige Länder der Europäischen Union (Dänemark, Schweden, Finnland) die An-wendung hoher Dosierungen von Zinkoxid als Arzneimittel.
Die Anwendung exzessiver Dosierungen von Zink und Kupfer in der Tierernährung hat zwangsläufig einen massiven Eintrag dieser Elemente in die Wirtschaftsdünger zur Folge. Diese Mengen werden in der Mischgülle zwar wieder verdünnt, hinterlassen aber dennoch ihre Spuren. So lässt sich beispiels-weise für Schweine anhand einfacher Kalkulationen ableiten, dass die Gabe exzessiver Mengen an Zink und Kupfer an Ferkel und Mastschweine die jeweiligen Gehalte der Mischgülle um etwa das 2- bis 3fache auf Werte um etwa 2000 mg Zn/kg T bzw. 300 mg Cu/kg T ansteigen lassen (Kickinger et al. 2008).

Windisch et al.: Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung?
8. BOKU-Symposium Tierernährung 2009 Seite 39
Zink und Kupfer aus der Sicht des Bodenschutzes Das oberste Ziel des Bodenschutzes ist die langfristige Erhaltung der lebenswichtigen Funktionen des Bodens als Puffer, Filter und Reinigungsmedium an der Schnittstelle zwischen Luft und Grundwasser, als Standort für Pflanzen und als Ausgangsort der Erzeugung von Nahrungs- und Futtermitteln. In der seit nahezu 3 Jahrzehnten praktizierten Konzeption des Bodenschutzes werden Akkumulationen von Schwermetallen auf landwirtschaftlichen Nutzflächen durchaus noch toleriert. Das Ausmaß der Akku-mulation wird jedoch durch Regelungen der Höchstgehalte an Schwermetallen in relevantem Material (z.B. Bioabfall, Klärschlamm, etc.) limitiert, so dass die Erreichung einer kritischen Konzentration im Boden (Vorsorgewerte) frühestens in einigen Jahrzehnten zu erwarten ist. Es ist jedoch festzuhalten, dass die bislang eingetretenen Akkumulationen zumindest mittelfristig irreversibel sind. Es wird somit immer vordringlicher, die Akkumulation an Schwermetallen im Boden künftig nicht mehr nur in ihrer Geschwindigkeit zu bremsen, sondern vielmehr endgültig zum Stillstand zu bringen. In diesem Zu-sammenhang ist es von erheblicher Bedeutung, dass Wirtschaftsdünger mit etwa drei Vierteln der Gesamtfracht an Zink und Kupfer auf die landwirtschaftliche Nutzfläche den mit Abstand stärksten Eintragspfad darstellen und deshalb auch als Hauptverursacher von Akkumulationen im Oberboden angesehen werden (Aichberger et al., 1995; Müller 1997; Dannenberg 1997; Kilian 1999; Bonnier 2002; Roth et al., 2002; Dersch und Hösch 2003; UBA, 2004; Nicholson et al., 2005).
Von Seiten des Bodenschutzes diskutiert man über die Einführung von Grenzwerten für die Gehalte an Zink, Kupfer und weiteren Schwermetallen im Wirtschaftsdünger, um weitere Akkumulationen auf den landwirtschaftlich genutzten Böden zu vermeiden. Zur Ableitung der hierfür erforderlichen Obergren-zen stehen verschiedene Konzepte im Raum, etwa die ausgeglichene Bilanz zwischen allen Ein- und Austragspfaden des Bodens, oder die Forderung, dass der langfristig im Boden verbleibende Anteil des aufgebrachten Materials keine höheren Konzentrationen an Schwermetallen aufweisen dürfe als der Boden selbst. Unter Berücksichtigung der besonderen Gegebenheiten der landwirtschaftlichen Praxis liegen die vorgeschlagenen Obergrenzen bei höchstens 450 mg Zink und 90 mg Kupfer je kg Wirt-schaftsdünger-Trockenmasse (UBA, 2001; Döhler et al. 2002; Schwarz und Freudenschuss, 2004). Sie sind damit durchaus vergleichbar mit den Regelungen für Bioabfall (400 mg Zink und 100 mg Kupfer je kg T; BIOABFV, 1998). Aus diesem Werten geht klar hervor, dass die Anwendung exzessiver Dosie-rungen an Zink und Kupfer im Tierfutter und die daraus resultierenden hohen Gehalte im Wirtschafts-dünger mit den Zielen des Bodenschutzes kollidieren.
Zink und Kupfer im Wirtschaftsdünger Tabelle 1 zeigt Messwerte der Zink- und Kupfergehalte von Wirtschaftsdüngern verschiedener Spezies landwirtschaftlicher Nutztiere aus Österreich und Deutschland. Bei Rindern und Geflügel entsprechen die beobachteten Werte durchaus den Erwartungen einer guten landwirtschaftlichen Praxis und kom-men auch den Vorstellungen des Bodenschutzes bereits sehr nahe. Bei diesen Spezies kann man so-mit davon ausgehen, dass die Gehalte an Zink und Kupfer im Wirtschaftsdünger kein grundsätzliches Problem darstellen.
Bei Schweinen liegen die Werte dagegen deutlich über denen von Rind bzw. Geflügel und überschrei-ten die Vorstellungen des Bodenschutzes für die Obergrenzen einer ökologisch verträglichen Fracht an Zink und Kupfer auf die landwirtschaftliche Nutzfläche bei Weitem. Darüber hinaus liegen die be-obachten Gehalte klar über den Werten, die bei Einhaltung der futtermittelrechtlichen Obergrenzen und Verzicht auf exzessive Dosierungen von Zink und Kupfer zu erwarten wären.
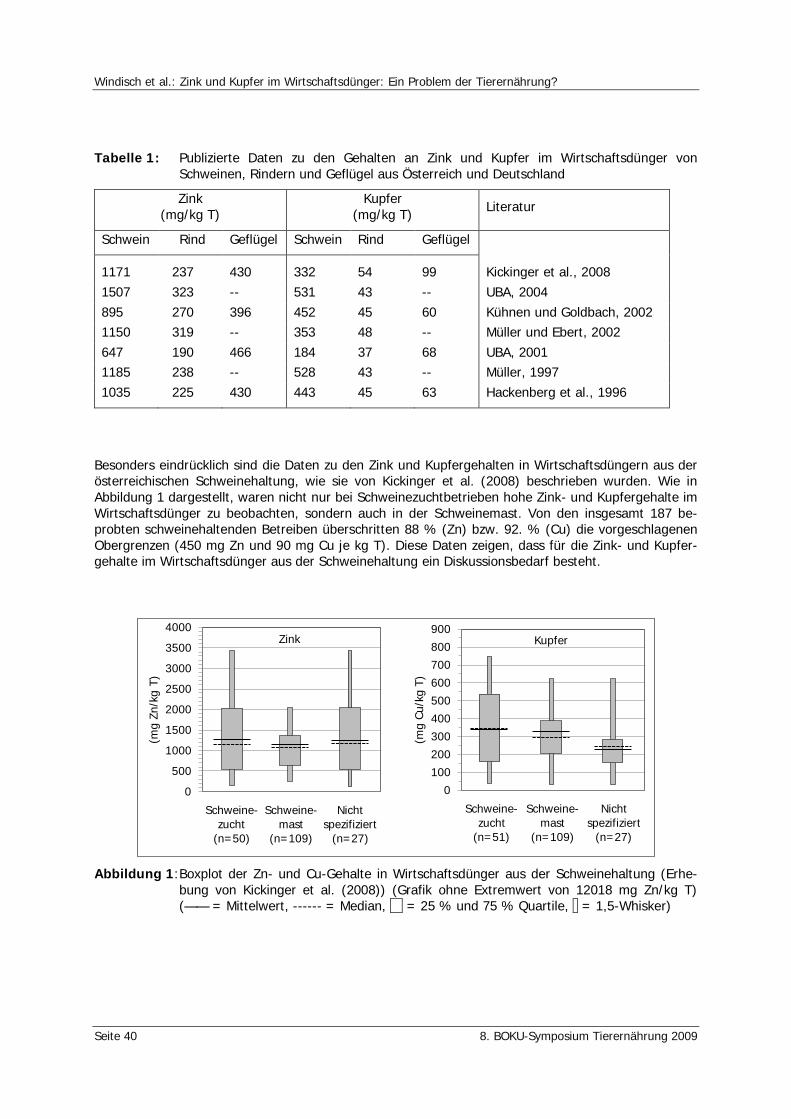
Windisch et al.: Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung?
Seite 40 8. BOKU-Symposium Tierernährung 2009
Tabelle 1: Publizierte Daten zu den Gehalten an Zink und Kupfer im Wirtschaftsdünger von Schweinen, Rindern und Geflügel aus Österreich und Deutschland
Zink (mg/kg T)
Kupfer (mg/kg T) Literatur
Schwein Rind Geflügel Schwein Rind Geflügel
1171 237 430 332 54 99 Kickinger et al., 2008 1507 323 -- 531 43 -- UBA, 2004 895 270 396 452 45 60 Kühnen und Goldbach, 2002 1150 319 -- 353 48 -- Müller und Ebert, 2002 647 190 466 184 37 68 UBA, 2001 1185 238 -- 528 43 -- Müller, 1997 1035 225 430 443 45 63 Hackenberg et al., 1996
Besonders eindrücklich sind die Daten zu den Zink und Kupfergehalten in Wirtschaftsdüngern aus der österreichischen Schweinehaltung, wie sie von Kickinger et al. (2008) beschrieben wurden. Wie in Abbildung 1 dargestellt, waren nicht nur bei Schweinezuchtbetrieben hohe Zink- und Kupfergehalte im Wirtschaftsdünger zu beobachten, sondern auch in der Schweinemast. Von den insgesamt 187 be-probten schweinehaltenden Betreiben überschritten 88 % (Zn) bzw. 92. % (Cu) die vorgeschlagenen Obergrenzen (450 mg Zn und 90 mg Cu je kg T). Diese Daten zeigen, dass für die Zink- und Kupfer-gehalte im Wirtschaftsdünger aus der Schweinehaltung ein Diskussionsbedarf besteht.
Abbildung 1: Boxplot der Zn- und Cu-Gehalte in Wirtschaftsdünger aus der Schweinehaltung (Erhe-bung von Kickinger et al. (2008)) (Grafik ohne Extremwert von 12018 mg Zn/kg T) (—— = Mittelwert, ------ = Median, = 25 % und 75 % Quartile, = 1,5-Whisker)
0
500
1000
1500
2000
2500
3000
3500
4000
Schweine-zucht
(n=50)
Schweine-mast
(n=109)
Nichtspezifiziert
(n=27)
(mg
Zn/k
g T)
0
100
200
300
400
500
600
700
800
900
Schweine-zucht
(n=51)
Schweine-mast
(n=109)
Nichtspezifiziert
(n=27)
(mg
Cu/k
g T)
Zink Kupfer

Windisch et al.: Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung?
8. BOKU-Symposium Tierernährung 2009 Seite 41
Die in Abbildung 1 dargestellten, hohen Cu-Gehalte im Wirtschaftsdünger der Schweinezuchtbetriebe sind durchaus mit der futtermittelrechtlich gestatteten Anwendung exzessiver Dosierungen im Ferkel-futter erklärbar. Dies gilt jedoch nicht für die hohen Cu-Gehalte im Wirtschaftsdünger von Schweine-mastbetrieben, sowie für die bei allen Kategorien beobachteten, im Einzelfall zum Teil sehr hohen Zinkgehalten (bis zu 12018 mg/kg T in der Mischgülle). Zwar stammen die Wirtschaftsdünger-Proben teilweise aus Zeiten, in denen der zulässige Zinkgehalt des Futters noch 250 mg/kg betrug (bezogen auf 88 % T). Dies hätte jedoch maximal nur 1000 mg Zn/kg Wirtschaftsdünger-T zur Folge gehabt. Damit verbleibt für die Mehrzahl der insgesamt 186 beprobten Betriebe immer noch ein gewisser Er-klärungsbedarf für die hohen Zinkgehalte, wobei die Anwendung exzessiver Zink-Dosierungen im Fut-ter sowohl von Ferkeln als auch von Mastschweinen am plausibelsten erscheint (siehe auch Eckel et al. 2007).
Die verbreitete Anwendung exzessiver Dosierungen von Zink und Kupfer in der Schweinefütterung außerhalb der futtermittelrechtlich zulässigen Grenzen bleibt eine Vermutung, solange nicht aufgezeigt wird, dass Wirtschaftsdünger bei nachweislicher Befolgung der Obergrenzen im Futter deutlich niedri-gere Zn- und Cu-Gehalte aufweisen. Darüber hinaus beantwortet diese Vermutung nicht die Frage, ob die Schweinefütterung bei Einhaltung einer guten fachlichen Praxis in Bezug auf die Einträge an Zink und Kupfer nicht doch ökologisch verträglich ist und den Vorstellungen des Bodenschutzes genügt. Aus diesem Grunde wurde in Österreich eine neue Erhebung an Schweinemastbetrieben durchgeführt, für die laut Futtermittelrecht auch im Falle des Kupfers keine exzessiven Dosierungen erlaubt sind (Kickinger et al., 2009). In diesem Datenpool befanden sich neben Selbstmischern auch solche Betrie-be, die ausschließlich kommerzielles Fertigfutter einsetzten. Letztere sollten ein gutes Modell darstel-len, welche Gehalte an Zink und Kupfer im Wirtschaftsdünger bei vollständig kontrolliertem Eintrag über das Futter zu erwarten sind.
Die Zink und Kupfergehalte im Futter der insgesamt 6 Betriebe mit ausschließlicher Verwendung von kommerziellem Mischfutter lagen erwartungsgemäß innerhalb der futtermittelrechtlich zulässigen Grenzen (bezogen auf 88 % Trockenmasse: 104 ±26 mg Zn/kg bzw. 20 ±2 mg Cu/kg). Bei den ins-gesamt 21 Selbstmischern mussten für Zink 2 und für Kupfer 3 Datensätze ausgeschlossen werden, da ungewöhnlich hohe Gehalte im Futter (bis zu 2138 mg Zn/kg und 90 mg Cu/kg) bzw. in der Gülle (bis zu 6210 mg Zn/kg) auf einen futtermittelrechtlich nicht abgedeckte Einsatz von Zink und Kupfer hinwiesen. Bei den verbliebenen Daten lagen die Zink- und Kupfergehalte des Mastfutters (bezogen auf 88 % Trockenmasse) im Mittel bei 129 ±39 mg/kg bzw. 25 ±6 mg Cu/kg). Die höheren Werte im Futter der Selbstmischer dürften hauptsächlich darauf beruhen, dass die nativen Gehalte der Fut-termittel bei der Zumischung des Mineralfutters in der Regel nicht berücksichtig werden.
Abbildung 2 zeigt die Verteilung der Zink- und Kupfergehalte in der Gülle der Schweinemastbetriebe. Die Werte bewegen sich weit unter dem Niveau, wie sie in der Literatur für Wirtschaftsdünger aus der Schweinehaltung beschrieben werden (vgl. Tabelle 1). Im Durchschnitt treffen sie sehr gut die theore-tisch erwarteten Zn und Cu-Gehalte, wobei Betriebe mit ausschließlicher Verwendung von kommerziel-lem Mischfutter aufgrund niedrigerer Gehalte im Futter erwartungsgemäß auch unter dem Niveau der Selbstmischer liegen. Die Rate des Transfers vom Futter in die Gülle betrug bei den Betrieben mit kommerziellem Mischfutter sehr einheitlich 84 % (Zink) bzw. 67 % (Kupfer). Dies bekräftigt die Ver-mutung, dass das Futter der einzige quantitativ relevante Eintragspfad von Zink und Kupfer in die Wirtschaftsdünger darstellt. Umgekehrt sind hohe Gehalte an Zink und Kupfer im Wirtschaftsdünger am plausibelsten mit hohen Gehalten im Futter zu erklären. Die Vermutung einer breiten Anwendung exzessiver Dosierungen an Zink und Kupfer als Hauptgrund für die ungewöhnlich hohen Gehalte an Zink und Kupfer im Wirtschaftsdünger aus der Schweinehaltung wird damit indirekt bekräftigt. Umge-kehrt zeigen diese Befunde, dass eine Schweinefütterung im Rahmen der guten fachlichen Praxis un-ter Verzicht auf exzessive Dosierungen den ökologischen Zielen des Bodenschutzes bereits sehr nahe kommt.
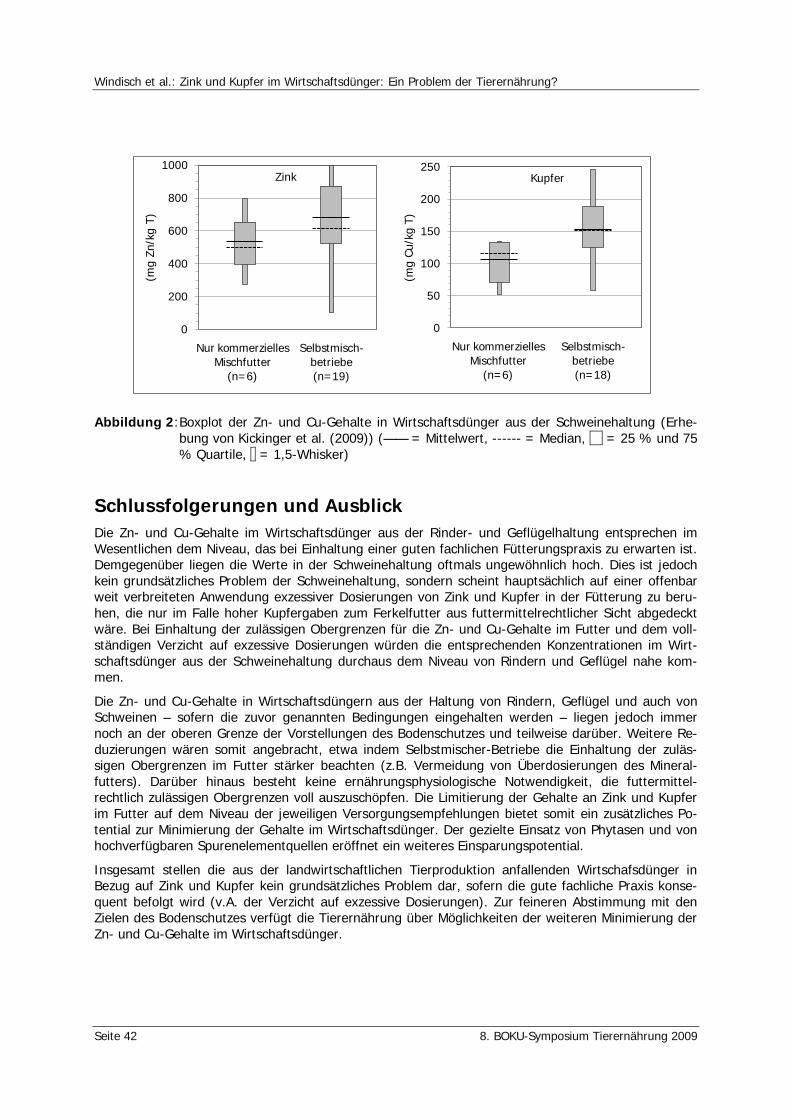
Windisch et al.: Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung?
Seite 42 8. BOKU-Symposium Tierernährung 2009
Abbildung 2: Boxplot der Zn- und Cu-Gehalte in Wirtschaftsdünger aus der Schweinehaltung (Erhe-bung von Kickinger et al. (2009)) (—— = Mittelwert, ------ = Median, = 25 % und 75 % Quartile, = 1,5-Whisker)
Schlussfolgerungen und Ausblick Die Zn- und Cu-Gehalte im Wirtschaftsdünger aus der Rinder- und Geflügelhaltung entsprechen im Wesentlichen dem Niveau, das bei Einhaltung einer guten fachlichen Fütterungspraxis zu erwarten ist. Demgegenüber liegen die Werte in der Schweinehaltung oftmals ungewöhnlich hoch. Dies ist jedoch kein grundsätzliches Problem der Schweinehaltung, sondern scheint hauptsächlich auf einer offenbar weit verbreiteten Anwendung exzessiver Dosierungen von Zink und Kupfer in der Fütterung zu beru-hen, die nur im Falle hoher Kupfergaben zum Ferkelfutter aus futtermittelrechtlicher Sicht abgedeckt wäre. Bei Einhaltung der zulässigen Obergrenzen für die Zn- und Cu-Gehalte im Futter und dem voll-ständigen Verzicht auf exzessive Dosierungen würden die entsprechenden Konzentrationen im Wirt-schaftsdünger aus der Schweinehaltung durchaus dem Niveau von Rindern und Geflügel nahe kom-men.
Die Zn- und Cu-Gehalte in Wirtschaftsdüngern aus der Haltung von Rindern, Geflügel und auch von Schweinen – sofern die zuvor genannten Bedingungen eingehalten werden – liegen jedoch immer noch an der oberen Grenze der Vorstellungen des Bodenschutzes und teilweise darüber. Weitere Re-duzierungen wären somit angebracht, etwa indem Selbstmischer-Betriebe die Einhaltung der zuläs-sigen Obergrenzen im Futter stärker beachten (z.B. Vermeidung von Überdosierungen des Mineral-futters). Darüber hinaus besteht keine ernährungsphysiologische Notwendigkeit, die futtermittel-rechtlich zulässigen Obergrenzen voll auszuschöpfen. Die Limitierung der Gehalte an Zink und Kupfer im Futter auf dem Niveau der jeweiligen Versorgungsempfehlungen bietet somit ein zusätzliches Po-tential zur Minimierung der Gehalte im Wirtschaftsdünger. Der gezielte Einsatz von Phytasen und von hochverfügbaren Spurenelementquellen eröffnet ein weiteres Einsparungspotential.
Insgesamt stellen die aus der landwirtschaftlichen Tierproduktion anfallenden Wirtschafsdünger in Bezug auf Zink und Kupfer kein grundsätzliches Problem dar, sofern die gute fachliche Praxis konse-quent befolgt wird (v.A. der Verzicht auf exzessive Dosierungen). Zur feineren Abstimmung mit den Zielen des Bodenschutzes verfügt die Tierernährung über Möglichkeiten der weiteren Minimierung der Zn- und Cu-Gehalte im Wirtschaftsdünger.
0
200
400
600
800
1000
Nur kommerziellesMischfutter
(n=6)
Selbstmisch-betriebe(n=19)
(mg
Zn/k
g T)
0
50
100
150
200
250
Nur kommerziellesMischfutter
(n=6)
Selbstmisch-betriebe(n=18)
(mg
Cu/k
g T)
Zink Kupfer

Windisch et al.: Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung?
8. BOKU-Symposium Tierernährung 2009 Seite 43
Literatur
Aichberger, K., Döberl, H., Eibelhuber, A., Froschauer, J., Hofer, G. (1995). Schriftenreihe der Abteilung Umweltschutz 7/95, Hrsg. Amt der Oberösterreichischen Landesregierung: Linz, Österreich.
Apgar, G.A., Kornegay, E.T., Lindemann, M.D., Notter, D.R. (1995): Evaluation of copper sulfate and a copper lysine complex as growth promoters for weanling swine. J. Anim. Sci. 73: 2640–2646.
Apgar, G.A., Kornegay, E.T. (1996): Mineral balance of finishing pigs fed copper sulfate or a copper- lysine complex at growth-promoting levels. J. Anim. Sci. 74: 1594–1600.
BIOABF (1998): Verordnung über die Verwertung von Bioabfällen auf landwirtschaftlich, forstwirtschaftlich und gärtnerisch genutzten Böden (Bioabfallverordnung - BIOABFV) vom 21. September 1998.
Bonnier, P. (2002): Anforderungen an die landwirtschaftliche Verwertung von Klärschlamm und anderen Düngemitteln in den Niederlanden – Gründe und Konsequenzen. In KTBL Schrift 404: 265–270. KTBL: Darmstadt, Deutschland.
Carlson, M.S., Hill, G.M., Link, J.E. (1999): Early and traditional weaned nurserey pigs benefit from phase-feeding pharmacol-ogical concentrations of zinc oxide: Effect on metallothionein and mineral concentrations. J. Anim. Sci. 77: 1199–1207.
Coffey, R.D., Cromwell, G.L., Monegue, H.J. (1994): Efficacy of a copper-lysine complex as a growth promoter for weanling pigs. J. Anim. Sci. 72: 2280–2886.
Commission Regulation (EC) No 1334/2003: Amending the conditions for authorisation of a number of additives in feedingstuffs belonging to the group of trace elements.
Cromwell, G.L., Stahly, T.S., Monegue, H.J. (1989): Effects of source and level of copper on performance and liver copper stores in weanling pigs. J. Anim. Sci. 67: 2996–3002.
Dannenberg, O. (1997): Bodenschutz in Österreich. Fachbeirat für Bodenfruchtbarkeit und Bodenschutz beim Bundesministerium für Land- und Forstwirtschaft, Wien, Österreich: 6-7.
Dersch, G., Hösch, J. (2003): Kupfer und Zink – In Zukunft stärker beachten. Der fortschrittliche Landwirt 11: 6–9.
Döhler, H., Henning, E., Uihlein, A., Wilcke, W. (2002): Bilanzierung von Spurenelementen bzw. Schwermetallen in der Milchviehhaltung, KTBL Schrift 410: 59–65. KTBL: Darmstadt, Deutschland.
Eckel, H., Roth, U., Döhler, H., Schultheiss, U. (2007): Assessment and reduction of heavy metal input into agro-ecosystems. In: Schlegel, P., Durosoy, S., Jongbloed, A.W. (Eds.), Trace elements in animal production systems, 187-201, Wageningen Academic Publishers, Wageningen,The Netherlands, pp 33-44
GfE (1999): Empfehlungen zur Energie- und Nährstoffversorgung der Legehennen und Masthühner (Broiler). Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie. Frankfurt am Main. DLG-Verlag.
GfE (2001): Empfehlungen zur Energie- und Nährstoffversorgung der Milchkühe und Aufzuchtrinder. Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie. Frankfurt am Main. DLG-Verlag.
GfE (2006): Empfehlungen zur Energie- und Nährstoffversorgung von Schweinen. Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie. Frankfurt am Main. DLG-Verlag.
Hahn, J.D., Baker, D.H. (1993): Growth and plasma zinc responses of young pigs fed pharmacological doses of zinc. J. Anim. Sci. 71: 3020–3024.
Hackenberg, S., Wegener, H.-R., Eurich-Menden, B. (1996): Herkunft der Schadstoffe in Komposten, Schadstoffgehalte in Komposten und anderen Dünge- und Bodenverbesserungsmitteln, Vor- und Nachteile beim Einsatz von Komposten in der Land- und Forstwirtschaft sowie im Landschafts- und Weinbau. Forschungsbericht Justus Liebig Universität Gießen, Deutschland.
Kickinger, T., Humer, J., Aichberger, K., Würzner, H., Windisch, W. (2008): Survey on zinc and copper contents in dung from Austrian livestock production. Bodenkultur 59, 101-110.
Kickinger, T., Würzner, H., Windisch, W. (2009): Zinc and copper in feeds, manure and soils from Austrian pig fattening farms feeding commercial complete feed or farm-based feed mixtures. Bodenkultur, submitted.
Kilian, W., (1999): Richtlinien für die sachgerechte Düngung. Wien: Bundesamt und Forschungszentrum für Landwirtschaft: 339-354.
Kühnen, V., Goldbach, H.E. (2002): Schwermetallbilanzen verschiedener Betriebstypen: Eintragswege, Flüsse, Minderungspotential. Forschungsbericht Nr. 118, Institut für Pflanzenernährung, Rheinische Friedrichs-Weilhelms-Universität Bonn, Deutschland.
Müller, C. (1997): Wirtschaftsdünger. In: Boden-Dauer-Beobachtungsflächen (BDF) - Bericht nach 10-jähriger Laufzeit 1985-1995. Teil II: Stoffeinträge – Stoffausträge – Schwermetallbilanzierungen verschiedener Betriebstypen. Schriftenreihe der bayrischen Landesanstalt für Bodenkultur und Pflanzenbau, 5/97 (Jg. 1).: 87–117.

Windisch et al.: Zink und Kupfer im Wirtschaftsdünger: Ein Problem der Tierernährung?
Seite 44 8. BOKU-Symposium Tierernährung 2009
Müller, C., Ebert, T. (2002): Schwermetall-Einträge durch Wirtschaftsdünger von 1996 bis heute – Ergebnisse aus dem bayerischen Bodenbeobachtungsprogramm. VDLUFA-Schriftenreihe 58: 635-639.
Nicholson, F., Boghal, A., Roth, U., Schultheiss, U. (2005): Assessment and reduction of heavy metal input into agro-ecosystems, KTBL Schrift 432: 43 – 53. KTBL: Darmstadt, Deutschland
Poulsen, H.D. (1995): Zinc oxide for weanling piglets. Acta Agric. Scand. 45: 159–167
Roth, U., Schultheiss, U., Döhler, H., Eckel, H., Kühnen, V., Früchtenicht, K., Uihlein, A. (2002): Spurenelement- bzw. Schwermetallgehalte in Futtermitteln und Wirtschaftsdüngern, KTBL Schrift 410: 50–59. KTBL: Darmstadt, Deutschland.
Smith, J.W., Tokach, M.D., Goodband, R.D., Nelssen J.L., Nessmith, W.B., Owen, K,Q., Richert, B.T. (1995): The effect of in-creasing zinc oxide supplementation on starter pig growth performance. J. Anim. Sci. 73 (abstr.): 72.
Schwarz, S., Freudenschuss, A. (2004): Referenzwerte für Schwermetalle in Oberböden. Auswertungen aus dem österreichweiten Bodeninformationssystem BORIS. Monographien M-170, Umweltbundesamt, Wien, Österreich.
UBA (Umweltbundesamt) (2001): Grundsätze und Maßnahmen für eine vorsorgeorientierte Begrenzung von Schadstoffeinträgen in landbaulich genutzten Böden. UBA, Berlin, Deutschland. UBA Texte 59–01.
UBA (Umweltbundesamt) (2004): Erfassung von Schwermetallströmen in landwirtschaftlichen Tierproduktionsbetrieben. UBA Berlin, Deutschland. UBA Texte 06–04.
Windisch, W., Schwarz, F.J., Gruber, K., Kirchgessner, M. (1999): Effect of Pharmacological Dietary Doses of Zinc Oxide on Performance and Fecal Characteristics of Weanling Piglets. Agribilol. Res. 51, 277–285.
Windisch, W., Gotterbarm, G.G., Roth, F.X. (2001): Effect of potassium diformate in combination with different amounts and sources of excessive dietary copper on production performance of weaning piglets. Arch. Anim. Nutr. 54: 87–100.
Autorenanschrift
Univ.Prof. DI Dr. Wilhelm Windisch Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Department für Lebensmittelwissenschaften und –technologie Universität für Bodenkultur Wien Gregor Mendel-Straße 33, 1180 Wien, Österreich E-Mail: [email protected]

Würzner: (Ent)warnung für Schwermetalle in Futtermitteln
8. BOKU-Symposium Tierernährung 2009 Seite 45
(Ent)warnung für Schwermetalle in Futtermitteln
Herbert Würzner
Österreichische Agentur für Gesundheit und Ernährungssicherheit, Wien
Einleitung Zu den Schwermetallen zählt man chemisch alle Elemente mit einem spezifischen Gewicht von über 5 g/cm3. In der Umwelt kommen sie meist in nur sehr geringen Spuren vor. Gemeinsam ist allen, dass sie in zu hohen Konzentrationen toxisch wirken. Die Schwermetalle Blei (Pb), Quecksilber (Hg), Cad-mium (Cd), Arsen (As) gehören in der Tierernährung und futtermittelrechtlich in die Gruppe der uner-wünschten Stoffe, da keine essentielle Wirkung für den Stoffwechsel bekannt ist und höhere Gehalte für Mensch, Tier und Umwelt schädlich sein können. Im Gegensatz dazu zählen Kupfer (Cu), Eisen (Fe), Zink (Zn), Mangan (Mn), Selen (Se), Kobalt (Co) und Molybdän (Mo) zu den essentiellen Spuren-elementen, die meist in geringen Mengen in Mischfuttermitteln zugesetzt werden, jedoch alle als Zu-satzstoffe mit einem gesetzlichen Höchstwert und unter bestimmten Bedingungen zugelassen sind.
Rechtliche Rahmenbedingungen Zur Tierernährung bestimmte Erzeugnisse können unerwünschte Stoffe enthalten, ihr Vorkommen lässt sich nicht völlig ausschließen. Um schädliche Wirkungen auf die Gesundheit von Tier und Mensch sowie auf die Umwelt zu minimieren, sieht die einschlägige Gesetzgebung für die wichtigsten uner-wünschten Elemente Höchstgehalte in Futtermitteln, Zusatzstoffen und Vormischungen vor. Die zuläs-sigen Höchstgehalte werden in der Richtlinie 2002/32/EG für unerwünschte Stoffe in der Tierernäh-rung geregelt und sind dort nach dem Verwendungszweck des Futters und der vorgesehenen Tierart abgestuft.
Verdünnungsverbot: Zur Tierernährung bestimmte Erzeugnisse mit einem über dem Grenzwert liegenden Gehalt dürfen nicht zu Verdünnungszwecken mit dem gleichen oder mit einem anderen zur Tierernährung bestimmten Erzeugnis gemischt werden (Artikel 5 der o. a. Richtlinie 2002/32).
Ergänzungsfuttermittel: Auch Ergänzungsfuttermittel dürfen, soweit keine Sonderbestimmungen gelten, unter Berücksichtigung ihres in einer Tagesration vorgeschriebenen Anteils, keine höheren Gehalte aufweisen, als sie für Alleinfuttermittel festgesetzt sind (Artikel 6 der o. a. Richtlinie 2002/32).
„Schutzklausel“: Hat ein Mitgliedstaat aufgrund neuer Informationen oder Erkenntnisse Gründe für die Annahme, dass ein festgesetzter Höchstgehalt eine Gefahr für die Gesundheit von Mensch oder Tier oder für die Umwelt darstellt, so kann er den bestehenden Höchstgehalt vorläufig herabsetzen (Vorsorgeprinzip !). Er teilt dies unter Angabe der Gründe unverzüglich der Kommission und den ande-ren Mitgliedstaaten mit (Artikel 7 der o. a. Richtlinie 2002/32).

Würzner: (Ent)warnung für Schwermetalle in Futtermitteln
Seite 46 8. BOKU-Symposium Tierernährung 2009
Tabelle 1: Höchstgehalte für unerwünschte Elemente in Futtermitteln (mg/kg FM mit 12 % Feuchte), Auszug aus Richtlinie 2002/32/EG, Anhang I (konsolidierte Fassung von 2009)
Arsen Blei Cadmium
Quecksilber
Futtermittel-Ausgangserzeugnisse, ausgenommen Grünmehl u.a. Phosphate u.a. Calciumcarbonat Magnesiumoxid .. aus Fischen .. aus Seealgen
2 4 10 15 20 15 40
10
30 15 20 - -
1 (pfl. Urspr.) 2 (tier. Urspr.) 2 (mineral. Urspr.)
10 - - -
0,1
- -
0,3 -
0,5 -
Alleinfuttermittel, ausgenommen für Fische, Pelztiere Rinder, Schafe, Ziegen Heimtiere Kälber, Schaf- und Zie-genlämmer und sonstige
2 6 - - -
5 - - - -
1 1 2
0,5
0,1 - -
0,4 -
Ergänzungs-futtermittel, ausgenommen Mineralfuttermittel
4
12
10
15
0,5
< 7% P 5 > 7% P 0,75 je % P,
max.7,5
0,2
-
Zusatzstoffe Spurenelemente Zinkoxid Manganoxid u.a. Bindemittel
- 100 400 200 30
10 30 30 2
-
Vormischungen - 200 15 -
Situation in Österreich - Ergebnisse Die Futtermittelkontrollen werden auf Grund eines statistisch abgesicherten, risikobasierten Stich-probenplanes durchgeführt. Zusätzlich werden von der Futtermittelkontrolle nachfassende- und ad-hoc-Kontrollen umgesetzt, welche ebenfalls im Kontrollplan abgebildet sind. Die nachfassenden Kontrollen erfassen Betriebe mit Beanstandungen aus dem Vorjahr bzw. Missstände aus Vorperio-den, welche erneut im Kontrollplan berücksichtigt werden.
Informationen aus dem Europäischen Schnellwarnsystem, Zollmeldungen, sowie Informationen aus laufenden Kontrollen bzw. zu futtermittelrechtlichen Aspekten finden als ad-hoc-Kontrollen ihren Eingang in den Kontrollplan. Nach diesem sind derzeit jährlich rund 600 Proben zur Untersuchung auf Schwermetalle vorgesehen.
In der folgenden Tabelle 2 werden die Analysenergebnisse der unerwünschten Elemente As, Pb, Cd und Hg, in Form der Grenzwert-Überschreitungen, in den letzten Jahren dargestellt. Bei allen vier Schadelementen zeigen die Datensätze eine stark asymmetrische Verteilung, was bedeutet, dass der überwiegende Anteil an Kontrollproben niedrige oder sehr niedrige Schwermetallwerte aufweist und

Würzner: (Ent)warnung für Schwermetalle in Futtermitteln
8. BOKU-Symposium Tierernährung 2009 Seite 47
nur wenige Proben durch höhere Konzentrationen bzw. Höchstgehaltüberschreitungen auffallen. Viele Gehalte liegen im Bereich der methodischen Nachweisgrenze und die Medianwerte sind speziell bei Allein- und Ergänzungsfuttermitteln mit den Gehalten von unbelastetem Getreide, Obst und Gemüse vergleichbar. Generell ist festzustellen, dass die Schwermetallkonzentrationen in Abhängigkeit vom Futtermitteltyp variieren und in der Reihenfolge Alleinfuttermittel < Ergänzungsfutter < Mineralfutter < Vormischungen ansteigen. Die höheren Pb-, Cd und As-Gehalte in Mineralfutter und Vormischungen sind weitgehend auf die eingesetzten Rohphosphate, Futterkalke oder sonstigen zugesetzten Mineral-stoffe und Spurenelementverbindungen zurückzuführen. Die vorgeschriebenen Höchstgehalte werden bei Quecksilber praktisch nie, bei Arsen, Blei und Cadmium nur in Ausnahmefällen überschritten.
Tabelle 2: Anzahl der Untersuchungen auf Arsen, Blei, Cadmium und Quecksilber (2004 – 2008). In Klammer die Anzahl der Beanstandungen wegen Überschreitung des gesetzlichen Höchstwertes.
As Pb Cd Hg 2004 384 (0) 408 (0) 534 (0) 116 (0) 2005 283 (1) 434 (0) 434 (0) 107 (0) 2006 198 (0) 353 (1) 353 (0) 88 (0) 2007 341 (0) 396 (0) 280 (0) 160 (0) 2008 741(2) 608 (0) 608 (1) 320 (0)
In den letzten Jahren gab es 2005 und 2006 je eine einzige Überschreitung (= 0,3 %) des gesetzli-chen Höchstgehaltes von Arsen in Pferdefutter bzw. Blei in Manganoxid. 2008 ergaben sich insgesamt 3 Überschreitungen (= 0,2 %), einmal in Form von Cadmium in Dicalciumphosphat, sowie Arsen ein-mal in Calciumcarbonat und einmal in einem Ergänzungsfuttermittel für Schweine.
2004 und 2007 gab es keine Überschreitungen.
Das jährliche Rohstoff-Monitoring auf unerwünschte Stoffe, zu dem wir von der Mischfutterwirtschaft beauftragt werden, zeigte 2008 bei 35 Proben auf jeweils diese 4 Schwermetalle ebenfalls keine Überschreitungen der gesetzlichen Grenzwerte.
Aus den Ergebnissen der Futtermittelkontrolle ist zu schließen, dass Futtermittel im allgemeinen sehr gering mit Schwermetallen belastet sind und die vorgeschriebenen Höchstgehalte weitgehend ein-gehalten werden.
Importprodukte und Europa Zum Vergleich möchte ich die Jahresstatistik 2008 über die amtliche Futtermittelüberwachung in Deutschland anführen, wo für die gleichen vier Schwermetalle bei rund 12.000 Untersuchungen nur 0,2 % Beanstandungen zu verzeichnen waren.
Dass es noch andere Ergebnisse auf diesem Gebiet gibt, zeigen die Meldungen aus dem europäischen Schnellwarnsystem RASFF, die meistens Importe verschiedener Mineralstoffe und Spurenelement-verbindungen betreffen. Beispielhaft sollen einige dieser Fälle, anonymisiert, in der Tabelle 3 ange-führt werden.
Das in der letzten Spalte angeführte Land muss nicht immer das Ursprungsland sein, sondern kann auch das Land, wo die Kontamination eines importierten Produktes festgestellt und gemeldet wurde, sein.
Die Fälle aus dem Schnellwarnsystem betrafen meist Cadmium (8x), 4x Blei und nur jeweils einmal Arsen und Quecksilber. Bezogen auf alle Meldungen im Futtermittelbereich hatten die vier wichtigsten Schwermetalle in den letzten Jahren nur einen Anteil von insgesamt etwa 2 bis 4 %.

Würzner: (Ent)warnung für Schwermetalle in Futtermitteln
Seite 48 8. BOKU-Symposium Tierernährung 2009
Tabelle 3: Meldungen betr. Schwermetallfälle in Futtermitteln (2007 – Juli 2009)
Nr. / Jahr Schwer-metall
Gehalt (mg/kg)
Gesetzlicher Grenzwert Produkt Ursprungsland
Jahr 2009 (bis Juli)
1007 Arsen 22 10 Austernschalen NL, BE 446 Cadmium 3,86 2 Fischmehl ESP 40 Quecksilber 0,24 0,1 Alleinfuttermittel F
248 Cadmium 12,4 10 Dicalciumphosphat PT 19 Cadmium 2,5/3,2 2 Tintenfischmehl Peru 35 Fluor 2500 2000 Monocalciumphosphat F
Jahr 2008 1484 Blei 105,8 30 Sepiolith ESP 1675 Cadmium 15,4 10 Dicalciumphosphat SLO 1065 Blei 22 10 Reisprotein Pakistan
746 Cadmium 357/454/ 349/173 30 Zinkoxid China
Jahr 2007 402 Blei 196 100 Kupfersulfat Russland BKD Blei 571 400 Zinkoxid Türkei
314 Cadmium 16/13,4/
12,1 10 Zinksulfat D/China 196 Cadmium 966 10 Zinksulfat (technisch) SK/PL AAR Cadmium 19,5 10 Monocalciumphosphat Türkei
Ein besonders gravierender Fall von Cadmium in Zinksulfat, den uns die Europäische Kommission im August zugespielt hat, möge beispielhaft für Ereignisse in China zeigen, was sich dort auf diesem Ge-biet abspielen kann:
Please find enclosed a media report making mention of high cadmium pollution in a zinc sul-phate manufacturing company (Changsha Xianghe Chemical plant). As can be seen in the media report, the company was ordered by the Chinese authorities to stop production in April 2009.
Nevertheless, it is appropriate and necessary to verify if zinc sulphate from that company has been imported into the EU and if this is the case, to control (or re-control) the zinc sulphate for the presence of cadmium before making further use of it for the production of feed. I remind you of the import in previous years of consignments of zinc sulphate originating from China containing very high levels of cadmium.
From Internet, following details from the concerned Chinese company could be found:
"China: Cadmium Pollution at Zinc Sulphate Facility Sickens Over 500, Kills 2 Source: Feedinfo News Ser-vice (dated 03/08/2009)
3 August 2009 - According to reports in the Chinese media today, 509 people have been sickened by pollution from a chemical plant in Liuyang country, Hunan province, which has also been blamed for two deaths.
It is understood that health checks carried out on nearly 3,000 people living near the plant revealed 509 had excessive levels of the toxic heavy metal cadmium - 33 of which are in need of hospitalisation.

Würzner: (Ent)warnung für Schwermetalle in Futtermitteln
8. BOKU-Symposium Tierernährung 2009 Seite 49
It is also believed that the cadmium may have been leaking for months before the two villagers died in May and June.
Local authorities have suspended two environmental officials and detained the head of the plant following a protest last week by 1,000 residents, reports the media.
The plant, owned by Changsha Xianghe Chemical, was ordered to stop production in April after the authorities discovered that manufacturing operations at the plant produced cadmium pollution. They said areas within a 1,200-meter radius of the factory were tainted.
The Changsha Xianghe Chemical plant mainly manufactures zinc sulphate, a trace element used in animal feed. According to a company factsheet, Changsha Xianghe Chemical distributes its zinc sulphate to North America, Asia, Europe and Oceania."
Schlussfolgerungen Dank eines funktionierenden, ausgeklügelten Kontroll- und Informationssystems in der Europäischen Union, werden mit Schwermetallen manchmal belastete Rohstoffe wie Mineralien, Fischmehl u. a. entweder schon an der Grenze oder bei Inlandskontrollen entdeckt und gelangen so nicht oder nur selten in die Futtermittelkette. Frühere Quellen aus Industrien im Inland wurden schon vor mehreren Jahren beseitigt, sodass man heute sagen kann, dass, bei Einhaltung einer guten Herstellungspraxis mit Eigenkontrollen sowie einer funktionierenden amtlichen Kontrolle im Land und an der Grenze, praktisch kein Risiko durch Schwermetall-Kontaminationen in Futtermitteln besteht.
Literatur
AGES – Österreichische Agentur für Gesundheit und Ernährungssicherheit GmbH (2009): Kontrollierte Futtermittel, gesunde Tiere, sichere Lebensmittel. http://www.ages.at/ages/landwirtschaftliche-sachgebiete/futtermittel/publikationen-vortraege/ futtermittel-broschuere/
RICHTLINIE 2002/32/EG über unerwünschte Stoffe in der Tierernährung. ABl. L 140 vom 30.05.2002, konsolidierte Fassung vom 20.04.2009 http://europa.eu/index_de.htm
Europ. Schnellwarnsystem – RASFF http://ec.europa.eu/food/food/rapidalert/index_en.htm
und http://ec.europa.eu/food/food/rapidalert/rasff_portal_database_en.htm
Jahresstatistik 2008 über die amtliche Futtermittelüberwachung in Deutschland (2009): http://www.bmelv.de/futtermittel
Autorenanschrift
Univ. Doz. Dipl.-Ing. Dr. Herbert Würzner Institut für Futtermittel Österreichische Agentur für Gesundheit und Ernährungssicherheit Bundesamt für Ernährungssicherheit Spargelfeldstrasse 191, A-1220 Wien E-Mail: [email protected]

Firmensponsoring – AGRANA Bioethanol GmbH, Industriegelände Pischelsdorf, A-3435 Pischelsdorf
Seite 50 8. BOKU-Symposium Tierernährung 2009
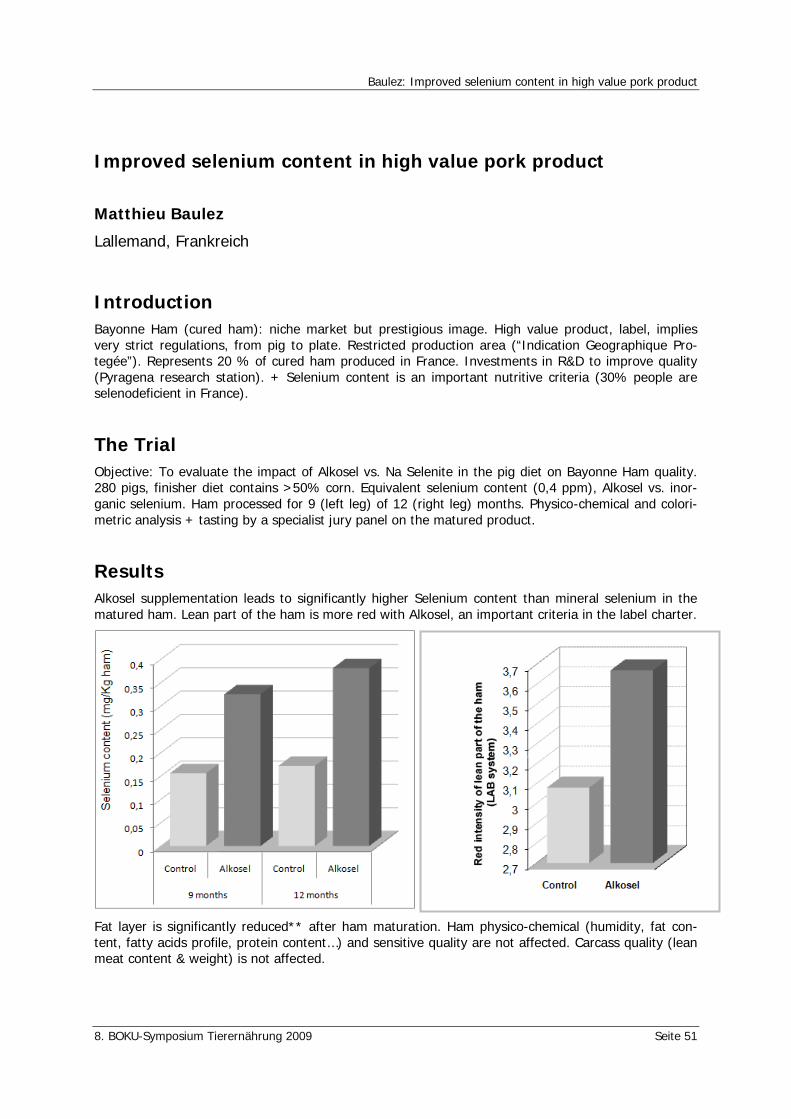
Baulez: Improved selenium content in high value pork product
8. BOKU-Symposium Tierernährung 2009 Seite 51
Improved selenium content in high value pork product
Matthieu Baulez
Lallemand, Frankreich
Introduction Bayonne Ham (cured ham): niche market but prestigious image. High value product, label, implies very strict regulations, from pig to plate. Restricted production area (“Indication Geographique Pro-tegée”). Represents 20 % of cured ham produced in France. Investments in R&D to improve quality (Pyragena research station). + Selenium content is an important nutritive criteria (30% people are selenodeficient in France).
The Trial Objective: To evaluate the impact of Alkosel vs. Na Selenite in the pig diet on Bayonne Ham quality. 280 pigs, finisher diet contains >50% corn. Equivalent selenium content (0,4 ppm), Alkosel vs. inor-ganic selenium. Ham processed for 9 (left leg) of 12 (right leg) months. Physico-chemical and colori-metric analysis + tasting by a specialist jury panel on the matured product.
Results Alkosel supplementation leads to significantly higher Selenium content than mineral selenium in the matured ham. Lean part of the ham is more red with Alkosel, an important criteria in the label charter.
Fat layer is significantly reduced** after ham maturation. Ham physico-chemical (humidity, fat con-tent, fatty acids profile, protein content…) and sensitive quality are not affected. Carcass quality (lean meat content & weight) is not affected.

Baulez: Improved selenium content in high value pork product
Seite 52 8. BOKU-Symposium Tierernährung 2009
Corresponding author
Matthieu Baulez, Monogastric Product Manager Lallemand SAS 19, rue des Briquetieres 31702 Blagnac Cedex, France E-mail: [email protected]

Firmensponsoring – Metall und Farben Ges.m.b.H., Ottakringerstraße 89, A-1160 Wien
8. BOKU-Symposium Tierernährung 2009 Seite 53
A-1
160
Wie
n/A
ust
ria
. Ott
akri
nge
rstr
. 89

Boulianne et al.: Effect of organic selenium on growth performances, feathering, skin strength and cellulitis prevalence of challenged broiler chickens
Seite 54 8. BOKU-Symposium Tierernährung 2009
Effect of organic selenium on growth performances, feathering, skin strength and cellulitis prevalence of challenged broiler chickens
Martine Boulianne1, Ghislaine Roch2, Matthieu Baulez3 1 Universität Montreal 2 Nutritionnist 3 Lallemand, Frankreich
Introduction Most of broilers ‘breeds used in North America are slow feathering, with short feathers and a low feathers weight at 48 days (Ajang et al. Br Poult Sci 34:73-91, 1993). Moreover, these birds have a poor skin resistance and elasticity (Granot et al. Poult Sci 70:1928-1935, 1991) and are thus more prone to skin scratches that might then be contaminated with Escherichia coli causing dermatitis.
It has been shown that organic Se yeast can have an influence on birds feathering and skin resistance providing a better protection of the animal against skin scratches.
Protocol Animals: 672 day-old broiler males were placed in 21 pens of 32 birds each.
Treatments: 3 groups receiving 0.3 ppm inorganic Se (A), or 0.15 ppm organic Se and 0.15 ppm inor-ganic Se (B), or 0.3 ppm organic Se (C).
Period of supplementation: Se added into the diet from day one to slaughter.
Measurements: daily pen feed consumption, weekly individual body weights, skin scratches-dorsoscapular-hip feather coverage and skin tensile strength and elasticity.

Boulianne et al.: Effect of organic selenium on growth performances, feathering, skin strength and cellulitis prevalence of challenged broiler chickens
8. BOKU-Symposium Tierernährung 2009 Seite 55
Results
Conclusion Feeding organic Se alone or in combination improved the average daily gain and feed conversion be-tween 28 and 35 days of age, as well as feed conversion between 0 and 35 days of age.
It also increased hip feather coverage at 22 and 29 days of age and skin resistance at 35 days of age, while improving skin elasticity.

Boulianne et al.: Effect of organic selenium on growth performances, feathering, skin strength and cellulitis prevalence of challenged broiler chickens
Seite 56 8. BOKU-Symposium Tierernährung 2009
Because only two birds from groups A and B were condemned for cellulitis, it was not possible to con-clude on the efficacy of the treatment to prevent this disease. However, since feathering and skin resistance are important risk factors involved in the development of cellulitis, supplementation with organic Se could potentially offer protection against this disease.
Corresponding author
Matthieu Baulez Lallemand SAS 19, rue des Briquetieres 31702 Blagnac Cedex, France E-mail: [email protected]

Bourdonnais: Intestinal absorption of zinc from zinc sulfate and zinc amino acid chelates in growing pigs
8. BOKU-Symposium Tierernährung 2009 Seite 57
Intestinal absorption of zinc from zinc sulfate and zinc amino acid chelates in growing pigs
Alain Bourdonnais
Albion Animal Nutrition, France
Abstract Three young pigs were surgically fitted with catheters to evaluate zinc absorption from various zinc sources. The study design was a 3X3 Latin square design. Two studies were conducted. The first study compared the absorption of zinc sulfate, zinc methionine chelate, and zinc amino acid chelate. It was found in this study that both the organic chelates had higher net portal absorption as determined by plasma zinc content than zinc sulfate over a period of 210 minutes. In the second study, two ratios of zinc sulfate and zinc amino acid chelate were compared to zinc sulfate alone. It was found that an equal mixture of inorganic zinc sulfate and zinc amino acid chelate had the greatest net portal absorp-tion. It was concluded that zinc amino acid chelates are better absorbed as compared to zinc sulfate.
Introduction Amino acid chelates have been used extensively as delivery systems for essential mineral nutrients. These compounds are characterized by an amino acid joined to a metal ion by both the oxygen from the carboxyl and the nitrogen from the amine group. This joint bonding results in a heterocyclic ring structure which is a distinctive feature of amino acid chelates. All naturally occurring amino acids have the same common backbone, and are capable of bonding metals in a similar fashion. Metal:ligand molar ratios can range from 1:1 to 3:1.1 A drawing of metal bisglycinate chelate is shown in Figure 1.
Figure 1. Diagram of metal bisglycinate chelate.

Bourdonnais: Intestinal absorption of zinc from zinc sulfate and zinc amino acid chelates in growing pigs
Seite 58 8. BOKU-Symposium Tierernährung 2009
In recent studies amino acid chelates have been shown to be more bioavailable in a broiler chick model.2 Iron amino acid chelates have been shown to increase reproduction efficiency in pigs as de-teremined by farrowing weight, mortality rates, and increased birth weights.3 Due to its unique chemi-cal properties, amino acid chelates have the capability to deliver minerals even under dietary chal-lenges of mineral antagonists.4 Additionally, due to their increased bioavailability, amino acid chelates have the potential of having a lesser impact on the environment than comparable inorganic minerals.5
Two studies were conducted to compare the net portal absorption (NPA) of zinc from various zinc sources in growing pigs. The first study compared two sources of zinc amino acid chelates to zinc sulfate. The second study compared various ratios of a zinc amino acid chelate and zinc sulfate. Both studies were similarly conducted.
Methods Three pigs (21.5±0.7 kg BW) were surgically fitted with catheters into carotid artery, portal vein, mesenteric vein, and pyloric stomach (See Figure 2.) and allotted to 3 x 3 Latin square design with 3 treatments. Each pig received treatment by injection through the pyloric stomach catheter and 3 peri-ods (48-h intervals). Each period was composed of 24-h feeding (0.09 kg x BW0.75), 19.5-h fasting, and 4.5-h infusion. A corn-soybean meal based diet with 18.2% CP and 3.35 Mcal ME/kg was fed to pigs before fasting. Para-aminohippuric acid (PAH) was infused (3.2 mg/min) into the mesenteric vein for a 4.5 h period. Zinc (230 mg) from one of the three treatments was injected into the lumen of the pyloric catheter 60 min after beginning the PAH infusion period. Blood samples (3 mL) were collected simultaneously from the carotid artery and portal vein catheters at -60, -30, 0, 15, 30, 45, 60, 90, 120, 150, and 210 min relative to zinc injection to measure PAH and zinc concentration in the plasma. Zinc NPA was calculated by multiplying the portal vein plasma flow rate by the porto-arterial plasma zinc concentration.
Figure 2. Schematic of catheter placement.
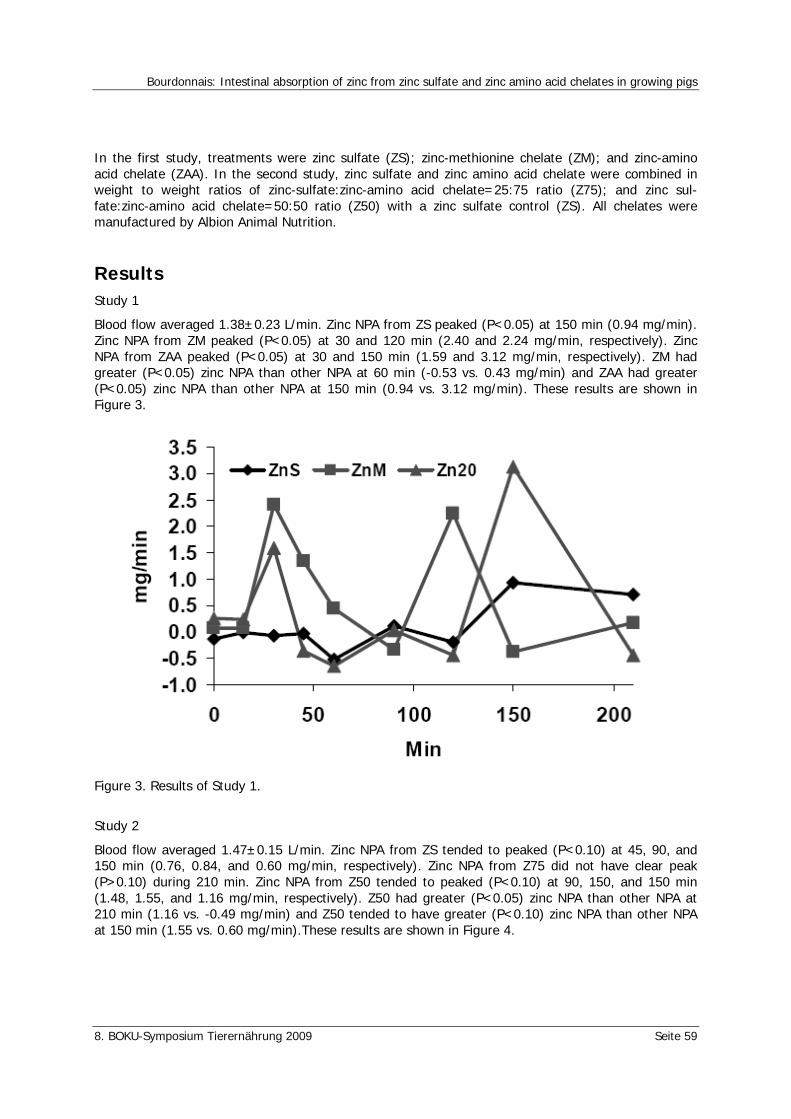
Bourdonnais: Intestinal absorption of zinc from zinc sulfate and zinc amino acid chelates in growing pigs
8. BOKU-Symposium Tierernährung 2009 Seite 59
In the first study, treatments were zinc sulfate (ZS); zinc-methionine chelate (ZM); and zinc-amino acid chelate (ZAA). In the second study, zinc sulfate and zinc amino acid chelate were combined in weight to weight ratios of zinc-sulfate:zinc-amino acid chelate=25:75 ratio (Z75); and zinc sul-fate:zinc-amino acid chelate=50:50 ratio (Z50) with a zinc sulfate control (ZS). All chelates were manufactured by Albion Animal Nutrition.
Results Study 1
Blood flow averaged 1.38±0.23 L/min. Zinc NPA from ZS peaked (P<0.05) at 150 min (0.94 mg/min). Zinc NPA from ZM peaked (P<0.05) at 30 and 120 min (2.40 and 2.24 mg/min, respectively). Zinc NPA from ZAA peaked (P<0.05) at 30 and 150 min (1.59 and 3.12 mg/min, respectively). ZM had greater (P<0.05) zinc NPA than other NPA at 60 min (-0.53 vs. 0.43 mg/min) and ZAA had greater (P<0.05) zinc NPA than other NPA at 150 min (0.94 vs. 3.12 mg/min). These results are shown in Figure 3.
Figure 3. Results of Study 1.
Study 2
Blood flow averaged 1.47±0.15 L/min. Zinc NPA from ZS tended to peaked (P<0.10) at 45, 90, and 150 min (0.76, 0.84, and 0.60 mg/min, respectively). Zinc NPA from Z75 did not have clear peak (P>0.10) during 210 min. Zinc NPA from Z50 tended to peaked (P<0.10) at 90, 150, and 150 min (1.48, 1.55, and 1.16 mg/min, respectively). Z50 had greater (P<0.05) zinc NPA than other NPA at 210 min (1.16 vs. -0.49 mg/min) and Z50 tended to have greater (P<0.10) zinc NPA than other NPA at 150 min (1.55 vs. 0.60 mg/min).These results are shown in Figure 4.
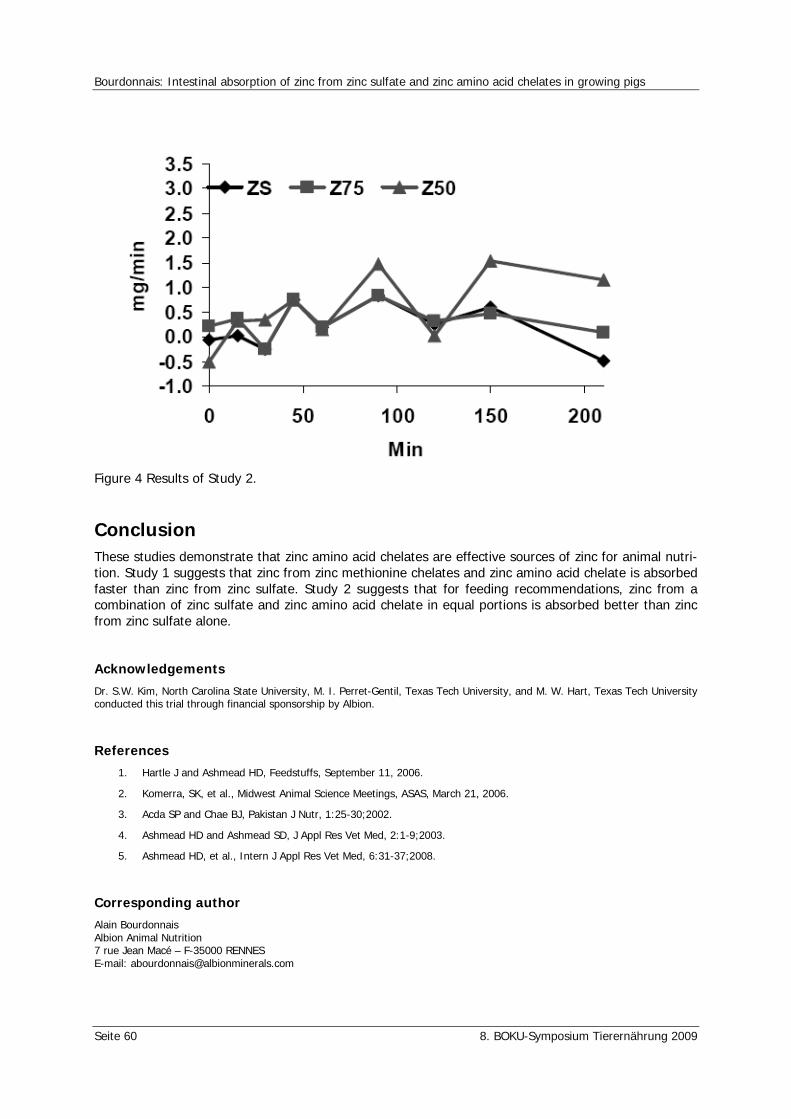
Bourdonnais: Intestinal absorption of zinc from zinc sulfate and zinc amino acid chelates in growing pigs
Seite 60 8. BOKU-Symposium Tierernährung 2009
Figure 4 Results of Study 2.
Conclusion These studies demonstrate that zinc amino acid chelates are effective sources of zinc for animal nutri-tion. Study 1 suggests that zinc from zinc methionine chelates and zinc amino acid chelate is absorbed faster than zinc from zinc sulfate. Study 2 suggests that for feeding recommendations, zinc from a combination of zinc sulfate and zinc amino acid chelate in equal portions is absorbed better than zinc from zinc sulfate alone.
Acknowledgements
Dr. S.W. Kim, North Carolina State University, M. I. Perret-Gentil, Texas Tech University, and M. W. Hart, Texas Tech University conducted this trial through financial sponsorship by Albion.
References
1. Hartle J and Ashmead HD, Feedstuffs, September 11, 2006.
2. Komerra, SK, et al., Midwest Animal Science Meetings, ASAS, March 21, 2006.
3. Acda SP and Chae BJ, Pakistan J Nutr, 1:25-30;2002.
4. Ashmead HD and Ashmead SD, J Appl Res Vet Med, 2:1-9;2003.
5. Ashmead HD, et al., Intern J Appl Res Vet Med, 6:31-37;2008.
Corresponding author
Alain Bourdonnais Albion Animal Nutrition 7 rue Jean Macé – F-35000 RENNES E-mail: [email protected]

Firmensponsoring – Albion Animal Nutrition International, 7 Rue Jean Macé, FR-35000 Rennes
8. BOKU-Symposium Tierernährung 2009 Seite 61

Chrenková et al.: The effect of heat treatment on phosphorus releasing from soya bean and pea
Seite 62 8. BOKU-Symposium Tierernährung 2009
The effect of heat treatment on phosphorus releasing from soya bean and pea
M. Chrenková, Z. Čerešňáková, P. Fľak, M. Poláčiková
Research Center of Animal Production – Department of Nutrition Nitra, Slovak Re-public
Introduction Important variation in phosphorus and calcium release kinetics was found among feeds (Südekum et al. 1999; Flachowsky et al. 1994; Čerešňáková et al. 2005; Čerešňáková et al. 2007a, b). Not only high fibre diets have the adverse effect on the bioavailability of minerals and trace elements but also phytate negatively affected P and Ca availability. Kenefick and Cashman (2000) determined that phy-tate in wheat fibre extract is the major inhibitory factor of Ca absorption. Rumen microbs produce phytase effectivevely release P from phytate (Morse, 1992).
Treatment of feeds by heating and formaldehyde suppressed not only ruminal crude protein degrada-tion but also releasing of P from wheat middlings, soybean meal, corn distillers grains, etc., and many authors (Morse 1992; Park et al. 1999; Bravo et al. 2000, Matsui 2002) explained this proccess of phytate degradation.
In our experiment we have used in sacco and mobile bag techniques for P release kinetics essessment in the rumen and in the intestinum from untreated and by heat treated soya bean and pea.
Material and Methods For CP, phosphorus (P) disappearance was used in sacco technique, and for intestinal digestibility mobile bag technique on three cows provided with a large rumen cannula and T-cannula in the duo-denum behind the pancreas. P disappearance was determined in untreated (SBUNT) and roasted soya bean (SBR), untreated (PUNT), flacked (PF) and extruded pea by in sacco method. Intestinal P releas-ing from undegraded residues after 16 h incubation were determined for untreated, flacked and ex-truded pea by mobile bag method.
Crude protein content was determined by STN 46 9072 and for mineral analysis (STN 46 9072) sam-ples of soybean and pea and in sacco residues were ashed at 550 ˚C and the ash was disolved in 10 ml of HCl (1:3). Content of phosphorus before and after incubations was determined colorimetrically using the vanadium-molybdate solution
The results were evaluated by One-way analysis of variance and significant differences were declared at P<0.01 and P<0.05.
Results and discussion The goal of balanced nutrition is to keep excretion of N and P as low as possible while maintaining optimum dairy cows performance. Field (1981) stated that mineral absorption as well as utilization depends on solubility in the rumen. Phosphorus disappearance is an inportant precondition of its utili-zation in animals. Our results documents that P release from SBUNT, SBR, PF and PE is very high and

Chrenková et al.: The effect of heat treatment on phosphorus releasing from soya bean and pea
8. BOKU-Symposium Tierernährung 2009 Seite 63
after 16 h incubation is about 80 % of P released. P release kinetics were similar from SBUNT, SBR but in extent were significant diferences. From SBR was released about 10 % units less P than from SBUNT (Fig 1). From literature is known that protein form stable complexes with phytate and chemical bonds must be broken before P can be released from the phytate molecules (Thomas et al. 1946) Treatments that lower protein degradability in the rumen such temperature or formaldehyde (Kanishi et al. 1999; Park et al. 1999; Bravo et al. 2000) might decrease the rate and extent of phytate degra-dation and releasing of P. Our results documented decreasing of P release in the rumen.
Soya
20
40
60
80
100
1 2 3 4 5 6Incubation time (h)
P re
leas
ing
(%)
P roasted P untreated
Fig. 1 Kinetic of P releasing after ruminal incubation of untreated and roasted soybean
Pea
0102030405060708090
100
0 3 6 9 16 24Incubation time (h)
P re
leas
ing
(%)
untreated flacked extruded
Fig. 2 Kinetic of P releasing after ruminal incubation of untreated, flacked and extruded pea
For pea are P release kinetics different (Fig. 2) one another. The P lost by washing were the lowest from PUNT and P releasing was the highest PE in every incubation time. More than 30 % of P was released from extruded pea like from flacked P at 6, 9, and 16 h of incubation.
Parameters of P release in the rumen are in Table 1. Edg of P is the highest for untreaterd soya bean and pea and the lowest is for flacked pea. The parameter „b“ is very high for untreated pea.
Roasting of soya bean had adverse effect on crude protein degradation in the rumen significantl (P<0.01) (Fig. 3). The difference is the highest for 16 h incubation time (67.8 % resp. 87.6 %). Heat treatment of pea did not influence so markedly CP disappearnce than soya bean. Extent of CP disap-pearnce by flacking was the lowest after 3, 6 and 9 h incubations (43.9 %, 54.2 % resp. 70.1 %). The differences among flacked pea and untreated and extruded pea were significant (P<0.05).

Chrenková et al.: The effect of heat treatment on phosphorus releasing from soya bean and pea
Seite 64 8. BOKU-Symposium Tierernährung 2009
Tab. 1 Parameters of phosphorus release from soya bean and pea
Feed Parameters a (%) b (%) Edg (%)
Soya bean Untreated 53.0a 47.0a 96.3a
Roasted 44.2a 55.8a 84.6a
Pea Untreated 8.0b 92.0b 99.0b
Flacked 49.1b,c 50.b,c 72.2b
Extruded 32.5b,c 67.5b,c 75.4b
a,b,c in the same column, means with the same superscripts differ significantly (P<0.01)
Nitrogen
20
30
40
50
60
70
80
90
100
0 3 6 9 16 24
Incubation time(h)
Dis
appe
arnc
e (%
)
SBR SBUNT PUNT PF PE
Fig. 3 Kinetic of N degradation after ruminal incubation of untreated and termically treated soya bean and pea
93
94
95
96
97
98
99
100
Pea untreated Pea f lacked Pea extruded
Inte
stin
al P
dis
appe
aran
ce (%
)
Fig. 4 Intestinal disappearance of phosphorus from ruminal undegraded resirues of untreated and treated pea

Chrenková et al.: The effect of heat treatment on phosphorus releasing from soya bean and pea
8. BOKU-Symposium Tierernährung 2009 Seite 65
Very important knowledge is very high P disappearance from ruminally undegraded residues in the intestinum. The highest is from flacked pea for which was the lowest ruminal releasing (Fig. 4).
The treatment of soya bean and pea suppressed protein and phoshporus degradation. Thus the avail-ability of P is decreased in the rumen.
Conclusion Our results confirmed that among the feeds are significant differences in phosphorus release in the rumen. Because a large quatity of P forms stable complexes with phytate in the feeds is it necessary to study the processes of P release from them. Our next research will be focused on this problem. Rumen P phosphorus measured by in sacco method is an interesting way to describe P quality of feeds.
References
Bravo, D., Meschy F., Bogaert C., Sauvant D., 2000: Ruminal phosphorus availability from several feedstiffs measured by the nylon bag technique. Reprod. Nutr. Dev., 40, pp.149-162.
Čerešňáková, Z., Fľak, P., Poláčiková, M., Chrenková, M., 2005: In sacco NDF degradability and minerals release from selected forages in the rumen. Czech. J.Anim.Sci., 50, pp. 320-328.
Čerešňáková, Z., Fľak, P., Poláčiková, M., Chrenková, M., 2007: In sacco macrominerals release from six selected forages, Czech. J.Anim.Sci., 52, pp.175-182.
Čerešňáková, Z., Fľak, P., Chrenková, M., Poláčiková, M., Gralak, M.A., 2008: Ruminal phosphorus release from selected forages determined by in sacco technique. Slovak Journal of Animal Science, 3, pp. 146-150.
Field, A.C., 1981: Some thoughts on dietary requirements of macroelements for ruminants. Proc. Nutr. Soc., 40, pp. 267-27.2
Flachowsky, G., Grün, M., Polzin S., Kronemann, H., 1994: In sacco dry matter degradability and Ca, Mg and P disappearance from italian ryegrass, alfalfa hay and wheat straw in sheep and goats. J Anim. Physiol. A. Anim. Nutr., 71, pp.57-64.
Kenefick, S., Cashman, K.D., 2000: Inhibitory effect of wheat fibre extract on calcium absorption in Caco-2 cells: evidence for role of associated phytate rather than fibre per se. Eur. J. Nutr., 39, pp,12-19.
Matsui, T., 2002: Relationship between mineral availabilities and dietary phytate in animals. Anim. Sci. J., 73, pp. 21-28.
Morse, D., Head, H., Wilcox, C.J., 1992: Disappearance of phosphorus in phytate from concntrates in vitro from rations fed to lactating dairy cows. J. Dairy Sci., 75, pp. 1979-1986.
Park, W.Y., Matsui, T., Konishi, C., Kim, S.W., Yano, F., Yano, H.,1999: Formaldehyde treatment suppresses ruminal degradation of phytate in soyabean meal and rapeseed meal. Br. J. Nutr., 81, pp. 467-471.
Corresponding author
Mária Chrenková Research Center of Animal Production Nitra Hlohovská 2 951 41 Lužianky, Slovak Republic E-mail: [email protected]

Doležal et al.: The Effect of Stage Maturity of Alfalfa (Medicago sativa L.) on the Content of Major Minerals
Seite 66 8. BOKU-Symposium Tierernährung 2009
The Effect of Stage Maturity of Alfalfa (Medicago sativa L.) on the Content of Major Minerals
P. Doležal, J. Skládanka, R. Poštulka, L. Zeman
Department of animal nutrition and forage production, Mendel university of Agricul-ture and Forestry in Brno, Zemědělská 1, 613 00 Brno
Introduction The alfalfa (Medicago sativa L.) is the oldest and the most significant perennial proteinous forage crops with a highest content of protein and nutrition value. Typical for alfalfa is a good protein com-position (BÍRO et al., 2006) and a lower lignification. It contents more vitamins and mineral treats (especially calcium, phosphor, kalium and magnesium) than grasses. The differences in the quality and nutrition value are determined through the variety of the forage (ZIMMER and HONIG 1987). The nutrition value is affected mainly through the sequence of cutting, the vegetation stage, temperature and through the amount of rainfall during the vegetation too. The higher temperature during the vegetation increases the content of NDF in the cell wall and through this fact decreases the organic matter digestibility (MÍKA et al., 1997). The optimal vegetation stage for the harvest of the alfalfa is the stage before the flowering (the formation of the flower-bud). In this time has the alfalfa the higher concentration of protein in dry matter (over 23 till 24 %) and the lowest content of fiber. This time is relatively short (5-7 days).
The aim of this work was an assessing the affect of the different vegetations stage of alfalfa on the changes in the content and composition of the mineral treats.
Material and Methods It was realized a model observing of the alfalfa chemical composition in the first cut in the different stages. The fodder, variety Pálava , from the first cut was taken in the regular intervals – 1) before the formation of the flower-bud, 2) in the beginning of the flower-bud formation, 3) in the maximal flower-bud formation, 4) in the end of the flower-but formation, 5) in the beginning of the flowering, 6) in the maximal flowering, 7) in the end of flowering, 8) after the flowering. The samples were taken away promptly after the taking in the laboratory of the Department of nutrition and forage pro-duction MZLU in Brno. The samples were analyzed on the content of the organic and mineral sub-stances through the law (public notice) nr.222/1996.
The content of calcium and magnesium was determined through the flame atomic absorb spectros-copy (wave length for calcium was 422,7; for magnesium 285,2). The determining of phosphorus and potassium was done by a spectroscope on the using λmax 420 nm. The measuring was made with „Spektr AA−30“. The particular rations of the taken samples were statistical evaluated through the method correlation and regression by the program Statistica 7.0 CZ.
Results and Discussion The values of the analyzed alfalfa are shown in tables I. and II. Is evidently that alfalfa has in the different vegetation stages other chemical composition. Between the observed stages were found the
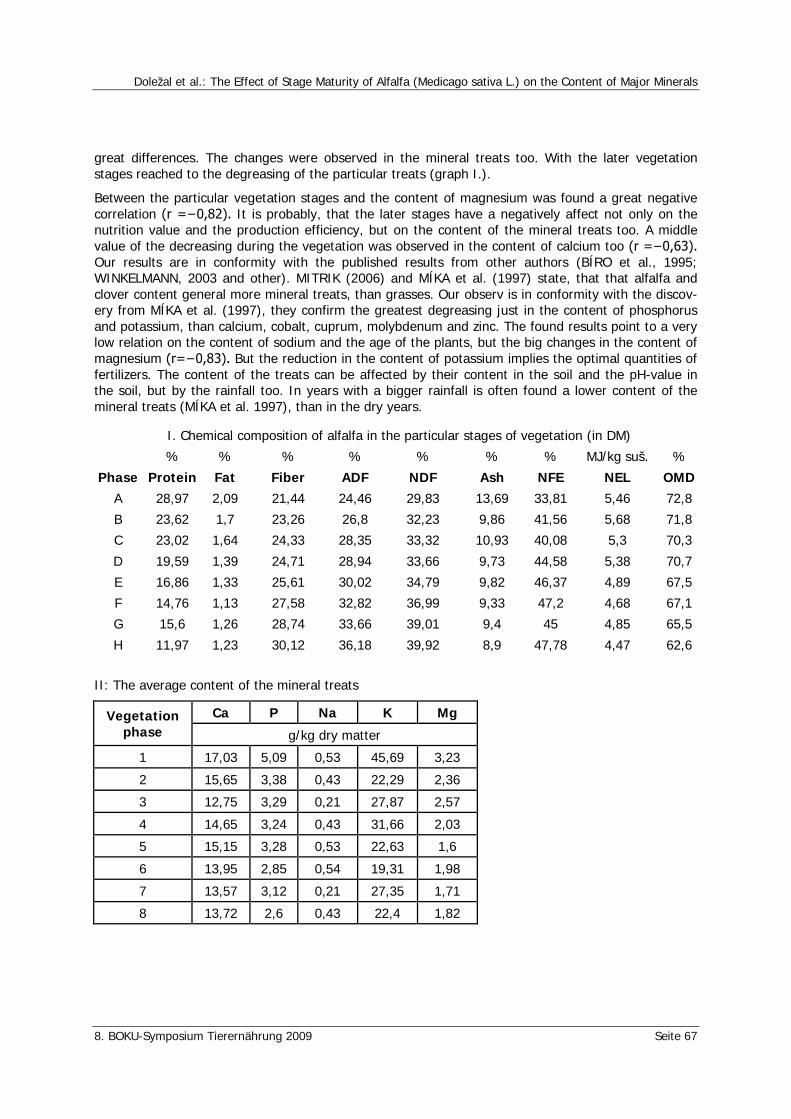
Doležal et al.: The Effect of Stage Maturity of Alfalfa (Medicago sativa L.) on the Content of Major Minerals
8. BOKU-Symposium Tierernährung 2009 Seite 67
great differences. The changes were observed in the mineral treats too. With the later vegetation stages reached to the degreasing of the particular treats (graph I.).
Between the particular vegetation stages and the content of magnesium was found a great negative correlation (r =−0,82). It is probably, that the later stages have a negatively affect not only on the nutrition value and the production efficiency, but on the content of the mineral treats too. A middle value of the decreasing during the vegetation was observed in the content of calcium too (r =−0,63). Our results are in conformity with the published results from other authors (BÍRO et al., 1995; WINKELMANN, 2003 and other). MITRIK (2006) and MÍKA et al. (1997) state, that that alfalfa and clover content general more mineral treats, than grasses. Our observ is in conformity with the discov-ery from MÍKA et al. (1997), they confirm the greatest degreasing just in the content of phosphorus and potassium, than calcium, cobalt, cuprum, molybdenum and zinc. The found results point to a very low relation on the content of sodium and the age of the plants, but the big changes in the content of magnesium (r=−0,83). But the reduction in the content of potassium implies the optimal quantities of fertilizers. The content of the treats can be affected by their content in the soil and the pH-value in the soil, but by the rainfall too. In years with a bigger rainfall is often found a lower content of the mineral treats (MÍKA et al. 1997), than in the dry years.
I. Chemical composition of alfalfa in the particular stages of vegetation (in DM)
% % % % % % % MJ/kg suš. %
Phase Protein Fat Fiber ADF NDF Ash NFE NEL OMD A 28,97 2,09 21,44 24,46 29,83 13,69 33,81 5,46 72,8
B 23,62 1,7 23,26 26,8 32,23 9,86 41,56 5,68 71,8
C 23,02 1,64 24,33 28,35 33,32 10,93 40,08 5,3 70,3
D 19,59 1,39 24,71 28,94 33,66 9,73 44,58 5,38 70,7
E 16,86 1,33 25,61 30,02 34,79 9,82 46,37 4,89 67,5
F 14,76 1,13 27,58 32,82 36,99 9,33 47,2 4,68 67,1
G 15,6 1,26 28,74 33,66 39,01 9,4 45 4,85 65,5
H 11,97 1,23 30,12 36,18 39,92 8,9 47,78 4,47 62,6
II: The average content of the mineral treats
Vegetation phase
Ca P Na K Mg
g/kg dry matter
1 17,03 5,09 0,53 45,69 3,23
2 15,65 3,38 0,43 22,29 2,36
3 12,75 3,29 0,21 27,87 2,57
4 14,65 3,24 0,43 31,66 2,03
5 15,15 3,28 0,53 22,63 1,6
6 13,95 2,85 0,54 19,31 1,98
7 13,57 3,12 0,21 27,35 1,71
8 13,72 2,6 0,43 22,4 1,82

Doležal et al.: The Effect of Stage Maturity of Alfalfa (Medicago sativa L.) on the Content of Major Minerals
Seite 68 8. BOKU-Symposium Tierernährung 2009
Graph I.
0
5
10
15
20
25
30
35
40
A B C D E F G HPhases
Cont
ent (
g/kg
dry
mat
ter)
Ca
P
NaK
Mg
Ca: y = 16,1375 - 0,3508*x; r = -0,6311; p = 0,0933 P: y = 4,4282 - 0,2382*x; r = -0,7803; p = 0,0223 Na: y = 0,4568 - 0,0093*x; r = -0,1674; p = 0,6919 K: y = 36,6379 - 2,0529*x; r = -0,5997; p = 0,1161 Mg: y = 2,9832 - 0,1824*x; r = -0,8288; p = 0,0110
CUME, et al (2000) indicates the relation between the concentration of mineral treats and the content of protein. He indicates the similarity of the relation (above-cited) with the relation between the high of the plants and the concentration of the minerals. In the first and second cut was detected the posi-tive correlation between the content of protein and the concentration of phosphorus and magnesium. The negative correlation was between the high of the plants and the content of phosphorus and mag-nesium.
Conclusion It comes with the maturing of the plants to significant worsening of the nutrition value (P<0,05), con-cretely the content of the minerals, especially magnesium (r =−0,82). In the particular vegetation phases was evidently the increasing of fiber (with the inclusion of the fraction ADF a NDF r =0,99; resp. r = 0,98). The relation of the energy content on the vegetation stages was very large (r = −0,92). The harvesting in the recommended stage is a premise for the optimal nutrition value, higher organic matter digestibility and higher efficiency.
Acknowledgment
This study was supported by the Research plan No. MSM6215648905 “Biological and technological aspects of sustainability of controlled ecosystems and their adaptability to climate change“, which is financed by the Ministry of Education, Youth and Sports of the Czech Republic.

Doležal et al.: The Effect of Stage Maturity of Alfalfa (Medicago sativa L.) on the Content of Major Minerals
8. BOKU-Symposium Tierernährung 2009 Seite 69
References
BÍRO, D., PETRIKOVIČ, P., ŠIMKO, J., 1995: Lucerna siata, Vysoká škola polnohospodárská Nitra, 26 s. ISBN 80−7137−226−9.
BÍRO, D., MICHÁLKOVÁ, J., JURÁČEK, J. et al., 2006: Zmeny aminokyselinového zloženia lucerny počas konzervačního procesu. Výživa zvířat 2006 – proteiny. MZLU Brno, s. 22–26. ISBN 80−7154−954−8.
CUME, S., et al. Relationships between crude protein and mineral concentrations in alfalfa and value of alfalfa silage as a min-eral source for periparturient cows. ANIMAL FEED SCIENCE AND TECHNOLOGY [online]. 2000, vol. 93, is. 3-4 [cit. 2009-08-05], s. 157-168. Dostupný z WWW: <Relationships between crude protein and mineral concentrations in alfalfa and value of alfalfa silage as a mineral source for periparturient cows>.
DOLEŽAL, P., DOLEŽAL, J., MIKYSKA, F. et al., 2006: Konzervace, skladování a úpravy objemných krmiv, skriptum MZLU v Brně, 247 s.
HAKL, J., KALISTA, J., ŠANTRŮČEK, J., 2005: Určení termínu sklizně vojtěšky s využitím sumy efektivních teplot. Úroda, roč. 53, č. 4, s. 6.
HOLÚBEK, R., GREGOROVÁ, H., ŠIMKO, J. et al., 1995: Ekologická, produkčná a kvalitatívna analýza kukurice na siláž a lucerny siatej. In 7th International Symposium Forage Conservation. Nitra: 18.−20. September, s. 27−30.
KALAČ, P., MÍKA, V., 1997: Přirozené škodlivé látky v rostlinných krmivech. ÚZPI Praha, 316 s., ISBN 80−85120−96−8.
KNOTEK, S., ŽILÁKOVÁ, J., 1996: Vplyv doby zberu a spôsobu ošetrenia lucerny na výživnú hodnotu siláží. In: Problémy bielkovinovej výživy zvierat. Nitra VÚŽV, s. 163−166.
KOVÁČ, A., ŠIMKO, J., BELOHOREC, R., 1980: Študium tvorby a redukcie faktorov úrodnosti lucerny siatej pestovanej na hmotu a semeno v rôzných agroekologických podmienkach. Záverečná sprava AF VŠP Nitra, 35 s.
MÍKA, V., HARAZIM, J., KALAČ, P. et al., 1997a: Kvalita píce. Ústav zemědělských a potravinářských informací, Praha, 227 s.
MÍKA, V., KALAČ, P., 1997b: Přirozené škodlivé látky v rostlinných krmivech. Ústav zemědělských a potravinářských informací, Praha, s. 98−28.
PETR, J., ČERNÝ, V., HRUŠKA, L. et al., 1980: Tvorba výnosu hlavích polních plodin. SZN Praha, s. 262−293.
SCEHOVIC, J., 1990: Vplyv rôznych rastlinnych druhov na chemické složenie a nutričnú hodnotu krmiva z prírodných trávných porastov. Agrochémia, 30, č. 9, s. 272−275.
ŠKULTÉTY, M., BENCOVÁ, E., JAŠKOVÁ, M., ŠKULTÉTYOVÁ, N. et al., 1995: Vplyv doby zberu a ošetrenia na kvalitu fermentácie lucernových siláží. In 7th International Symposium Forage Conservation. Nitra: 18. −20. September, s. 73−77.
VORLÍČEK, Z., 2004: Morfologické, biologické a hospodářské charakteristiky vojtěšky seté. In Trávy a jetelovinotrávy v zemědělské praxi. Agrární obzor Olomouc, s. 13−16. ISBN 80−903275−1−6.
Vyhl. 222/1996 Sb., kterou se stanoví metody odběru vzorků, metody laboratorního zkoušení krmiv, doplňkových látek a premixů a způsob uchovávání vzorků podléhajících zkáze.
WINKELMANN, J., 2003: Silážovat vojtěšku beze ztrát. Úspěch ve stáji, č. 1, s. 5.
ZELENKA, J., 2004: Výživná hodnota krmiv. In Výživa zvířat a nauka o krmivech. Skriptum MZLU v Brně, s. 184–213.
Corresponding author
Prof. MVDr. Ing. Petr Doležal, CSc., Doc. Ing. Jiří Skládanka, Ph.D Ústav výživy zvířat a pícninářství Mendelova zemědělská a lesnická univerzita v Brně Zemědělská 1, 613 00 Brno Česká republika E-mail: [email protected], [email protected]

Fry et al.: Effects of dietary zinc source and level on immune responses and health of cattle
Seite 70 8. BOKU-Symposium Tierernährung 2009
Effects of dietary zinc source and level on immune responses and health of cattle
Robert S. Fry1, Jerry W. Spears1, Patrick Schlegel2, and Stephane Durosoy3 1 North Carolina State University (USA) 2 Agroscope Liebefeld-Posieux Research Station ALP (Switzerland) 3 Pancosma (Switzerland)
Introduction Zinc (Zn) is known to play critical roles in the immune system (1). Stresses associated with weaning and transportation of calves reduces the immune response and increases the susceptibility of cattle to respiratory diseases. Some research (2, 3, 4) has indicated that calves supplemented with organic Zn sources have greater humoral immune response than those supplemented with inorganic Zn. A recent study with growing cattle suggested that Zn from a Zn glycinate complex (ZnGly) was more bioavail-able than Zn sulfate (5). The present study was conducted to determine the effect of dietary Zn level and source (ZnGly or ZnSO4) on health and immune response of calves.
Materials and Methods Receiving phase
One-hundred and five Angus and Angus-cross calves (steers and bulls) were purchased at an auction market and shipped to the North Carolina State University Beef Research facility. On arrival all calves were weighed, identified with an ear tag, bled via jugular puncture, vaccinated for respiratory (IBR, BVD, PI3, BSRV) and Clostridial diseases, and treated for internal and external parasites. Bull calves were also castrated by banding using a Calibander. Three calves were treated for respiratory disease on arrival at the feedlot and were not used in the experiment. The remaining calves were blocked by weight and initial sex (steer or bull) and randomly assigned to pens. Treatments were randomly as-signed to pens within a block. Treatments consisted of: 1) 15 mg Zn/kg DM from ZnSO4, 2) 30 mg Zn/kg DM from ZnSO4, 3) 15 mg Zn/kg DM from ZnGly (B-TRAXIM 2C Zn), and 4) 30 mg Zn/kg DM from ZnGly. Each treatment consisted of 5 replicate pens with 5 or 6 calves per pen, with the excep-tion of treatment 1 (15 mg Zn/kg from ZnSO4) which consisted of 4 replicate pens.
Steers were fed a corn silage-based diet for ad libitum intake (Table 1). The basal diet used in the receiving phase analyzed 22.7 mg Zn/kg DM and supplemental Zn was provided in a corn-Zn premix at 2% of the diet. Feed intake was measured daily during the 28-day receiving phase. Body weights and jugular blood was obtained from all calves on days 0, 7, 14, and 28 following arrival at the feed-lot. Plasma was analyzed for haptoglobin, an acute phase protein that is elevated in morbid animals. Haptoglobin was determined using an ELISA test kit obtained from Life Diagnostics.
Health of calves was visually evaluated twice daily during the receiving phase. Steers that appeared unhealthy were pulled from pens and rectal temperatures were taken. Antibiotic treatment was ad-ministered if rectal temperatures exceeded 39.9ºC. Treated steers were returned to their pens but temperatures were monitored and repeated treatments were given if needed. The first treatment con-sisted of injections of Nuflor, Banamine, and B-vitamin complex. Baytril was the second line antibiotic used if steers did not respond to Nuflor.

Fry et al.: Effects of dietary zinc source and level on immune responses and health of cattle
8. BOKU-Symposium Tierernährung 2009 Seite 71
Table 1. Ingredient composition of diets
Ingredient Receiving phase Growing phase ------------------------- % DM ------------------------- Corn silage 85.0 85.0 Soybean meal 10.9 9.4 Urea 0.51 0.75 Calcium carbonate 0.65 0.91 Monocalcium phosphate 0.55 --- Salt 0.40 0.40 Corn-zinc premix 2.00 2.00 Vitamin mix 0.03 0.03 Trace mineral mix 0.01 0.01 Monensin --- 0.02
Immune Measurements
At the end of the receiving phase steers continued on their same dietary treatment during an 80-day growing phase. Ingredient composition of the basal growing diet is presented in Table 1. The basal diet contained 25.1 mg Zn/kg DM.
The impact of dietary Zn treatments on immunity was assessed during the growing phase. The hu-moral immune response was assessed by measuring specific antibody production following an intra-muscular injection of a 25% pig red blood cell suspension. Two steers per replicate pen were injected with 10 mL of the pig red blood cell suspension on day 42 of the growing phase. Blood samples were obtained via jugular venipuncture from steers immediately prior to injection and on days 7, 14, and 21 following injection. Hemagglutination titers to pig red blood cells were determined using a mercap-toethnol-PBS microtitration technique as described by Droke et al. (6).
Cell mediated immune response was measured in vitro using a mitogen-induced lymphocyte blasto-genesis assay (7). On day 59 of the growing phase, jugular blood was obtained in heparinized tubes from 4 steers per treatment and lymphocytes were isolated. Isolated lymphocytes were incubated for 72 hours, and blastogenesis was determined by measuring incorporation of [3H] thymidine into lym-phocytes during the final 18 hours of incubation. Phytohemagglutinin (PHA) and Pokeweed mitogen (PWM) were used as mitogens in the blastogenesis assay and were added at a concentration of 10 µg/mL.
Statistical Analysis
Data were statistically analyzed as a randomized complete block design using the MIXED procedure of SAS (SAS Inst., Inc., Cary, NC). The model included Zn source, Zn level, Zn source x Zn level interac-tion and block. Pen means were used as the experimental unit for all variables except the lymphocyte blastogenesis data where individual animal served as the experimental unit. Plasma haptoglobin con-centrations and serum Ig titers were analyzed as repeated measures, with the model containing Zn level, Zn source, day and all possible interactions.
Results and Discussion Morbidity and antimicrobial treatment results are presented in Table 2. Incidence of respiratory dis-ease was high with approximately 75% of calves requiring at least one treatment. Percentage morbid-ity and number of antimicrobial treatments per steer were not significantly affected by treatment.
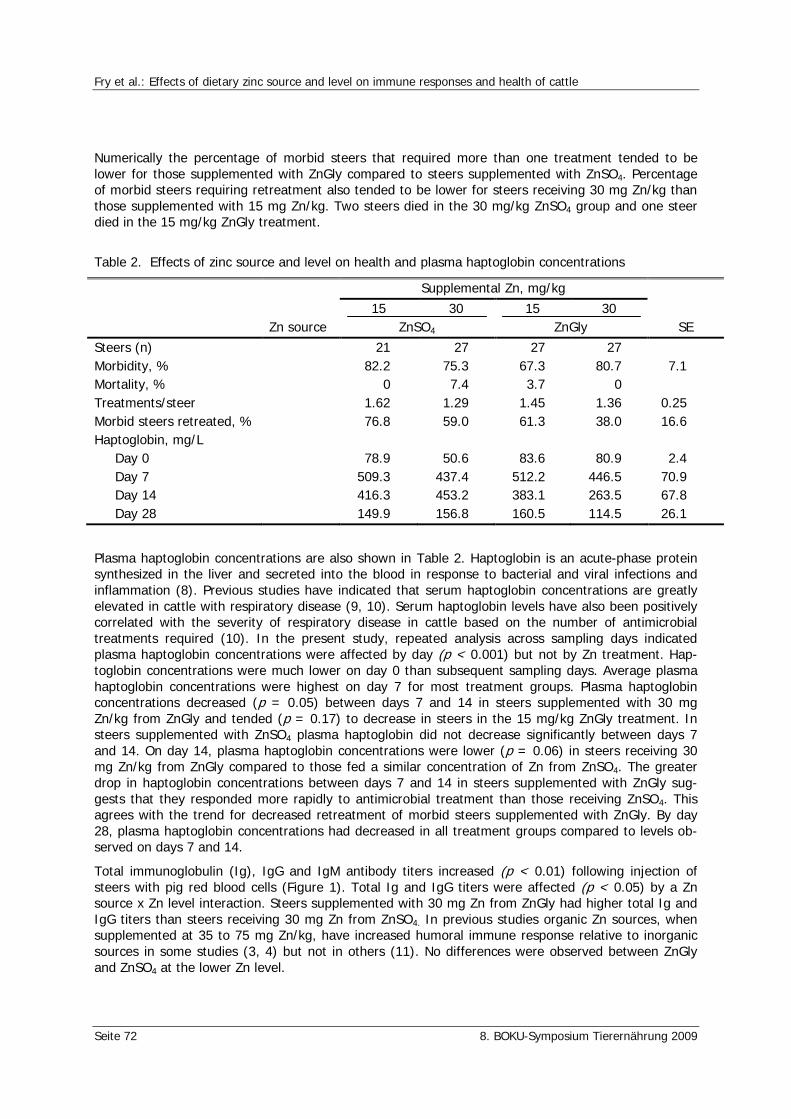
Fry et al.: Effects of dietary zinc source and level on immune responses and health of cattle
Seite 72 8. BOKU-Symposium Tierernährung 2009
Numerically the percentage of morbid steers that required more than one treatment tended to be lower for those supplemented with ZnGly compared to steers supplemented with ZnSO4. Percentage of morbid steers requiring retreatment also tended to be lower for steers receiving 30 mg Zn/kg than those supplemented with 15 mg Zn/kg. Two steers died in the 30 mg/kg ZnSO4 group and one steer died in the 15 mg/kg ZnGly treatment.
Table 2. Effects of zinc source and level on health and plasma haptoglobin concentrations
Supplemental Zn, mg/kg 15 30 15 30 Zn source ZnSO4 ZnGly SE Steers (n) 21 27 27 27 Morbidity, % 82.2 75.3 67.3 80.7 7.1 Mortality, % 0 7.4 3.7 0 Treatments/steer 1.62 1.29 1.45 1.36 0.25 Morbid steers retreated, % 76.8 59.0 61.3 38.0 16.6 Haptoglobin, mg/L Day 0 78.9 50.6 83.6 80.9 2.4 Day 7 509.3 437.4 512.2 446.5 70.9 Day 14 416.3 453.2 383.1 263.5 67.8 Day 28 149.9 156.8 160.5 114.5 26.1
Plasma haptoglobin concentrations are also shown in Table 2. Haptoglobin is an acute-phase protein synthesized in the liver and secreted into the blood in response to bacterial and viral infections and inflammation (8). Previous studies have indicated that serum haptoglobin concentrations are greatly elevated in cattle with respiratory disease (9, 10). Serum haptoglobin levels have also been positively correlated with the severity of respiratory disease in cattle based on the number of antimicrobial treatments required (10). In the present study, repeated analysis across sampling days indicated plasma haptoglobin concentrations were affected by day (p < 0.001) but not by Zn treatment. Hap-toglobin concentrations were much lower on day 0 than subsequent sampling days. Average plasma haptoglobin concentrations were highest on day 7 for most treatment groups. Plasma haptoglobin concentrations decreased (p = 0.05) between days 7 and 14 in steers supplemented with 30 mg Zn/kg from ZnGly and tended (p = 0.17) to decrease in steers in the 15 mg/kg ZnGly treatment. In steers supplemented with ZnSO4 plasma haptoglobin did not decrease significantly between days 7 and 14. On day 14, plasma haptoglobin concentrations were lower (p = 0.06) in steers receiving 30 mg Zn/kg from ZnGly compared to those fed a similar concentration of Zn from ZnSO4. The greater drop in haptoglobin concentrations between days 7 and 14 in steers supplemented with ZnGly sug-gests that they responded more rapidly to antimicrobial treatment than those receiving ZnSO4. This agrees with the trend for decreased retreatment of morbid steers supplemented with ZnGly. By day 28, plasma haptoglobin concentrations had decreased in all treatment groups compared to levels ob-served on days 7 and 14.
Total immunoglobulin (Ig), IgG and IgM antibody titers increased (p < 0.01) following injection of steers with pig red blood cells (Figure 1). Total Ig and IgG titers were affected (p < 0.05) by a Zn source x Zn level interaction. Steers supplemented with 30 mg Zn from ZnGly had higher total Ig and IgG titers than steers receiving 30 mg Zn from ZnSO4. In previous studies organic Zn sources, when supplemented at 35 to 75 mg Zn/kg, have increased humoral immune response relative to inorganic sources in some studies (3, 4) but not in others (11). No differences were observed between ZnGly and ZnSO4 at the lower Zn level.

Fry et al.: Effects of dietary zinc source and level on immune responses and health of cattle
8. BOKU-Symposium Tierernährung 2009 Seite 73
Figure 1. Effect of zinc source and level on antibody production following injection of pig red blood cells
0.01.02.03.04.05.06.0
0 7 14 21
15 ZnS04 30 ZnSO4 15 ZnGly 30 ZnGly
0.0
1.0
2.0
3.0
4.0
5.0
0 7 14 21
15 ZnS04 30 ZnSO4 15 ZnGly 30 ZnGly
Tota
l Ig
Tite
rs, l
og2
IgG
Tit
ers,
log 2
Day
Day
1A.
1B.
Table 3. Effects of zinc source and level on in vitro blastogenic response of peripheral blood lympho-cytes
Supplemental Zn, mg/kg 15 30 15 30 Zn source ZnSO4 ZnGly SE
--------------- CPM x 103 --------------- Phytohemagglutinina 110.2 66.4 80.1 92.1 9.8 Pokeweed mitogen 34.6 70.9 38.8 46.6 17.1 aZn source x Zn level interaction (p < 0.05).
The cell-mediated immune response was evaluated in vitro using a lymphocyte blastogenesis assay. Blastogenic response of lymphocytes to phytohemagglutinin was affected by a Zn source x Zn level interaction (p < 0.05). Lymphocytes isolated from steers fed 30 mg Zn/kg from ZnGly had a greater blastogenic response to phytohemagglutinin than those receiving ZnSO4 at 30 mg Zn/kg. However, at the lower level of Zn supplementation, blastogenesis was higher for lymphocytes obtained from ZnSO4 supplemented steers. Phytohemagglutinin stimulates predominately T-lymphocytes. Blastogenic re-

Fry et al.: Effects of dietary zinc source and level on immune responses and health of cattle
Seite 74 8. BOKU-Symposium Tierernährung 2009
sponse to pokeweed mitogen, a T-cell dependent B-lymphocyte mitogen, was not affected by dietary treatment.
Conclusion In the present study, supplementing ZnGly to weaned beef steers tended to reduce antimicrobial treatments per morbid steer. At the peak of morbidity, plasma haptoglobin concentrations decreased to a greater extent in ZnGly steers compared to steers receiving ZnSO4. Furthermore, the steers sup-plemented with 30 mg Zn/kg from ZnGly had greater production of total Ig and IgG following injection of pig red blood cell than steers supplemented with 30 mg Zn/kg from ZnSO4. Data from the present study suggests that supplementation of ZnGly at 30 mg/kg enhances the immune response in growing beef steers.
References
1 Shankar A. H. and Prasad A. S. 1998. Zinc and immune function: the biological basis of altered resistance to infection. Am. J. Clin. Nutr. 68:447S-463S.
2 Spears J. W., Harvey R. W. and Brown T. T. 1991. Effects of zinc methionine and zinc oxide on performance, blood characteristics, and antibody titer response to viral vaccination in stressed feeder calves. J. Am. Vet. Med. Assoc. 199:1731-1733.
3 Salyer G. B., Galyean M. L., Defoor P. J., Nunnery G. A., Parsons C. H. and Rivera J. D. 2004. Effects of copper and zinc source on performance and humoral immune response of newly received, lightweight beef heifers. J. Anim. Sci. 82:2467-2473.
4 Mandal G. P., Dass R. S., Isore D. P., Gang A. K. and Ram G. C. 2007. Effect of zinc supplementation from two sources on growth, nutrient utilization and immune response in male crossbred cattle. Amin. Feed Sci. Technol. 138:1-12.
5 Spears J. W., Schlegel P., Seal M. C. and Lloyd K. E. 2004. Bioavailability of zinc from zinc sulfate and different organic zinc sources and their effects on ruminal volatile fatty acid proportions. Livestock Prod. Sci. 90:211-217.
6 Droke E. A., Spears J. W., Brown T. T. and Qureshi M. A. 1993. Influence of dietary zinc and dexamethasone on immune responses and resistance to Pasturella hemolytica in growing lambs. Nutr. Res. 13:1213-1226.
7 Ward J. D., Spears J. W. and Kegley E. B. 1993. Effect of copper level and source (copper lysine vs copper sulfate) on copper status, performance, and immune response in growing steers fed diets with or without supplemental molybdenum and sulfur. J. Anim. Sci. 71:2748-2755.
8 Tizard I. R. 2000. Veterinary Immunology. 6th edition, W. B. Saunders Company, Philadelphia (USA).
9 Carter J. N., Meredith G. L., Montelongo M., Gill D. R., Krehbiel C. R., Payton M. E. and Confer A. W. 2002. Relationship of vitamin E supplementation and antimicrobial treatment with acute-phase protein responses in cattle affected by naturally acquired respiratory tract disease. Am. J. Vet. Res. 63:1111-1117.
10 Berry B. A., Confer A. W., Krehbiel C. R., Gill D. R., Smith R. A. and Monteongo M. 2004. Effects of dietary energy and starch concentrations for newly received feedlot calves: II. Acute-phase protein response. J. Amin. Sci. 82:845-850.
11 Nunnery G. A., Vasconcelos J. T., Parsons C. H., Salyer G. B., Defoor P. J., Valdez F. R. and Galyean M. L. 2007. Effects of source of supplemental zinc on performance and humoral immunity in beef heifers. J. Anim. Sci. 85:2304-2313.
Corresponding author
North Carolina State University Department of Animal Science Raleigh, NC 27695 USA E-mail : [email protected]

Firmensponsoring – Pancosma S.A., Voie-des-Traz 6, CH-1218 Le Grand-Saconnex
8. BOKU-Symposium Tierernährung 2009 Seite 75

Hammerl et al.: Experimentelle Untersuchungen einer hohen Jodversorgung auf die zootechnischen Leistungen sowie die Schlachtkörperqualität bei Mastschweinen
Seite 76 8. BOKU-Symposium Tierernährung 2009
Experimentelle Untersuchungen einer hohen Jodversorgung auf die zootechnischen Leistungen sowie die Schlachtkörperqualität bei Mastschweinen
Sabine Hammerl, Wilhelm Windisch, Qimeng Li, Karl Schedle
Universität für Bodenkultur Wien, Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie
Einleitung Das Spurenelement Jod [I] stellt einen essentiellen „Mikronährstoff“ für Menschen und landwirtschaft-liche Nutztiere dar. Jodmangel kann zu schwerwiegenden Erkrankungen wie Kropfbildung, Kretinismus und verminderter Fruchtbarkeit führen. Zur Vorbeugung dieser Krankheiten wird der Nahrung des Menschen und dem Futter der Tiere über verschiedene Quellen Jod zugesetzt. Ältere Studien bei Schweinen zeigen jedoch, dass zu hohe Joddosierungen negative Effekte auf die zootechnische Per-formance ausüben (Newton und Clawson 1974). Ähnliche Beobachtungen konnten auch bei Rindern festgestellt werden (Newton et al. 1974, Meyer et al. 2008). Beim Mastschwein liegt die maximale erlaubte Dosis des Jodgehaltes im Futter bei 10 mg/kg Futter. Diese Obergrenze wird in der prakti-schen Fütterung auch durchaus ausgenützt (Grünewald et al. 2006). In einer neueren Studie konnte Wagner (2008) zeigen, dass bereits Dosierungen unterhalb der zugelassenen Höchstgrenze zu Ver-schlechterungen der Mastleistungsparameter führen.
Um mögliche Auswirkungen unterschiedlicher Jodversorgungen innerhalb der gesetzlichen Grenzwerte erheben zu können, wurden im vorliegenden Versuch die Effekte verschieden hoher Jodversorgungen auf die Mast- und Schlachtleistungsparameter bei Mastschweinen untersucht.
Material und Methoden Der Versuch wurde an der österreichischen Schweineprüfanstalt in Streitdorf, Niederösterreich durch-geführt. Es wurden insgesamt 80 kastrierte Mastschweine (Dreirassenkreuzung, OEHYB) gleichmäßig gemäß ihrer Herkunft und ihres Anfangsgewichts auf 3 Versuchsgruppen, wie in Tabelle 1 dargestellt, aufgeteilt:
Tabelle 1: Jodgehalt der einzelnen Versuchsgruppen
Versuchsgruppe Anzahl der Tiere Joddosis (µg/kg) 1 16 150 (GfE Empfehlungen) 2 32 4000 3 32 10000
Jod wurde je zur Hälfte in Form von Kaliumjodid und Kaliumjodat supplementiert. Die Fütterung der Tiere erfolgte über eine automatische Fütterungsstation. Mit Hilfe von Transpondern konnte die tägli-che Futteraufnahme tierindividuell dokumentiert werden. Bis zu einem Lebendgewicht von 70 kg er-hielten die Tiere ein Vormastfutter, danach wurde auf ein Endmastfutter umgestellt. Die Futterratio-nen basierten auf Mais, Sojaextraktionsschrot, Gerste und Erbsen. Das geschrotete Futter, sowie Was-ser standen den Tieren ad libitum zur Verfügung.
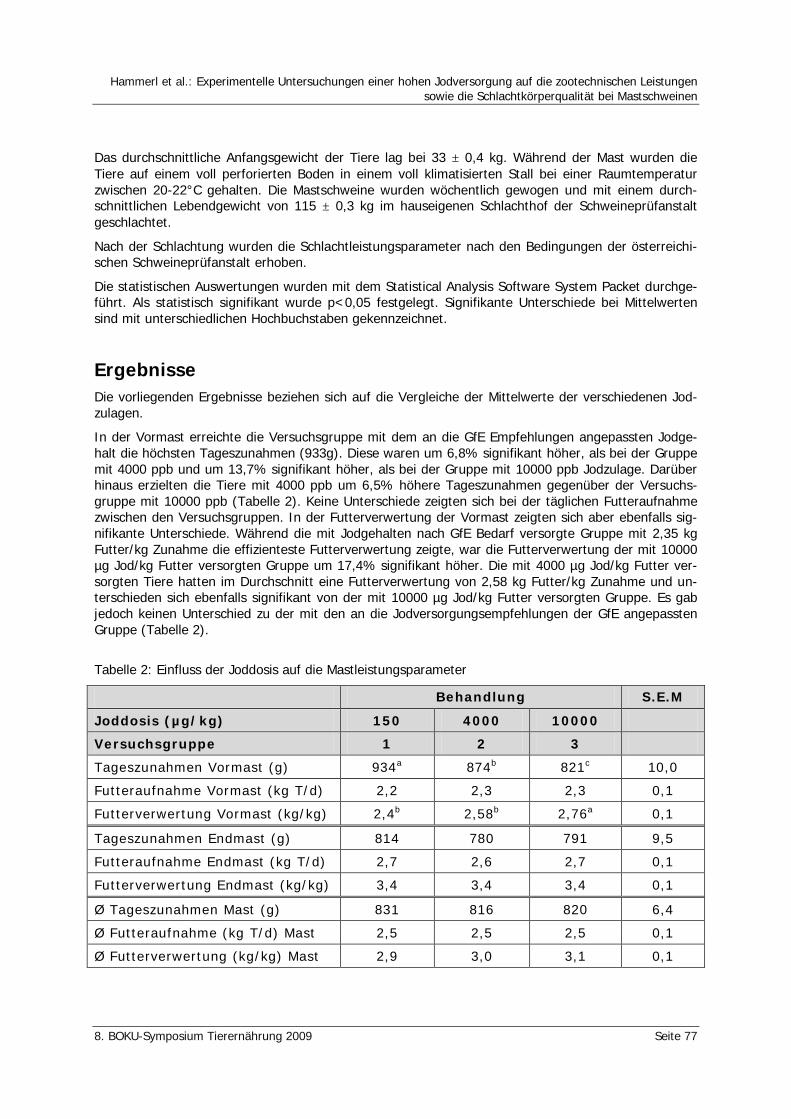
Hammerl et al.: Experimentelle Untersuchungen einer hohen Jodversorgung auf die zootechnischen Leistungen sowie die Schlachtkörperqualität bei Mastschweinen
8. BOKU-Symposium Tierernährung 2009 Seite 77
Das durchschnittliche Anfangsgewicht der Tiere lag bei 33 ± 0,4 kg. Während der Mast wurden die Tiere auf einem voll perforierten Boden in einem voll klimatisierten Stall bei einer Raumtemperatur zwischen 20-22°C gehalten. Die Mastschweine wurden wöchentlich gewogen und mit einem durch-schnittlichen Lebendgewicht von 115 ± 0,3 kg im hauseigenen Schlachthof der Schweineprüfanstalt geschlachtet.
Nach der Schlachtung wurden die Schlachtleistungsparameter nach den Bedingungen der österreichi-schen Schweineprüfanstalt erhoben.
Die statistischen Auswertungen wurden mit dem Statistical Analysis Software System Packet durchge-führt. Als statistisch signifikant wurde p<0,05 festgelegt. Signifikante Unterschiede bei Mittelwerten sind mit unterschiedlichen Hochbuchstaben gekennzeichnet.
Ergebnisse Die vorliegenden Ergebnisse beziehen sich auf die Vergleiche der Mittelwerte der verschiedenen Jod-zulagen.
In der Vormast erreichte die Versuchsgruppe mit dem an die GfE Empfehlungen angepassten Jodge-halt die höchsten Tageszunahmen (933g). Diese waren um 6,8% signifikant höher, als bei der Gruppe mit 4000 ppb und um 13,7% signifikant höher, als bei der Gruppe mit 10000 ppb Jodzulage. Darüber hinaus erzielten die Tiere mit 4000 ppb um 6,5% höhere Tageszunahmen gegenüber der Versuchs-gruppe mit 10000 ppb (Tabelle 2). Keine Unterschiede zeigten sich bei der täglichen Futteraufnahme zwischen den Versuchsgruppen. In der Futterverwertung der Vormast zeigten sich aber ebenfalls sig-nifikante Unterschiede. Während die mit Jodgehalten nach GfE Bedarf versorgte Gruppe mit 2,35 kg Futter/kg Zunahme die effizienteste Futterverwertung zeigte, war die Futterverwertung der mit 10000 µg Jod/kg Futter versorgten Gruppe um 17,4% signifikant höher. Die mit 4000 µg Jod/kg Futter ver-sorgten Tiere hatten im Durchschnitt eine Futterverwertung von 2,58 kg Futter/kg Zunahme und un-terschieden sich ebenfalls signifikant von der mit 10000 µg Jod/kg Futter versorgten Gruppe. Es gab jedoch keinen Unterschied zu der mit den an die Jodversorgungsempfehlungen der GfE angepassten Gruppe (Tabelle 2).
Tabelle 2: Einfluss der Joddosis auf die Mastleistungsparameter
Behandlung S.E.M
Joddosis (µg/kg) 150 4000 10000
Versuchsgruppe 1 2 3
Tageszunahmen Vormast (g) 934a 874b 821c 10,0
Futteraufnahme Vormast (kg T/d) 2,2 2,3 2,3 0,1
Futterverwertung Vormast (kg/kg) 2,4b 2,58b 2,76a 0,1
Tageszunahmen Endmast (g) 814 780 791 9,5
Futteraufnahme Endmast (kg T/d) 2,7 2,6 2,7 0,1
Futterverwertung Endmast (kg/kg) 3,4 3,4 3,4 0,1
Ø Tageszunahmen Mast (g) 831 816 820 6,4
Ø Futteraufnahme (kg T/d) Mast 2,5 2,5 2,5 0,1
Ø Futterverwertung (kg/kg) Mast 2,9 3,0 3,1 0,1

Hammerl et al.: Experimentelle Untersuchungen einer hohen Jodversorgung auf die zootechnischen Leistungen sowie die Schlachtkörperqualität bei Mastschweinen
Seite 78 8. BOKU-Symposium Tierernährung 2009
In der Endmast gab es keine Unterschiede zwischen den einzelnen Versorgungsniveaus bei den Para-metern Tageszunahme, tägliche Futteraufnahme und Futterverwertung (Tabelle 2).
Über die gesamte Mastperiode betrachtet verdünnten sich die in der Vormast beobachteten Effekte und so konnten nur mehr numerische Unterschiede festgestellt werden (Tabelle 2).
Die Schlachtleistungsparameter bei unterschiedlicher Jodzulage zeigten kaum Unterschiede. Der einzi-ge signifikante Unterschied lag in der Leitfähigkeit des Karrees eine Stunde nach der Schlachtung (Tabelle 3). Die Leitfähigkeit des Karrees eine Stunde nach dem Schlachten war bei den mit 4000 µg Jod/kg Futter versorgten Tiere mit 3,7 S/m-1 (Siemens/Meter) signifikant niedriger als bei den mit 10000 µg Jod/kg Futter versorgten Tiere mit einer Leitfähigkeit von 4,5 S/m-1. Die Leitfähigkeit des Karrees nach einer Stunde der nach GfE-Empfehlungen versorgten Gruppe unterschied sich weder von der mit 4000 µg Jod/kg Futter noch von der mit 10000 µg Jod/kg Futter versorgten Gruppe signifikant (Tabelle 3).
Tabelle 3: Einfluss der Joddosis auf die Schlachtleistungsparameter
Behandlung S.E.M
Joddosis (µg/kg) 150 4000 10000
Versuchsgruppe 1 2 3
Schlachtverluste (%) 20,3 20,3 20,8 0,2
Anteil wertvoller Teilstücke (%) 49,6 49,3 49,6 0,2
Magerfleischanteil (%) 58,8 58,5 58,7 0,2
Fettfleischverhältnis (1:…) 5,1 5,0 5,2 0,1
Rückenspeckdicke (mm) 24,8 24,9 23,4 0,3
pH Karree (nach 1 h) 5,9 5,9 5,8 0,1
pH Schinken (nach 1 h) 6,1 6,3 6,1 0,1
Leitfähigkeit Karree (nach 1 h) 3,9ab 3,7b 4,5a 0,1
Leitfähigkeit Schinken (nach 1 h) 4,0 3,8 4,1 0,1
Leitfähigkeit Karree (nach 24 h) 5,2 5,1 5,6 0,2
Leitfähigkeit Schinken (nach 24 h) 6,4 5,7 6,8 0,3
Fleischhelligkeit (U) 71,5 71,7 69,0 0,6
Dripverluste (%) 6,3 6,3 7,2 0,3
Intramuskulärer Fettgehalt (%) 1,2 1,2 1,1 0,1
Diskussion Jodsupplementierungen zum Schweinemastfutter über die Bedarfsempfehlungen der GfE hinaus sind gängige Praxis (Grünewald et al. 2006). Bereits 1974 beschrieben Newton und Clawson Verschlechte-rungen der Tageszunahme von Mastschweinen bei Jodsupplementierungen über dem gesetzlichen Grenzwert (800 und 1600 ppm). Wagner (2008) konnte bereits bei einem Versorgungsniveau von 4 ppm Jod negative Auswirkungen auf die Futterverwertung von Mastschweinen verzeichnen.

Hammerl et al.: Experimentelle Untersuchungen einer hohen Jodversorgung auf die zootechnischen Leistungen sowie die Schlachtkörperqualität bei Mastschweinen
8. BOKU-Symposium Tierernährung 2009 Seite 79
Auch unsere Studie zeigte, dass vor allem in der Vormast signifikante Unterschiede zwischen den drei Versorgungsniveaus auftraten. Die mit 150 ppb Futterjodgehalt versorgten Tiere wuchsen signifikant schneller, als die Tiere der anderen beiden Versorgungsniveaus. Beim Parameter Futteraufnahme zeigten sich keine Unterschiede zwischen den Versuchsgruppen in der Vormast, daraus resultierten aber signifikante Unterschiede in der Futterverwertung der unterschiedlichen Versorgungsniveaus für diese Leistungsperiode. In der Endmast gab es keine Unterschiede zwischen den verschiedenen Jod-versorgungsniveaus. Daraus ergaben sich über die gesamte Mastperiode nur mehr numerische Unter-schiede zwischen den Versuchsgruppen.
Die bei Schweinen beobachteten Effekte konnten auch bei anderen Tierarten beobachtet werden. Z.B. stellten Newton et al. (1974) bei ihrem Versuch mit Mastkälbern einen ähnlichen Trend fest. Auch hier verzeichneten die Versuchstiere während der ersten 49 Masttage schlechteres Wachstum, dass sie in weiterer Folge jedoch kompensieren konnten. Steigerungen der Jodsupplementation an die Grenze des gesetzlich erlaubten oder darüber hinaus verringerten die zootechnische Performance bei Rindern über die gesamte Mastperiode (Newton et al. 1974, Meyer et al. 2008).
Die auf niedrigen Jodversorgungsniveaus (z.B. bedarfsorientiert vs. 400ppb) herrschende Leistungs-homogenität kann durch den Zusatz goitrogener Substanzen zugunsten der höher versorgten Tiere verschoben werden (Cromwell et al. 1975). Aber auch im hohen Versorgungsbereich können goitroide Stoffe zu einer verminderten Jod Resorption aus dem Gastrointestinaltrakt bzw. zu einer erhöhten renalen Ausscheidung führen (Franke et al. 2009). Eventuell haben diese Substanzen Einfluss auf den für die Jodabsorption- bzw. die renale Jodausscheidung verantwortlichen Natrium-Jod Symporter. Dessen Expression wird unter anderem auch durch unterschiedliche Jodkonzentrationen im Futtermit-tel, oder durch Inhibitoren beeinflusst (Wagner 2008, Nicola 2009). Um diese Vermutung zu bekräfti-gen, sind jedoch weitere Studien notwendig. Unsere Studie zeigt, dass bei goitroidarmen Futterratio-nen die Bedarfsempfehlungen bereits ausreichen um optimale zootechnische Performance zu errei-chen.
Zusammenfassend kann gesagt werden, dass eine hohe Jodversorgung basierend auf einer Basisrati-on mit niedrigem Gehalt an goitroiden Substanzen, besonders in der Vormast zu einer Reduktion der Mastleistung geführt hat. Diesen Rückstand konnten die Tiere in der weiteren Mast jedoch teilweise kompensieren, so dass sich über die gesamte Mast gesehen nur mehr numerische Unterschiede in den Mastleistungsparametern zwischen den unterschiedlichen Versorgungsniveaus ergeben haben.
Schlussfolgerung Aufgrund unserer sowie anderer aktueller Studien sind die maximal erlaubten Jodkonzentrationen neu zu überdenken.
Literatur
Cromwell, G.L., Sihombing, D.T.H. & V.W. Hays (1975): Effects of Iodine Level on Performance and Thyroid Traits of Growing Pigs. Journal of Animal Science 41: 813-818.
Grünewald, K., Steuer, G. & Flachowsky, G. (2006): In: M. Rodehudskord (Hrsg.): 9. Tagung Schweine- und Geflügelernährung, 28.-30. November 2006, Institut für Agrar- und Ernährungswissenschaften, Universität Halle- Wittenberg. ISBN: 3-86010-833-6.
Franke, K., Meyer, U., Wagner, H., Hoppen, H.O. und Flachowsky G. (2009): Effect of various iodine supplementations, rapeseed meal application and two different iodine species on the iodine status and iodine excretion of dairy cows. Livestock Science doi:10.1016/j.livsci.2009.04.012.
Meyer, U., Weigel, K., Schöne, F., Leiterer M. & G. Flachowsky (2008): Effect of dietary iodine on growth and iodine status of growing fattening bulls. Livestock Science 115: 219-225.
Newton, G.L., Barrick, E.R., Harvey, R.W. & M.B. Wise (1974): Iodine Toxicity. Physiological Effects of Elevated Dietary Iodine on Calves. Journal of Animal Sciences 38. 449-455.

Hammerl et al.: Experimentelle Untersuchungen einer hohen Jodversorgung auf die zootechnischen Leistungen sowie die Schlachtkörperqualität bei Mastschweinen
Seite 80 8. BOKU-Symposium Tierernährung 2009
Newton, G.L. & A.J. Clawson (1974): Iodine Toxicity: Physiological Effects of Elevated Dietary Iodine on Pigs. Journal of Animal Science 39: 879-884.
Nicola, J.P., Basquin, C., Portulano, C., Reyna-Neyra, A., Paroder, M. und Carrasco, N. (2009): The Na+/I-Symporter (NIS) mediates active iodide uptake in the intestine. Am. J. Physiol. Cell Physiol. 296 (4), 654-662.
Wagner, V. (2008): Effects of High Dietary Iodine Supply as Iodide or Iodate on Tissue Iodine and Parameters of Iodine Metabolism in Growing Rats and Fattening Pigs. Dissertation an der Universität für Bodenkultur Wien.
Autorenanschrift
Bakk.techn. Sabine Hammerl Department für Lebensmittelwissenschaften und –technologie Abteilung: Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Universität für Bodenkultur Wien Gregor-Mendel-Straße 33, A-1180 Wien E-Mail: [email protected]

Firmensponsoring – BASF ChemTrade GmbH, Industriestraße 20, D-91593 Burgbernheiml
8. BOKU-Symposium Tierernährung 2009 Seite 81

Kozłowski et al.: Efficacy of different phytase preparations in broiler rations
Seite 82 8. BOKU-Symposium Tierernährung 2009
Efficacy of different phytase preparations in broiler rations
Krzysztof Kozłowski, Jan Jankowski and Heinz Jeroch
University of Warmia and Mazury in Olsztyn, Poland
Introduction The seeds of cereals, legumes and oil plants are poor in Ca and rich in total phosphorus (Jeroch et al. 2008). However, about 50 to 80% of total phosphorus is bound in organic form as phytate phospho-rus (Jeroch et al. 1999, Viveros et al. 2000). Phytic acid (IP6; myo-inositol 1,2,3,4,5,6-hexakis dihy-drogen phosphate) and its salts (phytates) are collectively referred to as “phytin” (Pallauf and Rim-bach 1997). In order to be utilized, the phytate P complex has to be first hydrolyzed by the enzyme phytase (myo-inositol-hexakisphosphate-phosphohydrolase) in the digestive tract. However, birds produce this enzyme in very low amounts (Maenz and Classen 1998). Phytase production by microor-ganisms in the digestive tract of birds is also very low (Marounek et al. 2008), in comparison with ruminants (Koddebusch and Pfeffer 1988). In products of plant origin (cereal grains, legume seeds, oil plants and their by-products), phytase activity was observed, but its concentrations in some plant feedstuffs are very low (Jeroch et al. 1999, Viveros et al. 2000), only the grains of rye, wheat and triticale were characterized by higher phytase activity. Plant phytase can also hydrolyze phytate phos-phorus (Barrier-Guillot et al. 1996, Oloffs et al. 1998), but different treatment methods (drying, pellet-ing) reduce phytase activity. Variation in phytase activity was also observed between genotypes of cereal species, e.g. in wheat (Jeroch et al. 1999). For several years phytases have been produced on an industrial scale with the help of microorganisms. According to a new EU recommendation (EU 1831/2003), these phytases have been included in the group of zootechnical feed additives. Their efficiency in the digestive tract of poultry has been demonstrated in many experiments (Nahm 2007, Selle and Ravindran 2007). The effectiveness of three different commercial microbial phytases in broi-ler rations based on wheat, corn and soybean meal was compared in the present study.
Materials and Methods The experiment was conducted with day-old Ross 308 male chicks on the University of Warmia and Mazury in Olsztyn. The birds were divided into the following groups:
Group I - positive control - standard P content Group II - negative control - lower P content (0.1% on aP basis) Group III - aP content as in group II + 500 FTU Aspergillus niger phytase per kg feed Group IV - aP content as in group II + 500 FTU Penicillium canescens phytase per kg feed Group V - aP content as in group II + 500 FTU Pichia (yeast strain) phytase per kg feed
The diets were provided as meal and the mean components were wheat, corn and soybean meal. As ingredients of the premixes, phytase preparations were added to starter (1-14 days) and grower (15-35 days) rations for groups III, IV and V, based upon the recommendations of the manufacturers and certificates of activity. Starter and grower rations (group I) contained 6.59 g or 6.09 g total P/kg and 3.98 g or 3.68 g av. P/kg, respectively. In the Starter and grower rations for groups II-V were 5.59 g or 5.12 g total P/kg and 2.98 g or 2.70 g av. P/kg, respectively. The experiment was conducted during 35 day-period in battery cages for growing chickens. 405 birds were randomly assigned to 5 treatment groups. Each treatment consisted of 9 replicates with 9 birds per replicate (i.e. 81 chickens per treat-ment). The stocking density in cages (0.5m2 each) was approximately 18 chickens per square meter,

Kozłowski et al.: Efficacy of different phytase preparations in broiler rations
8. BOKU-Symposium Tierernährung 2009 Seite 83
which corresponded to about 35 kg body weight/m2 at the end of the trial. Drinking water and feed was supplied ad libitum.
The body weight of chickens and feed intake were determined on a cage basis on day 1, 14 and 35. Based on these results, the feed conversion ratio and the growth rate were calculated on a cage basis. In each treatment, birds with average body weight were selected for carcass quality assessment (8 birds per treatment, 40 birds in total). Carcass weight, the proportions of breast muscles without skin, thigh and drumstick muscles, the proportion of abdominal fat and carcass yield were determined. Samples of the tibia were also taken from these chickens and specific gravity, DM, ash, Ca and P were analyzed.
The experimental results were processed statistically by ANOVA using Statistica for Windows software. Data in tables are given as mean values.
Results The birds of all experimental groups were characterized by good livability. Mortality rates were low, ranging from 0% (group IV) to 3.7% (groups II and III), and were not affected by the experimental treatments. Performance data for the entire experimental period are shown in Table 1. There were small differences in feed intake between the groups, indicating no clear effect of P reduction and phy-tase supplementation. The highest feed intake was recorded in broilers of group V (Pichia phytase). The feeding of rations with a reduced P content (group II) caused an approximately 8% decrease (P≤0.05) in the final body weights of birds, in comparison with group I fed the positive control diets. All three phytase preparations tested (groups III, IV and V) significantly improved the growth rate of birds. There were only small differences observed between the phytase-supplemented groups in this respect. Chickens of all phytase-supplemented groups almost reached the body weight of birds in the positive control group (I). The feed conversion ratio changed along the same lines as body weight. The highest value of this parameter was noted in group II (reduced P content). The difference be-tween group II and the other groups was significant. The three phytase preparations tested were equally effective, as confirmed by feed conversion ratios.
Carcass characteristics are presented in Table 2. Due to the lower final body weights of broilers in group II (lower P content), also their carcass weights were significantly lower in comparison with the other groups. The reduced P content and the addition of phytase had no effect on carcass propor-tions.
The results of tibia analysis are shown in Table 3. The rate of tibia mineralization was lower in birds of group II, compared with group I, but the differences were only partly significant. Diet supplementa-tion with two phytase preparations (groups III, V) enhanced the mineralization process. Penicillium canescens phytase (group IV) did not affect this process, but exerted a positive influence on perform-ance (Table 1).

Kozłowski et al.: Efficacy of different phytase preparations in broiler rations
Seite 84 8. BOKU-Symposium Tierernährung 2009
Discussion Phosphorus requirements in broilers are difficult to estimate due to both problems with accurate de-termination of the P content of feed and differences in recommendations. A high content of phytate phosphorus in total P and the varying levels of phytase activity in plant components of feed rations raise further doubts (Jeroch et al. 1999, Viveros et al. 2000). The addition of microbial phytases to diets enables higher amounts of otherwise hardly available phytate phosphorus to be utilized. The diets used in this experiment to test phytase efficacy contained considerably lower levels of total and available phosphorus than those recommended by the Polish Academy of Sciences (PAN) and the WPSA. Therefore, it should be kept in mind that the administration of such diets may cause a reduc-tion in performance, which was the case in the present study. However, the decrease in performance results was lower than expected as P levels were reduced by 25-30% (group II), compared with the PAN recommendations, relative to the positive control group (I) where P levels corresponded to the PAN recommendations.
The three microbial phytases tested hydrolyzed phytate P in the digestive tract of broilers, thus per-mitting performance results to be achieved consistent with those reported for broilers fed diets with the optimal P content. The effect of diet supplementation with phytase was also reflected in the con-centrations of crude ash, calcium and phosphorus in reference tibia samples. The results obtained are similar to the findings of other authors (Lee et al. 2000, Dilger et al. 2004).
The addition of phytase to broiler diets also brings other benefits, apart from reducing the level of P supplementation in the form of inorganic complexes. Higher phytate phosphorus utilization allows P excretion to decrease. This aspect is of environmental relevance on farms with high animal density levels. Studies have shown that the addition of phytase to broiler diets could reduce phosphorus ex-cretion by about 20 - 50% (Nahm 2007, Jeroch and Dänicke 2008).

Kozłowski et al.: Efficacy of different phytase preparations in broiler rations
8. BOKU-Symposium Tierernährung 2009 Seite 85
Conclusions The concentrations of total P or available P in broiler diets based on corn, wheat and soybean meal can be decreased by 1 g/kg provided that the diets are supplemented with microbial phytase (500 FTU/kg). The effect of phytases of bacterial and fungal origin, used in the present experiment, was found to be comparable.
References
Barrier-Guillot B, Casado P, Maupetit P, Jondreville C, Gatel F, Larbier M (1996) Wheat phosphorus availability: 2- In vivo study in broilers and pigs; relationship with endogenous phytasic activity and phytic phosphorus content in wheat. J Sci Food Agric 70: 69-74.
Dilger RN, Onyango EM, Sands JS, Adeola O (2004) Evaluation of microbial phytase in broiler diets. Poultry Sci 83: 962-970.
Jeroch H, Kluge H, Simon O, von Lengerken J (1999) Inhaltsstoffe und Futterwertdaten von Getreide und Körnererbsen. Martin-Luther-University Halle-Wittenberg Press, Halle.
Jeroch H, Dänicke S (2008) Faustzahlen zur Geflügelfütterung. Geflügeljahrbuch. Eugen Ulmer KG Publisher, Stuttgart, pp 148-182.
Jeroch H, Drochner W, Simon O (2008) Ernährung landwirtschaftlicher Nutztiere, 2nd ed., Eugen Ulmer KG Publisher, Stuttgart.
Koddebusch L, Pfeffer E (1988) Untersuchungen zur Verwertbarkeit von Phosphorus verschiedener Herkünfte an laktierenden Ziegen. J Anim Phys Anim Nutr 60: 269-275.
Lee SJ, Um JS, Paik IK, Lee JG (2000) Effect of crude phytase supplementation on performance of broilers fed different levels of phosphorus. Korean J Poult Sci 27: 169-179.
Maenz DD, Classen HL (1998) Phytase activity in the small intestinal brush border membrane of the chicken. Poult Sci 77: 557-563
Marounek M, Skrivan M, Dlouha B, Brenov N (2008) Availability of phytate phosphorus and endogenous phytase activity in the digestive tract of laying hens 20 and 47 weeks old. Anim Feed Sci Techn 146: 353–359.
Nahm KH (2007) Efficient phosphorus utilisation in poultry feeding to lessen the environment impact of excreta. Worlds Poult Sci J 63(4):625-654.
Oloffs K, Dolbusin A, Jeroch H (1998) Influence of microbial and native wheat phytase activity on phosphorus utilization in broilers. Arch Geflügelk 62: 260-263.
Pallauf J, Rimbach G (1997) Nutritional significance of phytic acid and phytase. Arch Anim Nutr 50: 301-319.
Viveros A, Centeno C, Brenes A, Canales R, Lozano A (2000) Phytase and acid phosphatase activity in plant feedstuffs. J Agric Food Chem 48: 4009-4013.
Corresponding author
Dr. Krzysztof Kozłowski Faculty of Animal Bioengineering Department of Poultry Science University of Warmia and Mazury in Olsztyn Oczapowskiego 5, 10-719 Olsztyn, Poland E-mail: [email protected]

Kulpys et al.: Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen
Seite 86 8. BOKU-Symposium Tierernährung 2009
Optimierung des Proteinstoffwechsels im Pansen mit Komplex-präparat von ätherischen Ölen und Spurenelementen
J. Kulpys1, R. Stankevičius1 und R. Urbonas2
1 Litauische Veterinärmedizinische Akademie 2 AG „Biofabrikas“
Einleitung Eiweißstoffen kommt in der Tiernährung im Vergleich mit den weiteren Hauptnährstoffen eine beson-dere Bedeutung zu. Neben Sauer-, Wasser- und Kohlenstoff sind im Eiweiß Stickstoff und Schwefel enthalten. Im Rahmen der Weender Futtermittelanalyse ist Rohprotein eine analytisch erfasste Frakti-on der Rohnährstoffe. Rohprotein besteht aus Reineiweiß und NPN-Verbindungen, deren Anteil von verschiedenen Faktoren (wie Futtermittelart, N-Düngung, Silierungsbedingungen u. a.) beeinflusst wird. Im jungen Gras kann NPN-Anteil beispielsweise bis 30 % reichen. NPN-Verbindungen (mit Aus-nahme der freien Aminosäuren) sind von monogastrischen Nutztieren nicht nutzbar. Proteinbausteine sind Aminosäuren (AS) – nichtessentiell und essentiell. Die essentiellen AS können durch keine Nähr-stoffe ersetzt werden. Für Monogastride wie Schweine sind sie eine Quelle für die Synthese der ent-behrlichen AS und der weiteren Stickstoffverbindungen im Tierkörper. Wiederkäuer können essentielle AS selbst nicht bilden, dennoch sind sie im Vergleich mit Monogastriden weniger auf Zufuhr einzelner AS mit dem Futter angewiesen, da im Pansen gebildetes Mikrobenprotein ein wichtiger Lieferant von essentiellen AS ist. Pansenmikroorganismen bauen Futterprotein und NPN-Verbindungen (wie z. B. Harnstoff) enzymatisch ab und liefern somit NH3 als Ausgangsstoff für die mikrobielle Proteinsynthese. In der Milchviehfütterung wird deshalb nach Möglichkeiten gesucht, Effizienz der Rohproteinver-wertung zu steigern (Roseler et. al., 1990). Darunter fällt Einsatz spezieller Präparate aus natürli-chen Stoffen.
Traditionell galt verdauliches Rohprotein als Maßstab für die Beurteilung der Proteinversorgung von Wiederkäuern. Es liefert jedoch keine konkreten Informationen über das mit dem Futter auf-genommene Protein.
In der EU werden Futtermittel gegenwärtig auf der Basis vom nutzbaren Rohprotein (nRP) beur-teilt. Das in den Dünndarm gelangende Protein unterscheidet sich in Menge und Zusammenset-zung von dem mit dem Futter zugeführten Protein, weil ein Teil der Futterproteine im Pansen abgebaut (aRP) und Mikrobenprotein (MP) gebildet wird. Dennoch sind Futterproteine nur zum Teil enzymatisch abbaubar. Ein Teil der Futterproteine gelangt unverändert in das Dünndarm und wird unabgebautes Rohprotein (uaRP) genannt. Der Anteil an uaRP hängt von den eingesetzten Futtermitteln und der Behandlungsart ab (Jeroch et al., 2004).
Futtermittel werden nach der ruminalen Proteinabbaubarkeit in drei Gruppen eingeteilt: 1) langsam abbaubar, im Mittel zu 65 % (55-75): Heu, Stroh, Soja- und Leinsamenku-
chen bzw. -schrot, Körner- und Silomais, Trockenschnitzel, Treber, Schlempe, Fischmehl; 2) mittelmäßig abbaubar, zu 75 % (65-85): Leguminosensilage, Anwelksilage, Kartoffel,
Futterrüben, Weizenkleie, Raps- und Sonnenblumenschrot bzw. -kuchen; 3) schnell abbaubar, im Mittel zu 85 % (75-95): Gras bzw. Grassilage, Zuckerrübenblatt-
silage, Gerste, Weizen, Hafer, Roggen, Erbsen, Ackerbohnen.
In der Praxis hat man meistens mit einem Überschuss an aRP zu tun: bei der Verfütterung von Grassi-lage und eigenem Getreide im Winter sowie bei der Verfütterung von Gras und dem gleichen Kraftfut-

Kulpys et al.: Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen
8. BOKU-Symposium Tierernährung 2009 Seite 87
ter im Sommer. Bei der Fütterung mit Mischfutter sollte man berücksichtigen, dass als wesentliche eiweißreiche Komponenten in den Futtermischungen in Litauen Soja-, Raps oder Sonnenblumenschrot bzw. -kuchen mit der ruminalen Proteinabbaubarkeit von 70-80 % verwendet werden. Einsatz lang-samer abbaubarer Eiweißfuttermittel wie Fischmehl erscheint dagegen nicht wirtschaftlich, spezielle Behandlungen (chemisch, thermisch u. a.) wirken sich negativ auf die Aufnahme von AS im Ver-dauungstrakt aus (Jukna et al., 1994; Juozaitienė et al., 2004).
Abb. 1. Proteinumsetzungen (N-Stoffwechsel) im Pansen
Es wurde festgestellt, dass bei Hochleistungskühen mikrobielle Poteinsynthese im Pansen mit 45-55% des Gesamtproteinbedarfs optimal ist. Restliches Protein soll als im Pansen unabbaubares Protein im Futter zur Verfügung stehen (Wallace, 1994; Williams et al., 1990).
Im modernen Proteinbewertungssystem für Wiederkäuer auf der nRP-Basis wird Proteingehalt von verschiedenen Futtermitteln und Rationen mittels Schätzgleichungen berechnet, wobei als Basisgrößen Umsetzbare Energie (UE) und verdauliche organische Substanz dienen. Damit wird gezeigt, dass mik-robielle Eiweißsynthese und Versorgung mit Stickstoff und Schwefel auch im Zusammenhang mit der Energieversorgung stehen. In den Schätzgleichungen wird auch uaRP-Gehalt mitberücksichtigt. Nutz-bares Rohprotein wird nach den folgenden Formeln berechnet (Jeroch et al., 2004):
1) nRP=11,93-[6,82×(uaRP:RP)]×UE+1,03×uaRP, beim Fettgehalt von < 7 % je kg TS; 2) nRP=13,06-[8,41×(uaRP:RP)]×(UE – UE RP)+1,03×uaRP, beim Fettgehalt von > 7 % je kg
TS; Anmerkungen: 1) nRP, RP und uaRP in g, UE in MJ, 2) nRP bei Rationsberechnung – pro Tag, bei Futterberechnung – je kg Futter-TS.
In den letzten Jahrzehnten ist die Milchleistung infolge genetischer Fortschritte und zielgerichteter Züchtung signifikant angestiegen, deshalb kann Pansenmikroflora nicht genug MP zur Realisierung des vorhandenen genetischen Leistungspotentials produzieren. Aus diesem Grund wird nach den Wegen gesucht, wie man Milchkühe möglichst effizient – insbesondere im Laktationshöhepunkt – mit Protein aus dem Futter versorgen und gute Milcherträge erreichen kann (Harris, 1996; Japertienė et al., 2006).

Kulpys et al.: Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen
Seite 88 8. BOKU-Symposium Tierernährung 2009
Zum Ausgleich des Verhältnisses von abbaubarem und unabbaubarem Protein werden biotechnologi-sche Verfahren zur Proteinstabilisierung im Pansen genutzt. Dazu zählt auch Einsatz spezieller Futter-zusätze. In der letzteren Zeit werden gute Ergebnisse erzielt, indem man spezielle Präparate zur Ver-langsamung des Proteinabbaus sowie zur Förderung der mikrobiellen Eiweißsynthese und der Ener-giegewinnung aus leicht verfügbaren Kohlehydraten sowie zur Normalisierung der NH3-Konzentration einsetzt (Roseler et al., 1993; Purwin et al., 2005).
Nach Angaben der japanischen Forscher (Hokkaido Research Center) (Kulpys et al., 2009) führte der Einsatz des Präparates „Rumitan“, das Rohproteinabbau verlangsamt, zu guten Ergebnissen (Tabel-le 1).
Tabelle 1. Vergrößerung des uaRP-Anteils mittels „Rumitan“ bei Holsteinkühen (n=50)
Parameter Kontrolle Versuchsgruppe Differenz Milchmenge, kg/d 35,1 36,0 + 2,6% Protein, % 3,2 3,4 +6,3 Fett, % 3,7 3,7 0 Harnstoff im Blut, mg/dl 0,41 0,28 -46%
Anmerkung: Zusammensetzung der Totalmischration (TMR): 45 kg Grassilage + 4 kg Zuckerrüben-schnitzel + 11,8 kg Kraftfutter (7,8 kg Trockentreber + 1,8 kg Weizen + 1,4 kg Sojaschrot + 0,9 kg Zitrustrester).
Material und Methoden Bei der Lösung des Problems von Proteinabbaubarkeit der Ration lenkte das von französischen For-schern entwickelte Komplexpräparat von ätherischen Ölen und Spurenelementen Novatan® die Auf-merksamkeit der litauische Tierernährungsspezialisten auf sich zu, mit dem man den Anteil an uaRP in der üblichen Ration für Milchkühe signifikant vergrößern kann. Sein Bioaktivitätsprinzip beruht auf dem Synergieffekt ätherischer Öle und Spurenelemente (Zn, Mn) auf enzymatische Aktivität der Pan-senbakterien sowie auf die Bildung elektrostatischer Bindungen zwischen Spurenelementen und Pro-tein. Die Bindungen werden später im sauren Labmagenmilieu leicht abgelöst (Toni et al., 2006).
Zusammensetzung des Präparates ist in der Abbildung 2 ausgewiesen.
Abb. 2. Zusammensetzung des verwendeten Komplexpräparates
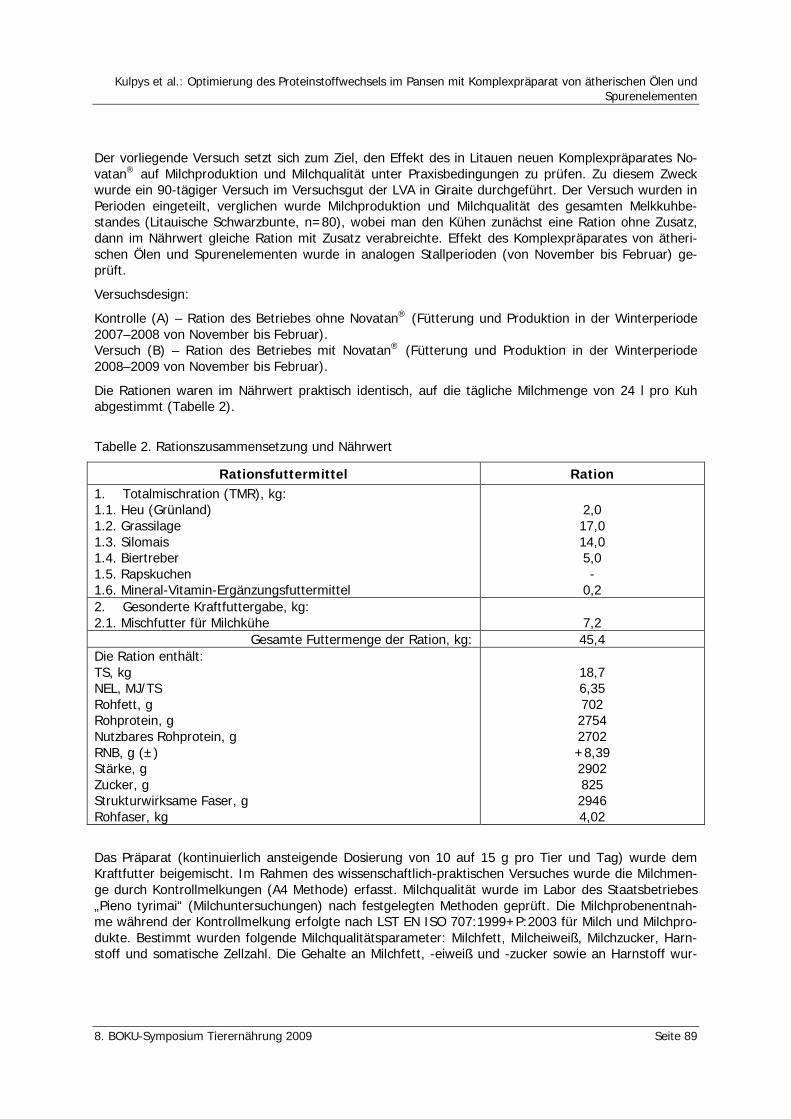
Kulpys et al.: Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen
8. BOKU-Symposium Tierernährung 2009 Seite 89
Der vorliegende Versuch setzt sich zum Ziel, den Effekt des in Litauen neuen Komplexpräparates No-vatan® auf Milchproduktion und Milchqualität unter Praxisbedingungen zu prüfen. Zu diesem Zweck wurde ein 90-tägiger Versuch im Versuchsgut der LVA in Giraite durchgeführt. Der Versuch wurden in Perioden eingeteilt, verglichen wurde Milchproduktion und Milchqualität des gesamten Melkkuhbe-standes (Litauische Schwarzbunte, n=80), wobei man den Kühen zunächst eine Ration ohne Zusatz, dann im Nährwert gleiche Ration mit Zusatz verabreichte. Effekt des Komplexpräparates von ätheri-schen Ölen und Spurenelementen wurde in analogen Stallperioden (von November bis Februar) ge-prüft.
Versuchsdesign:
Kontrolle (A) – Ration des Betriebes ohne Novatan® (Fütterung und Produktion in der Winterperiode 2007–2008 von November bis Februar). Versuch (B) – Ration des Betriebes mit Novatan® (Fütterung und Produktion in der Winterperiode 2008–2009 von November bis Februar).
Die Rationen waren im Nährwert praktisch identisch, auf die tägliche Milchmenge von 24 l pro Kuh abgestimmt (Tabelle 2).
Tabelle 2. Rationszusammensetzung und Nährwert
Rationsfuttermittel Ration 1. Totalmischration (TMR), kg: 1.1. Heu (Grünland) 1.2. Grassilage 1.3. Silomais 1.4. Biertreber 1.5. Rapskuchen 1.6. Mineral-Vitamin-Ergänzungsfuttermittel
2,0 17,0 14,0 5,0 -
0,2 2. Gesonderte Kraftfuttergabe, kg: 2.1. Mischfutter für Milchkühe
7,2
Gesamte Futtermenge der Ration, kg: 45,4 Die Ration enthält: TS, kg NEL, MJ/TS Rohfett, g Rohprotein, g Nutzbares Rohprotein, g RNB, g (±) Stärke, g Zucker, g Strukturwirksame Faser, g Rohfaser, kg
18,7 6,35 702 2754 2702 +8,39 2902 825 2946 4,02
Das Präparat (kontinuierlich ansteigende Dosierung von 10 auf 15 g pro Tier und Tag) wurde dem Kraftfutter beigemischt. Im Rahmen des wissenschaftlich-praktischen Versuches wurde die Milchmen-ge durch Kontrollmelkungen (A4 Methode) erfasst. Milchqualität wurde im Labor des Staatsbetriebes „Pieno tyrimai“ (Milchuntersuchungen) nach festgelegten Methoden geprüft. Die Milchprobenentnah-me während der Kontrollmelkung erfolgte nach LST EN ISO 707:1999+P:2003 für Milch und Milchpro-dukte. Bestimmt wurden folgende Milchqualitätsparameter: Milchfett, Milcheiweiß, Milchzucker, Harn-stoff und somatische Zellzahl. Die Gehalte an Milchfett, -eiweiß und -zucker sowie an Harnstoff wur-

Kulpys et al.: Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen
Seite 90 8. BOKU-Symposium Tierernährung 2009
den mit dem Gerät LactoScope FTIR (FT1.0. 2001; Delta Instruments) bestimmt; somatische Zellzahl mit SomaScope (CA-3A4.2004; Delta Instruments).
Rationszusammensetzung und Nährwert wurden in der Abteilung für Tierernährung der LVA nach standardisierten Methoden (AOAC) ermittelt, für Berechnungen kam die Hybrimin-Software (Futter-2006) zum Einsatz.
Die Daten wurden statistisch mit der Statistik-Software R 1.7.1 und WinExcel ausgewertet. Berechnet wurden folgende Parameter: Mittelwert, Standardabweichung, Varianz, Standardfehler des Mittel-werts. Realibilität (p) wurde mit dem t-Test geprüft (Juozaitienė und Kerzienė, 2001). Als Signifikanz-niveau wurde 0,05 gewählt.
Ergebnisse und Diskussion Der Versuch zeigte einen positiven Einfluss des Präparates auf die Milchproduktion (Abb. 3).
Abb. 3. Milchmenge (kg pro Kuh und Tag)
Wie aus der Abbildung 3 ersichtlich ist, ist die tägliche Milchmenge bereits im ersten Monat des Präpa-rateinsatzes um 6,8 % angestiegen, im zweiten und dritten jeweils um 16,4 % und 12,6 %. Am Ende des Versuches ist die tägliche Milchleistung zwar etwas gesunken (4,4 %), dennoch war sie über die gesamte Periode im Mittel um 2 Liter bzw. 9,7 % höher im Vergleich mit der Kontrolle. Darüber hinaus hatte das Präparat positive Wirkung auf die wesentlichen biochemischen Milchparameter (Tabelle 3).
Tabelle 3. Einfluss von Novatan® auf biochemische Milchparameter
Parameter Kontrolle (A)
Versuch (B) Im Mittel über die gesamte Versuchs-periode
Versuchs-beginn
(November)
Nach einem Monat
(Dezember)
Nach zwei Monaten (Januar)
Nach drei Monaten (Februar)
Fett, % 4,34±0,18 4,51±0,11 4,55±0,09 4,41±0,11 4,67±0,96 4,54 Protein, % 3,49±0,07 3,59±0,05 3,44±0,05 3,51±0,03 3,73±0,41 3,57
Milchzucker, % 4,63±0,06 4,61±0,03 4,61±0,05 4,67±0,03 4,67±0,26 4,64
Der Tabelle 3 ist zu entnehmen, dass nach einem Versuchsmonat Abnahme des Eiweißgehaltes (um 0,15 % geringer als zu Versuchbeginn) sowie geringere Milchzuckerkonzentration dokumentiert wur-den. Dies ist mit der signifikant angestiegenen (um 2,1 l) Milchleistung in Verbindung zu bringen (Abb. 2), wobei offensichtlich auch Unausgewogenheit der Ration hinsichtlich Protein- und Energiege-
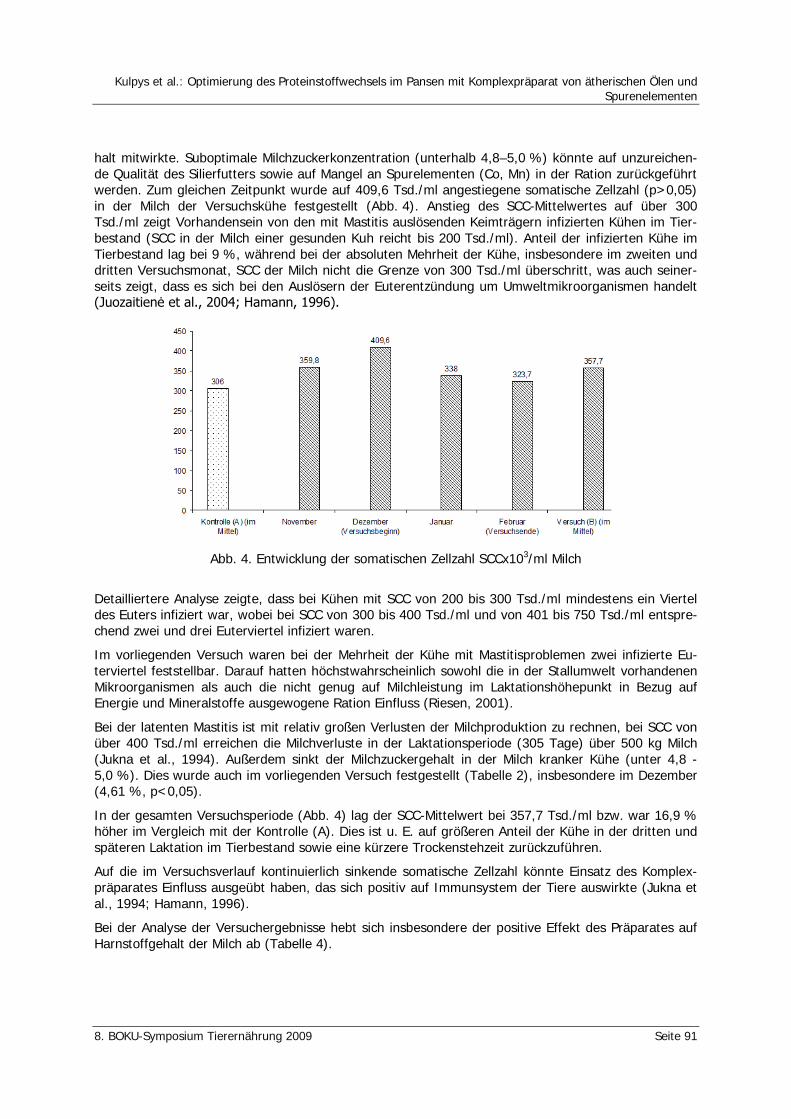
Kulpys et al.: Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen
8. BOKU-Symposium Tierernährung 2009 Seite 91
halt mitwirkte. Suboptimale Milchzuckerkonzentration (unterhalb 4,8–5,0 %) könnte auf unzureichen-de Qualität des Silierfutters sowie auf Mangel an Spurelementen (Co, Mn) in der Ration zurückgeführt werden. Zum gleichen Zeitpunkt wurde auf 409,6 Tsd./ml angestiegene somatische Zellzahl (p>0,05) in der Milch der Versuchskühe festgestellt (Abb. 4). Anstieg des SCC-Mittelwertes auf über 300 Tsd./ml zeigt Vorhandensein von den mit Mastitis auslösenden Keimträgern infizierten Kühen im Tier-bestand (SCC in der Milch einer gesunden Kuh reicht bis 200 Tsd./ml). Anteil der infizierten Kühe im Tierbestand lag bei 9 %, während bei der absoluten Mehrheit der Kühe, insbesondere im zweiten und dritten Versuchsmonat, SCC der Milch nicht die Grenze von 300 Tsd./ml überschritt, was auch seiner-seits zeigt, dass es sich bei den Auslösern der Euterentzündung um Umweltmikroorganismen handelt (Juozaitienė et al., 2004; Hamann, 1996).
Abb. 4. Entwicklung der somatischen Zellzahl SCCx103/ml Milch
Detailliertere Analyse zeigte, dass bei Kühen mit SCC von 200 bis 300 Tsd./ml mindestens ein Viertel des Euters infiziert war, wobei bei SCC von 300 bis 400 Tsd./ml und von 401 bis 750 Tsd./ml entspre-chend zwei und drei Euterviertel infiziert waren.
Im vorliegenden Versuch waren bei der Mehrheit der Kühe mit Mastitisproblemen zwei infizierte Eu-terviertel feststellbar. Darauf hatten höchstwahrscheinlich sowohl die in der Stallumwelt vorhandenen Mikroorganismen als auch die nicht genug auf Milchleistung im Laktationshöhepunkt in Bezug auf Energie und Mineralstoffe ausgewogene Ration Einfluss (Riesen, 2001).
Bei der latenten Mastitis ist mit relativ großen Verlusten der Milchproduktion zu rechnen, bei SCC von über 400 Tsd./ml erreichen die Milchverluste in der Laktationsperiode (305 Tage) über 500 kg Milch (Jukna et al., 1994). Außerdem sinkt der Milchzuckergehalt in der Milch kranker Kühe (unter 4,8 -5,0 %). Dies wurde auch im vorliegenden Versuch festgestellt (Tabelle 2), insbesondere im Dezember (4,61 %, p<0,05).
In der gesamten Versuchsperiode (Abb. 4) lag der SCC-Mittelwert bei 357,7 Tsd./ml bzw. war 16,9 % höher im Vergleich mit der Kontrolle (A). Dies ist u. E. auf größeren Anteil der Kühe in der dritten und späteren Laktation im Tierbestand sowie eine kürzere Trockenstehzeit zurückzuführen.
Auf die im Versuchsverlauf kontinuierlich sinkende somatische Zellzahl könnte Einsatz des Komplex-präparates Einfluss ausgeübt haben, das sich positiv auf Immunsystem der Tiere auswirkte (Jukna et al., 1994; Hamann, 1996).
Bei der Analyse der Versuchergebnisse hebt sich insbesondere der positive Effekt des Präparates auf Harnstoffgehalt der Milch ab (Tabelle 4).

Kulpys et al.: Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen
Seite 92 8. BOKU-Symposium Tierernährung 2009
Tabelle 4. Entwicklung des Harnstoffgehaltes der Milch
Parameter Kontrolle
(A) Versuch (B)
Oktober November Dezember Januar Februar Mittelwert 25,71 25,13 21,4 13,61 8,88 Standardfehler 1,14 0,67 1,50 0,52 0,64 Median 26 25 23 13 10 Modalwert 27 25 23 13 0 Standardabweichung 6,36 3,66 8,24 2,91 5,74 P<0,05 P<0,05
Der Tabelle 4 ist zu entnehmen, dass Harnstoffgehalt der Milch zu Versuchsbeginn (November–Dezember) optimal (20-30 mg%) war und in den weiteren Versuchsmonaten (Januar–Februar) auf unter 15 mg% absank. Wie aus anderen Versuchen (Harris, 1996) bekannt ist, sind auch beim opti-malen Harnstoffgehalt in der Milch ein niedriger Eiweißgehalt und Milchzuckerkonzentration unter 4,5 % eine Indiz dafür, dass Tiere mit Protein unterversorgt sind und Proteinabbau im Körper schneller als Eiweißsynthese erfolgt. Enthält die Milch weniger als 15 mg% Harnstoff und unter 3,6 % Eiweiß, so zeigt dies einen Mangel an eiweißreichen Komponenten und einen geringen Energieüberschuss in der Ration für Milchkühe (Gustafsson et al., 1993).
Schlussfolgerungen Versuchergebnisse lassen schlussfolgern, dass Einsatz von Novatan® Milchleistung (im Mittel 4,3 %) förderte und positiv den Milchfett- und Milcheiweißgehalt (ca. 7 %) beeinflusste sowie einen großen Effekt auf die Abnahme des Harnstoffgehalts (um ca. 65 %) hatte.
Die positiven Wirkungen sind in Verbindung mit verbesserten Fermentationsvorgängen im Pansen sowie mit einem ausgewogenen Verhältnis von aRP und uaRP im Verdauungstrakt zu bringen.
Im Versuchsverlauf konnte man feststellen, dass sich Präparatpulver (10–15 g pro Tier und Tag) dank der spezifischen Struktur von Mikroteilchen problemlos in die Kraftfuttermischung einmischen lässt und gute Lagerungseigenschaften besitzt.
Bei der Erstellung von Fütterungsprogrammen für Milchkühe ist die ruminale Proteinabbaubarkeit un-bedingt zu berücksichtigen. Bei begrenzten Futtermittelressourcen erscheint auch der Einsatz speziel-ler Zusätze sinnvoll, ein positives Beispiel dafür stellt die im vorliegenden Versuch benutzte Komplex-mischung von ätherischen Ölen und Spurenelementen mit relativ gutem zootechnischem Effekt in der Milchviehfütterung dar.
Literatur
AOAC. Official metods of Analysis. Vol. II. Association of official analitical chemists. 481. North Frederic Avenue, Suite 500. Gaithesburg, Maryland 20887-2417. USA, 1995.
Gyvūnų, skirtų eksperimiantiniams ir kitiems mokslo tikslams, laikymo, priežiūros ir naudojimo reikalavimai // VMVT įsakymas Nr B1-639. Žin., 2009-01-22 d. Nr. 8(287). P. 50-66.
Gustafsson A. H. and Palmquist D. L. Diurnal variation of rumen ammonia, serum urea and milk urea in dairy cows at high and low yields. J. Dairy Sci. 76, 1993. P. 475–484.
Hamann J. Nutrition in mastitis aetiology // Übersichten zur Tierernährung. 1996, 24(1). P. 141-148.
Harris B. Jr. Using milk urea nitrogen and blood urea values as management tools. In: Biotechnology in Feed Industry. Alltech Inc., Univ. Press, Nottingham, United Kingdom, 1996. P. 75–81.

Kulpys et al.: Optimierung des Proteinstoffwechsels im Pansen mit Komplexpräparat von ätherischen Ölen und Spurenelementen
8. BOKU-Symposium Tierernährung 2009 Seite 93
Japertienė R., Juozaitienė V. Analysis of the cows milk yield and mikability traits in diffferent lactation months // Proceedings of the 12 th Baltic animal breeding conference. Jurmala, Latvia, 2006. P. 47–50.
Jeroch H., Šeškevičienė J., Kulpys J. Žemės ūkio gyvulių ir paukščių mitybos fiziologinės reikmės. Kaunas: Naujasis lankas, 2004. P. 158.
Jukna Č., Andrus K., Alksninis A. Pieninė galvijininkystė JAV. Dairying in the U.S.A. Kaunas, LVA leidybos centras, 1994. P. 98–134.
Juozaitienė V., Juozaitis A. The Influence of somatic cell count in milk on reproductive traits and production of Black-and-White cows // Vetrerinarski arhiv, Univ. Zagreb Vet. Faculty, Zagreb, Croatia, 2005. N. 75(5). P. 407–414.
Juozaitienė V., Juozaitis A., Žakas A. Relationship of somatic cell count with milk yeild and composition in the herds of Black-and-White cattle. Medycyna Veterynaryjna PL ISSN 0025-8682003, 2004. Vol. 60 (7). P. 701–704.
Juozaitienė V., Kerzienė S. Biometrija ir kompiuterinė duomenų analizė. Kaunas, LVA leidybos centras, 2001. 115 p.
Juozaitis A., Juozaitienė V., Cukauskas V., Lavrinovičius J. Urėjos kiekis juodmargių karvių piene. Tarpt. konf. „Gyvulių mitybos indėlis įprastinės ir ekologinės gyvulininkystės vystymui Lietuvoje“. Pranešimų medžiaga. Kaunas, LVA, 2004. P. 63–65.
Kulpys J., Jerešiūnas A., Stankevičius R., Pikelis V. Zum Einfluss des Pansen-Modifikators S. cerevisiae1026 auf biochemische und hygienische Milchparameter. // Tagungsband 5. BOKU-Symposium Tierernährung „Qualitätsmindernde Futterinhaltsstoffe: Bedeutung – Vermeidung – Kontrolle”. Wien, Boku, 2006. S. 135–143.
Kulpys J., Urbonas R., Stankevičius R. Raciono baltymų pasisavinimo procesų stimuliavimo karvių organizme svarba ir būdai // Žemės ūkis / Verslas. 2009. Nr. 3(942). P. 369-37.
Miceikienė I., Pečiulaitienė N., Petraškienė R. Milk protein genotypes and their association with milk composition traits in the Lithuanian dairy cattle // Medycyna weterinaryjna. Lublin. Poland, 2005. N. 61(4). P. 394–397.
Pauliukas K., Šidiškis A., Urbonavičius A., Šerėnas K. Juodmargių karvių pieno sudėtis ir kokybės rodiklių kaita veikiant laktacijai ir kitiems faktoriams // Veterinarija ir zootechnika. Kaunas, 2005. T. 30 (52). P. 67–71.
Purwin C., Pysera B., Minakowski D., Sederevičius A., Traidaraitė A. Composition of milk and blood metabolites in high productivity dairy cows on pasture. // Veterinarija ir zootechnika. Kaunas, 2005. T. 32(54). P. 57-61.
Riesen G. Nutritional factors influencing milk quality // Use of organic compounds of trace elements to improve udder health. Milchpraxis. 2001. 39(4), S. 217-219.
Roseler D. K., Ferguson J. D. and Sniffen C. J. The effects of dietary protein dcgradability/undegradability on milk urea, milk NPN and blood urea in lactating dairy cows. J. Dairy Sci. 76, 1990, Suppl. 1. 168 p.
Roseler D. K., Ferguson J. D., Sniffen C. J. and Herrema J. Dietary protein degradability effects on plasma and milk urea nitrogen and milk nonprotein nitrogen in Holslein cows. J. Dairy Sci. 73, 1993. P. 525–534.
Šimkienė A., Juozaitienė V. Relation between somatic cell counts and lactose in milk cows // Moksl. konf. „Aktualios gyvulių ir paukščių mitybos problemos: alternatyvių pašarų priedų pašariniams antibiotikams panaudojimas gyvūnų mityboje“. Pranešimų medžiaga. Kaunas, LVA, 2006. P. 38–41.
Toni F., Grigoletto L., Rapp C.J., Socha M. T., Tolinson D. J. Auswirkung der Spurenelementform auf Milchleistungsparameter und Fruchtbarkeit von Milchkühen. Tagungsband 5. BOKU-Symposium Tierernährung „Qualitätsmindernde Futterinhaltsstoffe: Bedeutung – Vermeidung – Kontrolle”. Wien, Boku, 2006. S. 151–157.
Wallace R. J. Ruminal microbiology, biotechnology and ruminant nutrition: progress and problems. J. Anim. Sci. 72, 1994. P. 2992–3003.
Williams P. E. V. and Newbold C. J. Rumen probiosis: the effects of novel microorganisms on rumen fermentation and rumen productivity. In: Haresign W. and Cole W. J. A.(eds) Recent Advances in Animal Nutrition. Butterworth, London, 1990. 211 p.
Autorenanschrift
Dr. Jurgis Kulpys Abteilung für Tierernährung Litauische Veterinärmedizinische Akademie Tilzes 18, 47181 Kaunas, Litauen Tel +370 37 363408 E-Mail: [email protected]

Nitrayová et al.: Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs
Seite 94 8. BOKU-Symposium Tierernährung 2009
Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs
S. Nitrayová1, M. Brestenský1 , P. Patráš1, J. Brož2, J. Heger1 1 Animal Production Research Centre, Institute of Nutrition, Nitra, Slovakia 2 DSM Nutritional Products, Animal Nutrition and Health R&D, Basel, Switzerland
Introduction Plant ingredients commonly used in diets for pigs contain approximately two thirds of phosphorus in the form of salts of phytic acid that are unavailable for monogastric animals. There are many experi-ments demonstrating that the availability of plant phytin-bound P can be improved considerably in pigs by the addition of microbial phytase which is now an accepted feed additive (Selle and Ravindran, 2008). It effectively improves the availability of phytate phosphorus and decreases phosphorus excre-tion, thereby reducing environmental pollution (Vats et al., 2005). It has been suggested that the hydrolysis of phytate may also increase the digestibility of other nutrients.
Microbial phytases have been shown to have maximum activity at pH values ranging from 4.5 to 5.5 (Eeckhout and De Paepe, 1996). In contrast, pH values of common compound feeds as well as pig intestinal digesta range between 6.0 and 6.5 (Omogbenigun et al., 2003). Therefore, lowering the pH of gastrointestinal digesta might increase the efficacy of supplemental phytase. Indeed, there are several experiments demonstrating a synergistic effect of diet acidification and phytase supplementa-tion on P availability in pigs (Jongbloed et al., 1996; Kemme et al., 1997) even though it is unclear whether this effect is due to the lower pH or the presence of the acid, mostly citric acid, itself (Han et al., 1998; Boling et al., 2000). Moreover, organic acids are metabolized in the body and intestinal di-gesta have a relatively strong buffering capacity, which suggests that a significant effect of diet acidi-fication on the gastrointestinal tract pH can hardly be expected. An alternative strategy is the degra-dation of phytate outside the body under controlled conditions. In recent years, there has been an increasing interest in liquid feeding systems (Scholten et al., 2001) which allow to ferment pig diets, thus decreasing their pH to values close to a microbial phytase optimum (Jensen and Mikkelsen, 1998). In an in vitro study, Carlson and Poulsen (2003) found that within 48 hours of fermentation of phytase-supplemented, wheat- or barley-based diets at 20°C, pH decreased from 6.0 to about 4.5 and dietary phytate was almost completely degraded.
The purpose of the present study was to evaluate the effect of microbial phytase on apparent ileal and total tract digestibility of phosphorus (P), calcium (Ca), dry matter (DM) and nitrogen (N) in pigs fed dry or liquid fermented diets.
Material and methods Six Large White gilts of the Institute herd with an average initial body weight of 31.6 (SE 0.7) kg were used in the experiment. The pigs were surgically fitted with simple T- cannulas at terminal ileum and housed individually in metabolic cages. After a 14-d recovery period, during which a standard grower diet was offered, the pigs were randomly assigned to six dietary treatments according to a 6 x 6 Latin square design. Within the experiment, there were six consecutive periods, each consisting of a 5-d preliminary period followed by a 2-d collection period, during which faeces were collected by frequent

Nitrayová et al.: Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs
8. BOKU-Symposium Tierernährung 2009 Seite 95
grab sampling and stored as bulk for each individual. Ileal digesta was collected in 1-h intervals for 24 h starting at 6.00 h of the second day of the collection period. Digesta samples were acidified with 6M H2SO4 to pH 3.5, immediately frozen at -20 oC and stored for subsequent analysis.
Table 1. Ingredient and chemical composition of the basal diet (g/kg, air- dry basis)
Maize
590.0
Barley
170.5
Soyabean meal
201.0
L-lysine. HCl
1.94
L-threonine
0.16
Limestone
16.3
Monocalcium phosphate
0.5
Salt
3.6
Vitamin and micromineral premix1
3.0
Chromic oxide
3.0
Cellite
10.0
Dry matter
897.5
Crude protein
162.5
Ash
57.3
Ca
7.09
Total P
4.09
Digestible P (calculated)
1.10
Chromium
2.11
Acid insoluble ash
16.19
1 supplied per kg of diet: vitamin A 7 200 IU; vitamin D3 1350 IU; α-tocopherol 18 mg; vitamin B1 0.54 mg; vitamin B2 3.6 mg; vitamin B6 19.5 mg; Ca-pantothenate 10.5 mg; niacin 15 mg; vitamin K3 0.54 mg; biotin 0.06 mg; cyanocobalamin 0.021 mg; choline 102 mg; betaine 51 mg; Fe 60 mg; Zn 90 mg; Mn 42 mg; Cu 21 mg; I 0.42 mg; Co 0.54 mg; Se 0.21 mg
The basal diet (P0) with reduced content of total and calculated digestible P was formulated to contain maize, barley and soyabean meal as the main ingredients. The diet was supplemented with lysine and threonine to meet their respective requirements (NRC 1998). The calculated content of digestible P was approximately 50 % of the optimum requirement for a 40 kg gilt as given by NRC (1998). Chro-mic oxide and celite (as a source of acid insoluble ash) were included in the diet as digestibility mark-ers. The ingredient composition and chemical analysis of the basal diet is given in Table 1. Variant JP6500 of the microbial 6-phytase derived from Peniophora lycii (Ronozyme ® NP, DSM Nutritional Products Ltd., Switzerland) was added to the basal diet via premix at three levels equivalent to 1 000, 1 500 and 3 000 U/kg, thus forming three experimental diets (P1–P3). Other two diets (L0 and L1) were the same as diets P0 and P1, respectively, but were fed as liquid diets fermented in a laboratory setup simulating a commercial liquid feeding system. Three days before the start of the experiment, dry diets L0 and L1 were mixed with water at a feed:water ratio of 1:2.5 (w/w), 20 ml of lactic acid was added per kg of dry feed and the suspension was stirred for 1 h. The starting amounts of the diets corresponded to their requirements for 3 days. During the experiment, the diets were stirred for 5 min every 90 min and additionally for 20 min before each feeding. Every day after morning feeding, dry feed and water required for the next 24 h were added to the mixing vessel to maintain the

Nitrayová et al.: Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs
Seite 96 8. BOKU-Symposium Tierernährung 2009
feed:water ratio constant. Except for the first day, no lactic acid was added. The pH of the suspension was measured daily for the first 5 d and then every other day at 8.30 h.
The diets were fed twice daily at 6.00 and 16.00 h in two equal meals at a daily rate of 80 g/kg0.75. Diets P0-P3 were fed in a mash form (water:feed ratio 2:1, v/w). The spilled feed was dried and weighed to calculate actual feed intake. After each meal, water was offered ad libitum.
The diets, faeces and ileal digesta were analyzed for P, Ca, DM and N in accordance with standard methods of AOAC (1990). Chromic oxide was analyzed by atomic absorption spectrometry as de-scribed by Williams et al. (1962). The in-feed phytase activity was analyzed as described by Engelen et al. (1994). All analyses were performed in duplicate.
The data were subjected to ANOVA using Statgraphic Plus 3.1. package (version 3.1., Statistical Graphics Corp., Rockville, MD, USA). When a significant F-value for treatment means (P<0.05) was observed, the differences between means were assessed using Fisher's LSD procedure. Regression analysis was used to evaluate the relationship between supplemental phytase level (excluding liquid fermented diets) and P and Ca digestibility.
Results and discussion The intrinsic phytase activity of the basal diet was 109 U/kg, while the total phytase activities of diets P1, P2 and P3 were 954, 1419 and 2743 U/kg, respectively. Within the first five days of fermentation, pH value of the fermented diets decreased from 6.2 to about 4.5 and during the rest of the experi-ment it fluctuated between 4.1 and 4.7.
The mean values of apparent ileal digestibility of P, Ca, DM and N are presented in Table 2. Analysis of variance showed that, except for total P, phytase supplements or diet fermentation had no signifi-cant effect on ileal digestibility of nutrients. Ileal P digestibility of diet L1 was not significantly different from that of diet P3.
The data for apparent total tract digestibilities are summarized in Table 3. The digestibility of both P and Ca significantly increased as a result of phytase supplementation. Regression analysis (Table 4) showed significant linear and quadratic relationships between the phytase level and P digestibility as well as between the phytase level and Ca digestibility.
As expected, the supplementation of phytase to the maize- and barley-based diet significantly im-proved phosphorus digestibility. The maximum relative improvement for ileal and total tract P digesti-bility was approximately 30 and 60 %, respectively. A similar response was observed in other experi-ments using P-deficient, cereal-based diets (Omogbenigun et al., 2003; Sands and Kay, 2007). There was a positive linear and quadratic relationship between the phytase level and P digestibility. The quadratic terms in quadratic regression equations were not significant (Table 4), which suggested that there was no breakpoint or reduction in the response in the range of phytase activities examined. The effect of phytase supplementation on the digestibility of Ca was less apparent. Even though both ileal and total tract Ca digestibilities numerically increased with the increasing phytase level, a significant improvement was found only for total tract Ca digestibility in pigs receiving the highest phytase sup-plement. The results of experiments studying the effect of phytase on Ca digestibility in pigs are con-troversial. Some reports showed no effect (Harper et al., 1997; Sands et al., 2001) while others showed that phytase increased both ileal (Traylor et al., 2001) and total tract (Kemme et al., 1997) Ca digestibility. There are even studies demonstrating a greater effect of added phytase on Ca digestibil-ity than on P digestibility (Johnston et al., 2004). The Ca-releasing efficiency of phytase may have important consequences in the utilization of dietary P. It has been shown that the excess of Ca has an adverse effect on P digestibility, particularly at low P levels (Quian et al., 1996).

Nitrayová et al.: Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs
8. BOKU-Symposium Tierernährung 2009 Seite 97
Table 2. Effect of microbial phytase on apparent ileal digestibility of nutrients (%)
Diet
DM
N
P
Ca
P0 69.3 72.7 47.2 a 46.0 P1 70.4 73.8 49.5 a,b 47.6 P2 68.7 72.3 58.1 b,c 49.5 P3 70.1 74.2 61.3 c 54.3 L0 68.7 70.2 48.9 a,b 50.5 L1 71.1 74.3 64.3 c 50.8 Pooled SEM 1.0 1.4 3.5 4.6 a,b,c means within a column followed by different superscript are significantly different (P<0.05)
Table 3. Effect of microbial phytase on apparent total tract digestibility of nutrients (%)
Diet
DM
N
P
Ca
P0 86.2 87.3 41.1 a 47.9 a P1 86.4 86.6 54.4 b 51.2 a,b P2 86.9 87.5 55.3 b 53.9 a,b P3 87.2 87.9 65.7 c 59.7 c L0 86.6 86.3 43.0 a 48.2 a L1 87.3 87.5 62.1 b,c 54.7 a,b Pooled SEM 0.5 0.8 3.8 3.1
a,b,c means within a column followed by different superscript are significantly different (P<0.05)
Table 4. Parameters of regression equations relating apparent digestibility of P or Ca (Y, %) to dietary concentration of supplemental phytase (X, 10 3 U)1
Parameter
b0
b1
b2
P
R2
Ileal digestibility - linear2 P
0.472
0.050
0.002
37.2
Ca
0.446
0.041
0.014
24.0
Ileal digestibility - quadratic3 P
0.465
0.067
-0.0055
0.008
37.1
Ca
0.460
0.005
0.0114
0.042
26.1
Total tract digestibility - linear2 P
0.434
0.078
0.000
48.1
Ca
0.477
0.040
0.003
33.4
Total tract digestibility - quadratic3 P
0.416
0.125
-0.0150
0.001
49.8
Ca
0.478
0.037
0.0010
0.014
33.4
1 Excluding diets L0 and L1 2 Y = b0 + b1X 3 Y = b0 + b1X + b2X2

Nitrayová et al.: Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs
Seite 98 8. BOKU-Symposium Tierernährung 2009
In the liquid fermented diet, the phytate-degrading efficacy of phytase supplemented at the lowest level (diet L1) was not significantly different from that observed in the dry diet supplemented with the highest phytase level (diet P3). These results suggest that compound diets, designed for pig facilities using liquid feeding systems, may be formulated to contain lower phytase levels without reducing its effect on phytate degradation. There are other studies demonstrating a positive effect of soaking or fermenting single feeds or compound diets on the efficacy of exogenous phytase (Carlson and Poulsen, 2003; Blaabjerg et al., 2007). It seems that the main factors contributing to the increased degradation of phytate in liquid fermented diets are the acidification of fermentation media due to the proliferation of lactogenic bacteria population and the time of fermentation. It has been demonstrated that microbial phytase added to fermented high moisture maize remains active for a considerable time, thus maintaining its capacity to release phytate P (Niven et al., 2007). The mean pH value of the fermented diets was approximately 4.4, which was 0.9 units lower than the optimum for Peniophora lycii phytase (5.3). The lowered pH may also influence the solubility of phytate complexes. It is as-sumed that phytate is present in feedstuffs predominantly as an Mg phytate complex (Selle and Rav-indran, 2008), which is poorly soluble at pH above 5.0. At lower pH values, the solubility is considera-bly higher (Cheryan et al., 1983), thus making phytate more susceptible to enzymatic breakdown. In contrast to other reports (Carlson and Poulsen, 2003; Lyberg et al., 2006), fermentation itself had no significant effect on the phytate P availability. This might be due to different concentrations of native phytase in experimental diets. Unlike wheat and wheat byproducts, the intrinsic phytase activity in maize and soyabean meal, used as the main ingredients in the present study, is low (Viveros et al., 2000) and obviously insufficient to degrade phytate to a larger extent.
Table 5. Effect of microbial phytase on faecal P excretion
Diet
P intake (mg/kg BW)
Faecal P (mg/kg BW)
Faecal P (% of P intake)
P0
100.4
59.0
a
58.9
a
P1
104.4
47.1
b
45.6
b
P2
104.0
46.5
b
44.7
b
P3
100.1
34.6
c
34.3
c
L0
104.9
60.2
a
57.0
a
L1
104.9
39.7
b,c
37.9
b,c
Pooled SEM
3.9
4.1
3.6
a,b,c Means within a column followed by different superscript are significantly different (P<0.05)
Table 5 summarizes the data on P excretion expressed per kg body weight or as a percentage of P intake. In both cases, the reduction was dose-dependent, the highest effect being observed in pigs fed diet P3. As compared with diet P0, P excretion decreased by about 41 %. Faecal P excretion in fermented diet L1 was significantly lower than in diet L0 and was comparable with that of diet P3.
As a result of improved P digestibility due to phytase supplementation, faecal P excretion was reduced and similar results have been reported by other authors (Sands et al., 2001). However, the effect of phytase on total P excretion may be diverse, depending on the dietary concentration of available P. In our previous study using P-adequate diets (Patráš et al., 2006), we found that the phytase-induced reduction in faecal P excretion was partly counteracted by an increased urinary P excretion. It has been shown that urinary excretion is the main process responsible for P homeostasis in pigs, any ex-cess of absorbed P not used for anabolic purposes being excreted via urine (Rodehutscord et al., 1999). Therefore, to minimize total P excretion, pig diets should be formulated to meet available P

Nitrayová et al.: Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs
8. BOKU-Symposium Tierernährung 2009 Seite 99
requirement taking into account the P equivalency value of a given phytase product under given con-ditions.
Conclusion The total tract digestibility and ileal digestibility of P was significantly higher (P<0.05) in diets supple-mented with microbial phytase contrary diets without microbial phytase. The supplementation of mi-crobial phytase to the basal diets tended to improve the digestibility of Ca. The ileal and total tract digestibility of P was improved by phytase supplementation at 3000 U/kg to the dry diet, by 30 and 60% respectively. A similar effect was found in liquid fermented diet supplemented with phytase at 1000 U/kg.
As a result of improved P digestibility, faecal P excretion was reduced by 20 - 40 %. There was no significant effect of phytase addition or diet fermentation on the digestibility of DM and N.
References
AOAC (1990): Official Methods of Analysis, 15th edition. Association of Official Analytical Chemists, Arlington, Virginia.
Blaabjerg K., Carlson D., Hansen-Moeller J., Tauson HJ., Poulsen H.D. (2007): In vitro degradation of phytate and lower inositol phosphates in soaked diets and feedstuffs. Livestock Science, 109, 240–243.
Boling S.D., Webel D.M., Mavromichalis I., Parsons C. M., Baker D.H. (2000): The effects of citric acid on phytate-phosphorus utilization in young chicks and pigs. Journal of Animal Science, 78, 682–689.
Carlson D., Poulsen H.D. (2003): Phytate degradation in soaked and fermented liquid feed – effect of diet, time of soaking, heat treatment, phytase activity, pH and temperature. Animal Feed Science and Technology, 103, 141–154.
Cheryan M., Anderson F.W., Grynspan F. (1983): Magnesium- phytate complexes: effect of pH and molar ratio on solubility characteristics. Cereal Chemistry, 60, 235–237.
Eeckhout W., De Paepe M. (1996): In vitro and in vivo comparison of microbial and plant phytase. In: Phytase in Animal Nutrition and Waste Management, MB Coelho & ET Kornegay eds, BASF Corp. Mount Olive, NJ, 237-240.
Engelen A.J., van der Heeft F.C., Randsdorp P.H.G., Smit E.L.C. (1994): Simple and rapid determination of phytase activity. J. AOAC Int. 77, 760-764.
Han Y.M., Roneker K.R., Pond W.G., Lei X.L. (1998): Adding wheat middlings, microbial phytase, and citric acid to corn-soyabean diets for growing pigs may replace inorganic phosphorus supplementation. Journal of Animal Science, 76, 2649–2656.
Harper A.F., Kornegay E.T., Schell T.C. (1997): Phytase supplementation of low-phosphorus growing-finishing pig diets improves performance, phosphorus digestibility and bone mineralization and reduces phosphorus excretion. Journal of Animal Science, 75, 3174–3186.
Jensen B.B., Mikkelsen L.L. (1998): Feeding liquid diets for pigs. In: Garnsworthy P.C., Wiseman J. (eds.): Recent Advances in Animal Nutrition. Nottingham Univ. Press, Loughborough, UK, 107 pp.
Johnston S.L., Williams S.B., Southern L.L., Bidner T.D., Bunting L.D., Matthews J.O., Olcott B.M. (2004): Effect of phytase addition and dietary calcium and phosphorus levels on plasma metabolites and ileal and totaltract nutrient digestibility in pigs. Journal of Animal Science, 82, 705–714.
Jongbloed A.W., Kemme P.A., Mroz Z., Jongbloed R. (1996): The effects of organic acids in diets for growing pigs on the efficacy of microbial phytase. In: Coelho M.B., Kornegay E.T. (eds.): Phytase in Animal Nutrition and Waste Management, BASF Corp. Mount Olive, NJ, 515–524.
Kemme P. A., Jongbloed A.W., Mroz Z., Beynen A.C. (1997+1998): The efficacy of Aspergillus niger phytase in rendering phytate phosphorus available for absorption in pigs is influenced by pig physiological status. J. Anim. Sci. 75:2129–2138.
Lyberg K., Lundh T., Pedersen C., Lindberg J.E. (2006): Influence of soaking, fermentation and phytase supplementation on nutrient digestibility in pigs offered a grower diet based on wheat and barley. Animal Science, 82, 853–858.
Niven S.J., Zhu C., Columbus D., Pluske J.R., de Lange C.F.M. (2007): Impact of controlled fermentation and steeping of high moisture corn on its nutritional value for pigs. Livestock Science, 109, 166–169.
NRC (1998): Nutrient Requirements of Swine, 10th ed. National Academy Press, Washington, D.C.

Nitrayová et al.: Effect of diet fermentation and microbial phytase on apparent ileal and total tract digestibility of phosphorus, calcium, dry matter and nitrogen in growing pigs
Seite 100 8. BOKU-Symposium Tierernährung 2009
Omogbenigun F.O., Nyachoti C.M., Slominski B.A. (2003): The effect of supplementing microbial phytase and organic acids to a corn-soybean diet to early-weaned pigs. J. Anim. Sci. 81, 1806-1813.
Patráš P., Nitrayová S., Sommer A., Heger J. (2006): Effect of microbial phytase on apparent digestibility and retention of phosphorus and nitrogen in growing pigs. Czech J. Anim. Sci. 51, 437-443.
Qian H., Kornegay E.T., Conner D.E.Jr. (1996): Adverse effects of wide calcium:phosphorus ratios on supplemental phytase efficacy for weanling pigs fed two dietary phosphorus levels. J Anim Sci 74, 1288-1297.
Rodehutscord M., Faust M., Pfeffer E. (1999): The course of phosphorus excretion in growing pigs fed continuously increasing phosphorus concentrations after a phosphorus depletion. Arch. Anim. Nutr. 52: 323-334.
Sands J.S., Kay R.M. (2007): Phyzyme XP phytase improves growth performance and nutrient utilization in wheat-based diets fed to weaned pigs. Livestock Science, 109, 264–267.
Sands J.S., Ragland D., Baxter C., Joern B.C., Sauber T.E., Adeola O. (2001): Phosphorus bioavailability, growth performance, and nutrient balance in pigs fed high available phosphorus corn and phytase. J. Anim. Sci. 79, 2134-2142.
Scholten R.H.J., Rijnen M.M.J.A., Schrama J.W., Boer H., Vesseur P.C., Den Hartog L.A., van der Peet-Schwering C.M.C., Verstegen M.W.A. (2001): Fermentation of liquid coproducts and liquid compound diets: Part 1. Effects on chemical composition during a 6-day storage period. Journal of Animal Physiology and Animal Nutrition, 85, 111–123.
Selle, P.H., Ravindran, V. (2008): Phytate-degrading enzymes in pig nutrition. Livest. Sci. 113, 99-122.
Traylor, S.L., Cromwell, G.L., Lindemann, M.D., Knabe, D.A. (2001): Effects of level of supplemental phytase on ileal digestibility of amino acids, calcium, and phosphorus in dehulled soybean meal for growing pigs. J. Anim. Sci. 79:2634-2642.
Vats P., Bhattacharyya M.S., Benerjee U.C. (2005): Use of phytases (myo-Inositolhexakisphosphate phosphohydrolases) for combatting environmental pollution: A biological approach. Crit. Rev. Environ. Sci. Technol. 35, 469-486.
Viveros A., Centeno C., Brenes A., Canales R., Lozano A. (2000): Phytase and acid phosphatase activities in plant feedstuffs. Journal of Agricultural and Food Chemistry, 48, 4009–4013.
Williams C.H., David D.J., Lismoa O. (1962): The determination of chromic oxide in fecal samples by atomic absorption spectrophotometry. J. Agric. Sci. 59, 381-390.
Corresponding author
MVDr. Soňa Nitrayová, PhD. Animal Production Research Centre, Institute of Nutrition Hlohovecká 2, 95141 Lužianky, Slovakia E-mail: [email protected]

Firmensponsoring – DSM Nutritional Products Ltd, P.O. Box 2676, CH-4002 Basel
8. BOKU-Symposium Tierernährung 2009 Seite 101

Rapp et al.: Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen
Seite 102 8. BOKU-Symposium Tierernährung 2009
Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen
Christof Rapp1, Fried Flocke2, Josef Gövert3, Alfons Heseker4, Jochim Hopp5 1 Zinpro Animal Nutrition, Boxmeer, Niederlande 2 Raiffeisen Waren Zentrale Rhein Main e.G., Köln, Deutschland 3 Agravis Raiffeisen, Münster, Deutschland, 4 Essen (Oldenb.), Deutschland; 5 Hauptgenossenschaft Nord AG, Kiel, Deutschland
Einleitung Klauenverletzungen beeinträchtigen das Wohlbefinden der Sau, vermindern die Produktivität und kön-nen zu erheblichen wirtschaftlichen Einbußen führen. Erfahrungen aus der Praxis bei tragenden Sau-en lehren, dass beim Übergang von der Haltung in Kastenständen auf Gruppenhaltung die Häufigkeit von Klauenverletzungen zunimmt. Sämtliche tragenden Sauen innerhalb der Europäischen Union müs-sen ab dem 1. Januar 2013 in Gruppen gehalten werden (Richtlinie 2001/88/EG). Für die Bildung von widerstandsfähigem Klauenhorn sind Zink und Kupfer essentiell (Tomlinson et al., 2004). Mangan ist erforderlich für die Bildung von Knochen und Knorpel und kann somit indirekt, zum Beispiel über eine Fehlstellung der Gliedmaßen, die Klauengesundheit beeinflussen. Während die Würfe immer grösser werden, steigt die Futteraufnahme der Sau, die unter anderem für die Milchbildung erforderlich ist, nicht in gleichem Maße an. Die Spurenelementreserven der Sau nehmen mit steigender Wurfmasse ab (Mahan and Newton, 1995). Desweiteren sind durch den Gesetzgeber die Gesamtgehalte an Kupfer und Zink im Sauenfutter auf 25 bzw. 150 mg pro Kilogramm begrenzt (Verordnung [EG] Nr. 1334/2003). Eine Verminderung von Klauenverletzungen durch den Ersatz von anorganischen durch organisch gebundene Spurenelemente bei Kühen ist in der Literatur bereits beschrieben (Ballantine et al. 2002, Nocek et al. 2000, Nocek et al. 2006, Siciliano-Jones et al. 2008).
Ziel der vorliegenden Untersuchung war die Abschätzung der Wirkung einer Kombination von orga-nisch gebundenem Kupfer, Mangan und Zink auf die Häufigkeit sowie den Schweregrad von Klauen-verletzungen bei Sauen in Gruppenhaltung unter Praxisbedingungen.
Material und Methoden In sechs Sauenherden, mit einer Herdengröße zwischen 120 und 900 Sauen, wurden in den Abferkel-abteilen die Hinterfüße von Sauen nach einem einheitlichen Schema bonitiert (siehe Tabelle 1). Dabei wurden folgende Klauenverletzungen erfasst: Ballenwucherung und -risse, Risse im Übergang Bal-len/Sohle, Weiße Linie, waagrechte Wandrisse, senkrechte Wandrisse und lange Zehen. Der Schwere-grad der jeweiligen Verletzung wurde auf einer vierstufigen Skala wie folgt bonitiert: ohne Abwei-chung, leichte, mässige und schwere Abweichung. Bis zur ersten Bonitierung erfolgte die Spurenele-mentergänzung in sämtlichen Sauenfuttern in allen Herden allein durch anorganische Formen. Nach der ersten Bonitierung wurde in allen Herden sowohl im Futter für laktierende als auch tragende Sau-en ein Teil des anorganischen Kupfer (10 mg/kg), Mangan (20 mg/kg) und Zink (50 mg/kg) durch organische Formen (Availa®Sow, Zinpro, Eden Prairie, USA) ersetzt. Frühestens sechs Monate nach der ersten Bonitierung wurde eine weitere Beurteilung der Klauen nach dem gleichen Schema wie bereits beschrieben, durchgeführt. Es wurden insgesamt rund 700 Füße (350 Sauen) beurteilt.

Rapp et al.: Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen
8. BOKU-Symposium Tierernährung 2009 Seite 103
Ergebnisse und Diskussion Ballenwucherungen und Ballenrisse sind die in der vorliegenden Untersuchung am Häufigsten vor-kommenden Abweichungen. Bei einer Untersuchung aus den USA war der Ballen ebenfalls der Bereich in dem die meisten Abweichungen auftraten (Anil et al., 2007). Bevor ein Teils der anorganischen Spurenelemente durch organischen Quellen ersetzt wurde, kam keine der Herden über 20 % unver-sehrte Ballen Klauen hinaus (Abbildung 1). Nachdem die Fütterung der organischen Spurenelementen an Sauen über einen Zeitraum von mindestens sechs Monaten lang erfolgt war, hat sich der Anteil an Füssen mit unbeschädigten Ballen im Mittel der Herden von 9.4 auf 43.5 % erhöht.
Eine Verminderung der Häufigkeit von Rissen im Übergang vom Ballen zur Sohle nach Fütterung von organischen Spurenelementen wurde in fünf der sechs Herden beobachtet (Abbildung 2). Sechs Mo-nate nach Beginn der Fütterung der organischen Spurenelemente hat sich der Anteil von Füßen ohne Risse im Übergang vom Ballen zur Sohle im Mittel der Herden von anfänglich 56.4 auf 88.7 % erhöht.
Die Häufigkeit von Abweichungen der Weißen Linie hat sich nach der Fütterung von organischen Spu-renelementen in fünf von sechs Herden ebenfalls deutlich vermindert (Abbildung 3). Interessanterwei-se war Herde 2 wiederum die Herde bei der sich praktisch keine Verbesserungen zeigten. Im Mittel der Herden hat sich nach Fütterung der organischen Spurenelemente der Anteil an Füßen mit Abwei-chungen der Weißen Linie von anfänglich 60.4 auf 81.7 % erhöht.
Waagrechte Wandrisse traten bereits zu Beginn der Studie bei weniger als 20 % der Füße auf (Abbil-dung 4). Das heißt mehr als 80 % der Füsse waren ohne jegliche Abweichung. In der Hälfte der Her-den wurden nach der Fütterung von organischen Spurenelementen weniger waagrechte Wandrisse beobachtet. Bei der anderen Hälfte der Herden hat sich der Anteil an Füßen mit waagrechten Rissen geringfügig erhöht. Allerdings hat sich in den letztgenannten Herden der Anteil an Füssen mit schwe-ren Abweichungen vermindert. Der Anteil von Füßen ohne waagrechte Risse hat sich nach der Ergän-zung mit organischen Spurenelementen im Mittel der Herden von anfänglich 88.6 auf 91.6 % erhöht.
Der Anteil an Füßen mit senkrechten Wandrissen hat sich bei allen Herden nach der Fütterung von organischen Spurenelementen erhöht (Abbildung 5). Während die Erhöhung in einigen Herden deut-lich ausfiel, trat in anderen Herden nur eine geringfügige Veränderung auf. Im Mittel der Herden hat sich nach Fütterung der organischen Spurenelemente der Anteil an Füßen ohne senkrechte Risse in der Wand von anfänglich 82.2 auf 92.9 % erhöht.
Lange Zehen traten bereits bei der ersten Bonitierung in den meisten Herden bei weniger als 5 % der Füße auf (Abbildung 6). Der Anteil an langen Zehen hat sich in den meisten Herden praktisch nicht verändert. Der Anteil von Füßen ohne Abweichung der Zehen hat sich nach der Fütterung von organi-schen Spurenelementen im Mittel der Herden von anfänglich 93.7 auf 95.5 % erhöht.
Die oben dargestellte deutliche Verminderung von Klauenverletzungen an der Klauenunterseite bei der abschließenden Bonitierung deutet auf eine Verbesserung der Hornqualität durch die Zulage von or-ganisch gebundenem Zink, Mangan und Kupfer hin. Da die Rationen mit und ohne organisch gebun-denen Spurenelementen in zwei unterschiedlichen Zeiträumen gefüttert wurden, kann nicht ausge-schlossen werden, dass die Verbesserung der Klauengesundheit auch durch andere Einflussfaktoren bedingt war.

Rapp et al.: Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen
Seite 104 8. BOKU-Symposium Tierernährung 2009
Tabelle 1: System zur Bonitierung von Schweineklauen

Rapp et al.: Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen
8. BOKU-Symposium Tierernährung 2009 Seite 105
Abbildung 1: Ballenwucherungen und Ballenrisse in Füßen von Sauen mit anorganischer oder orga-nischer Spurenelementergänzung (Anteil Füße)
Abbildung 2: Risse im Übergang Ballen/Sohle in Füßen von Sauen mit anorganischer oder organi-scher Spurenelementergänzung (Anteil Füße)

Rapp et al.: Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen
Seite 106 8. BOKU-Symposium Tierernährung 2009
Abbildung 3: Risse in der Weißen Linie in Füßen von Sauen mit anorganischer oder organischer Spurenelementergänzung (Anteil Füße)
Abbildung 4: Waagrechte Wandrisse in Füßen von Sauen mit anorganischer oder organischer Spu-renelementergänzung (Anteil Füße)

Rapp et al.: Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen
8. BOKU-Symposium Tierernährung 2009 Seite 107
Abbildung 5: Senkrechte Wandrisse in Füßen von Sauen mit anorganischer oder organischer Spu-renelementergänzung (Anteil Füße)
Abbildung 6: Überlange Klauen in Füßen von Sauen mit anorganischer oder organischer Spuren-elementergänzung (Anteil Füße)

Rapp et al.: Wirkung einer Kombination von organischen Spurenelementen auf den Schweregrad von Klauenverletzungen bei Sauen
Seite 108 8. BOKU-Symposium Tierernährung 2009
Zusammenfassung In Zusammenfassung, sechs Monate nach Austausch von anorganischem Zink, Mangan und Kupfer durch organisch gebundene Herkünfte (Availa®Sow) war die Häufigkeit von Klauenverletzungen bei Sauen reduziert. Am meisten ausgeprägt war die Verminderung von Verletzungen an der Klauenunter-seite (Ballen, Übergang Ballen/Sohle und Weisse Linie).
Literatur
Anil, S.A., L. Anil, J. Deen, S. K. Baidoo, R. D. Walker. 2007. Factors associated with claw lesions in gestating sows. Journal of Swine Health and Production – March and April 2007. 15(2): 78-83.
Ballantine, H. T., M. T. Socha, D. J. Tomlinson, A. B. Johnson, A. S. Fielding, J. K. Shearer and S. R. Van Amstel. 2002. Effects of feeding complexed zinc, manganese, copper and cobalt to late gestation and lactating dairy cows on claw integrity, reproduction and lactation performance. The Professional Animal Scientist 18:211-218.
Mahan, D.C. und E.A. Newton. 1995. Effect of initial breeding weight on macromineral and micromineral composition of over a 3-parity period using a high-producing sow genotype. Journal of Animal Science, 73(1):151-158.
Nocek, J. E., A. B. Johnson and M. T. Socha. 2000. Digital characteristics in commercial dairy herds fed metal-specific amino acid complexes. J. Dairy Sci. 83:1553-1572.
Nocek, J. E., M. T. Socha and D. J. Tomlinson. 2006. The effect of trace mineral fortification level and source on performance of dairy cattle. J. Dairy Sci. 89:2679-2693.
Richtlinie 2001/88/EG des Rates vom 23. Oktober 2001 zur Änderung der Richtlinie 91/630/EWG über Mindestanforderungen für den Schutz von Schweinen. Amtsblatt Nr. L 316 vom 01/12/2001 S. 0001 – 0004.
Siciliano-Jones, J.L., M.T. Socha, D.J. Tomlinson, and J.M. DeFrain. 2008. Effect of Trace Mineral Source on Lactation Performance, Claw Integrity, and Fertility of Dairy Cattle . J. Dairy Sci. 91:1985-1995.
Tomlinson, D.J., C.H. Mülling und T.M. Fakler. 2004. Invited Review: Formation of keratins in the bovine claw: Roles of hormones, minerals and vitamins in functional claw integrity. J. Dairy Sci. 87:797-809.
Verordnung (EG) Nr. 1334/2003 der Kommission vom 25. Juli 2003 zur Änderung der Bedingungen für die Zulassung einer Reihe von zur Gruppe der Spurenelemente zählenden Futtermittelzusatzstoffen. Amtsblatt Nr. L 187 vom 26.7.2003 S. 11 – 15.
Autorenanschrift
Dr. Christof Rapp Zinpro Animal Nutrition Gerard Doustraat 4A 5831 CC Boxmeer, Niederlande E-Mail: [email protected]

Firmensponsoring – ZINPRO Performance MineralsTM, Gerard Doustraat 4a, NL-5831 CC Boxmeer
8. BOKU-Symposium Tierernährung 2009 Seite 109

Sager und Laguna Paredes: Selected nutrient- and trace element contents in hens´eggs – Nähr- und Spurenelemente in Hühnereiern
Seite 110 8. BOKU-Symposium Tierernährung 2009
Selected nutrient- and trace element contents in hens´eggs – Nähr- und Spurenelemente in Hühnereiern
Manfred Sager, Claudia Laguna Paredes
Competence Centre for elements, AGES – Austrian Agency for Health and Food Safety, 1220 Wien
Introduction Eggs are an important protein source in human nutrition. Eggs have to contain all nutrients, vitamins and trace elements which the chicken needs to develop healthy. About half of the egg consumption is not eaten as such, but hidden in processed food, like nooldes, cakes, sweets and ready made meals.
In order to protect the next generation from environmental influences, eggs from healthy hens might contain rather constant element levels of biologically more active elements, whereas whereas levels of less active elements are largely unknown, and might indicate the respective level in the poultry feed. Can the farmer influence the elementcomposition in his eggs by delivering designed food?
Material and methods 42 fresh and consumable egg samples were taken from the farmhouses together with the consumed feedstuffs. The mean weight of the sampled eggs was 57,4 ± 12,3 g, which closely appoaches the common standard of 60 g; the entire range was 39 - 93 g.
The eggs were separated into egg yolk and egg white, and 2-3 from the same farm were combined to yield mixed samples. The samples were freeze dried over 2 nights, ground in a mortar, and their wa-ter content was noted. The samples were digested with a K-chlorate solution in dilute nitric acid in microwave assisted pressure bombs, and made up to 25 ml. These digests could be used to determine most cations in the ICP-OES resp. ICP-MS, as well as the non- metals iodine, boron, and silicon. In the ICP-OES, the elemtns Al-Ba-Be-Ca-Cd-Co-Cr-Cu-Fe-Li-Mg-Mn-Na-Ni-P-Pb-Sb-Sn-Sr-Zn were read. The non-metals were read by another program yielding B-Ge-S-Si. Iodine was determined by ICP-MS by standard addition, as the signal is variable due to the oxidational status.
In a second run, all samples were digested with Mg-nitrate in the muffle furnace at 550°, and the ash dissolved in 1+1 hydrochloric acid, in order to look for selenium and arsenic by hydride AAS.
Finely ground feedstuffs were treated in the same way.
Resulting data Many elements in the egg yolk and egg white samples were below the detection limits of the ICP-OES, which were taken from the standard deviation of blank digests: Al < 2 mg/kg, Cd < 0,01 mg/kg, Cr < 0,15 mg/kg, Mo < 0,3 mg/kg, Ni < 0,8 mg/kg, Pb < 0,5 mg/kg, Sb < 1 mg/jkg, Sn < 1 mg/kg, and V < 0,1 mg/kg. Similarly, the non- metals boron (< 0,4 mg/kg) silicon (< 0,2 mg/kg) were not detect-able by the ICP-OES, either. We were satisfied to find none of the contaminants, and did not try the more sensitive ICP-MS. However, Ba-Be-Ca-Cu-Fe-Mg-Mn-Na-P-Sr-Zn were successfully determined,

Sager und Laguna Paredes: Selected nutrient- and trace element contents in hens´eggs – Nähr- und Spurenelemente in Hühnereiern
8. BOKU-Symposium Tierernährung 2009 Seite 111
besides the non metals P and s in a separate run at sufficient. Iodine was got from ICP-MS under oxidizing conditions and standard addition.
Table 1: Element contents for one egg
Egg yolk Egg white Egg in total Recommended daily intake
´mg ´mg ´mg DACH (mg) WHO (mg)
Sulfur 35,4 ± 7,4 74 ± 20 118 ± 31
Phosphorus 101 ± 22 5,3 ± 2,2 113 ± 27 700
Sodium 14,0 ± 3,8 87 ± 28 109 ± 35 550
Calcium 26,2 ± 5,5 2,39 ± 0,93 28,6 ± 5,9 1000 800
Magnesium 2,39 ± 0,59 4,8 ± 1,5 7,5 ± 2,1 300 - 400
Iron 1,04 ± 0,28 0,003 1,12 ± 0,33 10 // 15
Zinc 0,68 ± 0,15 0,005 0,73 ± 0,19 10 7 - 10 // 15
Copper 0,037 ±0,021 0,009 ±0,007 0,049 ± 0,024 1 - 1,5
Iodine 0,028 ± 0,016 0,003 0,031 ± 0,018 0,20 0,15
Strontium 0,020± 0,011 0,005 ±0,003 0,027 ± 0,015
Barium 0,023± 0,013 0,00042 0,023 ± 0,013
Manganese 0,016± 0,005 0,00026 0,017 ± 0,006 2 - 5
Selenium 0,0048 0,0023 0,0077 ± 0,0021
From the digests with Mg- nitrate, mainly selenium and arsenic were determined by hydride AAS in the batch mode. Arsenic was in the range of blanks throughout, smaller than 0,02 mg/kg. Selenium was higher than expected from literature data.
In respective feeds, most of the elements non detectable in the eggs, could be seen like Al, Cd, Cr, Co, Mo, Ni, Pb and V, whereas Sn and Sb were still too low. The feeds were not analyzed for boron and silicon.
From the concentrations in dry mass, the water contents, and the overall sample weights, the absoute amounts per egg were calculated, and given in table 1.
Discussion With respect to human nutrition, uptake of a few eggs delivers the recommended daily intake for io-dine and sodium. Contrary to older data, the total iodine contents in eggs was about 6-fold, preferably present in the egg yolk. Small eggs contained the same amount of Ba, Cu, and S than large eggs, whereas for all other elements, the absoute contents increased with egg weight.
Correlations between total element contents in the poultry feed and concentrations in dry egg mass were poor. The best correlation was found for strontium, whereas for calcium, there was none.
Similarly the composition of egg white and egg yolk was largely independent from each other. Just in case of strontium, copper, iodine and manganese, reliable relations were found.

Sager und Laguna Paredes: Selected nutrient- and trace element contents in hens´eggs – Nähr- und Spurenelemente in Hühnereiern
Seite 112 8. BOKU-Symposium Tierernährung 2009
References
M. Sager: Bestimmung von Jod in Pflanzen mit ICP- OES. Poster bei Colloquium Analytische Atomspektroskopie Konstanz, 23.03 - 27.03, 2003
M. Sager: Aktuelle Elementgehalte in Fleisch und Innereien aus Österreich. Ernährung/Nutrition 29(5), 199-206 (2005)
M. Sager: Selenium in agriculture, food, and nutrition. Pure Appl. Chem. 78(1), 43-65 (2006)
Druckaufschluss mit salpetersaurer Kaliumchloratlösung für die Spurenanalyse in pflanzlichem Material. M. Sager, Poster auf der ANAKON 07, 24.-27.3.2007 in Jena
M. Sager: Druckaufschluss mit salpetersaurer Kaliumchloratlösung für die Spurenanalyse in pflanzlichem Material. ALVA Jahrestagung 2007, Stadtschlaining
M. Sager: Jod und andere Spurenelemente in Hühnereiern. ALVA- Fachgruppe Futtermittel, Herbsttagung Wien, 2008
M. Sager: Spurenelemente in Lebensmitteln. Tagung Spurenelemente und Vitamine der GÖCh, Wien 2009
M. Sager, C. Laguna Paredes: Essential and Non- Essential Trace Elements in Sausages and Eggs. 3rd International IUPAC Symposium on Trace Elements in Food, Rom, 2. April 2009
M. Sager, C. Laguna Paredes: Einfluss des Legehennenfutters auf Jod- und andere Elementgehalte in Hühnereiern. 64. ALVA Jahrestagung in Salzburg, 2009
Corresponding author
Dr. Manfred Sager AGES Wien Analytik Kompetenzzentren, CC-Elemente Wien Spargelfeldstrasse 191 A-1220 Wien E-mail: [email protected]

Firmensponsoring – Daem eCo bvba, Nerumstraat 7, B-9340 Lede
8. BOKU-Symposium Tierernährung 2009 Seite 113

Sager und Werteker: Distribution of Elements in Flour Fractions – Elementverteilung in Mahlfraktionen von Weizen
Seite 114 8. BOKU-Symposium Tierernährung 2009
Distribution of Elements in Flour Fractions – Elementverteilung in Mahlfraktionen von Weizen
Manfred Sager, Manfred Werteker
Competence Centre of Elements, Institute for Crop Variety Testing, AGES – Austrian Agency for Health and Food Safety
Materials and Methods Samples of Austrian wheat varieties Antonius, Capo and Ludwig from the experimental sites in Sitzen-dorf and Staasdorf were milled on a Buhler MLU 202 roller mill. 6 Flour fractions and bran were pro-duced. Rests of endosperm particles were removed from the bran by a bran centrifuge. This centri-fuge produces a highly mineral enriched flour fraction.
The first three fractions (1 – 3) from the Buhler mill are milled by corrugated rollers. The bran result-ing from the first milling step is transported to the second pair of rollers and the bran of the second fraction to the third one, from where bran is collected as one of the milling products. The middlings of the first three milling steps are collected and milled by a system of three pairs of smooth rollers (4 – 6) in an analogous way. The flours from the first fraction of the corrugated rollers (1) and from the smooth rollers (4) represent the starchy inner parts of the endosperm while further fractions from corrugated (2, 3) and smooth rollers (5, 6) are from outer layers of the kernels as may be demon-strated by the ash levels of the different fraction, which are increasing from fraction 1 to 3 and from fraction 4 to 6.
The flour fractions were at first digested in the muffle furnace at 550° in presence of magnesium ni-trate, in order to obtain selenium by hydride AAS easily. In ICP-OES, however, these digests permit to determine mainly Cu, Fe, K, Mn, P, S, and Zn, whereas other lines are interfered by the matrix. Addi-tional digest by KClO3 in dilute nitric acid in the pressure bombe heated with microwaves, enabled to enlarge the element sprectrum to get boron, silicon, iodine, and also sodium, which was severely in-terfered from blanks in the Mg-nitrate digests.
Results and Discussion While S and Se were related to the protein and wheat gluten levels of the flour fractions, Zn, P, Mn and K were correlated very strictly (R²= 0,89 – 0,99) to the ash level. Other elements like Fe, Cu and Ca are correlated to the mineral level in a lower degree (R²= 0,67 – 0,84). For some Na and Si no significant correlation to mineral or protein components could be found.
For the distribution of most elements no differences by varietal or environmental influences could be found. But there are some interesting facts that should be mentioned. For example Cu is enriched in the outer layers of all three varieties (fraction 6, ash enriched flour and bran) from Staasdorf. Because no Cu-containing pesticides were applied in these trials other environmental effects might be respon-sible for this. The higher concentration of Se in protein in all fractions of all varieties seems also to be an environmental effect. The results of our investigations confirm, that there are no varietal effects on the mineral levels of flour or flour fractions. The concentration of most elements might only be influ-enced by environmental effects.

Sager und Werteker: Distribution of Elements in Flour Fractions – Elementverteilung in Mahlfraktionen von Weizen
8. BOKU-Symposium Tierernährung 2009 Seite 115
Much more interesting seems to be the distribution and the share on the whole ash level of different minerals in different flour fractions.
Si is one of the main mineral components of the bran. But it is also represented in the fractions 1 – 3, which are characteristic for inner and medium layers of the endosperm. An explanation of this phe-nomenon seems to be difficult.
The proportion of Ca to the whole ash level seems to decrease in the outer layer (fraction 6, ash en-riched flour and bran) while the one of K and Mn increases in these fractions. Zn seems to be weekly enriched in fractions 5 and 6 which correspond to the outer layers of endosperm.
The interpretation of the results might be very complex, but it seems that elements are enriched in these zones of the kernels, where they may participate in biochemical reactions for the synthesis of structure material as in the case of Si. Some results have to be confirmed by further investigations to discuss the biochemical background.
References
M. Sager: Einfluß der Vermahlung auf den Spurenelementgehalt von Pflanzenproben , ALVA- Jahrestagung 1994 Krems, Fachgruppe Futtermittel und Pflanzenanalyse, S. 21-31
M. Sager: Einfluß des Mahlens auf den Spurenelementgehalt einer Gerstenprobe (chinesisch), Szichuan Science and Technology Information on Environmental Protection 1995(4), 15-19
M. Sager: Zur Qualitätskontrolle von Reiben und Mahlen, ALVA - Arbeitsgruppe Pflanzenanalyse, Graz 1996
M. Sager, J. Mittendorfer: Influence of Milling or Cutting Procedures on Trace Element Contents of Plant Samples, Int. Journal of Environmental Analytical Chemistry 67, 59-71 (1997)
M. Sager: Statistische Betrachtungen zur Vermahlung von Getreide, ALVA Jahrestagung Fachgruppe Pflanzenanalyse, Hollabrunn 3.6. 1998
M. Sager, J. Hoesch: Macro- and microelement levels in cereals grown in Lower Austria. Journal of Central European Agriculture 2005, 6(4), 461-472
H. Spiegel, M. Sager: Elementzusammensetzung von Weizen und Kartoffeln in Österreich unter Berücksichtigung des Einflusses von Sorte und Standort. Ernährung/Nutrition 32,, 7/8, 297-308 (2008)
M. Sager, J. Hösch: Spurenelemente in Getreide, Vortrag auf der ALVA – Jahrestagung Wolfpassing, Fachgruppe Pflanzenanalyse und Futtermittel, 2001
M. Werteker und M. Sager: Selen und andere Elemente beim Backen von Semmeln, Vortrag auf der ALVA Jahrestagung 2007, 21. Mai 2007, Stadtschlaining
Corresponding author
Dr. Manfred Sager AGES Wien Analytik Kompetenzzentren, CC-Elemente Wien Spargelfeldstrasse 191 A-1220 Wien E-mail: [email protected]

Schlegel et al.: Effects of supplemental trace mineral (Fe, Cu, Mn, Zn) source and level on post-weaning piglet trace mineral excretion and status
Seite 116 8. BOKU-Symposium Tierernährung 2009
Effects of supplemental trace mineral (Fe, Cu, Mn, Zn) source and level on post-weaning piglet trace mineral excretion and status
P. Schlegel1, L. der Kinderen2, A. Mul2, M. Ubbink-Blanksma2, S. Durosoy3 and E. Bruininx2,4
1 Agroscope Liebefeld-Posieux Research Station, 1725 Posieux, Switzerland 2 CCL Research, N.C.B.-Laan 52, 5462 GE Veghel, The Netherlands 3 Pancosma S.A., Voie des Traz 6, 1218 Le Grand-Saconnex, Switzerland 4 Animal Nutrition Group, Wageningen University, Marijkeweg 40, 6709 PG Wagenin-
gen, The Netherlands
Introduction Trace minerals, such as copper, iron, manganese and zinc are essential for the piglet growth devel-opment as those are highly involved in metabolic functions. These minerals are generally supple-mented to the diet because native concentrations of available minerals in commonly used feedstuffs are generally insufficient to fulfill animal requirements. Mateos et al. (2005) investigated the common-ly practiced trace mineral supplementation on the Iberian Peninsula and concluded that dietary con-centrations are beyond official recommendations. To encourage reasonable trace mineral safety mar-gins (difference between applied and official recommended dietary concentrations) without impairing piglet performance, the partial or total replacement of inorganic by organic trace mineral sources were suggested (eg. Close, 2003; Mullan et al. 2005). Organic trace mineral sources are expected to be superior in their availability than inorganic sources. This potential was demonstrated using organic trace minerals with hydrolyzed protein as ligand (Schiavon et al. 2000; Close, 2003; Creech et al., 2004). Furthermore, organic trace minerals, with an amino acid such as glycine as ligand have dem-onstrated an increased apparent mineral absorbability in weaned piglets when directly compared with hydrolyzed soy based chelates (Ettle et al. 2008; Männer et al. 2008). This indicates that glycinates would have an additional potential for limiting trace mineral safety margins and/or limiting risk of trace mineral deficiencies on individuals within a herd.
The aim of the present study was to investigate the effects of two trace mineral sources (Cu, Fe, Mn and Zn as soy based chelates or glycinates, respectively) supplemented at two levels (in average 75% and 50% of commonly practiced supplementation of inorganic sources) on trace mineral excretion and status of piglets. Since the efficiency in using dietary trace minerals is regulated by homeostatic con-trol, which is highly dependent from the animal’s trace mineral status (eg. Windich and Ettle, 2008) this study was set up to limit initial trace mineral status of the weaning piglets. The growth perfor-mance data was presented previously (Schlegel et al., 2008).
Materials and methods According to body weight (BW), gender and litter, 342 piglets (8.6±0.4 kg BW; 26 d of age) were allocated to 36 pens in 6 rooms. As experimental predisposition, piglets had no access to creep feed and were injected with only half (0.5 ml) of the normal Fe-dextran dose on d 3 of age. These predis-positions were defined to artificially simulate non-optimal starting conditions with regards to trace mineral status.
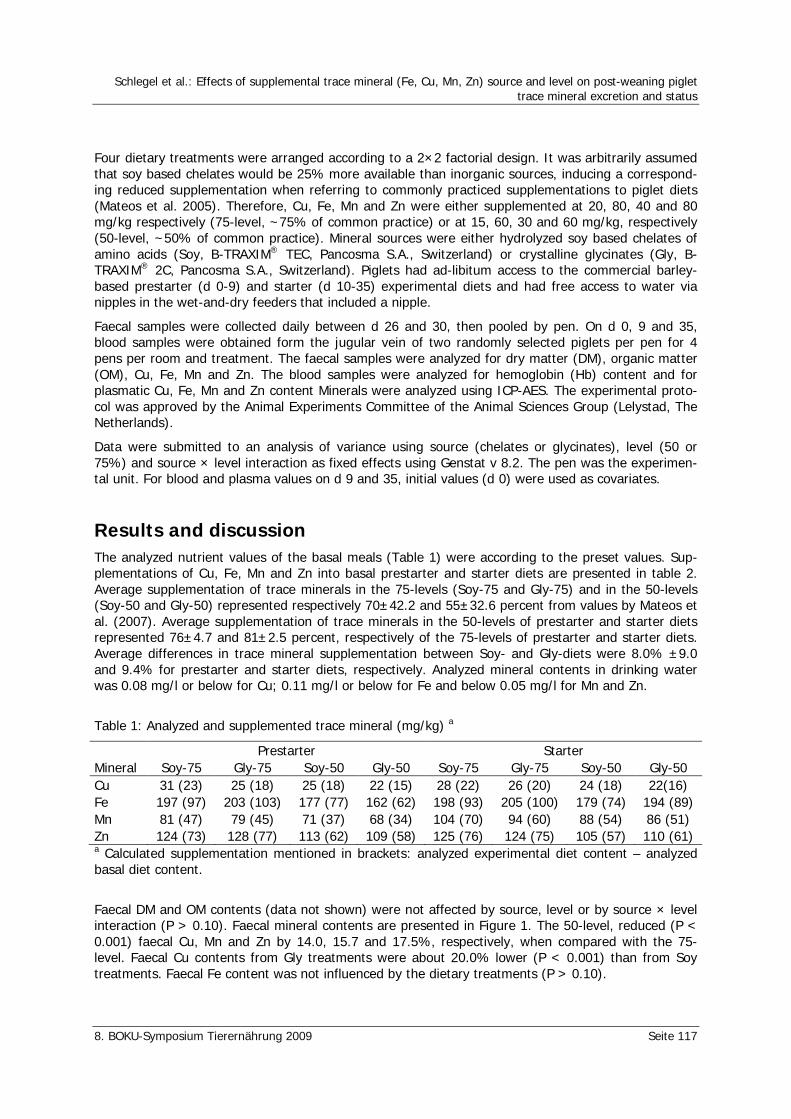
Schlegel et al.: Effects of supplemental trace mineral (Fe, Cu, Mn, Zn) source and level on post-weaning piglet trace mineral excretion and status
8. BOKU-Symposium Tierernährung 2009 Seite 117
Four dietary treatments were arranged according to a 2×2 factorial design. It was arbitrarily assumed that soy based chelates would be 25% more available than inorganic sources, inducing a correspond-ing reduced supplementation when referring to commonly practiced supplementations to piglet diets (Mateos et al. 2005). Therefore, Cu, Fe, Mn and Zn were either supplemented at 20, 80, 40 and 80 mg/kg respectively (75-level, ~75% of common practice) or at 15, 60, 30 and 60 mg/kg, respectively (50-level, ~50% of common practice). Mineral sources were either hydrolyzed soy based chelates of amino acids (Soy, B-TRAXIM® TEC, Pancosma S.A., Switzerland) or crystalline glycinates (Gly, B-TRAXIM® 2C, Pancosma S.A., Switzerland). Piglets had ad-libitum access to the commercial barley-based prestarter (d 0-9) and starter (d 10-35) experimental diets and had free access to water via nipples in the wet-and-dry feeders that included a nipple.
Faecal samples were collected daily between d 26 and 30, then pooled by pen. On d 0, 9 and 35, blood samples were obtained form the jugular vein of two randomly selected piglets per pen for 4 pens per room and treatment. The faecal samples were analyzed for dry matter (DM), organic matter (OM), Cu, Fe, Mn and Zn. The blood samples were analyzed for hemoglobin (Hb) content and for plasmatic Cu, Fe, Mn and Zn content Minerals were analyzed using ICP-AES. The experimental proto-col was approved by the Animal Experiments Committee of the Animal Sciences Group (Lelystad, The Netherlands).
Data were submitted to an analysis of variance using source (chelates or glycinates), level (50 or 75%) and source × level interaction as fixed effects using Genstat v 8.2. The pen was the experimen-tal unit. For blood and plasma values on d 9 and 35, initial values (d 0) were used as covariates.
Results and discussion The analyzed nutrient values of the basal meals (Table 1) were according to the preset values. Sup-plementations of Cu, Fe, Mn and Zn into basal prestarter and starter diets are presented in table 2. Average supplementation of trace minerals in the 75-levels (Soy-75 and Gly-75) and in the 50-levels (Soy-50 and Gly-50) represented respectively 70±42.2 and 55±32.6 percent from values by Mateos et al. (2007). Average supplementation of trace minerals in the 50-levels of prestarter and starter diets represented 76±4.7 and 81±2.5 percent, respectively of the 75-levels of prestarter and starter diets. Average differences in trace mineral supplementation between Soy- and Gly-diets were 8.0% ±9.0 and 9.4% for prestarter and starter diets, respectively. Analyzed mineral contents in drinking water was 0.08 mg/l or below for Cu; 0.11 mg/l or below for Fe and below 0.05 mg/l for Mn and Zn.
Table 1: Analyzed and supplemented trace mineral (mg/kg) a
Prestarter Starter Mineral Soy-75 Gly-75 Soy-50 Gly-50 Soy-75 Gly-75 Soy-50 Gly-50 Cu 31 (23) 25 (18) 25 (18) 22 (15) 28 (22) 26 (20) 24 (18) 22(16) Fe 197 (97) 203 (103) 177 (77) 162 (62) 198 (93) 205 (100) 179 (74) 194 (89) Mn 81 (47) 79 (45) 71 (37) 68 (34) 104 (70) 94 (60) 88 (54) 86 (51) Zn 124 (73) 128 (77) 113 (62) 109 (58) 125 (76) 124 (75) 105 (57) 110 (61) a Calculated supplementation mentioned in brackets: analyzed experimental diet content – analyzed basal diet content.
Faecal DM and OM contents (data not shown) were not affected by source, level or by source × level interaction (P > 0.10). Faecal mineral contents are presented in Figure 1. The 50-level, reduced (P < 0.001) faecal Cu, Mn and Zn by 14.0, 15.7 and 17.5%, respectively, when compared with the 75-level. Faecal Cu contents from Gly treatments were about 20.0% lower (P < 0.001) than from Soy treatments. Faecal Fe content was not influenced by the dietary treatments (P > 0.10).

Schlegel et al.: Effects of supplemental trace mineral (Fe, Cu, Mn, Zn) source and level on post-weaning piglet trace mineral excretion and status
Seite 118 8. BOKU-Symposium Tierernährung 2009
Figure 1: Faecal mineral contents
0
200
400
600
800
1000
1200
Cu Fe Mn ZnMineral
Feca
l con
tent
[m
g/kg
]
Level 50 Level 75 Soy Gly
Average initial blood Hb was 7.28±0.87 g/dl and average initial plasma Fe, Cu and Zn were 0.75±0.65, 2.01±0.18 and 1.40±0.24 mg/dl, respectively. The difference between mean reference values (Kaneko et al., 2008) and initial Hb, plasma Fe and Cu values were -44, -48 and -2%, respec-tively. This study design, similar to the one used by Gentry et al. (1997) and Schlegel et al. (2007) regarding pre-weaning dispositions, permitted to limit piglet’s Fe, Cu (to a certain extent) but not Zn status. Gérard (2000) reported that average piglet Hb on French farms was 10.6 g/dl but varied great-ly between 3.0 and 14.6 g/dl. The initial Fe and Cu status of the values from the present study can therefore be considered as non-optimal, but representative.
Hemoglobin was increased to concentrations above 8 g/dl on d 9 and above 9 g/dl on d 35 (Table 2). The dietary trace mineral level influenced Hb concentration as the 75-level had higher contents than the 50-level on d 9 (+7.7%, P < 0.05) and d 35 (+10.1%, P = 0.05). Trace mineral source did not influence Hb concentration at any moment (P > 0.10).
Compared to the 50-level, the plasma Fe concentration at d 9 was half (P = 0.05) of the average val-ue in the 75-level. On d 35, plasma Fe concentration was still numerically lower for the 50-level. Other plasma trace mineral concentrations were not influenced (P > 0.1) by the dietary treatments. Under limited trace mineral status the homeostatic regulation sets to maximize trace mineral retention (Spears and Hansen, 2008, Windisch and Ettle, 2008), which permits to express the potential advan-tages of a trace mineral source compared to another. This was not the case for Cu and Zn in the pre-sent study.
Reducing the amounts of supplemented trace minerals permitted to limit Cu, Mn and Zn excretion, but limited also Hb and plasma Fe. The final Hb and plasma Fe were even below the lower limit of the reference values (Kaneko et al., 2008), indicating Fe deficiency. The 50-level was therefore not pro-
* *
*
*
* : P < 0.001

Schlegel et al.: Effects of supplemental trace mineral (Fe, Cu, Mn, Zn) source and level on post-weaning piglet trace mineral excretion and status
8. BOKU-Symposium Tierernährung 2009 Seite 119
viding sufficient available trace minerals, especially Fe, to improve Fe status in piglets with initial low Fe and Cu plasma concentrations. The 50-level also presented less efficiency on growth and increased BW heterogeneity than the 75-level (Schlegel et al., 2008).
Table 2: Blood Hb and plasma mineral contents. a
Source Level Soy Gly P-value SEM 75 50 P-value SEM
Hb d9 (g/dl) 8.64 8.54 n.s. 0.174 8.91 8.27 < 0.05 0.174 Hb d35 (g/dl) 9.49 9.65 n.s. 0.263 10.02 9.10 0.05 0.263 Fe d9 (mg/dl) 0.66 0.86 n.s. 0.165 1.02 0.50 0.05 0.165 Fe d35 (mg/dl) 0.91 1.07 n.s. 0.160 1.14 0.84 n.s. 0.160 Cu d9 (mg/dl) 1.75 1.71 n.s. 0.046 1.70 1.76 n.s. 0.046 Cu d35 (mg/dl) 1.89 1.87 n.s. 0.069 1.87 1.89 n.s. 0.069 Zn d9 (mg/dl) 1.15 1.11 n.s. 0.030 1.15 1.10 n.s. 0.030 Zn d35 (mg/dl) 1.15 1.18 n.s. 0.034 1.19 1.14 n.s. 0.034 a Data adjusted with values from day 0 n.s. = P > 0.10
Conclusion The present data suggest that piglets presenting limited Fe and Cu status at weaning can improve their mineral status within 35 days when fed lower levels than commonly supplemented. Feeding lower levels of trace minerals permitted to reduce their faecal excretion. Under such conditions, feed-ing trace minerals as glycinates instead of soy based chelates, were beneficial regarding Cu excretion and comparable regarding blood contents.
Acknowledgments
We wish to thank the staff at Laverdonk Research farm and Hans Zwolschen DVM for their assistance during this study.
References
Close W.H. (2003). Trace mineral nutrition in pigs: working within the new recommendations. Feed Compounder, 14-18.
Creech B.L., Spears J.W., Flowers W.L., Hill G.M., Lloyd K.E., Armstrong T.A., and Engle T.E. (2004). Effect of dietary trace mineral concentration and source (inorganic vs. chelated) on performance, mineral status, and fecal mineral excretion in pigs from weaning through finishing. J. Anim. Sci. 82, 2140-2147.
Ettle T., Schlegel P. and Roth F.X. (2008). Investigations on iron bioavailability of different sources and supply levels in piglets. J. Anim. Physiol. Anim. Nutr. 92, 35-43.
Gentry J.L., Swinkels J.W.G.M., Lindemann M.D. and Schrama J.W. (1997). Effect of hemoglobin and immunization status on energy metabolism of weanling pigs. Journal of Animal Science 75, 1032 – 1040.
Gérard C. (2000). Les besoin de fer revus à la hausse. Réussir Porc, n°58, 26-27.
Kaneko JJ., Harvey JW. and Bruss ML. (2008). In: Clinical biochemistry of domestic animals. 6th Edition. Elsevier Academic Press. Burlington, MA, USA.

Schlegel et al.: Effects of supplemental trace mineral (Fe, Cu, Mn, Zn) source and level on post-weaning piglet trace mineral excretion and status
Seite 120 8. BOKU-Symposium Tierernährung 2009
Mahan D. (2005). Feeding sow and piglet to achieve maximum antioxidant and immunity protection. In: Re-defining Mineral Nutrition, J.A. Taylor-Pickard and L.A. Tucker (Eds). Nottingham University Press, Nottingham, United Kingdom. 63-73.
Männer K., Simon O. and Schlegel P. (2008). Bioavailability of trace minerals sources in swine. In: Trace elements in animal production systems, P. Schlegel, S. Durosoy and A.W. Jongbloed (Eds), Wageningen Academic Publishers, Wageningen, The Netherlands. 177-186.
Mateos G.G., Lazaro R., Astillero J.R. and Perrez Serano M. (2005). Trace Minerals: What text books don’t tell you. In: J.A. Re-defining Mineral Nutrition. J. A. Taylor-Pickard and L.A. Tucker (Eds), Nottingham University Press, Nottingham, United Kingdom. 21-61.
Mullan B.P., Hernandez A., D’Souza D.N. and Pluske J.R. (2005). Modern pig nutrition for performance: minerals, metabolism and the environment. In: Nutritional Biotechnology in the Feed and Food Industries, Proceedings of Alltech’s 21st Annual Symposium, T.P. Lyons and K.A. Jacques, (Eds) Nottingham University Press, United Kingdom. 185 - 200.
Schiavon S., Bailoni L., Ramanzin M., Vincenzi R., Simonetto A., and Bittante G. (2000). Effect of proteinate or sulphate mineral sources on trace elements in blood and liver of piglets. Anim. Sci. 71, 131-139.
Schlegel P., Durosoy S. and Dupas M. (2007). Iron status evolution of weaned piglets either fed iron sulfate or iron glycinate. In 6. BOKU Symposium Tierernährung, C. Plitzner, M. Kraft and W. Windisch (Eds), University of Natural Resources and Applied Life Sciences, Vienna, Austria. 273 - 278.
Schlegel P., der Kinderen L., Mul A., Ubbink-Blanksma M., Durosoy S. and Bruininx E. (2008). Effects of supplemental trace mineral (Fe, Cu, Mn, Zn) source and dose on growth performance in weaned piglets. In 7. BOKU Symposium Tierernährung, C. Plitzner, M. Kraft and W. Windisch (Eds), University of Natural Resources and Applied Life Sciences, Vienna, Austria. 225-228.
Spears J.W. and Hansen S.L. (2008). Bioavailability criteria for trace minerals in monogastrics and ruminants. In: Trace elements in animal production systems, P. Schlegel, S. Durosoy and A.W. Jongbloed (Eds), Wageningen Academic Publishers, Wageningen, The Netherlands. 161-176.
Windisch W. and Ettle T. (2008). Limitations and possibilities for progress in defining trace mineral requirements of livestock. In Trace elements in animal production systems, P. Schlegel, S. Durosoy and A.W. Jongbloed (Eds), Wageningen Academic Publishers, Wageningen, The Netherlands. 187 – 202.
Autorenanschrift
Patrick Schlegel Agroscope Liebefeld-Posieux Research Station ALP Tioleyre 4 1725 Posieux Switzerland E-Mail: [email protected]

Seresinhe et al.: Effect of concentrate–mineral supplements on the overall nutritional status of cross bred milking cows in two small farms in Sri Lanka
8. BOKU-Symposium Tierernährung 2009 Seite 121
Effect of concentrate–mineral supplements on the overall nutri-tional status of cross bred milking cows in two small farms in Sri Lanka
Thakshala Seresinhe1, A. Manawadu1, Christine Iben2 1 Dept. of Animal Science, Faculty of Agriculture, University of Ruhuna, Sri Lanka 2 Department of Farm Animals and Veterinary Public Health, Institute of Nutrition,
University of Veterinary Medicine, Vienna, Austria.
Introduction The primary constraint to livestock production in Sri Lanka is the scarcity and fluctuating quantity and quality of the year round feed supply. The country experience serious shortages of animal feeds and fodders of the conventional type especially during the dry spells. Fibre rich low protein forages and crop residues are the most abundant and appropriate feeds for ruminants during the times of feed scarcity. However, the energy value of ruminant feeds is generally affected by their fiber content be-cause this is often inversely related to digestibility. Since the utilization of fiber from these roughages depends primarily on microbial fermentation, supplementation of critical nutrients for microbial activity is the most important basic strategy towards reaching an optimum production.
In Sri-Lanka, the up grading of low quality roughages for ruminants is still a central issue as a strategy for improving ruminant production. Roughages are poor in protein, energy, minerals and vitamins. Addition of foliage from tree leaves or supplementation with urea and molasses can also improve the utilization of low quality roughages through the supply of nitrogen to rumen microbes. Therefore, it was identified that the strategy for improving production has been to maximize the efficiency of utili-zation of available feed resources and supplement dietary nutrients using locally available feeds. How-ever, farmers are often unaware of appropriate feeding strategies for combining poor quality feeds with high quality available green fodder resources and low cost concentrate-mineral supplements to improve ruminant production. Therefore, objectives of this study were to educate farmers to develop feeding strategies to boost their production level especially where feeds are scarce.
Materials and methods Two trials were carried out with milking cows on the principle of a 4 x 4 Latin square design. Cows were selected considering the breed, live weight, parity and average milk yield etc. The first trial was conducted in a small farm using Australian Frisian Sahiwal (AFS) cross bred milking cows of 185±5.4 kg body weight (BW). The farm was placed in Sapugoda, Kamburupitiya, in the low country wet zone of Sri Lanka.
Four milking cows were fed the following four diets. Diet 1 (farm diet): approximately 10 % of body weight fresh herbage - mainly rice field weeds Diet 2: Farm diet + low cost concentrate supplement (GAVA TRIPOSHA* - 250 g/head/day) Diet 3: Farm diet + 2 kg fresh forage of Gliricidia sepium (tree fodder) Diet 4: Farm diet + 2 kg fresh forage of Gliricidia sepium + low cost concentrate supplement (GAVA TRIPOSHA*)
*Low cost concentrate supplement (GAVA TRIPOSHA)- rice bran 35 %, urea 12 %, molasses 45 %, mineral mixture 8 %

Seresinhe et al.: Effect of concentrate–mineral supplements on the overall nutritional status of cross bred milking cows in two small farms in Sri Lanka
Seite 122 8. BOKU-Symposium Tierernährung 2009
The second trial was conducted in a medium scale coconut–cattle integrated farm in Gonapinuwala, Galle, in the low country wet zone of Sri Lanka using cross bred Jersy cows (300±4.5 kg BW). Four milking cows were fed on following diets on the principle of a Latin square design.
Diet 1 (farm diet): 25 kg fresh forages - Forage consists of Brachiaria ruziziensis and Pennisetum perpureum – var. Co3, 1:1 ratio Diet 2: Farm diet + low cost concentrate supplement (GAVA TRIPOSHA-750 g/head/day) Diet 3: Farm diet + 2 kg fresh forage of Gliricidia sepium (tree fodder) + low cost concentrate supplement (GAVA TRIPOSHA-750 g/head/day) Diet 4: Farm diet + farm made concentrate mixture** (2 kg/head/day)
** Rice bran 40 %, Coconut Poonac 40 %, Soya bean meal 16 %, Mineral mix. 4 %.
In both trials, the preliminary period lasted 20 days; the duration of the trial period was 7 days. The cows were weighed prior to each trial period using weigh band and actual weights were calculated using the regression equation of Seresinhe and Pathirana (2007).
In the trial period, feeds, feed residues and faeces were sampled and dried at 65 °C and milled. Dry matter (DM) was determined by drying at 105 °C to constant weight. Crude protein (CP) content was determined according to the Kjeldahl method (N x 6.25), crude fiber content by Fibertec system, eather extract content by Soxhlet system and ash content by combustion at 550 °C (AOAC 1985). Cell wall components were determined with detergent solutions according to the method by Van Soest and Robertson (1980).
In the trial period milk yields were recorded daily while milk samples were collected during the last three days of the trial period and analyzed for fat (Gerber method) and total solids (Lactometer method). During the trial period spot urine samples (at least 4 samples from each animal) were col-lected during the last two days of each trial period to cover 24 hrs, between 08-12, 12-15, 16-20 and 20 to 08 hrs next morning (Chen and Gomez, 1995). The urine samples were processed, preserved and analyzed for allantoin, uric acid and creatinine according to the procedures of IAEA TECHDOC (1997).
Total urinary and milk nitrogen contents were determined by Kjedahl procedure.
Follow ing calculations have been done
PD (purine derivatives):Creatinine ratio in urine (PDC INDEX) was calculated according to the follow-ing formulae (Seresinhe et al., 2004):
PDC INDEX = W is the body weight (kg). PD and Creatinine are their concentrations in mmol/l.
Using the PDC index, the PD excretion (mmol/d) was calculated using the following equation:
PD excretion (mmol/d) = (PDC index) C
C is the daily creatinine excretion (mmol/kg W0.75) for specific breed of animals of Sri Lankan cross bred cattle. The creatinine excretion averaged 0.96 mmol/kg W0.75/d (Seresinhe et al., 2004).
Using the above data, absorbed exogenous purine concentration (mmol/d) was derived as follows:
Y= (0.385kg W0.75) + 0.85X Y = Urinary PD excretion (mmol/d) X = absorbed exogenous purine as mmol/d W = live weight (kg)
[PD]x W0.75
[Creatinine]

Seresinhe et al.: Effect of concentrate–mineral supplements on the overall nutritional status of cross bred milking cows in two small farms in Sri Lanka
8. BOKU-Symposium Tierernährung 2009 Seite 123
Microbial nitrogen yield was calculated using above data as follows according to Chen et al. (1995):
Microbial N (g N / d) = Milk nitrogen efficiency was calculated according to Isler (2009): Milk nitrogen efficiency (%) for milk (MNE %) =
Table 1: Proximate composition of feeds
Items Natural forage
(rice field weeds)
G. sepium Low cost concentrate
Mixture
B. ruzizien-sis Co3 mixture
Farm made concentrate
mixture
Dry matter % 22.25 21.6 90 31.3 90 Composition of DM Crude protein % 9.69 23.2 37.6 10.21 22.24 Ash % 8.7 7.6 6.4 8.5 7.0 Crude fiber % 24.2 20.7 7.9 21.8 7.6
Results Trial 1
The dry matter intake of cows was highest with diet 4 (farm diet + concentrate supplement + tree fodder) followed by diet 2 (farm diet + concentrate supplement). The intake of digestible organic matter and nitrogen were also highest when the farm diet was supplemented with concentrate sup-plement and tree fodder (diet 4). The markedly higher N intake in diet 4 was more than two fold higher when compared with the N intake of diet 1 (farm diet). Highest milk yield was observed in cows when diet 4 was given together with highest amounts of milk fat, total solids and nitrogen per-centages. However, milk nitrogen efficiency (MNE) was highest when the farm diet was supplemented with low cost concentrate supplement (diet 2) followed by supplementation with concentrate and tree fodder (diet 4). Estimated total PD ranged from 18.58 mmol/day to 30.5 mmol/day for diet 1 and diet 4, respectively, in clear response to the supplementation. The amount of microbial nitrogen synthe-sized by experimental cows per day ranged from 8-15 g for diet 1 and 15-22 g for diet 4 respectively. The estimated PDC index was highest with diet 4 followed by diet 3 and diet 2 and was lowest with diet 1.
Trial 2
The highest dry matter intake was observed in milking cows fed diet 3 (farm diet + tree fodder + concentrate supplement) followed by cows fed diet 2 (farm diet + concentrate supplement). Digesti-ble organic matter intake was highest in cows fed farm diet + farm made concentrate ration (diet 4) followed by cows fed farm diet + concentrate supplement + tree fodder (diet 3). The Nitrogen intake was highest in cows fed concentrate supplement in addition to farm diet (diet 2) followed by cows fed concentrate mixture + tree fodder (diet 3) and supplementation of farm made concentrate rations (diet 4).
Higher milk yield was observed with cows supplemented with farm made concentrate ration (diet 4) followed by cows given concentrate supplement + tree fodder (diet 3). In contrast, feeding of concen-trate supplement + tree fodder together (diet 3) showed positive effects on increasing the milk fat
Daily N output in milkx 100
Daily N intake from feeds
X (mmol/d) x 700.727X
0.116 x 0.83 x 1000

Seresinhe et al.: Effect of concentrate–mineral supplements on the overall nutritional status of cross bred milking cows in two small farms in Sri Lanka
Seite 124 8. BOKU-Symposium Tierernährung 2009
percentage and total solids. Nitrogen percentage was highest in cows fed concentrate supplement (diet 2).
The milk nitrogen efficiency of cows was highest in diet 2 but substantially improved through supple-mentation of low cost concentrate mineral mixture and tree fodder (diet 3). The PD:Creatinine index (PDC Index) and estimated daily PD excretion (mmol/d) also responded significantly towards supple-mentation. PDC index and total PD (mmol/d) exctretion showed slight changes in diet 3 and diet 4. The amount of microbial nitrogen synthesized by experimental cows per day ranged from 22-32 g for diet 1 and 35-40 g for diet 4 respectively.
Discussion The results of this study suggest that the overall nutritional status of lactating cows were different in two farms due to breed characteristics, body weights of milking cows and quality of forages etc. The farm diet fed to cows in trial 1 mainly contained rice field weeds and the dry matter and crude protein contents were much lower than that of the farm diet used in trial 2 which consisted of improved grasses (Brachiaria ruziziensis and Pennisetum perpureum – var. Co3). However, in both trials the quality of “on farm” diet was substantially improved by supplementation of low cost concentrate-mineral mixture and tree fodder. A superior nutritional quality of supplements resulted in a higher intake of feed and N and other nutrients as well. Therefore, results confirm that supplementation is necessary to improve the milk production of local cows under small farmer conditions. Results of the trail 2 confirm that concentrate supplement and tree fodder were more effective as compared with high cost farm made concentrate ration (FMC) in improving the quality of milk rather than the yield. In both trials the values for the nitrogen efficiency for milk (MNE) was much lower than the sufficient value (>20) as indicated by Isler (2009). However, indications are that there are opportunities to im-prove the MNE through supplementation. Findings of Kauffman and St. Pierre (2005), suggested that in mature, non-growing lactating cows the N retentions should be near zero because N intake and N output (Milk N + faecal N + Urine N) amounts are very close to equal. Therefore, an increase in MNE could be expected if the quality of the diet improves. In both trials, investigations were done using mature lactation cows. It can be suggested that the effects of MNE in this trails would have been mainly associated with supplementation.
It was also clearly evident that supplementation of low cost concentrate mixture and tree fodder showed more or less similar response with Purine derivative excretion and microbial nitrogen produc-tion as well. Cantalapiedra et al. (2009) reported that urinary excretion of purine derivatives increased with increasing concentrate in animal fed diets with grass hay. Further, our findings are consistent with those reported by numerous workers, that the efficiency of microbial protein synthesis as meas-ured by urinary PD responded strongly to dry matter and CP intake (Puchala and Kulasek, 1992; Dapoza et al., 1990), while Antoniewicz et al. (1980) reported that the endogenous PD excretion can change as a result of alternations in the protein supply. PD excretion results of Nsahlai et al. (2000) and Long et al. (1999) are in close agreement with the present findings.

Seresinhe et al.: Effect of concentrate–mineral supplements on the overall nutritional status of cross bred milking cows in two small farms in Sri Lanka
8. BOKU-Symposium Tierernährung 2009 Seite 125
Table 2: Main results of the trails
Parameter Trial 1 Diet
1 2 3 4 FD FD+GT FD+TF FD+GT+TF
Trial 2 Diet
1 2 3 4 FD* FD+GT FD+GT+TF FD+FMC
Dry matter intake (kg/head/day)
4.58c 5.10b 5.05b 5.25a
±0.08 ±0.05 ±0.08 ±0.08
6.90a 6.99a 7.04a 6.92a ±0.45 ±0.33 ±0.23 ±0.36
Digestible organic matter intake (kg/head/day)
1.26c 1.45b 1.42b 1.82a ±0.03 ±0.04 ±0.05 ±0.06
3.53d 4.07c 4.52b 5.00a ±0.12 ±0.14 ±0.18 ±0.16
Nitrogen in-take (g/head/day)
60.1d 110.2b 90.3c 132.2a
±1.3 ±1.4 ±1.1 ±1.6
121.8c 178.4a 162.4b 165.5b ±2.0 ±3.0 ±2.0 ±1.0
Milk Parameters Milk yield (kg/head/day)
2.20c 3.30b 3.39b 4.26a
±0.10 ±0.19 ±0.21 ±0.13
4.0c 4.53b 4.50b 4.90a
±0.15 ±0.14 ±0.16 ±0.12
Milk fat g/kg
34c 36.3c 38b 41.3a
±0.57 ±0.53 ±0.49 ±0.48
44.4c 53a 49b 42c ±0.8 ±0.9 ±0.7 ±1.1
Total solids (g/kg)
119b 124a 122a 131a ±1.0 ±1.0 ±1.0 ±1.5
127b 137.8a 134.3a 126b ±1.3 ±1.2 ±1.4 ±1.3
Milk nitrogen (g/kg)
3.5a 4.0b 3.7a 4.4a ±0.2 ±0.4 ±0.3 ±0.4
4.7b 5.1a 4.7b 4.7b ±0.1 ±0.4 ±0.2 ±0.3
Milk Nitrogen efficiency (MNE)
12.8a 15.6a 13.9a 14.2a ±1.1 ±1.4 ±1.3 ±1.4
15.7a 17.5a 16.1a 14.5a ±1.1 ±1.4 ±1.2 ±1.3
Urinary excretion of Purine derivatives (PD) Estimated total PD (Mmol/day)
18.58c 22.0c 25.9b 30.5a ±1.21 ±1.43 ±1.23 ±1.65
40c 44b 54a 56a
±1.31 ±1.33 ±1.43 ±1.55 Microbial nitrogen (g/day)
8-15 15-22 15-22 22-32 20-24 25-27 30-39 35-40
PD/Creatinine Index (PDC index)
19d 23c 27 b 32a
27c 35b 55a 56a
Nutritional status
UF M M S M S G G
FD farm diet (mainly rice field weeds); FD* farm diet (Brachiaria ruziziensis); TF tree fodder (Gliricidia sepium); GT low cost concentrate supplement (GAVA TRIPOSHA); FMC farm made concentrate mixture; UF Under feeding; M Maintenance; S Satis-factory; G Good;

Seresinhe et al.: Effect of concentrate–mineral supplements on the overall nutritional status of cross bred milking cows in two small farms in Sri Lanka
Seite 126 8. BOKU-Symposium Tierernährung 2009
Low cost concentrate–mineral mixture and freely available tree fodders are best supplements to im-prove the production potential of local milking cows. Supplementation of TF or GT could enhance the microbial nitrogen production to reach the maintenance level of feeding. Supplementation both with GT and TF could boost the microbial nitrogen production to reach a satisfactory feeding level (ta-ble 2). In the second experiment, the better feeding value of improved grasses alone could meet the maintenance requirement but with tree fodder and concentrate supplement or farm made concentrate mixture a better level of nutritional status could be reached. It is a well known fact that the quality of diets may influence the microbial nitrogen supply (Seresinhe and Pathirana 2008). Therefore, the find-ings of this study suggest that feeding of tree fodder and GT could enhance the nitrogen content of herbage based diets thereby improving the microbial N production which is an indication of increased post ruminal microbial protein available to the animal.
The results of these two trials confirm that agricultural residues and commonly fed natural herbage to local cross bred cattle could be greatly improved by supplementation. Tree fodder compared to con-centrate type feed ingredients have greater potential in that regard particularly under resource limited small farmer conditions. However, the contribution of tree fodder and low cost concentrate –mineral mixture were the best. There is potential to increase the level of both types of supplements for higher level of microbial nitrogen production. To improve the animal performance, the rations can be formu-lated according to their actual nutrient requirements. However, by promoting farmers to grow more tree fodder, the amount of concentrate and therefore the cost of production could be reduced under tropical developing country situations for optimal rather than maximum levels of production under such conditions.
References
Antoniewicz A.N., Heinemann W.W. and Hanks E.M. (1980): The effect of changes in the flow of nucleic acids on allantoin excretion in the urine of sheep. J. of Agric. Science 95, 395-400.
AOAC Association of Official Agricultural Chemists, Official methods for analyses (13th Ed.) (1985): Association of the official analytical chemists, Washington, DC.
Cantalapiedra-Hijar G., Yanez-Ruiz D.R., Martin–Garcia A.I. and Molina-Alcaide E. (2009): Effects of forage: concentrate ratio and forage type on apparent digestibility, ruminal fermentation and microbial growth in goats J. Anim. Sci. 87,622-631.
Chen X.B., Mejia A.T., Kyle D.J. and Ørskov E.R. (1995): Evaluation of the use of purine derivative; creatinine ratio in Spot urine and plasma samples as an index of microbial protein supply in ruminants: in sheep. J. of Agric. Science 125, 137-143.
Chen X.B. and Gomez M.J. (1995): Estimation of microbial protein supply to sheep and cattle based on urinary excretion of purine derivatives. An overview of the technical details. Occasional publication international feed resources unit, Rowett Research Institute, Bucksburn, Aberdeen, U.K.
Dapoza C., Castrillo C., Balcells J., Martin-orue S. and Guada, J.A. (1999): On the variation of urinary excretion of creatinine and purine derivatives in pregnant and lactating ewes given diets with different protein contents. J. of Agric. Science 68, 555-556.
IAEA TECHDOC-945 (1997): International atomic energy agency, Vienna, Austria.
Isler V. (2009): Enhancing nitrogen efficiency by lactating dairy cows. http//www.agriculture.state.pc.US.agriculture/lib (browsed in Aug. 01, 2009).
Kauffmann A.J. and St-Pierre N.R. (2005): Effect of breed and concentrations of dietary crude protein and fiber on milk urea nitrogen. Research and Reviews: Dairy special circular 169-99, Ohio State University, USA.
Long R.J., Dong S.K., Chen X.B., Ørskov E.R. and Hu Z.Z: Preliminary studies on urinary excretion of purine derivatives and creatinine in yaks. J. of Agric. Science 133, 427-431.
Nsahlai I.V., Oosuji P.O. and Umunna N.N. (2000): Effect of form and quality of feed on the concentrations of purine derivatives in urinary spot samples, daily microbial N supply and predictability of intake. Animal Feed Science and Technology 85, 223-238.
Puchala R. and Kulasek G.W. (1992): Estimation of microbial protein flow from the rumen of sheep using microbial nucleic acid and urinary excretion of purine derivatives. Canadian Journal of Animal Science 72, 821-830.

Seresinhe et al.: Effect of concentrate–mineral supplements on the overall nutritional status of cross bred milking cows in two small farms in Sri Lanka
8. BOKU-Symposium Tierernährung 2009 Seite 127
Seresinhe T. and Pathirana K.K. (2008): Effect of supplementation of straw based diets on the digestibility and microbial nitrogen production of cross bred cattle. World Journal of Agricultural Sciences 4, 745-751.
Seresinhe T. and Pathirana K.K. (2007): Effect of feeding straw and supplements on the performance of cattle under small farmer conditions. Tropical Agricultural Research and Extension 9, 131-141.
Seresinhe T., Pathirana K.K. and Jayasuriya M.C.N. (2004): Urinary excretions of purine derivatives (PD) as a predictor of the nutritional status of local zebu-cattle and cross- bred milking cows. In: Harindra P.S., Makkar and Chen X. B. (Eds.): Estimation of microbial protein supply in ruminants using urinary purine derivatives. Kluwer Academic Publishers, pp 95-102.
Van Soest P.J., and Robertson J. B. (1980): Systems of analysis for evaluating fibrous feeds. In: Pigden W.J., Balch C.C. and Graham M. (eds): Standardization of Analytical Methodology in Feeds. International Research Development Center, Ottawa, Canada.
Corresponding author
Prof. Christine Iben Department of Farm Animals and Veterinary Public Health, Institute of Animal Nutrition, University of Veterinary Medicine, Vienna, Vaterinärplatz 1, 1210 Vienna, Austria. E-mail: [email protected]

Firmensponsoring – Inntaler Mischfutter GmbH & CO. KG, Klötzlmüllerstraße 140, D-84034 Landshut
Seite 128 8. BOKU-Symposium Tierernährung 2009

Vacchina et al.: Determination of Zn-glycinate complexes in diluted media
8. BOKU-Symposium Tierernährung 2009 Seite 129
Determination of Zn-glycinate complexes in diluted media
Véronique Vacchina1, Sébastien Oguey2, Ryszard Lobinski3, David Bravo2
1 UT2A, Pau, France 2 Pancosma, Geneva, Switzerland 3 CNRS, Pau, France
Introduction Some trace elements are known to be essential (Zn, Cu, Fe, Mn…) especially in the nutrition field. It is commonly admitted that their bioavailability highly depends on their chemical form. In the last dec-ade, the interest in using alternative mineral sources, particularly those chelated with proteins or amino acids, increased due to their higher bioavailability compared to inorganic sources (1). However, efficiency of organic sources themselves may vary in regard to the type of ligand involved (2). One of the most efficient seems to be glycinate complexes. Efficiency of supplementation by glycinate com-plexes have been shown for several micronutrients (3, 4, 5). It becomes therefore of primary impor-tance to prove the complexation of the metal with the glycine in the food supplements used.
The Zn-glycinate standard complex investigated in this work was previously characterized by X-ray diffraction. The metal was shown to be complexed with glycine, water and sulfate according to the formula detailed in the “materials and method” part of this paper. However, the stability of the mo-lecular structure in solution still needs to be checked. This must be addressed at three different levels: (i) stability in solution; (ii) stability after acidification to approach gastric conditions; (iii) stability dur-ing chromatographic separation in view of further speciation analysis.
The molecular structure of the glycinate complexes in solution, whatever the pH, can be assessed by ESI MS(/MS). Indeed, it is now a commonly used technique for the characterization of non-covalent organo-metallic species (6). Metal presence can be confirmed by its isotopic profile and structural in-formation on the ligands involved can be gained in the MS/MS mode. Moreover, it is more sensitive and less sample consuming than for example NMR which is another popular technique for analysis of such species (7). Complexation of trace elements with glycine have been widely studied by ESI MS(/MS) (8, 9, 10, 11, 12). But those results were obtained only after incubation of the trace elements with the glycine in-vitro and not from resolubilisation of solid complexes.
The stability during chromatographic process is the second parameter to check to establish the stabil-ity of the molecular integrity of the glycinate complexes. SE HPLC appears to be the most promising tool for this. Indeed SE HPLC has the advantage over other HPLC mechanisms of the high tolerance to biological matrices. However, dissociation of the complexes on the stationary phase of the column may occur (13). Therefore care must be taken to control the recovery from the column and elution conditions need to be optimized.
The aim of this work was therefore to characterize the Zn-glycinate complexes in solution by ESI MS(/MS) and to study their stability over acidification and during size-exclusion liquid chromatography.

Vacchina et al.: Determination of Zn-glycinate complexes in diluted media
Seite 130 8. BOKU-Symposium Tierernährung 2009
Materials and methods
Apparatus, reagents and standards
The electrospray QqTOF mass spectrometer was a QSTAR XL (Applied Biosystems, Foster City, CA, USA) used either in full scan TOF mode or in product ion (MS/MS) mode. Before the experiments, the mass accuracy of the instrument was calibrated with a reserpin standard : [M+H]+ 609.28121 (C33H40N2O9). The optimum settings were ion spray voltage : 4850 V; curtain gas : 25 V; GS1 : 17 V; GS2 : 0 V; collision gas : between 15 and 25 eV of N2 depending on the compound. TOF MS accurate mass spectra were recorded across the range m/z 70-2000.
Chromatographic separations were performed using a Model 1100 HPLC pump (Agilent, Wilmington, DE, USA) as the delivery system. Injections were made using a Rheodyne valve with a 100 µL loop. The exit of the column was directly connected to the Meinhardt nebulizer (Glass Expansion, Romain-motier, Switzerland) of the ICP MS equipped with a collision cell (Agilent 7500ce, Agilent, Tokyo, Ja-pan) by means of PEEK tubing.
Analytical reagent grade chemicals purchased from Sigma-Aldrich (Saint-Quentin Fallavier, France) and water (18 MΩ.cm) obtained with a Milli-Q system (Millipore, Bedford, MA) were used throughout unless stated otherwise.
The Zn-glycinate ([Zn(C2H5NO2)(H2O)2(SO4)]n) complexes were obtained from Pancosma (Geneva, Switzerland).
Size-exclusion chromatography
The size exclusion column was Superdex Peptide HR 10/30 (300 x 10 mm i.d.) (Pharmacia Biotech, Uppsala, Sweden). Mobile phases were prepared by dissolving the corresponding amount of ammo-nium acetate to reach the ionic strength aimed and by adjusting the pH by dropwise addition of acetic acid or ammonium hydroxide. The column was cleaned after each run by flushing with two bed vol-umes of 50 mM EDTA solution, followed by one bed volume of 0.5 M NaOH solution to remove EDTA and avoid artefacts of metal-EDTA in subsequent separations. The column was then stabilized with the following mobile phase to be used. The flow rate was set at 0.7 mL.min-1.
Results
Characterization of the Zn-glycinate complex in ESI QqTOF MS(/MS)
Direct infusion of Zn-glycinate complexes in the electrospray source leads to numerous peaks in the full mass spectrum. The peaks of interest were distinguished according to characteristic zinc isotopic patterns. Zinc has four isotopes which differ in mass by respectively 2-1-1 amu and with relative ab-undances 64Zn/66Zn/67Zn/68Zn of 49/28/4/19. This distinctive isotopic pattern enables a zinc-containing ion to be readily identified. This profile changes if the ion contains two or more zinc atoms according to the statistic arrangement between the different zinc isotopes.
Figures 1 show zooms of mass ranges presenting characteristic isotopic patterns in the mass spectra obtained for the Zn-glycinate complex. Two zinc characteristic isotopic patterns were found. The first one with m/z 253.91 for the 64Zn is characteristic for the presence of one zinc atom in the ion whereas the second one, with a most intense peak at m/z 472.84 is characteristic for the presence of two zinc atoms in the ion. Taking into account the expected chemical structure and the molecular mass expe-rimentally measured, the formulas of [Zn(Gly)(H2O)(SO4)] and [Zn(Gly)(SO4)]2 were respectively pro-posed.

Vacchina et al.: Determination of Zn-glycinate complexes in diluted media
8. BOKU-Symposium Tierernährung 2009 Seite 131
252.0 254.0 256.0 258.0 260.0
1.0
253.9106
255.9086
257.90380.5
x 104
253,9307 255,9277 257,9264 259,9253
a
469.0 471.0 473.0 475.0 477.0 479.0 481.0 483.0
1
2
x 103472.8467 474.8456
470.8490
476.8437
478.8456
470,8331 472,8301 474,8282 476,8260 478,8245
b
Figure 1: zooms of the characteristic Zn-containing patterns in the MS spectra of Zn-glycinate com-
plex. a) [Zn(Gly)(H2O)(SO4)+H]+. b) [Zn(Gly)(SO4)2+H]+. The theoretical isotopic pattern is given in inset.
The confirmation of the molecular structure of the Zn-glycinate complexes was obtained in the MS/MS mode. Product ion scan spectra were acquired for two parent ions containing different zinc isotopes to definitively identify metal-containing product ions. The reaction diagrams associated with the frag-mentation of the molecular ions of interest are shown Figures 2. Losses of mass of 98 and 80 amu are the most frequently observed. They correspond to the losses of H2SO4 and SO3 respectively which are characteristic to the presence of sulfate in the molecule. The presence of sulfate is also sustained by the loss of the neutral entity ZnSO4 from the n = 2 complex. The ion with m/z 470.85 contains Zn only as 64Zn and therefore can only loose 64ZnSO4 to give a single fragment at m/z 310.94. The corres-ponding molecular ion at m/z 472.86 contains two zinc atoms that can only be one 64Zn and one 66Zn. When 64ZnSO4 is the neutral entity lost, it gives a fragment at m/z 312.93. If it is 66ZnSO4 that is lost, the corresponding fragment is at m/z 310.94. This pair of fragments is well detected in the CID mass spectrum of the ion at m/z 472.86. When those fragments (m/z 312.93 and 310.94) loose one more ZnSO4, they can loose 64ZnSO4 or 66ZnSO4, depending on which one was lost first, to give a zinc-free ion that has the same mass in both tandem mass spectra, namely m/z 151.07.
The presence of glycine as a ligand is confirmed in different ways. Firstly by the loss of a fragment of 75 amu that is the mass of the neutral glycine. Moreover, in the n = 2 MS/MS spectra a peak is de-tected at m/z 76 amu that corresponds to the mass of protonated glycine. This peak has the same mass in both MS/MS spectra coming from two different isotopes which confirms that it is a metal-free ion. Finally, the losses of CO, CO2 or H2O (in the fully dehydrated ions) can be attributed to the pres-ence of an amino-acid like glycine as already observed by Hoppilliard et al. (9).

Vacchina et al.: Determination of Zn-glycinate complexes in diluted media
Seite 132 8. BOKU-Symposium Tierernährung 2009
When water is supposed to be present as a ligand, a loss of 18 amu is observed additionally to the one coming from the glycine. The fragment induced by the loss of this water molecule is one of the most abundant which confirms the weak link between the metal and the water molecule.
[ZnC2H8NO7S]+m/z : 253.9298 and 255.9283
[ZnC2H6NO6S]+m/z : 235.9190 and 237.9159
[ZnC2H6NO3]+m/z : 155.9580 and 157.9558
[ZnC2H4NO2]+m/z : 137.9777 and 139.9631
- H2O
- SO3
- H2O
[ZnCH4NO]+m/z : 109.9507 and 111.9491
-CO
- H2SO4
- H2SO4
a
[Zn2C4H11N2O12S2]+m/z : 470.8544 and 472.8671
[ZnC4H11N2O8S]+m/z : 310.9431 and 312.9396
or m/z : 310.9431 and 310.9409
- 64ZnSO4
or – 66ZnSO4
[Zn2C4H9N2O8S]+m/z : 372.8968 and 374.8967
- H2SO4
- NH3
[Zn2C4H6NO8S]+m/z : 355.9034 and 357.8956
b
- 64ZnSO4 or - 66ZnSO4
[C2H6NO2]+m/z : 76.0417 and 76.0407
- CO2- H2O
[Zn2C3H9N2O9S2]+m/z : 408.9204 and 410.9165
- SO3
[Zn2C3H9N2O6S]+m/z : 328.9563 and 330.9522
- CO2
- Gly
[C4H11N2O4]+m/z : 151.0790 and 151.0779 - SO3
[Zn2C4H6NO5]+m/z : 275.8323 and 277.9322
- H2O
[Zn2C3H7N2O5S]+m/z : 310.9431 and 312.9396
Figure 2: reaction diagrams of zinc-glycinate complexes. a) [Zn(Gly)(H2O)(SO4)]. b) [Zn(Gly)(SO4)]2.
Effect of the pH on the complexation of Zn-glycinate complexes
The second set of experiments aimed to check the stability of Zn-glycinate complexes upon pH de-crease. Indeed, during gastro-intestinal process, glycinate complexes can be in a solution at a pH down to 2. It is therefore important to check whereas the complexes are stable at this pH. The ESI MS spectra of the Zn-glycinate were recorded at pH ranging from 2 to 7, by step of 1 pH unit. Two pa-rameters were followed. The first one was the metal-glycinate complexes by checking the characteris-tic isotopic patterns previously determined. The second one was the metal-free glycine. Indeed, if the complexes are broken after acidification, metal-free glycine is going to appear and give a peak at m/z 76 for the protonated ion. The results obtained are presented Figure 3. The graph shows an increase of the metal-free glycine amount in acidic pHs, concomitant with the loss of a part of Zn-glycinate. The dissociation depends on the chain length. The intensity of the n = 1 complex is decreased by a

Vacchina et al.: Determination of Zn-glycinate complexes in diluted media
8. BOKU-Symposium Tierernährung 2009 Seite 133
factor of nearly two between pH 7 and 2 whereas the one of the n = 2 complex remains practically unchanged.
However, it has to be noted that ionisation efficiency in the ESI source differs when pH is changing. As a consequence, differences in intensity cannot directly be linked to differences in concentration. As protonation should be more favourable at acidic pHs, it is assumed that concentration is decreased more than the intensity. Additionally, the mass analyzer used in this study does not have a dynamic range optimum for quantitative analysis. Therefore the Figure 3 should be interpreted as a tendency more than quantitative results. It is nevertheless a fact that the Zn-glycinate complexes were at least partially detected at pH down to 2.
40
60
80
100
120
2 3 4 5 6 780
100
120
140
160
pH
inte
nsity
of c
ompl
exes
nor
mali
sed b
y pH
7, % intensity
of free glycine normalisedby pH 7, %
Figure 3: evolution of Zn-glycinate complexes and metal-free glycine as a function of the pH. (o and
) n = 1. ( and ) n = 2. (♦ and …) metal-free glycine.
Stability of the glycinate complexes in size-exclusion chromatography
As glycinate complexes can easily be degraded on the stationary phase of the size-exclusion column, elution conditions were optimized in terms of pH and ionic strength of the mobile phase in order to maximize the recovery level.
The molecular mass of glycinate complexes is in the range of several hundreds daltons. Therefore the column chosen for this study was Superdex Peptides HR 10/30 with an optimum separation range between 0.1 and 7 kDa. Keeping in mind that, in the whole procedure, analysis by ESI MS(/MS) will be run, ammonium acetate buffer was preferred to other commonly used non volatile buffers such as Tris. The ionic strength was then optimized starting from 10 mM and up to 200 mM that is the upper limit for ICP MS detection, setting the pH at the physiological value (pH = 7.4). The pH of the mobile was then optimized at the optimum ionic strength found. The lower limit selected for mobile phase pH optimisation was 6 and the upper one was 8.5 to stay in the range of physiological pHs. After injection of the glycinate complex in a given condition, the column was flushed with a mobile phase containing 50 mM of EDTA in order to remove the metal that have been dissociated on the stationary phase of the column. The signal was on-line recorded in order to check if at the end of the cleaning procedure the column was really metal-free (metal signal was back at the baseline level). The EDTA was then removed as specified in the “material and method” part and the column stabilized before a new injec-tion.
The chromatograms obtained for all the mobile phases investigated are shown Figures 4. They were voluntarily shifted for clarity of the figure. The Zn-glycinate was nearly entirely degraded on the col-umn with an elution at 10 mM and therefore Zn could only be eluted with EDTA. When increasing the

Vacchina et al.: Determination of Zn-glycinate complexes in diluted media
Seite 134 8. BOKU-Symposium Tierernährung 2009
ionic strength, the Zn-glycinate was less degraded on the stationary phase of the column and the corresponding peak was becoming sharper. However, when the pH was decreased down to 6 or in-creased up to 8.5, a shoulder on the chromatogram was observed suggesting instability of the com-plex. The optimum conditions chosen were therefore 200 mM ammonium acetate at pH 7.4. In those conditions the chromatograms show that most part of the metal elute before the cleaning step. The recovery level evaluated at 90% confirms it.
0
100000
200000
300000
400000
500000
600000
700000
0 20 40 60 80 100
2
x 106
6
4
Time, min
Inte
nsity
, cps
Figure 4: SE HPLC – ICP MS chromatograms of Zn-glycinate complexes. () 10 mM ammonium ace-
tate pH 7.4. () 40 mM ammonium acetate pH 7.4. () 100 mM ammonium acetate pH 7.4. (…) 200 mM ammonium acetate pH 7.4. (…) 200 mM ammonium acetate pH 6. (…) 200 mM ammonium ace-
tate pH 8.5. The oblique line represents the beginning of the cleaning procedure with EDTA.
In order to check whether the detected peaks could be Zn-glycinate or Zn2+, Zn2+ was injected in the same conditions and compared to the chromatogram obtained from glycinate complexes. Zn2+ does not elute in those conditions which confirms that the eluting compound after injection of Zn-glycinate is still a complex.
Discussion Two Zn-glycinate complexes were identified by mass spectrometry which was not expected in a first step. Some longer chain complexes may be present but not detected due to insufficient sensitivity of the ESI MS system. It was notably the case for other metal-glycinate complexes (data not shown). It is also possible that the shorter chain length complexes may be fragments formed in the ESI source from longer chain ones. It was already shown that for some compounds, formation of fragments in the ESI source could not be prevented (14). However, in such cases, those fragments were the most abundant ones in the MS/MS mode. Here, no fragments corresponding to the n = 1 complex were found in the MS/MS spectra of the n = 2 complex. Therefore, it is more likely that the glycinate sup-plements furnished contain of a mixture of complexes with different chain length. Speciation analysis may allow to clarify this issue. This could not be achieved by size-exclusion separation that is not powerful enough. A more powerful mechanism, such as CZE, is expected to bring more information.

Vacchina et al.: Determination of Zn-glycinate complexes in diluted media
8. BOKU-Symposium Tierernährung 2009 Seite 135
The experimental masses measured differ from 18 and 72 amu from the expected ones for n = 1 and 2 respectively from the theoretical ones. It corresponds to 1 or 4 water units respectively. This cannot be prevented whatever the ionization conditions applied. This phenomena was already observed by Seto et al. (10). They have shown that, for a complex Cu(Gly)(H2O)2, the loss of one molecule of wa-ter was easy but the attempts to cleave the two water molecules were unsuccessful. The same phe-nomenon is observed here for Zn. The n = 1 complex was shown to be linked to two water molecules by X-ray diffraction. In solution the most intense peak is observed for a compound with only one mo-lecule of water. The n = 2 complex was even observed fully dehydrated. This dehydratation pheno-menon is unlikely to be due to sample preparation as solid complex is simply dissolved in an aqueous solution. It is more likely due to a rearrangement in the ESI source. However, from the supplementa-tion field point of view, the number of molecules of water bound to the metal is not a key parameter for the efficiency of the glycinate complex as a food supplement. Therefore, finding conditions in mass spectrometry or sample preparation to get molecular ions of the hydrated complexes was not investi-gated furthermore.
Conclusion ESI QqTOF MS(/MS) allows the unambiguous characterization of glycinate complexes providing metal-specific isotopic pattern in the MS mode and characteristic fragments of glycine, water and sulfate in the MS/MS mode. Even if it was depending on the chain length, the complexes were shown to exist after acidification, as none of them was entirely dissociated at pH = 2. They were also shown to be stable upon size-exclusion liquid chromatography which shows potential further speciation analysis of those complexes.
References
1 Spears J.W., 1996, Animal Feed Sci. Technol., 58, 151-163
2 Cao J., Henry P. R., Guo R., Holwerda R. A., Toth J. P., Littell R. C., Miles R. D., Ammerman C. B., 2000, J. Anim. Sci., 78, 2039-2054
3 Spears J. W., Schlegel P., Seal M. C., Lloyd K. E., 2004, Livestock Production Science, 90, 211-217
4 Ettle T., Schlegel P., Roth F. X., 2008, Journal of Animal Physiology and Animal Nutrition, 92, 35-43
5 Hansen S. L., Schlegel P., Legleiter L. R., Lloyd K. E., Spears J. W., 2008, Journal of Animal Science, 86, 173-179
6 Lobinski R., Schaumlöffel D., Szpunar J., Mass Spectrometry Reviews, 2006, 25, 255-289
7 Zamora M., Herrerro S., Losada J., Cuadrado I., Casado C. M., Alonso B., Organometallics, 2007, 26, 2688-2693
8 Bouchonnet S., Hoppilliard Y., Ohanessian G., Journal of Mass Spectrometry, 30, 172-179
9 Hoppilliard Y., Rogalewicz F., Ohanessian G., International Journal of Mass Spectrometry, 2000, 204, 267-280
10 Seto C., Stone A. J., International Journal of Mass Spectrometry, 1999, 192, 289-302
11 Xu Y., Zhang X., Yergey A. L., J. Am. Soc. Mass Spectrom., 1996, 7, 25-29
12 Boudesocque S., Damaj Z., Dupont L., Behr J-B., Guillon E., Journal of Inorganic Biochemistry, 2008, 102, 1514-1522
13 Szpunar J., The Analyst, 2000, 25, 963-988
14 Vacchina V., Chassaigne H., Oven M., Zenk M. H., Lobinski R., The Analyst, 1999, 124, 1425-1430.

Vacchina et al.: Determination of Zn-glycinate complexes in diluted media
Seite 136 8. BOKU-Symposium Tierernährung 2009
Corresponding author
Véronique Vacchina UT2A, Hélioparc Pau-Pyrénées 2 avenue Pierre Angot 64053 Pau Cedex 9, France E-mail: [email protected]

Večerek et al.: Influence of Addition of Organic Manganese on its Digestibility in Sport Horses
8. BOKU-Symposium Tierernährung 2009 Seite 137
Influence of Addition of Organic Manganese on its Digestibility in Sport Horses
Michal Večerek, Petra Jancikova, Ladislav Zeman, Petr Mareš
Mendel University of Agriculture and Forestry Brno
Introduction One of important principle of horse’s nutrition is ensuring optimal contents of minerals in horses diets. Minerals in mixtures must be in sufficient amount and also in optimal ratio. (Dušek et al., 2007) Min-erals are indispendsable for reproduction, grow and for many function in digestion, physiological and biosynthetic processes. Trace minerals has influence on others minerals and theirs utilization. Bioa-lavability of trace minerals are influence of chemical structures of chemical compounds (HEMKEN, 1997). Daily intake of minerals is often higher then animals need and excrete of its can burding envi-ronment (CLOSE, 1998).
Several studies have reported varying results with regard to the digestibility of inorganic and organic trace mineral forms (BAKER, 2005). Minerals (Mn) chelated were used and it was concluded that the chelated minerals were effective in improving the performance of animals when the chelated minerals at the lowest supplementary level was compared to inorganic minerals, especially at the pharmacol-ogical levels (Paik, 2001).
Material and methods Eight clinically healthy Czech warm blood horses of farm Nosislav were in the experiment. Influence of addition of organic manganese (Mn) on its digestibility in sport horses.
Before experimental period the horses were fed by standard food staff and during experimental period of 28 day they were fed by premix with chelates of manganese. There were done three feaces sam-pling at the beginning experiment, 15th day and 28th day. Feeding of horses were twice a day by meadow hay and grain feed. The average hay intake was 10.37 kg of dry matter per horse and day. There were increased of content of manganese in diets. Average of nutrients of feed ration of two days before feaces sampling and bleeding are shown in tab. 1. and 2.
Tab. 1: The content of nutrient in basic diet. Dry mater g/kg 913.3
Crude protein g/kg 147.4 Fat g/kg 25.7
Fibre g/kg 98.7 Nitrogen free extract g/kg 692.7
Ash g/kg 35.6 Indicator g/kg 4.8 Energy MJ/kg 19.7
Mn mg/kg 13.3

Večerek et al.: Influence of Addition of Organic Manganese on its Digestibility in Sport Horses
Seite 138 8. BOKU-Symposium Tierernährung 2009
Tab. 2: The content of nutrient in experimental feeding additive.
Dry mater g/kg 23.0 Crude protein g/kg 3.2
Fat g/kg 0.4 Fibre g/kg 0.6
Nitrogen free extract g/kg 14.1 Ash g/kg 4.8
Indicator g/kg 0.6 Energy MJ/kg 0.3
Mn mg/kg 289.2
Particular specimens (feed, feaces) were dehydrated, mineralized and determinated content of Mn and iron (figure 1 and 2) by atomic absorbing spectrophotometry metod (MILESTONE ETHOS 1).
Results and discussion There were fed two variants of feed ration with different contents of nutrients (tab. 1 and 2.). Aim of our experiment was observed effect of feeding organic form manganese to content of manganese in feaces and digestibility on its. From tab. 1 and 2, there are contents of particular nutrients of feed rations and there are shown increase to content of Mn in diets. It is corresponding with NRC 1989. Daily intake of manganese was 481 mg/animal/day (in experimental mixture) versus 192 mg/animal/day (in basic diet). Degrease of digestibility of organic manganese was 20.97 % versus inorganic Mn. The content of manganese in feaces is represented in tab. 3. Increase of content of Mn in feaces was 29.5 % (figure 2).
Figure 1.: Digestion of manganese (%)

Večerek et al.: Influence of Addition of Organic Manganese on its Digestibility in Sport Horses
8. BOKU-Symposium Tierernährung 2009 Seite 139
Tab. 3.: The content of nutrients in feaces (mg/kg)
1st period 2nd period 3rd period Dry mater g/kg 930.0 931.0 921.9
Crude protein g/kg 99.2 100.1 77.8 Fat g/kg 25.6 19.0 13.8
Fibre g/kg 351.6 423.5 396.0 Nitrogen free extract g/kg 351.0 288.4 342.9
Ash g/kg 102.6 100.1 91.4 Indicator g/kg 52.1 51.1 39.7 Energy MJ/kg 17.7 17.7 17.8
Mn mg/kg 59.9 80.2 85.0
Figure 2.: Content of manganese in faeces (mg/kg)
Conclusion Differences between digestion of organic and inorganic manganese were not significantly in our ex-periment. Tendency of changes of digestion must be verify in other experiments.
References
BAKER, L. A.: Digestibility and retention of inorganic and organic forms of copper and zinc in yearling and mature geldings, Wageningen Academic Publishers, The Netherlands, 2005, ISBN 907699885X, p 11-26,
CLOSE, W. H.: New Developments in the use of Trace Mineral Proteinates to Improve Pig Performance and Reduce Enviromental Impact, European Lecture Tour, 1998, Alltech Brno, p 51-68
Dušek (2007): Chov koní (2 th Ed.). Nakladatelství Brázda, Praha, 404 p., ISBN 80-209-0352-6
HEMKEN, R. W.: Role of Organic Trace Minerals in Animal Nutrition, In: European and African Lecture Tour, Alltech, 1997, p 47 – 52
PAIK I. K.: Application of chelated minerals in animal production, Asian . Australasian J. Animals Sci., 2001, 14, 191-198.

Večerek et al.: Influence of Addition of Organic Manganese on its Digestibility in Sport Horses
Seite 140 8. BOKU-Symposium Tierernährung 2009
Corresponding author
Ing. Michal Večerek Department of Animal Nutrition and Forage Production Mendel University of Agriculture and Forestry Brno Zemědělská 1, 613 00, Brno, Czech Republic E-mail: [email protected]; tel.: + 420 545 133 170

Firmensponsoring – Trouw Nutrition Deutschland GmbH, Gempfinger Straße 15, D-86664 Burgheim
8. BOKU-Symposium Tierernährung 2009 Seite 141

Wetscherek und Wetscherek-Seipelt: Zweck und Durchführung einer Toleranzstudie bei tragenden und säugenden Zuchtsauen am Beispiel einer Phytase
Seite 142 8. BOKU-Symposium Tierernährung 2009
Zweck und Durchführung einer Toleranzstudie bei tragenden und säugenden Zuchtsauen am Beispiel einer Phytase
Wolfgang Wetscherek und Gabriela Wetscherek-Seipelt
Universität für Bodenkultur Wien, Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie
Einleitung Die Verwendung von Zusatzstoffen in der Tierernährung ist in der VO (EG) 1831/2003 rechtlich gere-gelt. Unter Futterzusatzstoffen versteht man Stoffe, Mikroorganismen oder Zubereitungen, die keine Futtermittel-Ausgangserzeugnisse oder Vormischungen sind und bewusst Futtermitteln oder Wasser zugesetzt werden, um insbesondere eine oder mehrere folgender Funktionen zu erfüllen:
• die Beschaffenheit des Futtermittels positiv beeinflussen;
• die Beschaffenheit der tierischen Erzeugnisse positiv beeinflussen;
• die Farbe von Zierfischen und -vögeln positiv beeinflussen;
• den Ernährungsbedarf der Tiere decken;
• die ökologischen Folgen der Tierproduktion positiv beeinflussen;
• die Tierproduktion, die Leistung oder das Wohlbefinden der Tiere, insbesondere durch Einwir-ken auf die Magen- und Darmflora oder die Verdaulichkeit der Futtermittel, positiv beeinflus-sen;
• eine kokzidiostatische oder histomonostatische Wirkung haben oder
• die Kontamination von Futtermitteln mit Mykotoxinen verringern.
Die EU-weit gültigen Regelungen der Zulassung von Futterzusatzstoffen verlangen neben dem Nach-weis der spezifischen Wirksamkeit auch Studien für die Sicherheit von Mensch, Tier, Umwelt und des erzeugten Lebensmittels. Die EFSA hat Durchführungsbestimmungen für die Zulassungsverfahren im Jahr 2009 neu überarbeitet und festgelegt. Dies gilt auch für die Durchführung von Toleranzstudien am Zieltier. Dabei werden neben einer Mindestversuchsdauer auch Anforderungen an das Versuchsde-sign bzw. bestimmte Analysenparameter genau vorgegeben. Neben einer Kontrollgruppe ohne Prüf-substanz sind eine Versuchsgruppe mit der empfohlenen Dosierung und eine Versuchsgruppe mit einer Überdosierung der Prüfsubstanz vorzusehen. Die übliche Überdosierung wird mit dem 10-fachen Wert der Höchstempfehlung der Prüfsubstanz vorgegeben. Bei diesen Untersuchungen sind die Tiere routinemäßig auf visuelle Auffälligkeiten und klinische Effekte zu untersuchen. Weiters sind die zoo-technischen Leistungen, die hämatologische und chemische Blutuntersuchung sowie andere relevante Parameter, welche durch die Prüfsubstanz beeinflusst werden könnten zu prüfen. Ebenso ist, wenn relevant, auch die Beeinflussung der Produktqualität der tierischen Lebensmittel zu testen.
Als Beispiel einer Toleranzstudie soll hier die Prüfung von einem Phytaseenzymes dargestellt werden. Ronozyme® NP (CT) ist ein Enzymprodukt, das 6-Phytase enthält und von Peniophora lycii hergeleitet wird.

Wetscherek und Wetscherek-Seipelt: Zweck und Durchführung einer Toleranzstudie bei tragenden und säugenden Zuchtsauen am Beispiel einer Phytase
8. BOKU-Symposium Tierernährung 2009 Seite 143
Material und Methoden Diese Studie wurde an der LSF Hatzendorf durchgeführt. Während der Trächtigkeit wurden die Sauen im Trächtigkeitsstall in Gruppenhaltung von max. 4 Sauen mit Einzelfuttertrögen gehalten. In der an-schließenden Laktationsperiode waren die Sauen in Einzelaufstallung in den Abferkelboxen unterge-bracht.
Ziel dieser Toleranzstudie war die Darstellung der Auswirkung einer 10-fachen Überdosis von Rono-zyme® NP (CT) der Firma DSM auf Sauen- und Ferkelleistung, sowie Blutparameter der Sauen am Ende des Versuches verglichen mit einer handelsüblichen Dosierung und einer Negativkontrolle.
Die Rationen der 3 Gruppen unterschied sich nur im Zusatz von Ronozyme® NP (CT)- Phytase, die gegen Gerste ausgetauscht wurde, und entsprachen in der Energie- und Nährstoffversorgung des Bedarfs für Schweine (Tabelle 1).
Tabelle 1 Versuchsplan
Merkmal
Gruppe A (negative Kontrolle)
Gruppe B Gruppe C
Phytasezusatz, U/kg - 3.000 30.000 Sauenanzahl, n 10 11 10
Tabelle 2 Zusammensetzung der Futtermischungen für die tragenden Sauen
Futtermittel, % Gruppe A Gruppe B Gruppe C Gerste 35.76 35.74 35.55 Mais 15.00 15.00 15.00 Weizenkleie 12.50 12.50 12.50 Weizen 12.00 12.00 12.00 Sonnenblumenextraktionsschrot 9.50 9.50 9.50 Trockenschnitzel 4.00 4.00 4.00 Vinasse 2.60 2.60 2.60 Sojaextraktionsschrot HP 2.00 2.00 2.00 Glycerin 1.93 1.93 1.93 Futterkalk 1.13 1.13 1.13 Melasse 1.00 1.00 1.00 Vitamin- und Spurenelementprämix 0.60 0.60 0.60 Monocalciumphosphat 0.57 0.57 0.57 Natriumbicarbonat 0.47 0.47 0.47 Fischöl 0.40 0.40 0.40 Lignocellulose 0.40 0.40 0.40 L-Lysin 98 0.14 0.14 0.14 Ronozyme® NP (CT) - 0.02 0.21
Den tragenden Sauen wurde das Futter abhängig von ihrem Lebendgewicht rationiert 2-mal täglich vorgelegt. Den säugenden Sauen wurde das Futter in Abhängigkeit von der Ferkelzahl ebenfalls 2-mal täglich angeboten.
Das Körpergewicht der Zuchtsauen wurde zu Versuchsbeginn, beim Wechsel in die Abferkelbox und beim Absetztermin der Ferkel erhoben. Die Futteraufnahme jeder Sau wurde täglich festgehalten. Die

Wetscherek und Wetscherek-Seipelt: Zweck und Durchführung einer Toleranzstudie bei tragenden und säugenden Zuchtsauen am Beispiel einer Phytase
Seite 144 8. BOKU-Symposium Tierernährung 2009
Ferkelzahl, lebend und tot geboren, das individuelle Gewicht bei der Geburt und beim Absetzen sowie die Ferkelsterblichkeit wurden aufgezeichnet. Am Tag des Absetzens wurde von jeder Sau eine Blut-probe genommen.
Table 3 Zusammensetzung der Futtermischungen für die säugenden Sauen
Futtermittel, % Gruppe A Gruppe B Gruppe C Gerste 28.97 28.95 28.76 Weizen 22.00 22.00 22.00 Mais 20.00 20.00 20.00 Sojaextraktionsschrot HP 14.00 14.00 14.00 Sonnenblumenextraktionsschrot 6.00 6.00 6.00 Weizenkleie 2.00 2.00 2.00 Trockenschnitzel 1.50 1.50 1.50 Futterkalk 1.50 1.50 1.50 Glycerin 1.00 1.00 1.00 Vitamin- und Spurenelementprämix 0.96 0.96 0.96 Monocalciumphosphat 0.90 0.90 0.90 Viehsalz 0.40 0.40 0.40 Vormischung - Org. Säuren 0.30 0.30 0.30 Fischöl 0.25 0.25 0.25 L-Lysin 98 0.22 0.22 0.22 Ronozyme® NP (CT) - 0.02 0.21
Table 4 Berechneter Nährstoffgehalt der Futtermischungen für tragende bzw. säugende Sauen
Nährstoffgehalt tragende Sauen Säugende Sauen Trockenmasse, % 88.8 88.6 Rohprotein, % 13.7 17.0 Rohfett, % 2.7 2.4 Rohfaser, % 6.9 4.1 Rohasche, % 5.8 6.3 Stärke, % 37.1 40.9 Zucker, % 3.6 3.4 Umsetzbare Energie, MJ/kg 11.90 12.89 Calcium, % 0.68 0.85 Phosphor, % 0.63 0.65 Natrium, % 0.23 0.22 Magnesium, % 0.22 0.22 Kalium, % 0.74 0.82 Eisen, mg/kg 237 175 Kupfer, mg/kg 31 30 Zink, mg/kg 130 113 Mangan, mg/kg 95 84 Selen, mg/kg 0.63 0.76
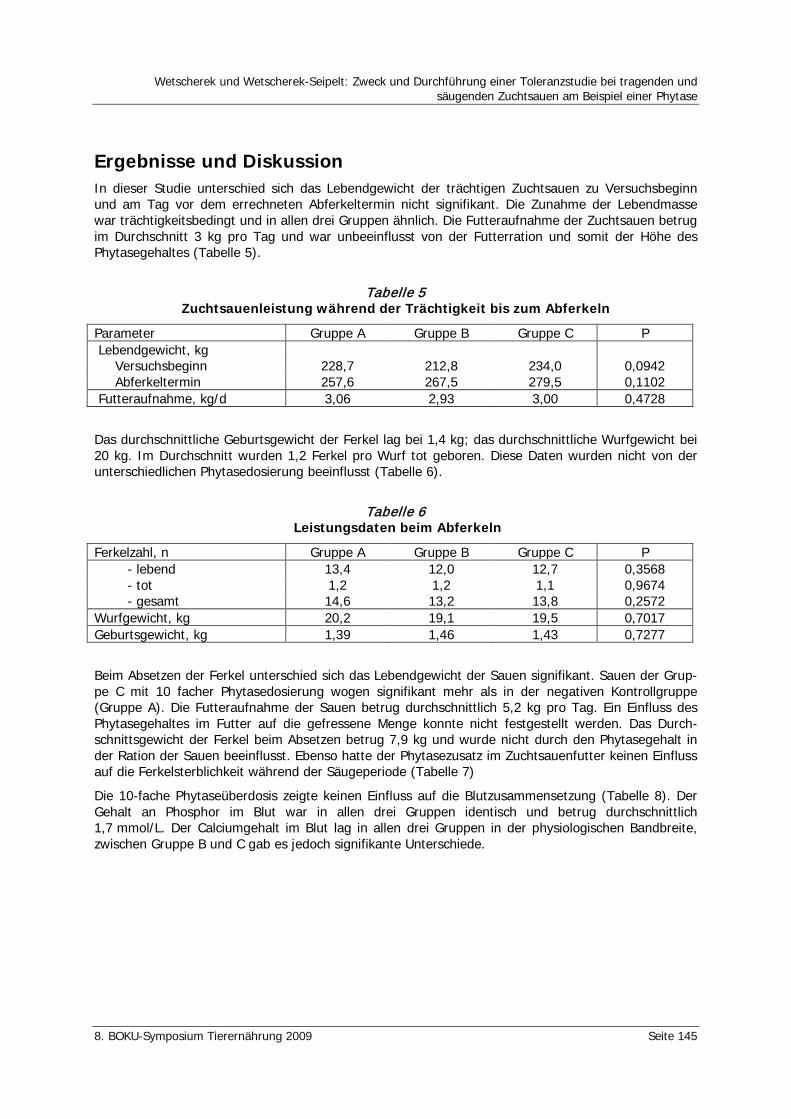
Wetscherek und Wetscherek-Seipelt: Zweck und Durchführung einer Toleranzstudie bei tragenden und säugenden Zuchtsauen am Beispiel einer Phytase
8. BOKU-Symposium Tierernährung 2009 Seite 145
Ergebnisse und Diskussion In dieser Studie unterschied sich das Lebendgewicht der trächtigen Zuchtsauen zu Versuchsbeginn und am Tag vor dem errechneten Abferkeltermin nicht signifikant. Die Zunahme der Lebendmasse war trächtigkeitsbedingt und in allen drei Gruppen ähnlich. Die Futteraufnahme der Zuchtsauen betrug im Durchschnitt 3 kg pro Tag und war unbeeinflusst von der Futterration und somit der Höhe des Phytasegehaltes (Tabelle 5).
Tabelle 5 Zuchtsauenleistung während der Trächtigkeit bis zum Abferkeln
Parameter Gruppe A Gruppe B Gruppe C P Lebendgewicht, kg Versuchsbeginn 228,7 212,8 234,0 0,0942 Abferkeltermin 257,6 267,5 279,5 0,1102 Futteraufnahme, kg/d 3,06 2,93 3,00 0,4728
Das durchschnittliche Geburtsgewicht der Ferkel lag bei 1,4 kg; das durchschnittliche Wurfgewicht bei 20 kg. Im Durchschnitt wurden 1,2 Ferkel pro Wurf tot geboren. Diese Daten wurden nicht von der unterschiedlichen Phytasedosierung beeinflusst (Tabelle 6).
Tabelle 6 Leistungsdaten beim Abferkeln
Ferkelzahl, n Gruppe A Gruppe B Gruppe C P - lebend 13,4 12,0 12,7 0,3568 - tot 1,2 1,2 1,1 0,9674 - gesamt 14,6 13,2 13,8 0,2572 Wurfgewicht, kg 20,2 19,1 19,5 0,7017 Geburtsgewicht, kg 1,39 1,46 1,43 0,7277
Beim Absetzen der Ferkel unterschied sich das Lebendgewicht der Sauen signifikant. Sauen der Grup-pe C mit 10 facher Phytasedosierung wogen signifikant mehr als in der negativen Kontrollgruppe (Gruppe A). Die Futteraufnahme der Sauen betrug durchschnittlich 5,2 kg pro Tag. Ein Einfluss des Phytasegehaltes im Futter auf die gefressene Menge konnte nicht festgestellt werden. Das Durch-schnittsgewicht der Ferkel beim Absetzen betrug 7,9 kg und wurde nicht durch den Phytasegehalt in der Ration der Sauen beeinflusst. Ebenso hatte der Phytasezusatz im Zuchtsauenfutter keinen Einfluss auf die Ferkelsterblichkeit während der Säugeperiode (Tabelle 7)
Die 10-fache Phytaseüberdosis zeigte keinen Einfluss auf die Blutzusammensetzung (Tabelle 8). Der Gehalt an Phosphor im Blut war in allen drei Gruppen identisch und betrug durchschnittlich 1,7 mmol/L. Der Calciumgehalt im Blut lag in allen drei Gruppen in der physiologischen Bandbreite, zwischen Gruppe B und C gab es jedoch signifikante Unterschiede.
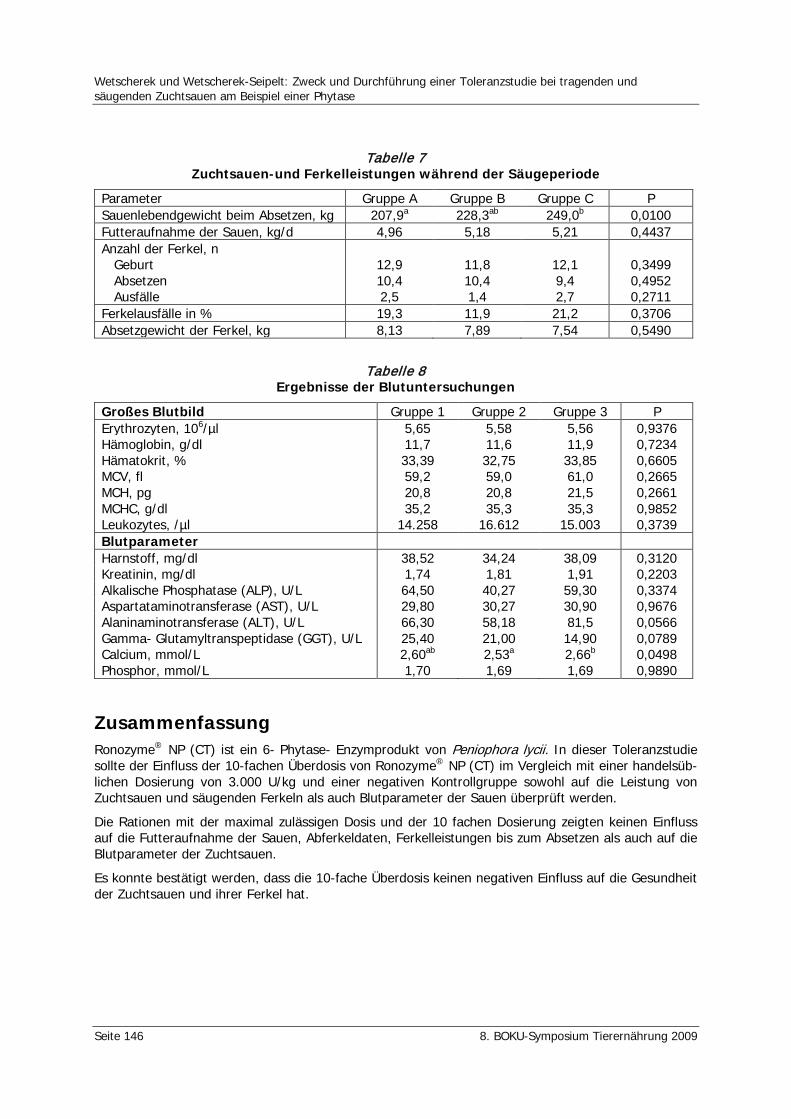
Wetscherek und Wetscherek-Seipelt: Zweck und Durchführung einer Toleranzstudie bei tragenden und säugenden Zuchtsauen am Beispiel einer Phytase
Seite 146 8. BOKU-Symposium Tierernährung 2009
Tabelle 7 Zuchtsauen-und Ferkelleistungen während der Säugeperiode
Parameter Gruppe A Gruppe B Gruppe C P Sauenlebendgewicht beim Absetzen, kg 207,9a 228,3ab 249,0b 0,0100 Futteraufnahme der Sauen, kg/d 4,96 5,18 5,21 0,4437 Anzahl der Ferkel, n Geburt 12,9 11,8 12,1 0,3499 Absetzen 10,4 10,4 9,4 0,4952 Ausfälle 2,5 1,4 2,7 0,2711 Ferkelausfälle in % 19,3 11,9 21,2 0,3706 Absetzgewicht der Ferkel, kg 8,13 7,89 7,54 0,5490
Tabelle 8 Ergebnisse der Blutuntersuchungen
Großes Blutbild Gruppe 1 Gruppe 2 Gruppe 3 P Erythrozyten, 106/µl 5,65 5,58 5,56 0,9376 Hämoglobin, g/dl 11,7 11,6 11,9 0,7234 Hämatokrit, % 33,39 32,75 33,85 0,6605 MCV, fl 59,2 59,0 61,0 0,2665 MCH, pg 20,8 20,8 21,5 0,2661 MCHC, g/dl 35,2 35,3 35,3 0,9852 Leukozytes, /µl 14.258 16.612 15.003 0,3739 Blutparameter Harnstoff, mg/dl 38,52 34,24 38,09 0,3120 Kreatinin, mg/dl 1,74 1,81 1,91 0,2203 Alkalische Phosphatase (ALP), U/L 64,50 40,27 59,30 0,3374 Aspartataminotransferase (AST), U/L 29,80 30,27 30,90 0,9676 Alaninaminotransferase (ALT), U/L 66,30 58,18 81,5 0,0566 Gamma- Glutamyltranspeptidase (GGT), U/L 25,40 21,00 14,90 0,0789 Calcium, mmol/L 2,60ab 2,53a 2,66b 0,0498 Phosphor, mmol/L 1,70 1,69 1,69 0,9890
Zusammenfassung Ronozyme® NP (CT) ist ein 6- Phytase- Enzymprodukt von Peniophora lycii. In dieser Toleranzstudie sollte der Einfluss der 10-fachen Überdosis von Ronozyme® NP (CT) im Vergleich mit einer handelsüb-lichen Dosierung von 3.000 U/kg und einer negativen Kontrollgruppe sowohl auf die Leistung von Zuchtsauen und säugenden Ferkeln als auch Blutparameter der Sauen überprüft werden.
Die Rationen mit der maximal zulässigen Dosis und der 10 fachen Dosierung zeigten keinen Einfluss auf die Futteraufnahme der Sauen, Abferkeldaten, Ferkelleistungen bis zum Absetzen als auch auf die Blutparameter der Zuchtsauen.
Es konnte bestätigt werden, dass die 10-fache Überdosis keinen negativen Einfluss auf die Gesundheit der Zuchtsauen und ihrer Ferkel hat.

Wetscherek und Wetscherek-Seipelt: Zweck und Durchführung einer Toleranzstudie bei tragenden und säugenden Zuchtsauen am Beispiel einer Phytase
8. BOKU-Symposium Tierernährung 2009 Seite 147
Summary Ronozyme® NP (CT) is an enzyme product containing 6-phytase derived from Peniophora lycii. The aim of the study was to evaluate the influence of the 10 fold overdose of Ronozyme® NP (CT), when compared to the normal dosage (3000 U/kg) and a negative control, on performance of sows and suckling piglets as well as on haematological and blood biochemical parameters of sows.
The dietary administration of Ronozyme® NP (CT) phytase at the maximum recommended dose and the 10 fold overdose did not show any influence on feed intake of sows, litter performance, growth parameters of piglets before weaning, haematological parameters and blood biochemistry of sows.
It has been confirmed that the 10 fold overdose of Ronozyme® NP phytase has no negative effects on health status of sows and performance of sows and their piglets as well.
Autorenanschrift
Ao. Univ.-Prof. Dr. Wolfgang Wetscherek Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Department für Lebensmittelwissenschaften und –technologie Universität für Bodenkultur Wien Gregor Mendel-Straße 33, 1180 Wien E-Mail: [email protected]

Bravo et al.: Evaluation of a mixture of carvacrol, cinnamaldehyde and capsicum oleoresin for improving growth performance and metabolizable energy for broiler chicks fed corn-soybean meal
Seite 148 8. BOKU-Symposium Tierernährung 2009
Evaluation of a mixture of carvacrol, cinnamaldehyde and capsi-cum oleoresin for improving growth performance and metabo-lizable energy for broiler chicks fed corn-soybean meal
D. Bravo1, P. Utterback2, C.M. Parsons2, A. Vikari1 1 Pancosma, Geneva, Switzerland 2 University of Illinois, Illinois, United States
Introduction Dietary energy represents up to 60% of the feed cost for broilers which eat to satisfy energy require-ment. The factorial method partitions energy requirement into energy required for maintenance, growth and production (Hurwitz et al., 1978). As the requirement of energy for maintenance is com-mon to all the feeding conditions, an imprecision on its evaluation could have an important impact. As a growing animal will never exhibit a zero energy balance, the energy requirement for maintenance for a broiler is the needs for all body functions and moderate activity (National Research Council, 1998). Among the body functions, the management of the digestive sphere, means digestive ecosys-tem and digestive mucosa, could represent a non negligible part, especially in young growing animals fed without antibiotic growth promoters. For example, Hughes (2003) demonstrated that up to 33% of the variation in metabolizable energy (ME) of broilers were associated with morphology of the small intestinal mucosa as with crypt depth. Moreover, the dietary factors that increase the activity of gut microflora depress energy utilization (Choct et al., 1996). The question could be whether the inverse assumption is correct meaning that the factors decreasing the activity of gut or improving the mor-phology of the small intestinal mucosa could be associated with improved energy utilization. Currently, there is a increasing interest towards feed additive based on plant extracts in broilers with evidences of effect on gut ecosystem and morphology (Mitsch et al., 2004, Cross et al., 2007). Our objective was to evaluate the effect of a blend of plant extracts on the ME of broilers fed a corn – soybean meal or corn – corn distiller – soybean meal diets (Experiment 1) and to check this value with a productive trial (Experiment 2).
Material and Methods Experiment 1: The objective of this trial was to evaluate if a mixture of plant extracts would have a positive effect on ME in broiler. Housing, handling, feeding, and killing procedures were in accord with policies of the University of Illinois Committee on Laboratory Animal Care. Birds, Housing, and Diets. Commercial broiler chicks (450) were obtained from a commercial hatchery. The chicks were housed in batteries with raised wire floors. They were weighed and sorted by weight and the 360 most uni-form chicks were assigned to 72 pens of 5 chicks each. 12 replicate groups of 5 chicks were assigned to each of the 4 treatments. Two factors were organized in a complete factorial design. Factor 1 was diet type (corn-soybean meal without versus with corn distillers dried grains with solubles, DDGS), factor 2 was XTRACT™ 6930 =XT (0 versus 100 ppm of XT). The compositions of the diets are shown in the Table 1 (main ingredients). Diets were fed from 0 to 21 days, body weight, feed intake, and gain to feed being measured at 7 and 21 days and excreta collected from each pen at 20-21 days for ME determination. The experimental diets were fed from 0 to 21 days of age. Body weights, feed intake, and gain to feed ratio were measured at 7 and 21 days. In addition, excreta were collected

2 et al.: Evaluation of a mixture of carvacrol, cinnamaldehyde and capsicum oleoresin for improving growth performance and metabolizable energy for broiler chicks fed corn-soybean meal
8. BOKU-Symposium Tierernährung 2009 Seite 149
from each pen at 20-21 days. The excreta were freeze-dried and ground. For ME determination, all diets and excreta samples were analyzed for gross energy, N, and acid insoluble ash.
Experiment 2: The objective of this trial was to evaluate whether the ME value obtained in the Expe-riment 1 was applicable in a feed. Put another way, the question was if the productive performances were similar between on the one hand, a diet with XT and with a reduction of ME of the extent de-termined in the Experiment 1 and on the other hand, a diet without XT and without reduction of ME content. 1 day old male chicks were kept in floor pen until day 9 post hatching. A total of 192 chick-ens were selected from a population of 250, discarding birds with extreme weights. Following 16 h of feed deprivation, chicks were individually weighed, and randomly distributed among 4 treatments groups with 48 replicates per treatment and 1 bird per replicate. Four replicate groups of 48 chicks were used. Chicks were placed in battery cages with wire floors for 25 days. Two factors were orga-nized in a complete factorial design with factor 1 (called Diet) was the dietary ME (3000 kcal/kg ver-sus 3000 kcal – ME, XT) and factor 2 (called XT) was the inclusion of 100 ppm of XT. Diet 2a was the basal corn-soybean meal diet (22% CP, 3050 kcal/kg). Diet 2b was the basal diet 2a plus 100 ppm of XT. Diet 2c was a basal corn-soybean meal diet with 3000 kcal – ME, XT. Diet 2d was the diet 2c plus 100 ppm of XT. The compositions of the diets are shown in the Table 1 (main ingredients). All diets were prepared with the same batch of ingredients, and they were formulated to meet or exceed the recommended nutrient requirements (National Research Council, 1994).
Table 1: Composition of the diets used in Experiment 1 and 2 (main ingredients)
Statistical analysis: Statistical analyses were conducted using SAS (Version 8.2, SAS Institute, Inc., Cary, NC). The results of daily gain, feed intake, gain to feed ratio (for Experiments 1 and 2) and ME (only for Experiment 1) were analyzed using the GLM procedure of SAS and the following model: Yijk= μ + αi + βj + (αβ)ij + εijk where Y = dependent variable, μ = overall mean of the population, α = mean effect of “XT” (i = 1, 2), β = mean effect of “Diet” (j = 1, 2), (αβ) = interaction between “XT” and “Diet”, and ε = residual error of the model assumed to be independent and normally. Significant differences between groups were determined by the Tukey’s test. The P-values were presented for all the effect, significance was declared at P < 0.05, trends were discussed at P < 0.15
Experiment 1 Experiment 2 Feedstuffs 1a 1b 1c 1d 2a 2b 2c 2d
Sorghum 61.17 61.16 62.37 62.36 Corn 63.58 63.58 57.76 57.76 Soybean Meal 31.50 31.50 27.00 27.00 Soybean Meal (46% CP) 32.5 32.5 32.3 32.3 DDGS 0 0 10.00 10.00 Soybean Oil 0.81 0.81 1.14 1.14 2.5 2.5 1.5 1.5 Dicalcium Phosphate 2.05 2.05 1.94 1.94 1.36 1.36 1.36 1.36 Limestone 1.19 1.19 1.23 1.23 1.25 1.25 1.25 1.25 Vitamin premix 0.20 0.20 0.20 0.20 0.025 0.025 0.025 0.025 Mineral Mix 0.15 0.15 0.15 0.15 0.15 0.15 0.15 0.15 XTRACT 6930 0 0.01 0 0.01 0 0.01 0 0.01 Calculated composition AMEn, kcal/kg 3002 3002 2947 2947 Crude fat, % 4.57 4.57 3.61 3.61 CP, % 20.5 20.5 20.5 20.5

2 et al.: Evaluation of a mixture of carvacrol, cinnamaldehyde and capsicum oleoresin for improving growth performance and metabolizable energy for broiler chicks fed corn-soybean meal
Seite 150 8. BOKU-Symposium Tierernährung 2009
Results Experiment 1: The results of the Experiment 1 are summarized in the Table 2.
From day 1 to day 8, neither experimental treatment influenced the feed intake (P = 0.832 and P = 0.827, respectively for XT and DDGS). On the contrary, XT and DDGS increased daily gain of respec-tively +6.4% (P = 0.055) and +6.7% (P = 0.047). Gain to feed was improved by XT (+6.6% with P = 0.081) and unaffected by DDGS (P = 0.181). From day 9 to day 22, neither experimental treatment influenced feed intake (P = 0.450 and P = 0.536, respectively for XT and DDGS) and daily gain (P = 0.157 and P = 0.293, respectively for XT and DDGS). XT did not affect gain to feed (P = 0.194) whe-reas DDGS improved it (+4.8%, P = 0.048). For the whole period (from day 1 to day 22), the treat-ments did not influence feed intake (P = 0.566 and P = 0.647, respectively for XT and DDGS) and daily gain (P = 0.117 and P = 0.209, respectively for XT and DDGS). The effect of XT on daily gain (P = 0.117) was comparable whatever the diet was with +4.8% improvement without DDGS and +4.9% improvement with DDGS. Gain to feed was increased by XT (+3.7%, P = 0.089) and by DDGS (+5.0%, P = 0.024). The interaction between the 2 treatments was not significant (P = 0.495), how-ever, the XT effect was more intense without DDGS (+5.4%) compared to the situation with DDGS (+2.0%). Finally, the ME was increased by DDGS (+155 kcal/kg, P < 0.001) and by XT (+41 kcal/kg, P = 0.003). The magnitude of the response of XT was higher for the diet without DDGS (+68 kcal/kg vs. +34 kcal/kg), even if the interaction between both treatments was not significant (P = 0.314). The results of the Experiment 1 indicated that the XT addition in a corn-soybean meal diet increased of 68 kcal/kg the ME content of the diet.
Table 2: Growth performance for chicks fed a diet containing either 0 vs. 10% DDGS1, or 0 vs. 100 ppm of XT2, Experiment 13
Experiment 2: The results of the Trial 2 are summarized in the Table 3. Neither any of the treatment nor their interaction influenced feed intake (P > 0.10). There was no interaction between treatments on daily gain (P = 0.808) and on gain to feed (P = 0.520). For this trial to be acceptable, the zootech-nical performance of the broilers should decrease when the ME content of the diet was decreased from 3000 to 2950 kcal/kg. This was observed with decreases of daily gain (-2.4%, P = 0.021) and of gain to feed ratio (-2.1%, P = 0.009). Inversely, the addition of XT increased the daily gain (+2.4%, P = 0.024) and gain to feed ratio (+2.3%, P = 0.006). The magnitude of the effect of XT was very close
Treatments Statistics Diet = 0% DDGS Diet = 10% DDGS Source of variation (P-value) Item XT = 0 ppm XT = 100 ppm XT = 0 ppm XT = 100 ppm SEM Diet XT Diet ×
XT day 1 to 8 post-hatching Feed intake (g/chick) 179.1 173.2 176.3 179.1 7.2 0.827 0.832 0.546 Daily gain (g/chick) 96.95 102.71 102.96 110.09 3.3 0.047 0.056 0.835 Gain to Feed (g/g) 554.9 595.7 586.6 621.6 21.2 0.181 0.081 0.893 day 9 to 22 post-hatching Feed intake (g/chick) 960.5 965.2 935.1 963.5 21.7 0.293 0.157 0.985 Daily gain (g/chick) 571.9 597.9 591.1 617.7 18.3 0.536 0.450 0.588 Gain : Feed (g/g) 593.2 621.7 632.1 641.7 14.5 0.048 0.194 0.515 day 1 to 22 post-hatching Feed intake (g/chick) 1139.6 1138.4 1111.3 1142.6 20.6 0.647 0.566 0.536 Daily gain (g/chick) 668.7 700.6 693.9 727.9 14.6 0.209 0.117 0.959 Gain to Feed (g/g) 585.2 616.6 624.3 637.9 12.8 0.024 0.089 0.495 AMEn (kcal/kg) 3099 3167 3271 3305 0.016 0.003 < 0.001 0.314 1 DDGS = distillers dried grains with solubles. 2 XT = Xtract 6930 (Pancosma S.A., Switzerland), mixture of 5% carvacrol, 3% cinnamaldehyde and 2% capsicum oleoresin. 3 Means represent 12 pens of 5 chicks per treatment; average initial weight was 37.3 g.

2 et al.: Evaluation of a mixture of carvacrol, cinnamaldehyde and capsicum oleoresin for improving growth performance and metabolizable energy for broiler chicks fed corn-soybean meal
8. BOKU-Symposium Tierernährung 2009 Seite 151
to the one of ME with daily gain changes of -2.4% for ME and +2.4% for XT and gain to feed changes of –2.1% for ME and +2.3% for XT.mean value was associated with a SEM of 0.011. The 90% confi-dence interval is 49.8 kcal/kg to 86.1 kcal/kg. The value of ME(XT) used in the Experiment 2 is 50 kcal/kg.
Table 3. Growth performance for chicks fed a diet containing either 2950 kcal/kg vs. 3000 kcal/kg of AMEn, or 0 vs. 100 ppm of XT1, Experiment 22
Discussion The present results indicate that the addition of 100 ppm of a mixture of 5% carvacrol, 3% cinnamal-dehyde and 2% capsicum oleoresin in a diet improved weight gain (+2.4% in Experiment 2, +4.7% in Experiment 1) and gain to feed ratio (+5.3% in Experiment 1, +2,3%, Experiment 2). The results figures are consistent with published zootechnical results with the same product on weight gain (+2.69% Jamroz et al, 2003, +3.5% for Hernandez et al., 2004) and of gain to feed ratio in a corn soybean meal (+3.9% Jamroz et al., 2006, +4.2% Jamroz et al., 2005, +4.5% Jamroz et al., 2003). Experiment 1 demonstrated that when added to a diet, this product increased the ME content by a mean effect of 51 kcal (mean of 34 and 69 kcal) which correspond to +1.6%. On the one hand this ME effect of XT could be due to an increase of dietary energy extraction based on improvement of the digestion itself or improvement of the absorption of the digested nutrients or a combination of both. Improved digestibility in broilers fed corn – soybean meal diet have been already published with XT. Jamroz et al (2003) measured that XT improved ileal apparent digestibility coefficients for DM (+3.9%), crude fat (+15.0%), crude ash (+49.2%) and N (+2.7%). Hernandez et al. (2004) reported improved apparent ileal DM digestibility in starter diet (+9.2%), in finisher diet (+6.9%) and im-provement of the total CP digestibility in finisher diet (+17.4%) and of ether extracts in starter diet (+2.3%). The reason of the enhanced digestibility with XT could be due to increased digestive secre-tions as reported with the effect of capsicum oleoresin on pancreatic and intestinal enzyme activity in rats (Platel et Scrinavan, 2001) and on bile flow and bile acid secretion (Ganesh et al., 1984). This is confirmed by in vivo enhanced digestive enzymes activity demonstrated with broilers fed corn soybean meal diet supplemented with XT (Jamroz et al., 2005). Improvement of the digestibility could also be partly mediated by a higher absorption capacity of the digestive mucosa. Individual components of XT demonstrated protective function in the gastro intestinal mucosa (Gonzalez et al., 1998, for capsaicin for example). Carvacrol and cinnamaldehyde have been shown to increase the activity of the antioxi-dant enzymes of the cells of the mucosa layer (Dhuley et al., 1999) which is know to be a protective system for the tissue. This would be the basis for a mucosa protective effect observed with such feed additive and illustrated by Jamroz et al., (2006). On the other hand, the enhanced ME with XT could also be due to a reduction of the part of energy used by the animal to maintain the digestive tract ecosystem and integrity. This could be mediated by the protective effect of cinnamaldehyde or carvac-rol on the mucosa which would have in turn decreased the necessity of repair. The same blend modu-lated the digestive microbial ecosystem in cocci-vaccinated broilers, avoiding drastic shifts after a mixed coccidian infection. (Oviedo-Rondon et al., 2006).
Treatments Statistics Diet = 2950 kcal/kg Diet = 3000 kcal/kg Source of variation P-value)
Item
XT = 0 ppm XT = 100 ppm XT = 0 ppm XT = 100 ppm SEM
Diet XT Diet × XT Feed intake (g/chick) 114.6 114.8 115.1 115 0.679 0.753 0.914 0.880 Daily gain (g/chick) 63.12 64.51 64.54 66.27 1.054 0.021 0.024 0.808 Gain to Feed (g/g) 552.1 561.8 561.1 576.6 4.579 0.009 0.006 0.520 1 XT = Xtract 6930 (Pancosma S.A., Switzerland), mixture of 5% carvacrol, 3% cinnamaldehyde and 2% capsicum oleoresin. 2 Means represent 48 pens of 1 chicks per treatment; average initial weight was 234.9 g (SE = 0.58).

2 et al.: Evaluation of a mixture of carvacrol, cinnamaldehyde and capsicum oleoresin for improving growth performance and metabolizable energy for broiler chicks fed corn-soybean meal
Seite 152 8. BOKU-Symposium Tierernährung 2009
Conclusion As a summary, the present trial indicated that 100 ppm of a mixture of carvacrol, cinnamaldehyde and capsicum oleoresin increased the performance of 21 days old broiler chickens. Simultaneously, the dietary ME was increased. In a second trial, the size of the increase of ME was tested in a productive trial and confirmed.
References
Choct, M., Hughes, R.J., Wang, J., Bedford, M.R., Morgan, A.J., and Annison, G., 1996. Increased small intestinal fermentation is partly responsible for the anti-nutritive activity of non-starch polysaccharides in chickens. Br Poult. Sci. 37: 609-621.
Cross D.E., McDevitt R.M., Hillman K., and T. Acamovic, 2007. The effect of herbs and their associated essential oils on performance, dietary digestibility and gut microflora in chickens from 7 to 28 days of age. Br. Poult. Sci. 48: 496-506.
Dhuley N. 1999. Anti-oxidant effects of cinnamon (Cinnamomum verum) bark and greater cardamon (Amomum subulatum) seeds in rats fed high fat diet. Indian J. Exp. Biol. 37:238-242.
Ganesh Bhat, B., Srinivasan, M.R., Chandrasekhara, N., 1984. Influence of curcumin and capsaicin on the composition and secretion of bile in rats. J. Food Sci. Technol. 21: 225-227.
Gonzalez, R., Dunkel, R., Koletzko, B., Schusdziarra, V., and H.D. Allescher. 1998. Effect of Capsaicin-Containing Red Pepper Sauce Suspension on Upper Gastrointestinal Motility in Healthy Volunteers. Dig. Dis. Sci. 43: 1165-1171.
Hernández, F., Madrid, J., García, V., Orengo, J., and M. D. Megías. 2004. Influence of Two Plant Extracts on Broilers Performance, Digestibility, and Digestive Organ Size. Poult. Sci. 83:169–174.
Hughes R.J. 2003. Energy Metabolism of Chickens Physiological Limitations. Rural Industries Research and Development Corporation. Roseworthy Campus, University of Adelaide.
Hurwitz, S., Sklan, D., and I. Bartov, 1978. New formal approaches to determination of energy and amino acid requirements of chicks. Poult. Sci. 57:197–205.
Jamroz, D., Wertelecki, T., Houszka, M., and C. Kamel. 2006. Influence of diet type on the inclusion of plant origin active subtances on morphological and histochemical characteristics of the stomach and jejunum walls in chickens. J Anim Physiol Anim Nutr (Berl) 90: 255.268.
Jamroz, D., Wiliczkiewicz, A., Wertelecki, T., Orda, J., and Skorupinska, J. 2005. Use of active substances of plant origin in chicken diets based on maize and locally grown cereals. Br Poult. Sci. 46: 485-493.
Jamroz, D., Orda, J., Kamel, C.m Wiliczkiewicz, A., Wertelecki, T., Skorupinska, J. 2003. The influence of phytogenic extracts on performance, nutrient digestibility, carcass characteristics, and gut microbial status in broiler chickens. J. Anim. Feed Sci. 12, 583-596.
Mitsch, P., K. Zitterl-Eglseer, B. Kohler, C. Gabler, R. Losa, and I. Zimpernik. 2004. The Effect of Two Different Blends of Essential Oil Components on the Proliferation of Clostridium perfringens in the Intestines of Broiler Chickens. Poultr. Sci. 83: 669–675.
National Research Council, 1994. Nutrient Requirements of Poultry. 9th rev. ed. National Academy Press, Washington, DC.
Oviedo-Rondon, E. O., Hume, M. E., Hernandez, C., and S. Clemente-Hernandez. 2006. Intestinal Microbial Ecology of Broilers Vaccinated and Challenged with Mixed Eimeria Species, and Supplemented with Essential Oil Blends. Poultr. Sci. 85: 854–860.
Platel, K., Srinivasan, K. 2001. A study of the digestive stimuland action of select spices in experimental rats. J. food. Sci. Technol., 38: 358-361.
Corresponding author
Dr. Armin Vikari, PANCOSMA S.A., Voie-des-Traz 6 1218 Le Grand-Saconnex Geneva-Switzerland E-mail: [email protected]
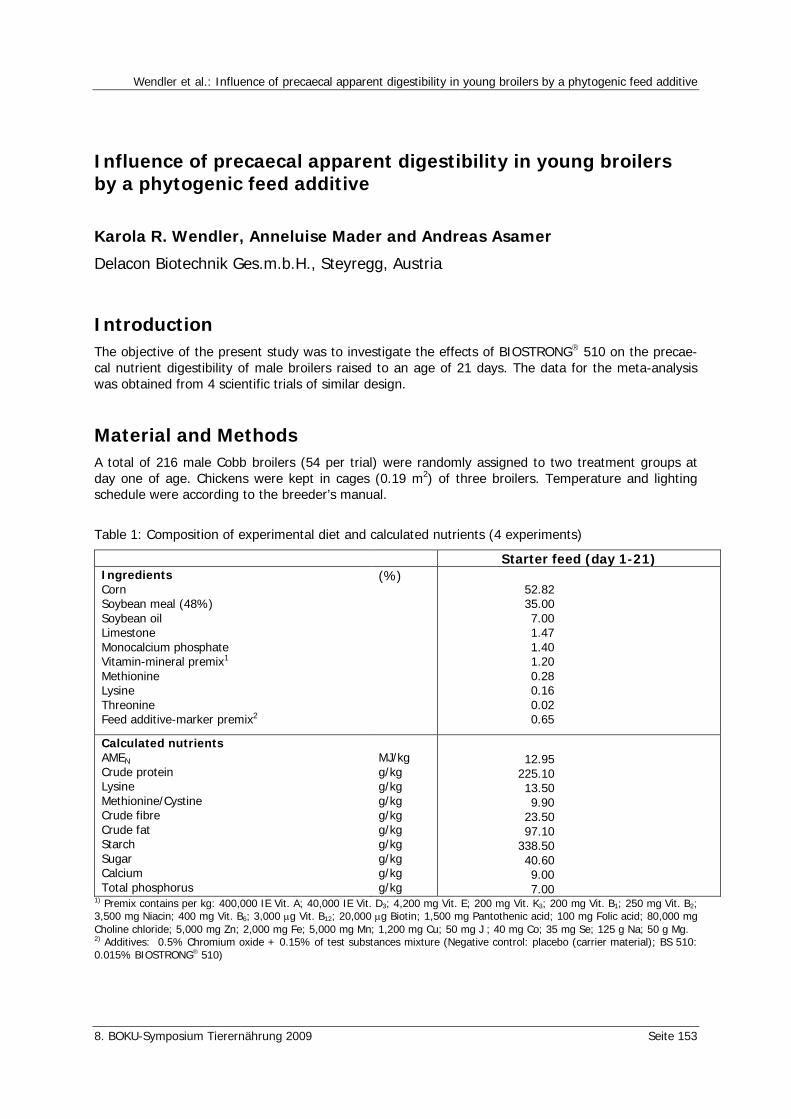
Wendler et al.: Influence of precaecal apparent digestibility in young broilers by a phytogenic feed additive
8. BOKU-Symposium Tierernährung 2009 Seite 153
Influence of precaecal apparent digestibility in young broilers by a phytogenic feed additive
Karola R. Wendler, Anneluise Mader and Andreas Asamer
Delacon Biotechnik Ges.m.b.H., Steyregg, Austria
Introduction The objective of the present study was to investigate the effects of BIOSTRONG 510 on the precae-cal nutrient digestibility of male broilers raised to an age of 21 days. The data for the meta-analysis was obtained from 4 scientific trials of similar design.
Material and Methods A total of 216 male Cobb broilers (54 per trial) were randomly assigned to two treatment groups at day one of age. Chickens were kept in cages (0.19 m2) of three broilers. Temperature and lighting schedule were according to the breeder’s manual.
Table 1: Composition of experimental diet and calculated nutrients (4 experiments)
Starter feed (day 1-21) Ingredients Corn Soybean meal (48%) Soybean oil Limestone Monocalcium phosphate Vitamin-mineral premix1
Methionine Lysine Threonine Feed additive-marker premix2
(%)
52.82 35.00 7.00 1.47 1.40 1.20 0.28 0.16 0.02 0.65
Calculated nutrients AMEN Crude protein Lysine Methionine/Cystine Crude fibre Crude fat Starch Sugar Calcium Total phosphorus
MJ/kg g/kg g/kg g/kg g/kg g/kg g/kg g/kg g/kg g/kg
12.95
225.10 13.50 9.90
23.50 97.10
338.50 40.60 9.00 7.00
1) Premix contains per kg: 400,000 IE Vit. A; 40,000 IE Vit. D3; 4,200 mg Vit. E; 200 mg Vit. K3; 200 mg Vit. B1; 250 mg Vit. B2; 3,500 mg Niacin; 400 mg Vit. B6; 3,000 µg Vit. B12; 20,000 µg Biotin; 1,500 mg Pantothenic acid; 100 mg Folic acid; 80,000 mg Choline chloride; 5,000 mg Zn; 2,000 mg Fe; 5,000 mg Mn; 1,200 mg Cu; 50 mg J ; 40 mg Co; 35 mg Se; 125 g Na; 50 g Mg. 2) Additives: 0.5% Chromium oxide + 0.15% of test substances mixture (Negative control: placebo (carrier material); BS 510: 0.015% BIOSTRONG 510)

Wendler et al.: Influence of precaecal apparent digestibility in young broilers by a phytogenic feed additive
Seite 154 8. BOKU-Symposium Tierernährung 2009
Chickens were fed on a starter feed with or without BIOSTRONG 510 supplementation at a dosage of 150 g per ton of feed (table 1). Feed and water were available ad libitum. Feed consumption (per cage) and body weight (individually) were recorded weekly and feed conversion was calculated.
All birds were sacrificed after 21 experimental days and precaecal digesta was collected. Feed and digesta samples were analysed for dry matter, crude protein, crude fat, and crude ash (VDLUFA methods). Nutrient digestibility was calculated by the determination of the indigestible marker chro-mium oxide (5 g/kg feed) in feed and digesta.
Data were analysed by analysis of variance (GLM) and comparison of LS-means, using the procedures of SAS for Windows. Limit for significance was set at P<0.1 and P<0.05.
Results and Discussion There were no animal losses during the experiments. Daily veterinary inspection did not show abnor-malities in faeces consistency.
At the beginning of the experiment the body weight was 47.2 g on average. Chickens fed on the con-trol diet gained 776 g during the 21 day experimental period, which corresponds with an average daily weight gain of 37.0 g. The dietary supplementation of the phytogenic feed additive BIOSTRONG 510 resulted in increased weight gain of chickens compared with the control. At the end of the experiment the daily weight gain was about 38.4 g and, therefore, 3.8 % (P=0.028) higher compared with the control. The improvement in weight gain was the result of improved nutrient conversion (table 2).
Table 2: Zootechnical performance of broilers during 21 experimental days
Negative Control (LS means)
BIOSTRONG 510 (LS means) SEM* %
Body weight, g Day 1 Day 7 Day 14 Day 21
46.7 158.3 405.4b
822.7B
47.7 163.0 421.1a 852.5A
0.39 2.66 5.95 9.35
99.9 103.0 103.9 103.6
Daily feed intake, g Day 1-7 Day 7-14 Day 14-21 Day 1-21
15.4A 45.2 78.8 46.5
16.8B 44.5 78.7 46.6
0.40 1.09 1.36 0.69
108.7 98.4 99.9 100.4
Feed conversion, g/g Day 1-7 Day 7-14 Day 14-21 Day 1-21
1.01 1.30a 1.34 1.27
1.01 1.22b 1.29 1.22
0.036 0.032 0.031 0.021
100.0 93.5 96.4 96.5
Means with different superscripts (a, b) differ significantly at P<0.05 * standard error of mean Means with different superscripts (A, B) differ significantly at P<0.1
Improved conversion of nutrients was proven by measurement of precaecal digestibility of broilers.
The application of the phytogenic feed additive significantly increased apparent digestibility of protein, fat, ash, and of organic matter (table 3).

Wendler et al.: Influence of precaecal apparent digestibility in young broilers by a phytogenic feed additive
8. BOKU-Symposium Tierernährung 2009 Seite 155
Table 3: Precaecal digestibility of nutrients
Coefficient of digestion, %
Negative Control (LS means)
BIOSTRONG 510 (LS means) SEM* %
Organic matter Crude protein Crude fat Crude ash
67.6b
78.6b
91.1b
39.0b
72.7a
82.9a
93.6a 43.7a
0.70 0.50 0.53 0.66
107.5 105.1 102.3 112.1
Means with different superscripts (a, b) differ significantly at P<0.05 * standard error of mean (n=72)
An increase of the coefficient of ash digestion and thus, mineral digestibility, can be pronounced (P<0.001). The improvement of nutrient digestibility is in agreement with a former study, where the phytogenic feed additive significantly increased protein and mineral digestibility (Loh et al., 2008).
Conclusion The results of the present study clearly show that the phytogenic feed additive BIOSTRONG 510 improves nutrient digestibility and growth performance in growing broilers.
The increase in the coefficients of digestion may be the result of positive effects on the secretion and/or activity of digestive enzymes.
Summary Four experiments with male Cobb broilers were conducted to investigate the effects of the phytogenic feed additive BIOSTRONG 510 on the nutrient digestibility and the performance until 21 days of age. Chickens were kept in cages (3 broilers per cage) and were fed on iso-nutritive diets, mainly consist-ing of corn and soybean meal. Broilers were sacrificed at an age of 21 days and ileal digesta was col-lected for the determination of the precaecal apparent digestibility of nutrients. The addition of the phytogenic feed additive significantly improved digestibility of nutrients (protein, fat, ash). This was reflected in the improvement of zootechnical performance (daily gain, feed conversion ratio). In con-clusion BIOSTRONG 510 can be used for growing broilers to enhance digestion and improve perfor-mance.
References
LOH, T.C., LING, H.G., THANH, N.T., FOO, H.L., RAJION, M.A., and DAVID, S.I. (2008): Effects of feeding phytogenic substances and phytase on growth performance and nutrient digestibility of young broilers. J. Appl. Anim. Res. 33, 187-192.
Corresponding author
Mag. Andreas Asamer, MBA Delacon Biotechnik Ges.m.b.H Weissenwolffstr. 14 A-4221 Steyregg E-mail: [email protected]

Firmensponsoring – DELACON BIOTECHNIK GmbH, Weissenwolffstraße 14, A-4221 Steyregg
Seite 156 8. BOKU-Symposium Tierernährung 2009

Cypriano et al.: Performance and tibia characteristics of broilers fed a phytogenic feed additive
8. BOKU-Symposium Tierernährung 2009 Seite 157
Performance and tibia characteristics of broilers fed a phyto-genic feed additive
L. Cypriano1, I. Piccini2, J. B. P. Filho2, N. S. Pastore2, A. Mader3 and A. Asamer3 1 Technical Department, Eurotec Nutrition, Palhoça, SC, Brazil 2 Animal Nutrition Department, Federal University of Parana (UFPR), Palotina, PR,
Brazil 3 Delacon Biotechnik GmbH, Steyregg, Austria
Introduction Several alternatives have been investigated to improve the zootechnical performance of broilers. Es-pecially phytogenic feed additives seem to be suitable products because spices and herbs can stimu-late secretion of digestive juices and enzymes (RAO et al., 2003; PLATEL & SRINIVASAN, 2004). Increased secretion of digestive enzymes might improve nutrient break down and thus, increase availability and absorption of nutrients and minerals (LOH et al, 2008). The phytogenic feed additive BIOSTRONG 510 is a standardized combination of selected essential oils with herbs and spices rich in pungent sub-stances, flavonoids, and saponins. This feed additive was developed for usage in broilers and laying hens for improvement of animal performance (BUCHANAN et al., 2008). The aim of this study was to investigate effects of BIOSTRONG 510 on performance and on mineralization of bones of growing broilers under commercial conditions.
Material and Methods From October to December 2008, field trials were conducted to investigate BIOSTRONG® 510 in a broiler integration in Brazil. In the trials the Integration Standard Diets (ISDT) were compared with diets containing 150 ppm of BIOSTRONG® 510 (FullBT), considering the Suggested Apparent Nutri-tional Matrix (SANM) for the product (1 kg Biostrong equals 35,000 g protein, 215,000 kcal AMEn, 2,300 g arginine, 1,300 g isoleucine, 2,000 g lysine, 750 g methionine, 1,400 g methionine and cyste-ine, 1,300 g threonine, 350 g tryptophan, 4,800 g Ca, and 4,500 g P-available).
A total of 5,048,589 broilers, allotted in farms/houses of 24,000 to 24,500 broilers each, were housed in dark-houses for commercial feeding trials. The nutritional program was composed of five phases: prestarter (days 1 to 7), starter (days 8 to 17), grower 1 (days 18 to 26), grower 2 (days 27 to 38) and finisher (day 39 to slaughter). Diets of both treatments were based on corn, soybean meal, bone meal and fat/soybean oil. Phytase and BIOSTRONG 510 were added to the diets considering the matrix values for both products.
The same coccidiostat program was used in both treatments. Temperature and lighting schedule were set according to the breeder’s manual. Broilers of ISDT group were slaughtered at the age of 45.06 days on average; birds of FullBT group were slaughtered at the age of 45.22 days on average. The slaughter was done in the facility of the integration.
Eight comparable houses of 24,500 Cobb broilers each were selected for tibia analyses in each treat-ment group. At 39 days of age, which is generally a period of increased incidence of leg problems in integrations, 10 males of the median weight of 50 birds per house were selected from each house (i.e.

Cypriano et al.: Performance and tibia characteristics of broilers fed a phytogenic feed additive
Seite 158 8. BOKU-Symposium Tierernährung 2009
10 birds per replication, 8 replications per treatment). The broilers were slaughtered, the left tibias removed and packed for laboratory analysis. The measurement was performed according to the me-thod of KOCABAĞLI (2001) at UFPR in the Animal Nutrition Laboratory. The robustness index of tibia was calculated from tibia length and the cube root of tibia weight. Furthermore, phosphorus and cal-cium contents were measured from bone ash.
The zootechnical data and the tibia parameters were done by analysis of variance (GLM) and compari-son of LS-means, using the procedures of SAS for Windows. Limit for significance was set at P<0.1 and P<0.05.
Results and Discussion The zootechnical results show typical data regarding the performance of farms of Brazil integrators and are presented in table 1.
Table 1: Zootechnical performance of broilers (commercial conditions)
ISDT (LS means)
FullBT (LS means)
SEM*
%
Production index 325 325 4.05 99.7
Mortality (%) 4.93 4.34 0.37 87.9
Live weight (kg) 2.73 2.70 0.02 99.0
Daily weight gain (g/d) 60.6 59.9 0.42 99.0
Feed conversion (kg/kg) 1.77 1.77 0.01 99.8
Feed conversion corrected (2.5 kg live weight) 1.70 1.71 0.01 100.3
* Standard error of the mean (n=31)
No statistical differences were identified for the investigated criteria. Chickens of the ISDT group showed the same performance as FullBT chickens in daily weight gain and live weight. The nutrient reduction by the matrix application of BIOSTRONG® 510 in the FullBT treatment did slightly reduce mortality (- 12 %) in comparison with ISDT. The obtained results of tibia parameters are presented in table 2.
Table 2: Tibia parameters of broilers
ISDT FullBT SEM * % Tibia parameters Robustness index 4.94a 4.90b 0.02 99.2 Mineral (%) 42.19 41.96 0.26 99.5 Calcium (%) 16.34B 17.15A 0.18 104.9 Phosphorus (%) 7.59 7.60 0.06 100.8 Means with different superscripts (a, b) differ significantly at P<0.05 * Standard error of the mean (n=8) Means with different superscripts (A, B) differ significantly at P<0.1
Tibia analyses show that phosphorus content was similar in both treatments, while calcium content was higher in the FullBT group, despite the lower dietary calcium and available phosphorus content. Tibia robustness index was lower in the FullBT group compared with the ISDT group (P<0.05), which indicates stronger bones (HALL et al., 2003).

Cypriano et al.: Performance and tibia characteristics of broilers fed a phytogenic feed additive
8. BOKU-Symposium Tierernährung 2009 Seite 159
Conclusion The absence of treatment differences in the zootechnical performance and the improvement in ro-bustness and calcium content of tibia confirm the nutritional matrix of BIOSTRONG® 510 in corn-soybean diets at concomitant use of phytases.
The application of BIOSTRONG® 510 and its nutritional matrix offer a reliable tool for cost reduction in the formulation of broiler diets.
Summary A commercial trial was conducted to study the effects of the inclusion of the phytogenic feed additive BIOSTRONG® 510 on performance and bone mineralization with application of the Suggested Appar-ent Nutritional Matrix (SANM) of the feed additive in feed formulation. Performance parameters were calculated from a total of approximately 5 million birds, fed a corn soybean diet (integrator standard diet) for 45 days. Tibia parameters of 160 male Cobb broilers (10 birds per replication, 8 replications per treatment) were measured at Animal Nutrition Laboratory of UFPR.
The dietary addition of BIOSTRONG® 510 and the application of the nutritional matrix led to similar performance as obtained with the standard diet. Bone calcium content and the robustness of tibia were increased in the supplemented group, showing better mineralization of bones and thus, in-creased mineral retention.
References
BUCHANAN, N. P., J. M. HOTT, S. E. CUTLIP, A. L. RACK, A. ASAMER and J. S. MORITZ (2008): The Effects of a Natural antibiotic alternative and a natural growth promoter feed additive on broiler performance and carcass quality. J. Appl. Poult. Res. 17, 202-210
HALL, L. E., R. B. SHIRLEY, R. I. BAKALLI, S. E. AGGREY, G. M. PESTI and H. M. EDWARDS et al. (2003): Power of two methods for the estimation of bone ash of broilers. Poultry Sci. 82, 414-418
KOCABAĞLI, N. (2001): The effect of dietary phytase supplementation at different levels on tibial bone characteristics and strength in broilers. Turk. J. Vet. Anim. Sci. 25:797–802.
LOH, T.C., LING, H.G., THANH, N.T., FOO, H.L., RAJION, M.A., and DAVID, S.I. (2008): Effects of feeding phytogenic substances and phytase on growth performance and nutrient digestibility of young broilers. J. Appl. Anim. Res. 33, 187-192.
PLATEL, K. & SRINIVASAN, K. (2004): Digestive stimulant action of spices: A myth or reality? Indian J Med Res 119, 167-179
RAO, R. R., K. PLATEL and K. SRINIVASAN (2003): In vitro influence of spices and spice-active principles on digestive enzymes of rat pancreas and small intestine. Nahrung/Food 47, No. 6, 408-412
Corresponding author
Mag. Andreas Asamer, MBA Delacon Biotechnik Ges.m.b.H Weissenwolffstr. 14 A-4221 Steyregg E-mail: [email protected]

Gärtner und Zentek: Untersuchungen zur Wirkung eines pflanzlichen Futterzusatzes auf einen enteropathogenen Escherichia coli- Stamm
Seite 160 8. BOKU-Symposium Tierernährung 2009
Untersuchungen zur Wirkung eines pflanzlichen Futterzusatzes auf einen enteropathogenen Escherichia coli- Stamm
Stefanie Gärtner, Jürgen Zentek
Institut für Tierernährung, Fachbereich Veterinärmedizin, Freie Universität Berlin
Einleitung Die Zeit nach dem Absetzen stellt beim Schwein auf Grund der Durchfallproblematik, verursacht durch enteropathogene Escherichia (E.) coli Stämme, noch immer eine kritische Phase dar. Wesentliche Voraussetzung für die Erkrankung ist die Adhäsion von pathogenen Keimen an Darmepithelzellen. Eine Proliferation darmpathogener Bakterien kann insbesondere dann stattfinden, wenn Abwehrmechanis-men beeinträchtigt sind (Kastner et al. 2002). In der Absetzphase sind diese durch verschiedene Fak-toren, z.B. Stresseinwirkungen, geschwächt, wodurch Infektionen begünstigt werden.
Die Adhäsion von Bakterien an der Darmwand ist ein komplexer Vorgang und zeichnet sich durch Gewebetropismus sowie Spezies- und Altersspezifität aus. Sie erfolgt bei E.coli Rezeptor vermittelt über Fimbrien und membranständige Adhäsine an der Darmschleimhaut (Kastner et al. 2002; Ofek et al. 1977). Biochemisch gesehen handelt es sich bei den membranständigen Rezeptoren um Glykokon-jugate mit verschiedenen Oligosaccharidsequenzen, die von den Bakterien als Rezeptorbindungsstelle erkannt werden (Neeser et al. 1986; Ofek et al. 1977).
Eine Blockierung dieser Adhäsionsprozesse an Darmepithelzellen hat in den letzten Jahren großes Interesse bei der Suche nach prophylaktischen Möglichkeiten gegen Durchfallerkrankungen geweckt. Besondere Aufmerksamkeit galt hier der Suche nach Rezeptoranaloga. Dabei erwiesen sich Mannose, Glykopeptide vom Oligomannosidtyp und Oligogalakturonoide hinsichtlich einer Blockierung der Adhä-sion von enteropathogenen und enterotoxischen E.coli- Stämmen und Salmonella enterica Stämmen an humanen und porzinen Epithelzellen als wirksam (Di Martino et al. 2006; Kastner et al. 2002; Naughton et al. 2001; Neeser et al. 1986; Ofek et al 1977).
Vor diesem Hintergrund erfolgten in der Studie Untersuchungen mit einem phytogenen Futterzusatz-stoff, in welchem neben ätherischen Ölen auch Kräuter und Gewürze (Wirkstoffe: Schleimstoffe (u.a. Galactomannane), Flavonoide, Scharfstoffe) enthalten sind, zur Adhäsion eines enteropathogenen E. coli- Stamms unter drei verschiedenen Gesichtspunkten. In einem in-vitro Infektionsmodell wurde der Einfluss von FRESTA® F Conc. (Fa. DELACON) auf die Adhäsion eines E. coli- Stamms an eine IPEC-J2 Zelllinie untersucht. In einem zweiten Modell erfolgten Untersuchungen zur Bindungskapazität des Produktes für den untersuchten E. coli Stamm. In einem dritten Ansatz sollte der Einfluss der Konzent-ration von FRESTA® F Conc. (Fa. DELACON) auf das Wachstumsverhalten des E. coli Stamms weite-ren Aufschluss über mögliche Wirkmechanismen pflanzlicher Materialien geben.
Material und Methoden Im ersten Modell wurde der Einfluss von FRESTA® F Conc. (Fa. DELACON, Steyregg, Austria) auf die Adhäsion eines enteropathogenen E.coli Stammes an einer permanenten porzinen Darmzelllinie (IPEC-J2) untersucht Der E.coli- Stamm wurde mittels Multiplex-PCR für die Expression von F4, estIa und estII positiv getestet und für alle drei Modelle verwendet.

Gärtner und Zentek: Untersuchungen zur Wirkung eines pflanzlichen Futterzusatzes auf einen enteropathogenen Escherichia coli- Stamm
8. BOKU-Symposium Tierernährung 2009 Seite 161
Die bei -80°C gelagerten Zellen wurden nach Auftauen in ein Zellmedium (MegaCellTM Dulbecco`s, Sigma-Aldrich, St. Louis; USA) verbracht und bei 37°C in einer 5%-igen CO2 Atmosphäre für 7 Tage inkubiert (Schierack et al. 2006). Nach Ausbildung eines beständigen Monolayers wurden die Zellen weiter passagiert und ab Passage 4 für den Versuch eingesetzt. Dafür wurden sie in einer Konzentra-tion von 1x105/ml Zellmedium und einem Volumen von 1ml in eine 6-Well Mikrotiter-Platte (Greiner BIO-One B.V.) pipettiert und für 48 Stunden erneut inkubiert. Die Anzüchtung des bei -80°C gelager-ten E.coli- Stamms erfolgte nach dem Auftauen für 24 Stunden bei 37°C auf LB (Luria-Bertini) –Agar (10g Trypton, 5g Hefe-Extrakt, 10g NaCl, 15g Agar auf 1l Aqua bidest., pH 7,5). Für den Versuch wurden Einzelkolonien in 3ml LB-Medium (10g Trypton, 5g Hefe-Extrakt, 10g NaCl auf 1l Aqua bidest., pH 7,0) verbracht und nochmals für 12 Stunden bei 37°C und 80rpm inkubiert.
Das zu testende Produkt wurde mit PBS (PBS Dulbecco Instamed 9,55g/l, BIOCHROM AG, Berlin; Germany) 1%-ig gelöst, der Überstand mit 0,2μm Filtern steril aufgereinigt und 1:100 in PBS ver-dünnt.
Die Färbung der E. coli erfolgte mit 2μl CFDA-Se- Farbstoff (10μl 100mM Carboxyfluorescein Diacetate Succinimidyl Ester auf 190μl Dimethylsulfoxid; Fa. Sigma Aldrich, St. Louis, USA) bei 37°C für 2 Stun-den. Anschließend wurden jeweils 1x106 Bakterien/ml und 1ml Testsubstrat als Vierfachansatz auf die IPEC-J2 Zelllinie pipettiert und die Mikrotiter-Platten für 90 Minuten bei 37°C inkubiert. Für die Nega-tivkontrolle wurde PBS, für die Positivkontrolle 1x106 Bakterien/ml auf die Zellen pipettiert. Schließlich wurden die Zellmonolayer vorsichtig mit einem sterilen Spatel aus den Wells gelöst und nach einmali-ger Zentrifugation (1200rpm, 10 Minuten, 4°C) am Durchflusszytometer (FACS Calibur, Fa. BD) ge-messen.
Für die Untersuchungen zur bakteriellen Bindungskapazität wurden in einem zweiten Modell Mikroti-ter-Platten mit dem Pflanzenmaterial beschichtet und die Bindungskapazität für den enteropathogenen E. coli Stamm untersucht (Becker et al. 2007). Für den Coating- Prozess wurden nach Herstellung einer 1%-igen wässrigen (PBS) Lösung von FRESTA® F Conc. (Fa. DELACON) und BSA (>98%; Fa. Roth, Karlsruhe, Germany) als Negativkontrolle, jeweils 350μl Substrat/Well in einem Vierfachansatz auf eine 96-Well Mikrotiter-Platte (Microlon F; Greiner Bio-One B.V.) pipettiert und für 12 Stunden bei 4°C inkubiert. Nach einer Waschung mit PBS zur Entfernung von nicht gebundenem Material wurde eine 1%-ige BSA/ 0,5%-ige Natriumazidlösung (in PBS) für 30 Minuten bei 4°C zugegeben. Danach erfolgte eine zweimalige Waschung mit PBS (Becker et al. 2007).
Die Kultivierung des E. coli Stamms erfolgte wiederum auf LB-Agar bei 37°C für 24 Stunden und an-schließend als Einzelkolonie in 3ml LB-Medium bei 37°C für 3,5 Stunden mit 80 rpm. Die Bakterien wurden in LB-Medium auf eine Konzentration von 1x107/ml eingestellt. In jedes Well kamen 300μl der Bakteriensuspension. Anschließend wurden die Platten bei Raumtemperatur (22°C) für 30 Minuten inkubiert. Für die Positivkontrolle wurden nicht gecoatete Wells benutzt. Um nicht gebundene Bakteri-en zu entfernen, erfolgte eine dreimalige Waschung mit PBS. Die anschließende Inkubation des Ansat-zes mit jeweils 300μl LB-Medium/Well erfolgte zur Abschätzung der Wachstumskurve der an die Sub-strate gebundenen Bakterien.
Die Messung der Bakteriendichte erfolgte semiquantitativ bei 620nm, 36°C-38°C über 24 Stunden mittels Photometer(sunrise, Fa. TECAN, Germany). Vor jeder Messung wurden die Platten jeweils 3 Sekunden geschüttelt.
Die Kultivierung der Bakterien für den dritten Modellansatz erfolgte unter Zusatz verschiedener Kon-zentrationen von FRESTA® F Conc (Fa. DELACON) zur Erfassung möglicher direkter Effekte auf das Wachstumsverhalten. Die Bakterien wurden dazu in LB-Medium auf eine Konzentration von 1x107/ml eingestellt. Für die geometrische Verdünnungsreihe wurde mit PBS eine 1%-ige wässrige Lösung der Testsubstanz hergestellt. Auf eine 96-Well Mikrotiter-Platte (Greiner BIO-One B.V.) wurde in einem Dreifachansatz folgende Verdünnungsreihe mit jeweils 100μl des wässrigen pflanzlichen Substrats pipettiert: 1%, 0,5%, 0,25%, 0,125%, 0,06%, 0,03%. Anschließend erfolgte die Zugabe der Bakteri-

Gärtner und Zentek: Untersuchungen zur Wirkung eines pflanzlichen Futterzusatzes auf einen enteropathogenen Escherichia coli- Stamm
Seite 162 8. BOKU-Symposium Tierernährung 2009
ensuspension mit 100μl/Well. Als Negativkontrolle dienten 100μl LB-Medium, als Positivkontrolle 100μl Bakteriensuspension in 100μl LB-Medium. Zusätzlich wurde eine Platte zur Messung der Leerwerte für die jeweiligen Konzentrationen der Testsubstanz pipettiert.
Die Messung erfolgte bei 620nm, 36°C-38°C über 24 Stunden mittels Photometer (sunrise, Fa. TECAN, Germany). Vor jeder Messung wurden die Platten jeweils 3 Sekunden geschüttelt.
Die Daten wurden mit Microsoft Office Excel 2003 und SPSS 15.0 für Windows nach Prüfung auf Nor-malverteilung mittels einfaktorieller Varianzanalyse (p<0,05) oder nichtparametrischen Testverfahren (Kruskal-Wallis-Test, Mann-Whitney-Test) ausgewertet.
Ergebnisse Adhäsion des pathogenen E. coli- Stamms an IPEC-J2
Die Ergebnisse der in-vitro Versuche zeigten deutliche Effekte durch den Einsatz von FRESTA® F Conc (Fa. DELACON) untersuchten Pflanzenrohstoffe auf die Adhäsion und das Wachstumsverhalten des geprüften enteropathogenen E. coli -Stamms.
Für die Adhäsion von E. coli an die IPEC-J2 Zelllinie vom Schwein konnte eine signifikante Reduktion (p<0,05) nachgewiesen werden.
Abbildung 1 verdeutlicht den Einfluss des Zusatzes auf die Fluoreszenz von IPEC-J2 Zellen. Die Fluo-reszenz dient als Messparameter für die Menge zellgebundener Bakterien. Der Fluoreszenzkoeffizient gibt an, um wie viel sich die Fluoreszenz nach Zusatz der Stoffe verändert. Bei niedrigen Werten ist davon auszugehen, dass weniger E. coli an die Zellen adhäriert haben. Unter dem Einfluss von FRESTA® F Conc stieg die Fluoreszenzstärke um das 0,96-fache. Zum Vergleich stieg die Fluoreszenz-stärke in der Positivkontrolle, also bei Koinkubation von Epithelzellen mit E. coli, um das 2-fache.
0,00
0,50
1,00
1,50
2,00
2,50
3,00
3,50
Flu
ore
sze
nzk
oe
ffiz
ien
t
Kontrolle (E. coli )
FRESTA® F Conc.
b
a
Abb.1: Einflluss von Fresta® F Conc. auf die Adhäsion CFDA-SE markierter enteropathogener E.coli an eine porzine IPEC-J2 Zelllinie (n=26). Mittelwerte mit unterschiedlichen Indizes unterscheiden sich signifikant (p< 0,05)
Untersuchungen zur Bindungskapazität des pflanzlichen Produkts für den untersuchten E. coli- Stamm
In den Untersuchungen zur Bindung des E. coli- Stamms an die Prüfsubstanzen war kein Unterschied in der Bakteriendichte zur Positiv- bzw. Negativkontrolle erkennbar. Die Messungen wurden nach 8 Stunden Inkubation ausgewertet, da sich in dieser Zeitphase eine logarithmische Wachstumskurve darstellte (Tabelle 1).

Gärtner und Zentek: Untersuchungen zur Wirkung eines pflanzlichen Futterzusatzes auf einen enteropathogenen Escherichia coli- Stamm
8. BOKU-Symposium Tierernährung 2009 Seite 163
Tab. 1: Extinktion bei 620 nm in der exponentiellen Wachstumsphase eines E. coli- Stamms (t=8 h) als Maß der bakteriellen Bindungskapazität für das getestete Produkt. (n=12)
OD mean± SD
FRESTA®F Conc. 0,33ab± 0,19
Positivkontrolle (E.coli) 0,48a± 0,05
Negativkontrolle (BSA) 0,04b± 0,03
Mittelwerte mit unterschiedlichen Indizes unterscheiden sich signifikant (p<0,05).
Einfluss unterschiedlicher Konzentration des Testprodukts auf das Wachstumsverhalten des E. coli- Stamms
Ein Einfluss des Pflanzenproduktes auf das Wachstumsverhalten des geprüften E. coli Stamms konnte weder bei einer Konzentration von 1% noch bei den weiteren Verdünnungsstufen beobachtet werden (Abbildung 2).
0
0,2
0,4
0,6
0,8
1
1,2
1,4
Ex
tin
kti
on
62
0 n
m
Positivkontrolle(E. coli)
FRESTA® F Conc.
Negativkontrolle (LB-Medium)
a
b
a
Abb.2: Einfluss von FRESTA® F Conc. (1% w/v) in der Phase des exponentiellen Wachstums (t= 10h) auf einen enteropatho-genen E. coli- Stamms. Mittelwerte mit unterschiedlichen Indizes unterscheiden sich signifikant (p<0,05).
Diskussion Die vorliegenden Resultate zeigen, dass das eingesetzte pflanzliche Präparat unter den gewählten in-vitro Bedingungen mit dem geprüften E. coli -Stamm interagiert. In der vorliegenden Studie führte der Einsatz von FRESTA® F Conc. zu einer Reduktion der Adhäsion des enteropathogenen E. coli -Stamms an eine porzine IPEC-J2 Zelllinie. Dieser Effekt ist möglicherweise auf eine Bindung der Bakterien durch Rezeptoranaloga wie die z. B. enthaltenen Galactomannane zurückzuführen. Andere Studien konnten Effekte von Kohlenhydratfraktionen im Sinne einer Reduktion bakterieller Adhärenz an Epi-thelzellen zeigen (Kastner et al. 2002; Naughton et al. 2001; Neeser et al. 1986; Ofek et al 1977).
Das Potential pflanzlicher Materialien, Bindungsstellen an Epithelzellen für Bakterien zu imitieren, wird durch die Kohlenhydratstruktur als auch durch deren Stabilität im Verdauungstrakt beeinflusst (Kastner et al. 2002). Die in FRESTA® F Conc. eingesetzten Pflanzenkomponenten enthalten ein brei-tes Wirkstoffspektrum. In dem Teilversuch zur Überprüfung der bakteriellen Adhäsion an gecoatete Mikrotiter-Platten konnte für FRESTA® F Conc. eine bakterielle Bindungskapazität beobachtet werden,

Gärtner und Zentek: Untersuchungen zur Wirkung eines pflanzlichen Futterzusatzes auf einen enteropathogenen Escherichia coli- Stamm
Seite 164 8. BOKU-Symposium Tierernährung 2009
die numerisch höher als in der Negativkontrolle (mit BSA gecoatete Wells) und unterhalb des als Posi-tivkontrolle gewählten direkten Coatings der Wells mit E. coli lag. Die von diesen Daten zu vermuten-de Eigenschaft zur Bindung von Bakterien ist auch für andere komplexe Pflanzenmaterialien wie Karot-tenpektine (Kastner et al. 2002), Mannan-Oligosaccharide (Becker et al. 2007) oder Preiselbeersaft (Di Martino et al. 2006) beschrieben worden. Die im dritten Versuchsansatz beobachtete Wachstumskine-tik bei Ko-Kultivierung des E. coli Stamms mit dem Produkt in unterschiedlichen Konzentrationen er-gab keine Hinweise auf eine vollständige wachstumshemmende Wirkung. Insofern ist dieser Wirkme-chanismus zumindest für die gewählten Prüfbedingungen auszuschließen. Aus den vorliegenden Re-sultaten können noch keine Aussagen hinsichtlich eines unter in in-vivo Bedingungen relevanten Wirkmechanismus getroffen werden. Die hier vorgestellten in-vitro Untersuchungen sind geeignet, prinzipielle Wirkmechanismen pflanzlicher Substanzen unter den spezifischen Prüfbedingungen zu beschreiben. Eine Reduktion der Bindung pathogener E. coli an Darmepithelzellen wäre von großem Interesse und könnte durch Interaktionen von Inhaltsstoffen pflanzlicher Produkte mit den Bakterien oder auch Bindungsstellen in den Materialien plausibel erklärt werden.
Schlussfolgernd weisen die Ergebnisse auf eine Bindungskapazität von FRESTA® F Conc. gegenüber einem enteropathogenen E. coli- Stamm hin. Eine Hemmung der Adhärenz an Darmepithelien bzw. kompetetive Bindung von E. coli an pflanzliche Substanzen könnten in-vivo wichtige Wirkmechanismen zur Prophylaxe absetzbedingter Durchfallerkrankungen sein, die in Fütterungsversuchen mit Ferkeln weiter evaluiert werden müssen.
Literatur
U. Kastner, S. Glasl, B. Follrich, JP. Guggenbichler und J. Jurenitsch (2002): [Acid Oligosaccharides as Active Principle of Aqueous Carrot Extracts in the Prophylaxis and Therapy of Gastrointestinal Infections] Saure Oligosaccharide als Wirkprinzip von wässrigen Zubereitungen aus der Karotte in der Prophylaxe und Therapie von gastrointestinalen Infektionen. Wien Med Wochenschr 152,379-381
P. J. Naughton, Lene Lind Mikkelsen, Bent Borg Jensen (2001): Effects of Nondigestible Oligosaccharides on Salmonella enterica Serovar Typhimurium and Nonpathogenic Escherichia coli in the Pig Small Intestine in Vitro. Appl Environ Microbiol 67, 3391-3395
J.-R. Neeser, B. Koellreutter, P. Wuersch (1986): Oligomannosid-Type Glycopeptides Inhibiting Adhesion of Escherichia coli Strains Mediated by Type 1 Pili: Preparation of Potent Inhibitors from Plant Glycoproteins. Infection and Immunity 52 (2), 428-436
A. Srichamroen, C.J. Field, A.B.R. Thomson, T.K. Basu (2008): The Modifying Effects of Galactomannan from Canadian-Grown Fenugreek (Trigonella foenum-graecum) on the Glycemic and Lipidemic Status in Rats. J Clin Biochem Nutr 43,167-174
I. Ofek, D. Mirelman, N. Sharon (1977): Adherence of Escherichia coli to human mucosal cells mediated by mannose receptors. Nature 265,623-625
P.M. Becker, S. Galletti, P.J. Roubos-van den Hil, P.G. van Wikselaar (2007): Validation of growth as measurand for bacterial adhesion to food and feed ingredients. J Appl Microbiol 103, 2686-2696
P. Di Martino, R. Agniel, K. David, C. Templer, J.L. Gaillard, P. Denys, H. Botto (2006): Reduction of Escherichia coli adherence to uroepithelial bladder cells after consumption of cranberry juice: a double-blind randomized placebo-controlled cross-over trial. World J Urol 24, 21-27
P. Schierack, M. Nordhoff, M. Pollmann, K.D. Weyrauch, S. Amasheh, U. Lodemann, J. Jores, B. Tachu, S. Kleta, A. Blikslager, K. Tedin, L. H. Wieler (2006): Characterization of a porcine intestinal epithelial cell line for in vitro studies of microbial patho-genesis in swine. Histochem Cell Biol 125, 293- 305
Autorenanschrift
Stefanie Gärtner Institut für Tierernährung, Fachbereich Veterinärmedizin Freie Universität Berlin Brümmerstr. 34, D- 14195 Berlin E- Mail: [email protected]

Kang et al.: Performance parameters and meat quality of broiler fed diets supplemented with a mixture of Quaternary Benzophenanthrinde and Protopine alkaloids (Sangrovit®)
8. BOKU-Symposium Tierernährung 2009 Seite 165
Performance parameters and meat quality of broiler fed diets supplemented with a mixture of Quaternary Benzophenan-thrinde and Protopine alkaloids (Sangrovit®)
Chang Won Kang1, Byung Ki Ahn1, Ji Sook Kim1, Kathrin Tschirner2 1 Animal Resources Research Centre, College of Animal Bioscience and Technology,
Konkuk University, Seoul, Korea 2 Phytobiotics Futterzusatzstoffe GmbH, Eltville; Germany
Introduction The European Union banned antibiotic growth promoters (AGP) in 2006. Since then other regions have been discussing this strategy and the search for alternatives to antibiotics has been critical. However, some questions still remain with regard to the efficacy and mode of action of the replace-ments.
Plant extracts, herbs and spices can play a good role in supporting both performance and health status of poultry (Gill, 2000; Manzanillo et al., 2001). Beneficial effects of secondary plant metabolites in poultry nutrition may include the stimulation of appetite and feed intake, the improvement of en-dogenous digestive enzyme secretion, activation of immune response and antibacterial, antiviral, anti-oxidant and antihelminthic actions (Wenk, 2003; Jamroz et al., 2005; Vieira et al., 2008). Conse-quently, they can be seen as candidates for inclusion in animal feed as an alternative to AGP’s.
The herbal feed additive Sangrovit® is a standardized mixture of Quarternary Benzo[c]phenanthridine and Protopine alkaloids (QBA/PA) from Papaveraceae plants. Therefore, the purpose of the present study was to evaluate the effect of Sangrovit® on growth performance, carcass characteristics and quality and cecal microflora in broiler.
Material and Methods A total number of 630 Ross male broiler chicks (average individual weight 46 g; 3 treatments x 7 rep-lication x 30 bird each) were used. The birds were randomly allocated to one of the three treatment groups: I) Negative-control without antibiotic growth promoter [NC]; II: Negative-control with Avila-mycin 10 ppm [PC]; III): Negative-control with Sangrovit® 20 mg/kg [S20]. All diets were formulated to be isonitrogenous and isocaloric and, to met or exceed the minimum requirements of NRC (1994). Diet composition is shown in Table 1. A two phase feeding system was applied (Starter: d1-21; Grower: d22-35). The birds were fed ad libitum and had free access to water.
Feed intake and body weight (BW) of each group were recorded weekly. At the end of the experimen-tal period, seven chicks from each treatment were selected and weighed individually. At necropsy, abdominal fat, breast muscle and right leg (thigh and drumstick) were immediately removed and weighed. The cecal digesta homogenates were serially diluted from 10-1 to 10-5 in phosphate buffered saline. Dilutions were subsequently plated on duplicate selective agar media for enumeration of target bacterial strains. Total microbes, coliforms and Lactobacillus spp. were enumerated using nutrient agar, MacConkey agar, and MRS agar, respectively; using the traditional method (Tuohy et al., 2002). Each plate was incubated at 37°C, for 24 hr anaerobically or aerobically, and colonies were then
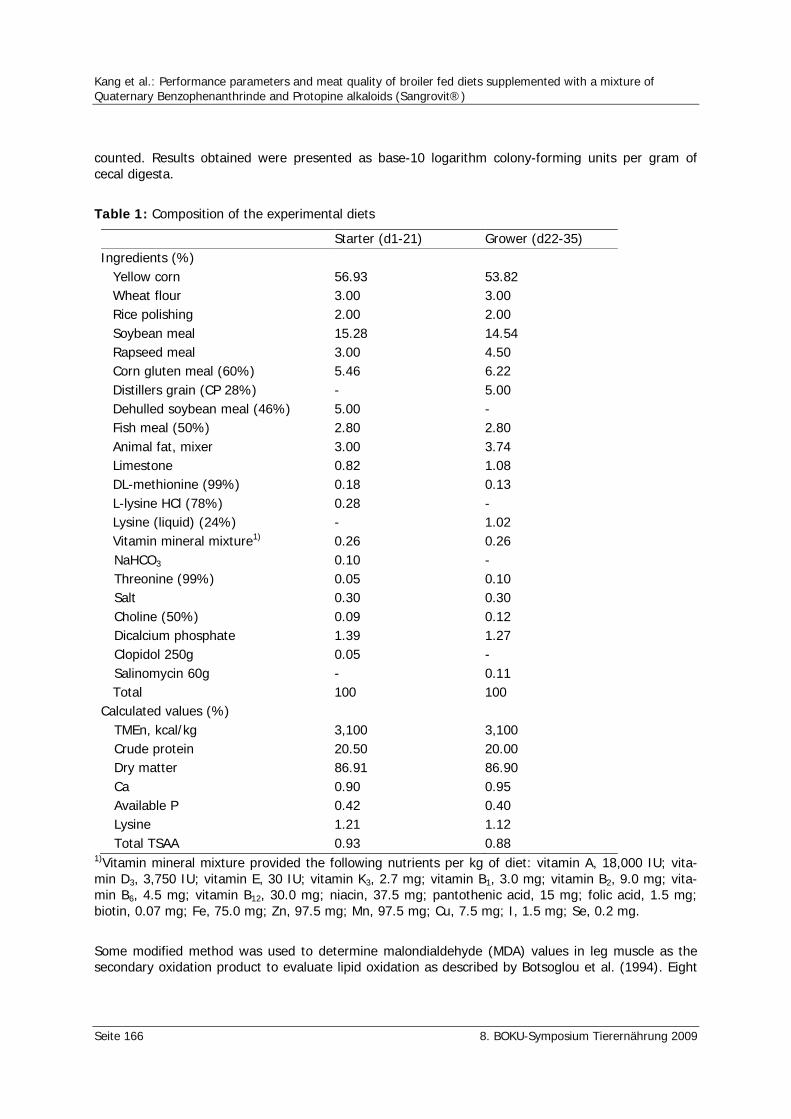
Kang et al.: Performance parameters and meat quality of broiler fed diets supplemented with a mixture of Quaternary Benzophenanthrinde and Protopine alkaloids (Sangrovit®)
Seite 166 8. BOKU-Symposium Tierernährung 2009
counted. Results obtained were presented as base-10 logarithm colony-forming units per gram of cecal digesta.
Table 1: Composition of the experimental diets
Starter (d1-21) Grower (d22-35) Ingredients (%)
Yellow corn 56.93 53.82 Wheat flour 3.00 3.00 Rice polishing 2.00 2.00 Soybean meal 15.28 14.54 Rapseed meal 3.00 4.50 Corn gluten meal (60%) 5.46 6.22 Distillers grain (CP 28%) - 5.00 Dehulled soybean meal (46%) 5.00 - Fish meal (50%) 2.80 2.80 Animal fat, mixer 3.00 3.74 Limestone 0.82 1.08 DL-methionine (99%) 0.18 0.13 L-lysine HCl (78%) 0.28 - Lysine (liquid) (24%) - 1.02 Vitamin mineral mixture1) 0.26 0.26 NaHCO3 0.10 - Threonine (99%) 0.05 0.10 Salt 0.30 0.30 Choline (50%) 0.09 0.12 Dicalcium phosphate 1.39 1.27 Clopidol 250g 0.05 - Salinomycin 60g - 0.11 Total 100 100
Calculated values (%) TMEn, kcal/kg 3,100 3,100 Crude protein 20.50 20.00 Dry matter 86.91 86.90 Ca 0.90 0.95 Available P 0.42 0.40 Lysine 1.21 1.12 Total TSAA 0.93 0.88
1)Vitamin mineral mixture provided the following nutrients per kg of diet: vitamin A, 18,000 IU; vita-min D3, 3,750 IU; vitamin E, 30 IU; vitamin K3, 2.7 mg; vitamin B1, 3.0 mg; vitamin B2, 9.0 mg; vita-min B6, 4.5 mg; vitamin B12, 30.0 mg; niacin, 37.5 mg; pantothenic acid, 15 mg; folic acid, 1.5 mg; biotin, 0.07 mg; Fe, 75.0 mg; Zn, 97.5 mg; Mn, 97.5 mg; Cu, 7.5 mg; I, 1.5 mg; Se, 0.2 mg.
Some modified method was used to determine malondialdehyde (MDA) values in leg muscle as the secondary oxidation product to evaluate lipid oxidation as described by Botsoglou et al. (1994). Eight

Kang et al.: Performance parameters and meat quality of broiler fed diets supplemented with a mixture of Quaternary Benzophenanthrinde and Protopine alkaloids (Sangrovit®)
8. BOKU-Symposium Tierernährung 2009 Seite 167
samples from each group were placed in a high temperature cabinet over 3 days of storage at 35°C. A 2 g sample was homogenized with 5% aqueous trichloroacetic acid solution containing 0.8% butylated hydroxytoluene, and then centrifuged for 3 min at 3,000 g. Following reaction with tiobarbituric acid reagent, MDA was directly quantified by third-derivative spectrophotometry against blank reaction mixture (Beckman DU-650, Beckman Counter, Inc.).
The main effects between treated groups were subjected to ANOVA using the GLM procedure of SAS (2002), and significant differences were determined using Duncan’s multiple range test at the level of p<0.05 (Duncan, 1955).
Results and Discussion The effects of treatments on performance parameters are presented in Table 2. Feed intake was sig-nificantly affected from d22-35. In that period, animals in S20 and PC group showed an increased feed consumption (p<0.05). In the starter phase and the whole trial period feed intake was also nu-merically higher in the S20 and PC group, but this was not significant. Final BW and daily weight gain of PC and S20 were increased compared to those of NC (p<0.05). Feed conversion was also improved by the dietary treatment, with improvements attributable to Sangrovit (S20) and the antibiotic growth promoter (PC) (p<0.05).
Table 2: The dietary effects of Sangrovit® on growth performance of broilers from day 1 to 35
NC PC S20
Initial BW, g 46.70 ±0.00 46.76 ±0.02 46.76 ±0.02
Final BW, g 1836.0 ±27.78b 2103.8 ±8.28a 2066.9 ±13.01a
Feed intake, g/d
d1-21 58.96 ±0.61 58.27 ±0.21 59.23 ±0.32
d22-35 127.60 ±2.50b 138.72 ±1.05a 136.17 ±0.67a
d1-35 86.83 ±1.15 89.59 ±0.39 89.84 ±0.47
BW gain, g/d
d1-21 45.07 ±0.41 46.35 ±0.30 46.40 ±0.40
d22-35 59.33 ±2.59b 77.75 ±0.52a 75.69 ±1.26a
d1-35 51.55 ±0.95b 58.82 ±0.33a 57.32 ±0.51a
FCR, feed/gain
d1-21 1.31 ±0.01a 1.26 ±0.01b 1.28 ±0.01ab
d22-35 2.16 ±0.07a 1.79 ±0.02b 1.80 ±0.03b
d1-35 1.68 ±0.01a 1.52 ±0.00c 1.57 ±0.02b
a,b Mean±SE values in a same row with no common superscripts are significantly different (p<0.05)
Feeding the herbal feed additive to broiler chicks resulted in to almost equal performance levels to the ones of the antibiotic growth promoter group. Similar results were obtained with other plant extracts by Jamroz and Kamel (2002), such as improvements of daily BW gain and feed conversion rate. Cross et al. (2007) also reported that chickens fed with diets supplemented with thyme oil and yarrow herb

Kang et al.: Performance parameters and meat quality of broiler fed diets supplemented with a mixture of Quaternary Benzophenanthrinde and Protopine alkaloids (Sangrovit®)
Seite 168 8. BOKU-Symposium Tierernährung 2009
showed greater live BW, average BW gain and better feed conversion. Their beneficial effects may be partly due to the improved efficiency in the utilization of feed nutrients, resulting in enhanced growth performance. Interestingly, Niewold (2007) recently suggested that the effects of AGP are mainly permitted by anti-inflammatory mechanisms. Various studies show that QBA/PA play a role in inflam-mation processes by suppressing it (Lenfeld et al., 1981; Agarwal et al., 1991). Putting in considera-tion that the principle way of QBA/PA clearance, is its elimination with faeces (Kosina et al., 2004), it can be suggested that, this activity is mainly located on the intestinal level. Consequently, beside a stimulation of feed intake and digestive processes, the anti-inflammatory properties of QBA/PA can be proposed as an explanation for its benefits on animal performance.
The effects of the different treatments on carcass characteristics are presented in Table 3. Numerically PC group showed the lowest relative weight of abdominal fat and the highest proportion of breast meat, but not significantly. The relative weight of abdominal fat and breast muscle of the Sangrovit treated birds were in between PC and NC groups, again this was not statistically significant at 5% level. As parameter for lipid peroxidation the content of MDA in the meat was determined. The MDA concentration in stored meat was significantly lowered by feeding 20 ppm Sangrovit® (S20; 0.12±0.02) when compared to those of NC (0.19±0.00) and PC (0.20±0.03) groups (p<0.05). In a previous study, Florou-Paneri et al. (2006) observed that dietary use of herb reduced lipid oxidation of egg yolks. Botsoglou et al. (2005) also reported that egg yolks in layers fed diets with saffron exhib-ited increased resistance to lipid oxidation. There are demands for using natural products that can prevent lipid oxidation in lipid enriched animal foods, due to consumer preferences and toxicological concerns of synthetic antioxidants. The present study suggested that Sangrovit® could be used as a valuable feed additive for reducing lipid oxidation during storage.
Table 3: The dietary effects of Sangrovit® on carcass characteristics in broiler chicken 1
NC PC S20
Abdominal fat 1.62 ±0.08 1.39 ±0.08 1.51 ±0.09
Leg muscle 9.38 ±0.12 9.32 ±0.18 9.41 ±0.19
Breast muscle 8.64 ±0.20 9.11 ±0.23 8.96 ±0.20
1Mean±SE
The effects of the different diets on the relative weight and length of small intestinal section are shown in Table 4. The weight of the small intestine of birds fed with the Sangrovit supplemented diet significantly decreased or tended to be decreased when compared with those of NC and PC groups. On the other hand, the length of the duodenum and jejunum in S20 was significantly enlarged when compared to those of the negative control (p<0.05). Between PC and S20 group numerical differ-ences, with higher values for S20 group, were shown (p>0.05). The measurements of intestinal weight and length may reflect the fact that Sangrovit® promoted the development of small intestinal sections and caused thinning of their epithelia, which subsequently reduce the maintenance needs for epithelial regeneration.
No significant differences in the number of total microbes and coliforms among the treatment groups could be observed. Nevertheless, the number of lactic acid bacteria in the chicks fed with Sangrovit® (6.62±0.09 log/cfu/g) was significantly higher than those of NC (6.01±0.20 log/cfu/g) and PC (6.01±0.17 log/cfu/g) groups (p<0.05). This result agreed with that of Jamroz et al. (2005) who re-ported that chicken fed diets with 100 ppm plant extract had higher numbers of Lactobacillus spp. in intestinal contents. Feeding Sangrovit® resulted in a beneficial modulation of cecal microflora as evi-denced by the numerical increases in the concentration of lactic acid bacteria.

Kang et al.: Performance parameters and meat quality of broiler fed diets supplemented with a mixture of Quaternary Benzophenanthrinde and Protopine alkaloids (Sangrovit®)
8. BOKU-Symposium Tierernährung 2009 Seite 169
Table 4: The dietary effects of Sangrovit® on the weight and length of parts of the small intestine in broiler chicken 1
1Abbreviation: DU, duodenum; JE, jejunum; IL, ileum. ; a,bMean±SE values in a same row with no common superscripts are significantly different (p<0.05).
Conclusion The overall results indicate that Sangrovit® is an effective feed additive in commercial broiler diets in view of enhancing animal performance and promoting beneficial intestinal microflora. In addition an improved meat quality by means of delaying lipid peroxidation during storage was observed. It is pos-tulated that the improved performance may be partly attributed to the stimulated appetite, digestive tract development and anti-inflammatory properties.
References
Agarwal, S., Reynolds, M. A., Pou, S., Peterson, D. E., Charon, J. A., Suzuki, J. B. (1991). The effect of sanguinarine on human peripheral blood neutrophil viability and functions. Oral Microbiol. Immuno. 6, 51-61.
Botsoglou, N. A., D. J. Fletouris, G. E. Papageorgiou, V. N. Vassilopoulos, A. J. Mantis, and A. G. Trakatellis, 1994. Rapid, sensi-tive, and specific thiobarbituri acid method for measuring lipid peroxidation in animal tissue, food, and feedstuff samples. J. Agri. Food Chem. 42:1931-1937.
Botsoglou, N. A., P. Florou-Paneri, I. Nikolakakis, J. Diannenas, V. Dotas, E. N. Botsoglou, S. Aggelopoulos, 2005. Effect of dietary saffron (Crocus sativus L.) on the oxidative stability on egg yolk. Br. Poult. Sci. 46:701-707.
Cross D. E., R. M. McDevitt, K. Hillman, and T. Acamovic, 2007. The effect of herbs and their associated essential oils on per-formance, dietary digestibility and gut microflora in chickens from 7 to 28 days of age. Br. Poult. Sci. 48:496-506.
Duncan, D. B., 1955. Multiple range and multiple F test. Biometric. 11:1-4.
Gill, C., 2000. Botanical feed additives. Feed Int. 14-17.
Florou-Paneri, P., D. Dotas, I. Mitsopoulos, V. Dotas, E. Botsoglou I. Nikolakakis, and N. Botsoglou, 2006. Effect of feeding rosemary and α-tocopheryl acetate on hen performance and egg quality. J. Poult. Sci. 43:143-149.
Jamroz, D., and C. Kamel, 2002. Palnt extarcts enhance broiler performance. J. Anim. Sci. 80. Suppl. 1: 4 [abstract]
Jamroz, D., A. Wiliczkiewicz, T. Wertelecki, J. Orda, and J. Skorupinska, 2005. Use of active substances of plant origin in chicken diets based on maize and locally grown cereals. Br. Poult. Sci. 46:485-493.
Kosina P., Walterová D., Ulrichová J., Lichnovský V., Stiborová M., Rýdlová H., Vicar J., Krecman V.,
Brabec MJ., Simánek V., 2004. Sanguinarine and chelerythrine: assessment of safety on pigs in ninety days feeding experiment. Food Chem Toxicol. 42 (1):85-91.
Lenfeld, J., Kroutil, M., Maršálek, E., Slavik, J., Preininger, V., Šimánek, V. (1981). Antiinflammatory activity of quaternary benzophenanthridine alkaloids from Chelidonium majus. Planta Med. 43, 161-165.
NC PC S20 Weight (g/100g BW) DU 0.34 ±0.03 0.32 ±0.02 0.27 ±0.02
JE 0.67 ±0.05a 0.62 ±0.03a 0.47 ±0.01b IL 0.55 ±0.04 0.56 ±0.01 0.47 ±0.04
Length (cm/100g BW) DU 1.13 ±0.03b 1.17 ±0.03ab 1.24 ±0.02a
JE 3.06 ±0.10b 3.21 ±0.11ab 3.50 ±0.08a IL 3.12 ±0.10 3.22 ±0.07 3.36 ±0.07

Kang et al.: Performance parameters and meat quality of broiler fed diets supplemented with a mixture of Quaternary Benzophenanthrinde and Protopine alkaloids (Sangrovit®)
Seite 170 8. BOKU-Symposium Tierernährung 2009
Manzanillo, E. G., F. Baucells, C. Kamel, J. Morales, J. F. Perez, and J. Gasa, 2001. Effects of plant extracts on the performance and lower gut microflora of early weaned piglets. J. Anim. Sci. Suppl. 1:473 [abstract]
National Research Council. 1994. Nutrients Requirements of Poultry. 9th rev. ed. National Academic Press, Washington, DC.
Niewold, T.A., 2007. The Nonantibiotic Anti-Inflammatory Effect of Antimicrobial Growth Promoters, the Real Mode of Action? A Hypothesis. Poultry Science 86:605–609
SAS. 2002. SAS User’s guide. Statistics, ver. 8.e, Statistical Analysis System Institute. Inc., Cary, NC.
Tuohy, K. M., C. J. Ziemer, A. Klinder, Y. Knobel, B. L. Pool-Zobel and G.. R. Gibson. 2002. A human volunteer study to deter-mine the probiotic effects of lactulose powder on human colonic microbiota. Microb. Ecol. Health Dis. 14:165-173.
Vieira, S. L., O. A. Oyarzabal, D. M. Freitase, J. Berres, J. E. M. Pena, C. A. Torres, and J. L. B. Coneglian, 2008. Performance of broilers fed diets supplemented with Sanguinarine-like alkaloids and organic acid. J. Appl. Poult. Res. 17:128-133.
Wenk, C., 2003. Herb and botanicals as feed additives in monogastric animal. Asian-Aust. J. Anim. Sci. 16: 282-289.
Corresponding author
Dr. Karin Tschirner Phytobiotics Futterzusatzstoffe GmbH Rosengasse 9 D-65343 Eltville E-mail: [email protected]

Mareš et al.: Spice Industry By-products in Pig Nutrition
8. BOKU-Symposium Tierernährung 2009 Seite 171
Spice Industry By-products in Pig Nutrition
Petr Mareš, Ladislav Zeman, Michal Večerek
Mendel University of Agriculture and Forestry Brno
Introduction Plants of family Apiaceae are commonly used in human nutrition as spices. The specific secondary metabolites in these plants are connected with positive effect in animal metabolism. By-products com-ing processing of fruits used as spice in human nutrition was used as feed additive in pig production. The base sources were fruits of cumin (Carum carvi) and coriander (Coriandrum sativum). The same type of by-product form allspice (Pimenta dioica) was used as the similar chemical content was ana-lysed. Pimenta dioica belongs to Myrtaceae family. The chemical that is common in these products (speaking about final by-product used in experiments) is limonene. In frame of secondary metabolites composition limonene is slightly supplemented by myrcene in allspice product and carvone and octa-deceoic acid in cumin product; in coriander product the limonene supplement higher content of alpha-pinene. Limonene (carven) is mono-terpene with pineapple or orange aroma depending on chemical isomery. The main occurrence of limonene is in citrus fruits but as this paper speaks about we can find it as well in other plants. Limonene support the activity of liver enzymes especially glutation S-tranferase that is tool of cancer development elimination. About anti-cancer activities speak authors e. g. Rabi and Bishayee, 2009. Anti-oxidative activities of limonene describe Lado, 2004, etc. Wagner and Elmadfa (2003) speak about carvone and its biofunctionality in positive sense (anti-carcinogenic, anti-oxidative, insecticide).
Material and methods The two – balance and feed - experiments run in frame of this project. In these experiments was used same feed mixture based on wheat, barley, high quality soya extract meal, mineral-vitamin additive, lysine additive, rapeseed fat and experimental treatment (0,5 %). Wheat meal was used as experi-mental treatment in control group. The by-products of cumin, allspice and coriander were added into feed mixture in rate 0.5 % as treatments B, C, D. The feed mixture was identical in nutrient composi-tion. The gas-chromatography analyses demonstrate the main secondary metabolites content (Fig. 1).
Fig. 1. – Chromatograms – experimental feed additives.
Balance experiment that did show effect of experimental treatments on digestibility of main nutrients (crude protein, energy, crude fat, crude fibber, ash, organic matter). Seven male pigs entered the experiment and during seven balance periods were the experimental feed mixture compared with

Mareš et al.: Spice Industry By-products in Pig Nutrition
Seite 172 8. BOKU-Symposium Tierernährung 2009
negative control. The feed mixture intake was semi-restricted and all pigs were fed by unified amount that increased between subsequent balance periods.
Feed experiment entered 80 male and female piglets (20 piglets in each group). Ratio between male and female piglets was 10:10 in all experimental groups. Young piglets one weak after weaning with average initial body weight 6.5 kg was used and the experiment started after one weak period in ex-perimental technology. The experimental feed mixture and water were fed ad-libitum for 28 days.
Results and discussion The balance experiment showed (Table 1.) that experimental treatment does not affect significantly the digestibility of monitored nutrients. It mean the selected level of feed additive content in feed mixture do not decrease the utilization of nutrients from feed mixture as well as there is no positive effect on this matter. We can compare this work with results of authors Pradeep and Gervani (1994) which speak about decreasing of digestibility coefficient when mixture of herbs including coriander was fed to laboratory rats. As well Cross et al. (2007) say that different herbs additives do not affect digestibility but the grow intensity was influenced.
Table 1. The digestibility (%) of main nutrients during balance experiment
Digestibility (%) Group A control Group B cumin Group C allspice Group D coriander
Dry matter 84,77 ± 1,67 84,73 ± 2,26 84,42 ± 1,06 84,23 ± 1,98
Crude protein 84,64 ± 1,48 84,63 ± 1,60 83,66 ± 0,85 83,99 ± 1,84
Energy 84,78 ± 1,54 84,40 ± 2,10 84,20 ± 1,02 84,14 ± 1,83
Crude fat 59,69 ± 5,70 61,42 ± 6,52 56,38 ± 8,72 59,05 ± 2,16
Crude ash 46,28 ± 9,88 49,39 ± 10,54 48,34 ± 6,15 45,59 ± 10,29
Crude fibber 34,46 ± 11,73 32,53 ± 11,22 34,06 ± 8,01 28,74 ± 11,44
In second experiment the growth intensity and feed conversion were monitored. Comparing with con-trol we can see statistical significance (P>0.9) in increasing of average daily gain in group B (cumin feed additive). As well other experimental treatment leads to the increasing of growth intensity with-out any statistical significance. The life body weight of animals was on the end of experiment was higher of 15.9, resp. 4.2 and 1.5 % comparing with control group which has average final weight on level of 15,94 kg. The feed mixture utilization has similar tendency. We can see improving of feed utilization mainly in second experimental group (feed conversion 2.34 kg feed/kg gain) in relation with the conversion that was found out in control group (2.91 kg feed/kg gain). The positive effect can be found in elimination of diarhea, anti-oxidative properties etc. that is described for example in work of Jabeen et al. (2009).
Table 2. Growth intensity of experimental animals
Group Initial weight (kg) Final weight (kg) Average daily gain (kg/animal/day)
A 6,46 ± 1,08 15,94 ± 4,06 0,34 ± 0,12 B 6,68 ± 0,90 18,48 ± 2,93 0,42 ± 0,08 C 6,40 ± 1,01 16,62 ± 2,87 0,37 ± 0,09 D 6,36 ± 1,18 16,18 ± 3,59 0,35 ± 0,09

Mareš et al.: Spice Industry By-products in Pig Nutrition
8. BOKU-Symposium Tierernährung 2009 Seite 173
Table 3. Feed utilization during experiment
Group Average daily feed intake (kg/animal/day)
Feed conversion (kg of feed/kg gain)
1/feed conversion (kg gain/kg feed)
A 0,987 2,91 0,34
B 0,987 2,34 0,43
C 1,038 2,84 0,35
D 0,987 2,81 0,36
Acknowledgement
Research project was supported by grant of Ministry of Agriculture Czech Republic under notification No. NAZV QG 60118
References
Rabi, T., Bishayee, A., 2009: d -Limonene sensitizes docetaxel-induced cytotoxicity in human prostate cancer cells: Generation of reactive oxygen species and induction of apoptosis. In Journal of Carcinogenesis. 8; p. 9. ISSN 1477-3163.
Wagner, K., H., Elmadfa, I., 2003: Biological relevance of terpenoids. Overview focusing on mono-, di- and tetraterpenes. In Annals of Nutrition and Metabolism. 47(3-4); p 95-106; ISSN: 0250-6807.
Lado, C., Then, M., Varga, I., Szoke, E., Szentmihályi, K., 2004: Antioxidant property of volatile oils determined by the ferric reducing ability. In Zeitschrift für Naturforschung. 59 (5-6); p 354-358; ISSN: 0341-0382.
Cross, D. E., Mc. Dewitt, R. M., Hillman, K., Acamovic, T., 2007: The Effect of Herbs and Their Associated Essentials Oils on Performance, Dietetary Digestibility and Gut Microflora in Chicken from 7 to 28 Days of Age. In British Poultry Science: Aug. 48 (4): s. 496 – 506. ISSN 0007 - 1668
Pradeep, K. U., Geervani, P., 1994: Influence of spices on protein utilization of Winter bean (Psophocarpus tetragonolobus) and horsefram (Dolichus biflorus) In Plant Foods Hum Nutr: Oct, 46 (3): s. 187 – 193. ISSN 0921-9668
Jabeen, Q., Bashir, S., Lyoussi, B., Gilani, A. H., 2009: Coriander fruit exhibits gut modulatory, blood pressure lowering and diuretic activities. Journal of Ethnopharmacology; 25; 122(1); p. 123-130; ISSN 0378-8741.
Corresponding author
Ing. Petr Mareš Mendel University of Agriculture and Forestry Brno Agronomy Faculty Department of Animal Nutrition and Forage Production Zemedelska street No. 1 613 00 BRNO E-mail: [email protected] Phone: 00 420 545 133 166

Reisinger et al.: Effects of essential oils on broiler performance and intestinal physiology during a coccidial vaccine challenge
Seite 174 8. BOKU-Symposium Tierernährung 2009
Effects of essential oils on broiler performance and intestinal physiology during a coccidial vaccine challenge
Nicole Reisinger1, Tobias Steiner2, Sabine Nitsch2, Gerd Schatzmayr1, Todd J. Applegate3
1 Biomin Research Center, Tulln, Austria
2 Biomin Holding GmbH, Herzogenburg, Austria
3 Purdue University, West Lafayette, IN, USA
Abstract A 2 x 2 experiment (8 pens / treatment, 26 birds / pen) was conducted with 2 doses of a coccidial vaccine (1X or 5X; Paracox® 5 administered at 1 d of age) with or without supplementation (125 g/1000 kg) with a phytogenic feed additive containing essential oils from oregano, anis and citrus peel (Biomin® P.E.P. 125 poultry). Within each pen, 13 birds received the coccidial vaccine while the re-mainder was naturally infected through recycling of oocysts in the litter. The higher (5X) Paracox® 5 dosage decreased broiler bodyweight (BW) up to 27% (P=0.08), but not FCR and mortality at 27 d of age (P > 0.05). Between d 14 and 27 within birds receiving the 1x Paracox® dosage, those that were fed the phytogenic additive had a 5.6% increased BW gain (P < 0.05). Supplementation with the phy-togenic additive did not affect feed intake or FCR (P > 0.05). Birds naturally infected in pens receiving the higher (5X) dosage of the coccidial vaccine had an 11% reduction in crypt depth (P < 0.05), but there was no effect on villus length or number of goblet cells (P > 0.05). The dosage of coccidial vac-cine had no effect on the apparent digestibility of energy or nitrogen. At Paracox® 1X dosage, birds fed the phytogenic feed additive had 12% longer villi than the control fed birds (P < 0.05) with 30% more goblet cells, and 23% more goblet cells per 10 microns villius length (P < 0.05). Birds fed the phytogenic feed additive at a Paracox® 5X dosage had 4% less apparent digestible energy (P < 0.05), however at the Paracox® 1X dosage there was no effect of the phytogenic feed additive (P > 0.05). In conclusion, the dosage of coccidial vaccine had an impact on BW and crypt depth, whereas dietary supplementation with the phytogenic feed additive during a coccidial vaccine challenge increased villi length and goblet cell density.
Introduction Plant-derived feed supplements have been shown to improve broiler performance (Windisch et al., 2008) and reduce clinical symptoms associated with coccidial infections (Applegate, 2009). Substantial focus has been brought to find alternatives to chemical and ionophore coccidostats as the EU is plan-ning to ban most of these feed additives. Phytogenic feed additives such as essential oils have a range of documented effects including growth promotion, modulation of the immune system, and therefore may help the bird cope with coccidial infection.
Knowledge about physiologic mechanisms associated with plant-derived supplements alleviating clini-cal and sub-clinical symptoms of coccidial infections and improved growth are limited. Plant-derived supplements, such as carvacrol, cinnamaldehyde or capsicum oleoresin, have been noted to have an effect on stimulating production and secretion of mucin in the intestine, thereby possibly impairing the adhesion of pathogens and having a positive effect on the gut of chicken (Jamroz et al, 2006). Further

Reisinger et al.: Effects of essential oils on broiler performance and intestinal physiology during a coccidial vaccine challenge
8. BOKU-Symposium Tierernährung 2009 Seite 175
benefits include influences on the nutrient digestibility, nutrient absorption, intestinal morphology, and a stabilizing effect on the intestinal microbiota (Windisch et al, 2008).
The objective of the present study was to evaluate the effect of a phytogenic feed additive containing essential oils from oregano, anis and citrus peel (Biomin® P.E.P. 125 poultry) during a mild coccidal vaccine (Paracox®5) challenge (direct and indirect exposure) on broiler performance, intestinal mor-phology, mucin production, and nutrient digestibility.
Experimental Design The study was designed as a 2 x 2 factorial experiment with the following treatments:
Paracox®5 dose Biomin® P.E.P. 125 poultry (PEP)
1 x none 1 x 125 g/1000 kg 5 x none 5 x 125 g/1000 kg
For this experiment, 32 pens with 26 birds per pen were used (8 replicate pens per treatment). Half of the broilers within a pen received Paracox at treatment dosage level. The non-vaccinated birds had a natural exposure to coccidia due to shedding of oocysts from the infected broilers. Paracox5® is a commercial vaccine, which contains Emeria acervulnia, E. maxima, E. mitis and E. tenella.
The phytogenic supplement was based on essential oils from oregano, anis and citrus peel (Biomin®
P.E.P. 125 poultry) and fructo-oligosaccharides.
Performance from 0-14 (starter) and 14-27 d (grower) of age was measured. 5 birds per pen (non-vaccinated) at 28 d of age were sacrificed for ileal digesta samples (apparent digestible energy, nitro-gen determination) and 2 birds per pen were injected (27 d of age) with 5-bromo-2’-deoxyuridine (BrdU, 10 mg/kg BW), and mid-ileum was collected for histological evaluation at 3 and 24 h post-injection.
Results The higher (5X) Paracox® 5 dosage decreased broiler BW (Figure 1), but the Paracox® dosage did not affect FCR. The effects of dietary supplementation with the phytogenic feed additive on bird perfor-mance during a coccidial vaccine challenge were numerical but not significant due in part to pen-to-pen variation. However, within the birds receiving the 1X Paracox® dosage, those receiving the phyto-genic feed additive gained 5.6% more (P < 0.05).
Birds fed PEP at a Paracox® 5X dosage had 4% less apparent digestible energy, which, however, did not affect performance. At Paracox® 1X dosage there was no effect of the phytogenic feed additive (P > 0.05). The dosage of coccidial vaccine had no effect on the apparent digestibility of energy or nitro-gen.
Birds naturally infected in pens receiving the higher (5X) dosage of the coccidial vaccine had an 11% reduction in crypt depth (P < 0.05), but there was no effect on villus length or number of goblet cells (P > 0.05). At Paracox® 1X dosage, birds fed the phytogenic feed additive had 12% longer villi than the control fed birds (P < 0.05) (Figure 2) with 30% more goblet cells, and 23% more goblet cells per 10 microns villius length (P < 0.05) (Figure 3).
Birds exposed to the 5X Paracox® dosage had a 1.3 day shorter intestinal turnover vs. 1X (5.4 vs 6.7 days, respectively) (P = 0.086), but there was no effect of the phytogenic feed additive.

Reisinger et al.: Effects of essential oils on broiler performance and intestinal physiology during a coccidial vaccine challenge
Seite 176 8. BOKU-Symposium Tierernährung 2009
Figure 1: BW at 27 days (Paracox P=0.08; PEP P = 0.29; Paracox*PEP P = 0.16)
Figure 2: Broiler morphology at day 27 (Crypt depth: Paracox P=0.05; PEP P = 0.03)
Figure 3: Goblet cell number per villus and goblet cells per 10 microns villus length.

Reisinger et al.: Effects of essential oils on broiler performance and intestinal physiology during a coccidial vaccine challenge
8. BOKU-Symposium Tierernährung 2009 Seite 177
Discussion Supplementation with the phytogenic feed additive (Biomin® P.E.P. 125 poultry) affected intestinal characteristics of broilers during a coccidial vaccine challenge and had minor effects on bird perfor-mance. The most pronounced physiological effects of the phytogenic supplement were noted on num-bers of goblet cells in the mid-ileum, which may provide more of a barrier against pathogens. The 5X recommended dosage of coccidial vaccine caused slight reduction in ileal crypts and increased mucos-al turnover rate, coinciding with a 3% reduction in 27 d BW.
References
Applegate, T.J. 2009. Influence of Phytogenics on the Immunity of Livestock and Poultry. In: Phytogenics in Animal Nutrition. Edit. T. Steiner. Nottingham University Press, Nottingham, United Kingdom. ISBN: 978-1-904761-71-6.
Jamroz, D., T. Wertelecki, M. Houszka, and C. Kamel. 2006. Influence of diet type on the inclusion of plant origin active substances on morphological and histochemical characteristics of the stomach and jejunum walls in chicken. J. Anim. Physiol. Anim. Nutr. (Berl.) 90:255–268.
Reisinger N., T. Steiner, S. Nitsch, G. Schatzmayr and T.J. Applegate.2009. Effects of plant-derived essential oils on broiler performance and intestinal morphology during a mild coccidial vaccine challenge. Poultry Science (Submitted)
Windisch W, Schedle K, Plitzner C and Kroismayr A (2008) Use of phytogenic products as feed additives for swine and poultry. J.Anim. Sci. 86:E140-E148
Corresponding author
Nicole Reisinger BSc. BIOMIN Holding GmbH BIOMIN Research Center Technopark 1 3430 Tulln, Austria E-mail: [email protected]

Firmensponsoring – BIOMIN GmbH, Industriestraße 21, A-3130 Herzogenburg
Seite 178 8. BOKU-Symposium Tierernährung 2009

Steiner et al.: Effects of Phytogenics on intestinal parameters and growth performance of broilers
8. BOKU-Symposium Tierernährung 2009 Seite 179
Effects of Phytogenics on intestinal parameters and growth per-formance of broilers
Tobias Steiner1, Jia-Chi Hong2, Tu-Fa Lien2 1 BIOMIN GmbH, Herzogenburg, Austria 2 National Chiayi University, Chiayi-City, Taiwan
Abstract A 42-d feeding experiment was carried out in order to determine the effects of Phytogenics and Anti-biotics on intestinal parameters and growth performance of broilers. In total 240 one-day old, male and female Arbor Acres broilers were randomly assigned to 3 treatments with 4 replicates (20 birds/replicate) each. The treatments were: (1) Control, (2) Antibiotics (50 ppm chlorotetracycline and 50 ppm oxytetracycline), (3) Phytogenics (125 ppm Biomin® P.E.P. 125 poultry). Basal starter and finisher diets were based on corn and soybean meal. Compared to the Control, villus height in the duodenum was numerically higher (P > 0.05) in birds fed Phytogenics or Antibiotics. There were no treatment effects (P > 0.05) on pH values in the duodenum, ileum, jejunum or cecum. Total concen-tration of volatile fatty acids was numerically lower (P > 0.05) in birds fed Phytogenics or Antibiotics, and ammonia concentration in the feces was reduced (P < 0.05) through Phytogenics and Antibiotics. In comparison with the Control, Antibiotics increased (P < 0.05) feed intake and weight gain, but did not affect (P > 0.05) feed conversion ratio (FCR). FCR was reduced (P < 0.05) by supplementation with Phytogenics. Overall FCR was 1.71, 1.60 and 1.69 in birds fed the Control diet, Phytogenics or Antibiotics, respectively. Feed intake and weight gain did not differ (P > 0.05) between the Control and Phytogenics treatment. Survival rate was increased (P < 0.05) by Phytogenics and Antibiotics. However, weight gain did not differ between birds fed Antibiotics and Phytogenics. In conclusion, supplementation of diets with Phytogenics affected intestinal parameters in broilers and improved feed conversion in broilers.
Introduction Recently, antibiotics residues and disease resistance have created public concern towards the usage of antibiotic growth promoters in the animal industry. “Natural” products including probiotics, prebiotics, enzymes, organic acids and plant extracts or their synthetic chemical principles have been used to as replacements for in-feed antibiotics. Essential oils may be used as feed additives to improve growth performance under intensive rearing systems and have demonstrated their biological functions such as antimicrobial and antioxidant activities (Lee and Ahn, 1998). In addition, dietary supplementation of essential oils has a beneficial effect on intestinal microflora (Helander et al., 1998) and the activity of digestive enzymes (Lee et al., 2003; Jang et al., 2004).
The objective of this trial was to evaluate the effect of a phytogenic additive based on essential oils and fructooligosaccharides in comparison with antibiotics on intestinal parameters and growth per-formance of broilers.

Steiner et al.: Effects of Phytogenics on intestinal parameters and growth performance of broilers
Seite 180 8. BOKU-Symposium Tierernährung 2009
Materials and Methods In total 240 one-day old, male and female Arbor Acres broilers were randomly assigned to 3 treat-ments with 4 replicates (20 birds/replicate) each. The treatments were:
(1) Control (2) Antibiotics (50 ppm chlorotetracycline and 50 ppm oxytetracycline) (3) Phytogenics (125 ppm Biomin® P.E.P. 125 poultry)
Birds were fed according to a two-stages feeding program, using a starter and finisher diet from day 1 to 21 and 21 to 42, respectively. The formulation and composition of the basal diets are shown in Table 1.
Table 1. Formulation and calculated chemical composition of basal diets Ingredients Starter
(Day 1-21) Finisher (Day 21-42)
--------------------------- % -------------------------- Formulation Corn, yellow 51.80 61.01 Soybean meal 31.04 23.97 Fish meal 7.00 5.00 Soybean oil 6.53 6.35 Salt 0.30 0.50 Dicalcium phosphate 1.71 1.39 Limestone, pulverized 0.45 0.66 Vitamin premix1 0.50 0.50 Mineral premix2 0.50 0.50 DL-Methionine 0.17 0.12 Calculated composition Crude protein, % 23 19 ME, kcal/kg 3200 3200 Calcium, % 1.0 0.9 Available phosphorus, % 0.45 0.35 1 Vitamin premix (content per kg): vitamin A, 12,500,000 IU; vitamin D3, 2,500,000 IU; vitamin E, 20,000 IU; vitamin K3, 2,500 mg; vitamin B1, 2,000 mg; vitamin B2, 5,000 mg; vitamin B6, 3,000 mg; vitamin B12, 12 mg; niacin, 35,000 mg; pantothenic acid, 12,000 mg; folic acid, 1,000 mg 2 Mineral premix (content per kg): Fe, 70 g; Zn, 90 g; Cu,10 g; Mn, 80 g; Se, 15 g; I, 0.4 g
Intestinal parameters
About 2 cm of duodenum, jejunum and middle ileum were placed in neutral formalin solution (includ-ing formalin [37-40%] 100 mL, distilled water 900 mL, NaH2PO4•H2O 4.0 g, NaHPO4•H2O 6.5 g) for 24 hours, followed by paraffin-embedded and hematoxylin purple-eosin staining. Each slice was selected 10 points for measuring the layer thickness, ileum villus height, villus area, villus perimeter, crypt depth and height of cecal mucosa and muscle thickness with a microscope.

Steiner et al.: Effects of Phytogenics on intestinal parameters and growth performance of broilers
8. BOKU-Symposium Tierernährung 2009 Seite 181
Intestinal content collection
Intestinal content was collected on the 4th, 5th and 6th week of experiment from the duodenum, jeju-num, ileum and cecum separately to provide material for the determination of intestinal volatile fatty acids (VFA), ammonia, primary amines and tissue examination.
Intestinal pH was determined immediately after sampling using a pH meter (Digital pH meter, Goodly, Taiwan). Total VFA content was determined following the method described by Parker and McMillan (1976) using 5 g cecal content.
For the determination of ammonia and primary amine content, samples were pre-treated: To the solid sample distilled water was added to 100 mL, mixed with 1 mL Na2SO3 (7.14 m M) and 1 mL ZnSO4•7H2O (0.348 M) in the samples and the standard solutions to precipitation. After filtration, liq-uid was taken and adjusted to pH 10.5. For measuring ammonia content, a 2 mL sample was used. To this sample 10 µL sodium tartrate (0.177 M) and 100 µL NaOH (6M) were added. NH4Cl was used as standard. 0.1 mL Nessler reagent was added for 1 min (0.02 M HgCl2 + 0.1 M KI , added 6 M NaOH to 100 mL), then ammonia content was measured using a spectrophotometer (Hitachi, U-2000, To-kyo, Japan) at a wavelength of 330 nm. To determine the content of primary amines, a standard curve was generated using Butylamine stock solution 740 μg/mL by dilution 2-fold thereafter. A 1 mL sample was mixed with 0.1 mL Borate buffer (0.5M,pH 10.8) and 0.9 mL OPA/NAC reagent (1:1, each 8.8 mM), then primary amine content was measured by a spectrophotometer (Hitachi, U-2000, Tokyo, Japan) at a wavelength 400 nm.
Results and Discussion Compared to the Control, villus height in the duodenum was numerically higher (P > 0.05) in birds fed Phytogenics or Antibiotics (Table 2).
Table 2. Effect of Antibiotics and Phytogenics on histological parameters of broilers Control Antibiotics Phytogenics S.E.M. Duodenum Villus height (μm) 1341.5 1383.9 1467.3 29.5 Crypt depth (μm) 206.2 240.5 244.2 7.5 Jejunum Villus height (μm) 857.0 919.9 886.6 23.4 Crypt depth (μm) 155.1 166.0 164.5 5.7 Ileum Villus height (μm) 553.6 479.6 553.3 12.9 Crypt depth (μm) 130.9 118.6 136.6 4.6
There were no treatment effects (P > 0.05) on pH values in the duodenum, ileum, jejunum or caecum (Table 3). Total concentration of volatile fatty acids was numerically lower (P > 0.05) in birds fed Phy-togenics or Antibiotics, and ammonia concentration in the faeces was reduced (P < 0.05) through Phytogenics and Antibiotics.
Performance parameters are shown in Table 4. In the starter period, birds fed Antibiotics were signifi-cantly (P < 0.05) heavier than birds in the other treatments. In the grower period, there was no dif-ference between treatments, probably due to higher variation within groups. Overall BW gain of birds fed Phytogenics were numerically (P > 0.05) between the Control and Antibiotics treatment. In all

Steiner et al.: Effects of Phytogenics on intestinal parameters and growth performance of broilers
Seite 182 8. BOKU-Symposium Tierernährung 2009
stages, feed intake was significantly (P < 0.05) enhanced in broilers fed Antibiotics. However, overall feed conversion in this treatment did not differ from the Control. In contrast, FCR was significantly improved (P < 0.05) in birds fed Phytogenics. Survival rate was increased (P < 0.05) through both Antibiotics and Phytogenics.
Table 3. Effect of Antibiotics and Phytogenics on intestinal parameters of broilers Control Antibiotics Phytogenics S.E.M. pH-value Duodenum 5.73 5.67 5.82 0.040 Jejunum 5.51 5.62 5.71 0.050 Ileum 5.50 5.51 5.93 0.120 Cecum 6.45 6.13 6.46 0.080 Total VFAs* (µmole/g) 102.54 86.67 88.95 3.470 Ammonia (M*) 0.256a 0.190b 0.200b 0.003 a, b, c Means within the same row with different superscripts differ significantly (P < 0.05). *VFA: Volatile Fatty Acids;M: Molar
Table 4. Effect of antibiotics and phytogenics on performance of broilers Age (days) Control Antibiotics Phytogenics S.E.M. Average daily weight gain (g/bird)
Starter (d 0-21) 710.92b 801.91a 746.34b 5.93 Finisher (d 21-42) 1319.00 1489.25 1386.75 31.37 Overall (d 0-42) 2030.05b 2291.18a 2133.10ab 32.13
Average daily feed intake (g/bird)
Starter (d 0-21) 1045.10b 1118.07a 1030.70b 11.23 Finisher (d 21-42) 2419.91b 2760.94a 2369.22b 51.76 Overall (d 0-42) 3465.61b 3879.01a 3399.92b 52.93
Feed conversion ratio (feed/gain)
Starter (d 0-21) 1.47b 1.39a 1.38a 0.006 Finisher (d 21-42) 1.84b 1.85b 1.71a 0.02 Overall (d 0-42) 1.71b 1.69b 1.60a 0.008
Survival rate (%) 88.75b 97.50a 98.75a 1.35 a, b Means within the same row with different superscripts differ significantly (P < 0.05).
Conclusion Supplementation of diets with Antibiotics or Phytogenics affected intestinal parameters, improved livability and increased growth performance in broilers. For Antibiotics the improvement in perform-ance was mainly due to increased weight gain, whereas for Phytogenics, better performance was achieved mainly through improved feed conversion.

Steiner et al.: Effects of Phytogenics on intestinal parameters and growth performance of broilers
8. BOKU-Symposium Tierernährung 2009 Seite 183
References
Helander M, Alakomi H, Latva-Kala K, Mattila-Sandholm T, Pol I, Smid EJ, Gorris LGM and Wright AV (1998) Characterization of the action of selected essential oil components on Gram-negative bacteria Journal of Agricultural and Food Chemistry 46 3590–3595.
Jang IS, Ko YH, Yang HY, Ha JS, Kim YI, Kang SY, Yoo DH, Nam DS, Kim DH and Lee CY (2004) Influence of essential oil components on growth performance and the functional activity of the pancreas and small intestine in broiler chickens. Asian-Australasian Journal of Animal Science 17: 394–400.
Lee HS, Ahn YJ (1998) Growth-inhibiting effects of Cinnamomum cassia bark-derived materials on human intestinal bacteria. Journal Agricultural Food Chemistry 46: 8–12.
Lee KW, Everts H, Kappert HJ, Frehner M, Losa R and Beynen AC (2003) Effects of dietary essential oil components on growth performance, digestive enzymes and lipid metabolism in female broiler chickens. British Poultry Science 44: 450–457.
Parker DC and McMillan RT (1976) The determination of volatile fatty acids in the caecum of the conscious rabbit. British Journal of Nutrition 35:365–371.
Corresponding author
Dr. Tobias Steiner BIOMIN Holding GmbH Industriestrasse 21 3130 Herzogenburg Austria E-mail: [email protected]

Jerešiūnas et al.: Comparable assessment of the alternative zinc oxide growth promoters, organic acids and probiotics in the rations of weaned piglets
Seite 184 8. BOKU-Symposium Tierernährung 2009
Comparable assessment of the alternative zinc oxide growth promoters, organic acids and probiotics in the rations of weaned piglets
A. Jerešiūnas1, J. Kulpys1, G. Sudikas1, O. Jerešiūnienė2
1 Lithuanian veterinary academy 2 Public institution Abromiškės Rehabilitation Hospital
Introduction Feed and its quality have a great influence on piglet productivity and their health. Feed constitutes approximately 70% of all animal breeding production expenses. Therefore, feed composition for pigs has a main economic role in pig breeding.
Pig weaning is a strong and stressful factor which has a negative influence on both pig productivity and their immunity (Simon, 2001). The security of optimal pig feeding conditions is recommended within this period. Normal intestine microflora of piglets influences their wellness, formation of immu-nity and resistance to infections. The stress of weaning negatively affects the population of bifidobac-teria by decreasing their number in the intestine of piglets. Precisely the change of microflora in the intestine of piglets determines their diseases, decrease of productivity as well as deaths. The decrease of intestine microflora (Bifidobacteria, Enterococcus) population of piglets at the time of stress leads to the pathogenic microorganism increase. The security of stable microflora in the intestine of piglets after weaning is one of the main factors determining their health and productivity.
The guarantee of stable microflora in the intestine was feeding antibiotics for a long time. Currently antibiotics can be used only for the treatment. Other feed supplements replaced antibiotic growth stimulants, i.e. organic acids (acidifiers), probiotics and zinc oxide. Regardless that Zink oxide has negative effect to the environment, it has been used in Lithuania underground yet in the feeding of weaned piglets as the antibiotic alternative.
A review of the scientific literature suggest that, in general, alternative additives provide little consis-tent growth benefits [2,3,9,10]. Among the more promising are organic acids and probiotics.
The addition of high levels of zinc oxide to creep feed reportedly has beneficial effects on the inci-dence and severity of diarrhea and on performance in post weaning piglets. Other reports indicate tendency toward improved weight gain or no effects [1].
Organic acids are considered a promising alternative to replace antibiotic growth promoter in animal nutrition (Gauthier, 2005). Several reports have confirmed that organic acids effectively increase growth performance in different animal species. As reviewed by Freitag et al. (1999), supplementation of diets pigs and poultry may improve weight gain and feed efficiency by 3-5 and 2-10 %, respec-tively.
Effects of supplementation with probiotics on performance parameters in pigs were summarized in a review by Simon (2005). It can be derived from this review that, in most cases, supplementation with probiotics tended to improve average daily gain and feed efficiency. However, improvements in these parameters were significant only in 7 or 5%, respectively, of the experiments that were reviewed.
The aim of the work will be to ascertain the influence of zinc oxide, BASF Formi acidifier and probiotic BioPlus 2B on the growth, feed consumption and health of the weaned piglets.

Jerešiūnas et al.: Comparable assessment of the alternative zinc oxide growth promoters, organic acids and probiotics in the rations of weaned piglets
8. BOKU-Symposium Tierernährung 2009 Seite 185
Material and methods The experiment was conducted with the crossbreeds of Lithuanian Great white (mother) and Norwe-gian Landrass and Yorkshire (father) in the pig farm located in central region of Lithuania in 2008. The weaned piglets of the age of 30-35 days will be selected for the experiment. On the grounds of ana-logical principal considering the breed, gender, age and weight, three groups of 40-50 weaned pigs in each will composed. The duration of the experiment aimed at 31 days in total. During the experiments the piglets of both groups were fully fed by the dry powder compound feed from automatic feeders. Automatic separate water-troughs were applied in order to get enough water. Every stall was equipped with one automatic water-trough and feeder.
The piglets from the first group were given the compound feed with the 4 kg per one zinc oxide, the piglets from second group were given the feed with BASF Formi acidifier and the piglets from the third group were given the feed with probiotic BioPlus 2B. Biochemical composition and nutritive value of feeds are presented in Table 1.
Table 1. Composition and nutritive value of compound feeds
Item Group I
Group II Group III
Wheat % 46,2 46,6 46,6 Barley % 30,0 30,0 30,0 Soyaschrot % 7,5 7,5 7,5 Fish meal % 2,0 2,0 2,0 Soy protein concentrate AK 530 agro-korn % 7,5 7,5 7,5
Monocalciumphosphate % 0,6 0,6 0,6 Canola oil % 2,0 2,0 2,0 Mycofix Plus 0,1 0,1 0,1 Mineral Vitamin Supplement % 3,7 3,7 3,7 Zinc oxide (Zn 75%) % 0,4 - - BASF Formi, kg/t - 10 - BioPlus 2B (Bacillus licheniformis and Bacillus subtilis), g/t
- 400
Analytical data, 1 kg: metabolizable energy – 13 MJ, crude protein – 17,8%, fibre – 3,13 %, fat – 3,81 %, calcium – 1,07%, phosphorus – 0,74%, sodium – 0,28%, lysine – 0,93%, methionine+cistine – 0,63%, threonine – 0,80%, triptophane – 0,22 %
In order to ascertain mass growth, the piglets were weighed at the beginning of the experiment and at the end of the experiment. The consumption of feeds was determined during the experiments as well as record of feeds was kept.
In order to ascertain piglets’ health condition at the beginning of the experiment and at the end of experiment, the main data of the research conducted, except the indices of chemical feed composition and consumption, were assessed by the methods of statistical analysis. The reliability of statistical data was estimated according to the criterion t. The data are regarded reliable on the grounds of the Student when p<0,05 (Juozaitienė and Kerzienė, 2001). The computer programme used for the data processing was Microsoft Excel.

Jerešiūnas et al.: Comparable assessment of the alternative zinc oxide growth promoters, organic acids and probiotics in the rations of weaned piglets
Seite 186 8. BOKU-Symposium Tierernährung 2009
Results and discussion The trial results are presented in Table 2. It was estimated that during the trial the piglets from II group, fed by the compound feed with acidifier, gained 116 g weight per day on average or 29,9% more (p>0,05) than the piglets from the first group and gained 104 g weight per day on average or 26% more (p>0,05) than the piglets from the second group.
Table 2. Trial results
Item Group 1
Group 2 Group 3
No of piglets at the start of trial 51 41 51 No of piglets at the end of trial 51 41 50 Average daily weight gain, g/day 388 504 400 Compound feed consumption per kg gain
1,69 1,63 1,72
Feed intake kg/day/pig 0,655 0,640 0,690 Dozes price of preparation €/t 9,9 15,5 4,7
After the calculation of the feed consumption per kg gain it was estimated that BASF Formi Acidifier influence this index. The II group piglets of 1kg weight gain consumed 3,6% less feed than the I group piglets and 5,2% less than the II group piglets.
The research results show that the best results are achieved when the weaned piglets were feeding with acidifier. The use of probiotic is also useful. Thus, the piglets of the 3rd group put on 22 g of weight per day or 3,1% more than the piglets of the 1st group. The results of the experiment also show that the use of probiotic in the piglets feeding increases the feed consumption per kg gain. The 3rd group piglets for 1 kg weight gain consumed 1,8% and 5,5% compound feeds more than the pig-lets of the 1st and the 2nd groups respectively.
The experiment results show that probiotic has a great influence on the feed intake per day. Thus, BioPlus 2 B supplement increased the daily feed intake up to 5,3% and 7,8% respectively in compari-son with the results of the 1st and 2nd groups. Positive influence of probiotic upon the feed intake is proved by other authors [5,8,9].
During the experiment no health disorders were identified within the groups. Zink oxide and Acidifier did not have any essential influence upon the mortality of the piglets during the experiment, which continued 31 day. However, during the experiment one piglet from the 3rd group died. We could not identify the reason of the death. We still could not deny the opinion that Zink oxide influences posi-tively the weaned piglets. Taking into account the literature information [1,4,7,11] one can state that Zink oxide betters the piglets’ state of health after the weaning. However, according to the data of our experiment, Acidifier influences positive upon the piglets’ state of health.
Basing upon the results of the conducted experiment one can state that the use of Zink oxide in the feeding of the piglets is not reasonable from the economic point of view. Our results show that probi-otic increases minimally the price of feed. The price is mainly increased by BASF Formi. However, the use of this preparation gives the best results of the piglets’ growth.
So the illegal use of Zink oxide in several Lithuanian pig farms is an anarchism, which should be extir-pated from the experience as it has neither zoo technical nor economic benefits, but pollutes the envi-ronment.

Jerešiūnas et al.: Comparable assessment of the alternative zinc oxide growth promoters, organic acids and probiotics in the rations of weaned piglets
8. BOKU-Symposium Tierernährung 2009 Seite 187
Conclusion 1. Acidifiers and probiotics can guarantee good results of the piglets’ growth.
2. The use of Zink oxide in the fodders of the piglets does not increase the growth and has no eco-nomic benefit.
3. The use of Basf Formi increases the average weight per day of the piglets up to 29,9% in compari-son with the piglets fed with Zink oxide.
4. Probiotic BioPlus 2B increases the piglets’ average weight per day only up to 3,1%, however its’ use is economically more profitable and it increases the feed intake per day up to 5,3%.
References
1. Broom L., Miller H. Et al. (2006). Effects of zink oxide and Enterococcus faecium SF68 dietary supplementation on the performance, intestinal microbiota and immune status of weaned piglets. Research in Veterinary Science. 80(1): 45-54.
2. Freitag, M., Hensche, H.U., Schulte-Siencbeck, H. and Reichelt, B. (1999). Biologische Effekte konventioneller und alternativer Leistingsforderer. Kraftfutter/Feed Magazine 2: 49-57.
3. Gauthier, R. (2005). Organic acids and essential oils, a realistic alternative to antibiotic growth promoters. I. Forum Interna-cional de Avicultura, August 17-19 2005, Foz do Iguacu, PR, Brasil, 148-157.
4. Hojberg O., Canibe N. et al. (2005). Influence of dietary zinc oxide and copper sulfate on the gastrointestinal ecosystem in newly weaned piglets. Applied and Environmental Microbiology. 71 (5): 2267-2277.
5. Jeroch H., Drochner W., Simon O. (1999). Ernährung landwirtschaftlicher Nutztiere. Stuttgard, Ulmer, 90-91; 352-354.
6. Juozaitienė V., Kerzienė S. (2001): Biometrija ir kompiuterinė duomenų analizė. Kaunas. 114.
7. Ryselis S., Baranauskienė D., Abdrachmanovas O., Naginienė R., Stepaniukas A., Šernienė L. (2007). Sulfato, chlorido, cinko ir kadmio jonų poveikis 5-aminolevulino rūgšties dehidratazės aktyvumui bandomųjų gyvūnų kraujyje in vitro. Veterinarija ir zootechnika. ISSN 1392-2130. T. 39 (61), 53-59.
8. Simon, O. (2001). Probiotika aus der Sicht der Tierernährung. Vitamine und Zusatzstoffe in der Ernährung von Mensch und Tier. 8. Symposium 26. und 27. September 2001, Jena/Thüringen, 118-127.
9. Simon, O. (2005). Mikroorganismen als Futterzusatzstoffe: Probiotika – Wirksamkeit und Wirkungsweise. 4 Boku-Symposium Tierernährung: Tierernährung ohne Antibiotische Leistungsförderer. Vienna, Austria, 10-16.
10. Ulbrich M., Hoffmann M., Drochner W. (2004) Fütterung und Tiergesundheit. Stuttgard, Ulmer, 90-91.
11. Zentek J., Hellweg P., Khol-Parisini A. (2007). Spurenelemente als Faktoren der Tiergesundheit. 6. BOKU-Symposium Tierernährung: Sekundärwirkungen von Futterinhaltsstoffen - vom Nährstoff zum Wirkstoff. Vienna, Austria, 42-46.
Corresponding author
Dr. Andrejus Jerešiūnas Lithuanian Veterinary Academy Animal Nutrition Department Tilžės street 18, Kaunas LT-47181 E-mail: [email protected]

Roth et al.: Antibacterial activity of formic and propionic acids against Salmonella and E. coli and their influence on broiler performance
Seite 188 8. BOKU-Symposium Tierernährung 2009
Antibacterial activity of formic and propionic acids against Sal-monella and E. coli and their influence on broiler performance
Nataliya Roth1, Thomas Germ2, Alfred Klimitsch2, Renata Urbaityte1, Sabine Nitsch1
1 BIOMIN Holding GmbH, Industriestrasse 21, 3130 Herzogenburg, Austria 2 BIOMIN Holding GmbH, BIOMIN Research Center, Technopark 1, 3430 Tulln, Aus-
tria
Abstract The inhibitory effect of acids on bacterial growth has long been used to preserve feed from spoilage. The use of organic acids as acidifiers for swine and poultry is also well known. In the present study the antimicrobial activities of formic and propionic acids and their combination were investigated in vitro on E. coli and Salmonella typhimurium, as these pathogens cause problems in animal production. The combination of acids was more effective than the effect of individual acids together. The reason of the synergy between organic acids blends can be the combination of acid with a low pKa, which reduces the pH of the environment of bacterial cells and acid with a high pKa and low molecular weight to ensure an antimicrobial effect inside the cell.
The efficacy of the combination of formic and propionic acid (Biotronic® SE forte) in vivo was investi-gated on the performance of broilers in Center of Applied Animal Nutrition, Austria. Body weight gain (BWG) and feed conversion ratio (FCR) of broiler chicks were significantly (P<0.001) improved by the supplementation of a blend of acids at the dosage 3 kg/t of feed.
Introduction Organic acids have been used to ensure water hygiene and to reduce contamination in feed. Organic acids exhibit two different modes of action with respect to their antimicrobial properties. The first is a reduction of the pH value in the environment, which is achieved by dissociation of the acid. The lower pH level slows down microbial replication. The second model of antibacterial activity is the direct an-timicrobial property of the undissociated acid molecule, which is able to penetrate the bacterial cell and dissociate there. This results in disturbance of the ionic balance inside the cell by the H+ and the disruption of DNA synthesis by the anion. As a result replication of bacterial cells is disturbed (Lambert and Stratford, 1999).
With respect to antimicrobial activity of organic acids there are two important parameters: the mole-cular weight (MW) and the pKa value (pH at which 50% of acid is in undissociated form). The lower the MW the higher is the amount of H+ and the stronger is the ability of the acid to reduce the pH level of the environment. Organic acids with antimicrobial activity have a pKa value between 3 and 5. The higher the pKa value the larger is the part of undissociated acid and the better is the antimicrobial activity of the acid.
It is generally well-known that acidifiers improve animal performance. Acids start their work by reduc-ing microbial load, buffering capacity and the pH level in the feed. The level of reduction depends a lot on the acid used because each acid has its own specific range of efficacy. By incorporating differ-ent acids, there is a broader spectrum of activity and their combination is considerably more effective.

Roth et al.: Antibacterial activity of formic and propionic acids against Salmonella and E. coli and their influence on broiler performance
8. BOKU-Symposium Tierernährung 2009 Seite 189
The influence of acidifiers in the upper intestinal tract of animals has been investigated and described extensively in the scientific literature. The pH reducing effect of acidifiers in the stomach supports protein digestion (Eidelsburger et al., 1992; Overland et al., 2000). A change in the upper digestive tract influences the lower digestive tract, and in case of acidifiers this means a reduction of pathogen loads, improved protein digestion and a slowdown in the emptying of the gut which leads to improved digestibility and lower diarrhea rates (Kluge et al., 2004). In addition, acids reduce the buffering ca-pacity of the feed, which also improves nutrient digestibility.
Formic and propionic acids are used as feed acidifiers. The efficacy of the individual acids and their combination was investigated in vitro against E. coli and Salmonella and in vivo on broiler perfor-mance.
Material and methods
In vitro trial
Formic acid, propionic acid and their blend were used to determine the antibacterial effect of acids on pathogenic microorganisms [Salmonella typhimurium (Bio 60) and Escherichia coli O55:K59 (B5) (Bio 114)]. Each acid as well as the combination were diluted to 0.05% and then used for the micro plate assay. According to the used microorganisms, nutrient broth was used for S. typhimurium and E. coli O55:K59 (B5). The media were prepared according to the manual instructions.
An overnight culture in the appropriate medium was prepared. For this 10 ml of the medium was in-oculated with 100 µl bacterial suspension and incubated at 37°C overnight. To ensure that the growth was in the exponential phase, the culture was reincubated for an additional four hours the next day.
The optical density (OD) was adjusted to 0.1 at a wave length of 690 nm. Pure medium served as zero value.
The micro plates (96 well) were prepared under sterile conditions. The first row was filled with bacte-rial suspension (which was adjusted before to an OD of 0.1) and served as positive control (PC). The second row was filled with pure medium and appears as negative control (NC). 100 µl pure medium was put forward starting from row D till the last row. Afterwards 200 µl of a 2% acid dilution of each acid and their blend were poured into the wells of row C. The test series were performed in triplicates. The plates were incubated at 37°C for 24 hours. The OD was measured photometrically at 620 nm using a micro plate reader. To be sure that there is no contamination in the negative control the plates were checked after incubation. After the measurement the blank values were subtracted from the test values. The positive control served as optimum (100%) value (medium + substrate without growth inhibition).
In vivo trial on the broiler grow th performance
The trial was conducted in the Center of Applied Animal Nutrition (CAN), Austria. A number of 354 one-day old mixed sex broiler chickens Ross 308 were divided into two groups with 177 birds each. Each application has been repeated 8 times with 25 chicks per replication. Both groups received the same commercial broiler diet. The experimental diets were fed for 35 days. The control group re-ceived no growth promoting feed additives while the experimental group received 0.3% commercial acidifier Biotronic® SE forte produced by Biomin Holding GmbH with the proportion of formic and pro-pionic acids 1:1. Feeding was done manually twice a day. The trial feed was given to the chicks ad libitum and their need for water was supplied through nipple drinkers. The feed composition is shown in the Table 1.

Roth et al.: Antibacterial activity of formic and propionic acids against Salmonella and E. coli and their influence on broiler performance
Seite 190 8. BOKU-Symposium Tierernährung 2009
Table 1. The composition of broiler diet (g/kg)
Ingredients Starter (0-14 days) Grower (14-35 days) Corn 579.3 597.5
Soybean 312.5 296.0
Soybean oil 25.0 20.0
Plant oils and fats 12.5 25.0
Premix for broiler 62.5 60.0
L-Lysin 3.8 1.5
DL-Methionin 0.8 -
Threonin 1.3 -
Limestone 2.5 -
Total 1000.2 1000.0
Nutrients
ME-G, MJ 12.65 12.96
Crude protein, g 208.05 197.37
Crude fat, g 65.2 73.18
Crude fiber,g 25.43 25.23
Crude ash, g 75.57 68.68
Methoinin, g 5.49 4.59
Met + Cys, g 9.02 8.02
Lysine, g 14.06 11.78
Threonine, g 9.27 7.77
Tryptophan, g 2.44 2.34
Ca, g 11.96 11.10
P, g 8.47 7.68
Na, g 2.02 1.94
Birds were kept on wood shavings. Climate conditions as well as a lighting program were computer operated and regulated according to the breeding company’s standard recommendations.
All animals were controlled by the responsible veterinarian at the beginning of the trial, at day 21 (Salmonella-control) and at the end of the trial. Throughout the whole trial period all animals re-mained healthy and free of clinical disease symptoms and no medication was necessary.
Clinical observation was done twice a day and all incidents recorded. Weight of the individual animal was measured and recorded at the beginning and at the end of the trial (day 35). Average daily feed intake (ADFI) and mortality were measured and recorded. Average daily weight gain (ADWG), feed conversion ratio (FCR) as well as the EBI (European Broiler Index = Liveability [%] x Live weight [g]/age [d] / FCR x 10) were calculated. All data generated out of the trial was subjected to statistical analysis by means of ANOVA (SPSS® statistics 17.0).

Roth et al.: Antibacterial activity of formic and propionic acids against Salmonella and E. coli and their influence on broiler performance
8. BOKU-Symposium Tierernährung 2009 Seite 191
Results and discussion The results of the present study showed that the inhibitive effect of formic acid was lower than pro-pionic acid. The pH level of media was neutral; therefore propionic acid had stronger antimicrobial activity due to higher pKa value. Antibacterial activity of formic and propionic acids was better due to the use of the combination of these two acids. The reason for the synergy between formic and pro-pionic acids can be the combination of acid with a low pKa, which reduces the pH of the environment of the bacteria and acid with a high pKa and low molecular weight to ensure an antimicrobial effect inside the cell.
The inhibition level of formic, propionic acids and their blends on E. coli is shown in Figure 1 and on Salmonella thyphimurium in Figure 2. The acid concentration of all tested materials was on the same level.
Figure 1. Inhibition of E.coli by formic, propionic acids and their combination
Figure 2. Inhibition of Salmonella thyphimurium by formic, propionic acids and their combination
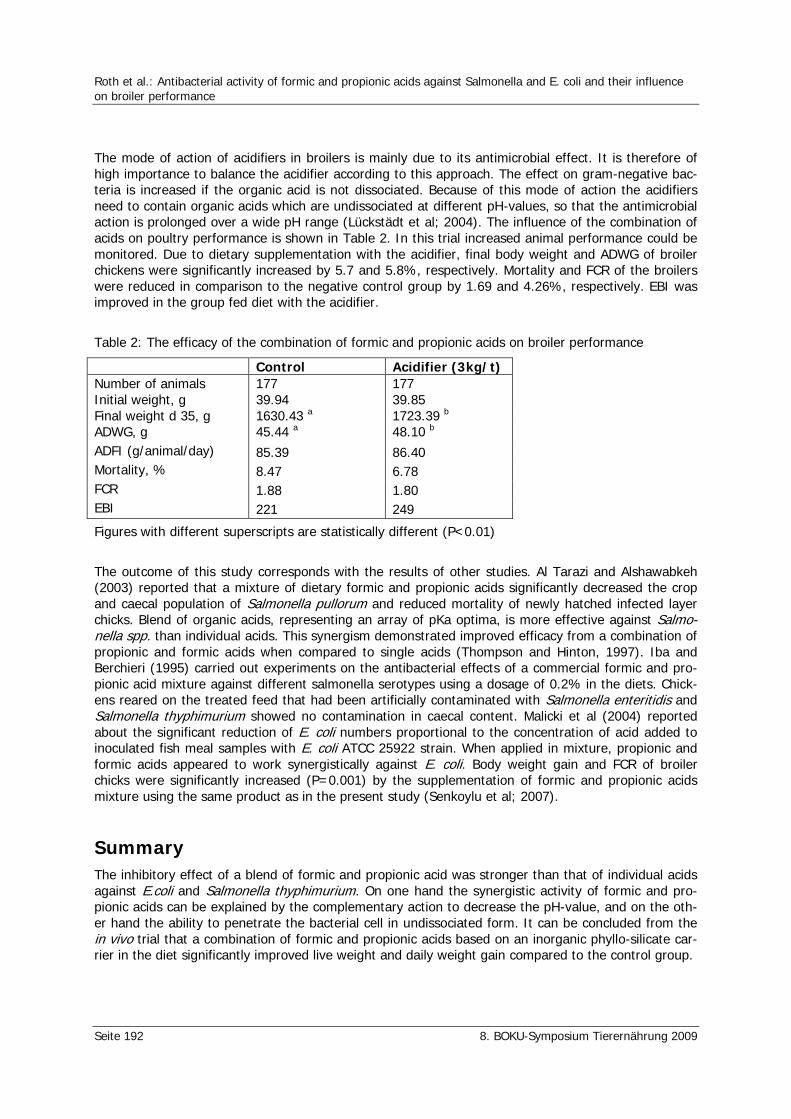
Roth et al.: Antibacterial activity of formic and propionic acids against Salmonella and E. coli and their influence on broiler performance
Seite 192 8. BOKU-Symposium Tierernährung 2009
The mode of action of acidifiers in broilers is mainly due to its antimicrobial effect. It is therefore of high importance to balance the acidifier according to this approach. The effect on gram-negative bac-teria is increased if the organic acid is not dissociated. Because of this mode of action the acidifiers need to contain organic acids which are undissociated at different pH-values, so that the antimicrobial action is prolonged over a wide pH range (Lückstädt et al; 2004). The influence of the combination of acids on poultry performance is shown in Table 2. In this trial increased animal performance could be monitored. Due to dietary supplementation with the acidifier, final body weight and ADWG of broiler chickens were significantly increased by 5.7 and 5.8%, respectively. Mortality and FCR of the broilers were reduced in comparison to the negative control group by 1.69 and 4.26%, respectively. EBI was improved in the group fed diet with the acidifier.
Table 2: The efficacy of the combination of formic and propionic acids on broiler performance
Control Acidifier (3kg/t) Number of animals 177 177 Initial weight, g 39.94 39.85 Final weight d 35, g 1630.43 a 1723.39 b ADWG, g 45.44 a 48.10 b ADFI (g/animal/day) 85.39 86.40 Mortality, % 8.47 6.78 FCR 1.88 1.80 EBI 221 249
Figures with different superscripts are statistically different (P<0.01)
The outcome of this study corresponds with the results of other studies. Al Tarazi and Alshawabkeh (2003) reported that a mixture of dietary formic and propionic acids significantly decreased the crop and caecal population of Salmonella pullorum and reduced mortality of newly hatched infected layer chicks. Blend of organic acids, representing an array of pKa optima, is more effective against Salmo-nella spp. than individual acids. This synergism demonstrated improved efficacy from a combination of propionic and formic acids when compared to single acids (Thompson and Hinton, 1997). Iba and Berchieri (1995) carried out experiments on the antibacterial effects of a commercial formic and pro-pionic acid mixture against different salmonella serotypes using a dosage of 0.2% in the diets. Chick-ens reared on the treated feed that had been artificially contaminated with Salmonella enteritidis and Salmonella thyphimurium showed no contamination in caecal content. Malicki et al (2004) reported about the significant reduction of E. coli numbers proportional to the concentration of acid added to inoculated fish meal samples with E. coli ATCC 25922 strain. When applied in mixture, propionic and formic acids appeared to work synergistically against E. coli. Body weight gain and FCR of broiler chicks were significantly increased (P=0.001) by the supplementation of formic and propionic acids mixture using the same product as in the present study (Senkoylu et al; 2007).
Summary The inhibitory effect of a blend of formic and propionic acid was stronger than that of individual acids against E.coli and Salmonella thyphimurium. On one hand the synergistic activity of formic and pro-pionic acids can be explained by the complementary action to decrease the pH-value, and on the oth-er hand the ability to penetrate the bacterial cell in undissociated form. It can be concluded from the in vivo trial that a combination of formic and propionic acids based on an inorganic phyllo-silicate car-rier in the diet significantly improved live weight and daily weight gain compared to the control group.

Roth et al.: Antibacterial activity of formic and propionic acids against Salmonella and E. coli and their influence on broiler performance
8. BOKU-Symposium Tierernährung 2009 Seite 193
References
Al Tarazi Y.H. and K. Alshawabkeh 2003. Effect of dietary formic and propionic acids on Salmonella pullorum shedding and mortality in layers chicks after experimental infection. Journal of Veterinaty Medicine. 50 (3):112-117.
Eidelsburger U. and M. Kirchgessner. 1994. Zum Einfluß organischer Säuren und Salze im Futter auf die Mastleistung von Broilern. Archiv für Geflügelkunde 58: 268-277.
Iba A.M. and A.Berchieri. 1995. Studies on the use of a formic acid - propionic acid mixture to control experimental Salmonella infection in broiler chickens. Avian Pathology 24: 303-311.
Kluge H., Broz J. and K. Eder. 2004. Studies on the influence of benzoic acid as a feed additive on growth performance, digestibility of nutrients, nitrogen balance, microflora and parameters of the microbial metabolism in the gastrointestinal tract of weaned piglets. 8. Tagung für Schweine und Geflügelernährung Halle (saale) Germany: 42-45.
Lambert R.J. and M. Stratford 1999. Weak acid preservatives: modeling microbial inhibition and response. Journal of Applied Microbiology 86: 157-164.
Lückstädt C., Senköylü N., Akyürek H. and A. Agma. 2004. Acidifier - a modern alternative for anti-biotic free feeding in livestock production, with special focus on broiler production. Veterinarija ir Zootechnika. ISSN 1392-2130. T.27 (49)
Malicki A., Zawadzki W., Bruzewicz S., Graczyk S. and A. Czerski 2004. Effect of formic and propionic acid mixture on Escherichia coli in fish meal stored at 12°C. Pakistan Journal of Nutrition 3(6): 353-357.
Overland M., Granli T., Kjos N.P., Fjetland O., Steien S.H. and M. Stokstad. 2000. Effect of dietary formates on growth performance, carcass traits, sensory quality, intestinal microflora and stomach alterations in growing-finishing pigs. Journal of Animal Science 78: 1875-1884.
Senkoylu N., Samli H.E., Kanter M. and A. Agma. 2007. Influence of a combination of formic and propionic acids added to wheat- and barley- based diets on the performance and gut histomorphology of broiler chickens. Acta Veterianaria Hungarica 55(4): 479-490
Thompson J.L. and M. Hinton. 1997. Antibacterial activity of formic and propionic acids in the diet of hens on salmonellas in the crop. British Poultry Science 38 (1):59
Corresponding author
Dr. Nataliya Roth BIOMIN Holding GmbH Industriestrasse 21 3130 Herzogenburg, Austria Tel: +43 2782 803 314 Mobile: +43 664 8254 318 Fax: +43 2782 803 40 E-mail: [email protected]

Wetscherek et al.: Einsatz von organischen Säuren in der Ferkelaufzucht
Seite 194 8. BOKU-Symposium Tierernährung 2009
Einsatz von organischen Säuren in der Ferkelaufzucht
Wolfgang Wetscherek, Elisabeth Rohrer und Gabriela Wetscherek-Seipelt
Universität für Bodenkultur Wien, Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie
Einleitung Der Einsatz von organischen Säuren in der Fütterung von Ferkeln wird in vielen Versuchen mit positi-ven Effekten zur Verhinderung von Verdauungsstörungen in der Absetzphase beschrieben.
Die organischen Säuren senken die Pufferkapazität des Futters und reduzieren somit auch den pH-Wert im Magen. Das führt zur verbesserten Eiweißverdauung und somit zu einer verringerten Anrei-cherung unverdauter Proteine im Verdauungstrakt. Weiters wirkt ein tieferer pH-Wert im Magen als Barriere gegen Mikroorganismen bzw. wird durch organische Säuren die mikrobielle Aktivität im Ver-dauungstrakt vermindert. Nimmt die mikrobielle Stoffwechseltätigkeit ab, wird das Tier weniger durch bakterielle Toxine (biogene Amine, Ammoniak), die wachstumsdepressiv wirken können, belastet.
In dieser Untersuchung wurde der Einsatz von verschiedenen Mischungen von organischen Säuren auf die Futteraufnahme, Aufzuchtleistung und den Gesundheitsstatus von Ferkeln im Vergleich zu einer negativen Kontrollgruppe geprüft.
Material und Methoden Aus den Ferkeln von 8 Zuchtsauen wurden beim Absetzen mit etwa 28 Lebenstagen 48 Ferkel ausge-wählt. Bei der Aufteilung auf die vier Versuchsgruppen wurde auf eine gleichmäßige Verteilung von Geschlecht, Wurf und Gewicht geachtet. Der Versuch begann nach der 4. Lebenswoche mit etwa 9,5 kg Lebendgewicht und endete nach 7 Versuchswochen mit einem Lebendgewicht von etwa 33 kg. Während der Versuchsperiode wurden die Tiere in 8 strohlosen Boxen mit Spaltenböden gehalten.
Die Zusammensetzung der Futtermischungen der drei Versuchsgruppen unterschied sich nur durch den Zusatz von einer unterschiedlichen organischen Säuremischung der Firma Dr. Eckel GmbH.
Gruppe 1: Anta cid LFA ist eine Mischung aus Milch- und Ameisensäure Gruppe 2: Anta cid LFAS ist eine Mischung aus Milch-, Ameisen-, Essig- und Sorbinsäure Gruppe 3: Anta cid LFA Coa ist eine Mischung aus Milch-, Ameisen-, Essig-, Fumar- und Zitronen-säure
Die jeweiligen Dosierungen waren in der Absetz- und Aufzuchtperiode gleich und betrugen 8g/kg. In der negativen Kontrollgruppe (Gruppe 4) wurde nur das Trägermaterial eingemischt. Die Umstellung von der Absetzfuttermischung auf die Aufzuchtfuttermischung erfolgte nach zwei Versuchswochen. Der Versuchsplan ist in der Tabelle 1 dargestellt.
Die Rezepturen der Basismischungen für die Ferkelabsetz- und Ferkelaufzuchtphase werden in den Tabellen 2 und 3 dargestellt. Die wichtigsten Nährstoffgehalte sind in der Tabelle 4 aufgelistet.

Wetscherek et al.: Einsatz von organischen Säuren in der Ferkelaufzucht
8. BOKU-Symposium Tierernährung 2009 Seite 195
Tabelle 1 Versuchsplan
Gruppe 1 Gruppe 2 Gruppe 3 Gruppe 4 Anta cid
LFA Anta cid
LFAS Anta cid LFA Coa
Neg. Kontrolle
8 g / kg 8 g / kg 8 g / kg 4 g / kg Trägermateri-al
2 Boxen 2 Boxen 2 Boxen 2 Boxen 6 Ferkel / Box 6 Ferkel / Box 6 Ferkel / Box 6 Ferkel / Box
Box 1 + 5 Box 2 +6 Box 3 +7 Box 4 +8
Tabelle 2 Zusammensetzung der Basisfuttermischung des Absetzfutters
Futtermittel Anteil in % Mais 22,67 Triticale 15,00 Erbsen 12,00 Sonnenblumenkuchen 10,30 Gerste 10,00 Sojabohnen getoastet 8,90 Molkenpulver- süß 6,25 Kartoffeleiweiß 5,90 Molkeneiweißpulver- 45% 3,00 Mineralstoffprämix S 2,01 Rübenmelasse 2,00 Melasseschnitte 12% 1,50 Kohlensaurer Kalk „Grieß“ 0,30 Monocalciumphosphat Aliphos 22,7% 0,11 Natriumchlorid 0,06
Datenerhebung Die Ferkel wurden bei der Einstellung und danach jede Woche gewogen. Die pro Box und Abschnitt gefressenen Futtermengen wurden wöchentlich ermittelt und notiert. Die Relation der je Box und Ab-schnitt aufgenommenen Futtermengen zur Summe der Lebendgewichtszunahmen der Tiere je Box wurde berechnet. Behandlungen, Ausfälle und Krankheiten wurden mit Tiernummer, Datum, Gewicht und Ursache festgehalten.
Auftretende Durchfälle wurden nach dem nachstehenden 3- Punkteschema beurteilt.
• Leichter Durchfall: In der Box ist sehr weicher Kot zu finden, Tiere sind nicht ver-schmutzt und machen keinen kranken Eindruck, sondern sind sehr lebendig und auf-geweckt.
• Mittlerer Durchfall: In der Box ist flüssiger Kot zu finden, Tiere sind nicht ver-schmutzt und machen keinen kranken Eindruck, sondern sind lebendig und aufge-weckt.
• Schwerer Durchfall: In der Box ist flüssiger Kot zu finden, Tiere sind verschmutzt und machen einen kranken Eindruck.

Wetscherek et al.: Einsatz von organischen Säuren in der Ferkelaufzucht
Seite 196 8. BOKU-Symposium Tierernährung 2009
Tabelle 3 Zusammensetzung der Basisfuttermischung des Aufzuchtfutters
Futtermittel Anteil in % Gerste 40,57 Mais 20,00 Sojaextraktionsschrot HP 17,83 Weizen 10,67 Weizenkleie 5,00 Futterfett 1,40 Monocalciumphosphat 1,10 Calciumcarbonat 0,67 L-Lysin 0,47 Viehsalz 0,40 Fischöl 0,35 Lignocellulose 0,30 L-Threonin 0,14 DL-Methionin 0,10 Prämix 1,00
Tabelle 4 Errechnete wertbestimmende Nährstoffgehalte der Basismischungen
Energie- und Nährstoffgehalt Ferkelabsetzfutter Ferkelaufzuchtfutter ME, MJ/kg 13,0 13,4 Rohprotein, % 19,7 17,5 Rohfett, % 4,8 3,8 Rohfaser, % 5,0 3,8 Rohasche, % 5,9 5,2 Stärke, % 34,2 40,0 Zucker, % 8,4 3,7 Lysin, % 1,11 1,2 Methionin, % 0,35 0,4 Methionin und Cystin, % 0,70 0,7 Threonin, % 0,84 0,7 Vitamin A, IE/kg 6800 16.000 Vitamin D3, IE/kg 1100 2.000 Vitamin E, mg/kg 65 160 Kupfer, mg/kg 6 12
Bei der Aufzuchtleistung wurden der Gruppen- und der Geschlechtseinfluss mittels zweifacher Vari-anzanalyse unter der Berücksichtigung der Wechselwirkung von Gruppe und Geschlecht berechnet. Neben den Gruppenmittelwerten werden die Ergebnisse der biometrischen Auswertung als P-Wert angegeben. Ein P-Wert von 0,05 und kleiner bedeutet, dass sich die Gruppen signifikant unterschei-den. Gruppen, welche sich signifikant unterscheiden, werden mit unterschiedlichen hochgestellten Buchstaben gekennzeichnet.

Wetscherek et al.: Einsatz von organischen Säuren in der Ferkelaufzucht
8. BOKU-Symposium Tierernährung 2009 Seite 197
Versuchsergebnisse Der Nährstoffbedarf war bei den vier Gruppen durch die Futtermischungen gedeckt und wird in der Tabelle 5 und 6 dargestellt. Die Energie- und Nährstoffgehalte unterschieden sich zwischen den Grup-pen nicht wesentlich.
Zu Versuchsbeginn traten Durchfallsprobleme auf, die in Folge zu einem Ausfall in der Gruppe 2 am Beginn der 2. Versuchswoche führten. Die Durchfallsprobleme wurden nicht mit Medikamenten be-handelt. Eine genaue Darstellung wird in der Tabelle 7 gegeben.
Für die statistische Auswertung wurde jeweils ein Tier der Gruppe 2 und ein Tier der Gruppe 3 mit einem geringen Gesamttageszuwachs von 369 g bzw. 332 g ausgeschlossen, weil diese mehr als 2 Standardabweichungen unter dem Gruppenmittelwert lagen.
Tabelle 5 Analysenergebnisse Absetzfuttermischungen
V e r s u c h s g r u p p e Merkmal 1 2 3 4 Trockensubstanz g/kg 891 891 884 880 Rohprotein g/kg 192 193 191 190 Rohfett g/kg 45 45 45 46 Rohfaser g/kg 42 42 43 43 Rohasche g/kg 60 60 57 55 Stärke g/kg 376 375 374 375 Zucker g/kg 66 67 66 63 ME MJ/kg 13,46 13,44 13,32 13,30
Tabelle 6 Analysenergebnisse Aufzuchtfuttermischungen
V e r s u c h s g r u p p e Merkmal 1 2 3 4 Trockensubstanz g/kg 885 886 886 888 Rohprotein g/kg 168 168 165 163 Rohfett g/kg 36 36 36 34 Rohfaser g/kg 35 36 35 36 Rohasche g/kg 57 58 58 58 Stärke g/kg 433 427 429 431 Zucker g/kg 39 39 39 39 ME MJ/kg 13,41 13,35 13,38 13,31

Wetscherek et al.: Einsatz von organischen Säuren in der Ferkelaufzucht
Seite 198 8. BOKU-Symposium Tierernährung 2009
Tabelle 7 Durchfallsbeobachtungen
Gruppe 1 2 3 4 Box 1 5 2 6 3 7 4 8
10.02.2009 - ++ - ++ - ++ - - 11.02.2009 - ++ - ++ - ++ - - 12.02.2009 ++ +++ ++ +++ ++ +++ ++ ++
+ leichter Durchfall ++ mittlerer Durchfall +++ schwerer Durchfall
Die Gruppe 1 (Anta cid LFA) erreichte im Vergleich zur negativen Kontrollgruppe in der Absetzpha-se (Versuchswoche 1 und 2) tendenziell bessere Tageszunahmen. Auch in den beiden darauf folgen-den Versuchswochen erreichte diese Versuchsgruppe um 7,5% tendenziell höhere Tageszunahmen. In der 5. Versuchswoche erreichte diese Gruppe signifikant bessere Tageszuwächse als die Gruppen 2 und 3. In den folgenden Versuchswochen waren keine Vorteile dieser Gruppe im Vergleich zur negati-ven Kontrollgruppe mehr feststellbar. Betrachtet man den gesamten Versuchszeitraum, erreichte die Gruppe 1 um etwa 3,5% tendenziell höhere Tageszunahmen im Vergleich zur negativen Kontrollgrup-pe. Der Futterverbrauch und die Futterverwertung wurden durch den Futterzusatzstoff nicht beeinf-lusst.
Die Gruppe 2 (Anta cid LFAS) erreichte im Vergleich zur negativen Kontrollgruppe in der Absetz-phase (Versuchswoche 1 und 2) tendenziell bessere Tageszunahmen. Auch in den beiden darauf fol-genden Versuchswochen erreichte diese Versuchsgruppe um 6,5% tendenziell höhere Tageszunah-men. Im Durchschnitt der drei letzten Versuchswochen sowie über den Zeitraum der gesamten Ver-suchsperiode lagen die Tageszunahmen auf gleichem Niveau wie die der negativen Kontrollgruppe. Die Futteraufnahme lag etwas unter den Werten der negativen Kontrollgruppe. Dies führte bei etwa gleichen Zunahmen zu einer um etwa 3% geringeren Futterverwertung.
Die Gruppe 3 (Anta cid LFA Coa) erreichte im Vergleich zur negativen Kontrollgruppe in der Ab-setzphase (Versuchswoche 1 und 2) tendenziell bessere Tageszunahmen. Die tendenziell positiven Effekte in den beiden Folgewochen in der Höhe von etwa 3% waren tendenziell geringer als in den beiden anderen Versuchsgruppen. In den letzten beiden Versuchswochen lagen die Tageszunahmen auf gleicher Höhe, in der 5. Versuchswoche tendenziell unter dem Niveau der negativen Kontrollgrup-pe. Betrachtet man den gesamten Versuchszeitraum, erreichte die Gruppe 3 nahezu die gleichen Ta-geszunahmen wie die negative Kontrollgruppe. Die Futteraufnahme war im Vergleich zu den anderen Gruppen am niedrigsten. Dies führte bei etwa gleichen Zunahmen zu einer um etwa 6 % geringeren Futterverwertung im Vergleich zur negativen Kontrollgruppe.
Insgesamt zeigten die drei Versuchsgruppen tendenziell positive Effekte in den ersten beiden Ver-suchswochen nach dem Absetzen. In den folgenden 2 Wochen verflachte sich der positive Effekt. Wobei hier die Gruppen 1 und 2 besser abschnitten als die Gruppe 3. In der 5. Versuchswoche wurde eine signifikante Überlegenheit der Gruppe 1 gegenüber den Gruppen 2 und 3 registriert. Betrachtet man die Durchschnittswerte der letzten drei Versuchswochen, so lag die Gruppe 1 auf annähernd gleichen Niveau wie die negative Kontrollgruppe. Die Gruppen 2 und 3 schnitten um etwa 2% schlech-ter ab. Insgesamt war die Gruppe 1 mit einem tendenziellen Plus von 3,5% gegenüber der negativen Kontrollgruppe bezüglich der Tageszunahmen am Besten. Die beiden anderen Versuchsgruppen erziel-ten ähnliche Ergebnisse wie die negative Kontrollgruppe.
Bezüglich der Futteraufnahme gab es geringere Werte für die Gruppen 2 und 3, welche zu einer Ver-besserung der Futterverwertung um 3 bzw. 6% führten.
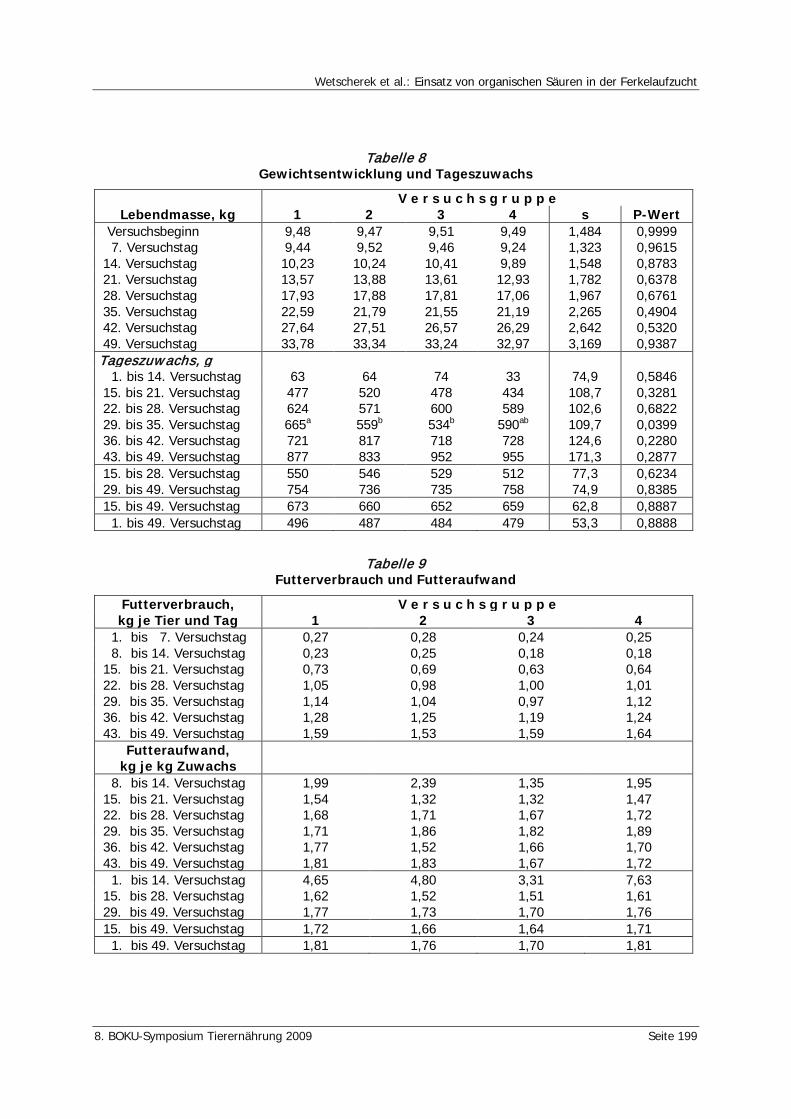
Wetscherek et al.: Einsatz von organischen Säuren in der Ferkelaufzucht
8. BOKU-Symposium Tierernährung 2009 Seite 199
Tabelle 8 Gewichtsentwicklung und Tageszuwachs
V e r s u c h s g r u p p e Lebendmasse, kg 1 2 3 4 s P-Wert
Versuchsbeginn 9,48 9,47 9,51 9,49 1,484 0,9999 7. Versuchstag 9,44 9,52 9,46 9,24 1,323 0,9615 14. Versuchstag 10,23 10,24 10,41 9,89 1,548 0,8783 21. Versuchstag 13,57 13,88 13,61 12,93 1,782 0,6378 28. Versuchstag 17,93 17,88 17,81 17,06 1,967 0,6761 35. Versuchstag 22,59 21,79 21,55 21,19 2,265 0,4904 42. Versuchstag 27,64 27,51 26,57 26,29 2,642 0,5320 49. Versuchstag 33,78 33,34 33,24 32,97 3,169 0,9387 Tageszuwachs, g 1. bis 14. Versuchstag 63 64 74 33 74,9 0,5846 15. bis 21. Versuchstag 477 520 478 434 108,7 0,3281 22. bis 28. Versuchstag 624 571 600 589 102,6 0,6822 29. bis 35. Versuchstag 665a 559b 534b 590ab 109,7 0,0399 36. bis 42. Versuchstag 721 817 718 728 124,6 0,2280 43. bis 49. Versuchstag 877 833 952 955 171,3 0,2877 15. bis 28. Versuchstag 550 546 529 512 77,3 0,6234 29. bis 49. Versuchstag 754 736 735 758 74,9 0,8385 15. bis 49. Versuchstag 673 660 652 659 62,8 0,8887 1. bis 49. Versuchstag 496 487 484 479 53,3 0,8888
Tabelle 9 Futterverbrauch und Futteraufwand
Futterverbrauch, kg je Tier und Tag
V e r s u c h s g r u p p e 1 2 3 4
1. bis 7. Versuchstag 0,27 0,28 0,24 0,25 8. bis 14. Versuchstag 0,23 0,25 0,18 0,18 15. bis 21. Versuchstag 0,73 0,69 0,63 0,64 22. bis 28. Versuchstag 1,05 0,98 1,00 1,01 29. bis 35. Versuchstag 1,14 1,04 0,97 1,12 36. bis 42. Versuchstag 1,28 1,25 1,19 1,24 43. bis 49. Versuchstag 1,59 1,53 1,59 1,64
Futteraufwand, kg je kg Zuwachs
8. bis 14. Versuchstag 1,99 2,39 1,35 1,95 15. bis 21. Versuchstag 1,54 1,32 1,32 1,47 22. bis 28. Versuchstag 1,68 1,71 1,67 1,72 29. bis 35. Versuchstag 1,71 1,86 1,82 1,89 36. bis 42. Versuchstag 1,77 1,52 1,66 1,70 43. bis 49. Versuchstag 1,81 1,83 1,67 1,72 1. bis 14. Versuchstag 4,65 4,80 3,31 7,63 15. bis 28. Versuchstag 1,62 1,52 1,51 1,61 29. bis 49. Versuchstag 1,77 1,73 1,70 1,76 15. bis 49. Versuchstag 1,72 1,66 1,64 1,71 1. bis 49. Versuchstag 1,81 1,76 1,70 1,81

Wetscherek et al.: Einsatz von organischen Säuren in der Ferkelaufzucht
Seite 200 8. BOKU-Symposium Tierernährung 2009
Abbildung 1 Vergleich des Tageszuwachses zur negativen Kontrollgruppe
über die gesamte Versuchsperiode
Tageszuwachs - Relationen zur negativen Kontrollgruppe = 100 %
80
90
100
110
120
130
140
150
160
170
180
190
200
210
220
230
240
1.-2. Woche
1.-3. Woche
1.-4. Woche
1.-5. Woche
1.-6. Woche
1.-7. Woche
Anta cid LFA Anta cid LFAS Anta cid LFA Coa Neg. Kontrolle
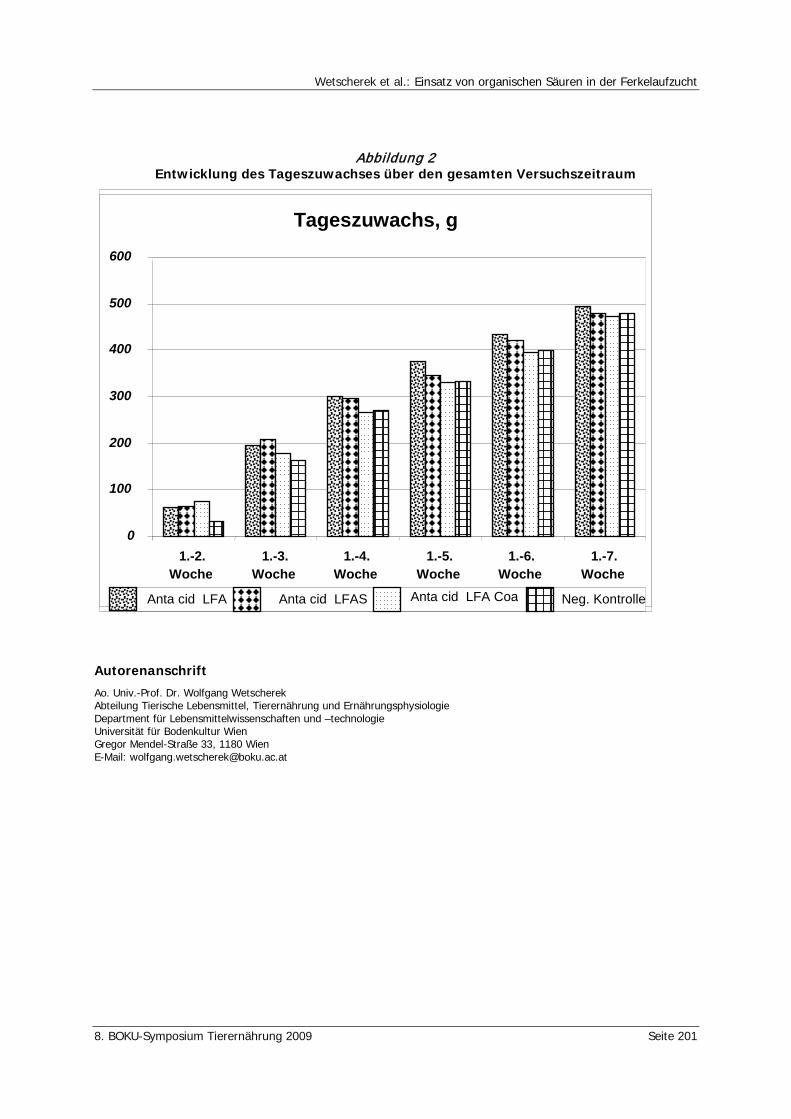
Wetscherek et al.: Einsatz von organischen Säuren in der Ferkelaufzucht
8. BOKU-Symposium Tierernährung 2009 Seite 201
Abbildung 2 Entwicklung des Tageszuwachses über den gesamten Versuchszeitraum
Autorenanschrift
Ao. Univ.-Prof. Dr. Wolfgang Wetscherek Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Department für Lebensmittelwissenschaften und –technologie Universität für Bodenkultur Wien Gregor Mendel-Straße 33, 1180 Wien E-Mail: [email protected]
Tageszuwachs, g
0
100
200
300
400
500
600
1.-2. Woche
1.-3. Woche
1.-4. Woche
1.-5. Woche
1.-6. Woche
1.-7. Woche
Anta cid LFA Anta cid LFAS Neg. Kontrolle Anta cid LFA Coa

Firmensponsoring – Dr. Eckel GmbH, Im Stiefelfeld 10, D-56651 Niederzissen
Seite 202 8. BOKU-Symposium Tierernährung 2009

Urbaityte et al.: Effect of urinary acidifier on reproduction performance in sows
8. BOKU-Symposium Tierernährung 2009 Seite 203
Effect of urinary acidifier on reproduction performance in sows
Renata Urbaityte, Diego Padoan, Nataliya Roth
BIOMIN GmbH, Industriestrasse 21, 3131 Herzogenburg, Austria
Introduction Urinary tract infections (UTI) and Postpartum Disgalactia Syndrome (PPDS) syndrome are among the most common health problems in sow herds. Both diseases are caused by bacterial infections and influence the fertility of the sow herd. From 17 to 40% of sows suffer from UTI, which consequentially leads to increased non-productive days, early cullings and decreased litters/sows/year (Saoulidis et al., 2000). The UTI include cystitis, inflammation of the urinary bladder, and pyelonephritis, ascendant inflammation into kidneys. Furthermore UTI are considered as predisposing factor for the upper geni-tal tract inflammations (oviduct and uterus), which also lead to PPDS problems. The bacteria attach to the mucosa of urogenital tract by interactions between bacterial surface adhesions and complemen-tary epithelial cell receptors, and stimulate cytokine release resulting in an inflammatory response and symptoms. However, the clinical signs of UTI are commonly overlooked in commercial farms or they are absent. Typically sows with significant bacteriuria are more often seen in the dog sitting position, have inferior body condition, longer weaning-to-service interval, low fertility rates and tend to wean small litters. Various health, environmental and management factors can contribute to UTI. For exam-ple, constipation, traumas to the urethra and bladder at parturition, assistance at farrowing, retained placenta or fluids, poor sanitized floor in farrowing crates, poor water quality, which leads to reduced water intake in sows, and ultimately reduced urination (Maes et al., 1998).
The flora of the vagina of healthy sows consists of a wide range of bacteria, including Streptococcus spp., Staphylococcus spp., Enterobacteria, Corynbacterium, Micrococcus spp. and Actinobacillus spp. Many of these bacteria especially E. coli, Actinobaculum suis, Pseudomonas aeruginosa, Klebsiella spp., Streptococcus faecalis, Streptococcus suis are frequently reported in clinical cases of urogenital tract infections. The highest isolation rates of these bacteria were obtained on the day of farrowing and the second highest number of positive samples was found immediately after mating. It is reported that parturition and a dietary change result in an upgrowth of E. coli and other bacteria in the repro-ductive tract of sows (MacLean and Thomas, 1974). The bacteria invasion is favored by acid-base unbalance in sows’ diets, bringing out an increase of urine pH. Various studies have confirmed that acid-base balance (ACB) of the diet directly correlates with urinary pH value and total bacteria colony forming units in urine (Dee et al., 1994; DeRouchey et al., 2003). The ACB of conventional sows diets have a range of +200 to +600 mmol/kg feed, which makes the urine pH greater than 7. It is known that a urinary pH below 6.5 can significantly reduce the number of bacteria excreted with urine. The alkaline environment also inhibits the growth of competitive microflora and promotes the precipitation of the urinary salts and crystals. Such precipitates not only further increase inflammatory changes in the bladder mucosa but also provide a nidus for the bacterial growth and protection from antibiotics and host defense mechanisms.
Diets supplemented with urinary tract acidifier based on a combination of natural plant extracts, inor-ganic acids and anionic substances decreases the urinary pH via lowering the ACB of the diet by means of the anionic substances and inorganic acids. Moreover, due to the mode of action of inor-ganic acid and anionic substances Ca-mobilization mechanisms is triggered resulting in the higher Ca2+ availability in the blood. High Ca2+ amounts are necessary for the uterus muscle contraction at farrow-ing, preventing laborious, long lasting farrowing and saving healthy piglets from condemnation to

Urbaityte et al.: Effect of urinary acidifier on reproduction performance in sows
Seite 204 8. BOKU-Symposium Tierernährung 2009
death. Additionally it is known that the natural plant extracts containing proanthocanidins inhibit the adhesion of pathogenic bacteria, especially E. coli to urinary tract cell walls.
In the present study, the efficacy of the urinary tract acidifier on the decrease of urine pH and repro-ductive performance in sows was evaluated.
Materials and methods A number of 48 sows were assigned to two different treatments. The control group received commer-cial gestation and lactation diets, whereas the trial group received the same gestation and lactation diets supplemented with 0.5 % of a blend of natural plant extracts, inorganic acids and anionic sub-stances (Biomin® pHD, BIOMIN, Austria) (PEx_InA_AS). The gestation diet was provided from d 108 of pregnancy until farrowing and then from d 1 weaning to d 1 mating, whereas the lactation feed from d 1 farrowing to d 1 weaning. On d 108 of gestation the sows (parities 3-4) were moved into farrowing facility and assigned to their treatments. The sows were penned individually within farrow-ing crates. The sows were allowed to access feed and water ad libitum. The conventional gestation and lactation diets were based on corn, soybean meal and rice bran, and they were formulated to contain 16.50 % crude protein.
The farrowing duration, intervals between litter mates, number of born alive, mummified, stillborn and condemned piglets, litter weight at birth and weaning were determined. The sows’ urinary pH was measured at the beginning of the trial, d 108 and d 112 of pregnancy, at farrowing, d 14 and d 21 post-farrowing and d 1 after weaning and d 1 after insemination. All urine samples were collected in the morning between 7:00 and 8:00 before feeding. The urinary pH value was measured with pH-meter (Checker® 1, Hanna Instruments) in samples taken from the mid stream urine.
Results and discussion The lower urine pH is a natural barrier against the pathogens. A urinary pH below 6.5 can significantly reduces the number of bacteria excreted with urine. The urinary pH is a function of ACB of the diet, therefore it may vary depending on the composition of the diet and time ingested. Urine samples after a meal may have an alkaline pH. The electrolyte balance greater than 350 kEq/kg results in the respi-ratory and metabolic alkalosis and in an alkaline pH in the urine (pH > 7), thus increases the risk of UTI. The alkaline urine could be caused by certain bacteria, e.g. Staphylococcus, Proteus, producing urease, which metabolize the urea into ammonia and alkalize the urine. During these infections the urinary pH might reach values between 8 to 9, which leads to the further bacterial multiplication (Su-zuki et al., 1979; Dee, 1992). The results of the present study showed that supplementation with the blend of PEx_InA_AS decreased the urinary pH ranging from 0.08 to 0.54 pH units during gestation and lactation periods (Table 1). It might be assumed that the dietary supplementation with the blend of PEx_InA_AS decreased the ACB of the diet consequentially reducing the urine pH. The highest de-crease in pH was on d 112 of gestation, following d 1 post-artificial insemination (AI) and d 14 post-farrowing. The results of the present study are in good agreement with other studies, where decrease in ACB of the diet led to reduction of the urinary pH (DeRouchey et al., 2003). The reduction of uri-nary pH at farrowing and mating is crucial, because during that time the cervix is opened, therefore the sows are more susceptible to infections. Moreover, the reduced counts of pathogenic bacteria in urogenital tract lead to healthier farrowing crates and benefit newborn piglets ultimately encountering lower counts of Streptococcus suis, E. coli after birth and before suckling the colostrum.
Additionally the plant extracts have reduced the risk of bacterial infection preventing the bacterial adhesion to the urinary tract walls. For example, the adherence of E. coli is a critical step in the pathogenesis of UTI. The bacteria been found to have specific binding sites which can attach the spe-

Urbaityte et al.: Effect of urinary acidifier on reproduction performance in sows
8. BOKU-Symposium Tierernährung 2009 Seite 205
cific receptors on urothelial cells of urinary tract wall. It is well known that, cranberry extracts contain high amounts of proanthocanidins, which prevent the attachment of E. coli to the urinary tract wall by occupying the specific binding cites for the uroepithelial cells in bacteria (Foo et al., 2000).
Table 1. Efficacy of the bled of of natural plant extracts, inorganic acids and anionic substances on sows urine pH reduction.
Collection time Control group1 Trial group2 Δ No. of sows n = 24 n = 24 108 d gestation 6.29 6.17 0.12 112 d gestation 6.42 5.88 0.54 1 d post-farrowing 6.04 5.96 0.08 14 d post-farrowing 7.61 7.35 0.26 21 d post-farrowing 7.48 7.57 0.09 1 d post-weaning 7.15 7.10 0.05 1 d post-AI3 7.35 6.94 0.41 1 Control group = no blend of natural plant extracts, inorganic acids and anionic substances 2 Trial group = with blend of natural plant extracts, inorganic acids and anionic substances 3 AI = artificial insemination
In the present study the duration of the farrowing time between the first and the last piglet was shortened by 37.16 min and the interval between the littermates was reduced by 7.87 min (P<0.10) in the group supplemented with the blend of PEx_InA_AS compared to those in the control group (Table 2). Due to supplementation of the blend PEx_InA_AS the reproduction performance of sows in the trial group was improved. In the trial group the total number and born alive were increased by 8.4 and 16.2 percentage units, respectively, whereas the number of the stillborn decreased by 26.1 per-centage units compared to the control group. The shorter farrowing period and the reduction of the stillborn number are related to the activation of the Ca-mobilization mechanism, which is triggered by supplementation with the inorganic and anionic substances.
The blend of PEx_InA_AS through the light acidification of the blood stimulates the parathyroid gland to release parathyroid hormone, which attempts to rebalance blood pH value 7.4. Parathyroid hor-mone is deputed to control Ca2+ levels, leading to enhanced Ca2+ absorption from gut lumen and its mobilization from the bones, a decrease phosphate re-adsorption in the kidneys, and less Ca2+ excre-tion through kidneys. Ca-mobilization into blood stream is important for the uterus muscle contraction. It is known that hypocalcemia decreases the strength of uterine contractions and contribute to a pro-longed farrowing, thus predisposing higher number of stillborns and lactation failure (Cline and Richert, 2001).The lack of Ca2+ at farrowing results in laborious farrowing and economic losses due to higher number of fully developed piglets born dead. The stillborns are frequent in sow units, favored mainly by low motility during gestation, stress and constipation.
Moreover, in the trial group litter weight at birth and at weaning were increased by 8.4 and 20.4 per-centage units, respectively, compared to the control group. The higher litter size at weaning and sur-vival rate in the group supplemented with the blend of PEx_InA_AS was improved (P<0.10), which is in good agreement with other studies (Saoulidis et al., 2000; DeRouchey et al. 2003). In the trial group the higher litter size and survival rates at weaning could be related to the higher amount of Ca2+ in the blood stream, which is essential for the lactogenesis (Miller et al., 1994). It is known that the high piglet mortality is associated with low milk production during the first days after the farrow-ing, because the glycogen stores decrease rapidly 12 to 18 h. Thus improvement of milk yield and milk composition during the 1st week of lactation improved vigor and survivability of piglets.

Urbaityte et al.: Effect of urinary acidifier on reproduction performance in sows
Seite 206 8. BOKU-Symposium Tierernährung 2009
Table 2. Efficacy of the bled of of natural plant extracts, inorganic acids and anionic substances on farrowing duration and reproduction performance.
Collection time Control group1 Trial group2 Δ No. of sows n = 24 n = 24 1st-last piglet, min 191.33 154.17 37.16 Interval between littermates, min 22.00A 14.13B 7.87 Litter size at birth
Total 9.92 10.75 0.83 Born alive 8.50 9.88 1.38 Mummified 1.70 2.50 0.80 Stillborn 1.80 1.33 0.47 Condemned 2.00 1.00 1.00
Litter weight at birth, kg 11.91 12.92 1.01 Litter size at weaning 7.33A 9.08B 1.75 Survival at weaning, % 85.00A 92.29B 7.29 Litter weight at weaning, kg 54.22 65.29 11.07 1 Control group = no blend of natural plant extracts, inorganic acids and anionic substances 2 Trial group = with blend of natural plant extracts, inorganic acids and anionic substances A,B Different superscript between groups differed at P<0.10
Conclusions The dietary supplementation with the blend of natural plant extracts, inorganic acids and anionic sub-stances in the diets improved the farrowing and reproduction performance of sows. This blend could be considered as efficient in promoting a more efficient farrowing process, guaranteeing presence of Ca2+ in blood, preventing partly stillborn animals, and rebounding positive effects along lactation. Moreover, the acidification of urine concomitant with good farm management practice can be a pre-ventive measure that minimizes or eliminate the risk factor for occurrence the urogenital tract infec-tions and PPDS.
References
Cline T.R. and Richert B.T. 2001. Feeding growing-finishing pigs. In Swine Nutrition (ed. A.J. Lewis, L.L Southern). pp. 717-724.
Dee S.A. 1992. Porcine urogenital disease. The Veterinary Clinics of North America Food animal Practice 8:641-660.
Dee S.A., Tracy J.D. and King V. 1994. Using citric acid to control urinary tract disease in swine. Veterinary Medicine 89:473-476.
DeRouchey J.M., Hancock J.D., Hines R.H., Cummings K.R., Lee D.J., Maloney C.A., Dean D.W., Park J.S. and Cao H. 2003. Effects of dietary electrolyte balance on the chemistry of blood and urine in lactating sows and sows litter performance. Journal of Animal Science 81:3067-3074.
Foo Y.L., Lu Y., Honwell A.B. and Vorsa N. 2000. A-typer proanthocyanidin trimers from cranberry that inhibit adherence of uropathogenic P-fimbriated Escherichia coli. Journal of Natural Products 63: 1225-1228.
MacLean C.W. and Thomas N.D. 1974. Faecal and vaginal bacteriology of sows during the reproductive cycle. British Veterinary Journal 130:230-237.
Maes D., Verdonck M. and de Kruif A. 1998. Vaginal microecology and vulval discharge in swine. In Vaginal Flora in Health and Disease (ed. P. Heidt, P.B. Carter, V.D. Rusch, D. van der Waaij). pp. 39-48.
Miller M.B., Hartsock T.G., Erez B., Douglass L. and Alston-Mills B. 1994. Effect of dietary calcium concentrations during gestation and lactation in the sow on milk composition and litter growth. Journal of Animal Science 72:1315-1319.

Urbaityte et al.: Effect of urinary acidifier on reproduction performance in sows
8. BOKU-Symposium Tierernährung 2009 Seite 207
Saoulidis K., Kritas S.K., Sarris K., Roubies N. and Kyriakis S.C. 2000. Case report of urinary tract infection in sows of a commercial pig unit. Journal of Hellenic Veterinary Medical Society 51:272-276.
Suzuki K., Benno Y., Mitsuoka T., Takebe S., Kobashi K. and Hase J. 1979. Urease-producing species of intestinal anaerobes and their activities. Applied and Environmental Microbiology 37:379-382.
Corresponding author
Dr. Renata Urbaityte BIOMIN GmbH Industriestrasse 21 A-3130 Herzogenburg E-mail: [email protected]

Matusevicius und Asmenskaite: Zum Einfluss von Zichoriewurzelmehl auf Leistungsparameter von Mastkaninchen
Seite 208 8. BOKU-Symposium Tierernährung 2009
Zum Einfluss von Zichoriewurzelmehl auf Leistungsparameter von Mastkaninchen
P. Matusevicius und L. Asmenskaite
Litauische Veterinärmedizinische Akademie in Kaunas
Einleitung Als eine alternative Substanzgruppe für die verbotenen Fütterungsantibiotika in der EU werden auch Präbiotika diskutiert (Flickinger et al., 2003; Falcao-e-Cunha et al., 2007). Zu den präbiotisch wirken-den Kohlenhydraten zählt Inulin, das u. a. in der Topinamburknolle und in den Wurzeln der Zichorie (Chichorium intybus L.) reichlich enthalten ist. In Versuchen mit wachsenden Kaninchen wurde des-halb ein Trockenprodukt aus der Zichoriewurzel in 2 unterschiedlichen Mengen im pelletierten Allein-futter geprüft, nachdem in Modellversuchen mit dem Labortier Ratte mit verschiedenen Zichoriepro-dukten sich das Wurzelmehl am günstigsten auf die Prozesse im Caecum auswirkte (Juskiewicz et al., 2006).
Material und Methoden Mit dem Zichoriewurzelmehl (ZM) wurden 2 Wachstumsversuche mit Kaninchen der Rasse Weiße Neu-seeländer im Vivarium der LVA in Kaunas durchgeführt. Versuch 1 bestand aus 24 und Versuch 2 aus 21 Tieren, die jeweils auf 3 Gruppen bei Einzeltierhaltung der Kaninchen aufgeteilt waren:
Gruppe I (Kontrollgruppe), Gruppe II mit 25 g ZM/kg Futtermischung, Gruppe III mit 50 g ZM/kg Fut-termischung.
Die Prüfsubstanz wurde anstelle von Trockenschnitzel aus der Zuckerrübenverarbeitung in den Mi-schungen integriert. Die Basisration hatte die folgende Zusammensetzung je kg: 179,5 g Gersten-schrot, 135,2 g Haferschrot, 100 g Weizenkleie, 165 g Sonnenblumenextraktionsschrot, 80 g Sojaext-raktionschrot, 260 g Grasmehl, 50 g Trockenschnitzel, 21 g Calciumcarbonat, 2,3 g Natriumchlorid, 2 g Natriumbicarbonat und 5 g Spurenelement-Vitamin-Prämix. An Inhaltsstoffen wurden in der Basisra-tion u.a. berechnet (je kg TS): 9,83 MJ Umsetzbare Energie, 175 g Rohprotein, 145 g Rohfaser, 8,3 g Lysin, 8,0 g Methionin + Cystin, 6,6 g Threonin, 2,6 g Tryptophan, 10,1 g Calcium, 5,3 g Phosphor, 2,0 g Natrium. Die chemische Analyse des Prüfpräparates Zichoriewurzelmehl ergab folgende Zusam-mensetzung (je kg TS): 965 g Trockensubstanz, 36 g Rohasche, 49 g Rohprotein, 50 g Cellulose, 771 g Inulin, 30 g Saccharose, 29 g Fructose.
Versuch 1 begann am 54. und Versuch 2 am 36. Lebenstag der Tiere; die Versuchsdauer betrug 36 (Versuch 1) bzw. 56 Tage (Versuch 2). Folgende Parameter wurden erfaßt: Lebendmassen je Tier zu Versuchsbeginn, nach 28 Tagen (Versuch 2) und am Versuchsende, Futterverzehr durch Rückwaage der Futterreste an den Wägeterminen, Schlachtkörpergewicht und Schlachtköperzusammensetzung der Tiere von Versuch 2. Der Futteraufwand wurde auf der Basis der verzehrten Futtermengen und der erzielten Zunahmen berechnet. Des Weiteren wurde der Gesundheitszustand beobachtet. Außer-dem erfolgten verschiedene biochemische, physiologische und histologische Untersuchungen am Ver-suchsende mit dem Verdauungstrakt und dessen Inhalt sowie Blutuntersuchungen, deren Ergebnisse von Versuch 1 inzwischen publiziert sind (Juskiewicz et al., 2008; Asmenskaite et al., 2007). Zur statis-tischen Auswertung der Versuchsdaten wurde das Programm STATISKA für WINDOWS (STATSOFT, INC., 2007) benutzt.

Matusevicius und Asmenskaite: Zum Einfluss von Zichoriewurzelmehl auf Leistungsparameter von Mastkaninchen
8. BOKU-Symposium Tierernährung 2009 Seite 209
Ergebnisse und Diskussion Beide Versuche verliefen normal. Verluste und gesundheitliche Störungen (z.B. Durchfall) traten nicht auf. Sicherlich haben hierzu die optimalen Haltungsbedingungen maßgeblich beigetragen.
Im Versuch 1 wurde kein Effekt der Prüfsubstanz festgestellt. Die Ursache könnte sein, daß die Tiere bei Versuchsbeginn 54 Tage alt waren und eine mittlere Lebendmasse von 2,43 ± 0,17 kg bei Ver-suchsbeginn aufwiesen.
Die Leistungsdaten von Versuch 2 enthält Tabelle 1 für den gesamten Prüfzeitraum zusammengefaßt. Im Vergleich zu den Tieren in Gruppe I verzehrten die Kaninchen von beiden Zichorie-Gruppen deut-lich mehr Futter und wiesen am Versuchsende höhere Lebendmassen auf. Die Zunahmeverbesserung durch den präbiotischen Futterzusatz beträgt 8% (Gruppe II) bzw. 11% (Gruppe III). Eine von Fal-cao-e-Cunha et al. (2007) erfolgte Literaturauswertung zeigt sehr unterschiedliche Wirkungen bei den zootechnischen Parametern, die z.B. bei der Lebendmassezunahme vom Nulleffekt bis 11% Verbesse-rung reichen.
Tabelle 1: Leistungsdaten der Kaninchen im Versuch 2
Gruppe Futterverzehr g/Tier
Anfangsmasse g/Tier
Endmasse g/Tier
Zunahme g/Tier
Futteraufwand kg Futter je 1 kg Zunahme
I (Kontrolle) 3776 799±21 2150±60a 1351a 2,794 II (25 g ZM/kg Futter
4117 797±42 2260±31ab 1463ab 2,814
III (50 g ZM/kg Futter)
4126 798±22 2300±33b 1502a 2,747
Mittelwerte mit unterschiedlichen Hochbuchstaben unterscheiden sich statistisch signifikant (P<0,05)
Infolge der Lebendmassesteigerung durch das Zichoriepräparat sind gleichfalls die mittleren Schlacht-körpermassen in den Gruppen II und III gegenüber I insignifikant höher. Bei der Schlachtkörperzu-sammensetzung (prozentuale Anteile von Vorderteile, Rücken und Keulen am Schlachtkörper) unter-scheiden sich die Gruppen nicht oder nur minimal.
Zusammenfassung In 2 Wachstumsversuchen mit Kaninchen der Rasse Weiße Neuseeländer wurde Zichoriewurzelmehl in Anteilen von 25 bzw. 50 g/kg Futter geprüft. Das Produkt ist reich an Inulin, einem Kohlenhydrat mit präbiotischer Wirkung. Während mit älteren und schwereren Tieren im Versuch 1 ein Effekt der Prüf-substanz ausblieb, wurde im Versuch 2 das Wachstum numerisch (25 g/kg) bzw. signifikant verbessert (50 g/kg). Ein positiver Effekt von Zichoriewurzelmehl bestand auch auf die Schlachtkörpermasse.
Literatur
Asmenskaite, L., Juskiewicz, J., Zdunczyk, Z., Staniskiene, B., Budreckiene. R., Sinkeviciene, I., Zilinskiene, A. and Matusevicius, P. (2007): Influence of chicory flour (Cichorium intybus L.) on physiology of digestive tract and health in rabbits. Veterinarija ir Zootechnika T. 40 (62), 3-8.
Falcao-e-Cunha, L., Castro-Solla, L., Maertens, L., Marounek, M., Pinheiro, V., Freire, J. and Nourao, J.L. (2007): Alternatives to antibiotic growth promoters in rabbit feeding: a review. World Rabbit Sc. 15, 127-140.
Flickinger, E.A., Loo, J.V. and Fahey, G.C. (2003): Nutritional response of the presence of inulin and oligofructose in the diets of domestical animals. A review. Food Science and Nutrition 43, 19-60.

Matusevicius und Asmenskaite: Zum Einfluss von Zichoriewurzelmehl auf Leistungsparameter von Mastkaninchen
Seite 210 8. BOKU-Symposium Tierernährung 2009
Juskiewicz, J., Asmenskaite, L., Zdunczyk, Z., Matusevicius, P., Wroblewska, M. and Zilinskiene, A. (2008): Metabolic response of the gastrointestinal tract and serum parameters of rabbits to diets containing chicory flour rich in inulin. J. of Animal Physilogy and Animal Nutrition 92, 113-120.
Juskiewicz, J., Galazka,I., Krol, B. and Zdunczyk, Z. (2006): Effect of chicory products with different inulin content on rat caecum physiology. J. of Animal Physilogy and Animal Nutrition 90, 200-2007.
StatSoft, Inc. (2007): Statisca (data analysis software system), version 8.0. www.statsoft.com.
Autorenanschrift
Doz. Dr. Paulius Matesevicius Litauische Veterinärmedizinische Akademie Tilzes g. 18 LT-47181 Kaunas, Litauen E-Mail: [email protected]

Firmensponsoring – JRS J.Rettenmaier & Söhne GmbH+Co.KG, Geschäftsbereich Tierernährung Holzmühle 1, D-73494 Rosenberg
8. BOKU-Symposium Tierernährung 2009 Seite 211

Liu et al.: Einfluss von Lignocellulose in der Broilerfütterung
Seite 212 8. BOKU-Symposium Tierernährung 2009
Einfluss von Lignocellulose in der Broilerfütterung
Jin Liu1, Julia Trautwein1, Manfred Pietsch2 und Georg Dusel1 1 Fachhochschule Bingen, Fachbereich 1 - Life Sciences – Agrarwirtschaft, Germany 2 J. Rettenmaier & Söhne GmbH + Co. KG, Holzmühle, Germany
Einleitung Beschäftigt man sich mit der Qualität von Exkrementen in der Geflügelfütterung so steht das soge-nannte „wet-litter-syndrom“ (feuchte Einstreu) im Fokus der Betrachtung. Das Syndrom verursacht erhebliche wirtschaftliche Verluste in der Geflügelproduktion. Daher gilt es Alternativen aufzuzeigen die Trockensubstanz des Kotes zu erhöhen. Der Einfluss von Rohfaser auf die Kotkonsistenz wird in Arbeiten von HARTINI et al. (2003) und POTTGÜTER (2008) umfassend dargestellt. Voraussetzung zur Beeinflussung der Kotkonsistenz ist das Wasserbindevermögen der Rohfaserquelle. Faserstoffe, die in hohem Ausmaß lignifiziert sind scheinen besonders zur Modifikation der Kotkonsistenz geeignet zu sein, da das Lignin die Fasersubstanzen vor mikrobiellem Abbau schützt. Ausserdem wird der Einsatz von unlöslicher cellulosischer Faser in Geflügelfutterrationen aufgrund einer besseren Entwicklung des Muskelmagens beim wachsenden Geflügel und einer verbesserten Stärkeverdaulichkeit von internatio-nal tätigen Geflügelzuchtunternehmen empfohlen. ISA (2007).
Das Rohfaserkonzentrat ARBOCEL® (natürliche, reine, auffibrillierte, mykotoxin- und rindenfreie Ligno-cellulose) mit einem Rohfasergehalt von über 65 % erfüllt diese Voraussetzung, da es überwiegend unlösliche Fasern liefert und zudem mit ca. 25% Lignin relativ stark lignifiziert ist (PIETSCH, 2008). Das Wasserbindevermögen von ARBOCEL® Lignocellulose liegt mit ca. 800 % weit über dem Wasserbinde-vermögen von vielen konventionellen Rohfaserträgern. Dieses hohe Wasserbindevermögen bedingt sich durch die fein fibrillierte Faserstruktur von ARBOCEL® Lignocellulose, welche durch eine spezielle Vermahlungs- und Fibrillierungstechnologie (HPC Fibrillierung) erreicht wird.
In nachfolgender Untersuchung sollte dieser Effekt HPC fibrillierter Lignocellulose unter Versuchsbe-dingung (Institutsbedingungen) an Broilerküken getestet werden.
Material und Methoden Die Studie wurde mit 200 Mastküken (Genetik Cobb-500) durchgeführt. Die Versuchsdurchführung erfolgte in der Broilerkäfiganlage der Fachhochschule Bingen. Dort stehen für Versuchsfragen 100 Broilerkäfige (Doppelbelegung) zur Verfügung. Versuchsbeginn war am dritten Lebenstag, bei einem mittleren Durchschnittsgewicht von 85g. Je 40 Tiere wurden nach Gewicht balanciert und zufällig den 5 Fütterungsvarianten zugeteilt:
A kommerzielles Broilerfutter ohne Zusatz B kommerzielles Broilerfutter + 0,3% ARBOCEL®
C kommerzielles Broilerfutter + 0,6% ARBOCEL®
D kommerzielles Broilerfutter + 0,9% ARBOCEL®
E kommerzielles Broilerfutter + 1,2% ARBOCEL®
Die gesamte Versuchsdauer betrug 35 Tage, bis zu einem Ausstallgewicht von ca. 2,3-2,4 kg LM. Die Futterzuteilung erfolgte in pelletierter Form über Futterautomaten. Als Basisfutter diente kommerziel-les Broilerstarter- (Tag 1-14, Cobb 500-Starter) und Broilermastfutter (Tag 15-35, Cobb 500-Grower)

Liu et al.: Einfluss von Lignocellulose in der Broilerfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 213
der Muskator GmbH, Mannheim. In dortiger Mahl- und Mischanlage erfolgte auch die Supplementie-rung des zu prüfenden Faserkonzentrates ARBOCEL® im Austausch mit Weizenkleie (vgl. Tab. 1).
Tabelle 1: Zusammensetzung der Futterrationen (Hauptkomponenten Starter- und Aufzuchtfutter- Angaben Muskator-Werke GmbH)
Rohwaren (in %)
Starter (Tag 1-14)
Grower (Tag 15-35)
A B C D E A B C D E
Körnermais Weizen Sojaextr.schrot 46 Pflanzenfett Pflanzenöl Rapsextr.schrot Lysin 50 W.Kleie ARBOCEL®
1,200 ---
0,900 0,300
16,0 48,8 22,6 3,8 --- 2,0
1,29
0,600 0,600
0,300 0,900
--- 1,200
1,200 ---
0,900 0,300
15,4 49,0 19,0 4,0 1,6 5,0
1,11 0,600 0,600
0,300 0,900
--- 1,200
Die Futteraufnahme wurde wöchentlich pro Käfig erfasst. Das Tränkewasser stand den Tieren in Scha-lentränken ad lib. zur Verfügung. Die Tiere wurden zu Versuchsbeginn und dann im weiteren Verlauf wöchentlich einzeln gewogen. Der Futteraufwand wurde pro Woche und Käfig errechnet.
Am Versuchstag 22 und zu Versuchsende (Tag 36) wurde von 4 Broilern je Variante, nach cervicaler Dislokation der Colon entnommen und der Inhalt zur Bestimmung der Trockenmasse gewonnen. Tro-ckenmassebestimmung erfolgte auch ab Beginn der dritten Versuchswoche wöchentlich mittels Kot-sammlung (10 Käfige pro Variante).
Am Ende der dritten Versuchswoche wurde die Tierzahl halbiert, so dass die Tiere in den letzten bei-den Versuchswochen einzeln gehalten wurden.
Die Futtermittelanalyse der Rohnährstoffe erfolgt nach Weender-Analyse + Stärke- und Zuckerbe-stimmungsmethode im Labor der Fachhochschule Bingen. Die Metabolische Energie (MJ ME) wurde nach geltender Mischfutterformel aus den Rohnährstoffgehalten errechnet.
Die statistische Auswertung der Daten erfolgte mit dem Programm Statistica for Windows (StatSoft, Version 8) anhand einer Varianzanalyse (General Linear Model Procedure) unter Berücksichtigung der Behandlungsgruppe als fixer Faktor. Die korrigierten Mittelwerte (LS-Means) wurden mittels Tukey HSD-Test auf signifikante Unterschiede geprüft. Signifikante Mittelwertunterschiede (p<0,05) werden durch unterschiedliche Kleinbuchstaben (a, b, c) gekennzeichnet.
Ergebnisse Futteranalysen:
Die analysierten Rohnährstoffgehalte sind Tabelle 2 zu entnehmen.

Liu et al.: Einfluss von Lignocellulose in der Broilerfütterung
Seite 214 8. BOKU-Symposium Tierernährung 2009
Tabelle 2: Analysierte Inhaltsstoffe (g/kg bei 88% TS) der Versuchsdiäten
Inhaltsstoffe (g/kg 88%)
Starterfutter (Tag 1-14)
Mastfutter (Tag 15-35)
A B C D E A B C D E
Rohprotein Rohfett Rohfaser Rohasche Stärke Zucker ME - G (in MJ)
217 40,8 26,2 57,1 421 58,6 11,9
213 41,0 25,0 61,2 419 54,3 11,7
215 39,9 28,7 65,1 415 55,5 11,6
211 39,9 26,0 71,3 411 52,3 11,5
215 39,4 36,1 75,7 409 53,9 11,5
196 42,9 32,7 49,9 415 48,2 11,8
194 42,4 34,0 50,1 425 50,5 12,0
200 42,7 32,6 51,4 429 50,5 12,1
200 43,0 34,8 50,5 432 51,5 12,2
197 43,4 36,4 49,9 424 50,2 12,1
Gewichtsentwicklung:
Die Küken wurden mit einem mittleren Gewicht von 85 g den Behandlungsgruppen zugeteilt. Die Kon-trollgruppe erreichte nach 35 Versuchstagen ein durchschnittliches Ausstallgewicht von 2.295g (Tab. 3). Gegenüber der Kontrollgruppe erreichten die Tiere der Versuchsvariante B ein um 2,3% höheres Ausstallgewicht. Variante C lag um 4,8% über dem Niveau der Kontrollgruppe, Variante D um 3,6% und Variante E um 3,1%.
An Versuchstag 15 konnte Variante D gegenüber der 0-Kontrolle eine signifikant verbesserte Lebend-masseentwicklung aufweisen, an Tag 22 war die Differenz zwischen der Kontrolle und den Zulagestu-fen 0,3%, 0,6% und 0,9% statistisch abzusichern.
Tabelle 3: Broilergewichte LM der Versuchsvarianten in g (n=40/20 Tiere je Variante)
LM in g A B C D E Varianz- Analyse
Tag Kontrolle 0,3% 0,6% 0,9% 1,2% SEM p-Wert
LM 1 85,3 ± 7,5 85,1 ± 7,5 85,4 ± 7,5 85,4 ± 7,5 85,0 ± 7,6 0,53 0,999
LM 8 315 ± 28 325 ± 32 324 ± 28 328 ± 32 317 ± 31 2,18 0,308
LM 15 619 a ± 81 661 ab ± 74 659 ab ± 76 672 b ± 71 647 ab ± 91 5,73 0,041
LM 22 1065 a ± 182 1163 bc ± 157 1165 bc ± 133 1168bc ± 131 1132 ac ± 164 11,29 0,018
LM 29 1717 ±215 1724 ± 160 1797 ± 152 1754 ± 92 1768 ± 182 16,52 0,562
LM 36 2295 ± 357 2347 ± 225 2406 ± 299 2377 ± 230 2366 ± 304 29,14 0,824
Über den Versuchszeitraum Woche 1 bis 3 erreichten die Tiere der Zulagestufen 0,3%, 0,6% und 0,9% Arbocel® eine signifikant verbesserte Tageszunahme gegenüber der Kontrollvariante (Tab. 4).

Liu et al.: Einfluss von Lignocellulose in der Broilerfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 215
Tabelle 4: Tägliche Zunahme TZ der Versuchsvarianten in g (n=40 Tiere je Variante)
TZ in g A B C D E Varianz- Analyse
Woche Kontrolle 0,3% 0,6% 0,9% 1,2% SEM p-Wert
TZ 1-3 44,5 a ± 8,3 49,4 b ± 6,7 49,1 b ± 5,9 49,4 b ± 5,8 47,8 ab ± 7,4 0,51 0,010
TZ 4-5 90,0 ± 18,9 84,1 ± 13,7 91,8 ± 12,7 88,0 ± 11,0 87,0 ± 18,2 1,68 0,590
TZ 1-5 61,4 ± 10,0 62,8 ± 6,2 64,5 ± 8,2 63,6 ± 6,4 63,3 ± 8,4 0,81 0,822
Futteraufnahme und Futteraufwand:
Über die gesamte Versuchsperiode nahm die Kontrollgruppe A durchschnittlich 115 g Futter/Tier/Tag auf. Bei den Versuchsvarianten B, C, D und E lag die Futteraufnahme mit 114 g, 120 g, 116 g sowie 119 g/Tier/Tag auf vergleichbarem Niveau (Tab. 5). In Versuchswoche 3 zeigten die Tiere der Zulage-stufen 0,6% und 1,2% gegenüber der Kontrollvariante eine erhöhte Futteraufnahme von 3,8% bzw. 3,3%.
Tabelle 5: Futteraufnahme FA der Versuchsvarianten je Tier und Tag in g (n=20 Käfige je Variante)
FA in g A B C D E Varianz- Analyse
Woche Kontrolle 0,3% 0,6% 0,9% 1,2% SEM p-Wert
FA 1 44 ± 4 42 ± 5 43 ± 4 44 ± 5 41 ± 5 0,47 0,247
FA 2 78 ± 13 82 ± 11 86 ± 9 84 ± 9 83 ± 12 1,10 0,264
FA 3 116 a ± 18 127 ab ± 15 133 b ± 11 129 ab ± 13 132 b ± 17 1,59 0,006
FA 4 160 ± 29 153 ± 24 155 ± 23 145 ± 14 162 ± 22 2,33 0,148
FA 5 185 ± 40 171 ± 27 189 ± 17 184 ± 37 183 ± 28 3,20 0,454
FA 1-3 78 ± 11 82 ± 9 85 ± 7 84 ± 7 83 ± 11 0,91 0,117
FA 4-5 173 ± 33 162 ± 19 174 ± 15 164 ± 22 173 ± 23 2,40 0,335
FA 1-5 115 ± 16 114 ± 10 120 ± 8 116 ± 10 119 ± 14 1,23 0,415
Die Berechnung der Futterverwertung (FVW) je Käfig erfolgt an Hand der Futteraufnahme pro Käfig und der Einzeltiergewichte.
Die Futterverwertung der Kontrollvariante A lag während der 5-wöchigen Versuchsphase durchschnitt-lich bei 1,83g Futter pro g LM-Zuwachs. Bei den Varianten B, C, D und E lag die Futterverwertung bei 1,77g, 1,79g, 1,74g und 1,80g Futter pro g Zuwachs. Errechnet man die relativen Veränderungen der Futterverwertung im Verhältnis zur Kontrollvariante A, so konnte die Futterverwertung bei 0,3% Arbo-cel® um 3,3% verbessert werden, bei 0,6% um 2,2%, bei 0,9% um 4,9% und bei 1,2% um 1,6% (Tab. 6). Diese Effekte konnten jedoch statistisch nicht abgesichert werden. Statistisch abzusichern

Liu et al.: Einfluss von Lignocellulose in der Broilerfütterung
Seite 216 8. BOKU-Symposium Tierernährung 2009
war jedoch eine verbesserte FVW in der Variante B (-11,9%) gegenüber A in der ersten Versuchswo-che sowie zwischen Variante D (-7,2%) und A in der zweiten Versuchswoche.
Tabelle 6: Futterverwertung FVW der Versuchsvarianten in g Futter pro g Zunahme (n=20 Käfige je Variante)
FVW A B C D E Varianz- Analyse
Woche Kontrolle 0,3% 0,6% 0,9% 1,2% SEM p-Wert
FVW 1 1,60 a ± 0,35 1,41 b ± 0,11 1,45 ab ± 0,09 1,46ab±0,11 1,43ab± 0,07 0,020 0,017
FVW 2 1,81 a ± 0,16 1,73 ab ± 0,08 1,79 ab ± 0,14 1,68 b±0,12 1,76ab± 0,11 0,014 0,023
FVW 3 1,85 ± 0,15 1,80 ± 0,08 1,84 ± 0,09 1,84 ± 0,16 1,88 ± 0,15 0,013 0,404
FVW 4 1,79 ± 0,24 1,90 ± 0,26 1,72 ± 0,17 1,78 ± 0,20 1,81 ± 0,14 0,022 0,094
FVW 5 2,16 ± 0,37 2,07 ± 0,21 2,16 ± 0,44 2,05 ± 0,29 2,21 ± 0,43 0,038 0,635
FVW 1-3 1,79 ± 0,13 1,70 ± 0,09 1,74 ± 0,08 1,70 ± 0,09 1,76 ± 0,13 0,011 0,056
FVW 4-5 2,01 ± 0,32 1,95 ± 0,22 1,91 ± 0,18 1,90 ± 0,16 1,98 ± 0,20 0,023 0,591
FVW 1-5 1,83 ± 0,13 1,77 ± 0,08 1,79 ± 0,09 1,74 ± 0,07 1,80 ± 0,09 0,011 0,129
Trockensubstanzgehalt von Colon-Digesta und Faeces:
An den Versuchstagen 22 und 36 wurden vier Tiere je Variante geschlachtet und der Trockensub-stanzgehalt vom Dickdarminhalt (Colon) analysiert. Signifikant vermindert zeigte sich der Colon-TS-Gehalt der Versuchsvariante E gegenüber A und B an Versuchstag 22. Zu Versuchsende konnte kein Behandlungseinfluss auf den Colon-TS-Gehalt festgehalten werden.
Tabelle 9: Trockensubstanzgehalt des Dickdarminhaltes (n=4 pro Variante)
Colon -TS A B C D E Varianz-
Analyse
Tag Kontrolle 0,3% 0,6% 0.9% 1,2% SEM p-Wert
22 23,0 a ± 1,86 23,7 a ± 2,09 22,5 ab ± 1,21 21,6 ab ± 2,01 19,7 b ± 2,30 0,492 0,119
36 21,5 ± 2,60 20,8 ± 1,33 23,3 ± 1,10 21,6 ± 1,95 23,0 ± 1,77 0,422 0,302
Bei der Bestimmung der Kot-Trockenmasse konnte an Versuchstag 13 ein geringerer TS- Gehalt in den Varianten D und E gegenüber A sowie E gegenüber B und C festgehalten werden. An den darauf-folgenden Messzeitpunkten konnte kein Behandlungseffekt auf die Kot-TM abgesichert werden.

Liu et al.: Einfluss von Lignocellulose in der Broilerfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 217
Tabelle 10: Trockensubstanzgehalt im Kot (n=10 pro Variante)
Kot-TS A B C D E Varianz- Analyse
Tag Kontrolle 0,3% 0,6% 0,9% 1,2% SEM p-Wert
13 25,1 a ± 1,4 23,1 ac ± 1,6 23,3 ac ± 1,7 21,4 bc ± 1,3 20,7 b ± 2,3 0,32 0,000
20 25,4 ± 4,1 24,6 ± 1,3 25,8 ± 2,2 25,7 ± 2,8 28,9 ± 5,7 0,53 0,085
27 26,0 ± 4,1 24,3 ± 2,8 26,8 ± 1,4 24,0 ± 3,4 25,6 ± 3,5 0,46 0,289
34 24,1 ± 2,6 25,4 ± 1,8 24,5 ± 2,5 23,0 ± 3,9 23,4 ± 3,7 0,43 0,428
Gesundheitsstatus:
Über die gesamte Versuchsdauer waren 6 Tierverluste zu verzeichnen. Die Verluste waren gleichmäßig über alle Versuchsgruppen verteilt, so dass ein Behandlungseinfluss ausgeschlossen werden kann. Eine Verlustrate von 3% liegt in der Broileraufzucht auf niedrigem bis üblichem Niveau. Der Gesund-heitszustand der Tiere war über die gesamte Versuchsperiode als positiv und stabil zu bewerten. Da-her war über die Versuchsperiode keine Behandlung und Medikation der Versuchstiere notwendig.
Diskussion und Schlussfolgerung Effekte der Lignocellulosesupplementierung zeigten sich in diesem Versuch im Hinblick auf die Ge-wichtsentwicklung sowie den Futteraufwand. Der Trockensubstanzgehalt vom Colon-Inhalt sowie der Faeces wurde nicht nachhaltig beeinflusst. Sicherlich bedingt durch die Käfighaltung stellt sich zu feuchter Kot in der eingesetzten Versuchsanlage nicht als auffallend oder gar problematisch dar. Den-noch könnte die Lignocellulose auf Grund der im Dickdarm wirkenden unlöslichen Fasern regulierend auf die Darmfunktion wirken. Die unlöslichen Fasern erhöhen die Darmperestaltik und fördern damit einen ungestörten und regelmäßigen Kotabsatz und erschweren die Ansiedlung pathogener Erreger. Somit tragen sie zur Darmgesundheit bei (LEIBETSEDER und NEUFELD, 2008). Da die Leistungsparameter positive Effekte der Lignocellulose auf den Futteraufwand zeigen, könnte daher dennoch –wenn auch nicht durch den Kot-TS-Gehalt erkennbar- eine gute Darmgesundheit die Ausnutzung der Nährstoffe begünstigen. Des Weiteren können die fein fibrilliereten Fasern des Rohfaserkonzentrates eine Art Netzwerk ausbilden, welches den Nahrungsbrei auflockert und damit die körpereigene Enzympenetra-tion und somit die Nährstoffausnutzung verbessern kann.
In einem 5-wöchigen Mastversuch mit 200 Broilern zeigte der Einsatz eines Rohfaserkonzentrates aus HPC fibrillierter Lignocellulose (Arbocel®) positive Effekte auf die Gewichtsentwicklung und den Futter-aufwand. Die Tiere wurden im Alter von 3 Tagen und mit einem mittleren Gewicht von 85g fünf Be-handlungsgruppen zugeteilt. Diese differenzierten sich durch unterschiedlich hohe Gehalte an Ligno-cellulose (0%, 0,3%, 0,6%, 0,9%, 1,2%) welche im Austausch mit Weizenkleie in die Futterrationen (2-phasig, pelletiert) eingemischt wurde. Die Tiere –Cobb-500-Genetik- wurden in Käfigen gehalten (Tag 1-21 Doppelbelegung, ab Tag 22 Einzeltierhaltung) und erhielten das Futter über Trockenfutter-automaten und Wasser über Schalentränken.
Eine höhere Tagezunahme der Behandlungsgruppen 0,3% (49,4g), 0,6% (49,1g) und 0,9% Arbocel® (49,4g) gegenüber der 0-Kontrolle (44,5g) konnte in den ersten 3 Versuchswochen abgesichert wer-den. Nicht abzusichern war ein höheres Mastendgewicht von 2,3% (0,3% Arbocel®), 4,8% (0,6%

Liu et al.: Einfluss von Lignocellulose in der Broilerfütterung
Seite 218 8. BOKU-Symposium Tierernährung 2009
Arbocel®), 3,6% (0,9% Arbocel®) und 3,1% (1,2% Arbocel®) gegenüber der 0-Kontrolle. Eine Beein-flussung der Futteraufnahme durch die Supplementierung konnte in diesem Versuch nicht nachgewie-sen werden. In den ersten beiden Versuchswochen zeigte sich in den Dosierungsstufen 0,3% und 0,9% signifikant positive Effekte auf den Futteraufwand. Über den gesamten Versuchszeitraum wurde tendenziell eine verbessert Futterverwertung von 3,3%, 2,2%, 4,9% und 1,6% mit ansteigender Ar-bocel®-Supplementierung dokumentiert. Gerichtete Effekte auf den Kottrockensubstanzgehalt konnten nicht festgehalten werden. Errechnet man an Hand eines quadratischen Modells (r=0,95) der Tages-zunahmen und des Futteraufwandes über den gesamten Versuch die „Optimalkonzentration“ an Lignocellulose in der Ration, so ergibt sich eine Arbocel®-Konzentration von 0,67 bis 0,7%.
Literatur
HARTINI S., M. CHOCT, G. HINCH AND J. NOLAN (2003): Effect on diet composition, gut microbial status and fibre forms on “Cannibalim in Layers”; Australian Egg Corporation Limit, April
LEIBETSEDER und NEUFELD, 2008: Mehr als nur Rohfaser, SUS 02/2008, S. 48
PIETSCH M., (2009): Rohfaser, die vernachlässigte Komponente in der Geflügelfütterung (unveröffentlichte Mitteilung)
POTTGÜTER R., (2008): Fibre in Layer diets, Lohmann Information, Vol 43 (2). Oct. 2008, p. 22-31
ISA A HENDRIX GENETICS COMPANY (2007) Energy levels and feed presentation for laying hens: Effects on performance and intake
Autorenanschrift
Prof. Dr. Georg Dusel Fachhochschule Bingen/University of Applied Sciences Fachbereich 1 – Life Sciences and Engineering Berlinstrasse 109, D- 55411 Bingen, Germany E-Mail: [email protected]

Kroismayr et al.: Lignocellulose als Nahrungsfaserquelle in der Zuchtsauenfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 219
Lignocellulose als Nahrungsfaserquelle in der Zuchtsauenfütte-rung
Arthur Kroismayr1, Fabian Braunsberger2, Klaus Neufeld3 1 Agromed Austria GmbH, Bad Haller Straße 23, A-4550 Kremsmünster, Österreich 2 Höhere landwirtschaftliche Bundeslehranstalt St. Florian, Fernbach 37, A-4490
St. Florian, Österreich 3 Animal Nutrition Research Center, Sattelbach 13, A-2532 Heiligenkreuz, Österreich
Einleitung Traditionell werden in der heimischen Zuchtsauenfütterung Produkte wie Kleien oder Zuckerrüben-schnitte als Faserquelle eingesetzt. In modernen Rationen finden sich vermehrt Produkte basierend auf Holz, sogenannte Lignocellulose - Produkte, welche der ausreichenden Versorgung mit Nahrungs-faser dienen sollen. Die positive Wirkung von Nahrungsfaser in der Fütterung tragender Zuchtsauen ist durchaus bekannt und die Notwendigkeit einer ausreichenden Versorgung mit Rohfaser im Bereich der Zuchtsauenernährung ist unbestritten. Das Wissen um die Wichtigkeit dieser Futtermittelfraktion für Gesundheit und Wohlbefinden der Sauen wurde zum Beispiel in der Bundesrepublik Deutschland durch eine gesetzliche Richtlinie, die am 4. August 2006 in Kraft getreten ist, umgesetzt [Bundesge-setzblatt Jahrgang 2006 Teil I Nr. 37 §25(6)]. Diese Richtlinie schreibt einen Rohfasergehalt von min-destens 8% in der Trockenmasse im Alleinfutter für trächtige Jungsauen und Sauen vor.
Generell wird der diätetische Einfluss von Nahrungsfasern in der Tierernährung vorrangig mit einem negativen Effekt auf Futteraufnahme und Leistungsdaten in Verbindung gebracht und der diätetische Nutzen, vor allem jener von fermentierbarer Nahrungsfaser, vernachlässigt.
Eine Unterscheidung zwischen fermentierbarer und nicht - fermentierbarer Nahrungsfaser scheint notwendig, weil der ernährungsphysiologische Effekt dieser Futtermittelkomponenten auf völlig ver-schiedenen Wirkmechanismen basiert. Spricht man in traditioneller Weise von Rohfaser oder Ballast-stoffen, so meint man in der Regel die Effekte nicht - fermentierbarer Nahrungsfaser. Diese sind nicht löslich weisen aber teils ein beträchtliches Wasserbindungsvermögen auf. Im Dickdarm bewirken nicht fermentierbare Nahrungsfasern durch den Volumenreiz eine Verkürzung der Verweildauer des Futter-breis, folglich wird die regelmäßige Darmentleerung gefördert und somit Verstopfung vorgebeugt (Wil-fart et al, 2007). Eine zügige Darmpassage erschwert das „Aufsteigen“ pathogener Keime aus dem Dickdarm in die vorderen Darmabschnitte und trägt auf diese Weise zur Aufrechterhaltung von Darm-gesundheit und Durchfallprophylaxe bei.
Eine ausgewogene Kombination fermentierbarer und nicht - fermentierbarer Nahrungsfaser ist auch insbesondere deshalb notwendig, weil nach neuen wissenschaftlichen Erkenntnissen nicht fermentier-bare Nahrungsfaser dazu beiträgt, dass die fermentierbaren Komponenten in jene hinteren Abschnitte des Dickdarms verlagert werden, wo die Fermentation besonders positive Effekte nach sich zieht (Go-vers et al., 1999). Die nicht fermentierbare Nahrungsfaser schiebt die fermentierbaren Anteile faktisch vor sich her bis in das hintere Colon, wo sie die Darmflora stabilisieren und die Zellen der Darmwand unterstützen, welche die Wasserrückresorption aus dem Darminhalt gewährleisten.
Produkte basierend auf Lignocellulose enthalten sehr hohe Rohfasergehalte und werden seit geraumer Zeit zur Nahrungsfaserergänzung eingesetzt. Im Vergleich zu traditionellen Lignocellulose Produkten, welche mehr oder weniger aus nicht - fermentierbarer Faser bestehen, gibt es auch neuartige Ligno-

Kroismayr et al.: Lignocellulose als Nahrungsfaserquelle in der Zuchtsauenfütterung
Seite 220 8. BOKU-Symposium Tierernährung 2009
cellulose Anwendungen die auch fermentierbare Faserbestandteile enthalten und sich somit positiv auf Verdauungsprozesse und Darmgesundheit von Nutztieren auswirken sollten.
Aus diesem Grund werden im Folgenden zwei Studien vorgestellt bei denen eine definierte Kombinati-on aus Lignocellulose (OptiCell®, Agromed Austria GmbH) welche fermentierbare und nicht fermen-tierbare Nahrungsfaserkomponenten enthält, einmal mit Weizenkleie und einmal mit Weizenkleie und Luzernepellets in der Zuchtsauenfütterung verglichen wurde.
Material und Methoden In Versuch I (Braunsberger, 2009) wurden 18 tragende Zuchtsauen gleicher Genetik und Wurfzahl ab dem 70. Tragendtag auf 2 Versuchsgruppen aufgeteilt. Die Tiere beider Gruppen erhielten die gleiche Menge einer Basisration bestehend aus Gerste, Mais, Weizen, Hafer, Fischmehl und Mineralergänzung. Je nach Futtergruppe wurde diese Basisration mit 300g Weizenkleie (12% XF) bzw. 60g Lignocellulo-seprodukt (59% XF) je Tier und Tag on top ergänzt. Ab der Umstellung auf Laktationsfutter (4 Tage vor dem Abferkeln) erhielten alle Tiere identisches Laktationsfutter (XF: 2,7%).
Nach bzw. während der Geburt wurden die Paramater Lebend geborene Ferkel und Wurfdauer in Mi-nuten aufgezeichnet. Des Weiteren wurde am 22. Versuchstag (Beginn 14. Tragendwoche) der Tro-ckensubstanzgehalt des Sauenkots analysiert.
Die statistischen Berechnungen erfolgten mit dem Statistikpaket SAS. Um die Versuchsgruppen zu vergleichen wurde ein T-Test durchgeführt.
In Versuch II (Sarandan, 2009) wurde selbige Lignozellulose wie bei Versuch I mit Weizenkleie und Luzernepellets über den gesamten Reproduktionszyklus von Zuchtsauen verglichen. Dazu wurden 65 Zuchtsauen auf 2 Gruppen aufgeteilt. Die Kontrollgruppe erhielt Tragendfutter auf Basis Mais, Son-nenblumenschrot, Spurenelement – und Aminosäurenergänzung und als Faserergänzung Weizenkleie (10%) und Luzernepellets (7%). Das Laktationsfutter der Kontrollgruppe enthielt Weizen, Sojaschrot, Sonnenblumenschrot, Fischmehl, Spurenelement – und Aminosäurenergänzung Weizenkleie (10%) als Faserergänzung.
Das Tragendfutter der Versuchsgruppe auf Basis Mais, Sojaschrot, Sonnenblumenschrot Spurenele-ment – und Aminosäurenergänzung enthielt außerdem 3% Lignocelluloseprodukt. Das Säugendfutter der Versuchsgruppe auf Basis Weizen, Sojaschrot, Sonnenblumenschrot, Fischmehl, Spurenelement – und Aminosäurenergänzung enthielt 0,5% Lignocelluloseprodukt bei gleichzeitig verminderten Gehal-ten an Weizenkleie (7%).
Kotproben der Zuchtsauen wurden zu Beginn der Trächtigkeit und dann in monatlichen Abständen während der Tragezeit gesammelt, außerdem bei Einstallung in der Abferkelbucht, drei Tage nach dem Abferkeln sowie beim Absetzen. Die Kotproben wurden auf ihren Trockensubstanzgehalt unter-sucht.
Mit dem Umstallen in die Abferkelbucht bis drei Tage nach dem Abferkeln wurde bei den Sauen mor-gens und abends die innere Körpertemperatur (Rektaltemperatur) gemessen. Am Tag 1 und 3 nach dem Abferkeln wurde das Gesäuge adspektorisch und palpatorisch beurteilt und Mastitissymptome sowie Milchmangel protokolliert. Des Weiteren wurde die Anzahl lebend geborener Ferkel dokumen-tiert.
Statistik: Die Ergebnisse wurden mittels Vierfelder- Χ2-Test bzw. t-Test überprüft.

Kroismayr et al.: Lignocellulose als Nahrungsfaserquelle in der Zuchtsauenfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 221
Ergebnisse Versuch I zeigte, dass die Verfütterung von Lignocellulose im Vergleich zu Weizenkleie zu signifikant (P< 0,05) höherer Kottrockensubstanz führte (35,5% vs. 32,0%). Signifikant verkürzt (p<0,01) konn-te ebenso die Wurfdauer in der Versuchsgruppe (151 min vs. 215 min in der Kontrollgruppe) werden. Die Anzahl lebend geborener Ferkel konnte in der Lignocellulose – Gruppe (11,56) im Vergleich zur Kontrollgruppe mit Weizenkleie (11,0) numerisch verbessert werden. (siehe Abbildungen 1-2)
Abbildung 1: Anzahl lebend geborener Ferkel
Abbildung 2: Wurfdauer in Minuten
a,b p≤0,01
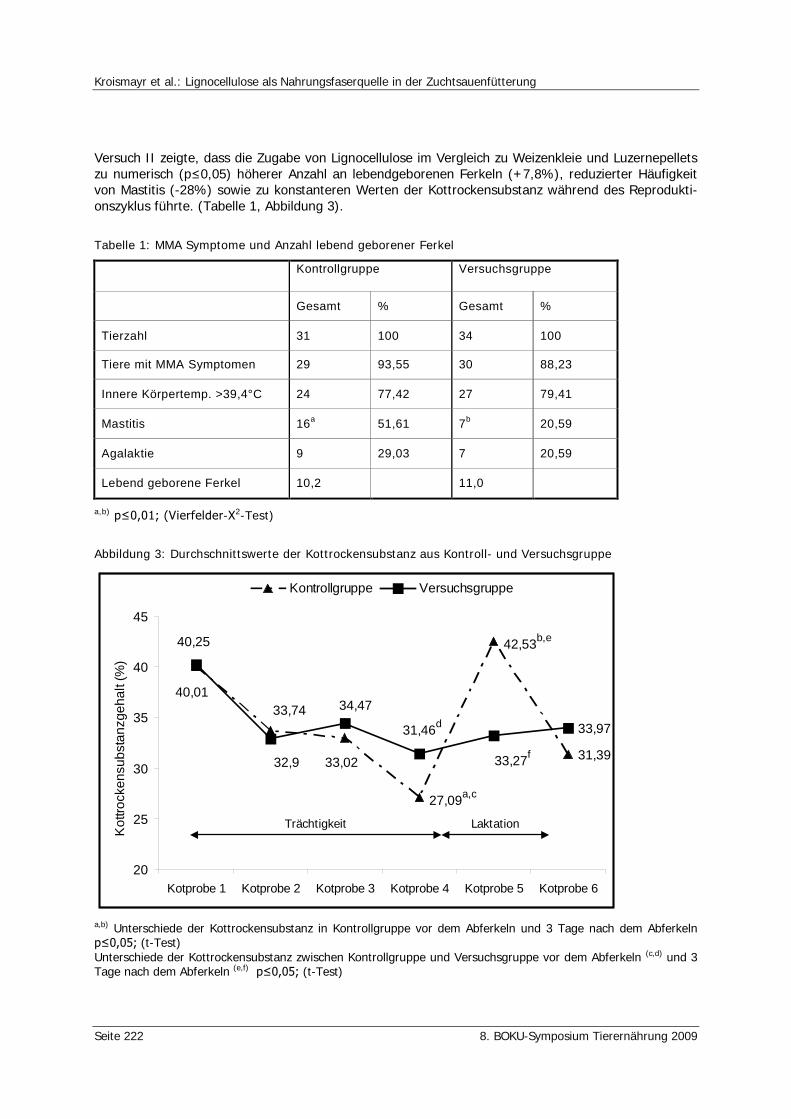
Kroismayr et al.: Lignocellulose als Nahrungsfaserquelle in der Zuchtsauenfütterung
Seite 222 8. BOKU-Symposium Tierernährung 2009
Versuch II zeigte, dass die Zugabe von Lignocellulose im Vergleich zu Weizenkleie und Luzernepellets zu numerisch (p≤0,05) höherer Anzahl an lebendgeborenen Ferkeln (+7,8%), reduzierter Häufigkeit von Mastitis (-28%) sowie zu konstanteren Werten der Kottrockensubstanz während des Reprodukti-onszyklus führte. (Tabelle 1, Abbildung 3).
Tabelle 1: MMA Symptome und Anzahl lebend geborener Ferkel
Kontrollgruppe Versuchsgruppe
Gesamt % Gesamt %
Tierzahl 31 100 34 100
Tiere mit MMA Symptomen 29 93,55 30 88,23
Innere Körpertemp. >39,4°C 24 77,42 27 79,41
Mastitis 16a 51,61 7b 20,59
Agalaktie 9 29,03 7 20,59
Lebend geborene Ferkel 10,2 11,0
a,b) p≤0,01; (Vierfelder-Χ2-Test)
Abbildung 3: Durchschnittswerte der Kottrockensubstanz aus Kontroll- und Versuchsgruppe
33,97
33,02
33,7440,01
31,39
27,09a,c
42,53b,e
32,9
40,25
34,47
31,46d
33,27f
20
25
30
35
40
45
Kotprobe 1 Kotprobe 2 Kotprobe 3 Kotprobe 4 Kotprobe 5 Kotprobe 6
Kot
trock
ensu
bsta
nzge
halt
(%)...
..
Kontrollgruppe Versuchsgruppe
a,b) Unterschiede der Kottrockensubstanz in Kontrollgruppe vor dem Abferkeln und 3 Tage nach dem Abferkeln p≤0,05; (t-Test) Unterschiede der Kottrockensubstanz zwischen Kontrollgruppe und Versuchsgruppe vor dem Abferkeln (c,d) und 3 Tage nach dem Abferkeln (e,f) p≤0,05; (t-Test)
Trächtigkeit Laktation

Kroismayr et al.: Lignocellulose als Nahrungsfaserquelle in der Zuchtsauenfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 223
Diskussion Eine Ration mit hohem Rohfasergehalt soll eine gute Sättigung der Zuchtsauen während der Tragezeit bewirken und somit Aggressionsverhalten und Stereotypien vorbeugen. In der Bundesrepublik Deutschland wurde daher ein minimaler Rohfasergehalt für das Tragefutter gesetzlich festgelegt (Bundesgesetzblatt der Bundesrepublik Deutschland 2006). Bergeron et al (2000) demonstrierten, dass eine Futterration mit hohem Rohfasergehalt bei zweimal täglicher Fütterung stereotypes Verhal-ten reduziert. Noch bessere Effekte konnten bei ad libitum Fütterung erreicht werden. Andere Unter-suchungen widersprechen diesen Ergebnissen. So sahen Holt et al. (2006) bei Verfütterung einer Ra-tion mit hohem Anteil an Sojaschalen keinen positiven Einfluss auf das Wohlbefinden oder die Repro-duktionsleistung. Die widersprüchlichen Ergebnisse können ihre Ursache in der Verwendung unter-schiedlicher Rohfaserquellen haben. Bei der Auswahl der Rohfaserquellen ist darauf zu achten, dass sich fermentierbare und nicht fermentierbare Nahrungsfasern ergänzen sollten, da sie verschiedene Wirkmechanismen und Effekte haben (Govers et al 1999). Um die Eubiose des Darmtraktes zu unter-stützen ist eine ausgewogene Kombination von fermentierbarer und nicht fermentierbarer Nahrungs-faser von Bedeutung.
Neue wissenschaftliche Untersuchungen (Begum et al 2004) haben belegt, dass Lignin als Precursor von Säugerlignanen wie Enterolacton und Enterodiol fungiert, die phytoöstrogene Wirkung haben. Um die Wirkung von Nahrungsfasern und einzelnen Nahrungsfaserkomponenten besser zu verstehen, sind weitere wissenschaftliche Untersuchungen nötig.
In den beschriebenen Studien wurde ein Nahrungsfasersupplement aus unbehandeltem Frischholz eingesetzt. Die Testsubstanz zählt somit zu den sogenannten Lignocelluloseprodukten, welche sowohl fermentierbare als auch nicht fermentierbare Nahrungsfaser enthält, während herkömmliche Lignocel-luloseprodukte ausschließlich nicht fermentierbare Nahrungsfaser aufweisen. Die Testsubstanz wurde bereits in einer großen Anzahl an Feldversuchen in unterschiedlichen Dosierungen und Einsatzberei-chen untersucht. Unter Praxisbedingungen zeigte die Testsubstanz positiven Einfluss auf Darmge-sundheit und Leistungsdaten beim Absetzferkel, Mastschwein, Broiler und Mastkaninchen (Neufeld und Leibetseder, 2008).
Die beiden wissenschaftlichen Studien zeigen, dass das untersuchte Lignocellulose Produkt die Leis-tung von Zuchtsauen im Vergleich zu traditionellen Rohfaserprodukten (Weizenkleie, Luzernepellets) positive beeinflusst. Daraus ist zu schließen, dass die Höhe des Rohfasergehalts selbst, welcher in den Versuchs- und Kontrollgruppen auf annähernd selben Niveau war, wenig über die physiologische Wir-kung im Darm aussagt. Vielmehr scheint die Zusammensetzung der Faser (Anteil lösliche und unlösli-che Faser) eine wichtige Rolle zu spielen. Dadurch ließen sich jedenfalls die Unterschiede in der Dauer des Geburtsprozesses, die erhöhte Zahl lebend geborener Ferkel, das reduzierte Auftreten von MMA und die konstantere Kot – Trockenmasse erklären. Weitere Studien über die genau physiologische Wirkung des untersuchten Lignocellulsoe Produkts sollten folgen.
Schlussfolgerung Das getestete Nahrungsfaserprodukt (OptiCell®, Agromed Austria GmbH) führte in den beschriebenen Studien verglichen mit traditionellen Rohfaserprodukten (Weizenkleie, Luzernepellets) zu einem positi-ven Einfluss auf MMA Symptome, Reproduktionsparameter und Kotqualität. Die Zusammensetzung von Faserprodukten (v.a. löslicher und nicht löslicher Anteil) scheinen eine wichtige Rolle im Bezug auf die physiologische Wirksamkeit zu spielen.

Kroismayr et al.: Lignocellulose als Nahrungsfaserquelle in der Zuchtsauenfütterung
Seite 224 8. BOKU-Symposium Tierernährung 2009
Literatur
Begum AN, Nicolle C, Mila I, Lapierre C, Nagano K, Fukushima K, Heinonen SM, Adlercreutz H, Rémésy C, Scalbert A (2004): Dietary lignins are precursors of mammalian lignans in rats. J. Nutr. 134: 120-127
Bergeron R, Bolduc J, Ramonet Y, Meunier-Salaun MC, Robert S (2000): Feeding motivation and stereotypes in pregnant sows fed increasing levels of fibre and/or food. Appl. Anim. Behav. Sci 70(1): 27-40
Braunsberger F (2009): Einfluss verschiedener Rohfaserträger auf die Reproduktionsleistung von Zuchtsauen. Diplomarbeit, HFLS St. Florian, Austria
Bundesgesetzblatt Bundesrepublik Deutschland 2006, Teil I Nr. 37 §25(6)
Göransson L (1989): The effect of dietary crude fibre content on the frequency of post partum agalactia in the sow. Zentralbl. Veterinärmed. A. 36(6): 474-479
Govers MJ, Gannon NJ, Dunshea FR, Gibson PR, Muir JG (1999): Wheat bran affects the site of fermentation of resistant starch and luminal indexes related to colon cancer risk: A study in pigs. Gut 45(6): 840-847
Holt JP, Johnston LJ, Baidoo SK, Shurson GC (2006): Effects of high-fiber diet and frequent feeding on behavior, reproductive performance and nutrient digestibility in gestating sows. J. Anim. Sci. 84: 946-955
Neufeld K, Leibetseder J (2008): Nahrungsfaser in der Nutztierernährung. Feed Magazine 5-6: 21-27
Sarandan H., Leibetseder J., Neufeld K. (2009): Effects of a new Lignocellulose product for fibre supplementation on MMA symptoms, reproductive performance and faecal quality in a pig farm with evident MMA problems in Romania. Unpublished Study.
Wilfart A, Montagne L, Simmins H, Noblet J, Milgen J (2007): Digesta transit in different segments of the gastrointestinal tract of pigs as affected by insoluble fibre supplied by wheat bran. Br J Nutr 98(1): 54-62
Autorenanschrift
Dr. Arthur Kroismayr Bad Haller Straße 23 A-4550 Kremsmünster (Österreich) E-Mail: [email protected]

Firmensponsoring – agromed Austria GmbH, Bad Hallerstraße 23, A-4550 Kremsmünster
8. BOKU-Symposium Tierernährung 2009 Seite 225

Wetscherek-Seipelt und Wetscherek: Einsatz von Weizenkleie bzw. Trockenschnitzel bei Ferkel
Seite 226 8. BOKU-Symposium Tierernährung 2009
Einsatz von Weizenkleie bzw. Trockenschnitzel bei Ferkel
Gabriela Wetscherek-Seipelt und Wolfgang Wetscherek
Universität für Bodenkultur Wien, Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie
Einleitung Es wurde die Auswirkung des Einsatzes von Trockenschnitzel und Trockenschnitzel-Plus auf die Fut-teraufnahme, Aufzuchtleistung und den Gesundheitsstatus von Ferkeln im Vergleich zu einer negati-ven Kontrollgruppe, sowie einer Gruppe mit Weizenkleie geprüft. Bei den Trockenschnitzel-Plus han-delt es sich um eine Ergänzungsfuttermischung bestehend aus 82% Trockenschnitzel, 12% teil-entzuckerte Melasse, 4% Kartoffelrestfruchtwasser-Konzentrat und 2% Carbonatationskalk.
Material und Methoden Aus den Ferkeln von 6 Zuchtsauen der LFS-Hatzendorf wurden beim Absetzen mit etwa 28 Lebensta-gen 48 Ferkel ausgewählt. Bei der Aufteilung auf die vier Versuchsgruppen wurde auf eine gleichmä-ßige Verteilung von Geschlecht, Wurf und Gewicht geachtet. Der Versuch begann nach der 4. Le-benswoche mit etwa 8,5 kg Lebendgewicht und endete nach 8 Versuchswochen mit einen Lebendge-wicht von etwa 28 kg. Die Tiere wurden in 8 strohlosen Boxen mit Spaltenböden gehalten.
Die Zusammensetzung der Futtermischungen der vier Gruppen unterschied sich nur durch den Zusatz von Trockenschnitzel, Trockenschnitzel-Plus bzw. Weizenkleie. Die jeweiligen Dosierungen waren in der Absetz- und Aufzuchtperiode gleich. Die Umstellung von der Absetz- auf die Aufzuchtfuttermi-schung erfolgte nach zwei Versuchswochen. Der Versuchsplan wird in der Tabelle 1 dargestellt.
Tabelle 1 Versuchsplan
Gruppe 1 Gruppe 2 Gruppe 3 Gruppe 4 Neg. Kontrolle = Ba-
sismischung Basismischung
+3% Weizenkleie Basismischung
+3% Trockenschnitzel Basismischung +3% Trocken-schnitzel-Plus
2 Boxen 2 Boxen 2 Boxen 2 Boxen 6 Ferkel / Box 6 Ferkel / Box 6 Ferkel / Box 6 Ferkel / Box
Box 1 + 5 Box 2 + 6 Box 3 + 7 Box 4 + 8
Die Rezepturen der Basismischungen für die Ferkelabsetz- und Ferkelaufzuchtphase werden in den Tabellen 2 und 3 dargestellt. Die wichtigsten Nährstoffgehalte sind in der Tabelle 4 aufgelistet.

Wetscherek-Seipelt und Wetscherek: Einsatz von Weizenkleie bzw. Trockenschnitzel bei Ferkel
8. BOKU-Symposium Tierernährung 2009 Seite 227
Tabelle 2 Zusammensetzung der Basisfuttermischung des Absetzfutters
Futtermittel Gehalt in % Frischsubstanz Mais 22,655 Triticale 15,000 Erbse 12,000 Sonnenblumenkuchen 10,300 Gerste 10,000 Sojabohne getoastet 8,900 Molkenpulver- süß 6,250 Kartoffeleiweiß 5,900 Molkeneiweißpulver- 45% 3,000 Mineralstoffprämix S 2,010 Rübenmelasse 2,000 Melasseschnitte 12% 1,500 Kohlensaurer Kalk „Grieß“ 0,300 Monocalciumphosphat Aliphos 22,7% 0,110 Natriumchlorid 0,060 LBC-ME 10 0,015
Tabelle 3 Zusammensetzung der Basisfuttermischung des Aufzuchtfutters
Futtermittel Gehalt in % Frischsubstanz Weizen 25,00 Mais 24,49 Erbse 17,50 Sojabohne 12,50 Gerste 8,00 Kürbiskernkuchen 5,20 Kartoffeleiweiß 4,30 Mineralstoffprämix S 3,00 LBC-ME 10 0,01
Datenerhebung Die Ferkel wurden bei der Einstellung und anschließend jede Woche gewogen. Pro Box und Abschnitt wurden die gefressenen Futtermengen wöchentlich ermittelt. Aus der Relation der je Box und Ab-schnitt aufgenommenen Futtermengen zur Summe der Lebendgewichtszunahmen der Tiere je Box wurde die Futterverwertung berechnet.
Behandlungen, Ausfälle und Krankheiten wurden mit Tiernummer, Datum, Gewicht und Ursache fest-gehalten.
Bei der Aufzuchtleistung wurden der Gruppen- und Geschlechtseinfluss mittels zweifacher Varianz-analyse unter der Berücksichtigung der Wechselwirkung von Gruppe und Geschlecht berechnet. Ne-ben den Gruppenmittelwerten werden die Ergebnisse der biometrischen Auswertung als P-Wert ange-geben. Ein P-Wert von 0,05 und kleiner bedeutet, dass sich die Gruppen signifikant unterscheiden.

Wetscherek-Seipelt und Wetscherek: Einsatz von Weizenkleie bzw. Trockenschnitzel bei Ferkel
Seite 228 8. BOKU-Symposium Tierernährung 2009
Tabelle 4 Errechnete wertbestimmende Nährstoffgehalte der Basismischungen
Energie- und Nährstoffgehalt Ferkelabsetzfutter Ferkelaufzuchtfutter ME, MJ/kg 13,0 13,8 Rohprotein, % 19,7 19,5 Rohfett, % 4,8 4,9 Rohfaser, % 5,0 3,6 Rohasche, % 5,9 5,5 Stärke, % 34,2 43,7 Zucker, 8,4 3,3 Lysin, % 1,11 1,05 Methionin, % 0,35 0,32 Methionin und Cystin, % 0,70 0,67 Threonin, % 0,84 0,76 Vitamin A, IE/kg 6800 10200 Vitamin D3, IE/kg 1100 1600 Vitamin E, mg/kg 65 100 Kupfer, mg/kg 6 8
Ergebnisse und Diskussion Der Nährstoffbedarf war bei den vier Gruppen durch die Futtermischungen gedeckt und wird in den Tabellen 5 und 6 dargestellt. Die Energie- und Nährstoffgehalte unterschieden sich zwischen den Gruppen nicht wesentlich. Der Gehalt an Deoxynivalenol lag bei den eingesetzten Testfuttermitteln Weizenkleie, Trockenschnitzel, Trockenschnitzel-Plus jeweils unter der Nachweisgrenze von 35 ppb und somit deutlich unter dem Richtwert für ein Alleinfuttermittel von 900 ppb. Der Gehalt an Zearale-non lag für Weizenkleie und Trockenschnitzel unter der Nachweisgrenze von 15 ppb. Die Proben der Trockenschnitzel-Plus enthielt einen Zearalenongehalt von 88 ppb. Auch dieser liegt unter dem Richt-wert für ein Alleinfutter von 100 ppb (Tabelle 7).
Tabelle 5 Analysenergebnisse Absetzfuttermischungen
V e r s u c h s g r u p p e Merkmal 1 2 3 4
Trockensubstanz g/kg 897 885 887 885 Rohprotein g/kg 204 193 192 191 Rohfett g/kg 50 48 47 46 Rohfaser g/kg 39 45 46 47 Rohasche g/kg 49 49 48 48 Stärke g/kg 406 386 392 393 Zucker g/kg 53 57 57 57 ADF g/kg 56 61 63 61 NDF g/kg 188 169 167 151 ME MJ/kg 14,00 13,47 13,46 13,37

Wetscherek-Seipelt und Wetscherek: Einsatz von Weizenkleie bzw. Trockenschnitzel bei Ferkel
8. BOKU-Symposium Tierernährung 2009 Seite 229
Tabelle 6 Analysenergebnisse Aufzuchtfuttermischungen
V e r s u c h s g r u p p e Merkmal 1 2 3 4
Trockensubstanz g/kg 894 894 896 895 Rohprotein g/kg 207 203 202 202 Rohfett g/kg 58 55 52 53 Rohfaser g/kg 39 38 38 42 Rohasche g/kg 58 59 60 61 Stärke g/kg 393 394 402 381 Zucker g/kg 32 33 35 35 ADF g/kg 58 55 63 63 NDF g/kg 207 190 195 205 ME MJ/kg 13,97 13,91 13,88 13,69
Tabelle 7 Analysenergebnisse der zugesetzten Futtermittel
Merkmal Weizenkleie Trockenschnitzel Trockenschnitzel-Plus
Trockensubstanz g/kg 897 916 920 Rohprotein g/kg 150 150 129 Rohfett g/kg 45 13 11 Rohfaser g/kg 100 161 151 Rohasche g/kg 74 82 102 Stärke g/kg 108 196 181 Zucker g/kg 55 84 84 ADF g/kg 151 192 186 NDF g/kg 486 440 399 ME MJ/kg 10,52 8,07 7,99 Deoxynivalenol
unter der Nachweis-grenze von 35 ppb
unter der Nachweis-grenze von 35 ppb
unter der Nachweis-grenze von 35 ppb
Zearalenon
unter der Nachweis-grenze von 15 ppb
unter der Nachweis-grenze von 15 ppb
88 ppb
Während des Versuches traten keine Ausfälle oder Erkrankungen auf. Bei allen Gruppen konnten keine Durchfallserscheinungen oder Unterschiede in der Kotkonsistenz zwischen den vier Gruppen beobach-tet werden.
Die Differenzen bei der Gewichtsentwicklung zwischen den vier Gruppen waren sowohl in der Absetz-phase als auch in der Aufzuchtperiode nicht signifikant unterschiedlich (Tabelle 8).
Sehr wohl zeigten sich in der Absetzphase (1. und 2. Versuchswoche) tendenziell positive Effekte auf die Gewichtsentwicklung durch den Einsatz von Weizenkleie und Trockenschnitzel. Der Einsatz Tro-ckenschnitzel-Plus führte in dieser Phase zu einer etwas geringeren Gewichtsentwicklung als bei der Kontrollgruppe.
In der Aufzuchtperiode erreichten die Ferkel der Gruppe mit Weizenkleie die gleichen Zuwachsleistun-gen wie in der Kontrollgruppe. Die beiden Gruppen mit Trockenschnitzel lagen auf ähnlichem Niveau,
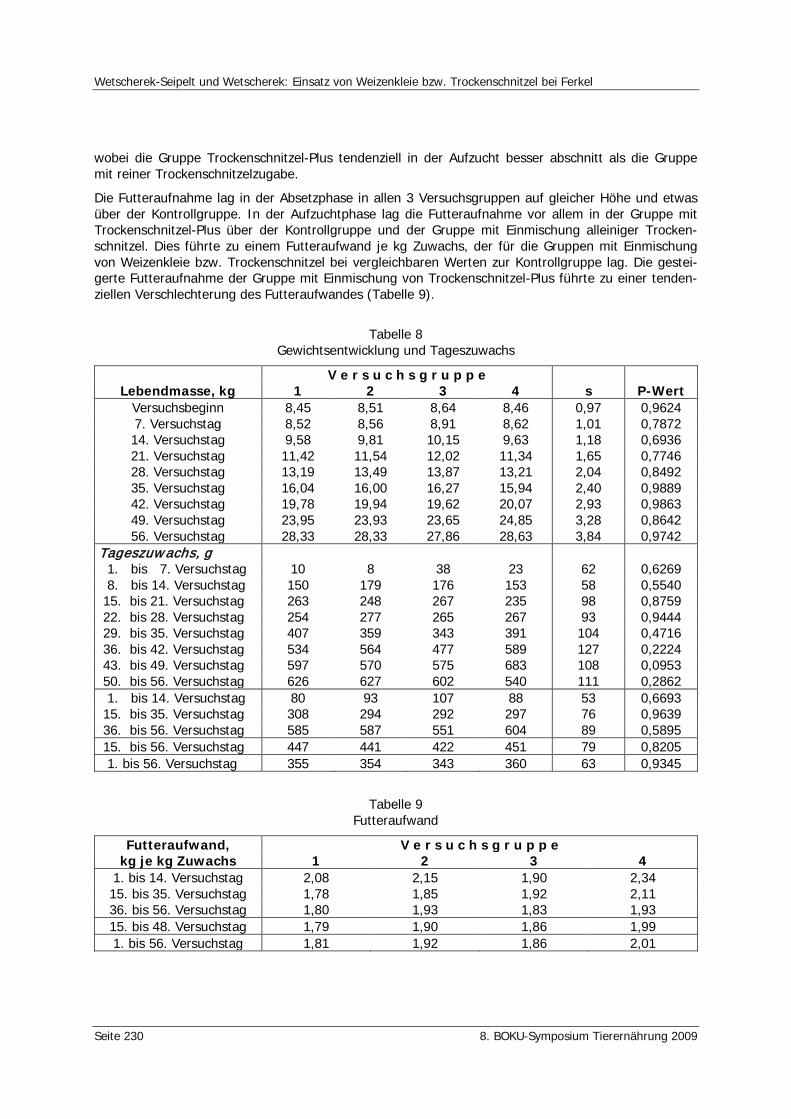
Wetscherek-Seipelt und Wetscherek: Einsatz von Weizenkleie bzw. Trockenschnitzel bei Ferkel
Seite 230 8. BOKU-Symposium Tierernährung 2009
wobei die Gruppe Trockenschnitzel-Plus tendenziell in der Aufzucht besser abschnitt als die Gruppe mit reiner Trockenschnitzelzugabe.
Die Futteraufnahme lag in der Absetzphase in allen 3 Versuchsgruppen auf gleicher Höhe und etwas über der Kontrollgruppe. In der Aufzuchtphase lag die Futteraufnahme vor allem in der Gruppe mit Trockenschnitzel-Plus über der Kontrollgruppe und der Gruppe mit Einmischung alleiniger Trocken-schnitzel. Dies führte zu einem Futteraufwand je kg Zuwachs, der für die Gruppen mit Einmischung von Weizenkleie bzw. Trockenschnitzel bei vergleichbaren Werten zur Kontrollgruppe lag. Die gestei-gerte Futteraufnahme der Gruppe mit Einmischung von Trockenschnitzel-Plus führte zu einer tenden-ziellen Verschlechterung des Futteraufwandes (Tabelle 9).
Tabelle 8 Gewichtsentwicklung und Tageszuwachs
V e r s u c h s g r u p p e Lebendmasse, kg 1 2 3 4 s P-Wert
Versuchsbeginn 8,45 8,51 8,64 8,46 0,97 0,9624 7. Versuchstag 8,52 8,56 8,91 8,62 1,01 0,7872 14. Versuchstag 9,58 9,81 10,15 9,63 1,18 0,6936 21. Versuchstag 11,42 11,54 12,02 11,34 1,65 0,7746 28. Versuchstag 13,19 13,49 13,87 13,21 2,04 0,8492 35. Versuchstag 16,04 16,00 16,27 15,94 2,40 0,9889 42. Versuchstag 19,78 19,94 19,62 20,07 2,93 0,9863 49. Versuchstag 23,95 23,93 23,65 24,85 3,28 0,8642 56. Versuchstag 28,33 28,33 27,86 28,63 3,84 0,9742
Tageszuwachs, g 1. bis 7. Versuchstag 10 8 38 23 62 0,6269 8. bis 14. Versuchstag 150 179 176 153 58 0,5540 15. bis 21. Versuchstag 263 248 267 235 98 0,8759 22. bis 28. Versuchstag 254 277 265 267 93 0,9444 29. bis 35. Versuchstag 407 359 343 391 104 0,4716 36. bis 42. Versuchstag 534 564 477 589 127 0,2224 43. bis 49. Versuchstag 597 570 575 683 108 0,0953 50. bis 56. Versuchstag 626 627 602 540 111 0,2862 1. bis 14. Versuchstag 80 93 107 88 53 0,6693 15. bis 35. Versuchstag 308 294 292 297 76 0,9639 36. bis 56. Versuchstag 585 587 551 604 89 0,5895 15. bis 56. Versuchstag 447 441 422 451 79 0,8205 1. bis 56. Versuchstag 355 354 343 360 63 0,9345
Tabelle 9 Futteraufwand
Futteraufwand, kg je kg Zuwachs
V e r s u c h s g r u p p e 1 2 3 4
1. bis 14. Versuchstag 2,08 2,15 1,90 2,34 15. bis 35. Versuchstag 1,78 1,85 1,92 2,11 36. bis 56. Versuchstag 1,80 1,93 1,83 1,93 15. bis 48. Versuchstag 1,79 1,90 1,86 1,99 1. bis 56. Versuchstag 1,81 1,92 1,86 2,01

Wetscherek-Seipelt und Wetscherek: Einsatz von Weizenkleie bzw. Trockenschnitzel bei Ferkel
8. BOKU-Symposium Tierernährung 2009 Seite 231
Zusammenfassung • Der Einsatz von Weizenkleie, Trockenschnitzel bzw. Trockenschnitzel-Plus in der Höhe von 3%
hatte keinen Einfluss auf den Gesundheitsstatus und die Kotkonsistenz der Ferkel.
• Der Einsatz von 3% Trockenschnitzel-Plus führte zu einer Erhöhung der Futteraufnahme. So-mit können negative sensorische Effekte des Carbonatationskalk in der vorgenommenen Do-sierung ausgeschlossen werden.
• Die Gewichtsentwicklung wurde durch die Einmischung ballaststoffhältiger Futtermittel nicht negativ beeinflusst. Ein positiver Effekt auf die Leistung und Tiergesundheit durch die Einmi-schung ballaststoffhältiger Futtermittel, wie er in der Literatur beschrieben wird, konnte für die Absetzphase für Weizenkleie bzw. Trockenschnitzel tendenziell bestätigt werden.
Summary • The use of wheat bran, dried sugar beet pulp as well as dried sugar beet pulp-plus in the
amount of 3 percent had none influence on health status and faeces structure of piglets.
• The use of 3 percent of dried sugar beet pulp-plus led to an increase of the feed intake. Con-sequently, negative sensory effects of the carbonation lime can be excluded in the performed dosage.
• The weight gain development was not influenced negatively by feed with dietary fibre. A posi-tive effect on performance and animal health through the addition dietary fibre, as described in literature, could be confirmed tendency for weaned piglets for wheat bran as well as dried sugar beet pulp.
Autorenanschrift
Ao. Univ.-Prof. Dr. Wolfgang Wetscherek Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Department für Lebensmittelwissenschaften und –technologie Universität für Bodenkultur Wien Gregor Mendel-Straße 33, 1180 Wien E-Mail: [email protected]

Maevis et al.: Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen
Seite 232 8. BOKU-Symposium Tierernährung 2009
Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen
G. Maevis1, C. Koch2, F.-J. Romberg3, K. Landfried2, J. Kühl4, H. Steingaß5 und K.-H. Südekum1 1 Institut für Tierwissenschaften, Abteilung Tierernährung, Rheinische Friedrich-
Wilhelms-Universität Bonn 2 Lehr- und Versuchsanstalt für Viehhaltung Hofgut Neumühle, Münchweiler an der
Alsenz 3 Dienstleistungszentrum Ländlicher Raum (DLR) Westpfalz 4 LUFA Speyer 5 Universität Hohenheim
Einleitung Die chemische Verbindung Glycerin, ein dreiwertiger Alkohol mit der Summenformel C3H5(OH)3, kommt in allen pflanzlichen und tierischen Fetten und Ölen als alkoholischer Esterbestandteil vor. Gly-cerin fällt heute in großen Mengen, je Tonne Biodiesel 100-110 kg Glycerin, als klassisches Nebenpro-dukt der Biodieselherstellung an. Glycerin ist ein wertvoller Grundstoff für die chemische, pharmazeu-tische und technische Industrie.
Hauptrohstoff für die Biodieselherstellung in Deutschland ist Raps, vielmehr das aus der Rapssaat gepresste oder extrahierte Rapsöl. Chemisch exakt wird aus Rapsöl hergestellter Biodiesel als Rapsöl-methylester (RME) bezeichnet. Seit Mitte der 90er Jahre hat sich der Absatz von Biodiesel bis zum Jahr 2005 fast verfünfzigfacht und ist auf 3,3 Mio. t im Jahr 2007 gestiegen (UFOP 2005; VDB 2009). Diese Situation bewirkte einen Angebotsüberschuss von Glycerin am Markt. Der Markt von Glycerin ist relativ eng, da es von den verschiedenen Industriezweigen mengenmäßig nur begrenzt aufgenommen werden kann. Der Angebotsüberschuss führte zu Absatzproblemen und einem schnellen Preisverfall von Glycerin. Deshalb mussten neue Absatzgebiete erschlossen werden. Ein Teil des Angebotsüber-schusses wird in der Landwirtschaft abgesetzt. Zum einen wird Glycerin als Co-Substrat in der Biogas-produktion genutzt. Zum anderen setzt die Tierernährung Glycerin in der Fütterung und der Herstel-lung von Mischfuttermitteln ein.
Der Einsatz von Glycerin in Rationen für Wiederkäuer ist bereits in verschiedenen Fütterungsversu-chen erprobt worden. Einige Versuche haben gezeigt, dass Glycerin die Futteraufnahme positiv beein-flussen kann. Im September 2008 wurde an der Lehr- und Versuchsanstalt (LVA) für Viehhaltung Hof-gut Neumühle in D-67728 Münchweiler an der Alsenz ein Fütterungsversuch durchgeführt, mit dem Ziel die Auswirkungen einer Glycerinzulage in einer Gesamtmischration (TMR) auf die Futteraufnahme sowie auf die in vitro-Pansenfermentation und Leistungskenngrößen zu untersuchen.
Material und Methoden Der Fütterungsversuch zum Einsatz von Rohglycerin in Rationen für hochleistende Milchkühe gliederte sich in eine 14-tägige Vorperiode zur Anfütterung und eine 12-wöchige Versuchsperiode. Zu Beginn des Fütterungsversuches in der 37. Kalenderwoche (KW) 2008 standen 60 laktierende Kühe zu Verfü-

Maevis et al.: Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen
8. BOKU-Symposium Tierernährung 2009 Seite 233
gung. Die Tiere wurden nach den Kriterien Laktationsnummer, Laktationstag und Milchleistung in zwei vergleichbare Gruppen aufgeteilt. Um die Umwelteffekte und mögliche stallbauliche Effekte zu mini-mieren, erfolgte nach drei und nach neun Wochen ein Platztausch der Gruppen. Außerdem wurde nach sechs Wochen die Ration getauscht, d. h. die bisherige Kontrollgruppe bekam jetzt die Ration mit der Glycerinzulage. Im Versuchszeitraum ausscheidende Tiere wurden durch abkalbende Tiere ersetzt.
Die Rationen in der Kontroll- und Glyceringruppe waren gleich konzipiert und bestanden aus Maissila-ge, Grassilage, Heu, einer Getreidemischung (70 % Wintergerste und 30 % Weizen), Sojaextraktions-schrot, Körnermais, Rapsextraktionsschrot, Futterkalk, Mineralsalz und Harnstoff. Die Zusammenset-zung der gefütterten Rationen ist aus Tab. 1 zu entnehmen. Die Versuchsration unterschied sich von der Kontrollration nur durch die Glycerinzulage. Die Futtervorlage erfolgte zweimal täglich (morgens: 5:30 Uhr, nachmittags: 15:30 Uhr) in Fress-Wiegetröge, wobei morgens 40 % und nachmittags 60 % der Tagesmenge gefüttert wurde. Der selbstfahrende Horizontalmischwagen Typ R.M.H. 350-CS (Fir-ma Lachish Industries Ltd., Israel) stellt die TMR mit vier horizontalen Mischschnecken her. Nach dem Befüllen und Mischen erhielt zunächst die Kontrollgruppe die Ration. Anschließend wurden für die Fütterung der Versuchsgruppe 1,25 kg Rohglycerin pro Tier und Tag in die TMR eingemischt. Das von der Firma Inter-Harz GmbH (D-25314 Elmshorn) bezogene Rohglycerin hatte einen deklarierten Glyce-ringehalt von >80 %. Mit einer Dosierpumpe wurde es aus dem Kunststofftank in den Mischbehälter gepumpt.
Die Futteraufnahme wurde über 24 Stunden erfasst. Um 24 Uhr eines jeden Tages wurde die FM-Aufnahme zu einer Tagessumme addiert und in einer tierindividuellen Liste gespeichert. Um die auf-genommene TM bestimmen zu können, wurden tgl. homogene Proben von jeder TMR aus den Wiege-trögen gezogen und zu einer Wochenprobe zusammengestellt. Anhand der Wochenproben wurde die Trockensubstanz (TS) der Kontroll- und der Versuchsration bestimmt.
Während des gesamten Versuchzeitraumes erfolgten repräsentative Futtermittelprobenahmen der gefütterten Rationskomponenten. Insgesamt wurden die Grobfuttermittel Mais-, Grassilage und Heu viermal beprobt. Von den anderen Rationskomponenten wurden je Charge zwei Proben gezogen.
Tabelle 1: Zusammensetzung der Ration
Versuchsgruppen (Kontrolle und Glycerin) Nr. Komponenten (% i. d. TM) 1 Maissilage 32,2 2 Grassilage 16,1 3 Heu 6,9 4 Getreidemischung 16,1 5 Sojaextraktionsschrot 6,9 6 Körnermais 15,7 7 Rapsextraktionsschrot 4,7 8 Futterkalk 0,7 9 Mineralfutter 0,3 10 Harnstoff 0,5 11 Reinglycerin 5 % Glycerin je Kuh u. Tag in der Versuchsgruppe
Die LVA für Viehhaltung Hofgut Neumühle ist dem Landeskontrollverband (LKV) Rheinland-Pfalz e. V. angeschlossen. Zusätzlich zur monatlichen Milchkontrolle wurden wöchentliche Probenahmen Sonntag Abend und Montag Morgen durchgeführt. Aus den abends und morgens in den Kontrollen abgelese-nen Milchmengen wurde die Summe kalkuliert, welche für die Milchleistung des Einzeltieres in der jeweiligen Versuchswoche herangezogen wurde. Die Proben wurden zeitnah in das Labor des LKV in

Maevis et al.: Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen
Seite 234 8. BOKU-Symposium Tierernährung 2009
Thalfang verbracht und dort analysiert. Die Analyse umfasste die Milchinhaltsstoffe Fett und Eiweiß, somatische Zellzahl, fettfreie TM sowie den Harnstoff- und Acetongehalt.
Die Tiere wurden wöchentlich gewogen. Im 14-tägigen Abstand wurde die Körperkondition der Herde mittels Body-Conditon-Score (BCS) eingestuft (EDMONDSON et al. 1989). Ebenfalls 14-tägig wurde die Rückenfettdicke (RFD) der gesamten Herde nach der Methode von STAUFENBIEL (1997) per Ultra-schallgerät Aloka SSD 500 (Fa. Aloka GmbH, D-40670 Meerbusch) gemessen.
Am Institut für Tierwissenschaften, Abteilung Tierernährung der Rheinischen Friedrich-Wilhelms-Universität in Bonn wurden zusätzlich zum Fütterungsversuch an der LVA für Viehhaltung Hofgut Neumühle in vitro-Untersuchungen durchgeführt.
Die Fermentationseigenschaften der eingesetzten Futtermittel wurden mit Hilfe der Gasbildung im Hohenheimer Futterwerttest (HFT) charakterisiert. Die Inkubation der Futtermittel mit Pansensaft erfolgte gemäß der Methode nach MENKE UND STEINGASS (1986). Von den Rationskomponenten 1 bis 3 wurden je vier Proben, von Rationskomponente 4 wurden zwei Proben und von jeder Charge Kraft-futter (Komponenten 5 bis 7) wurde eine Probe inkubiert. Für jede Futterprobe wurden drei Präzisi-onsglasspritzen mit Kapillarrohr-Ansatz vorbereitet. Nach 4, 8, 12, 16, 24, 32, 48, 56, 72, 80 und 96 h wurde das Volumen jeder Präzisionsglasspritze auf 0,5 ml genau abgelesen und dokumentiert.
Aus den gemahlenen Futterproben wurde die TMR zusammengestellt, die im Fütterungsversuch an der LVA für Viehhaltung Hofgut Neumühle gefüttert wurde. Für den Anteil der Rationskomponenten Futterkalk, Mineral- und Harnstoff wurde ein gemahlenes Mineralfutter der Firma Höveler eingemischt. Um die Qualität und Mischgenauigkeit der im Labor gemischten TMR beurteilen zu können, wurden in den einzeln vorgemischten Rationskomponenten und in der gemischten TMR die Gehalte an Rohprote-in (XP) in der FM bestimmt. In den Rationen wurde im modifizierten HFT nach STEINGASS et al. (2001) der Gehalt an nutzbarem Rohprotein (nXP) geschätzt und die flüchtigen FS, als Endprodukte der Fermentation, bestimmt. Ziel war es, die Auswirkungen unterschiedlicher Glycerinzulagen in der Ration auf die nXP-Synthese und die Endprodukte der Fermentation zu untersuchen. Die Inkubation erfolgte über 24 h, getestet wurden vier Varianten, eine ohne Glycerinzulage und drei mit unterschied-lichen Glycerinzulagen:
1.) ohne Glycerinzulage (Kontrolle) 2.) 4,4 mg Glycerinzulage = 500 g Reinglycerin im Fütterungsversuch (LG) 3.) 8,8 mg Glycerinzulage = 1000 g Reinglycerin im Fütterungsversuch (MG) 4.) 13,2 mg Glycerinzulage = 1500 g Reinglycerin im Fütterungsversuch (HG)
Für jede Variante wurden parallel drei Präzisionsglasspritzen in dreifacher Wiederholung vorbereitet. Zum einen zur nXP-Bestimmung nach 8 und 24 h und zum anderen zur Bestimmung der flüchtigen FS nach 24 h. Die Destillation der Proben erfolgte mit einem automatischen Schnelldestilliersystem Vapo-dest 50 s (Fa. Gerhardt, D-53639 Königswinter) mit Titration, das auch für die N-Bestimmung nach Kjeldahl benutzt wird. Bis zur Destillation wurden die Proben gekühlt aufbewahrt.
Die flüchtigen FS in den Proben wurden mit dem Gaschromatographen (GC) AutoSys (Fa. Perkin El-mer, Massachusetts, USA) bestimmt.
Ergebnisse und Diskussion Das im Fütterungsversuch eingesetzte Rohglycerin von der Firma Interharz wurde mit einem deklarier-ten Glyceringehalt von >80 % geliefert. Die LUFA in Speyer hat folgende Inhaltsstoffe im Rohglycerin ermittelt: Glycerin 83,3 %, Wasser 9,8%, Na 1,86 %, K < 0,01 %.
Die gefütterte Ration hat einen Energiegehalt von 7,2 MJ NEL/kg TM und einen nXP-Gehalt von 159 g/kg TM. Sie ist ausgelegt für eine Milchleistung von 35,5 kg Milch nach NEL und 35,2 kg Milch

Maevis et al.: Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen
8. BOKU-Symposium Tierernährung 2009 Seite 235
nach nXP, bei einer unterstellten TM-Aufnahme von 21,7 kg. Hinsichtlich der RNB liegt die berechnete Ration im positiven Bereich, mit einem RNB-Wert von 24 g. Der XF-Gehalt beträgt 15,7 % in der TM (Tab. 2).
Tabelle 2: Rationsberechnung der Kontrollration über den Versuchszeitraum
Kontrolle Futtermittel %
in TM FM (kg)
TM (kg)
NEL (MJ)
nXP (g)
RNB (g)
XF (g)
XS (g)
XP (g)
XL (g)
Maissilage 32,2 20,71 7,00 44,3 917 -49 1519 1750 616 203 Grassilage 16,1 7,87 3,50 22,2 502 14 868 0 590 137
Heu 6,9 1,61 1,50 7,7 155 -6 503 0 118 31 Getreidemischung 16,1 4,05 3,50 29,3 603 -16 201 n.a. 503 132
Sojaextraktionsschrot 6,9 1,71 1,50 12,8 442 53 75 0 771 43 Körnermais 15,7 3,90 3,40 32,1 598 -42 96 n.a. 334 174
Rapsextraktionsschrot 4,6 1,12 1,00 7,1 235 24 143 0 387 40 Futterkalk 0,7 0,15 0,15 0,0 0 0 0 0 0 0
Mineralfutter 0,3 0,07 0,07 0,0 0 0 0 0 0 0 Harnstoff 0,5 0,10 0,10 0,0 0 46 0 0 288 0 Summe 100 41,3 21,7 155,4 3452 24 3405 1750 3606 759
Ration pro kg TM 7,2 158,9 1,1 156,8 80,6 166,0 34,9 ECM (680 kg LM) 35,5 35,2
Beim Futterverzehr besteht ein geringer Unterschied zwischen Kontroll- und Glyceringruppe. Kühe in der Glyceringruppe haben gegenüber Kühen in der Kontrollgruppe 0,22 kg TM/Tier und Tag mehr gefressen. Dieser geringe Unterschied konnte statistisch nicht abgesichert werden. Die Tiere der Kon-trollgruppe erzielten mit 0,14 kg Milch/Tier und Tag eine höhere Milchleistung als die Tiere der Glyce-ringruppe. Durch die Korrektur der Milchmenge auf energiekorrigierte Milch (ECM) mit 4 % Fett- und 3,4 % Eiweißgehalt verringert sich der Leistungsunterschied auf 0,12 kg Milch/Tier und Tag. Die Fett- und Eiweißgehalte der Milch liegen in der Glyceringruppe um 0,01 Prozentpunkte niedriger als in der Kontrollgruppe. Mengenmäßig gibt es keinen Unterschied zwischen Kontroll- und Glyceringruppe in den Milchinhaltstoffen Fett und Eiweiß. Die Acetonuntersuchung zeigt, dass der Acetongehalt [mmol/l] in der Milch der Glyceringruppe etwa 30 % niedriger ist als in der Kontrollgruppe. Diese Unterschiede sind alle nicht statistisch zu sichern.
Nur der Harnstoffgehalt in der Milch charakterisiert einen statistisch abgesicherten Unterschied zwi-schen den Versuchsvarianten. Die Kontrollgruppe hat einen um 41,53 mg/l höheren Harnstoffgehalt als die Glyceringruppe. Dieser Unterschied ist mit p < 0,0001 höchst signifikant. Tendenziell sind die Tiere in der Glyceringruppe etwa 3,3 kg schwerer als in der Kontrollgruppe.
Aus den Ergebnissen des Fütterungsversuches geht hervor, dass sich die TM-Aufnahmen der Kontroll- und Glyceringruppe trotz der Rohglycerinzulage in Höhe von 1250 g bzw. 1041 g Reinglycerin pro Tier und Tag zu der TMR der Glyceringruppe nicht signifikant unterscheiden. Dieses Ergebnis stimmt mit der Mehrzahl jüngerer Forschungsergebnisse zum Glycerineinsatz in der Fütterung von Milchkühen überein, die in den Jahren 2000 bis 2008 durchgeführt worden sind (DEFRAIN et al. 2004; KRISTEN-SEN UND RAUN 2007; HARZHEIM et al. 2008; DONKIN UND DOANE 2008; WANG et al. 2008). Andere Untersuchungen berichten von einer Steigerung der Futteraufnahme durch die Glycerinzulage oder –ergänzung (BODARSKI et al. 2005; ENGELHARDT et al. 2006; MAHLKOW-NERGE 2006; ALERT UND FRÖHLICH 2008). Diese Ergebnisse sind nicht direkt miteinander vergleichbar, weil Versuchsbedingun-gen, Versuchsdauer und Tierzahlen in den Versuchen oft stark voneinander abweichen. Mögliche Ur-sachen für die widersprüchlichen Effekte des Glycerins in der Fütterung können einerseits unterschied-

Maevis et al.: Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen
Seite 236 8. BOKU-Symposium Tierernährung 2009
lich hohe Einsatzmengen und Qualitäten sein. Andererseits unterscheiden sich in den Versuchen die gefütterten Rationen in den Komponenten, den Inhaltsstoffen und im Grob- zu Kraftfutterverhältnis.
Die Glycerinzulage von 1041 g pro Tier und Tag hat die Milchmenge und –inhaltsstoffe nicht signifi-kant beeinflusst und den Harnstoffgehalt in der Milch signifikant gesenkt. Diese Reaktionen auf eine Glycerinzulage in der Fütterung von Milchkühen bestätigen mehrere Untersuchungen (DEFRAIN et al. 2004; DONKIN UND DOANE 2008; HARZHEIM et al. 2008). HARZHEIM et al. (2008) führen den geringe-ren Harnstoffgehalt bei Glycerinfütterung auf die beinahe quantitative Verdrängung der Grundration zurück und begründen damit die ausgebliebene Wirkung von Glycerin auf die Futteraufnahme und die Leistung.
Die Reduzierung des Harnstoffgehaltes kann auch andere Ursachen haben. Das mit dem Futter aufge-nommene Protein gelangt zu 15-30 % als pansenstabiles Protein (UDP) unabgebaut in den Dünn-darm. Die restlichen 70-85 % stehen nach dem Abbau zu Peptiden und Aminosäuren bis hin zu Am-moniak der mikrobiellen Proteinsynthese zu Verfügung. Limitierender Faktor für die mikrobielle Prote-insynthese ist die Energie. Überschüssiges Ammoniak wird in der Leber entgiftet und zu Harnstoff umgewandelt, der über Harn und Milch ausgeschieden wird. Glycerin als Energieträger erhöht die Energieaufnahme. Dadurch steht für die Mikroorganismen mehr Energie zur Proteinsynthese bereit. Die Nutzung des Proteins wird verbessert und die Ammoniakverluste reduziert. Eine weitere Möglich-keit ist, dass durch die Glycerinzulage die Passagegeschwindigkeit des Futters gesteigert wird. Eine schnellere Passage hat einen geringeren Proteinabbau zur Folge, so dass weniger Harnstoff gebildet wird.
Das verfütterte Glycerin wurde zusätzlich zur Ration dosiert, somit wird der Proteingehalt dieser Ration um etwa 10 g/kg TM niedriger sein, womit sich ein RNB-Wert von etwa 0 bis –4 g N je Tier und Tag ergibt. Diese Veränderungen in der ruminalen Stickstoffbilanz können ebenfalls als Erklärung für die niedrigen Harnstoffwerte in der Glyceringruppe gesehen werden.
Die Kontrollgruppe hat im Mittel einen Harnstoffgehalt von 242,78 mg/l und die Glyceringruppe einen von 201,25 mg/l. Die Reduzierung des Harnstoffgehaltes durch die Glycerinzulage entlastet den Stoff-wechsel der Leber. Da der Gewichtsunterschied nur eine Tendenz zeigt, kann angenommen werden, dass die Glycerinzulage keine direkte Auswirkung auf die Körperkondition im durchgeführten Versuch hat.
Tabelle 3: Mittelwert und Standardabweichung (s) der Gasbildung (Gb) (ml/200 mg T) in 24 h, mit Korrekturfaktor (KF), korrigierter Gb und geschätzter NEL im 1. Lauf
Präzisionsglasspritze Mittelwert s KF Gb 24 korr. MJ NEL [MJ/kg TM] Ration 1 2 3
Kontrolle 51,7 53,9 54,7 53,4 1,5 1,07 57,2 7,01 LG 53,7 52,9 53,2 53,2 0,4 1,07 57,0 6,99 MG 55,0 54,3 54,8 54,7 0,4 1,07 58,5 7,17 HG 52,3 54,3 53,8 53,5 1,0 1,07 57,2 7,02
Tabelle 4: Mittelwert und Standardabweichung (s) der Gasbildung (Gb) (ml/200 mg T) in 24 h, mit Korrekturfaktor (KF), korrigierter Gb und geschätzter NEL im 2. Lauf
Präzisionsglasspritze Mittelwert s KF Gb 24 korr. MJ NEL
[MJ/kg TM] Ration 1 2 3 Kontrolle 51,5 52,6 54,2 52,8 1,4 1,08 57,0 6,99
LG 53,1 52,8 53,9 53,3 0,6 1,08 57,5 7,06 MG 54,4 54,7 53,4 54,2 0,7 1,08 58,5 7,17 HG 53,5 53,5 53,5 53,5 0,0 1,08 57,8 7,08

Maevis et al.: Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen
8. BOKU-Symposium Tierernährung 2009 Seite 237
Auf eine statistische Auswertung der in vitro-Untersuchungen wurde aus zeitlichen Gründen verzichtet. Während des gesamten Versuchszeitraumes weisen alle Rationskomponenten gleichmäßige Fermenta-tionseigenschaften auf. Die im Labor gemischte Kontrollration hat einen Gehalt von 155 g XP/kg TM und 27 g XL/kg TM. Die in den Tabellen 4 und 5 dargestellten Energiegehalte zeigen keine großen Abweichungen voneinander. Im Mittel der beiden Läufe hat die Kontrollration einen Energiegehalt von 7 MJ NEL/kg TM, die Ration mit der geringen Glycerinzulage (LG) einen von 7,02 MJ NEL/kg TM, die Ration mit der mittleren Glycerinzulage (MG) einen von 7,17 MJ NEL/kg TM und die Ration mit der hohen Glycerinzulage (HG) einen von 7,05 MJ NEL/kg TM.
Fazit Unterschiede zwischen den Versuchsvarianten in Futteraufnahme und Leistungskenngrößen konnten nicht statistisch abgesichert werden. Die vor Versuchsbeginn angenommene verzehrsfördernde und gleichzeitig leistungtungssteigernde Wirkung einer Glycerinzulage, die aus den Versuchergebnissen von ENGELHARDT et al. (2006), sowie von MAHLKOW-NERGE (2006) und aus der im Jahr 2008 von ALERT UND FRÖHLICH durchgeführten Studie resultierte, ist im vorliegenden Versuch ausgeblieben.
Die Glycerinzulage hatte aber auch keine negativen Einflüsse auf die untersuchten Parameter. Im Gegenteil, in der Fütterungsvariante mit Glycerin wurde eine höchst signifikante Reduzierung des Milchharnstoffgehaltes beobachtet. Die gefütterte Ration war mit einem Milcherzeugungswert > 30 kg für hochleistend laktierende Kühe ausgelegt. Diese basierte auf hohen Grobfutterqualitäten der Mais- und Grassilagen, kombiniert mit Kraft-, Ausgleich- und Mineralfuttermitteln zu einer gut fermentierba-ren und schmackhaften TMR.
Abschließend ist festzuhalten, dass die Zulage von Rohglycerin in der Fütterung hochleistender Milch-kühe unter den gegebenen Bedingungen keine Auswirkungen auf die Futteraufnahme und Leistungs-kenngrößen hat und somit nicht erforderlich und empfehlenswert ist.
Literatur
ALERT, H.-J. UND FRÖHLICH, B. (2008): Propylenglykol bzw. Glycerin als TMR - Bestandteil oder Futterabrufkomponente bei Milchkühen. In: Forum angewandte Forschung in der Rinder- und Schweinefütterung, Fulda, Tagungsunterlage 09./10.04.2008: 62-66.
BODARSKI, R., WERTELECKI, T., BOMMER, F. UND GOSIEWSKI, S. (2005): The changes of metabolic status and lactation per-formance in dairy cows under feeding TMR with glycerin supplement at periparturient period. Electronic Journal of Polish Agri-cultural Universities, Animal Husbandary, 8: 1-9. Online in Internet: URL: http://www.ejpau.media.pl/volume8/issue4/art-22.html [Stand: 03.10.08]
DEFRAIN, J. M., HIPPEN, A. R., KALSCHEUR K. F. UND JARDON, P. W. (2004): Feeding Glycerol to Transition Dairy Cows: Effects on Blood Metabolites and Lactation Performance. J. Dairy Sci. 87: 4195-206.
DONKIN, S. S. UND DOANE, P. H. (2008): Glycerol from Biodiesel Production: The New Corn for Dairy Cattle. In: Southwest Nutrition and Management Conference Arizona 2008. Online in Internet: URL: http://animal.cals.arizona.edu/swnmc/2008/08proceedings-/14%20Donkin%20Glycerol%20from%20Biodiesel%20Production.pdf [Stand: 05.10.08]
EDMONDSON, A. J., LEAN, L. J., WEAVER, L. D., FARVER, T. UND WEBSTER, G. (1989): A body condition chart for Holstein dairy cows. J. Dairy Sci. 72: 68-78.
ENGELHARDT, T., RIEMANN E., ANDERT G., ZWARWEL H., MEYER A., STAUFENBIEL R., GENZEL A., KANITZ W. und STÜRMER G. (2006): Vergleich des Einsatzes von Propylenglykol und Glycerin in Rationen für Hochleistungskühe. Versuchsbericht, Landesanstalt für Landwirtschaft, Forsten und Gartenbau Sachsen Anhalt, Zentrum für Tierhaltung und Technik, 39606 Iden.
HARZHEIM, J., STEINGAß, H. UND DROCHNER, W. (2008): Wirkung von Glycerinzulagen auf Futteraufnahme und Leistung bei Milchkühen. 120. VDLUFA-Kongress: 38.

Maevis et al.: Auswirkungen von Glycerin in einer Gesamtmischration (TMR) für hochleistende Milchkühe auf Futteraufnahme, in vitro-Pansenfermentation und Leistungskenngrößen
Seite 238 8. BOKU-Symposium Tierernährung 2009
KRISTENSEN, N. B. UND RAUN, B. M. L. (2007): Ruminal fermentation, portal absorption, and hepatic metabolism of glycerol infused into the rumen of lactating dairy cows. In: Energy and Protein Metabolism and Nutrition – Proceed-ings of the 2nd International Symposium on Energy and Protein Metabolism and Nutrition, EAAP Publication, No. 124: 355-356.
MAHLKOW-NERGE, K. (2006): Vergleich des Einsatzes von Propylenglykol und Glycerin in Rationen für Hochleistungskühe. In: Forum angewandte Forschung in der Rinder- und Schweinefütterung, Fulda, Tagungsunterlage 05./06.04.2006. 30-34
MENKE, K. H. UND STEINGASS, H. (1986): Schätzung des energetischen Futterwerts aus der in vitro mit Pansensaft bestimmten Gasbildung und der chemischen Analyse. I. Untersuchungen zur Methode. Übers. Tierernährung 14: 251-270.
STAUFENBIEL, R. (1997): Konditionsbeurteilung von Milchkühen mit Hilfe der sonographischen Rückenfettdickenmes-sung. Prakt. Tierarzt, Coll. Vet. 27: 87-92.
STEINGASS H., NIBBE D., SÜDEKUM K.-H., LEBZIEN P. UND SPIEKERS H. (2001): Schätzung des nXP-Gehaltes mit Hilfe des modifizierten Hohenheimer Futterwerttests und dessen Anwendung zur Bewertung von Raps- und Sojaextraktionsschroten. 113. VDLUFA-Kongress: 114.
UFOP (2005): Statusbericht Biodieselproduktion und Vermarktung in Deutschland 2005. Union zur Förde-rung von Öl- und Proteinpflanzen e. V., Berlin. Online in Internet: URL: http://www.ufop.de/publikationen_allgemein.php#Statusbericht [Stand: 27.09.08]
VDB, Verband der Deutschen Biokraftstoffindustrie e. V. (2009): Biodiesel/Marktdaten. Online in Internet: URL: http://www.biokraftstoffverband.de/de-/biodiesel/marktdaten.html [Stand: 20.02.2009]
WANG, C., LIU, Q., YANG, W. Z., HUO, W. J., DONG, K. H., HUANG, Y. X., YANG, X. M., UND HE, D. C. (2008): Effects of glycerol on lactation performance, energy balance and metabolites in early lactation Holstein dairy cows. Anim. Feed Sci. Technol., doi: 10.1016/j.anifeedsci.2008.10.009
Autorenanschrift
Prof. Dr. K.-H. Südekum Institut für Tierwissenschaften Endenicher Allee 15 D-53115 Bonn E-Mail: [email protected] Dipl.-Ing. agr. Christian Koch LVA für Viehhaltung Hofgut Neumühle D-67728 Münchweiler E-Mail: [email protected]

Vielhaber et al.: Einsatz von Oligogalakturoniden zur Reduzierung des Absetzdurchfalles
8. BOKU-Symposium Tierernährung 2009 Seite 239
Einsatz von Oligogalakturoniden zur Reduzierung des Absetz-durchfalles
B. Vielhaber1, K. Zitterl-Eglseer2, M. Gallnböck1, A. Tichy3 und W. Hagmüller1 1 LFZ Raumberg-Gumpenstein, Institut für Biologische Landwirtschaft und Biodiversi-
tät der Nutztiere 2 Veterinärmedizinische Universität Wien, Institut für Angewandte Botanik und Phar-
makognosie 3 Veterinärmedizinische Universität Wien, Institut für Medizinische Physik und Biosta-
tistik
Einleitung Absetzdurchfall einhergehend mit verminderten Lebendmassezunahmen gefährdet sowohl die Tierge-sundheit, als auch die Wirtschaftlichkeit der Ferkelproduktion (STEINER, 2006). Das Absetzen bedeu-tet aufgrund von Trennung von der Muttersau, abrupter Futterumstellung, Einstallung in eine neue Umgebung und Neugruppierung großen Stress für die Ferkel. Infolgedessen kippt das Gleichgewicht der Mikroflora im Darm in Richtung Pathogene (FULLER, 1989; PLUSKE, 2001; ROLLE u. MAYR, 2007).
Der wichtigste infektiöse Erreger des Absetzdurchfalles bei Ferkeln ist E. coli. Die Colidiarrhoe wird durch Adhäsion des Bakteriums an das Wirtsgewebe ausgelöst. Die Anheftung wird durch Lektine, die sich auf der Oberfläche von infektiösen Organismen befinden, vermittelt. Diese binden an bestimmte Kohlenhydrate auf der Oberfläche des Wirtsgewebes (SHARON u. OFEK, 2000). Sind sie einmal ge-bunden, können sich die Bakterien vermehren, Toxine ausschütten, das Tier infizieren und klinische Erkrankungen auslösen (PUSZTAI, 1992; KASTNER et al., 2004).
Da die Anzahl antibiotikaresistenter Colistämme steigt, ist eine Prophylaxe des Absetzdurchfalles mit Chemotherapeutika abzulehnen (ROLLE u. MAYR, 2007). Die Resistenzproblematik war der Anlass für ein allgemeines Verbot antibiotischer Leistungsförderer (aLF) in der EU ab 2006 (VERORDNUNG (EG) Nr. 1831/2003 und 1334/2003). Das Bestreben Alternativen zu finden ist groß, bisher konnte damit aber noch nicht die Effizienz antibiotischer Leistungsförderer erreicht werden, weshalb ernährungs-physiologischen und hygienischen Aspekten in der Tierhaltung größere Aufmerksamkeit geschenkt werden soll (VANBELLE, 2001). Durch diese Entwicklung eröffnet sich ein aufnahmebereiter Markt für jegliche „natürliche“ Systeme, die eine effiziente Kontrolle der Darmflora in Aussicht stellen (CHESSON u. STEWART, 2001). Ein Beispiel für ein „natürliches“ System ist die „Antiadhäsionstherapie“. Darunter versteht man eine Verhinderung der Anheftung von Bakterien durch Gabe von Rezeptoranaloga (SHA-RON u. OFEK, 2000). Laut Guggenbichler et al. (1995) sind Oligogalakturonide aus Pektinen von Ka-rotten, Äpfeln, Orangen und anderen Pflanzen wirksam in der Adhärenzblockierung. KASTNER et al. (2002) sind der Meinung, dass derartige Oligogalakturonide vielfältig klinisch einsetzbar sind, nicht nur prophylaktisch und in der Therapie von Durchfallerkrankungen, sondern auch zum Beispiel um die Mikroflora des Darms bei langfristiger Antibiotikagabe stabil zu halten.
Im vorliegenden Versuch wurde die Wirkung eines mit Oligogalakturoniden angereicherten Apfeltres-ters in einer Dosierung von 0,5 %, 1 % und 5 % auf das Durchfallgeschehen bei Absetzferkeln unter-sucht.

Vielhaber et al.: Einsatz von Oligogalakturoniden zur Reduzierung des Absetzdurchfalles
Seite 240 8. BOKU-Symposium Tierernährung 2009
Material und Methoden Der Fütterungsversuch wurde mit 196 Ferkeln der Rasse [(Edelsau x Landrasse) x Pietrain] des Ver-suchsstalls des LFZ Raumberg-Gumpenstein, Institut für Biologische Landwirtschaft in Wels/Thalheim durchgeführt. Am Absetztag waren die Ferkel mindestens 40 Tage alt, was den Vorgaben der EU VO 834/2007 entspricht. Folgende zootechnische Maßnahmen und Impfungen wurden in der Säugezeit durchgeführt: intramuskuläre Injektion von 200 mg Eisendextran innerhalb der ersten drei Lebensta-ge, Kastration ohne Narkose innerhalb der ersten Lebenswoche und Mykoplasmenschutzimpfung in der 2. und 4. Lebenswoche.
Drei Tage vor dem Absetzen wurden die Ferkel durch ausgleichende Randomisierung nach den Para-metern Gewicht, Haptoglobin, Geschlecht und Wurfzugehörigkeit auf drei Versuchs- und eine Kontroll-gruppe aufgeteilt. Beim Absetzen wurden die 4 Gruppen in 4 gleiche, nebeneinanderliegende Buchten abwechselnd in 2 Aufzuchtställen eingestallt (Tabelle 1). Es ergaben sich insgesamt 5 Wiederholungen mit ca. 10 Tieren pro Gruppe. Jeder Durchgang dauerte 22 Tage und gliederte sich in 3 Tage vor dem Absetzen, Absetztag und 18 Tage nach dem Absetzen.
Tabelle 1: Gruppenplan
Gruppe 1. Durchgang 2. Durchgang 3. Durchgang 4. Durchgang 5. Durchgang Anzahl Bucht Anzahl Bucht Anzahl Bucht Anzahl Bucht Anzahl Bucht
0,5 % 10 1 II 7 2 II 8 4 I 13 3 II 11 1 I 1 % 10 2 II 7 3 II 8 1 I 13 4 II 11 3 I 5 % 10 3 II 7 4 II 8 2 I 13 1 II 11 4 I KG 10 4 II 7 1 II 8 3 I 13 2 II 11 2 I 0,5 %, 1 % und 5 %: Versuchsgruppen KG: Kontrollgruppe I, II: Aufzuchtstall 1 bzw. 2
Ab Tag -3 vor dem Absetzen bis inklusive Tag 0 (= Absetztag) erhielten die Ferkel der Versuchsgrup-pen den mit Wasser vermischten, enzymatisch behandelten Apfeltrester als Paste oral in den 3 unter-schiedlichen Dosierungen. Die Kontrollgruppe erhielt Wasser als Placebo. Von Tag 0 bis Tag 18 wurde den Ferkeln der Versuchsgruppen der enzymatisch behandelte Apfeltrester in einer Dosierung von 0,5 %, 1 % und 5 % ins Aufzuchtfutter gemischt. Das Ferkelaufzuchtfutter entsprach den Vorgaben der EU VO 834/2007 und enthielt keine zugesetzten Enzyme, Aminosäuren oder organische Säuren.
Am Absetztag und am Tag danach wurde allen Ferkeln eine Elektrolytlösung zur freien Aufnahme angeboten. Dies diente dazu, den Tieren während der Angewöhnungsphase im Aufzuchtstall genü-gend Flüssigkeit zur Verfügung zu stellen und somit optimale Startbedingungen zu garantieren.
Die Ferkel wurden an den Tagen -3, 0, 4, 11 und 18 gewogen. Gleichzeitig erfolgte an den Tagen 4, 11 und 18 eine Futterrückwaage, um die durchschnittliche tägliche Futteraufnahme und Futterverwer-tung berechnen zu können.
Der Kot wurde an den Tagen 4 bis 8 und 13 mittels Kotscore von 0 (geformter Kot) bis 2 (flüssiger Kot) beurteilt. Für die Auswertung wurde der Begriff „Gruppensumme“ eingeführt. Er bezeichnet die addierten Kotscores innerhalb einer Gruppe am jeweiligen Untersuchungstag. Weiters wurde der Be-griff „Durchfalltag“ verwendet. Aus Vorarbeiten (HAGMÜLLER, unveröffentlicht) geht hervor, dass Ferkel, die zu keinem Untersuchungszeitpunkt einen Kotscore von > 1 hatten, als gesund zu werten sind, da diese Tiere keine Wachstums- oder Leistungseinbußen zeigten. Dementsprechend wurde ein „Durchfalltag“ folgendermaßen definiert: jeder Tag, an dem der Scorewert eines Ferkels 1,5 oder 2,0 war bzw. die Tage ab dem Tod eines Ferkels, sofern die Todesursache mit akutem Durchfallgesche-hen in Zusammenhang stand. Solange Tiere mit Durchfall keine Abweichungen von einem normalen

Vielhaber et al.: Einsatz von Oligogalakturoniden zur Reduzierung des Absetzdurchfalles
8. BOKU-Symposium Tierernährung 2009 Seite 241
Allgemeinverhalten sowie erhaltene Fresslust zeigten, wurde von einer antibiotischen Behandlung abgesehen.
Blutproben von allen Ferkeln wurden durch Punktion der Vena cava cranialis am Tag -3 und 6 gewon-nen. Serum diente zur Analyse des Entzündungsparameters Haptoglobin. Hohe Haptoglobinwerte im Serum deuten auf unspezifische Gewebsschädigungen hin, die z.B. schon vor dem Auftreten von Durchfall stattfinden können.
Die statistische Auswertung erfolgte mit den Programmen SPSS und EXCEL 2007 für Windows Vista. Unterschiede zwischen den Gruppen wurden beim Kotscore mittels Chi²-Test ermittelt, bei den ande-ren Parametern mittels einfacher Varianzanalyse bzw. T-Test. Es galt als Signifikanzniveau p < 0,05.
Ergebnisse Aus der Darstellung des Kotscores als Gruppensumme (Abbildung 1) war ersichtlich, dass die 5 % Versuchsgruppe an den Tagen 5 bis 8 die niedrigste Gruppensumme hatte, was auf eine höhere An-zahl an Tieren mit normal geformten Kot hindeutet. Statistisch waren keine Unterschiede zwischen den Gruppen nachweisbar (p = 0,123).
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
Tag 4 Tag 5 Tag 6 Tag 7 Tag 8 Tag 13
Tage nach dem Absetzen
Gru
ppen
sum
me
KG
0,5%
1%
5%
Abbildung 1: Kotscore, dargestellt als Gruppensumme
Rund 80 % der Ferkel erkrankten in diesem Versuch an Durchfall (Tabelle 2). Somit waren die Ver-suchsbedingungen optimal, um herauszufinden, ob der Futterzusatz wirkt oder nicht. In der 1 % Ver-suchsgruppe waren bereits am Tag 4 mehr als die Hälfte der Tiere krank, während in den anderen Gruppen erst am Tag 5 diese Marke überschritten wurde. Von insgesamt 245 möglichen Durchfallta-gen (Kotscore Tag 4 bis 8 = 5 Tage * 49 Ferkel pro Gruppe) gab es in der Kontrollgruppe am meisten, in der 5 % Versuchsgruppe am wenigsten. Dieser Unterschied ließ sich nicht statistisch absichern (p = 0,282).
Die Ferkelverluste betrugen über den gesamten Versuch 6,63 % (13 Tiere). Die meisten Ferkel fielen in der 5 % Versuchsgruppe (12,24 %, 6 Ferkel) aus. Darauf folgte die Kontrollgruppe mit 4 Ferkeln (8,16 %) und die 0,5 % Versuchsgruppe mit 3 Ferkeln (6,12 %). In der 1 % Versuchsgruppe überleb-ten alle Ferkel (0 % Verlust). Alle verendeten Tiere zeigten akute Durchfallsymptomatik. Die Laborun-tersuchung in der AGES Linz ergab ausschließlich hämolysierende E. coli.

Vielhaber et al.: Einsatz von Oligogalakturoniden zur Reduzierung des Absetzdurchfalles
Seite 242 8. BOKU-Symposium Tierernährung 2009
Tabelle 2: Durchfallhäufigkeit (bei den Berechnungen wurde Tag 13 nicht berücksichtigt)
KG (N = 49)
0,5 % (N = 49)
1 % (N = 49)
5 % (N = 49)
Anzahl Ferkel mit Durchfall 42 42 41 36 Durchschnittliche Anzahl an Ferkeln mit Durchfall an den Kotscore-Tagen
4 40,8 % 34,7 % 59,2 % 36,7 % 5 65,3 % 57,1 % 53,1 % 51,0 % 6 69,4 % 63,3 % 59,2 % 51,0 % 7 63,3 % 69,4 % 61,2 % 57,1 % 8 67,3 % 67,3 % 69,4 % 63,3 %
(13) 26,5 % 34,7 % 22,4 % 44,9 % Anzahl Durchfalltage 150 143 148 127 Durchfalltage pro Ferkel 3,1 2,9 3,0 2,6 N: Anzahl Ferkel KG: Kontrollgruppe 0,5 %, 1 % und 5 %: Versuchsgruppen Anzahl Ferkel mit Durchfall: ein Ferkel hat dann „Durchfall“, wenn es einmal einen Scorewert von 1,5 bzw. 2,0 aufweist oder eine Gruppensumme > 5,0 oder wenn es verstorben ist Durchfalltage pro Ferkel: ist die durchschnittliche Anzahl der Tage (von den 5 Kotscore-Tagen), die ein Ferkel krank war
Tabelle 3: Biologische Leistungsdaten
Tag KG 0,5 % 1 % 5 % M SD N M SD N M SD N M SD N
Körpergewicht (kg)
-3 12,08 2,40 49 12,08 2,39 49 12,08 2,45 49 12,08 2,43 49 0 13,13 2,67 49 13,13 2,64 49 13,11 2,65 49 13,26 2,75 49 4 13,56 2,78 49 13,72 2,89 49 13,69 3,10 49 13,80 2,89 49 11 15,16 3,57 45 15,50 3,40 46 15,46 3,84 49 15,76 3,96 44 18 19,36 4,34 45 19,85 3,83 46 19,75 4,80 49 19,86 4,15 43
Tageszunahme (g)
-3 – 0 350 205 49 352 170 49 345 132 49 392 186 49 1 – 4 108 213 49 146 175 49 143 214 49 136 162 49 5 – 11 216 192 45 248 212 46 254 204 49 268 192 44 12 – 18 601 172 45 621 127 46 612 173 49 569 137 43
gesamt (1 – 18) 342 138 45 369 108 46 367 154 49 361 109 43 M: Mittelwert SD: Standardabweichung N: Anzahl Ferkel
KG: Kontrollgruppe 0,5 %, 1 % und 5 %: Versuchsgruppen
Die Ferkel der Versuchsgruppen zeigten eine höhere Lebendmassezunahme (Tabelle 3) und wogen am Tag 18 um 484 g (0,5 % Versuchsgruppe), 388 g (1 % Versuchsgruppe) bzw. 501 g (5 % Ver-suchsgruppe) mehr als die Tiere der Kontrollgruppe, was jedoch durch statistische Prüfung nicht ab-gesichert werden konnte (p > 0,05).

Vielhaber et al.: Einsatz von Oligogalakturoniden zur Reduzierung des Absetzdurchfalles
8. BOKU-Symposium Tierernährung 2009 Seite 243
Die Ferkel der Kontrollgruppe nahmen insgesamt etwas weniger Futter auf (0,67 kg/Ferkel/Tag), als die der Versuchsgruppen (rund 0,71 kg/Ferkel/Tag), was sich in der niedrigsten Tageszunahme wider-spiegelt. Auch die Futterverwertung war in dieser Gruppe am schlechtesten (2,10). Die 1 % Versuchs-gruppe zeigte die beste Futterverwertung (1,92).
Bei der Analyse von Haptoglobin ergab sich am Tag -3 kein Unterschied zwischen den Gruppen (p = 1,0), was zeigt, dass die Gruppeneinteilung für diesen Parameter ausgeglichen war. Die Haptoglobin-konzentration stieg in allen Gruppen von Tag -3 bis Tag 6. Es wurde kein signifikanter Unterschied (p > 0,05) zwischen den Gruppen gefunden.
Im Verlauf des Versuchs wurden in der Kontrollgruppe und in der 0,5 % Versuchsgruppe je 2 Ferkel antibiotisch behandelt, eines davon starb am darauffolgenden Tag. In der 5 % Versuchsgruppe wur-den 3 Ferkel mit Antibiotika therapiert und alle drei starben in den folgenden Tagen. Nur die mit Anti-biotika behandelten Ferkel der 1 % Versuchsgruppe (2 Ferkel) überlebten alle. Sämtliche Ferkel, die behandelt wurden und überlebten blieben mit ihren Tageszunahmen innerhalb der doppelten Stan-dardabweichung vom Gruppenmittelwert. Deshalb wurden sie bei den Berechnungen mitberücksich-tigt.
Diskussion Der vorliegende Fütterungsversuch diente dazu aufzuzeigen ob ein mit Oligogalakturoniden angerei-cherter Apfeltrester zur Prophylaxe des Absetzferkeldurchfalles eingesetzt werden kann und wenn ja welche Konzentration zu empfehlen wäre.
Aus den Ergebnissen ist zu entnehmen, dass es im Kotscore keine signifikanten Unterschiede zwischen den Gruppen gab. Am Tag 13 war die Gruppensumme des Kotscores in allen Gruppen deutlich niedri-ger, als an den Tagen 4 bis 8. Eine mögliche Erklärung dafür ist, dass in den Durchgängen 3, 4 und 5 an den Tagen 8 (nach der Beurteilung des Kots), 9 und 10 den Ferkeln aller Gruppen zusätzlich die gleiche Menge Torf oder Erde gegeben wurde, da sie noch hochgradigen Durchfall hatten. Durch die Wirkung von Torf bzw. Erde kann es zu einer Verbesserung des Kots kommen. Eine weitere Erklärung ist, dass sich die Ferkel an die neue Situation (Stall, neue Gruppe, Futter) angepasst haben. Vorarbei-ten von HAGMÜLLER (unveröffentlicht) und HAGMÜLLER et al. (2009) zeigten, dass der Absetzdurch-fall an den Tagen 4 bis 8 nach dem Absetzen am stärksten ist und sich dann wieder bessert. Unter Berücksichtigung der Tatsache, dass alle Ferkel dieselbe Behandlung (gleiche Menge Torf bzw. Erde an den Tagen 8, 9 und 10) erhalten haben, kann die Gruppensumme des Kotscores am Tag 13 fol-gendermaßen interpretiert werden: die 1 % Versuchsgruppe zeigte die geringste Anzahl an Durchfall-tieren, was jedoch statistisch nicht abgesichert werden konnte (p = 0,186).
Es gab keine signifikanten Unterschiede im Gewicht und den Tageszunahmen. Zu berücksichtigen ist hier jedoch der Einfluss der verstorbenen Ferkel in der Kontrollgruppe, der 0,5 % und 5 % Versuchs-gruppe. Hätten diese überlebt, wären die Zunahmen je nach Leistung gestiegen oder gefallen. Die tatsächliche Auswirkung ist schwer abschätzbar.
Betrachtete man die Durchgänge getrennt, so fiel im 3. Durchgang auf, dass die 1 % Versuchsgruppe ein signifikant höheres Gewicht (p < 0,01) als die Kontrollgruppe aufwies, sowie signifikant bessere Tageszunahmen als die Kontrollgruppe und die 0,5 % Versuchsgruppe (p < 0,05). Auch beim Ver-gleich der Durchfalltage war die 1 % Versuchsgruppe mit nur 16 Durchfalltagen besser als die 5 % Versuchsgruppe mit 22, die Kontrollgruppe mit 32 und die 0,5 % Versuchsgruppe mit 33 Durchfallta-gen. Dieses Ergebnis wiederholte sich aber nicht in den anderen Durchgängen. Die wechselnden Er-gebnisse bestätigen die Meinung von CHESSON (1994), dass alle intestinalen Mediatoren, dazu zählen von Antibiotika über Oligosaccharide bis hin zu Antikörpern, Probiotika usw. alle Futterzusätze, die einen Einfluss auf den Darm haben, eines gemeinsam haben: die Variabilität und Unvorhersehbarkeit ihrer Effekte.

Vielhaber et al.: Einsatz von Oligogalakturoniden zur Reduzierung des Absetzdurchfalles
Seite 244 8. BOKU-Symposium Tierernährung 2009
Unter den gegebenen Versuchsbedingungen erreichte der mit Oligogalakturoniden angereicherte Ap-feltrester keine statistisch nachweisbare Senkung des Anteils an Ferkeln mit Durchfall. JUGL et al. (2001) konnten in ihrem Versuch zeigen, dass Karottenpulver mit einem Anteil von 10 % ins Futter eingemischt als Durchfallprophylaktikum wirksam ist. Weitere Untersuchungen, bei denen Oligogalak-turonide in höherer Dosierung oder gemischt mit anderen Futterzusätzen, die sich mit dem Wirkme-chanismus der Oligogalakturonide kombinieren lassen (z.B. Kräuter, Tonmineralien, Torf, Präbioti-ka, …), erscheinen sinnvoll.
Literatur
CHESSON, A. (1994): Probiotics and other intestinal mediators. In: COLE, D.J.A., WISEMAN, J., VARLEY, M.A. (Eds.): Principles of Pig Science. Nottingham University Press, Nottingham, p. 197-214.
CHESSON, A., STEWART, C. (2001): Modulation of the gut microflora by enzyme addition. In: PIVA, A., BACH KNUDSEN, K.E.,
FULLER, R. (1989): Probiotics in man and animals. Journal of Applied Bacteriology 66, 365-378.
GUGGENBICHLER, J.P., MEISSNER, P., JURENITSCH, J., DE BETTIGNIES-DUTZ, A. (1995): Blockierung der Anlagerung von Keimen an Epithelzellen. Deutsches Patent DE 4 330 773 A1.
HAGMÜLLER, W., VIELHABER, B., GALLNBÖCK, M., HAHN, I., FRANZ, C. (2009): Einsatz von Kräutern, Tonmineralien und Effektiven Mikroorganismen zur Prophylaxe des Absetzdurchfalles. Beiträge zur 10. Wissenschaftstagung ökologischer Landbau. Zürich 2009, S. 198-201.
JUGL, M., ZITTERL-EGLSEER, K., BEIER, T., SCHILCHER, F., GABLER, C., SCHUH, M., KASTNER, U., GUGGENBICHLER, J.P., FRANZ, C. (2001): Experimentelle Feldstudie über den Einsatz von Karottenpektinen als Futterzusatz zur Durchfallprophylaxe in der Ferkelaufzucht. Tierärztliche Praxis 29 (G), 308-312.
KASTNER, U., GLASL, S., FOLLRICH, B., GUGGENBICHLER, J.P., JURENITSCH, J. (2002): Saure Oligosaccharide als Wirkprinzip von wäßrigen Zubereitungen aus der Karotte in der Prophylaxe und Therapie von gastrointestinalen Infektionen. Wiener Medizinische Wochenschrift 152, 379-381.
KASTNER, U., KECK, B., MATTHES, S., GUGGENBICHLER, J.P., PETERS, C., GADNER, H. (2004): Prophylaxe enteraler Infektio-nen in der pädiatrischen Onkologie: Orale Dekontamination versus anti-adhäsiver Prophylaxe mit Oligosacchariden aus der Karotte. Antibiotika Monitor 5/6.
PLUSKE, J.R. (2001): Morphological and functional changes in the small intestine of the newly-weaned pig. In: PIVA, A., BACH
PUSZTAI, A. (1992): Lectins in Probiosis. UK Patent PCT GB91 02236.
ROLLE, M., MAYR, A. (2007): Medizinische Mikrobiologie, Infektions- und Seuchenlehre. 8. Aufl., Enke Verlag, Stuttgart.
SHARON, N., OFEK, I. (2000): Safe as mother’s milk: Carbohydrates as future anti-adhesion drugs for bacterial diseases. Glycoconjugate journal 17, 659-664.
STEINER, T. (2006): Managing Gut Health – Natural Growth Promoters as a Key to Animal Performance. Nottingham University Press, Nottingham.
VANBELLE, M. (2001): Current Status and future Perspectives in E.U. for Antibiotics, Probiotics, Enzymes and organic Acids in Animal Nutrition. In: PIVA, A., BACH KNUDSEN, K.E., LINDBERG, J.E. (Eds.): Gut Environment of Pigs. Nottingham University Press, Nottingham, p. 231-256.
Autorenanschrift
Mag. Barbara Vielhaber LFZ Raumberg-Gumpenstein Institut für Biologische Landwirtschaft und Biodiversität der Nutztiere Außenstelle Wels Austraße 10, A-4600 Wels/Thalheim E-Mail: [email protected]

Firmensponsoring – Reisenberger GmbH, Stuttgarterstraße 2, A-2380 Perchtoldsdorf
8. BOKU-Symposium Tierernährung 2009 Seite 245

Wehr und von Rosenberg: Lanthan-Verbindungen wirken einem erhöhten Knochenverlust im Osteoporosemodell der ovariektomierten Ratte entgegen
Seite 246 8. BOKU-Symposium Tierernährung 2009
Lanthan-Verbindungen wirken einem erhöhten Knochenverlust im Osteoporosemodell der ovariektomierten Ratte entgegen
Ulrich Wehr1 und Sylvia von Rosenberg2 1 Landesinstitut für Tiergesundheit und Futtermittel am Bayerischen Landesamt für
Gesundheit und Lebensmittelsicherheit, Oberschleißheim 2 Lehrstuhl für Tierernährung und Diätetik des Veterinärwisschenschaftlichen De-
partments der Tierärztlichen Fakultät, Ludwig-Maximilians-Universität, München
Einleitung Lanthan zählt zu einer Gruppe von 17 Übergangsmetallen, die als Seltenerdmetalle bezeichnet werden und in der dritten Nebengruppe des Periodensystems stehen. Dabei handelt es sich im Wesentlichen um die Gruppe der Lanthanoide, welche um die Elemente Scandium und Yttrium erweitert ist. Die Gruppe der Lanthanoide umfasst 14 Elemente, welche im Periodensystem auf das Lanthan folgen und mit diesem eine enge chemische Verwandtschaft aufweisen. Diese sind Cer, Praseodym, Neodym, Promethium, Samarium, Europium, Gadolinium, Terbium, Dysprosium, Holmium, Erbium, Thulium, Ytterbium und Lutetium. Der Begriff „Seltene Erden“ bezeichnet im engeren Sinne die Oxide der Sel-tenerdmetalle.
Neben ihrer Verwendung in der Metallurgie, der Glas- und Keramikindustrie sowie der Radiologie wer-den Seltene Erden seit über 40 Jahren in der chinesischen Landwirtschaft zu Zwecken der Leistungs-förderung eingesetzt. Aufgrund ihrer vielfältigen pharmakologischen Eigenschaften wurden Seltene Erden aber bereits auch früher in der Humanmedizin unter anderem als Antiemetika, Antikoagulantien und Antiinfektiva verwendet. Seit mehreren Jahren findet die Verbindung Lanthancarbonat als nicht calcium- und nicht aluminiumhaltiger Phosphatfänger in der Behandlung von Patienten mit Nie-ren¬insuffizienz Anwendung. Als Nebeneffekt wurde von D`HAESE et al. (2003) in einer Studie beo-bachtet, dass bei 80 % der Patienten während der Einnahme von Lanthancarbonat der erhöhte Kno-chenturnover normalisiert werden konnte.
Diese Beobachtung ist insofern von Bedeutung, da die Therapie bzw. Prävention der manifesten Oste-oporose bis heute schwierig ist und teilweise nur unbefriedigende Ergebnisse liefert. So ist die Hor-monersatztherapie, die immer noch als Mittel der Wahl gilt, aufgrund ihrer Nebenwirkungen oft in Diskussion. Daher werden seit Jahren alternative Substanzen oder Stoffgruppen auf eine knochenmo-dulierende Wirkung mit dem Ziel untersucht, sie in der Therapie der postmenopausalen Osteoporose als Ersatz der Hormonersatztherapie oder zumindest als hormonsparende Zusatztherapie einzusetzen.
Ziel dieser Untersuchung war es daher, die oben beschriebenen positiven Effekte von Lanthan auf den Knochen im Osteoporosemodell der ovariektomierten Ratte näher zu untersuchen.
Material und Methoden 40 weibliche Wistar-Ratten wurden im Alter von 4 Monaten entweder scheinoperiert (SHAM, n=10) oder ovariektomiert (OVX, n=30). Den ovariektomierten Tieren zweier Gruppen (n=10/Gruppe) wurde entweder Lanthancarbonat oder Lancer® (ein Lanthan-Cer-Gemisch, mit einem Lanthan-Cer-Verhältnis von 1 zu 1,8) in einer Dosierung von 12 mg Lanthan/100 g Körpermasse/Tag für 24 Wochen über das

Wehr und von Rosenberg: Lanthan-Verbindungen wirken einem erhöhten Knochenverlust im Osteoporosemodell der ovariektomierten Ratte entgegen
8. BOKU-Symposium Tierernährung 2009 Seite 247
Futter verab¬reicht. Die SHAM-Gruppe diente als positive und eine OVX-Gruppe als negative Kontrol-le.
Nach 0, 4, 8, 12, 16, 20 und 24 Wochen wurden zur Beurteilung des Knochenstoffwechsels die Bone Marker Pyridinolin (PYD in nmol/mmol Crea) als Parameter der Knochenresorption im 24h-Urin und Osteocalcin (Oc in ng/ml) als Parameter der Knochenformation im Serum gemessen.
Am Ende der Untersuchung wurde sowohl der Knochenmineralstoffgehalt der Tibia (Calcium in mg/g Asche) als auch deren Knochendichte (BMD in mg/cm3) mittels peripherer quantitativer Computerto-mographie (pQCT) bestimmt.
Ergebnisse In beiden Versuchsgruppen führte die Gabe von Lanthan dazu, dass die nach Ovariektomie stark er-höhte Exkretion des Knochenresorptions¬parameters Pyridinolin gesenkt (p<0,05) und die Konzentra-tion des Knochenformationsparameters Osteocalcin erhöht wurde (siehe Tabelle 1).
Tabelle 1: Osteocalcingehalt im Serum und Pyridinolinausscheidung im Urin (berechnet als nmol/mmol Crea) der Tiere in den Versuchsgruppen (MW ± SD) als Flächenberechnung unter der Kurve (Area under the Curve, AUC) im Versuchszeitraum zwischen Woche 0 und 20
AUC Oc AUC PYD/Crea
SHAM 643,3±147,4 124,6±25,9*
OVX 653,2±219,1 192,4±14,7#
OVX+LaCO3 902,2±218,7***### 187,3±5,6#
OVX+Lancer® 1016,3±300,5***### 160,3±31,3*
• * (p <0,05), ** (p<0,01) , *** (p<0,001) vs OVX
• # (p <0,05), ## (p<0,01), ### (p<0,001) vs SHAM
Neben diesen positiven Effekten auf den Knochenstoffwechsel, konnte in beiden Therapiegruppen der Knochenmineralstoffverlust (Calcium in der Knochenasche) nach Ovariektomie verhindert werden (p<0,05). Des Weiteren lag die Knochendichte in beiden Lanthan-Gruppen oberhalb der Knochendich-te der unbehandelten OVX-Gruppe, wobei speziell in der Lancer®-Gruppe die trabekuläre Dichte im Vergleich zur OVX-Gruppe signifikant erhöht war (p<0,01) (siehe Tabelle 2).
Um die positiven Effekte auf die Knochendichte optisch darzustellen, wurde die Knochenmikroarchitek-tur mittels Mikro-Computertomographie (µCT) dreidimensional erfasst, was in Abbildung 1 beispielhaft von jeweils einem Tier pro Gruppe dargestellt ist. Besonders gut sichtbar wird hierbei die im Vergleich zur unbehandelten OVX-Kontrolle höhere Trabekeldichte in den beiden Therapiegruppen.

Wehr und von Rosenberg: Lanthan-Verbindungen wirken einem erhöhten Knochenverlust im Osteoporosemodell der ovariektomierten Ratte entgegen
Seite 248 8. BOKU-Symposium Tierernährung 2009
Tabelle 2: Calciumgehalt (mg Ca/g Asche), Gesamtdichte und trabekuläre Dichte (mg/cm3, pQCT-Messung) der Knochen (gemessen an der Tibia) der Tiere in den Versuchsgruppen (MW ± SD)
Ca Tibia (mg/g Asche)
Gesamtdichte Tibia (mg/cm3)
Trabekuläre Dichte Tibia (mg/cm3)
SHAM 430,7±28,8* 865,0±42,7* 469,5±37,0***
OVX 361,2±41,1# 685,8±32,8# 222,8±58,5###
OVX+LaCO3 432,5±64,4* 757,0±99,7 227,4±68,1###
OVX+Lancer® 433,89±12,3* 693,3±46,9# 310,4±82,1###**
* (p <0,05), ** (p<0,01) , *** (p<0,001) vs OVX # (p <0,05), ## (p<0,01), ### (p<0,001) vs SHAM
Abbildung 1: Dreidimensionale Darstellung der Knochendichte (gemessen per µCT) jeweils eines Tie-res der positiven (Sham) und negativen (OVX) Kontrollgruppe, sowie der beiden Thera-piegruppen (OVX+LaCO3 und OVX+Lancer®)

Wehr und von Rosenberg: Lanthan-Verbindungen wirken einem erhöhten Knochenverlust im Osteoporosemodell der ovariektomierten Ratte entgegen
8. BOKU-Symposium Tierernährung 2009 Seite 249
Schlussfolgerung Die Ergebnisse dieser Studie zeigen, dass die eingesetzten Lanthan-Verbindungen sowohl auf den Knochenstoffwechsel als auch auf Knochendichte und –struktur positive Effekte haben. Somit stellen sie eine erste Grundlage für weiterführende Untersuchungen zu knochenprotektiven Effekten von Lan-than bzw. evtl. auch anderer Lanthanoide dar. Möglicherweise werden die knochenbildenden Osteozy-ten stimuliert, was in dieser Studie durch den im Vergleich zur OVX-Negativkontrolle höheren Gehalt an Osteocalcin belegt wurde. Ob dies jedoch auf einer Zunahme der Aktivität oder einer zunehmenden Anzahl an Osteozyten oder auch einer Kombination von beidem beruht, ist bislang unklar.
Es kann jedoch postuliert werden, dass der höhere Osteocalcinspiegel nicht auf einem lediglich höhe-ren Knochen-Turnover beruht, da der Knochenresorptionsparameter Pyridinolin im Urin der Therapie-gruppen gleichzeitig niedriger war als in der OVX-Negativkontrolle. Der Calciumgehalt der Knochen-asche war bei den Therapiegruppen signifikant höher als bei der OVX-Negativkontrolle und unter-schied sich nicht von der Positivkontrolle Sham.
Weiterhin bleibt zu klären, welche Mechanismen die Lanthanoide im Körper aktivieren oder welche Rezeptoren im Organismus möglicherweise stimuliert werden, die zu den hier gezeigten knochenpro-tektiven Effekten im postmenopausalen Osteoporosemodell der ovariektomierten Ratte führen.
Literatur
D´HAESE, P. D., SPASOVSKI, G. B., SIKOLE, A., HUTCHISON, A., FREEMONT, T. J., SULKOVA, S., SWANPOEL, C., PEJANOVIC, S., DJUKANOVIC, L., BALDUCCI, A., COEN, G., SULOWICZ, C., FERREIRA, A., TORRES, A., CURIC, S., POPOVIC, M., DIMKOVIC, N. (2003): A multicenter study on the effects of lanthanum carbonate (FosrenolTM) and calcium carbonate on renal bone disease in dialysis patients. Kidney International 63:73–78
Autorenanschrift
Dr. Ulrich Wehr Sachgebiet Futtermittel Landesinstitut Tiergesundheit und Futtermittel Bayerisches Landesamt für Gesundheit und Lebensmittelsicherheit D-85762 Oberschleißheim E-Mail: [email protected] Dr. Sylvia von Rosenberg Lehrstuhl für Tierernährung und Diätetik Veterinärwissenschaftliches Department Ludwig-Maximilians-Universität München Schönleutnerstr. 8 D-85764 Oberschleißheim E-Mail: [email protected]

Kluge et al.: Eine Valinsupplementierung ergibt neue Möglichkeiten bei der Proteinreduzierung im Ferkelfutter
Seite 250 8. BOKU-Symposium Tierernährung 2009
Eine Valinsupplementierung ergibt neue Möglichkeiten bei der Proteinreduzierung im Ferkelfutter
Holger Kluge1, Joerg Bartelt2, Gabriele Stangl1 1 Martin-Luther-Universität Halle-Wittenberg, Institut für Agrar- und Ernährungswis-senschaften
2Lohmann Animal Health GmbH & Co.KG
Einleitung Proteinreduzierte Rationen für Ferkel reduzieren nicht nur die Stickstoffexkretion sondern senken die Säurebindungskapazität der Ration, die zu einer besseren Futterumstellung nach dem Absetzen bei-trägt. Während der Aufzucht im Flatdeck wird die Futteraufnahme und Lebendmassezunahme verbes-sert und die Durchfallhäufigkeit reduziert. Diese Befunde wurden durch Absenkung des Proteingehal-tes in der Ration von 22 % auf 18 % in der Literatur hinreichend beschrieben. Weitere Reduzierungen der Proteinkonzentration führten in den meisten Versuchen zu keinen Leistungs-verbesserungen. In Rationen auf Basis Getreide und Sojaextraktionsschrot und einer Proteinkonzentration von <16 % wirkt zusätzlich zu den Aminosäuren Lysin, Threonin, Methionin + Cystein und Tryprophan auch Valin limitierend auf das Wachstum von Jungsauen, wie die Untersuchungen von Figuerora et al (2003) gezeigt haben. Neuere Untersuchungen von Baera et al. (2009) empfehlen für Absetzferkel ein opti-males Verhältnis an standardisiert ileal verdaulichen (siv) Val:Lys von 70 %. Inwieweit unter Einhal-tung dieses Val:Lys Verhältnisses eine proteinreduzierte Fütterung Leistungsparameter von Absetzfer-keln beeinflusst, war Gegenstand dieser Untersuchung.
Material und Methoden Für die Untersuchung wurden 126 Kreuzungsferkel (DLxDE)xPI mit einem Absetzalter von 28 Tagen auf 9 Gruppen zu je 14 Tieren aufgeteilt. Die neun Gruppen wurden randomisiert zu 3 Varianten zu-geordnet. Bis zum Versuchsbeginn über 14 Tage erhielten alle Gruppen ein kommerzielles Ferkelstar-terfutter. Danach erhielten die Gruppen der Variante 1 eine Grundmischung I und eine Ergänzung an Sojaextraktionsschrot, Mineralfutter und Aminosäuren mit einem Rohproteingehalt in der Gesamtrati-on von 18 % (Tab. 1 u. 2). Tiere der Varianten 2 und 3 erhielten die proteinreduzierte Grundmischung II und eine Ergänzung an Sojaextraktionsschrot, Mineralfutter und Aminosäuren (Tab. 1 u. 2). In Vari-ante 3 erfolgte zusätzlich eine Ergänzung an Valin, um das gleiche siv Val:Lys Verhältnis wie in Varian-te 1 zu erreichen. Das Durchschnittliche Anfangsgewicht zu Versuchsbeginn betrug 11,1 ± 1,7 kg. Über einen Versuchszeitraum von 4 Wochen wurden die Tiere wöchentlich einzeln gewogen und der Futterverbrauch und Futteraufwand pro Gruppe ermittelt. Den Tieren standen pro Gruppe zwei Fut-terautomaten und 3 Zapfentränken zur Verfügung. Das Futter wurde pelletiert verabreicht. Die Fut-termischungen wurden auf die Gehalte an Rohprotein (N x 6,25) nach der Dumas Methode und die Gehalte an Aminosäuren mit einem Aminosäurenanalysator (Jeol, Croissy-sur-Seine, Frankreich) in einem kommerziellen Labor (Ajinomoto Eurolysine SAS, Amiens, Frankreich) analysiert. Tryptophan wurde im gleichen Labor nach Hydrolyse mit Bariumhydroxid mit einem HPLC und Fluoreszenzdetektor bestimmt.
Die Daten wurden einer Varianzanalyse (Statistica 7.0 for Windows Operating System) unterzogen. Signifikante Unterschiede zwischen den Mittelwerten (p<0,05, Tukey HSD-Test) wurden mit Hoch-buchstaben gekennzeichnet.

Kluge et al.: Eine Valinsupplementierung ergibt neue Möglichkeiten bei der Proteinreduzierung im Ferkelfutter
8. BOKU-Symposium Tierernährung 2009 Seite 251
Tabelle 1: Zusammensetzung und Inhaltsstoffe der Grundmischungen
Grundmischung I Grundmischung II Weizen 25,0 25,0 Gerste 23,4 28,65 Mais 30,0 30,0 Sojaextraktionsschrot 20,5 15,0 Sojaöl 0,45 0,70 Monocalciumphosphat 0,65 0,65 Inhaltsstoffe (g/kg Futter) und Umsetzbare Energie Rohprotein 177 152 Lysin 8,0 6,5 Methionin+Cystin 5,4 4,8 Threonin 6,1 5,1 Tryptophan 2,2 2,0 Valin 7,5 6,5 ME (MJ/kg Futter) 13,6 13,6
Tabelle 2: Zusammensetzung und Inhaltsstoffe der Versuchsrationen
Variante 1 Variante 2 Variante 3 Grundmischung I 93,40 0 0 Grundmischung II 0 93,13 93,05 Sojaextraktionsschrot 4,0 4,0 4,0 Mineralfutter 2,0 2,0 2,0 Lysin HCl 0,32 0,47 0,47 DL-Methionin 0,12 0,16 0,16 L-Threonin 0,12 0,18 0,18 L-Tryptophan 0,04 0,06 0,06 L-Valin 0 0 0,08 Rohprotein- und Aminosäurenkonzentration (g/kg Futter) und Umsetzbare Energie Rohprotein 180 170 171 Lysin 10,9 11,1 11,2 siv. Lysin1 10,0 10,0 10,0 Methionin/Cystin 6,6 6,5 6,6 Threonin 7,3 7,2 7,3 Tryptophan 2,7 2,6 2,6 Valin Isoleucin
7,8 7,0
7,2 6,4
7,9 6,3
siv. Valin1 7,0 6,3 7,1 siv Val:Lys (%) 70 63 70 ME (MJ/kg)2 13,4 13,4 13,4 1 kalkuliert aus der standardisiert praecaecalen Verdaulichkeit der Einzelkomponenten (GfE, 2006) 2 kalkuliert nach DLG Futterwerttabellen Schweine (1991)
Ergebnisse Die geplanten Aminosäuren- und RP-Gehalte wurden durch die Analyse bestätigt. Der Fütterungsver-such verlief über den ganzen Versuchszeitraum ohne nennenswerte Komplikationen. Es fielen insge-samt 2 Tiere in der Fütterungsvariante 1 und 1 Tier in der Fütterungsvariante 3 aus. Wie aus den Leistungsdaten in Tabelle 3 ersichtlich ist, wurde durch die Reduzierung der Proteinkonzentration in

Kluge et al.: Eine Valinsupplementierung ergibt neue Möglichkeiten bei der Proteinreduzierung im Ferkelfutter
Seite 252 8. BOKU-Symposium Tierernährung 2009
den Varianten 2 und 3 die Lebendmassezunahme und Futterverwertung verbessert. Die Futterauf-nahme stieg leicht an. Eine zusätzliche Supplementierung der proteinabgesenkten Ration mit Valin in Variante 3, auf das gleiche Niveau wie in Variante 1, führte zu einer signifikanten Verbesserung der Futterverwertung.
Tabelle 3: Leistungsparameter über den gesamten Versuchszeitraum (4 Wochen)
Variante 1 Variante 2 Variante 3 p-Wert Lebendmasse Versuchsbeginn (kg) 11,2 ±1,8 11,0 ±1,8 11,0 ±1,6 0,904 Lebendmasse Versuchsende (kg) 24,2 ±4,1 24,3 ±3,9 25,2 ±3,5 0,429 Lebendmassezunahme (g/Tag) 467 ±99 474 ±100 508 ±91 0,118 Futteraufnahme (g/Tag) 788 ±84 789 ±33 818 ±63 0,807 Futteraufwand (kg/kg) 1,69 ±0,04a 1,66 ±0,04ab 1,61 ±0,02b 0,054 a,b signifikante Unterschiede zwischen den Varianten sind durch unterschiedliche Hochbuchstaben gekennzeichnet
Diskussion Im vorliegenden Versuch wurde die Proteinkonzentration der Ration von 18 % auf 17 % reduziert. Trotz dieser geringen Absenkung konnten ähnliche Effekte auf Zunahmen und Futterverwertung beo-bachtet werden wie in Versuchen, die die Proteinabsenkung von einem höheren Proteinniveau durch-geführt hatten (Le Bellego et al. 2001; Htoo et al. 2007). Untersuchungen mit Absetzferkeln (Theil et al. 2004) haben gezeigt, dass bei einer Proteinkonzentration von 18,6 % und ein Verzicht auf eine Valinsupplementierung die Futteraufnahme um 15 % und die Lebendmassezunahme um 21 % gegen-über einer mit Valin supplementierten Ration signifikant gesenkt wurde. Das Verhältnis an siv Va-lin:Lysin betrug in der unsupplementierten Ration 59 % und in der supplementierten Ration 72 %. Im vorliegenden Versuch konnte durch Reduzierung der Proteinkonzentration von 18% auf 17 % unter Einhaltung eines siv Val:Lys Verhältnisses von 70 % eine signifikante Verbesserung der Futterver-wertung erzielt werden. Auch gegenüber der proteinreduzierten aber ohne Valin ergänzten Ration war die Lebendmassezunahme und Futterverwertung tendenziell verbessert. Für den Futteraufwand bei Ferkeln ermittelten (Wiltafsky et al. 2009) ein optimales siv Valin:Lysin Verhältnis von 61% während die Lebendmassezunahme bzw. Futteraufnahme bei einem entsprechenden Verhältnis von 66% bzw. 67% optimiert wurde. Zu berücksichtigen ist jedoch, dass in dieser Untersuchung keine optimale Tryp-tophanversorgung vorlag. Die tägliche Aufnahme an siv Valin im vorliegenden Versuch betrug in der Valin supplementierten Variante 5,8 g/Tag und in der nicht supplementierten Variante 5,0 g/Tag. Nach den Empfehlungen der GfE (2006) wird für Absetzferkel im Gewichtsbereich von 10-20 kg und einer Lebendmassezunahme von 500 g/Tag eine tägliche Aufnahme an siv Valin von 6,2 g empfohlen. Diese Aufnahme an siv Valin ist in proteinreduzierten Rationen (17 % Rohprotein) nur durch eine Valiner-gänzung in der Ration erreichbar. Bei einem Verhältnis an siv Valin:Lysin von 70% werden die Leis-tungsparameter von Ferkeln nicht negativ beeinflusst, wenn der Rohproteingehalt auf 17% abgesenkt wird. Gleichzeitig sollten die Aminosäuren Methionin, Threonin und Tryptophan entsprechend dem idealen Protein ergänzt sein. Ökonomisch betrachtet verteuert sich die Ration durch die Proteinredu-zierung und den zusätzlichen Einsatz kristalliner Aminosäuren um 0,74 Euro/100 kg Futter. Durch die bessere Futterverwertung der proteinreduzierten und mit Valin supplementierten Ration wurde ge-genüber der Ration mit einem um 1 % höheren Proteingehalt eine Einsparung an Futterkosten pro Tier von 0,08 Euro erzielt.

Kluge et al.: Eine Valinsupplementierung ergibt neue Möglichkeiten bei der Proteinreduzierung im Ferkelfutter
8. BOKU-Symposium Tierernährung 2009 Seite 253
Literatur
Barea R., Brossard L., Le Floc’h N., Primot Y., Melchior D. and van Milgen J. (2009): The standard ileal digestible valine-to-lysine requirement ratio is at least seventy percent in postweaned piglets. J. Anim. Sci. 87, 935-947
Deutsche Landwirtschafts-Gesellschaft (DLG), 1991: DLG-Futterwerttabellen – Schweine. DLG-Verlag, Frankfurt (Main)
Gesellschaft für Ernährungsphysiologie (GfE), 2006: Empfehlungen zur Energie- und Nährstoffversorgung von Schweinen. DLG-verlag, Frankfurt (Main)
Figueroa J.L., Lewis A.J., Miller P.S., Fischer R.L. and Diedrichsen R.M. (2003): Growth, carcass traits, and plasma amino acid concentrations of gilts fed low-protein diets supplemented with amino acids including histidine, isoleucine, and valine. J. Anim. Sci. 81, 1529-1537
Htoo J.K., Araiza A., Sauer W.C., Rademacher M., Zhang Y., Cervantes M. und Zijlstra R.T. (2007): Effect of dietary protein content on ileal amino acid digestibility, growth performance, and formation of microbial metabolites in ileal and cecal digesta of early-weaned pigs. J. Anim. Sci. 85, 3303-3312
Le Bellego L., van Milgen J., Dubois S. and Noblet J. (2001): Energy utilization of low-protein diets in growing pigs. J. Anim. Sci. 79, 1259-1271
Theil P.K., Fernandez J.A. and Danielsen V. (2004): Valine requirement for maximal growth rate in weaned pigs. Livestock Production Science 88, 99-106
Wiltafsky M.K., Schmidtlein B. and Roth F.X. (2009): Estimates of the optimum dietary ratio of standardized ileal digestible valine to lysine for eight to twenty-five kilograms of body weight pigs. J. Anim. Sci. 87, 2544-2553
Danksagung
Die Autoren bedanken sich bei Ajinomoto Eurolysine S.A.S. für die Bereitstellung des L-Valins und der finanziellen Unterstützung dieser Untersuchungen
Autorenanschrift
Holger Kluge Martin-Luther-Universität Halle-Wittenberg Naturwissenschaftliche Fakultät III Institut für Agrar- und Ernährungswissenschaften Von Danckelmann Platz 2, 06120 Halle-Saale E-Mail: [email protected]

Firmensponsoring – LOHMANN ANIMAL HEALTH GmbH & Co.KG, Heinz-Lohmann-Straße 4, D-27472 Cuxhaven
Seite 254 8. BOKU-Symposium Tierernährung 2009

Rutzmoser et al.: Vorschlag eines Systems von nutzbaren Aminosäuren bei Mastbullen
8. BOKU-Symposium Tierernährung 2009 Seite 255
Vorschlag eines Systems von nutzbaren Aminosäuren bei Mast-bullen
Karl Rutzmoser, Thomas Ettle, Anton Obermaier
Institut für Tierernährung und Futterwirtschaft Grub
Einleitung Der Proteinbedarf für Milchkühe wird in Form des nutzbaren Rohproteins (nXP) ausgewiesen und be-zieht sich auf die Anflutung von Protein am Dünndarm. In der Arbeit von Schröder u. a. (2008) wird das nXP-System für Milchkühe auf die nutzbaren Aminosäuren Lysin und Methionin erweitert (nXAS-System). Dieses Verfahren soll auf Mastbullen angewendet werden. In diesem Beitrag werden die Grundlagen des Systems der nutzbaren Aminosäuren bei Mastbullen dargestellt, in einer weiteren Arbeit wird dieses Verfahren auf einen Fütterungsversuch angewendet.
Grundlagen des Verfahrens Ein Futterbewertungssystem umfasst die beiden Bereiche des Bedarfes des Tieres und der Lieferung durch Futtermittel. Der Bedarf wird hier auf der Ebene des Nettobedarfes an nutzbaren Aminosäuren gesehen, dieser teilt sich in Erhaltungs- und Leistungsbedarf (Ansatz im Körperprotein) auf. Bei der Lieferung durch die Futtermittel muss beim Wiederkäuer auf den im Pansen nicht abgebauten Teil des Futterproteins und auf das Mikrobenprotein abgehoben werden. Die Bewertung zielt wie bei Milchkü-hen auch bei Mastbullen auf Lysin und Methionin als begrenzende Aminosäuren ab.
Ableitung des Bedarfes Der Nettobedarf für die Leistung eines Mastbullen ergibt sich aus Zuwachsleistung, Proteinanteil im Zuwachs und Aminosäureanteil im Protein. Es können die Anteile von 6,4 % Lysin und 2,0 % Methio-nin des Rohproteins im Körpergewebe von Rindern angesetzt werden, welche von Schwab u. a. (2005) nach O’Connor u. a. (1993) genannt wurden. Diese können mit Gehalten im Schweinekörper verglichen werden. Nach GfE (2006) steigt Lysin von 6,3 % (5 bis 15 kg Lebendgewicht) auf 7,1 % im Rohprotein (60 bis 115 kg Lebendgewicht), was einen Anteil von 7,2 % Lysin im Rohprotein des Zu-wachses ergibt. Für Methionin wird mit 2,0 % im Rohprotein (ausser der oberen Gewichtsstufe 2,2 %) der gleiche Gehalt wie für Rinder angegeben.
Der Proteinanteil im Zuwachs wird in den Bedarfsableitungen für Fleckviehbullen (GfE 1995) über einen weiten Gewichtsbereich mit 175 g/kg Zuwachs angesetzt. Wegen des üblicherweise mit dem Gewicht steigenden Fettanteils ist im Regelfall ein leicht abfallender Proteinanteil im Zuwachs zu er-warten. Ein konstanter Wert von 175 g Rohprotein je kg Zuwachs kann jedoch als hinreichend genau angesehen werden, ausserdem erübrigt sich damit eine Zuordnung nach Leistung oder Gewicht.
Da Bedarfswerte auf Leistungen nach Tageszunahme angesetzt werden, sind die erforderlichen Ami-nosäuren auf den jeweiligen Zuwachs zu beziehen.
Der Erhaltungsbedarf ist als Nettobedarf an Rohprotein für die unvermeidlichen Verluste beschrieben (GfE 1995). Die unvermeidlichen Verluste setzen sich aus endogenem Harn- und Kot-N sowie den

Rutzmoser et al.: Vorschlag eines Systems von nutzbaren Aminosäuren bei Mastbullen
Seite 256 8. BOKU-Symposium Tierernährung 2009
Oberflächenverlusten zusammen. Auf Rohprotein berechnet (Faktor N * 6,25 = RP) werden folgende Gleichungen verwendet:
Endogene Harnverluste (Bezugsgröße: log10 des Lebendgewichtes W in kg): RP in Harn, endogen: ( 5,9206 * log( W ) – 6,76 ) * 6,25.
Endogene Kotverluste (Bezugsgröße: Futtertrockenmasseaufnahme TM in kg): RP in Kot, endogen: ( 2,19 * TM ) * 6,25.
Oberflächenverluste (Bezugsgröße: metabolisches Lebendgewicht W in kg): RP in Oberflächenverlusten: ( 0,018 * W**0,75 (exp) ) * 6,25.
Für die Aminosäurenanteile im Erhaltungsbedarf an Rohprotein sind nur Näherungen möglich. Der von Schröder u. a. abgeleitete Bedarf an nutzbarem Lysin und Methionin für Milchkühe entspricht nahezu dem Gehalt in der Milch, so dass keine deutliche Abweichung des Erhaltungsbedarfes erkennbar ist. Aus der Ableitung des Erhaltungsbedarfes von Schweinen (GfE 2006) ergibt sich auf der Ebene der Dünndarmverdaulichkeit ein Anteil von 2,4% Lysin und 0,6 % Methionin am Rohprotein für Erhaltung. Dies ist wesentlich durch den niedrigen Anteil von 17 % essentiellen Aminosäuren am Erhaltungs-Rohprotein bedingt. Rechnet man den Erhaltungsbedarf an Rohprotein vom Schwein auf das metabo-lische Körpergewicht von Bullen um, ist dieser etwa 2 bis 3-fach niedriger als die genannte Berech-nung aus den unvermeidlichen Verlusten bei Bullen. Werden die Aminosäurenanteile am Rohprotein vom Schwein auf den Erhaltungsbedarf von Bullen übertragen, führt das zu einer etwa 2 bis 3-fach höheren Aminosäurenzufuhr verglichen mit der Ableitung auf vergleichbarem metabolischen Körper-gewicht beim Schwein. Somit scheint gerechtfertigt, auf den Erhaltungsbedarf aus den unvermeidli-chen Verlusten der Bullen (GfE 1995) die Anteile von 2,4 % Lysin und 0,6 % Methionin (GfE 2006, Erhaltung Schwein) anzuwenden.
Berechnung der Aminosäurenzufuhr Die Zufuhr an nutzbarem Lysin und Methionin wird aus der Summe von unabgebautem Futterprotein und angeflutetem Mikrobenprotein am Dünndarm abgeleitet. Die Anflutung an nutzbarem Protein ergibt sich nach DLG (1997, Formel VIa) aus UDP (g) * 1,03. Nach Schröder u. a. (2008) kann ange-nommen werden, dass das Aminosäureprofil des unabgebauten Futterproteins dem des Futterproteins entspricht. Somit können mit entsprechenden Tabellenwerten von UDP und Aminosäuren, ggf. Unter-suchungswerten davon die Mengen aus dem unabgebauten Futterprotein berechnet werden.
Die Menge an Mikrobenprotein in g wird nach DLG (1997) aus dem ME-Gehalt und UDP-Anteil errech-net mit Formel VIa: (11,93-(6,82*UDP%))*ME MJ. Nach Schröder u. a. (2008) kann ein mittlerer An-teil von 7,79 g Lysin und 2,43 g Methionin je 100 g Mikrobenprotein angesetzt werden. Mit der kalku-lierten Menge an Mikrobenprotein ergibt sich damit die Zufuhr an nutzbaren Aminosäuren am Dünn-darm.
Darstellung von Ergebnissen Die dargestellten Werte von Leistungs- und Erhaltungsbedarf beziehen sich auf den Nettobedarf an Aminosäuren. Aus den aufgenommen Futtermitteln wird die Zufuhr bzw. die Anflutung am Dünndarm berechnet. Die ermittelten Nettobedarfswerte können darauf bezogen werden. Dies wird als Ausnut-zung der Anflutung für den Nettobedarf in Prozent dargestellt und ist so definiert:
Ausnutzung AS % = Nettobedarf g AS / Anflutung g AS * 100
Der erreichte Ausnutzungsgrad wird sinnvollerweise nach Gewichtsklassen abgestuft. Von der Modell-vorstellung her wird erwartet, dass bei limitierenden Aminosäuren die Zuwachsleistung davon be-grenzt wird und die Ausnutzung ein gewisses Maximum nicht überschreitet. Zusätzlich sollten die er-

Rutzmoser et al.: Vorschlag eines Systems von nutzbaren Aminosäuren bei Mastbullen
8. BOKU-Symposium Tierernährung 2009 Seite 257
reichten Zunahmen mit der errechneten Versorgung in Form der Ausnutzung verglichen werden. Als Kenngrößen bieten sich die Ausnutzung von Lysin, Methionin, Rohprotein und nutzbarem Protein an.
Wenn ein hinreichender Umfang an Daten verfügbar ist, könnte man aus Nettobedarf und erreichba-rer Ausnutzung (ohne Leistungsminderung) den Bruttobedarf an Aminosäuren in der Bullenmast, ge-gliedert nach Lebendgewicht (Erhaltungsbedarf) und Tageszunahme (Leistung) ableiten.
Berechnungsbeispiel Anhand eines frei gesetzten Beispieles soll der vorgeschlagene Rechengang aufgezeigt werden. Die Zunahmeleistung eines Mastbullen mit 300 kg Lebendgewicht sei 1300 g je Tag. Die Fütterung beste-he aus 12,5 kg Maissilage, 1 kg Weizen und 1 kg Sojaschrot.
In der Tabelle 1 wird die Anflutung von nutzbaren Aminosäuren aus den aufgenommenen Futtermit-teln errechnet. Dabei wird aufgeteilt in die Anflutung aus unabgebautem Protein aus den Futtermitteln mit deren UDP-Anteil und Gehalt an Aminosäuren sowie aus dem Mikrobenprotein, abhängig von ME (und UDP-Anteil) mit deren Aminosäurengehalten.
Tabelle 1: Berechnung der Anflutung aus den Futtermitteln (Beispiel Tagesfutter Mastbulle 300 kg, 1300 g Zunahme)
Maissilage Weizen Sojaschrot Gesamt- aufnahme
Frischmasse Aufnahme kg/Tag 12,5 1,0 1,0 Trockenmasse g/kg, Aufnahme 350 880 880 6135 Rohprotein g/kg TM, Aufnahme 82 138 477 900 UDP % 25 20 30 ME MJ/kg TM, Aufnahme 10,88 13,40 13,49 71,26 Lysin g/kg TM 1,98 3,90 28,87 Methionin g/kg TM 1,25 2,20 6,93 Unabgebautes Protein, UDP g 92 25 130 247 Lysin Anflutung aus UDP g 2,2 0,7 7,9 10,8 Methionin Anflutung aus UDP g 1,4 0,4 1,9 3,7 Mikrobenprotein g 487 125 117 729 Lysin Anflutung aus MP g 37,9 9,7 9,1 56,8 Methionin Anflutung aus MP g 11,8 3,0 2,9 17,7 Nutzbares Protein g 579 150 247 976 Nutzbares Lysin g 40,1 10,4 17,0 67,5 Nutzbares Methionin. G 13,2 3,4 4,7 21,4
Der Nettobedarf an nutzbaren Protein und Aminosäuren wird in der Tabelle 2 der Lieferung gegen über gestellt und als Ausnutzung gezeigt. Zunächst wird der Nettobedarf an nutzbarem Protein für Leistung und Erhaltung berechnet:
Leistungsbedarf: 1300 g Zunahme * 175 g Rohprotein/kg Zuwachs = 228 g Erhaltungsbedarf Harnverluste: ( 5,9206 * log( 300 kg ) – 6,76 ) * 6,25 = 49,4 Kotverluste: ( 2,19 * 6,135 kg TM ) * 6,25 = 84,0 Oberflächenverluste: ( 0,018 * 300 kg**0,75 ) * 6,25 = 8,1

Rutzmoser et al.: Vorschlag eines Systems von nutzbaren Aminosäuren bei Mastbullen
Seite 258 8. BOKU-Symposium Tierernährung 2009
Tabelle 2: Berechnung des Nettobedarfes und Ausnutzung von nutzbarem Protein und Aminosäuren (Beispiel Mastbulle 300 kg, 1300 g Zunahme)
Rohprotein nXP nLys nMet Leistungsbedarf 1300 g Zunahme 228 228 14,6 4,6 Erhaltungsbedarf Verluste 142 141 3,4 0,8 Summe Nettobedarf 369 369 18,0 5,4 Aufnahme, Anflutung 900 976 67,5 21,4 Ausnutzung % 41 38 27 25
In diesem einfachen Beispiel erreicht das nXP mit 38 % eine niedrigere Ausnutzung als das Rohprote-in, was durch die höhere Anflutung an nXP bedingt ist. Folglich kann das Rohprotein in diesem Bei-spielsfall als stärker begrenzend angesehen werden. Die Werte der Ausnutzung liegen bei den nutzba-ren Aminosäuren deutlich niedriger. Dies deutet darauf hin, dass im Vergleich zum Bedarf relativ viel nLys und nMet zur Verfügung steht.
Literatur
Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie (GfE) (1995): Energie- und Nährstoffbedarf landwirtschaftlicher Nutztiere. Nr. 6 Empfehlungen zur Energie- und Nährstoffversorgung der Mastrinder. DLG-Verlag Frankfurt (Main)
Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie (GfE) (2006): Empfehlungen zur Energie- und Nährstoffversorgung von Schweinen. DLG-Verlag Frankfurt (Main)
O’Conor, J. D., Sniffen, C. J., Fox, D. G., Chalupa, W. (1993): A net carbohydrate and protein system for evauating cattle diets. IV. Predecting amino acid adequacy. Journal of Animal Science 71, 1289 – 1311.
Schröder, A., Bennett, R., Rulquin, H. (2008): Rationsgestaltung mit Aminosäuren: Das nXAA-System – eine Erweiterung des nXP-Systems. Kongressband 2008 Jena, VDLUFA-Schriftenreihe Bd. 64/2008, ISBN 978-3-941273-05-4, 305 – 312.
Schwab, C. G., Huhtanen, B., Hunt, C. W., Hvelund, T. (2005): Nitrogen Requirements of Cattle. In: Pfeffer, E., Hristov, A. N.: Nitrogen and Phosporus Nutrition of Cattle – Reducing the Environmental Impact of Cattle Operatations.CABI Publishing, ISBN 0 85199 013 4, 13 – 70.
Autorenanschrift
Dr. Karl Rutzmoser Institut für Tierernährung und Futterwirtschaft Grub Prof.-Dürrwaechter-Pl. 3, 85586 Poing E-Mail: [email protected]

Ettle et al.: Anwendung des Systems von nutzbaren Aminosäuren bei einem Fütterungsversuch mit Mastbullen
8. BOKU-Symposium Tierernährung 2009 Seite 259
Anwendung des Systems von nutzbaren Aminosäuren bei einem Fütterungsversuch mit Mastbullen
Thomas Ettle, Karl Rutzmoser, Anton Obermaier
Institut für Tierernährung und Futterwirtschaft Grub
Einleitung Während in Deutschland der Proteinbedarf für Milchkühe in Form des nutzbaren Rohproteins (nXP) ausgewiesen wird (GfE, 2001), beziehen sich Versorgungsempfehlungen für Mastbullen auf das Roh-protein (GfE, 1995). In anderen Ländern wie Frankreich oder den Niederlanden werden in Futterwert-tabellen für Wiederkäuer Angaben zu verfügbaren Aminosäuren gemacht (z.B. Sauvant et al. 2002). Im INRA-System beziehen sich Bedarfsangaben für nutzbare Aminosäuren beim Milchvieh auf Arbeiten von Rulquin et al. (2003). Da die wesentlichen Grundlagen der Proteinbewertungssysteme von INRA bzw. GfE (2001) für Milchvieh ähnlich sind, zeigten Schröder et al. (2008) einen Ansatz, auf Basis der französischen Daten das deutsche nXP System auf die nutzbaren Aminosäuren Lysin und Methionin zu erweitern. Rutzmoser et al. (2009) stellen in einem Beitrag dieses Tagungsbandes dar, wie das Ver-fahren der nutzbaren Aminosäuren auf Mastbullen übertragen werden kann. In der vorliegenden Ar-beit werden diese Bedarfsableitungen und die Zufuhr an nutzbarem Lysin und Methionin auf Daten aus einem Fütterungsversuch mit Mastbullen angewendet.
Material und Methoden Im neuen Rindermastversuchsstall in Grub wurde ein Fütterungsversuch mit 135 Mastbullen der Rasse Fleckvieh im Lebendmassebereich von 220 bis ca. 730 kg durchgeführt. Dazu wurden drei Futtergrup-pen gebildet, die sich jeweils auf 4 Buchten verteilten, davon 2 Buchten Vollspaltenboden (jeweils eine Bucht mit und ohne Gummiauflage) und 2 Buchten Tretmist je Futtergruppe. Die Tiere wurden zeitlich versetzt in mehreren Wellen im Abstand von etwa 2 Wochen aufgestallt. In den Futtergruppen wur-den getrocknete Weizen-Gerste-Zucker-Schlempe (Schlempe), Rapsextraktionsschrot (Raps) und So-jaextraktionsschrot (Soja) als alleinige Eiweißfuttermittel in einer Mischration auf Basis Maissilage und Stroh eingesetzt. Absicht war es, für die drei Versuchsgruppen isonitrogene und isoenergetische Rati-onen zu erstellen. Die Mischrationen wurden für die Mastabschnitte Anfangs-, Mittel- und Endmast hinsichtlich der Energie- und Rohproteinversorgung angepasst. Tabelle 1 zeigt die Zusammensetzung der Rationen für die jeweiligen Mastabschnitte und Fütterungsgruppe. Das Kraftfutter in den Mischra-tionen setzte sich aus Körnermais, Weizen, Mineralfutter und dem jeweiligen Proteinträger zusammen. In Mastabschnitt 1 lag der Anteil an Schlempe, Raps- und Sojaextraktionsschrot am Kraftfutter bei 63,4; 63,4 und 51,3 %, in den Mastabschnitten 2 und 3 lagen die entsprechenden Werte jeweils bei 50,0; 50,0 und 38,0 %. Nähere Angaben zu den Nährstoff- und Energiegehalten finden sich bei Ettle et al. (2009).
Zur Auswertung wurden Daten von 130 Tieren herangezogen. Im Gegensatz zur Arbeit von Ettle et al. (2009), bei der als Folge der Anwendung eines komplexeren statistischen Modelles LSQ-Werte darge-stellt werden, beziehen sich die hier gezeigten Mittelwerte auf die unkorrigierten Daten.
Tabelle 2 zeigt die für die eingesetzten Futtermittel kalkulierten Gehalte an nXP, nutzbarem Lysin (nLys) und nutzbarem Methionin (nMet) sowie die XP-Gehalte. Für die Proteinträger Schlempe, Raps- bzw. Sojaextraktionsschrot wurden analysierte Aminosäurengehalte (1,80; 5,42 und 5,95 % Lys im XP

Ettle et al.: Anwendung des Systems von nutzbaren Aminosäuren bei einem Fütterungsversuch mit Mastbullen
Seite 260 8. BOKU-Symposium Tierernährung 2009
und 1,39; 1,97 und 1,33 % Met im XP für Schlempe, Raps- und Sojaextraktionsschrot) verwendet. Für Maissilage und Stroh sowie die anderen Kraftfutterkomponenten (Degussa, 1996) wurden Tabellen-werte für die Bruttogehalte an Lysin und Methionin angesetzt. Die prozentualen UDP-Gehalte wurden den DLG-Tabellen für Wiederkäuer (1997) entnommen, die XP-und ME-Versorgung ergibt sich aus den Weender Analysen der Einzelkomponenten.
Tabelle 1: Zusammensetzung der Mischrationen (% der TM) in den jeweiligen Mastabschnitten
Behandlung Schlempe Raps Soja Anfangsmast Maissilage 68,4 69,6 69,6 Stroh 3,9 2,6 3,9 Kraftfutter 27,7 27,8 26,5 Mittelmast Maissilage 69,3 69,5 69,5 Stroh 3,9 3,9 3,9 Kraftfutter 26,8 26,6 26,6 Endmast Maissilage 72,8 73,0 73,1 Stroh 3,7 3,7 3,7 Kraftfutter 23,5 23,3 23,2
Tabelle 2: Gehalte an XP, nXP, nLys und nMet (g/kg T) in Maissilage, Stroh und den eingesetzten Kraftfuttermischungen für die Gruppen Schlempe, Raps und Soja
Gehalte, g/kg TM XP nXP nLys nMet
Maissilage 80,0 135,5 9,5 3,1 Stroh 45,0 79,8 5,1 1,6 Kraftfutter, Anfangsmast Schlempe 286,3 220,4 10,6 4,3 Raps 288,2 199,3 13,3 4,5 Soja 290,8 210,2 14,4 4,3 Kraftfutter, Mittelmast Schlempe 249,6 206,9 10,6 4,2 Raps 251,2 190,3 12,7 4,3 Soja 245,1 196,8 13,4 4,1 Kraftfutter, Endmast Schlempe 249,6 206,9 10,6 4,2 Raps 251,2 190,3 12,7 4,3 Soja 245,1 196,8 13,4 4,1
Ergebnisse und Diskussion Die TM-Aufnahme lag über den gesamten Versuch betrachtet im Mittel aller Versuchsgruppen bei 9,4 kg/Tier und Tag mit einem nur numerischen Vorteil für die Gruppe Raps. Die Tiere der Gruppe Soja hatten im Mittel des gesamten Versuches deutlich (p<0,05) höhere Zunahmen (1602 g/d) als die Tie-re der Gruppe Schlempe (1493 g/d), während der tägliche Zuwachs der Tiere der Gruppe Raps (1549 g/d) sich nicht von den Vergleichsgruppen unterschied. Eine detaillierte Darstellung zu Mast- und Schlachtleistungen findet sich bei Ettle et al. (2009).

Ettle et al.: Anwendung des Systems von nutzbaren Aminosäuren bei einem Fütterungsversuch mit Mastbullen
8. BOKU-Symposium Tierernährung 2009 Seite 261
Tabelle 3: Leistungsdaten und Parameter der Proteinversorgung in der Anfangsmast
Versuchsgruppe Schlempe Raps Soja Lebendgewicht kg 324 304 317 Zuwachs g/d 1584 1666 1787 TM-Aufnahme kg/d 7,37 7,20 7,42 nxP-Anflutung g/d 1157 1093 1137 nLys-Anflutung g/d 71,1 75,2 78,8 nMet-Anflutung g/d 25,0 24,5 24,5 nxP-Bedarf g/d 438 448 473 nLys-Bedarf g/d 23,8 24,8 26,4 nMet-Bedarf g/d 6,51 6,77 7,22 nXP-Ausnutzung % 37,9 41,2 41,8 nLys-Ausnutzung % 33,5 33,1 33,6 nMet-Ausnutzung % 26,0 27,8 29,6
Tabelle 4: Leistungsdaten und Parameter der Proteinversorgung in der Mittelmast
Versuchsgruppe Schlempe Raps Soja Lebendgewicht kg 478 470 485 Zuwachs g/d 1388 1503 1501 TM-Aufnahme kg/d 9,27 9,66 9,32 nxP-Anflutung g/d 1414 1429 1395 nLys-Anflutung g/d 89,2 98,3 96,6 nMet-Anflutung g/d 31,0 31,9 30,8 nxP-Bedarf g/d 439 464 459 nLys-Bedarf g/d 22,2 23,8 23,6 nMet-Bedarf g/d 6,03 6,46 6,43 nXP-Ausnutzung % 31,1 32,5 33,1 nLys-Ausnutzung % 24,9 24,2 24,6 nMet-Ausnutzung % 19,5 20,3 21,0
Tabelle 5: Leistungsdaten und Parameter der Proteinversorgung in der Endmast
Versuchsgruppe Schlempe Raps Soja Lebendgewicht kg 634 638 640 Zuwachs g/d 1393 1475 1481 TM-Aufnahme kg/d 11,1 11,2 11,2 nxP-Anflutung g/d 1674 1635 1653 nLys-Anflutung g/d 107,0 112,7 115 nMet-Anflutung g/d 36,9 36,9 37,0 nxP-Bedarf g/d 472 487 489 nLys-Bedarf g/d 23,0 24,1 24,2 nMet-Bedarf g/d 6,25 6,54 6,56 nXP-Ausnutzung % 28,2 29,9 29,6 nLys-Ausnutzung % 21,5 21,4 21,1 nMet-Ausnutzung % 16,9 17,8 17,8

Ettle et al.: Anwendung des Systems von nutzbaren Aminosäuren bei einem Fütterungsversuch mit Mastbullen
Seite 262 8. BOKU-Symposium Tierernährung 2009
In Tabelle 3 bis 5 sind die mittleren Lebendgewichte je Abschnitt, die Tageszunahmen und Futterauf-nahmen und die daraus resultierenden Anflutungs- und Bedarfswerte (Nettobedarf) für nXP, nutzbares Lysin und Methionin dargestellt.
Mit 1500 – 1600 g wurden im Mittel der Gesamtmast in allen Gruppen sehr hohe Zuwachsraten er-reicht. Dabei war die Rohproteinversorgung vor allem in der Anfangsmast gegenüber derzeit gültigen Versorgungsempfehlungen in allen Gruppen relativ niedrig (Ettle et al. 2009). Ein Grund für die hohen Zuwachsleistungen zu Beginn der Mast könnten relativ verhaltene Zuwachsraten während der Fres-seraufzucht sein.
Die Anflutung an nXP und nMet lag innerhalb der einzelnen Mastabschnitte im Vergleich der Versuchs-gruppen auf ähnlichem Niveau (Tab. 3 bis 5). Die Anflutung an nLys war in der Gruppe Schlempe tendentiell niedriger als in den Vergleichsgruppen, erklärbar aus den niedrigeren Brutto-Lysingehalten in der Schlempe.
Der errechnete Nettobedarf an nXP, nLys und nMet folgt in gewissem Maße den Zunahmen, wobei die Unterschiede durch den Erhaltungsbedarf abgeschwächt sind. Die Ausnutzung an nXP, aber auch an nMet, war lediglich in der Anfangsmast in der Gruppe Schlempe erniedrigt. In diesem Mastabschnitt waren jedoch auch die Unterschiede in den täglichen Zunahmen zur Gruppe Soja am höchsten.
In allen Mastabschnitten war die Ausnutzung des nXP höher, als die der nutzbaren Aminosäuren, was darauf hindeutet, dass nXP die Leistung am ehesten begrenzte. Dabei lag die Versorgung mit Rohpro-tein um etwa 150 g/Tag (Anfangsmast) bis 300 g/Tag (Endmast) unter der errechneten Anflutung an nXP. Daraus leitet sich die Frage ab, ob für die Mikrobenproteinsynthese im Pansen tatsächlich genü-gend rezyklierter Stickstoff verfügbar war.
Abbildung 1: Zusammenhang zwischen täglicher Aufnahme an XP (g), Anflutung an nXP (g), nLys (g) sowie nMet (g) und täglichem Zuwachs (g)

Ettle et al.: Anwendung des Systems von nutzbaren Aminosäuren bei einem Fütterungsversuch mit Mastbullen
8. BOKU-Symposium Tierernährung 2009 Seite 263
Abbildung 1 zeigt anhand der Tiermittelwerte über die gesamte Versuchsperiode den Zusammenhang zwischen Zuwachs und Zufuhr an Rohprotein bzw. Anflutung an nutzbarem Rohprotein, nutzbarem Lysin und nutzbarem Methionin. Der Zusammenhang zwischen nXP-Anflutung und Zuwachs ist mit einem R2 von 0,12 relativ gering, die Beziehung zwischen XP-Zufuhr und Zuwachs erhöht das R2 auf 0,17. Das nutzbare Lysin als erklärende Variable erbringt eine Verbesserung des R2 auf 0,24. Die Ver-wendung des nutzbaren Methionins ergibt eine nahezu gleich niedrige Erklärung des Zuwachses, wie die Verwendung des nXP. Für den Zusammenhang zwischen Zuwachsraten und täglicher TM- bzw. ME-Aufnahme (Daten hier nicht dargestellt) errechnet sich ein R2 von 0,15 bzw. 0,16. Obwohl die hier dargestellten Bestimmtheitsmaße insgesamt nicht sehr hoch sind, scheint die Einschätzung der Ver-sorgungslage über das nLys bei vorliegendem Datenmaterial doch eine verbesserte Erklärung der Zuwachsleistungen zu bieten.
Schlussfolgerungen In vorliegender Untersuchung wurde ein System der nutzbaren Aminosäuren auf einen Fütterungsver-such bei Mastbullen, in dem 3 unterschiedliche Eiweißträger eingesetzt wurden, angewandt. Insbe-sondere aus dem Zusammenhang zwischen Angebot an XP, nXP oder den nutzbaren Aminosäuren Lysin und Methionin und den Zuwachsleistungen lässt sich erkennen, dass die Bewertung auf Basis des nLys die Variation der täglichen Zunahmen besser erklären kann.
Bei der Kalkulation der Anflutung von nutzbarem Protein und Aminosäuren ist zu prüfen, ob die Sum-me an zugeführtem Rohprotein oder das im Pansen abgebaute Rohprotein bedeutsam sein kann. Für eine abschließende Einordnung erscheinen weitere Auswertungen, beispielsweise über enger gefasste Mastabschnitte oder mit einem größeren Datenmaterial angebracht.
Literatur
DLG (1997): Futterwerttabellen für Wiederkäuer. DLG-Verlags-GmbH, Frankfurt a. Main.
Ettle, T., Obermaier, A., Preissinger, W., Hitzlsperger, L., Meiser, H., Spiekers, H., Potthast, C. (2009): Einsatz von getrockneter Getreideschlempe (Weizen, Gerste, Zucker), Raps- oder Sojaextraktionsschrot in der intensiven Mast von Bullen der Rasse Fleckvieh. Forum angewandte Forschung in der Rinder- und Schweinefütterung. 58-61.
GfE (1995): Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie. Energie- und Nährstoffbedarf landwirtschaftlicher Nutztiere. Nr. 6 Empfehlungen zur Energie- und Nährstoffversorgung der Mastrinder. DLG-Verlag Frankfurt (Main).
GfE (2001): Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie. Energie- und Nährstoffbedarf landwirtschaftlicher Nutztiere. Nr. 8 Empfehlungen zur Energie- und Nährstoffversorgung der Milchkühe und Aufzuchtrinder. DLG-Verlag Frankfurt (Main).
Schröder, A., Bennett, R., Rulquin, H. (2008): Rationsgestaltung mit Aminosäuren: Das nXAA-System – eine Erweiterung des nXP-Systems. Kongressband 2008 Jena, VDLUFA-Schriftenreihe Bd. 64/2008, ISBN 978-3-941273-05-4, 305 – 312.
Sauvant, D., Perez, J.-M., Tran, G. (2002): Tables of composition and nutritional value of feed materials.
Rulquin, H., Pisulewski, P.M., Verite, R., Guinard, J. (1993): Milk production and composition as a function of postruminal lysine and methionine supply: a nutrient-response approach. Livest. Prod. Sci. 37, 69-90.
Rutzmoser, K., Ettle, T., Obermaier, A. (2009): Vorschlag eines Systems von nutzbaren Aminosäuren bei Mastbullen. Tagungsband 8. BOKU-Symposium Tierernährung.
Autorenanschrift
Dr. Thomas Ettle Institut für Tierernährung und Futterwirtschaft Grub Prof.-Dürrwaechter-Pl. 3, 85586 Poing E-Mail: [email protected]

Hošková et al.: The effect of feeding DDGS on broiler meat composition
Seite 264 8. BOKU-Symposium Tierernährung 2009
The effect of feeding DDGS on broiler meat composition
Š. Hošková, A. Vašátková, M. Balabánová, M. Hošek, L. Zeman
Mendel University of Agriculture and Forestry in Brno
Introduction Distiller´s dried grains with solubles (DDGS) is a by-product of the fuel ethanol industry. During the production of alcohol, starch is removed from the grain and converted to alcohol and carbon dioxide. As a result of starch removal, the concentration of the remaining nutrients in the grain increases ap-proximately threefold (SPIEHS et al., 2002). A vast increase in ethanol production over the last 5 to 10 years has led to an increased supply of DDGS that is available for livestock feed (Noll et al., 2007). DDGS is increasing rapidly and poultry nutritionists are making increasing use of it in their diets (WANG et al., 2007). LUMPKINS et al. (2004) concluded that broilers can be fed 6 % DDGS in the starter period and 12 to 15 % DDGS in the grower and finishing stages, respectively, without affecting growth. DALE and BATAL (2003) used 0, 6, 12 and 18 % DDGS in a 42 d growout study and reported that 12 % DDGS resulted in a slight decrease in performance during the started period while 18 % DDGS had a negative impact on body weight and feed conversion over 42 d period.
Factors such as selection of grains, type of fermentation, and drying temperature and duration can influence the nutritional and physical properties of DDGS (SPIEHS et al., 2002). In the process of dry-ing DDGS the material is exposed to temperatures of approximately 315 °C. It has been reported that excessive heating leads to a decrease in amino acid (AA) availability, specifically Lys (LUMPKINS and BATAL, 2005). According to BATAL and DALE (2006) the average total concentration and digestibility coefficients of several most limiting AA for the 8 DDGS samples were as follows: Lys, 0.71 % (70); Met, 0.54 % (87); cystine, 0.56 % (74); Thr, 0.96 % (75); Val, 1.33 % (80); Ile, 0.97 % (83); and Arg, 1.09 % (84). Correlations were found among digestible Lys, Thr, Arg, His and Trp and the yel-lowness and lightness of the DDGS samples. In general, DDGS samples that were more yellow and lighter in color had higher total and digestible AA levels.
The aim of our experimental supervision was detected the effect of feeding DDGS on broiler meat composition – dry matter, content of crude protein in dry matter and content of fat in dry matter in breast and thigh meat.
Materials and Methods Growth experiment was realized on fattening poultry station Lípa u Havlíčkova Brodu. There were used 1 day old male broilers of Ross 308 hybrid combination. There were used 500 chickens which were divided into 5 stalls by 100 pieces. Average weight of chicks was 39 g. The birds were housed on deep litter. There were installed corresponding numbers of drinker and bunk feeders. Feed and water were available ad libitum. Temperature and humidity were measured by Comet L3120 on two points of animal house.
Broilers were fed by a starter until 12 day of experiment; it was a standard commercial diet BR1. Ex-periment feed mixtures BR2 were fed from 13 day to 35 day. Experimental diets were formulated to contain: 1) 0 % DDGS (control group), 2) 10 % DDGS, 3) 15 % DDGS, 4) 20 % DDGS, 5) 25 % DDGS. There were used wheat-triticale DDGS from ethanol plant in the Czech Republic. Composition of the feed mixtures is presented in the Table 1.
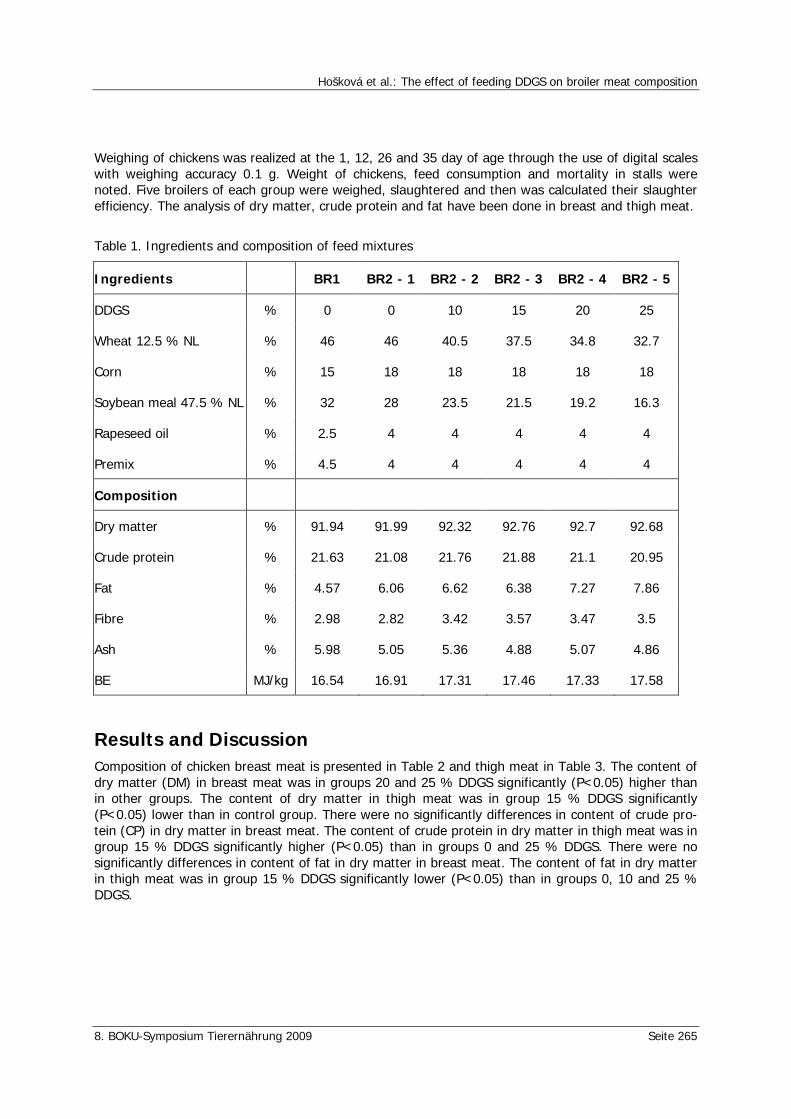
Hošková et al.: The effect of feeding DDGS on broiler meat composition
8. BOKU-Symposium Tierernährung 2009 Seite 265
Weighing of chickens was realized at the 1, 12, 26 and 35 day of age through the use of digital scales with weighing accuracy 0.1 g. Weight of chickens, feed consumption and mortality in stalls were noted. Five broilers of each group were weighed, slaughtered and then was calculated their slaughter efficiency. The analysis of dry matter, crude protein and fat have been done in breast and thigh meat.
Table 1. Ingredients and composition of feed mixtures
Ingredients BR1 BR2 - 1 BR2 - 2 BR2 - 3 BR2 - 4 BR2 - 5
DDGS % 0 0 10 15 20 25
Wheat 12.5 % NL % 46 46 40.5 37.5 34.8 32.7
Corn % 15 18 18 18 18 18
Soybean meal 47.5 % NL % 32 28 23.5 21.5 19.2 16.3
Rapeseed oil % 2.5 4 4 4 4 4
Premix % 4.5 4 4 4 4 4
Composition
Dry matter % 91.94 91.99 92.32 92.76 92.7 92.68
Crude protein % 21.63 21.08 21.76 21.88 21.1 20.95
Fat % 4.57 6.06 6.62 6.38 7.27 7.86
Fibre % 2.98 2.82 3.42 3.57 3.47 3.5
Ash % 5.98 5.05 5.36 4.88 5.07 4.86
BE MJ/kg 16.54 16.91 17.31 17.46 17.33 17.58
Results and Discussion Composition of chicken breast meat is presented in Table 2 and thigh meat in Table 3. The content of dry matter (DM) in breast meat was in groups 20 and 25 % DDGS significantly (P<0.05) higher than in other groups. The content of dry matter in thigh meat was in group 15 % DDGS significantly (P<0.05) lower than in control group. There were no significantly differences in content of crude pro-tein (CP) in dry matter in breast meat. The content of crude protein in dry matter in thigh meat was in group 15 % DDGS significantly higher (P<0.05) than in groups 0 and 25 % DDGS. There were no significantly differences in content of fat in dry matter in breast meat. The content of fat in dry matter in thigh meat was in group 15 % DDGS significantly lower (P<0.05) than in groups 0, 10 and 25 % DDGS.

Hošková et al.: The effect of feeding DDGS on broiler meat composition
Seite 266 8. BOKU-Symposium Tierernährung 2009
Table 2. Composition of breast meat
dry matter (%) CP in DM (%) fat in DM (%)
0 % DDGS 25.05±0.201a 93.50±2.503a 3.63±0.928a
10 % DDGS 24.61±0.449a 91.75±0.790a 3.93±0.419a
15 % DDGS 24.65±0.207a 92.60±0.940a 3.67±0.752a
20 % DDGS 26.65±0.147b 91.65±1.014a 4.16±1.023a
25 % DDGS 26.21±0.332b 90.86±1.152a 4.78±1.200a
Table 3. Composition of thigh meat
dry matter (%) CP in DM (%) fat in DM (%)
0 % DDGS 27.73±0.964b 65.32±2.517a 30.84±2.665b
10 % DDGS 26.75±2.518ab 65.63±7.977ab 34.78±4.261b
15 % DDGS 25.72±0.350a 70.78±1.344b 25.60±1.146a
20 % DDGS 28.62±0.922ab 66.99±3.757ab 28.79±3.875ab
25 % DDGS 29.12±0.486ab 64.73±2.526a 32.74±2.058b
Conclusion The level of DDGS in broiler diets didnot affect in principle the composition of chicken breast and thigh meat.
Acknowledgments
This project was supported by IGA of MZLU as project no. IG290101.
References
BATAL, A. B., DALE, N. M. True metabolize energy and amino acid digestibility of distillers dried grains with solubles. Journal of Applied Poultry research. 2006, vol. 15, no. 1, s. 89-93.
DALE, N., BATAL, A B. Nutritional value of distillers dried grains and solubles fo poultry. In 19th Annual Carolina Nutrition Conf.. NC : Research Triangle Park, 2003. s. 1-6.
LUMPKINS, B. S., BATAL, A. B. The Bioavailability of Lysine and Phosphorus in Distillers Dried Grains with Solubles. Poultry Science. 2005, no. 84, s. 581-586.

Hošková et al.: The effect of feeding DDGS on broiler meat composition
8. BOKU-Symposium Tierernährung 2009 Seite 267
LUMPKINS, B. S., BATAL, A. B., DALE, N. M. Evaluation of dried distillers grain solubles as a feed ingredient for broilers. Poultry Science. 2004, no. 83, s. 1891-1896.
NOLL, S. E., PARSONS, C. M., DOZIER, W. A. Formulating poultry diets with DDGS-How far can we go. In Proceedings 5th Mid Atl. Nutr. Conf. N. G. Zimmerman. [s.l.] : Univ. Maryland College Park, 2007.
SPIEHS, M. J., WHITNEY, M. H., SHURSON, G. C. Nutrient database for distiller´s dried grains with solubles produced from new ethanol plants in Minnesota and South dakota. Journal of Animal Science. 2002, no. 80, s. 2639-2645.
WANG, Z., et al. Effect of Rapid Multiple Changes in Level of Distillers Dried Grains with Solubles (DDGS) in Broiler Diets on Performance and Carcass Characteristics. International Journal of Poultry Science. 2007, vol. 6, no. 10, s. 725-731.
ZEMAN, L. et al. Katalog krmiv. VÚVZ Pohorelice, 1995, s. 465.
Corresponding author
Ing. Sarka Hoskova Department of Animal Nutrition and Forage Production Mendel University of Agriculture and Forestry in Brno Zemedelska 1, 613 00, Brno, Czech Republic E-mail: [email protected]

Brunauer et al.: Qualität des intramuskulären Fettes von Grauviehalmochsen im Vergleich zum durchschnittlichen österreichischen Rindfleisch mit besonderem Augenmerk auf essentielle n6- und n3-Fettsäuren
Seite 268 8. BOKU-Symposium Tierernährung 2009
Qualität des intramuskulären Fettes von Grauviehalmochsen im Vergleich zum durchschnittlichen österreichischen Rindfleisch mit besonderem Augenmerk auf essentielle n6- und n3-Fettsäuren
Christoph Brunauer1, Christiane Mair1, Matthias Schreiner2, Wilhelm Windisch1 1 Department für Lebensmittelwissenschaften und -technologie, Abteilung Tierische
Lebensmittel, Tierernährung und Ernährungsphysiologie, Universität für Bodenkul-tur Wien
2 Department für Lebensmittelwissenschaften und -technologie, Abteilung Lebensmit-telchemie, Universität für Bodenkultur Wien
Einleitung Die Summe der Eigenschaften, die den Genuss-, Gesundheits- und Verarbeitungswert des Magerflei-sches beeinflussen, wird als Fleischqualität bezeichnet. Den ernährungsphysiologischen Wert von Fleisch bestimmen hauptsächlich die Amino- und Fettsäurezusammensetzung. Sensorisch bedeutend sind beim Rind die Zartheit des Fleisches und der intramuskuläre Fettgehalt. Parameter, welche zur Bestimmung der Fleischqualität herangezogen werden, sind der intramuskuläre Fettgehalt, pH-Wert (24 h nach der Schlachtung), Scherwert nach Warner-Bratzler (24 h nach der Schlachtung und 14 Tage nach der Reifung), Dripverlust, Farbhelligkeit und Marmorierung. Den höchsten intramuskulären Fettgehalt hat der Musculus longissimus [Brandscheid 2007].
Fett erfüllt eine wichtige Funktion zum Beispiel als Trägersubstanz von Aromen und lebensnotwendi-gen Vitaminen. Die Beschaffenheit des Nahrungsfettes selbst gewinnt aber in den letzten Jahren im-mer mehr an Bedeutung. Besonders hervorzuheben ist hier die Wirkung der mehrfach ungesättigten Fettsäuren (Polyunsaturated Fatty Acids = PUFA) der n3- (oder ω3) und n6- (ω6) Reihe, welche zum Beispiel vor Herz- Kreislauferkrankungen schützen [Djoussé 2001].
Verschiedene Beurteilungskriterien wurden entwickelt, um die Qualität der Fette vergleichbar zu ma-chen. Drei wichtige Parameter sind das n6:n3-Fettsäureverhältnis, der P:S-Quotient (Verhältnis zwi-schen PUFAs und gesättigten Fettsäuren (= SFA)) und das P:M:S-Verhältnis. Letztgenanntes stellt PUFAs und MUFAs (Monounsaturated Fatty Acids) den SFAs gegenüber.
Es wurde nachgewiesen, dass durch Grasfütterung bei Rindern ein signifikant höherer Gehalt an PU-FAs und n3-Fettsäuren intramuskulär eingelagert wird, als durch Fütterung mit Maissilage und Kraft-futter [Dannenberger et al. 2007].
Die Kategorie beeinflusst ebenfalls den Fettgehalt. So haben Jungstiere eine geringere Marmorierung (Fettverteilungsstruktur im Fleisch) als die anderen Kategorien und Ochsen erzielen bei gleicher Fütte-rung einen höheren Fetteinlagerungsgrad als Stiere [Brandscheid 2007].
In der vorgelegten Arbeit sollte der Frage nachgegangen werden, ob eine extensive Weidemast mit mindestens einer Alpung (Tiroler Grauviehalmochsen) im Vergleich zur intensiven Stallmast von durchschnittlichem österreichischen Rindfleisch (Fleckviehstiere) unterschiedliche Auswirkungen auf die Zusammensetzung/Qualität des intramuskulären Fettes hat. Im Zuge dessen sollte auch der Tro-ckenmasse-, Rohfett- und Rohproteingehalt des Muskelfleisches ermittelt werden.

Brunauer et al.: Qualität des intramuskulären Fettes von Grauviehalmochsen im Vergleich zum durchschnittlichen österreichischen Rindfleisch mit besonderem Augenmerk auf essentielle n6- und n3-Fettsäuren
8. BOKU-Symposium Tierernährung 2009 Seite 269
Material und Methoden Die 33 Muskelproben des Tiroler Grauviehalmochsen stammen vom Zerlegebetrieb Hörtnagl (Hall in Tirol). Die Fleischproben der 22 Fleckviehmaststiere, welche für das durchschnittliche Rindfleischan-gebot in Österreich stehen, wurden von der Firma Alpenrind Salzburg (Schlachthof Bergheim) zur Verfügung gestellt. Das Schlachtgewicht lag bei den Maststieren mit 397 kg deutlich über dem der Almochsen (283 kg), obwohl sie mit einem Durchschnittsalter von 18 Monaten um 8 Monate jünger als die Almochsen waren. Als standardisiertes Probenteilstück wurde der Musculus longissimus aus dem linken Hinterviertel, unter Aussparung des Anschnittes, entnommen. Die Muskelfleischproben wurden vom äußeren Binde- und Fettgewebe befreit und die Trockenmasse im Zuge der Gefriertrock-nung bestimmt. Nach dem Homogenisieren des getrockneten Probenmaterials erfolgte die Rohfett- (Soxhletextraktion) und Rohproteinbestimmung (Stickstoffbestimmung nach Kjeldahl) nach Standard-methoden [Naumann und Bassler 1997].
Für die Bestimmung des Fettsäuremusters wurden je 500 mg Probenmaterial in Doppelbestimmung in 16 x 160 mm Pyrex Röhrchen genau eingewogen. Die Veresterung wurde nach der „One-Step Methy-lation“-Methode [Sukhija und Palmquist 1988] durchgeführt. Dabei finden die Fettextraktion aus dem Pro-benmaterial und die Transmethylierung der Fette in einem Schritt statt. Toluol wurde als Extraktions-mittel verwendet. Als Interner Standard wurde ein C19:0-Fettsäurestandard (Fluka Analytical Nonade-cansäure 1 g) verwendet und zur Überprüfung der Genauigkeit der Methode ein zertifizierter BCR-Standard (Referenz-Material Nr. 163 Beef/Pig FAT Blend) herangezogen. Je 1 µl Fettsäuremethylester (FAME) wurde mit dem Gaschromatographen (Agilent Technologies 7890A) analysiert.
Die statistische Auswertung der gesammelten Daten erfolgte mit dem Statistikprogramm SAS 9.1.3 (SAS Inst., Inc., Cory., NC). Die Werte wurden mit dem Kolmogorov-Smirnov-Test auf Normalvertei-lung geprüft und als arithmetische Mittelwerte +/- Standardfehler der Mittelwerte (SEM) angeben. Für die verschiedenen Einflussgrößen kam der t-Test zum Einsatz, um signifikante Unterschiede zwischen den beiden Gruppen heraus zu arbeiten. Bei der Fleisch- (E, U, R, O, P) und Fettklassifizierung (1, 2, 3, 4, 5) wurde jedoch anstatt des t-Tests der Wilcoxon-Mann-Whitney-Test durchgeführt. Das Signifi-kanzniveau α wurde mit 0,05 angenommen.
Ergebnisse Im Mittel entsprach das Fleisch der Almochsen der Fleischklasse „R“ und der Fettklasse „2“ in der EU-ROP-Klassifizierung. Im Gegensatz dazu wiesen die Maststiere durchschnittlich die Fleischklasse „U“ und Fettklasse „3“ auf. Die ermittelten Daten aus der Trockenmasseanalyse, der Stickstoffbestimmung nach Kjeldahl und der Fettextraktion nach Soxhlet sind in Abbildung 1-3 dargestellt. Der Trockenmas-segehalt war im Fleisch der Almochsen signifikant niedriger als bei den Fleckviehstieren. Bei den Alm-ochsen wurde weiters um knapp 4 % weniger intramuskuläres Fett (TM) als bei den Maststieren vor-gefunden. Auf die Frischmasse bezogen ergibt das einen Rohfettgehalt von 1,7 % bei den Almochsen und 2,8 % bei den Maststieren (p<0,05). Ebenfalls gab es einen signifikanten Unterschied im Bezug auf den Proteingehalt in der Trockenmasse, der im Almochsenfleisch deutlich höher war. Auf die Frischmasse bezogen ist der Unterschied jedoch mit 22,1 % bei den Almochsen und 22,2 % bei den Maststieren vernachlässigbar gering. Ein signifikanter Unterschied bestand zwischen den langkettigen PUFAs (LC-PUFAs) (Abbildung 6) und zwischen den n3-FS (Abbildung 4). Im Fleisch des Almochsen waren diese relativ um 40,6 % bzw. um 111,6 % gegenüber den Maststieren erhöht. Der Unterschied des Verhältnisses zwischen der Summe der n6- gegenüber den n3-Fettsäuren ist ebenfalls signifikant. Es gibt keinen signifikanten Unterschied zwischen den Versuchsgruppen im Bezug auf den P:S-Fettsäurequotient (Abbildung 5). Abbildung 7 stellt das P:M:S-Fettsäureverhältnis der Versuchsgrup-pen dem Idealverhältnis gegenüber. Deutlich zu erkennen ist der hohe Gehalt an SFAs und der niedri-ge PUFA-Gehalt bei beiden Versuchsgruppen im Vergleich zum Idealverhältnis. Signifikant ist hier nur der um 2,4 % höhere Anteil an MUFAs bei den Maststieren.
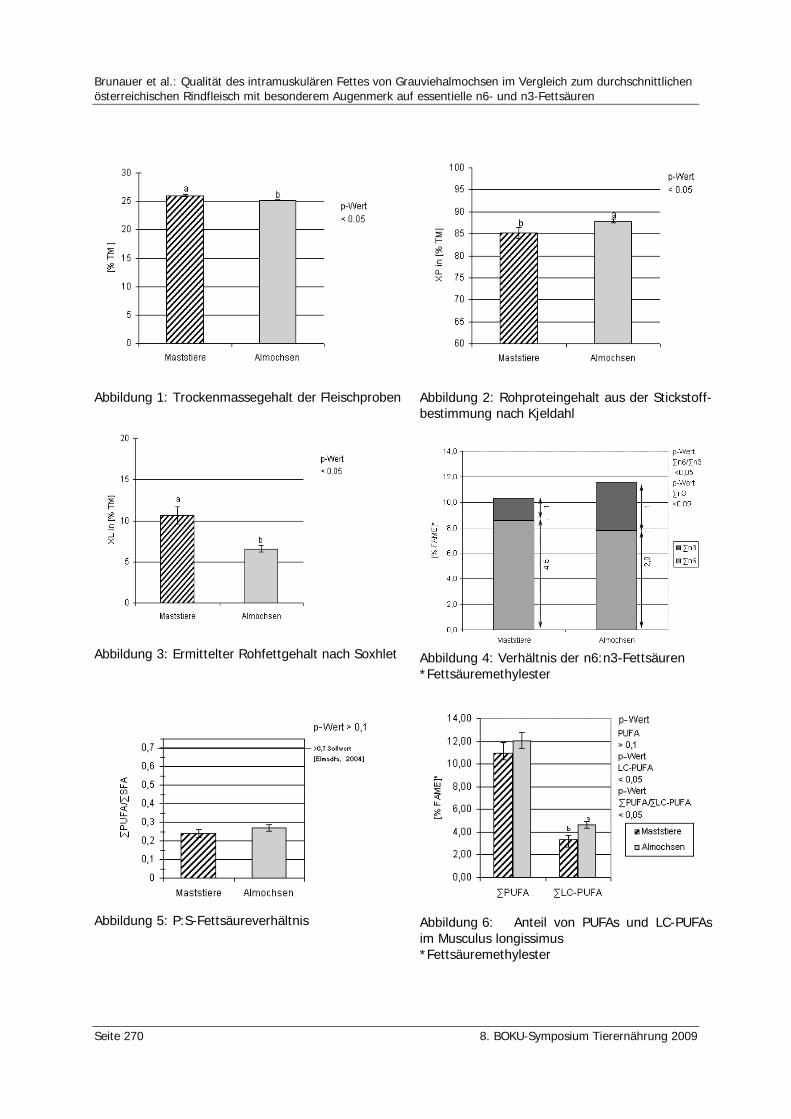
Brunauer et al.: Qualität des intramuskulären Fettes von Grauviehalmochsen im Vergleich zum durchschnittlichen österreichischen Rindfleisch mit besonderem Augenmerk auf essentielle n6- und n3-Fettsäuren
Seite 270 8. BOKU-Symposium Tierernährung 2009
Abbildung 1: Trockenmassegehalt der Fleischproben
Abbildung 2: Rohproteingehalt aus der Stickstoff-bestimmung nach Kjeldahl
Abbildung 3: Ermittelter Rohfettgehalt nach Soxhlet
Abbildung 4: Verhältnis der n6:n3-Fettsäuren *Fettsäuremethylester
Abbildung 5: P:S-Fettsäureverhältnis
Abbildung 6: Anteil von PUFAs und LC-PUFAs im Musculus longissimus *Fettsäuremethylester
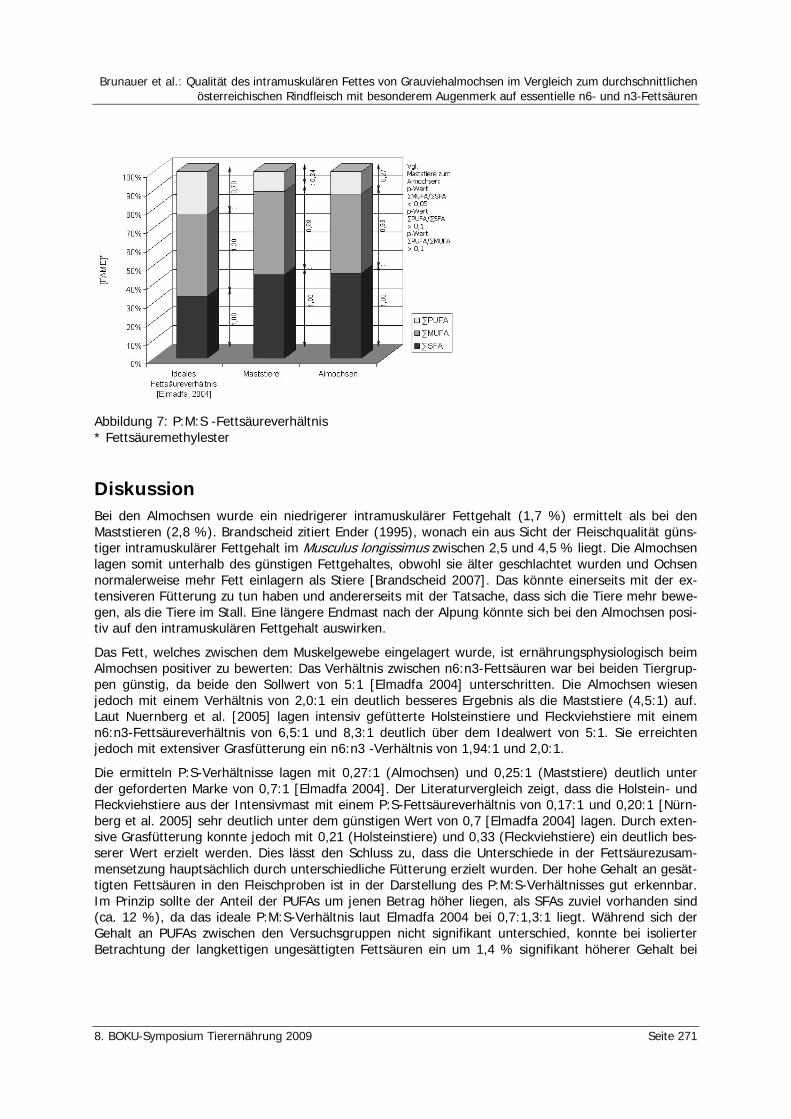
Brunauer et al.: Qualität des intramuskulären Fettes von Grauviehalmochsen im Vergleich zum durchschnittlichen österreichischen Rindfleisch mit besonderem Augenmerk auf essentielle n6- und n3-Fettsäuren
8. BOKU-Symposium Tierernährung 2009 Seite 271
Abbildung 7: P:M:S -Fettsäureverhältnis * Fettsäuremethylester
Diskussion Bei den Almochsen wurde ein niedrigerer intramuskulärer Fettgehalt (1,7 %) ermittelt als bei den Maststieren (2,8 %). Brandscheid zitiert Ender (1995), wonach ein aus Sicht der Fleischqualität güns-tiger intramuskulärer Fettgehalt im Musculus longissimus zwischen 2,5 und 4,5 % liegt. Die Almochsen lagen somit unterhalb des günstigen Fettgehaltes, obwohl sie älter geschlachtet wurden und Ochsen normalerweise mehr Fett einlagern als Stiere [Brandscheid 2007]. Das könnte einerseits mit der ex-tensiveren Fütterung zu tun haben und andererseits mit der Tatsache, dass sich die Tiere mehr bewe-gen, als die Tiere im Stall. Eine längere Endmast nach der Alpung könnte sich bei den Almochsen posi-tiv auf den intramuskulären Fettgehalt auswirken.
Das Fett, welches zwischen dem Muskelgewebe eingelagert wurde, ist ernährungsphysiologisch beim Almochsen positiver zu bewerten: Das Verhältnis zwischen n6:n3-Fettsäuren war bei beiden Tiergrup-pen günstig, da beide den Sollwert von 5:1 [Elmadfa 2004] unterschritten. Die Almochsen wiesen jedoch mit einem Verhältnis von 2,0:1 ein deutlich besseres Ergebnis als die Maststiere (4,5:1) auf. Laut Nuernberg et al. [2005] lagen intensiv gefütterte Holsteinstiere und Fleckviehstiere mit einem n6:n3-Fettsäureverhältnis von 6,5:1 und 8,3:1 deutlich über dem Idealwert von 5:1. Sie erreichten jedoch mit extensiver Grasfütterung ein n6:n3 -Verhältnis von 1,94:1 und 2,0:1.
Die ermitteln P:S-Verhältnisse lagen mit 0,27:1 (Almochsen) und 0,25:1 (Maststiere) deutlich unter der geforderten Marke von 0,7:1 [Elmadfa 2004]. Der Literaturvergleich zeigt, dass die Holstein- und Fleckviehstiere aus der Intensivmast mit einem P:S-Fettsäureverhältnis von 0,17:1 und 0,20:1 [Nürn-berg et al. 2005] sehr deutlich unter dem günstigen Wert von 0,7 [Elmadfa 2004] lagen. Durch exten-sive Grasfütterung konnte jedoch mit 0,21 (Holsteinstiere) und 0,33 (Fleckviehstiere) ein deutlich bes-serer Wert erzielt werden. Dies lässt den Schluss zu, dass die Unterschiede in der Fettsäurezusam-mensetzung hauptsächlich durch unterschiedliche Fütterung erzielt wurden. Der hohe Gehalt an gesät-tigten Fettsäuren in den Fleischproben ist in der Darstellung des P:M:S-Verhältnisses gut erkennbar. Im Prinzip sollte der Anteil der PUFAs um jenen Betrag höher liegen, als SFAs zuviel vorhanden sind (ca. 12 %), da das ideale P:M:S-Verhältnis laut Elmadfa 2004 bei 0,7:1,3:1 liegt. Während sich der Gehalt an PUFAs zwischen den Versuchsgruppen nicht signifikant unterschied, konnte bei isolierter Betrachtung der langkettigen ungesättigten Fettsäuren ein um 1,4 % signifikant höherer Gehalt bei

Brunauer et al.: Qualität des intramuskulären Fettes von Grauviehalmochsen im Vergleich zum durchschnittlichen österreichischen Rindfleisch mit besonderem Augenmerk auf essentielle n6- und n3-Fettsäuren
Seite 272 8. BOKU-Symposium Tierernährung 2009
den Almochsen detektiert werden, was wiederum aus ernährungsphysiologischer Sicht positiv zu be-werten ist.
In wie fern die im Fleisch enthaltene Menge an n3-Fettsäuren tatsächlich für die menschliche Ernäh-rung von Bedeutung ist, zeigt folgendes Rechenbeispiel: Der Energiebedarf eines durchschnittlichen Mannes (74 kg, 1,76 m) liegt bei etwa 12 MJ/Tag. Die n3-Fettsäuren sollten dabei 0,5 % der zuge-führten Energie darstellen (1,54 g/Tag) [DGE 2001]. Beim Verzehr eines 183 g Steaks/Tag (66,8 kg Fleisch/Jahr = durchschnittlicher Fleischkonsum laut Statistik Austria 2008) vom Musculus longissimus der Almochsen sind das mit 0,12 g n3-Fettsäuren 7,5 % und mit 0,09 g (Maststiere) 5,9 % des Ta-gesbedarfes. Der Beitrag der im Muskelfleisch enthaltenen essentiellen n3-FS für die menschliche Er-nährung ist somit relativ gering.
Schlussfolgerung Der Vergleich mit Literaturdaten lässt die Hypothese zu, dass die Fettsäurequalität des intramuskulä-ren Fettes maßgeblich durch die Fütterung beeinflusst wird und der Einfluss der Rasse und Kategorie etwas geringer ist. Eine extensive Weidefütterung im Vergleich zur Intensivmast kann sich aus ernäh-rungsphysiologischer Sicht positiv auf das eingelagerte Muskelfett auswirken. Dies wurde im Versuch mit Tiroler Grauviehalmochsen und Fleckviehmaststieren, welche für das durchschnittliche Österreichi-sche Rindfleischangebot stehen, bestätigt.
Literatur
W. Brandscheid, K.O. Hanikel, G. v. Lengerken, K.Troeger (Hrsg.): Qualität von Fleisch und Fleischwaren; 2. A. Band 1,157-205, 2007
L. Djoussé, J. S. Pankow, J. H. Eckfeldt, A. R. Folsom, P. N. Hopkins, M. A. Province, Y. Hong, and R C. Ellison: Relation be-tween dietary linolenic acid and coronary artery disease in the National Heart, Lung, and Blood Institute Family Heart Study; Am J Clin Nutr,74:612–9, 2001
D. Dannenberger, G. Nuernberg, N.l. Scollan, K. Ender, K. Nuernberg: Diet Alters teh Fatty Acid Composition of Individual Phos-pholipid Classes in Beef Muscle; J. Agric. Food Cehm. 55, 452-460, 2007
C. Naumann, R. Bassler: Die chemische Untersuchung von Futtermitteln; VDLUFA-Verlag, Darmstadt, Deutschland, 1997
P. S. Sukhija and D. L. Palmquist: Rapid method for Determination of Total Fatty Acid Content and Composition of Feedstuffs and Feces; J. Agric. Food Chem., 36, 1202-1206, 1988
I. Elmadfa: Ernährung des Menschen: Lipide und Begleitstoffe; Verlag Eugen Ulmer GmbH & Co, 93-103, 2004
K. Nuernberg, D. Dannenberger, G. Nuernberg, K. Ender, J. Voigt, N.D. Scollan, J.D. Wood, G.R. Nute, R.I. Richardson: Effect of a grass-based and a concentrate feeding system on meat quality characteristics and fatty acid composition of longissimus muscle in different cattle breeds; Livestock Production Science 94, 137-147, 2005
Statistik Austria 2008: Versorgungsbilanz für Fleisch nach Arten 2002 bis 2007 (Zugriff am 29.07.09) http://www.statistik.at/web_de/statistiken/land_und_forstwirtschaft/preise_bilanzen/versorgungsbilanzen/index.html,
Deutsche Gesellschaft für Ernährung (DGE): Referenzwerte für die Nährstoffzufuhr; 1. Aufl. -Frankfurt am Main, 2.korr. Nachdr., Verl. Umschau Braus, 23-53, 2001
Autorenanschrift
Christoph Brunauer Bakk.techn. Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Department für Lebensmittelwissenschaften und -technologie Universität für Bodenkultur Wien Gregor Mendel-Straße 33, A-1180 Wien E-Mail: [email protected]

Firmensponsoring – H. Wilhelm SCHAUMANN GmbH & CO. KG Jakob-Fuchs-Gasse 25-27, A-2345 Brunn am Gebirge
8. BOKU-Symposium Tierernährung 2009 Seite 273

Wetscherek-Seipelt et al.: Stichprobenuntersuchung von österreichischen Schweinefleischproben zum Fettgehalt und zur Fettzusammensetzung
Seite 274 8. BOKU-Symposium Tierernährung 2009
Stichprobenuntersuchung von österreichischen Schweine-fleischproben zum Fettgehalt und zur Fettzusammensetzung
Gabriela Wetscherek-Seipelt1, Rudolf Stückler2, Joachim Mandl2 und Wolfgang Wetscherek1 1 Universität für Bodenkultur Wien, Abteilung Tierische Lebensmittel, Tierernährung
und Ernährungsphysiologie 2 Agrarmarkt Austria Marketing GesmbH
Einleitung Die Anteile der Fettsäuren im tierischen Fett sind seit einiger Zeit Gegenstand intensiver Forschung und werden in zahlreichen Veröffentlichungen thematisiert. Dabei wurden die Einflüsse der Fettzu-sammensetzung auf die sensorische Merkmale der Fleischqualität, auf Gesundheitsaspekte, ernäh-rungsphysiologische Faktoren und Aspekte der Verarbeitungseignung untersucht. Es tritt immer wie-der ein Zielkonflikt zwischen den Beschaffenheitsanforderungen für die Dauerwarenherstellung und der diätischen Qualität von tierischen Lebensmitteln zu Tage.
Für die verarbeitende Industrie, die überwiegend Dauerwaren herstellt, ist die Fettqualität und Zu-sammensetzung ein bedeutender Qualitätsfaktor bei Schweinefleisch. Während in Österreich keine Vorgaben für den Landwirt mit finanziellen Zu- oder Abschlägen existieren, wurden in der Schweiz bereits seit längerer Zeit Grenzen für die Schweinefettqualität festgelegt, die Auswirkungen auf die Bezahlung der Schlachtkörper haben. Ein Polyenfettsäuregehalt von unter 12 Mol% wird als gut, ein Gehalt zwischen 12 und 13 Mol% als tolerierbar und höhere Anteile als unbefriedigend qualifiziert (Seewer, 1996). Während also nach den Schweizer Qualitätsanforderungen mehr als 13 Mol% Polyen-fettsäuren im Rückenfett für die Verarbeitung als nicht mehr geeignet betrachtet werden, sehen FI-SCHER et al. (1990) 15 Mol% Polyensäuren als obere Grenze für die Herstellung von ungegarten Fleischprodukten an.
In einer Vielzahl von Fütterungsversuchen wurde gezeigt, dass sich der Fettsäuregehalt des Futters bei landwirtschaftlichen Nutztieren auf die Zusammensetzung der endogenen Lipide verschiedener Organe und Gewebe auswirkt. Dies wurde insbesondere bei Monogastriern (Schwein, Geflügel), aber auch in geringerem Umfang bei Wiederkäuern (Rind und Schaf) nachgewiesen (Honikel et al., 2006). Beispiele dafür sind beispielsweise in Arbeiten von Fischer et al., 1992; Leibetseder, 1996; Reichart et al. 2003 oder Zettl 1995 zu finden.
Soll der Gehalt von 12 % Polyenfettsäuren im Karreeauflagenfett nicht überschritten werden, darf der Polyensäuregehalt im Mastfutter nur 10 - 16 g/kg betragen. Schweizer Autoren geben eine obere Grenze von 0,8 g Polyensäuren / MJ verdauliche Energie beim Schwein (ca. 12 g PUFA/kg Futter) für die Sicherung einer guten Qualität des Rückenspecks von Schweinen vor (Reichardt et al., 2002).
Material und Methoden Die Probenziehung wurde von der AMA bei drei verschiedenen Schlachthöfen bei der Schlachtung durchgeführt. Dabei wurden Proben vom Karreefleisch mit Auflagenfett genommen, Vakuum verpackt und tief gefroren. Um einen repräsentativen Querschnitt der österreichischen Schweine-fleischproduktion zu erhalten wurden die Proben an den Schlachthöfen Schreiner GesmbH Co KG mit

Wetscherek-Seipelt et al.: Stichprobenuntersuchung von österreichischen Schweinefleischproben zum Fettgehalt und zur Fettzusammensetzung
8. BOKU-Symposium Tierernährung 2009 Seite 275
dem Einzugsgebiet der Grenzregion Nieder- und Oberösterreich, Rudolf Großfurtner GmbH, von Land-wirten aus Oberösterreich und Schirnhofer Fleischwarenvertriebsgesellschaft mbH, welche von steiri-schen Betrieben beliefert wurde, gezogen.
Zur Probenvorbereitung wurden aus den gefrorenen Koteletts Fleischquadrate an einer definierten Stelle herausgeschnitten, und mit einem Fleischmixer homogenisiert. Nach erfolgter Einwaage für die Analysen der Rest der Probe wieder vakuumverpackt eingefroren. Vom inneren Auflagenfett wurden mit einem Ausstecher Proben zur Fettsäurenbestimmung gezogen.
Für die Trockensubstanzbestimmung wurden Porzellanschalen mit Seesand gefüllt, im Trockenschrank tariert, 5 g homogenisierte Probe eingewogen, im Sand zerrieben und im Trockenschrank bei 103oC 4 Stunden getrocknet. Die Porzellanschale mit den getrockneten Proben wurde im Exsikkator zum Auskühlen gebracht und rückgewogen.
Für die Rohfettbestimmung wurden die getrockneten Proben von der Trockensubstanzbestimmung in Extraktionshülsen quantitativ übergeführt und über 6 Stunden mit Diäthylether mit einer Soxlethappa-ratur extrahiert. Das so gewonnene Fett wurde in einem tarierten Stehrundkolben gesammelt und die Menge durch Differenzwägung bestimmt.
Für die Fettäurenanalyse wurde das Auflagenfett bzw. das durch Extraktion gewonnene Fett des Mus-kelfleisches in Röhrchen übergeführt, mit 0,8 ml Hexan und 0,5 ml methanolischer KOH versetzt, 1 Minute geschüttelt und 10 Minuten stehen gelassen, die oberste Schicht vorsichtig abgehoben und über 1 g Na2SO4 getrocknet, mit 1 ml Hexan versetzt, geschüttelt und die klare Lösung ab pipettiert im 1,5 ml Vials gefüllt und mit einem Agilent 7890 A GC-System mit Autosampler analysiert. Dabei wurde eine 60 m DB 23 Kapillarsäule, mit 35 ml/min Wasserstoff als Trägergas und einem Tempera-turprogramm von 50 bis 230° C verwendet.
Ergebnisse und Diskussion Wie in der Abbildung 1 dargestellt streute der Fettgehalt im Karreefleisch stark. Der Mittelwert lag bei 2,8 % mit einer Standardabweichung von 0,94. Bemerkenswert dabei ist, dass die meisten Proben einen Fettgehalt zwischen 2 und 4 % aufwiesen.
Die Fettzusammensetzung des Karreefleisches hatte nur etwa 40 % des Anteils an Polyenfettsäuren im Vergleich zum Karreeauflagenfett. Diese Verschiebung ging vor allem zu Lasten der Monoenfett-säuren. Die genaue Zusammensetzung des Fettes wird in den Tabellen 1 und 2 dargestellt. Die sehr große Streuung der Zusammensetzung des Fettes sowohl im Fleisch als auch im Auflagenfett lässt auf große Unterschiede in der Fütterung rückschließen. Zur Veranschaulichung dieser Streuung wird in der Abbildung 2 der Polyenfettsäurenanteil der Fleischproben und in der Abbildung 3 der Polyenfettsäu-renanteil des Auflagenfettes dargestellt. Von den 60 Auflagenfettproben entsprachen fast die Hälfte (28 Proben) den strengen Qualitätsanforderungen der Schweiz von maximal 12 Mol% Polyenfettsäu-renanteil. Etwa ¼ der Proben (13 Proben) lag über dem Wert von 15 Mol% Polyenfettsäuren (Richt-wert von Fischer et al., 1990), eine Fettzusammensetzung, welche die Eignung für Dauerwarenher-stellung nicht mehr sicher gewährleisten kann. Der Maximalwert der untersuchten Proben lag bei ei-nem Polyenfettsäurenanteil von 24,3 Mol%. Aus der Literatur ist bekannt, dass die Gehalte an Polyen-fettsäuren durch den Gehalt der Polyenfettsäuren im Futter bestimmt werden. Daher zeigt dieses Er-gebnis die breite Streuung der Rationszusammensetzung in den österreichischen Schweinemastbetrie-ben. Aus Sicht der fleischverarbeitenden Betriebe, die Dauerwaren herstellen, wäre es natürlich wün-schenswert, dass auch in Österreich der Polyenfettsäurenanteil mehr Beachtung findet.
Neben den Grenzwerten für den Gehalt von Polyensäuren wurden von Prabucki und Häusler (1990) noch weitere Qualitätskriterien, wie der Gehalt an 2, 3 bzw. 4-fach ungesättigten Fettsäuren, für den Karreespeck festgehalten. Wie in Tabelle 3 dargestellt, zeigen 70 % der untersuchten Proben Über-schreitungen bei den Diensäuren bzw. 45 % der Proben eine Überschreitung bei den Polyensäuren.

Wetscherek-Seipelt et al.: Stichprobenuntersuchung von österreichischen Schweinefleischproben zum Fettgehalt und zur Fettzusammensetzung
Seite 276 8. BOKU-Symposium Tierernährung 2009
Abbildung 1 Fettgehalt des Karreefleisches
0
1
2
3
4
5
6
7
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49 51 53 55 57 59
Proz
ent
Tabelle 1 Fettzusammensetzung des Karreefleisches
Fettsäure, relativ in Mol% des Fettes
Mittelwert Median Standardab-weichung
Minimum Maximum
C 10:0 0,1 0,1 0,04 <0,1 0,2 C 12:0 0,1 0,1 0,02 0,0 0,1 C 14:0 1,4 1,4 0,10 1,3 1,7 C 14:1 0,0 0,0 0,09 <0,1 0,6 C 16:0 25,6 25,7 0,79 23,9 27,3 C 16:1 3,6 3,6 0,51 2,5 4,9 C 18:0 13,9 13,9 1,54 10,9 19,7 C 18:1 49,2 49,7 2,21 43,9 53,1 C 18:2 4,4 4,3 1,15 2,5 7,4 C 18:3 0,2 0,2 0,10 <0,1 0,5 C 20:0 0,1 0,1 0,08 <0,1 0,3 C 20:1 0,8 0,8 0,14 0,4 1,4 C 20:2 0,2 0,2 0,09 <0,1 0,4 C 20:4 0,4 0,3 0,15 <0,1 0,8
Monoenfettsäuren 53,6 54,0 2,41 48,2 58,4 Polyenfettsäuren 5,1 5,0 1,18 3,1 8,4

Wetscherek-Seipelt et al.: Stichprobenuntersuchung von österreichischen Schweinefleischproben zum Fettgehalt und zur Fettzusammensetzung
8. BOKU-Symposium Tierernährung 2009 Seite 277
Abbildung 2 Anteil der Polyensäuren im Fett des Karreefleisches
0
1
2
3
4
5
6
7
8
9
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49 51 53 55 57 59
Proz
ent
Tabelle 2 Fettzusammensetzung des inneren Karreeauflagenfettes
Fettsäure, relativ in Mol% des Fettes
Mittelwert Median Standardab-weichung
Minimum Maximum
C 10:0 0,1 0,1 0,02 < 0,1 0,1 C 12:0 0,1 0,1 0,03 < 0,1 0,1 C 14:0 1,3 1,3 0,15 1,0 1,6 C 14:1 0,0 0,0 0,02 < 0,1 0,1 C 16:0 25,7 25,7 1,42 22,0 28,0 C 16:1 2,1 2,1 0,40 1,3 3,1 C 18:0 15,7 15,7 1,59 11,7 18,6 C 18:1 41,3 41,4 2,30 36,9 46,4 C 18:2 11,6 11,1 3,03 6,2 22,0 C 18:3 0,6 0,6 0,20 0,2 1,3 C 20:0 0,1 0,0 0,09 < 0,1 0,3 C 20:1 0,8 0,8 0,21 0,3 1,5 C 20:2 0,5 0,5 0,17 < 0,1 1,0 C 20:4 0,2 0,2 0,07 < 0,1 0,4
Monoenfettsäuren 44,2 44,3 2,52 39,5 49,6 Polyenfettsäuren 12,9 12,2 3,33 7,2 24,3

Wetscherek-Seipelt et al.: Stichprobenuntersuchung von österreichischen Schweinefleischproben zum Fettgehalt und zur Fettzusammensetzung
Seite 278 8. BOKU-Symposium Tierernährung 2009
Abbildung 3 Anteil der Polyensäuren im inneren Karreeauflagenfett
0
5
10
15
20
25
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49 51 53 55 57 59
Proz
ent
Tabelle 3 Anteil der untersuchten Proben, die nicht den Schweizer Qualitätsanforderungen
für Karreespeck entsprechen
Merkmal
Schweizer Qualitätsanforderungen
gut
tolerierbar
unbefriedigend Anteil der Proben mit unbefriedigenden Er-
gebnis ungesättigte Fett-
säuren < 59 % 59 – 61 % > 61 % 12 %
Polyensäuren < 12 % 12 – 13 % > 13 % 45 % Diensäuren < 10 % 10 % > 10 % 70 % Triensäuren < 1 % 1 % > 1 % 3 %
Tetraenfettsäuren < 0,5 % 0,5 % > 0,5 % 0 %
Zusammenfassung • Das Ergebnis der Stichprobenuntersuchung zeigt eine sehr große Varianz in den Ergebnissen
des Fettgehaltes im Karreefleisch aber auch in der Fettzusammensetzung im Fleisch und im inneren Auflagenfett.
• Aus zahlreichen Fütterungsversuchen liegt der Schluss nahe, dass eine unterschiedliche Fütte-rung der Tiere dafür verantwortlich ist. Vor allem die Zufuhr an Polyenfettsäuren durch direkte Öleinmischungen oder indirekten Fettgehalt in verschieden Futtermitteln kann als Hauptursa-

Wetscherek-Seipelt et al.: Stichprobenuntersuchung von österreichischen Schweinefleischproben zum Fettgehalt und zur Fettzusammensetzung
8. BOKU-Symposium Tierernährung 2009 Seite 279
che angesehen werden. Die derzeit weit verbreitete Rationsgestaltung mit Mais als energierei-che Hauptgetreidekomponente (durch ihren Ölgehalt) und eventueller Kombination mit Ölku-chen oder Expellern führt zwangsläufig zu einer sehr hohen Aufnahme an Polyenfettsäuren.
• Einerseits ist der hohe Anteil an Proben, die den Grenzwert an Polyensäuren bzw. Diensäuren überschreiten, nicht zufriedenstellend für die verarbeitungstechnische Eignung des österrei-chischen Schweinefleisches. Andererseits zeigen die Proben, welche die Anforderungen erfül-len, dass eine Erfüllung dieser Qualitätsanforderungen bei richtigem Management und ange-passter Fütterung auch in Österreich durchaus möglich ist.
• Nicht unerwähnt soll bleiben, dass aus ernährungsphysiologischer Sicht ein hoher Gehalt an ungesättigten Fettsäuren durchaus wünschenswert ist. Somit sind die Proben mit geringen Gehalten an gesättigten Fettsäuren gerade für die Frischfleischverwendung und nicht lange zu lagernden Verarbeitungsprodukten sehr gut geeignet und aus gesundheitlicher Sicht positiv zu bewerten.
• Zur endgültigen Abschätzung der Einsatzgrenzen von Futtermittel mit hohen Polyensäuren sollten Versuche mit dem heutigen genetischen Material durchgeführt werden.
Literatur
Fischer, K.; P. Freudenreich, K.-H Hoppenbrock,.; W. Sommer, 1990: Einflüsse von Fütterung, Mastendgewicht und Geschlecht auf die Fettqualität beim Schwein. Mitteilungsblatt der BASF 108, 130-139.
Fischer, K.; P. Freudenreich, K.-H Hoppenbrock,.; W. Sommer, 1992: Einfluss produktionstechnischer Bedingungen auf das Fettsäurenmuster im Rückenspeck von Mastschweinen. Fleischwirtschaft 72, 200-205
Honikel, K.-O., H. Rosenbauer und D. Kühne. 2002: Einfluss der Fütterung auf die Qualität von Schweinefleisch und daraus hergestellter Schweinefleischerzeugnissen.- Schriftenreihe des Bundesministeriums für Ernährung, Landwirtschaft und Forsten. Reihe A: Angewandte Wissenschaft. Heft 495, 167-182
Leibetseder, J. 1996: Beeinflussung der Zusammensetzung tierischer Fette durch die Fütterung. Arch. Tierz., Dummerstorf 39, 333-345
Prabucki, A. L. und A. Häuser, 1990: Die Fettqualität bei Mastschweinen (1988/1989). Landwirtschaft Schweiz 3, 199-200.
Reichardt, W.; E. Gernand, S. Müller, H. Hartung, B. Eckert, U. Braun, 2003: Erhebungen zur Fettsäurenzusammensetzung von Rückenfett bei Thüringer Schweinen sowie zum Fett von Thüringer Knackwürsten aus dem Einzelhandel. Arch. Tierz., Dummerstorf 46, 257-267
Reichard, W.; S. Müller; H. Hartung, B. Eckert und U. Braun (2002): Erhebungen zur Fettsäurenzusammensetzung sowie zur Iodzahl von intramuskulärem Fett und Rückenspeck bei Thüringer Schweinen. Kolloquium zur Fleischqualität. Schriftreihe der TTL 2-2002, 15-23.
Seewer, G.J.F. 1996: Qualität des Fettgwebes beim Schwein und beim Mastrind. Agrarforschung 3, 435-438, 1
Zettl, A. 1995: Einsatz von vollfetten Sojabohnen in der Schweinemast. Dissertation, Universität für Bodenkultur Wien.
Autorenanschrift
Ao. Univ.-Prof. Dr. Wolfgang Wetscherek Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Department für Lebensmittelwissenschaften und –technologie Universität für Bodenkultur Wien Gregor Mendel-Straße 33, 1180 Wien E-Mail: [email protected]

Firmensponsoring – Agrarmarkt Austria Marketing GesmbH, Dresdner Straße 68a, A-1200 Wien
Seite 280 8. BOKU-Symposium Tierernährung 2009

Wetscherek et al.: Nährstoffgehalt verschiedener getrockneter Topinamburkrautsorten
8. BOKU-Symposium Tierernährung 2009 Seite 281
Nährstoffgehalt verschiedener getrockneter Topinamburkraut-sorten
Wolfgang Wetscherek1, Laura Kocsis2, Peter Liebhard3 1 Universität für Bodenkultur Wien, Abteilung Tierische Lebensmittel, Tierernährung
und Ernährungsphysiologie 2 Brenntage Cee GmbH, Bahnstraße 13, A-2353 Guntramsdorf 3 Universität für Bodenkultur Wien, Department für Pflanzenwissenschaften und
Pflanzenbiotechnologie
Einleitung Das relativ schnelle Krautwachstum und die zum Teil hohen Erträge des oberirdischen Aufwuchses führen dazu, dass das Kraut des Topinamburs als frisches Grünfutter für Nutztiere ins Gespräch ge-kommen ist. Es kann auch zur Silagegewinnung und als Wildfutter in Form von Wildäckern, Schwei-neweide eingesetzt werden. In diesem Versuch wurde die Nährstoffzusammensetzung von 6 verschie-denen Sorten getrocknetem Topinamburkraut untersucht.
Bereits Pätzold (1957) hat festgestellt, dass für überwiegende Krautnutzung des Topinamburs statt der Knollen andere Produktionsmaßnahmen notwendig waren. Die Qualität für die Grünfütterung ist demnach vom Blatt-Stängel-Verhältnis abhängig. Der Gehalt an wertbestimmenden Inhaltsstoffen ist daher neben der Sortenwahl vom Erntetermin abhängig. Der Gesamtpflanzenertrag von Topinambur ist sortenspezifisch unterschiedlich hoch und erreicht zwischen Mitte September und Mitte November das Maximum (Liebhard 1991, Kocsis et al. 2007).
Material und Methoden Mehrjährige Feldversuche wurden in den Jahren von 2000 bis 2002 und 2007 an der Außenstelle der Universität für Bodenkultur Wien, an der Versuchswirtschaft in Groß Enzersdorf durchgeführt. In die-sem Versuch wurden 7 verschiedene Topinambursorten in einer zweifaktorielle Blockanlage mit neun Ernteterminen in 2-facher Wiederholung getestet. Der Erntetermin Oktober 2007 wurde für ergän-zende Futteranalysen herangezogen. Das Kraut wurde bei 105oC getrocknet, gemahlen und pelletiert.
Für die enzymatische Zuckerbestimmung wurden 200 mg gefriergetrocknete Probe wurden mit UHQ Wasser bei 80 °C 2 Stunden extrahiert, danach wurden die Proben zentrifugiert und je nach Bedarf verdünnt. Aus dieser Lösung wurden die Glukose, die Fruktose und die Saccharose enzymatisch mit-tels der Glukose/Fruktose/Saccharose –UV- Testkombination von Boehringer –Mannheim (Nr.716 260) bestimmt.
Für die enzymatische Inulinbestimmung wurden Glukose und Fruktose nach Säurehydrolyse der Fruk-tane mit der Glukose/Fruktose UV – Testkombination von Boehringer – Mannheim (Nr.716 269) nach Abzug der freien Glukose, Fruktose und Saccharose als Inulin berechnet.
WEENDER Analyse, ADF- und NDF-Bestimmung wurden nach der Standardmethode VDLUFA (Nau-mann und Bassler 1997) bestimmt.

Wetscherek et al.: Nährstoffgehalt verschiedener getrockneter Topinamburkrautsorten
Seite 282 8. BOKU-Symposium Tierernährung 2009
Ergebnisse und Diskussion Vor der maschinellen Ernte der Topinamburknollen muss die Zerkleinerung, Ernte oder Entfernung des Krautes erfolgen. Je nach Sorte und Erntezeitpunkt ist der Trockensubstanz unterschiedlich hoch Auch der Rohfasergehalt schwankt durch unterschiedlich fortgeschrittene Verholzung der Stängel stark. Das Kraut nimmt bei der Herbsternte einen Anteil von 25-56% an der Gesamtbiomasse ein und erreicht je nach Sorte und Standort Hektarerträge zwischen 5000 kg in semiariden bis 12000 kg Trockenmasse in semihumiden Produktionsgebieten (Liebhard 1991).
Aufgrund der Witterung kam es in den verschiedenen Erntejahren zu unterschiedlichen Wachstums-verläufen, die sich auch im Knollenertrag zeigten. Jedoch war ersichtlich, das späte Sorten wie Fuseau und Waldspindel schon im Jugendstadium, bereits ab ca. 20 Tagen nach Feldaufgang, ein deutlich stärkeres Krautwachstum im Vergleich zu den frühen Sorten Bianka und Topstar aufwiesen (Wuchs-höhenunterschied bis zu 15 cm, der sich bis Vegetationsende bis zu 1,0 m erhöhte).
Trotz der höheren mittleren Stängelzahl je Pflanze der frühen Sorten Bianka und Topstar (4 bis 8) gegenüber den mittelspäten und späten Sorten (mittlere Stängelzahl/Pflanze 2 bis 3) unterscheiden sich die Krauterträge nicht deutlich (Abb.1) (Kocsis et al., 2009).
Abbildung 1
Einfluss der Sorte, der Jahreswitterung und des Erntetermins auf den Krautertrag (Stän-gel und Blätter) in kg TM/ha,
Ergebnisse aus den Vegetationsjahren 2000, 2001 und 2002
0100020003000400050006000700080009000
10000
2000 2001 2002 2000 2001 2002 2000 2001 2002 2000 2001 2002 2000 2001 2002 2000 2001 2002 2000 2001 2002
Bianka Topstar Gigant Violet Rozo Waldsp. Fuseau 60
Sorte & Jahr
kg T
M h
a-1
Termin 1 Termin 2 Termin 3 Termin 4 Termin 5 Termin 6
Die Ursache für das stärkere späte Krautwachstum der späten Sorten ist durch den späteren Zeitpunkt des Knollenansatzes und die dadurch vorgegebene spätere Knollenbildung bedingt. Die Photo-syntheseprodukte werden daher in das Krautwachstum, d.h. in die Stängelmassebildung verlagert.
Ein wesentlich beeinflussender Parameter für den Krautertrag ist die Wuchshöhe. Sie wurde durch die Sorte, den unterschiedlichen Temperaturverlauf und vor allem durch verschieden hohes Wasserange-bot signifikant beeinflusst. Die mittelspäten und späten Sorten erreichten erwartungsgemäß im Ver-gleich zu den frühen Sorten eine im Trend höhere Wuchshöhe (Abb. 2) Kocsis et al., 2009).
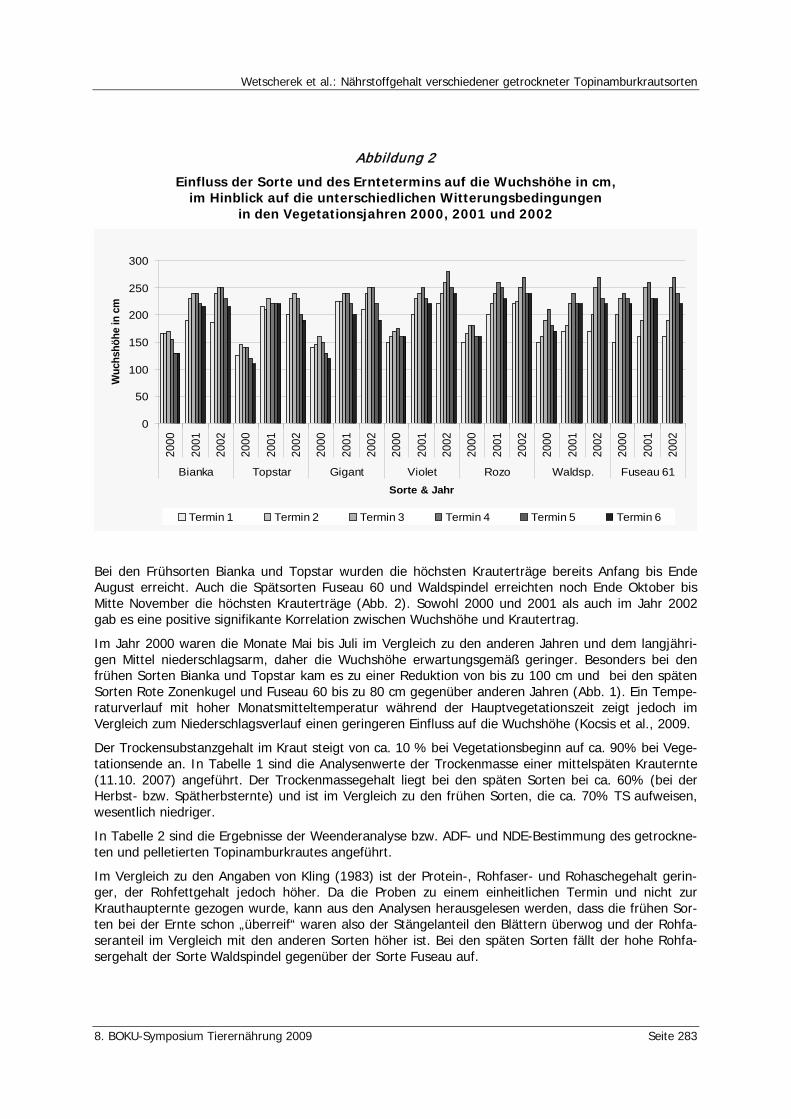
Wetscherek et al.: Nährstoffgehalt verschiedener getrockneter Topinamburkrautsorten
8. BOKU-Symposium Tierernährung 2009 Seite 283
Abbildung 2
Einfluss der Sorte und des Erntetermins auf die Wuchshöhe in cm, im Hinblick auf die unterschiedlichen Witterungsbedingungen
in den Vegetationsjahren 2000, 2001 und 2002
0
50
100
150
200
250
300
2000
2001
2002
2000
2001
2002
2000
2001
2002
2000
2001
2002
2000
2001
2002
2000
2001
2002
2000
2001
2002
Bianka Topstar Gigant Violet Rozo Waldsp. Fuseau 61Sorte & Jahr
Wuc
hshö
he in
cm
Termin 1 Termin 2 Termin 3 Termin 4 Termin 5 Termin 6
Bei den Frühsorten Bianka und Topstar wurden die höchsten Krauterträge bereits Anfang bis Ende August erreicht. Auch die Spätsorten Fuseau 60 und Waldspindel erreichten noch Ende Oktober bis Mitte November die höchsten Krauterträge (Abb. 2). Sowohl 2000 und 2001 als auch im Jahr 2002 gab es eine positive signifikante Korrelation zwischen Wuchshöhe und Krautertrag.
Im Jahr 2000 waren die Monate Mai bis Juli im Vergleich zu den anderen Jahren und dem langjähri-gen Mittel niederschlagsarm, daher die Wuchshöhe erwartungsgemäß geringer. Besonders bei den frühen Sorten Bianka und Topstar kam es zu einer Reduktion von bis zu 100 cm und bei den späten Sorten Rote Zonenkugel und Fuseau 60 bis zu 80 cm gegenüber anderen Jahren (Abb. 1). Ein Tempe-raturverlauf mit hoher Monatsmitteltemperatur während der Hauptvegetationszeit zeigt jedoch im Vergleich zum Niederschlagsverlauf einen geringeren Einfluss auf die Wuchshöhe (Kocsis et al., 2009.
Der Trockensubstanzgehalt im Kraut steigt von ca. 10 % bei Vegetationsbeginn auf ca. 90% bei Vege-tationsende an. In Tabelle 1 sind die Analysenwerte der Trockenmasse einer mittelspäten Krauternte (11.10. 2007) angeführt. Der Trockenmassegehalt liegt bei den späten Sorten bei ca. 60% (bei der Herbst- bzw. Spätherbsternte) und ist im Vergleich zu den frühen Sorten, die ca. 70% TS aufweisen, wesentlich niedriger.
In Tabelle 2 sind die Ergebnisse der Weenderanalyse bzw. ADF- und NDE-Bestimmung des getrockne-ten und pelletierten Topinamburkrautes angeführt.
Im Vergleich zu den Angaben von Kling (1983) ist der Protein-, Rohfaser- und Rohaschegehalt gerin-ger, der Rohfettgehalt jedoch höher. Da die Proben zu einem einheitlichen Termin und nicht zur Krauthaupternte gezogen wurde, kann aus den Analysen herausgelesen werden, dass die frühen Sor-ten bei der Ernte schon „überreif“ waren also der Stängelanteil den Blättern überwog und der Rohfa-seranteil im Vergleich mit den anderen Sorten höher ist. Bei den späten Sorten fällt der hohe Rohfa-sergehalt der Sorte Waldspindel gegenüber der Sorte Fuseau auf.
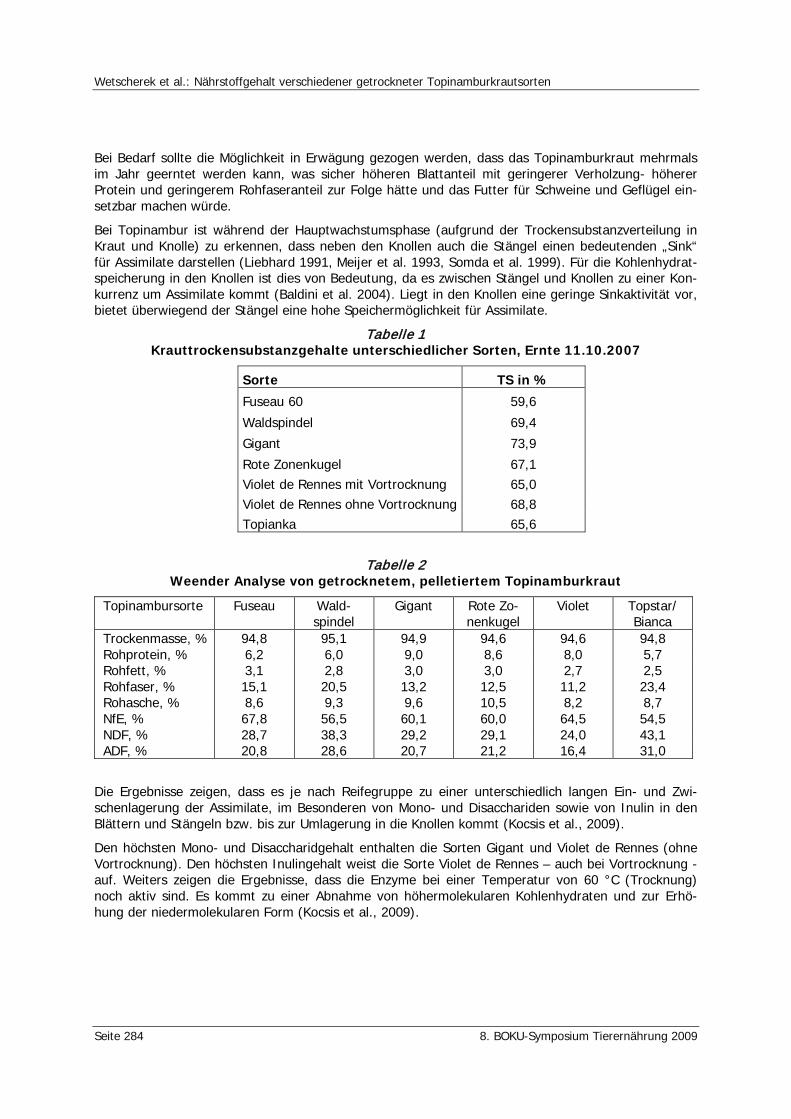
Wetscherek et al.: Nährstoffgehalt verschiedener getrockneter Topinamburkrautsorten
Seite 284 8. BOKU-Symposium Tierernährung 2009
Bei Bedarf sollte die Möglichkeit in Erwägung gezogen werden, dass das Topinamburkraut mehrmals im Jahr geerntet werden kann, was sicher höheren Blattanteil mit geringerer Verholzung- höherer Protein und geringerem Rohfaseranteil zur Folge hätte und das Futter für Schweine und Geflügel ein-setzbar machen würde.
Bei Topinambur ist während der Hauptwachstumsphase (aufgrund der Trockensubstanzverteilung in Kraut und Knolle) zu erkennen, dass neben den Knollen auch die Stängel einen bedeutenden „Sink“ für Assimilate darstellen (Liebhard 1991, Meijer et al. 1993, Somda et al. 1999). Für die Kohlenhydrat-speicherung in den Knollen ist dies von Bedeutung, da es zwischen Stängel und Knollen zu einer Kon-kurrenz um Assimilate kommt (Baldini et al. 2004). Liegt in den Knollen eine geringe Sinkaktivität vor, bietet überwiegend der Stängel eine hohe Speichermöglichkeit für Assimilate.
Tabelle 1 Krauttrockensubstanzgehalte unterschiedlicher Sorten, Ernte 11.10.2007
Sorte TS in %
Fuseau 60 59,6
Waldspindel 69,4
Gigant 73,9
Rote Zonenkugel 67,1 Violet de Rennes mit Vortrocknung 65,0 Violet de Rennes ohne Vortrocknung 68,8 Topianka 65,6
Tabelle 2 Weender Analyse von getrocknetem, pelletiertem Topinamburkraut
Topinambursorte Fuseau Wald-spindel
Gigant Rote Zo-nenkugel
Violet Topstar/ Bianca
Trockenmasse, % 94,8 95,1 94,9 94,6 94,6 94,8 Rohprotein, % 6,2 6,0 9,0 8,6 8,0 5,7 Rohfett, % 3,1 2,8 3,0 3,0 2,7 2,5 Rohfaser, % 15,1 20,5 13,2 12,5 11,2 23,4 Rohasche, % 8,6 9,3 9,6 10,5 8,2 8,7 NfE, % 67,8 56,5 60,1 60,0 64,5 54,5 NDF, % 28,7 38,3 29,2 29,1 24,0 43,1 ADF, % 20,8 28,6 20,7 21,2 16,4 31,0
Die Ergebnisse zeigen, dass es je nach Reifegruppe zu einer unterschiedlich langen Ein- und Zwi-schenlagerung der Assimilate, im Besonderen von Mono- und Disacchariden sowie von Inulin in den Blättern und Stängeln bzw. bis zur Umlagerung in die Knollen kommt (Kocsis et al., 2009).
Den höchsten Mono- und Disaccharidgehalt enthalten die Sorten Gigant und Violet de Rennes (ohne Vortrocknung). Den höchsten Inulingehalt weist die Sorte Violet de Rennes – auch bei Vortrocknung - auf. Weiters zeigen die Ergebnisse, dass die Enzyme bei einer Temperatur von 60 °C (Trocknung) noch aktiv sind. Es kommt zu einer Abnahme von höhermolekularen Kohlenhydraten und zur Erhö-hung der niedermolekularen Form (Kocsis et al., 2009).

Wetscherek et al.: Nährstoffgehalt verschiedener getrockneter Topinamburkrautsorten
8. BOKU-Symposium Tierernährung 2009 Seite 285
Abbildung 3 Einfluss der Sorte auf den Glukose-, Fruktose- und Saccharosegehalt
in g/100 g TM im Kraut, Ernte 11.10. 2007
Abbildung 4 Einfluss der Sorte auf den Inulingehalt und Polymerisationsgrad
in g/100 g TM im Kraut, Ernte 11.10. 2007
0
1
2
3
4
5
6
Fuseau Waldspindel Gigant RoteZonenkugel
Violet deRennes withpre-drying
Violet deRennes
without pre-drying
Topianka
g/10
0 g
DM
glucose fructose sucrose
0
5
10
15
20
25
30
35
40
45
Fuseau Waldspindel Gigant RoteZonenkugel
Violet deRennes withpre-drying
Violet deRennes
without pre-drying
Topianka
g/1
00 g
DM
fructan DP

Wetscherek et al.: Nährstoffgehalt verschiedener getrockneter Topinamburkrautsorten
Seite 286 8. BOKU-Symposium Tierernährung 2009
Zusammenfassung • Die Futterzusammensetzung zeigt ein Potential des getrockneten Topinamburkrautes als Roh-
faserträger in Futtermischungen beispielsweise für tragende Zuchtsauen auf.
• Eine andere Einsatzrichtung wäre die Nutzung des Gehaltes an präbiotischen Inhaltsstoffen z.B. Inulin.
• Zur besseren Abschätzung des Futterwertes sind noch weitere gezielte Untersuchungen über den Sorteneinfluss und den Einfluss des optimalen Schnittzeitpunktes notwendig. Eventuell wäre auch eine Mehrschnittnutzung aus der Sicht des Futterwertes interessant.
• Zur endgültigen Abschätzung der Eignung des Futtermittels müssten auch noch Fütterungs-versuche durchgeführt werden.
Literatur
Baldini, M., F. Danuso, M. Turi and P. Vanozzi, 2004: Evaluation of new clones of Jerusalem artichoke (Helianthus tuberosus L.) for inulin and sugar yield from stalks and tubers. Industrial Crops and Products 19, 25 – 40.
Kling, Max, 1983, Handelsfuttermittel Band 2A.
Kocsis, Laura, H. –P. Kaul, W. Praznik und P. Liebhard, 2007: Einfluss des Erntetermins auf den Kraut und Knollenertrag unter-schiedlicher Sorten von Topinambur (Helianthus tuberosus L.) im semiariden Produktionsgebiet Österreichs. Pflanzenbauwissenschaften, 11 (2), 67-76.
Kocsis; Laura, P. Liebhard, C. Zeitlhofer und W. Wetscherek, 2009: Einfluss der Sorte und des Erntezeitpunktes auf den Topi-namburkrautertrag sowie auf ernährungsphysiologische Aspekte in der Schweinefütterung, LTZ Augustenburg (Hrsg.), Topi-nambur- eine Pflanze mit vielen Verwertungsmöglichkeiten, 9.1- 9.8
Liebhard, P., 1991: Kulturartenvergleich von Nutzpflanzen als Rohstoffe für die industrielle Verarbeitung und zur Alternativen Energieversorgung. Forschungsbericht. Im Auftrag der Arbeitsgemeinschaft „Erneuerbare Energien“. Universität für Bodenkultur Wien.
Meijer, W.J.M., E.W.J.M. Mathijssen and G.E.L. Borm, 1993: Crop characteristics and inulin production of Jerusalem artichoke and cichory: A. Fuchs, S. Schittenhelm and L. Frese (Eds.): Proceedings of the sixth seminar on Inulin and Inulin containing Crops, Studies in Plant science, 3. Elsevier, 29-38.
Naumann, C, und Bassler, R., 1997 : Die chemische Untersuchung von Futtermitteln. In: Methodenbuch, Band III. 3. Ausgabe, 4. Ergänzungslieferung, VDLUFA – Verlag Darmstadt.
Pätzold, Chr. (1957): Die Topinambur als landwirtschaftliche Kulturpflanze. BM. Für Landwirtschaft und Forsten in Zusammenarbeit mit dem land- und hauswirtschaftlichen Auswertungs- und Informationsdienst. E. V. (AID) hekt.
Somda Z.C., W. J. Mc Laurin and S.J. Kays, 1999: Jerusalem Artichoke Growth, Development and Field Storage. II. Carbon and Nutrient Element Allocation and Redistribution. Journal of Plant Nutrition 22, 1315-1334.
Autorenanschrift
Ao. Univ.-Prof. Dr. Wolfgang Wetscherek Abteilung Tierische Lebensmittel, Tierernährung und Ernährungsphysiologie Department für Lebensmittelwissenschaften und –technologie Universität für Bodenkultur Wien Gregor Mendel-Straße 33, 1180 Wien E-Mail: [email protected]

Mayrhofer et al.: Evaluierung von Laktobazillen-Stämmen bezüglich erworbener Antibiotikaresistenzgene
8. BOKU-Symposium Tierernährung 2009 Seite 287
Evaluierung von Laktobazillen-Stämmen bezüglich erworbener Antibiotikaresistenzgene
Sigrid Mayrhofer1, Konrad J. Domig1, Angela H.A.M. van Hoek2, Sebastian Aigner1, Wolfgang Kneifel3 1 BOKU, Universität für Bodenkultur Wien, Department für Lebensmittel-
wissenschaften und –technologie, Abteilung Lebensmittelmikrobiologie und –hygiene, Wien
2 RIKILT, Institut of Food Safety, Department of Microbiology and Novel Foods, Wageningen
3 BOKU, Universität für Bodenkultur Wien, Department für Lebensmittel-wissenschaften und –technologie, Abteilung Lebensmittelqualitätssicherung, Wien
Einleitung Laktobazillen sind in der Natur weit verbreitet. Dabei sind sie vor allem in tierischen und pflanzlichen Produkten sowie im Gastrointestinal- und Vaginaltrakt von Säugern zu finden (Holt et al., 1994). Auf-grund ihrer, in erster Linie auf einer Ansäuerung beruhenden, konservierenden Wirkung haben Lakto-bazillen in der Landwirtschaft große wirtschaftliche Bedeutung erlangt. Hierbei werden sie vorwiegend als Starterkulturen für die Haltbarmachung von Viehfutter eingesetzt. Seit den siebziger Jahren des vorigen Jahrtausends werden sie zusätzlich als Probiotika im Tierfutter verwendet, um die Darmmicro-biota der Wirtstiere im positiven Sinne zu beeinflussen. Dabei sind sie auch als Ersatz für nutritive Antibiotika gedacht (Reuter, 2001).
Bei der Produktion von Futtermitteln werden Laktobazillen in hohen Zahlen zugefügt (Kastner et al., 2006). Um das bereits bestehende Resistenzgenreservoir in der Darmmikrobiotia des Wirtstiers und schlussendlich des Menschen nicht weiter zu erhöhen, setzt das Gremium für Zusatzstoffe, Erzeugnis-se und Substanzen in der Tierernährung (FEEDAP) der europäischen Behörde für Lebensmittelsicher-heit (EFSA) das Fehlen erworbener Antibiotikaresistenzgene für die Zulassung tierischer Probiotika und Starterkulturen voraus. Dabei wird in einer Stellungnahme des Gremiums detailiert auf die Überprü-fung des Vorhandenseins von erworbenen Antibiotikaresistenzgenen eingegangen (FEEDAP, 2008).
Zunächst sollte dabei die Identität des zu testenden Stammes mit Hilfe von molekularbiologischen Methoden überprüft werden. Hierbei ist auch der zusätzliche Einsatz von phänotypischen und chemo-taxonomischen Methoden („polyphasic approach“) empfehlenswert, um eine eindeutige Identifizierung ermöglichen zu können. Anschließend wird phänotypisch auf das Vorhandensein von Resistenzen mit-tels Dilutionsmethoden getestet. Die Detektion einer atypischen, nicht natürlich vorkommenden Resis-tenz erfordert weitere Analysen. Zur genaueren Charakterisierung der Resistenz werden üblicherweise auf PCR basierende Methoden eingesetzt. In letzter Zeit werden auch verstärkt Mikroarrays verwen-det, da man mit diesen auf viele Resistenzgene gleichzeitig untersuchen kann.
Anhand dieser Studie wird demonstriert, wie man bei der Überprüfung des Vorhandenseins von er-worbenen Antibiotikaresistenzgenen nach den Richtlinien des FEEDAP Gremiums vorgehen soll. Zu-sätzlich wurde die Lokalisation vorkommender Resistenzgene genauer bestimmt, da das Vorhanden-sein dieser auf übertragbaren Elementen (z.B.: Plasmid, Transposon) das höchste Risiko für einen Gentransfer darstellt.

Mayrhofer et al.: Evaluierung von Laktobazillen-Stämmen bezüglich erworbener Antibiotikaresistenzgene
Seite 288 8. BOKU-Symposium Tierernährung 2009
Material und Methoden Insgesamt standen 101 Laktobazillen-Isolate für die Untersuchung zur Verfügung, die zuvor mittels PCR oder ARDRA (Amplified Ribosomal DNA Restriction Analysis) identifiziert wurden (Danielsen et al., 2008). Dabei waren 53 Isolate tierischer Herkunft (2 L. acidophilus, 25 L. amylovorus, 3 L. crispatus, 7 L. gallinarum, 16 L. johnsonii) und 40 Isolate menschlicher Herkunft (7 L. acidophilus, 2 L. amylovo-rus, 4 L. crispatus, 25 L. gasseri, 2 L. johnsonii). Zusätzlich stammten vier Isolate aus Lebensmitteln (1 L. acidophilus, 1 L. amylovorus, 1 L. gasseri, 1 L. johnsonii), drei aus Futtermitteln (3 L. amylovo-rus) und eines aus einem pharmazeutischen Produkt (1 L. johnsonii).
Das Resistenzverhalten der Isolate wurde mittels Mikrodilutionsverfahren gegenüber Chloramphenicol überprüft. Dabei wurde zunächst eine auf eine bestimmte Dichte eingestellte und verdünnte Bakteri-ensuspension hergestellt. Jeweils 50 µl dieser Suspension wurden anschließend in die Vertiefungen einer mit je 50 µl einer aufsteigenden Chloramphenicolkonzentration befüllten Mikrotiterplatte pipet-tiert. Nach erfolgter Inkubation wurde jene Antibiotikumkonzentration, die benötigt wird um ein sicht-bares Wachstum des Keimes zu hemmen (Minimale Hemmstoffkonzentration – MHK), ermittelt.
Isolate, deren MHK-Werte von denen der empfindlichen Population abwichen, wurden mittels Mikroar-ray auf das Vorhandensein unterschiedlicher Chloramphenicolresistenzgene überprüft. Anschließend wurde die Präsenz des ermittelten Genes mit Hilfe der PCR bestätigt.
Um die Lokalisierung der Resistenzgene auf dem Chromosom oder einem Plasmid genauer bestimmen zu können, wurde die DNA der resistenten Isolate mit speziellen Enzymen geschnitten und anschlie-ßend mit Pulsfeldgelelektrophorese aufgetrennt. Nach erfolgter Übertragung der geschnittenen DNA-Fragmente auf eine Membran (Southern Blotting) und Hybridisierung der Membran mit einer DIG-markierten Sonde wurde das Resistenzgen mittels Chemieluminiszenz detektiert.
Ergebnisse Bei der Überprüfung des Resistenzverhaltens von insgesamt 101 Stämmen wies der Großteil der Isola-te (97 %) gegenüber Chloramphenicol MHK-Werte im Bereich von 1 – 8 µg/ml auf. Bei drei L. johnso-nii-Isolaten tierischer Herkunft (3 %) wurde ein MHK-Wert von 32 µg/ml ermittelt.
Abb. 1: Verteilung der minimalen Hemmstoffkonzentration (MHK) gegenüber Chloramphenicol bei 101 Laktobazillen-Isolaten.
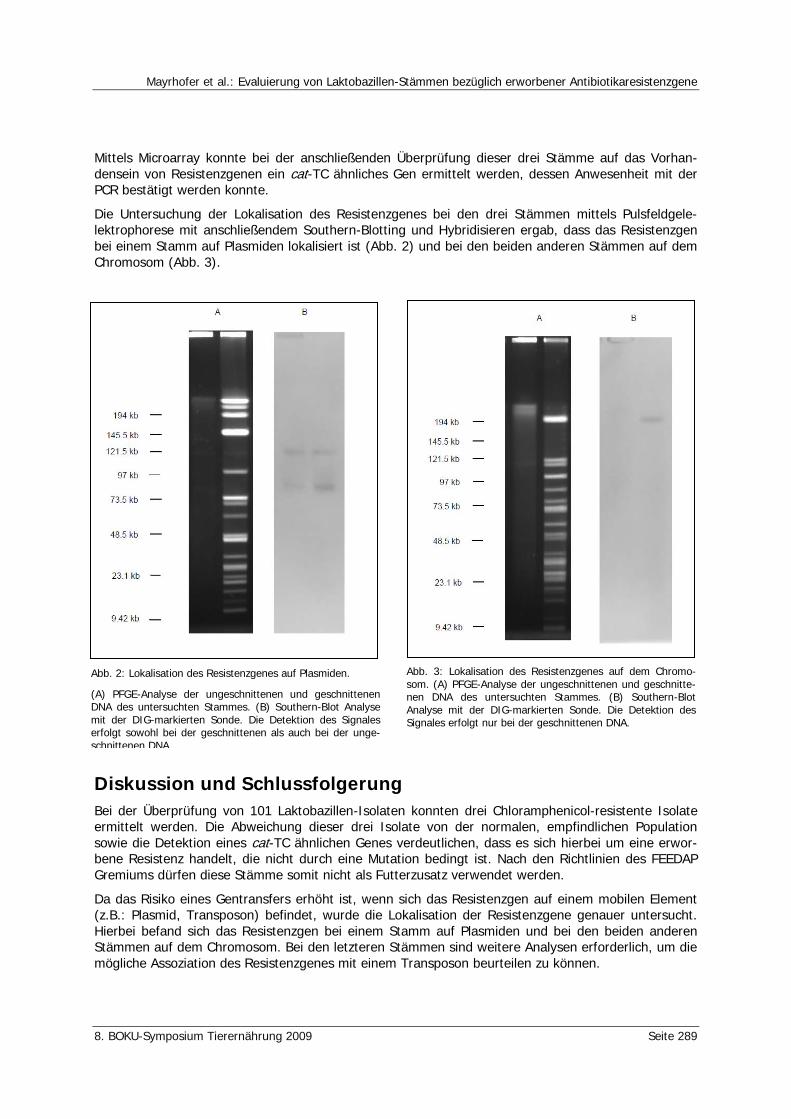
Mayrhofer et al.: Evaluierung von Laktobazillen-Stämmen bezüglich erworbener Antibiotikaresistenzgene
8. BOKU-Symposium Tierernährung 2009 Seite 289
Mittels Microarray konnte bei der anschließenden Überprüfung dieser drei Stämme auf das Vorhan-densein von Resistenzgenen ein cat-TC ähnliches Gen ermittelt werden, dessen Anwesenheit mit der PCR bestätigt werden konnte.
Die Untersuchung der Lokalisation des Resistenzgenes bei den drei Stämmen mittels Pulsfeldgele-lektrophorese mit anschließendem Southern-Blotting und Hybridisieren ergab, dass das Resistenzgen bei einem Stamm auf Plasmiden lokalisiert ist (Abb. 2) und bei den beiden anderen Stämmen auf dem Chromosom (Abb. 3).
Diskussion und Schlussfolgerung Bei der Überprüfung von 101 Laktobazillen-Isolaten konnten drei Chloramphenicol-resistente Isolate ermittelt werden. Die Abweichung dieser drei Isolate von der normalen, empfindlichen Population sowie die Detektion eines cat-TC ähnlichen Genes verdeutlichen, dass es sich hierbei um eine erwor-bene Resistenz handelt, die nicht durch eine Mutation bedingt ist. Nach den Richtlinien des FEEDAP Gremiums dürfen diese Stämme somit nicht als Futterzusatz verwendet werden.
Da das Risiko eines Gentransfers erhöht ist, wenn sich das Resistenzgen auf einem mobilen Element (z.B.: Plasmid, Transposon) befindet, wurde die Lokalisation der Resistenzgene genauer untersucht. Hierbei befand sich das Resistenzgen bei einem Stamm auf Plasmiden und bei den beiden anderen Stämmen auf dem Chromosom. Bei den letzteren Stämmen sind weitere Analysen erforderlich, um die mögliche Assoziation des Resistenzgenes mit einem Transposon beurteilen zu können.
Abb. 2: Lokalisation des Resistenzgenes auf Plasmiden.
(A) PFGE-Analyse der ungeschnittenen und geschnittenen DNA des untersuchten Stammes. (B) Southern-Blot Analyse mit der DIG-markierten Sonde. Die Detektion des Signales erfolgt sowohl bei der geschnittenen als auch bei der unge-schnittenen DNA.
Abb. 3: Lokalisation des Resistenzgenes auf dem Chromo-som. (A) PFGE-Analyse der ungeschnittenen und geschnitte-nen DNA des untersuchten Stammes. (B) Southern-Blot Analyse mit der DIG-markierten Sonde. Die Detektion des Signales erfolgt nur bei der geschnittenen DNA.

Mayrhofer et al.: Evaluierung von Laktobazillen-Stämmen bezüglich erworbener Antibiotikaresistenzgene
Seite 290 8. BOKU-Symposium Tierernährung 2009
Zusätzlich könnte man mit Hilfe von Konjugationsexperimenten die mögliche Übertragbarkeit der de-tektierten Resistenzgene testen. Hier wirkt sich jedoch das Fehlen entsprechender Methoden für be-stimmte Bakterienarten negativ aus. Ebenso ist es notwendig bereits vorhandene, phänotypische Me-thoden für die Empfindlichkeitstestung zu harmonisieren und zu standardisieren. Aufgrund des Feh-lens einer einheitlichen, international anerkannten Methodik wurden nämlich unterschiedliche Metho-den und Testmedien eingesetzt, was sich erschwerend auf die Evaluierung des Resistenzverhaltens relevanter Stämme auswirkt. Zusätzlich konnte eine Diskrepanz zwischen phänotypischen und genoty-pischen Resistenzprofilen festgestellt werden (Woodford und Sundsfjord, 2005). Vor allem das Vor-handensein unbekannter Resistenzgene und -mechanismen macht dabei die Evaluierung schwierig.
Zusätzliche Untersuchungen bezüglich der Natur der Resistenzen sowie die Harmonisierung und Wei-terentwicklung verwendeter Methoden sind daher von eminenter Bedeutung.
Literatur
Danielsen, M., Mayrhofer, S., Domig, K.J., Amtmann, E., Mayer, H.K., Flórez, A.B., Mayo, B., Korhonen, J., Tosi, L. (2008): Assessment of the antimicrobial wild-type minimum inhibitory concentration of species of the Lactobacillus delbrueckii group. Dairy Science and Technology 88, 183-191
FEEDAP Panel (2008): Update of the criteria used in the assessment ob bacterial resistance to antibiotics of human or veterinary importance. The EFSA Journal 732, 1-15
Holt, J.G., Krieg, N.R., Sneath, P.H.A., Staley, J.T., Williams, S.T. (1994): Bergey’s Manual of Determinative Bacteriology. 9. Auflage. Williams & Wilkins: Baltimore
Kastner, S., Perreten, V., Bleuler, H., Hugenschmidt, G., Lacroix, C., Meile, L. (2006): Antibiotic susceptibility patterns and resistance genes of starter cultures and probiotic bacteria used in food. Systematic and Applied Microbiolology 29, 145-155
Reuter, G. (2001): Probiotika – Möglichkeiten und Grenzen ihres Einsatzes in Lebensmitteln, im Tierfutter und in pharmazeuti-schen Präparaten für Mensch und Tier. Berliner und Münchener Tierärztliche Wochenschrift 114, 410-419
Woodford, N., Sundsfjord, A. (2005): Molecular detection of antibiotic resistance: When and Where? Journal of Antimicrobial Chemotherapy 56, 259-261
Autorenanschrift
Dr. Sigrid Mayrhofer Universität für Bodenkultur Wien Department für Lebensmittelwissenschaften und –technologie Abteilung für Lebensmittelmikrobiologie und –hygiene E-Mail: [email protected]

Firmensponsoring – Alltech (Deutschland) GmbH, Wiesenstraße 21 (W21), D-40549 Düsseldorf
8. BOKU-Symposium Tierernährung 2009 Seite 291

Petersson et al.: Einsatz der Denaturierenden Gradientengelelektrophorese (DGGE) zur Untersuchung der Darmmikrobiota des Schweins
Seite 292 8. BOKU-Symposium Tierernährung 2009
Einsatz der Denaturierenden Gradientengelelektrophorese (DGGE) zur Untersuchung der Darmmikrobiota des Schweins
Agnes Petersson1, Konrad J. Domig1, Verka Dombazyan1, Philipp Nagel2, Werner Zollitsch2, Werner Hagmüller3, Wolfgang Kneifel1 1 Universität für Bodenkultur Wien, Department für Lebensmittelwissenschaften und -
technologie, Abteilung Lebensmittelmikrobiologie und –hygiene 2 Universität für Bodenkultur Wien, Department für Nachhaltige Agrarsysteme, Insti-
tut für Nutztierwissenschaften 3 LFZ- Raumberg Gumpenstein, Außenstelle Wels, Institut für Biologische Landwirt-
schaft und Biodiversität der Nutztiere
Einleitung Die Ziele der biologischen Landwirtschaft sind in der Gesundheit der Tiere und deren Wohlergehen behaftet, um somit die Produktion sicherer tierischer Lebensmittel zu gewährleisten. Der Untersu-chung der Darmmikrobiota wird aufgrund ihres Einflusses auf die Gesundheit des Tieres eine wichtige Rolle zugeordnet (Richards et al., 2005; Sørensen et al., 2006).
Im Rahmen des EU-Projektes "Quality Low Input Food" wurden die Effekte der Verabreichung von ballaststoffreichem Grundfutter bzw. eines Probiotikums auf Leistung, Ökonomik und Tiergesundheit untersucht. Hierzu wurden 76 Bio-Mastschweine vier unterschiedlichen Futterbehandlungen ausge-setzt. Während die Kontrollgruppe keine Zusätze zur Basalnahrung bekam, wurden zwei Gruppen zusätzlich mit Mais- bzw. Grassilage gefüttert. Die vierte Behandlung bestand aus einer zusätzlichen Gabe eines Probiotikums (Bifidobacterium animalis subsp. lactis).
Ziel der Arbeit war, den Einfluss der Futterbehandlung auf die Zusammensetzung der Darmmikrobiota zu untersuchen und mögliche Unterschiede zwischen den vier Gruppen sichtbar zu machen. Hierzu kam die Denaturierende Gradientengelelektrophorese (DGGE) zum Einsatz. Durch die Applikation die-ser Methode ist es möglich, Populationsveränderungen in komplexen Umweltproben, wie zum Beispiel dem Gastrointestinaltrakt des Schweins, zu visualisieren (Muyzer und Smalla, 1998, Ercolini, 2004). Markante Banden wurden aus den Gelen ausgeschnitten und nach Klonierung mit anschließender Se-quenzierung, sowie Sequenzsuche in Datenbanken, identifiziert.
Material und Methoden Im Zuge eines Fütterungsversuchs, der Teil des EU-Projekts „Quality Low Input Food“ ist, wurden 76 Schweine vier verschiedenen Futterbehandlungen (Kontrollgruppe (KG), KG + Grassilage, KG + Mais-silage und KG + Probiotikum ausgesetzt (Nagel et al., 2007).
In Abständen mehrerer Wochen wurden von jedem Tier drei Faezesproben, sowie nach der Schlach-tung fünf Intestinalproben (Magen, Duodenum, Ileum, Caecum und Colon) gezogen und in sterilen Plastiksäcken in anaerober Atmosphäre (GENbag® anaer, Biomerieux, Marcy l'Etoile, Frankreich) bei - 80°C gelagert. Aus diesen Proben wurde die gesamte mikrobielle DNA mit Hilfe eines Ready-to-use-Isolierungskits, QIAamp® DNA Stool Mini Kits (Qiagen, Hilden, Deutschland), extrahiert, um die Pro-ben für nachfolgende molekularbiologische Untersuchungen einsatzfähig zu machen.

Petersson et al.: Einsatz der Denaturierenden Gradientengelelektrophorese (DGGE) zur Untersuchung der Darmmikrobiota des Schweins
8. BOKU-Symposium Tierernährung 2009 Seite 293
Anschließend erfolgte die Amplifizierung der DNA mittels PCR in einem Mastercycler (Eppendorf, Ham-burg, Deutschland). Hierzu kamen die in Tabelle 1 erwähnten Primerpaare Bif164f/Bif662-GC sowie Lac1f/Lac2r-GC zum Einsatz. Das Primerpaar Bif164f/Bif662-GC wurde unter Einsatz folgendes Pro-grammes zum Nachweis der Gattung Bifidobacterium eingesetzt (Satokari et al., 2001): 94°C für 5 Minuten initiale Denaturierung, 35 Zyklen mit Denaturierung bei 94°C für 30 Sekunden, Annealing-temperatur bei 62°C für 20 Sekunden und Extension bei 68°C für 40 Sekunden, anschließend 62°C für 20 Sekunden und 68°C für 7 Minuten. Die DNA der Lactobacillus Gruppe wurde mit dem Primerpaar Lac1f/Lac2r-GC mit folgendem Programm amplifiziert (adaptiert von Scanlan et al., 2006): initiale Denaturierung bei 95°C für 5 Minuten, 35 Zyklen mit Denaturierung bei 95°C für 30 Sekunden, Annea-lingtemperatur bei 61°C für 40 Sekunden und Extension bei 72°C für 1 Minute mit finalen Extension bei 72°C für 5 Minuten. Die amplifizierten PCR-Produkte mit einer ungefähren Länge von 380 bp (Lac1f/Lac2r-GC) sowie 520 bp (Bif164f/Bif662-GC) wurden in einer Agarosegelelektrophorese bestä-tigt.
Anschließend wurden die PCR-Produkte im Bio-Rad DCode™ Universal Mutation Detection System (Bio-Rad, München, Deutschland) mittels DGGE aufgetrennt. Hierzu wurden 8%ige Polyacrylamidgele mit einem linearen denaturierenden Gradient (Tabelle 1) eingesetzt und die Elektrophorese bei 60°C und 70 V für 16 h ausgeführt. Daraufhin wurden die DGGE-Bandenprofile mit Ethidiumbromid gefärbt und dadurch sichtbar gemacht. Markante Banden wurden aus den Gelen ausgeschnitten und über Nacht bei 4°C in 1xPCR-Puffer eluiert. Anschließend wurde erneut eine PCR mit den oben genannten Primern, jedoch ohne GC-Klammer, und eine weitere Agarosegelelektrophorese durchgeführt, um die Länge der PCR-Produkte zu kontrollieren. Dann erfolgte die Klonierung der PCR-Produkte in hoch effi-ziente JM109 E. coli Zellen mit Hilfe eines kommerziellen Klonierungskits, pGem®-T Vector System (Promega, Mannheim, Deutschland). Nach Ligation und Transformation (laut Anleitung) wurden aus-gewählte Zellen mit eingebautem Insert abgeerntet und jeweils in 40 µl UHQ-Wasser eluiert und bei 96°C für 10 Minuten lysiert. Erneute PCR-Amplifizierung erfolgte mit den Sequenzierungsprimern T7/SP6 mit folgendem Programm: 94°C für 2 Minuten, 20 Zyklen mit 94°C für 1 Minute, 55°C für 1 Minute und 72°C für 1 Minute, sowie finale Extension bei 72°C für 1 Minute. Die PCR-Produkte wurden wieder mittels Agarosegelelektrophorese auf den Einbau des Inserts kontrolliert. Produkte, mit einge-bautem Insert, wurden mit Hilfe des QIAquick PCR Purification Kits (Qiagen) gereinigt und anschlie-ßend extern sequenziert (Eurofins MWG Operon (Ebersberg, Deutschland)). Die erhaltenen Sequenzie-rungsergebnisse wurden mit öffentlich zugänglichen Sequenzen in einer Sequenzdatenbank unter Zuhilfenahme des BlastN Werkzeuges auf der NCBI Homepage verglichen (http://www.ncbi.nlm.nih.gov/blast/Blast.cgi).
Tabelle 1: eingesetzte Primer
Primer Sequence (5´- 3´) Specificity Annealing temp. (°C)
DGGE gra-dient (%)
Bif164f GGG TGG TAA TGC CGG ATG2 Bifidobacterium 62 45 – 55
Bif662r-GC1 CCA CCG TTA CAC CGG GAA2 Bifidobacterium 62 45 - 55
Lac1f AGC AGT AGG GAA TCT TCC A3 LAB4 61 30 - 55
Lac2r-GC1 ATT (CT)CA CCG CTA CAC ATG3 LAB4 61 30 - 55
1: GC-Klammer (5´-CCC GCC GCG CCC CGC GCC CGT CCC GCC GCC CCC GCC CG-3´) angehängt an das 5´-Ende des Primers 2: Satokari et al., 2001 3: Scanlan et al., 2006 4: Lactic acid bacteria - Milchsäurebakterien

Petersson et al.: Einsatz der Denaturierenden Gradientengelelektrophorese (DGGE) zur Untersuchung der Darmmikrobiota des Schweins
Seite 294 8. BOKU-Symposium Tierernährung 2009
Ergebnisse und Diskussion Im Zuge einer Fütterungsstudie wurden die Fäzes- und Colonproben von 76 Bio-Mastschweinen mit den Primerpaaren Bif164f/Bif662-GC sowie Lac1f/Lac2r-GC auf ihr DGGE Bandenprofil untersucht und verglichen. Interessante Banden wurden aus den Gelen ausgeschnitten und mit Hilfe einer Klonie-rungsstrategie mit anschließender Sequenzierung identifiziert. Die angegebenen PCR- sowie DGGE-Bedingungen eigneten sich zur Darstellung der Bandenmuster. Alle PCR-Produkte konnten durch den Einsatz einer Agarosegelelektrophorese bestätigt werden. Die gezeigten Abbildungen 1 und 2 zeigen typische Bilder an DGGE Profilen ausgewählter Fäzes- und Colonproben des Schweins. Die markierten Banden wurden aus dem Gel ausgeschnitten und zur Klonierung herangezogen.
Einzelne Banden, die DNA Fragmente bzw. Bakterienarten repräsentieren, können aus den DGGE Ge-len ausgeschnitten, kloniert und anschließend zur Identifizierung sequenziert werden. Somit können die untersuchten Proben nicht nur auf ihre Diversität, gezeigt durch die Komplexität des Bandenpro-fils, getestet, sondern auch einzelne Bakterienarten, die in der Umweltprobe vorhanden sind, identifi-

Petersson et al.: Einsatz der Denaturierenden Gradientengelelektrophorese (DGGE) zur Untersuchung der Darmmikrobiota des Schweins
8. BOKU-Symposium Tierernährung 2009 Seite 295
ziert werden. Die Denaturierende Gradientengelelektrophorese (DGGE) stellt eine verlässliche, schnelle Methode mit guter Reproduzierbarkeit dar und wird gerne zur Untersuchung komplexer Bakterienge-meinschaften eingesetzt (Muyzer und Smalla, 1998; McCartney et al., 2002; Namsolleck et al., 2004; Ben Amor et al., 2007).
Tabelle 2: Sequenzierergebnisse
Probe Klon Nächster Verwandter (% Sequenzähnlichkeit)
Lb1 a
L. amylovorus (99) L. acidophilus (99) L. sobrius (99) L. ultunensis (99)
Lb1 b
L. acidophilus (99) L. amylovorus (99) L. gallinarum (99) L. sobrius (99) L. ultunensis (99) L. kitasatonis (99)
Lb3 a
L. acidophilus (100) L. amylovorus (100) L. gallinarum (100) L. sobrius (100) L. ultunensis (100)
Lb3 b L. amylovorus (99) L. gallinarum (99) L. sobrius (99)
Lb3 c
L. acidophilus (99) L. amylovorus (99) L. gallinarum (99) L. sobrius (99) L. kitasatonis (99) L. ultunensis (99) L. helveticus (97)
Bif5 a
B. boum (99) B. thermacidophilum (98) B. pseudocatenulatum (98) B. thermophilum (97)
Bif8 a
B. thermophilum (99) B. thermacidophilum (99) B. ruminantium (99) B. adolescentis (99)
Bif8 b B. animalis (97) B. boum (97)
Bif8 c
B. thermacidophilum (99) B. pseudocatenulatum (98) B. boum (98) B. thermophilum (97) B. dentium (97) B. adolescentis (97)
Die eingesetzten Methoden geben einen guten Überblick über die Lactobacillen und Bifidobakterien Populationen in den untersuchten Fäzes- und Colonproben der Bio-Mastschweine. Durch die ange-wendete Strategie konnten allerdings keine eindeutigen Sequenzierergebnisse ermittelt werden. Daher war auch eine eindeutige Identifizierung der Klone und damit der aus dem DGGE Gel ausgeschnitte-nen Banden nicht möglich. Sowohl bei den Bifidobakterien, als auch bei den Lactobacillen kommen jeweils mehrere mögliche Arten als Ergebnisse in Frage (Tabelle 2). Möglicherweise können die einge-setzten Primer zwar gut zur Identifizierung auf Gattungsebene herangezogen werden, zeigen aber bei den einzelnen Arten zu wenige Unterschiede in den amplifizierten Sequenzabschnitten, um diese ein-

Petersson et al.: Einsatz der Denaturierenden Gradientengelelektrophorese (DGGE) zur Untersuchung der Darmmikrobiota des Schweins
Seite 296 8. BOKU-Symposium Tierernährung 2009
deutig einzelnen Arten zuordnen zu können. Durch die Auswahl und Verwendung anderer konservati-ver Gene bzw. Primer könnte in Zukunft eine genauere Aussage über die Lactobacillen und Bifidobak-terien Populationen in Intestinalproben gemacht werden.
Danksagung
Mein Dank gilt der H. Wilhelm Schaumann Stiftung (Hamburg, Deutschland) für die Finanzierung des Forschungsstipendiums sowie des Laborverbrauchsmaterials.
Literatur
Ben Amor K, Vaughan EE, De Vos WM. 2007. Advanced molecular tools for the identification of lactic acid bacteria. The Journal of Nutrition 137:741-747.
Ercolini D. 2004. PCR-DGGE fingerprinting: novel strategies for detection of microbes in food. Journal of Microbiological Methods 56:297– 314.
McCartney AL. 2002. Application of molecular biological methods for studying probiotics and the gut flora. British Journal of Nutrition 88:29-37.
Muyzer G, Smalla K. 1998. Application of denaturing gradient gel electrophoresis (DGGE) and temperature gradient gel electrophoresis (TGGE) in microbial ecology. Antonie van Leeuwenhoek. 73:127–141.
Nagel P, Domig KJ, Hagmüller W, Pfalz S, Kronsteiner S, Ortner B, Sundrum A, Zollitsch W. 2007. Effects of silage or probiotics on performance and gut microbial composition of organic growing-finishing pigs. Paper presented at: 6. BOKU-Symposium Tierernährung, Sekundärwirkungen von Futterinhaltsstoffen – vom Nährstoff zum Wirkstoff; 15 November 2007; Vienna, Aus-tria.
Namsolleck P, Thiel R, Lawson P, Holmstrøm K, Rajilic M, Vaughan EE, Rigottier-Gois L, Collins MD, de Vos WM, Blaut M. 2004. Molecular methods fort he analysis of gut microbiota. Microbial Ecology in Health and Disease 16:71-85.
Richards JD, Gong J, De Lange CFM. 2005. The gastrointestinal microbiota and its role in monogastric nutrition and health with an emphasis on pigs: Current understanding, possible modulations, and new technologies for ecological studies. Canadian Journal of Animal Science 85:421-435.
Satokari RM, Vaughan EE, Akkermans ADL, Saarela M, De Vos WH. 2001. Bifidobacterial diversity in human feces detected by genus-specific PCR and Denaturing Gradient Gel Electrophoresis. Applied and Environmental Microbiology 67:504–513.
Scanlan PD, Shanahan F, O´Mahony C, Marchesi JR. 2006. Culture-independent analyses of temporal variation of the dominant fecal microbiota and targeted bacterial subgroups in Crohn’s disease. Journal of Clinical Microbiology 44:3980–3988.
Sørensen JT, Edwards S, Noordhuizen J, Gunnarsson S. 2006. Animal production systems in the industrialized world. Revue Scientifique et Technique (International Office of Epizootics) 25:493-503.
Autorenanschrift
DI Agnes Petersson Abteilung Lebensmittelmikrobiologie und –hygiene Department für Lebensmittelwissenschaften und -technologie Universität für Bodenkultur Wien Gregor Mendel-Strasse 33, A-1180 Wien E-Mail: [email protected]

Müller et al.: Gut Microbiota – Studying bacterial DGGE profiles
8. BOKU-Symposium Tierernährung 2009 Seite 297
Gut Microbiota – Studying bacterial DGGE profiles
Anneliese Müller1, Corinna Kern1, Verity Ann Sattler1, Gertrude Wegl1, Gerd Schatzmayr2 and Viviana Klose1 1 BOKU – University of Natural Ressources and Applied Life Sciences, Vienna, Dept.
for Agrobiotechnology, Div. Environmental Biotechnology, Tulln, Austria 2 Biomin GmbH, Industriestraße, Herzogenburg, Austria
Introduction Gut Microbiota The intestinal tracts of both, humans and animals, are colonized by a great number of different bacte-rial species. Important effects of these natural inhabitants on the host have been described, ranging from metabolic activities improving nutrient absorption and energy conservation, to stimulation of epithelial gut cells as well as the immune system and protection against pathogenic bacteria (Guarner F. and Malagelada J.R. 2003). Nevertheless, also pathogens colonize the gut.
Despite of the importance of gut microbiota, many of these natural inhabitants are still unknown and a good part might be uncultivable requiring culture-independent techniques for investigation.
Aim of the study Goal of this study was to find out whether varying nutrition results in a changing gut microflora in chicken as a profound impact of nutrition on occurrence of different bacteria can be assumed. There-fore, three groups of chicken were fed with a standard poultry diet containing either 0.1% or 0.2% yeast cell derivative (YCW∗
Although changes of the whole bacterial community were documented by using a universal primer pair for the amplification step at the beginning of an analysis with DGGE, special attention was paid on the genus Bifidobacterium (Actinobacteria), which is assumed to have particularly beneficial effects on the host.
) or 0.1% YCW plus Poultry Star (Biomin), which contains the putative beneficial gut bacteria Lactobacillus reuteri, Lactobacillus salivarius, Enterococcus faecium, Bifidobac-terium animalis and Bifidobacterium thermophilum. Gut samples have been taken from these trial groups plus an additional control group, which received the control diet without any of the two addi-tives.
A further goal was to compare the microflora along the intestinal tract. The gut consists of various sections, which provide distinct environmental conditions requiring specific adaptations of the inhabi-tants (Kleerebezem, M. and Vaughan, E.E. 2009). Changes in histology as for instance the presence of microvilli in the small and the absence in the large intestines as well as changes in pH and oxygen concentration might influence colonization behaviour. As a consequence, samples were taken from the lower part of the small intestine (ileum), the caeca and the upper part of the large intestine (colon). Samples from ileum and colon were taken from both, chyme and mucus. Gut bacteria, which are able to attach to the mucus, a slime layer covering the inner epithelial cells, might be long-term residents of the gut and might thus be dissimilar from bacteria inhabiting chyme, a mixture of half-digested food and gastric secretions, which is passing the intestines. Furthermore, faeces was investigated to test how far faecal communities represent gut communities.
∗ YCF...constituent of the yeast cell wall; believed to increase the number of lactobacilli attached to the mucosa and to decrease the number of coliforms; overall believed to improve growth performance of broilers (Yang Y. et al. 2008)

Müller et al.: Gut Microbiota – Studying bacterial DGGE profiles
Seite 298 8. BOKU-Symposium Tierernährung 2009
Approach Different communities were characterized by applying Denaturing Gradient Gel Electrophoresis (DGGE), a DNA fingerprinting technique. Thereby the DNA of all bacteria within each sample gets extracted, the region of interest amplified and subsequently separated according to its sequence. Varying GC-contents of the DNA result in dissimilar migration behaviour of the amplicons along a de-naturing gradient within an electric field, leading to specific banding patterns. Divergent banding pat-terns between the samples are believed to represent differing communities.
Material and Methods Feeding trial Four groups of 8 broiler chicken each, were fed with either feed containing 0.1% YCF, 0.2% YCF, 0.1% YCF plus Poultry Star or with feed containing no additives (control group).
Sampling and storage Samples were taken from the mucosa and chyme of the ileum and the colon and from the caecal chyme of each chicken. Each group within the feeding trial consisted of 8 chicken which gave a total sample number of 140 plus 4 additional faecal samples (one from each feeding group). All samples were stored at -20°C.
Establishing reference systems Reference strains from various sources were sub-cultured in the respective media (see table 1) until they were dense enough to be harvested. Thereby the cultures were centrifuged for 10 min with 3000 rpm. After a washing step with 1ml 1xPBS, the centrifugation step was repeated.
DNA extraction Environmental DNA was extracted by using the QIAamp DNA Stool Kit according to the manufacturer’s instructions. DNA of the reference strains was extracted by using the ArchivePure DNA Yeast and Gram-+ Kit or by performing a Phenol/Chloroform/Isoamylalkohol-extraction (Chan et al. 2003). For the faecal samples washing steps were introduced to increase the yield of DNA. Thereby approximate-ly 2 g of each faecal sample were weighed in a 15 ml Greiner tube. 10 ml sterile 1xPBS (phosphate buffered saline) were added to each sample and vortexed. Centrifugation steps were slightly modified from the protocol given in N.D.Phillips et al. (2006): centrifugation was done at 3000 g for 10 min and the supernatant discarded. The washing step was repeated and around 500 mg of each remaining final pellet were transferred to a 2 ml tube. Subsequent extraction was done with the QIAamp DNA Stool Kit. Quantity and quality of DNA was controlled photometrically with a Nanodrop and by gel electrophoresis. Eluted DNA was stored at -20 °C.
Amplification of DNA For the amplification of all bacteria the universal primer pair 341f (5´-CCT ACG GGA GGC AGC AG-3´) and 518r (5´-CGT ATT ACC GCG GCT GCT GG-3´) (Muyzer et al. 1998) specific for the V3 region of the 16SrDNA was used. A GC-clamp was attached to the 5`-end of the forward primer for further analysis with DGGE. A 50 µl PCR reaction mix consisted of 5 µl 10x PCR buffer (100 mM Tris-HCl (pH 8,8), 500 mM KCl, 0.1 % Tween 20 and 15 mM MgCl2; peqlab, Austria), 0.5 µl Taq Polymerase (2 U/µl; peqlab, Austria), 0.5 µl 10 mM dNTPs (peqlab, Austria), 0.5 µl of each primer (12.5 pmol; VBC-Biotech, Austria), 41 µl distilled H2O and 1 µl of the template (~50 ng/µl). The cycling program of the applied Touch down PCR was as follows: first denaturation: 94 °C for 5 min, then 10 cycles of denatu-ration for 30 s at 94 °C, annealing for 20 s at 64 °C -0.5 °C/cycle, elongation for 40 s at 72 °C; fol-lowed by 20 cycles of denaturation for 30 s at 94 °C, annealing for 20 s at 56 °C, elongation for 40 s

Müller et al.: Gut Microbiota – Studying bacterial DGGE profiles
8. BOKU-Symposium Tierernährung 2009 Seite 299
at 72 °C; a final elongation step of 7 min at 72 °C was performed and the ending temperature in the cycler was 15 °C. Products were frozen until further use.
Table 1: Reference strains with the respective media and growth conditions
representative strain code additional information
Clostridium perfringens CCUG 47895 37 °C, anaerobic, DRCM (Oxoid)
Selenomonas ruminantium DSM 2150 37 °C, anaerobic, isolate from bovine rumen
Lactobacillus reuteri DSM 16350 37 °C, anaerobic, MRS (Oxoid), chicken strain
Lactobacillus salivarius DSM 16351 37 °C, anaerobic, MRS, chicken strain
Pediococcus acidilactici DSM 16210 37 °C, anaerobic, MRS, chicken strain Enterococcus cecorum DSM 20682 37 °C, anaerobic, poultry caecum
Bifidobacterium animalis DSM 16284 37 °C, anaerobic, trypticase-phytone-yeast ex-tract (Rada et al. 2000), chicken strain
Bifidobacterium gallinarium DSM 20105 37 °C, anaerobic, trypticase-phytone-yeast ex-tract, chicken cecum
Bifidobacterium choerinum DSM 20434 37 °C, anaerobic, trypticase-phytone-yeast ex-tract, piglet faeces
Bifidobacterium longum DSM 20219 37 °C, anaerobic, trypticase-phytone-yeast ex-tract, intestine of adult
Bifidobacterium longum DSM 20211 37 °C, anaerobic, trypticase-phytone-yeast ex-tract, pig faeces
Bifidobacterium thermoacidophilum subsp.porcinum DSM 17755 37 °C, anaerobic, trypticase-phytone-yeast ex-
tract, pig faeces
Bifidobacterium thermophilum DSM 20210 37 °C, anaerobic, trypticase-phytone-yeast ex-tract, pig faeces
Bifidobacterium boum DSM 20423 37 °C, anaerobic, trypticase-phytone-yeast ex-tract, bovine rumen
Bifidobacterium bifidum DSM 20239 37 °C, anaerobic, trypticase-phytone-yeast ex-tract
Bifidobacterium breve DSM 20213 37 °C, anaerobic, trypticase-phytone-yeast ex-tract, intestine of infant
Bifidobacterium pullorum DSM 27685 37 °C, anaerobic, trypticase-phytone-yeast ex-tract, chicken faeces
Salmonella choleraesuis subsp. cho-leraesuis (S. enterica subsp. enteri-ca)
DSM 554 37 °C, nutrient broth or agar + 25µg/ml tetracyc-line
Salmonella enteritidis Serover Ente-riditis USDA 59 37 °C, nutrient broth
Salmonella typhimurium Serovar Typhimurium USDA 554 37 °C, nutrient broth
E. coli O1:K1:H7 DSM 30083 37 °C, nutrient broth; isolate from urine; patho-genic to chicken
Campylobacter jejuni CCUG 25903 37 °C, microaerobic, blood medium, isolate from chicken
Bacteroides fragilis DSM 2151 37 °C, anaerobic; chopped meat medium, isolate from appendix abscess

Müller et al.: Gut Microbiota – Studying bacterial DGGE profiles
Seite 300 8. BOKU-Symposium Tierernährung 2009
For the amplification of bacteria belonging to the genus Bifidobacterium two Nested PCR approaches were conducted: in the first one the primer pair 164f (5´-GGG TGG TAA TGC CGG ATG-3´)/662r (5`-CCA CCG TTA CAC CGG GAA-3´) (Langendijk et al. 1995) specific for bifidobacterial 16SrDNA and in the second round the universal primer pair 341f-GC and 518r was used as described previously. The reaction mix of the first PCR was alike the amplification with 341f-GC/518r apart from minor modifica-tions: the concentration of MgCl2 was increased to 2.5 mM and the total reaction volume was 25 µl (volumes of the other reactants were adapted to this final volume). 1 µl of the product of the first round was used as template in the second round.
The second approach consisted of two specific amplification steps: in the first round the primer pair lm3 (5´-CGG GTG CTI CCC ACT TTC ATG-3´) and lm26 (5`-GAT TCT GGC TCA GGA TGA ACG-3`) specific for bifidobacterial 16SrDNA and rendering a fragment of 1420 bp was used. Reaction mix and volume were according to the amplification with the primer pair 164f/662r. The cycling program con-sisted of a primary denaturation step of 5 min, followed by 35 cycles of denaturation for 30 s at 94 °C, annealing for 30 s at 57 °C, elongation for 1 min 30 s at 68 °C and a final elongation step for 7 min at 68 °C (following Satokari R.M. et al. 2001). 1 µl of the first PCR were used in the second PCR per-formed with the primer pair 164f/662r-GC with total volumes of 50 µl. The cycling program was: de-naturation for 5 min at 94 °C followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 62 °C for 30 s and elongation at 68 °C for 40 s and a final elongation step of 7 min at 68 °C. For amplifi-cation of environmental DNA with the second approach, conditions were altered for optimization as for instance the concentration of MgCl2, the annealing temperature and the template concentration within the reaction mix. Products were frozen until further use.
DGGE DGGE analysis was performed by using a Biorad- or Ingeny-System. Polyacrylamid gels were prepared with Acrylamid/Bis (40 % 37.5:1) (final concentration of 8 %), Glycerol (final conc. of 2 %), 50xTAE-buffer (2 M Tris, 1 M Sodium Acetate, 50 mM Na2EDTA, dH2O; final conc. of 0.5x), Formamid (end conc. 40 %) and Urea (7 M). For the universal analysis with the primer pair 341f-GC/518r, a gradient of 30-60 % was used. For an analysis specific for Bifidobacteria with the second approach (1st PCR with the primer pair lm3f/lm26r; 2nd PCR with the primer pair 164f/662r-GC), the optimal gradient was 45-60 %. Gels were run at 60 °C for 16 h with 96 V (Biorad-System) or 120 V (Ingeny-System) in 0.5xTAE-buffer. The gel was stained with SYBRGreen for approximately 30 min DNA fragments were visualized by using a transilluminator.
Results Universal approach A reference system was established. DNA of the samples is currently under investigation. The de-tected results of this study will be discussed later on.
Bifidobacterium specific approach A reference system using the PCR approach with two specific primer pairs has been established but needs further verification (Fig.1). The size of the amplicon in the first round of the Nested PCR ap-proach was 1420 bp, after the second amplification round it was 520 bp. The optimal gradient for this assay was 45-60 % and allowed to distinguish between all reference strains apart from, firstly, B.thermoacidophilum and B.thermophilum, and, secondly, B.choerinum and B.pullorum.

Müller et al.: Gut Microbiota – Studying bacterial DGGE profiles
8. BOKU-Symposium Tierernährung 2009 Seite 301
Fig.1: DGGE-gel showing the banding pattern of the reference strains for the Bifidobacterium specific approach
B.thermoacidoph. ... B.thermoacidophilum subsp.porcinum
While no products could be obtained with the new very specific PCR approach using the environ-mental DNA as a template, all of the samples screened with the less specific approach so far (24 out of 144), showed bands at the appropriate height when controlling the PCR by gel electrophoresis.
Discussion So far no final answers of the actual questions of the study can be given as the analysis is still running and investigation techniques partly have to be optimized. As soon as the DGGE analysis is completed, DGGE profiles can be compared by using the software Gel Compare II (Applied Maths, Belgium). Sta-tistical analysis should enable the detection of significant dissimilarities and allow conclusions on influ-ence of nutrition on gut microbiota. Furthermore, new information about the communities in different gut sections will be available.
Although bacteria belonging to the genus Bifidobacterium seem to occur within the samples, they could not be detected using the very specific approach. This is contrary to results of Martín R. et al. (2009), who investigated breast milk and infant faeces by performing DGGE with the same Nested PCR approach. An increase of the MgCl2 concentration and the template concentration as well as a decrease of the annealing temperature did not give better results. A possible reason for the absence of PCR products might be the size of the amplicon in the first round (about 1420 bp). A longer tem-plate is probably more difficult to copy as the chance of inaccessibility due to secondary and tertiary structure of the DNA, which could block the elongation step of the Taq Polymerase, is higher. DNA could also be affected by freezing and thawing. Disruptions of the DNA are more likely to appear within the area of interest the longer the amplicon is. However, disruptions due to freezing-thawing of the isolated DNA are not very likely to occur in all gut samples screened so far. The main advantage of this approach is, besides being more accurate in amplifying Bifidobacteria, that longer amplicons give more phylogenetic information. Prominent bands could thus be cut out of the gel, eluted, se-quenced and used for phylogenetic analysis and identification. A PCR product of only 177 bp does not provide the same phylogenetic information, rendering identification impossible. Therefore, further

Müller et al.: Gut Microbiota – Studying bacterial DGGE profiles
Seite 302 8. BOKU-Symposium Tierernährung 2009
approaches using different primer pairs shall be considered instead of performing the analysis with the less specific system right away.
Conclusion Generally DGGE seems to be a good choice for tracking population changes; a major drawback is, nevertheless, that no quantitative data is available and that, even though the reference system allows conclusions on the appearance of certain bacteria, no real identification of the occurring bacteria is possible, unless for single bands. Thus it might be highly interesting to focus more on the identifica-tion of gut bacteria in future studies.
References
Chan R.K., Wortman,C.R., Smiley B.K., Hendrick C.A. (2003): Construction and use of a computerized DNA fingerprint database for lactic acid bacteria from silage. Journal of Microbiological Methods 55(3):565-574.
Guarner F., Malagelada J.-R. (2003): Gut flora in health and disease. The Lancet. 360: 512-519
Kleerebezem M., Vaughan E.E. (2009): Probiotic and Gut Lactobacilli and Bifidobacteria: Molecular Approaches to Study Diversity and Activity. Annu. Rev. Microbiol. 63, 269-290.
Langendijk P.S., Schut F., Jansen G.J., Raangs G.C., Kamphuis G.G., Wilkinson M.H.F., Weeling G.W. (1995): Quantitative Fluorescence In Situ Hybridization of Bifidobacterium spp. With Genus-Specific 16S rRNA-Targeted Probes and Its Application in Fecal Samples. Applied and Environmental Microbiology 61(8): 3069-3075.
Martín R., Jiménez E., Heilig H., Férnandez L., Marín M.L., Zoetendal E.G., Rodríguez J.M. (2009): Isolation of Bifidobacteria from Breast Milk and Assessment of the Bifidobacterial Population by PCR-Denaturing Gradient Gel Electrophoresis and Quantitative Real-Time PCR. Applied and Environmental Microbiology 75(4): 965-969.
Muyzer G., Smalla K. (1998): Application of denaturing gradient gel electrophoresis (DGGE) and temperature gradient gel elec-trophoresis (TGGE) in microbial ecology. Antonie van Leeuwenhoek 73: 127-141.
Phillips N.D., La T., Hampson D.J. (2006): Development of a two-step nested duplex PCR assay for the rapid detection of Bra-chyspira pilosicoli and Brachyspira intermedia in chicken faeces. Veterinary Microbiology 116: 239–245.
Rada V., Petr J. (2000): A new selective medium for the isolation of glucose non-fermenting bifidobacteriafrom hen caeca. J. Microbiol. Methods 43:127-132.
Satokari R.M., Vaughan E.E., Akkermans A.D.L., Saarela M., de Vos W.W. (2001): Bifidobacterial Diversity in Human Feces Detected by Genus-Specific PCR and Denaturing Gradient Gel Electrophoresis. Applied and Environmental Microbiology 67(2): 504-513.
Yang, Y., Iji PA., Kocher A., Mikkelsen LL., Choct M. (2008): Effects of dietary mannanoligosaccharide on growth performance, nutrient digestibility and gut development of broilers given different cereal-based diets. J. Anim. Physiol. Anim. Nutr. (Berl.). 92(6), 650-659.
Corresponding author
Mag. Anneliese Müller Universität f. Bodenkultur Interuniversitäres Department für Agrarbiotechnologie, IFA Tulln Abteilung für Umweltbiotechnologie Konrad Lorenz Straße 20, A -3430 Tulln E-mail: [email protected]

Wegl et al.: Evaluation and selection of a stable endogenous reference gene for studying immunological host response with quantitative reverse transcription PCR (qRT- PCR) in coccidiosis challenged chickens
8. BOKU-Symposium Tierernährung 2009 Seite 303
Evaluation and selection of a stable endogenous reference gene for studying immunological host response with quantitative re-verse transcription PCR (qRT- PCR) in coccidiosis challenged chickens
Gertrude Wegl1, Todd J. Applegate2, Gerd Schatzmayr3 and Viviana Klose1 1 University of Natural Resources and Applied Life Sciences Vienna, Dep. for Agrobio-
technology, Tulln, Austria 2 Purdue University, Dep. of Animal Science, West Lafayette, IN, USA 3 Biomin Research Center, Tulln, Austria
Introduction To analyze the expression of inflammatory parameters during infectious diseases, qRT-PCR has be-come a method of to analyze the expression of inflammatory genes by quantification of mRNA due to its high sensitivity, reproducibility and large dynamic of range (Bustin et al. 2000). However, in addi-tion to careful reaction set-up and optimization, accurate and reliable normalization to some standard is highly recommended to adjust for differences in RNA extraction and cDNA synthesis or for the pres-ence of inhibitors in different sample materials (Bustin et al. 2002). In relative quantification internally expressed reference genes (RG), mainly housekeeping genes (HKG) like e.g. glyceraldehydes -3- phosphate dehydrogenase (GAPDH) or ß-Actin (ßAct), are commonly used for standardization. Since it is well documented, that some of these genes are somehow regulated (Goidin et al. 2001, Lupberger et al. 2002, Schmittgen et al. 2000), their expression stability should be validated to ensure accurate quantification results.
The aim of the present study was to identify suitable HKG as internal control to normalize the expres-sion of immunological marker genes in spleen and cecal tonsils in a mild coccidial stressor model for feed additive evaluation in broiler chickens.
Materials and Methods The feeding trial was designed as a 2 x 2 factorial experiment (summarized in Tab.1.), 1 day old broi-ler chicken were grouped in 4 groups and supplemented with either control diet or diet with phytogen-ic feed supplementation (Biomin® P.E.P. 125). Half of the birds were vaccinated with 2 different doses of Paracox® 5 while the remaining broiler chicken within each group remained unvaccinated to investi-gate ‘natural’ exposure to oocyst due to shedding.
Tab. 1. Experimental design of the coccidial stressor model Treatment (n total= 24) Coccidial vaccine dose1 PEP
1 1x None 2 1x PEP 3 5x None 4 5x PEP
1Half of the birds within each treatment group remained unvaccinated

Wegl et al.: Evaluation and selection of a stable endogenous reference gene for studying immunological host response with quantitative reverse transcription PCR (qRT- PCR) in coccidiosis challenged chickens
Seite 304 8. BOKU-Symposium Tierernährung 2009
At day 28, samples were taken from spleen and cecal tonsils from vaccinated and naturally exposed birds and immediately stored in RNAlater® (Ambion). Total RNA extraction was performed using the RNeasy Mini Kit (QIAGEN) according to the instructions of the manufacturer. Contaminating DNA was removed by DNAse digestion (Fermentas) and reverse transcription was performed using the Rever-tAidTM H Minus First Strand cDNA Synthesis Kit from Fermentas. For RT-PCR a fixed amount of RNA was used and oligo-dT primer and random hexamer primer were added in equal concentrations.
Primers were designed with the Primer Premier software using mRNA sequences for GAPDH, ribosom-al protein L4 (RPL4), TATA box binding protein (TBP) and hypoxanthine phosphoribosyltransferase (HPRT) obtained from the NCBI database (Tab.2.).
Tab.2. Characteristics of gene specific Real-Time PCR assays
Gene Accession no. Sequence (5' - 3') Prod-uct length
Pri-mer
MgCl2
Ta Efficien-cy1
GAPDH K01458.1 F: GCC CAG AAC ATC ATC CCA 137 bp
150mM 4mM 60°
C 0.96
R: CGG CAG GTC AGG TCA ACA 150mM 4mM
RPL4 NM_001007479.1
F: GTT ATG CCA TCT GTT CTG CC 236 bp
150mM 4mM 60°
C 0.89 R: GCG ATT CTT CAT CTT ACC CT
150mM 4mM
TBP NM_205103 F: TGC CCG AAA TGC TGA ATA 289 bp
150mM 4mM 58°
C 1.01
R: GCT GCT GAA CTG CTG GTG 150mM 4mM
HPRT AJ_132697 F: TTA CGA CCT GGA CTT GTT
174 bp
150mM 4mM 55°
C 0.98
R: AAT TTA TAG CCA CCC TTG 150mM 4mM
1 Primer efficiencies are given as mean efficiency values of repeated standard curve measurements
Real Time PCR was performed in the Real-Plex 2 S system (Eppendorf) using the MESA Green qPCR Master Mastermix (Eurogentec). Cycle conditions were set as follows: 5 min at 95°C followed by 45 PCR cycles with a denaturation step at 95°C for 15 sec, annealing at primer dependent temperature (see Tab.2.) for 15 sec and elongation at 72°C for 40 sec. Different dilutions of cDNA were used in qPCR (1:8 dilution of cDNA for TBP and HPRT assays and 1:16 dilution for GAPDH and RPL4 assays). Samples were analysed in triplicate and all assays were quantified including a non-template control (NTC) and a standard curve at each plate.
To analyze the expression stability of the chosen candidate RG, the software geNorm (Vandesompele et al. 2002), NormFinder (Andersen et al. 2004) and BestKeeper (Pfaffl et al. 2004) were used.
Results The expression stability of the four RG was assessed for each sample group. For analysis of gene ex-pression with geNorm, raw expression values were used for calculating the expression stability meas-ure (M). The M value for each RG is the average pairwise variation for that gene with all other tested

Wegl et al.: Evaluation and selection of a stable endogenous reference gene for studying immunological host response with quantitative reverse transcription PCR (qRT- PCR) in coccidiosis challenged chickens
8. BOKU-Symposium Tierernährung 2009 Seite 305
control genes. Stepwise exclusion of the genes with the highest M-value allowed ranking of the tested genes according to their expression stability from least to most stable: HPRT, GAPDH, RPL4 and TBP (Fig.1.).
Fig.1. Average expression stability values(m) of control genes analyzed by the geNorm software plot-ted from least stable (left) to most stable (right). NormFinder uses a model-based approach (mixed linear effect modeling) to estimate expression sta-bility and ranks the candidates with minimal estimated intra- and intergroup variations. TBP was iden-tified as the most stable expressed gene, whereas taking the different experimental groups into ac-count, TBP and GAPDH were identified as best combination (data not shown).
The software program BestKeeper analyses variability in expression of candidate RG by calculating the cycle threshold (Ct) data variation. The founding principle for identification of stable expressed refer-ence genes is that proper reference genes should display a similar expression pattern.
Tab.3. BestKeeper descriptive statistical analysis for reference genes based on cycle threshold (Ct) values
GAPDH HPRT RPL4 TBP N1 48,00 48,00 48,00 48,00 geo mean [Ct] 1 21,20 26,61 20,35 25,42 ar mean [Ct] 1 21,22 26,65 20,37 25,43 min [Ct] 1 19,72 24,20 19,00 24,36 max [Ct] 1 23,51 31,21 22,53 27,97 std dev [± Ct] 1 0,82 1,10 0,53 0,55 CV [ % Ct] 1 3,85 4,15 2,62 2,18 1Abbreviations: N: Number of samples; geo mean [Ct]: the geometric mean of Ct; ar mean [Ct]: the arithmetric mean of Ct; min [Ct] and max [Ct): the extreme values of Ct, SD [±Ct]: the standard deviation of the Ct; Cv [Ct]: the coefficient of variance expressed as a percentage on the Ct level In this context RPL4 and TBP were found to be the most stable expressed reference genes with CV ± SD of 2.62 ± 0.53 or 2.18 ± 0.55, respectively. HPRT and GAPDH exhibited the highest coefficient of variance with 4.15 ± 1.1 or 3.85 ± 0.82. Results are summarized in Tab.3. Heterogeneous variance

Wegl et al.: Evaluation and selection of a stable endogenous reference gene for studying immunological host response with quantitative reverse transcription PCR (qRT- PCR) in coccidiosis challenged chickens
Seite 306 8. BOKU-Symposium Tierernährung 2009
between the groups of differently expressed genes invalidated use of the pearson correlation coeffi-cient in the BestKeeper tool.
Discussion Normalization of gene expression is used to correct sample to sample variation in order to identify gene specific variation. Therefore real–time PCR data are usually normalized against a reference gene. Since even the expression of so called housekeeping genes has been shown to vary with experimental treatment or tissue type, careful selection of an endogenous control gene is necessary for the acquisi-tion of accurate quantification data. This aspect of controlling is essential, especially in approaches, where only slight differences in gene expression are expected. Obtained data may be misinterpreted as no change of expression although slight, but biologically significant induction might have relevance (Taylor et al. 2007). Therefore, Vandesompele et al. proposed the use of multiple reference genes in normalization for a good performance of qPCR analysis (Vandesompele et al. 2004). However, norma-lization to multiple reference genes requires additional resources and depending on the experimental system, might not be necessary.
In this preliminary study, the free available software programs geNorm, NormFinder and BestKeeper were used to evaluate the expression stability of 4 candidate RG in spleen and cecal tissue of chicken exposed to different experimental conditions. All approaches generated the same output, anyway HPRT showed greatest inconsistencies. Among the remaining HKG TBP was outlined as the most sta-ble gene, indicated by the lowest M value in geNorm, lowest stability value in NormFinder and low SD variation in BestKeeper. Differences in the particular ranking of TBP, RPL4 and GAPDH might be due to different calculation algorithms, e.g. NormFinder also takes inter- and intra- group variations into account. In order to check the value of this preliminary validation, these results have to be confirmed by repeating the qPCR and software analysis. Furthermore, inclusion of HKG involved in other cell processes (e.g. structural related-genes like ß-Actin or α-tubulin) might enlarge the information on the stability of this system and confirm results of this experiment.
In conclusion, although quite laborious, the necessity of carefully selecting stable reference genes for ensuring quality-assurance in normalization can be outlined and recent publications in animal science research (De Boever et al. 2008, Nygard et al. 2007) support this demand. Although results are only preliminary, this experiment presents useful tools for evaluation of an appropriate HKG and is, so far, the first study on reference gene stability in different chicken tissues.
References
Andersen,C.L., Jensen,J.L., and Orntoft,T.F. (2004) Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res 64: 5245-5250.
Bustin,S.A. (2000) Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J Mol Endocrinol 25: 169-193.
Bustin,S.A. (2002) Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): trends and problems. J Mol Endocrinol 29: 23-39.
De Boever, S., Vangestel,C., De Backer, P., Croubels,S., and Sys,S.U. (2008) Identification and validation of housekeeping genes as internal control for gene expression in an intravenous LPS inflammation model in chickens. Vet Immunol Immunopathol 122: 312-317.
Goidin,D., Mamessier,A., Staquet,M.J., Schmitt,D., and Berthier-Vergnes,O. (2001) Ribosomal 18S RNA prevails over glyceraldehyde-3-phosphate dehydrogenase and beta-actin genes as internal standard for quantitative comparison of mRNA levels in invasive and noninvasive human melanoma cell subpopulations. Anal Biochem 295: 17-21.

Wegl et al.: Evaluation and selection of a stable endogenous reference gene for studying immunological host response with quantitative reverse transcription PCR (qRT- PCR) in coccidiosis challenged chickens
8. BOKU-Symposium Tierernährung 2009 Seite 307
Lupberger,J., Kreuzer,K.A., Baskaynak,G., Peters,U.R., le,C.P., and Schmidt,C.A. (2002) Quantitative analysis of beta-actin, beta-2-microglobulin and porphobilinogen deaminase mRNA and their comparison as control transcripts for RT-PCR. Mol Cell Probes 16: 25-30.
Nygard,A.B., Jorgensen,C.B., Cirera,S., and Fredholm,M. (2007) Selection of reference genes for gene expression studies in pig tissues using SYBR green qPCR. BMC Mol Biol 8 (6)
Pfaffl,M.W., Tichopad,A., Prgomet,C., and Neuvians,T.P. (2004) Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper Excel-based tool using pair-wise correlations. Biotechnology Letters 26: 509-515.
Schmittgen,T.D. and Zakrajsek,B.A. (2000) Effect of experimental treatment on housekeeping gene expression: validation by real-time, quantitative RT-PCR. Journal of Biochemical and Biophysical Methods 46: 69-81.
Taylor,D.L., Thomson,P.C., de Silva,K., and Whittington,R.J. (2007) Validation of endogenous reference genes for expression profiling of RAW264.7 cells infected with Mycobacterium avium subsp. paratuberculosis by quantitative PCR. Veterinary Immunology and Immunopathology 115: 43-55.
Vandesompele,J., De,P.K., Pattyn,F., Poppe,B., Van,R.N., De,P.A., and Speleman,F. (2002) Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol 3: 0034.1-0034.11
Corresponding author
Mag. Gertrude Wegl Universität f. Bodenkultur Interuniversitäres Department für Agrarbiotechnologie, IFA Tulln Abteilung für Umweltbiotechnologie Konrad Lorenz Straße 20, A -3430 Tulln E-mail: [email protected]

Balabánová et al.: Effect of sample conservation on blood parameters analyses
Seite 308 8. BOKU-Symposium Tierernährung 2009
Effect of sample conservation on blood parameters analyses
M. Balabánová, Š. Hošková, A. Vašátková, M. Večerek, L. Zeman
Faculty of Agronomy, Mendel University of Agriculture and Forestry Brno
Introduction The good health status is base of livestock keeping systems. Production diseases have subclinical symptom very often (MATĚJÍČEK, 2004), they persist for a long time and when they are diagnostic on the base of clinical symptoms than organism is very harmed generally and each tendency of remedy is difficult. Dairy cow is set aside from drove for consequential reasons like sterility of cow, lower milk usefulness, disease of hooves, diseases of organs as digestive tract and liver.
For example, disturbances of metabolism energy, sugars, lipids represent the most frequent distur-bances of time after calve and at the beginning of lactation (STAUFENBIEL, 2007).
These disturbances give rise to considerable economical worse therefore it is necessary solve these problems earlier than clinical symptoms evolve into illness.
HOFÍREK et al. (2004) say that it is important take into account concrete conditions of farm. It should be economic analysis of implied trust costs and financial profitability of preventative equipment. On the basis of this facts, farmer knows that very important observe health condition of his animals.
Metabolism dysfunction can be caused by stress which are caused by more physiological and metabol-ism changes. These changes could be at the bottom of metabolism disfunction (VAN SAUN, 2004).
In this time, when lot of farmers tries to save any money, for example from veterinary corps of their animal, there is very important pay to health. Production disturbances are possible observe on changes of blood components.
There are many ways how it is possible of specify disturbance. The most important of them are based on blood analysis. The basic characteristic of blood is its ability of making blood clot (it means coagu-lation). In case that we cannot do analysis in time (from native x fresh blood) so we must use test tube for bleed blood with any preparation which avoid inception of blood clot (anticoagulant).
An anticoagulant is a substance that prevents coagulation; that is, it stops blood from clotting. A group of pharmaceuticals called anticoagulants can be used in vivo as a medication for thrombotic disorders. Some chemical compounds are used in medical equipment, such as test tubes, blood trans-fusion bags, and renal dialysis equipment.
Apart from heparin, most of these chemicals work by binding calcium ions, preventing the coagulation proteins from using them.
• Citrate is usually in blue Vacutainer tube. It is in liquid form in the tube and is used for coa-gulation tests, as well as in blood transfusion bags. It gets rid of the calcium, but not as strongly as EDTA. Correct proportion of this anticoagulant to blood is crucial because of the dilution. It can be in the form of sodium citrate or ACD.
• Oxalate has a mechanism similar to that of citrate. It is the anticoagulant used in fluoride (grey top) tubes.
• EDTA is denoted by mauve or purple caps on Vacutainer brand test tubes. This chemical strongly and irreversibly binds calcium (WIKIPEDIA, 2009).

Balabánová et al.: Effect of sample conservation on blood parameters analyses
8. BOKU-Symposium Tierernährung 2009 Seite 309
EDTA is a widely used acronym for the chemical compound ethylenediaminetetraacetic acid (which has many other names like EDTA, H4EDTA, Diaminoethanetetraacetic acid, Edetic acid, Edetate, Ethy-lenedinitrilotetraacetic acid, Versene, Ethylene diamine tetraacetic acid, ethylenediaminetetraacetate, 2-[2-(Bis(carboxymethyl)amino) ethyl-(carboxymethyl)amino]acetic acid). EDTA is a polyamino car-boxylic acid. This colorless, water-soluble solid is widely used to dissolve scale. Its usefulness arises because of its role as a chelating agent, i.e. its ability to "sequester" metal ions such as Ca2+ and Fe3+. After being bound by EDTA, metal ions remain in solution but exhibit diminished reactivity
EDTA finds many specialized uses in the biomedical laboratories, such as in veterinary ophthalmology as an anticollagenase to prevent the worsening of corneal ulcers in animals. In tissue culture EDTA is used as a chelating agent, which binds to calcium and prevents joining of cadherins between cells, preventing clumping of cells grown in liquid suspension, or detaching adherent cells for passaging (WIKIPEDIA, 2009).
Heparin is currently the most widely used anticoagulant (DAVENPORT,2009). Heparin, a highly-sulfated glycosaminoglycan, is widely used as an injectable anticoagulant, and has the highest nega-tive charge density of any known biological molecule (COX et al., 2004). It can also be used to form an inner anticoagulant surface on various experimental and medical devices such as test tubes. Phar-maceutical grade heparin is derived from mucosal tissues of slaughtered meat animals such as porcine (pig) intestine or bovine (cow) lung (LINHARDT et al., 1999).
Although used principally in medicine for anticoagulation, the true physiological role in the body re-mains unclear, because blood anti-coagulation is achieved mostly by endothelial cell-derived heparin sulfate proteoglycans (MARCUM et al., 1986). Heparin is usually stored within the secretory granules of mast cells and released only into the vasculature at sites of tissue injury. It has been proposed that, rather than anticoagulation, the main purpose of heparin is in a defensive mechanism at sites of tissue injury against invading bacteria and other foreign materials (NADER et al., 1999). In addition, it is preserved across a number of widely different species, including some invertebrates that lack a similar blood coagulation system (WIKIPEDIA, 2009).
The aim of our experimental was determined if there is some difference in blood analysis in depen-dence on using different anticoagulation preparation. We would like to find out which of watched anti-coagulants are the best for using to blood taking of cows.
Materials and Methods Our experiment was realized on school agricultural farm Žabčice, which is possession of Mendel Uni-versity of Agriculture and Forestry in Brno. We had 10 cows of Holstein breed for our observation. These cows were at the same stage lactation, at the third lactation.
Our endeavour was eliminate influence of stress factors therefore monitoring cows stayed in their drove and they had the same conditions of humidity, temperature, air flow, feeding ration or place for their movement as they had after our experiment.
We had 2 different anticoagulants for our observation – heparin and EDTA. We took blood to 1 test tube with lithium heparin (test tube with blue stopper) and 1 test tube with K3 EDTA (test tube with green stopper) each of ten cows from vena caudalis mediana. These specimens of blood had to be mixed with contained anticoagulants in the test tubes. After we had marked each ones of tubes, we went to laboratory of Mendel University and we did blood analysis there.
The ABC Vet (Animal Blood Counter Veterinary) was used for our blood analysis. It is fully automated (Microprocessor controlled) hematology analyzer used for the in vitro diagnostic testing of blood spe-cimens. The ABC Vet is available as 8 or 16 parameters hematology system. We were decided that we are monitored only 8 from them. Minimum volume must be 100 µl blood.

Balabánová et al.: Effect of sample conservation on blood parameters analyses
Seite 310 8. BOKU-Symposium Tierernährung 2009
We analyzed: WBC…….White blood cell count RBC……..Red blood cell count HGB…….Hemoglobin HCT…….Hematocrit PLT……..Platelet count MCV……Mean cell volume MCH……Mean corpuscular hemoglobin MCHC….Mean corpuscular hemoglobin concentration
Results and Discussion Results of 8 monitoring parameters of heparins blood were compared with parameters of EDTA´s blood each cow. Measured values of blood parameters are represented in Table 1 and 2.
Table 1. Blood parameters of samples with heparin
Number Heparin
WBC RBC HGB HCT PLT MCV MCH MCHC
[103/mm3] [106/mm3] [g/dl] [%] [103/mm3] [µm3] [pg] [g/dl]
1 6.4 6.58 10.1 31.4 506 48 15.3 32.1
2 8.9 6.68 9.9 30.7 263 46 14.8 32.2
3 4.3 5.7 9.5 28.7 281 50 16.6 33.0
4 8.5 6.34 9.5 29.8 444 47 15.0 32.0
5 7.8 6.27 9.1 28.1 435 45 14.5 32.5
6 6.9 5.77 9.2 28.4 485 49 15.9 32.3
7 7.2 6.43 9.5 29.4 321 46 14.8 32.4
8 11.0 8.56 10.9 34.6 405 40 12.7 31.5
9 5.6 5.91 9.9 31.0 402 52 16.8 32.0
10 6.7 6.11 9.8 30.1 316 49 16.0 32.5

Balabánová et al.: Effect of sample conservation on blood parameters analyses
8. BOKU-Symposium Tierernährung 2009 Seite 311
Table 2. Blood parameters of samples with EDTA
Number EDTA
WBC RBC HGB HCT PLT MCV MCH MCHC
[103/mm3] [106/mm3] [g/dl] [%] [103/mm3] [µm3] [pg] [g/dl]
1 6.4 6.43 9.9 30.6 461 48 15.4 32.2
2 8.9 6.73 9.9 31.0 369 46 14.7 31.9
3 4.5 5.77 9.6 29.1 288 50 16.7 33.1
4 8.3 6.26 9.4 29.5 420 47 15.0 31.8
5 7.9 6.20 9.1 27.8 438 45 14.6 32.6
6 6.7 5.73 9.1 28.2 526 49 15.8 32.2
7 7.3 6.27 9.5 28.8 343 46 15.1 32.9
8 10.7 8.15 10.5 32.8 466 40 12.9 32.1
9 5.7 5.84 10.0 30.6 411 52 17.2 32.7
10 7.2 6.03 9.5 29.4 378 49 15.7 32.2
Table 3. Averages and standard deviations of blood parameters
Heparin
WBC RBC HGB HCT PLT MCV MCH MCHC
[103/mm3] [106/mm3] [g/dl] [%] [103/mm3] [µm3] [pg] [g/dl]
Average 7.3 6.4 9.7 30.2 385.8 47.2 15.2 32.3
SD 1.77 0.77 0.49 1.80 81.16 3.12 1.13 0.38
EDTA
WBC RBC HGB HCT PLT MCV MCH MCHC
[103/mm3] [106/mm3] [g/dl] [%] [103/mm3] [µm3] [pg] [g/dl]
Average 7.4 6.3 9.7 29.8 410.0 47.2 15.3 32.4
SD 1.64 0.67 0.41 1.41 64.93 3.12 1.13 0.41
In Table 3, there are survey of averages and standard deviations of blood elements. For example mean cell volumes (MCV) are same both sample blood in test tube with heparin and EDTA. Either of them have 47.2 ± 3.12 µm3 counted to plate counts (PLT) are diverse. PLT heparin´s blood has 385.8 ± 81.16 103/mm3 and PLT blood with EDTA has 410 ± 64.93 103/mm3.

Balabánová et al.: Effect of sample conservation on blood parameters analyses
Seite 312 8. BOKU-Symposium Tierernährung 2009
Figure 1 - 8. Illustrations of summary observed blood parameters
There are not any statistically conclusive differences between two diverse methods of blood mainten-ance. So that it is possible to say generally that it can be used like anticoagulant both heparin and EDTA.
Conclusion Wrong nourishment is a cause of origin many production diseases. Nutrition deficiencies are evocated by low concentration of nutrients, mineral substances, trace elements and vitamins a lot of times. Therefore is important maintenance of good nutrition and health for good cow´s keeping which are connected closely.
In case of non-performance than it would be cause of metabolism disturbances and negative effects to efficiency, milk usefulness or fertility.
It is necessary veterinary prevention of drove and timely resolution of problem. Since lot of diseases is possible observe from changes of bloods component. We are detected that anticoagulant which was used in test tube, had not establishable effect on blood analysis. So we could say that price is only the aspect which is deciding.
References
Anticoagulant [online]. 2001-2009 , this page was last modified on 12 July 2009 at 01:23 [cit. 2009-08-10]. Available from WWW: < http://en.wikipedia.org/wiki/Anticoagulant >.
COX, M.; NELSON D. Lehninger, Principles of Biochemistry. Freeman. 2004. pp. 1100. ISBN 0-71674339-6.DAVENPORT, A. Low-molecular-weight heparin as an alternative anticoagulant to unfractionated heparin for routine outpatient haemodialysis treatments. NEPHROLOGY [online]. 2009, vol. 14, is. 5 [cit. 2009-08-10], s. 455-461. Dostupný z WWW: <http://apps.isiknowledge.com/full_record.do?product=WOS&search_mode=Refine&qid=2&SID=3BFiE5FH9h@pBAfKgP2&page=1&doc=1>. ISSN 1320-5358.
EDTA [online]. 2001-2009 , this page was last modified on 4 August 2009 at 17:07 [cit. 2009-08-10]. Available from WWW: < http://en.wikipedia.org/wiki/EDTA >.
Heparin [online]. 2001-2009 , this page was last modified on 30 July 2009 at 02:36 [cit. 2009-08-10]. Available from WWW: < http://en.wikipedia.org/wiki/Heparin >.

Balabánová et al.: Effect of sample conservation on blood parameters analyses
8. BOKU-Symposium Tierernährung 2009 Seite 313
HOFÍREK, B., PECHOVÁ A., DOLEŽEL R., PAVLATA L., DVOŘÁK R., FLEISCHER P. et al.: Produkční a preventivní medicína v chovech mléčného skotu. 1. vyd. Brno : VFU, 2004. 184 p. ISBN 80-7305-501-5
LINHARDT, RJ.; GUNAY, NS. Production and Chemical Processing of Low Molecular Eieght Heparins, Sem. Thromb. Hem.. 1999. 3, p. 5-16
MARCUM, JA. et al. Anticoagulantly active heparin-like molecules from mast cell-deficient mice, Am. J. Physiol. 1986. 250 (5 Pt 2): p. 879-888
MATĚJÍČEK, M. Využití metabolických testů k hodnocení výživy u skotu . Informační magazín VVS Veměřovice [online]. 2004 [cit. 2009-08-10], p. 2-3. Available from WWW: <http://www.vvs.cz/vvs_info/jaro2004>.
NADER, HB. et al. Heparin sulfates and heparins: similar compounds performing the same functions in vertebrates and inverterbrates?, Braz. J. Med. Biol. Res. 1999. 32 (5): p. 529-538
STAUFENBIEL, R. Metabolické testy [online]. 2007 [cit. 2009-08-10]. Dostupný z WWW: <http://www.genoservis.cz/layout.php>.
VAN SAUN, R. J. Výživa a management krav v přechodovém období : klíč k úspěšné reprodukci stáda. In Sborník referátů z odborného semináře . Hradec Králové : [s.n.], 17. 4. 2004.
Corresponding author
Ing. Marie Balabánová Department of Animal Nutrition and Forage Production Mendel University of Agriculture and Forestry Brno Zemědělská 1, 613 00, Brno, Czech Republic E-mail: [email protected] tel.: + 420 545 133 170

Finck et al.: Untersuchung der Energie- und Nährstoffbilanzen in der Vorbereitungs- und Säugephase von Sauen
Seite 314 8. BOKU-Symposium Tierernährung 2009
Untersuchung der Energie- und Nährstoffbilanzen in der Vorbe-reitungs- und Säugephase von Sauen
Elmar Finck1, Georg Dusel1, Markus Klaßen2, Karl Landfried2 1 Fachhochschule Bingen 2 Lehr- und Versuchsanstalt für Viehhaltung, Hofgut Neumühle
Einleitung Die Rentabilität der Zuchtsauenhaltung hängt ganz entscheidend von den biologischen Leistungen der Sauen ab. Die Erhöhung der biologischen Leistungen definiert sich überwiegend in einer Steigerung der Zahl lebend geborener, beziehungsweise abgesetzter Ferkel. Auch eine größere Wurfmasse und damit verbunden ein höheres durchschnittliches Geburtsgewicht der Ferkel wird angestrebt, da schwe-rere Ferkel vitaler sind und dadurch geringere Ferkelverluste aufweisen. Bei steigender Ferkelzahl je Wurf und steigender Aufzuchtleistung in der Säugephase bedeutet dies, dass die Milchmenge der Sauen steigen muss. Somit steigt auch die erforderliche metabolische Leistung, wobei die Futterauf-nahmekapazität der Sauen den Nährstoff- und Energieinput limitiert. Es kann ein Nährstoff- und Ener-giedefizit entstehen, das durch den Abbau an Körpermasse kompensiert werden muss. Eine energeti-sche Aufwertung des Futters durch Fette oder Öle, sowie das Einschmelzen von Körperfett belastet den Leberstoffwechsel und es kann zu einer ketogenen Stoffwechsellage kommen (IBEN und SCHNURR-BUSCH, 1999).
Ziel dieser experimentellen Untersuchung war es, den Energieversorgungsstatus, sowie die Nährstoff-aufnahme und den Nährstoffbedarf von Zuchtsauen zu ermitteln um mögliche ketogene Stoffwechsel-lagen der Zuchtsauen anhand von ausgewählten Blutparametern zu bestätigen. Besondere Beachtung fanden die Blutparameter, die bei Milchkühen zur Feststellung einer Ketose (verursacht durch Ener-giemangel) herangezogen werden. Die von NEWCOMB et al. (1991) ermittelten Werte für ß-hydroxibuttersäure (ß-HBS, Indikatorsubstanz für Ketose) bei Sauen, mit bis zu 253 µmol/l, reichen an den von DRACKLEY et al. (1992) ermittelten Schwellenwert einer Ketose bei Kühen. Dieser wird mit ~390 µmol/l angegeben. Da Labore für klinische Diagnostik zur Untersuchung der Stoffwechselbelas-tung bei Milchkühen sogenannte "Blutanalyse-Pakete" anbieten, wurde ein solches zur Untersuchung des Schweineblutes herangezogen.
Material und Methoden Es erfolgten zwei Versuchsdurchgänge mit insgesamt 14 Sauen, die an der Lehr- und Versuchsanstalt für Viehhaltung Hofgut Neumühle durchgeführt wurden. Als Sauengenetik wurden die Rassen DL, BHZP und JSR ausgewählt. Die Sauen befanden sich im zweiten bis achten Reproduktionszyklus. Nachfolgende Parameter wurden erfasst:
Futter-, Nährstoff und Energieaufnahme der Sauen und Ferkel (täglich) Rückenfettdicke (Stambock-Methode) und Gewicht der Sauen (wöchentlich ab 7. Tag a.p.) tägl. Ferkellebendmasse/-entwicklung Blutparameter der Sauen

Finck et al.: Untersuchung der Energie- und Nährstoffbilanzen in der Vorbereitungs- und Säugephase von Sauen
8. BOKU-Symposium Tierernährung 2009 Seite 315
Futteraufnahme der Sauen und der Ferkel
Die täglich aufgenommene Futtermenge der Sauen wurde ab dem ersten Tag p.p. (post partum) er-fasst. Hierzu wurde zweimal täglich das in den Trog gegebene Futter gewogen. Alle Futterreste die aus dem Trog entfernt wurden, wurden gewogen und in die Tages-Futterbilanz eingerechnet.
Die Ferkel wurden erst ab dem vierzehnten Lebenstag zugefüttert, damit der Ferkelzuwachs aus-schließlich aus der Milchaufnahme resultierte. Das ab dann genommene Ferkelfutter wurde in Ferkel-zuwachs umgerechnet und floss somit indirekt in die Kalkulation der Energiebilanz der Sauen ein.
Rückenfettdicke und Gewicht der Sauen
Diese beiden Parameter wurden wöchentlich erfasst, beginnend eine Woche vor dem Abferkeltermin mit der Einstallung in das Abferkelabteil. Am Absetztag (28. Tag p.p.) wurden zum sechsten und letz-ten mal gemessen. Die Rückenfettdicke wurde mit einem Bild-gebenden Ultraschall Gerät gemessen (Aloka SSD 500, Micrus). Es wurden sechs Messpunkte je Sau erfasst und zu einem Mittelwert ver-rechnet. Die Messpunkte wurden farblich markiert, damit immer an der gleichen Stelle gemessen wur-de. Die Lage der Punkte wurde in Anlehnung an die Stambock-Methode (SIVERDING, 2000) gewählt. Hierbei liegen jeweils drei Messpunkte links, beziehungsweise rechts, der Wirbelsäule. Die Distanz zur Wirbelsäule betrug 10 cm. Der Abstand zwischen den Messpunkten betrug 15 cm. Das Gewicht der Sauen wurde mit einer geeichten Viehwaage ermittelt.
Blutparameter
Im Laufe jedes Durchganges wurden drei Blutproben je Sau gezogen. Die Proben wurden vier Stun-den nach der morgendlichen Fütterung durch Punktion der Vena jugularis externa entnommen. Zeit-punkt der Probenahme war eine Woche a.p., vier und elf Tage p.p. bzw. 28 Tage p.p. Die Blutent-nahme am 28.-Tag p.p. erfolgte nur bei der zweiten Versuchsgruppe, da hier die Proben des elften Tages nach der Geburt nicht verwendet werden konnten. Alle Proben wurden im Diagnostik-Labor LABOKLIN (97668 Bad Kissingen) auf zwölf Parameter untersucht, siehe Tabelle 1.
Tabelle 1: Untersuchte Blutparameter
Parameter: Einheit
Gesamteiweiß g/l Harnstoff mmol/l Cholesterin mmol/l GLDH U/l NEFA mmol/l AST (GOT) U/l g-GT U/l Bilirubin ges. µmol/l Calcium mmol/l Phosphat-anorg. mmol/l ß-HBS mmol/l Insulin µU/ml
Dieses Spektrum an Parametern wird unter der Bezeichnung "Leistungsparameter Wiederkäuer" zu einem Analyseblock zusammengefasst und beinhaltet unter anderem die ketose-typischen Parameter NEFA (freie Fettsäuren), ß-HB (beta-Hydroxy-Buttersäure) und Leberenzymkonzentrationen. Zusätzlich wurden alle Proben auf den Insulingehalt untersucht.

Finck et al.: Untersuchung der Energie- und Nährstoffbilanzen in der Vorbereitungs- und Säugephase von Sauen
Seite 316 8. BOKU-Symposium Tierernährung 2009
Berechnung der (Energie-)bilanzen
Die Berechnung der Nährstoff- und Energiebilanz erfolgte aus der aufgenommenen Futtermenge (In-put), dem Erhaltungsbedarf der Sau sowie der aufgezogenen Wurfmasse (Ferkelzunahmen). Der Energiebedarf wurde auf Futteräquivalente bezogen. Das heißt, dass die metabolische Energie der Milch, die zur Erzeugung von einem Kilogramm Ferkelmasse notwendig ist, in von der Sau aufzuneh-mende Futterenergie umgerechnet wurde. Hierzu wurde der Verwertungsfaktor der Energie für die Milchproduktion (kl) von 0,7 angesetzt. Der Energiegehalt der Milch wurde mit 5,1 MJ/kg angenom-men, der Milchbedarf für ein kg Ferkelzuwachs mit 4,1 kg Milch/kg. Die Beifütterung der Ferkel, die ab dem vierzehnten Lebenstag erfolgte, wurde bei der Berechnung der Milchleistung der Sau beachtet. Für die Berechnung des Erhaltungsbedarfes wurde mit einem täglichen Bedarf von 0,44 MJ ME je Kilogramm metabolischer Körpermasse (LM0,75) gerechnet. Das Gewicht der Sauen vom ersten Tag post partum wurde dieser Rechnung zugrunde gelegt.
Bei einem Körpermasseabbau von einem Kilogramm Körpersubstanz wird eine Energiemenge von durchschnittlich 22 MJ ME freigesetzt. Bei einem Verwertungsfaktor von 0,8 für die Umsetzung dieser Energie in Milchenergie ergibt dies eine Milchmenge von etwa 3,4 Litern. Zur Produktion dieser Milch-menge müsste die Sau alternativ 25 MJ Energie über das Futter aufnehmen. (GfE, 2006)
Ergebnisse und Diskussion Blutuntersuchungen
Die die Analyseergebnisse aller Blutproben wurden auf eine Korrelation mit den errechneten Energie-bilanzen zum Zeitpunkt der Blutprobennahme überprüft. Bezüglich des Energiedefizits besteht die größte Beziehung mit dem Gehalt an Cholesterin (r=0,75). Auch der Gehalt an Harnstoff ist, bei einem Korrelationskoeffizient von 0,63, mit dem Wert der Energiebilanz korreliert. Die höchste Korrelations-koeffizient zwischen zwei Blutparametern besteht zwischen dem Bilirubingehalt und dem NEFA-Gehalt (r=0,66). In Tabelle 2 sind alle ermittelten Korrelationskoeffizienten dargestellt. In dieser Matrix sind die Analyseergebnisse vom 28.-Tag p.p., der zweiten Sauengruppe, nicht eingeflossen. Dadurch er-höht sich der Korrelationskoeffizient zwischen der Energiebilanz und dem Cholesteringehalt auf 0,87.
Anhand der analysierten Werte wurde keine signifikante Beziehung zwischen den ermittelten Energie-bilanzen und den ketose-typischen Blutparameter ß-HBS und NEFA festgestellt. Die in der Untersu-chung verwendete Analysegenauigkeit dieses Parameters (mmol/l) wird bei der Diagnostik von Ketose bei Milchkühen verwendet. REVELL et al. (1998) und NEWCOMB et al. (1991) analysierten das Blut bei ihren Untersuchungen an Zuchtsauen ebenfalls auf ß-HBS, jedoch im µmol-Bereich. REVELL et al. (1998) ermittelten bei ihren Untersuchungen Werte von 6-42 µmol ß-HBS/l. Die Werte von NEWCOMB et al. (1991) lagen zwischen 11 und 253 µmol ß-HB/l. Die von uns ermittelten geringen Werte deuten darauf hin, dass die verwendete Analysegenauigkeit bei ß-HBS nicht ausreichend war, um Korrelatio-nen mit der Energiebilanz zu ermitteln. Zu dem traten keine klinischen Anzeichen bei den Sauen auf, die auf eine akute Ketose und damit erhöhte Werte hindeuteten.
Die Freien Fettsäuren (NEFA, nonesterified fatty acids), die im Bereich der Milchviehhaltung ebenfalls zur Diagnose ketotischer Stoffwechsellagen herangezogen werden, zeigten in vorliegender Untersu-chung ebenfalls keinen Zusammenhang mit den ermittelten Energiebilanzen. WELDON et al. (1994) konnten bei Untersuchungen an Sauen, die während der Laktation unterschiedlich energetisch ver-sorgt wurden, eine Beziehung zwischen dem NEFA-Gehalt im Blut und der Energieversorgung feststel-len. Die von WELDON et al. (1994) verwendete Analysegenauigkeit (µEq/l) war höher als die in der vorliegender Untersuchung verwendete Einheit (mmol/l). Die Übertragung der Analysegenauigkeit für Blutproben aus dem Bereich der Milchkuhhaltung auf den Bereich der Zuchtsauenhaltung ist mögli-cherweise nicht gegeben. Zur Bewertung der Blutinhaltstoffe bei Schweinen hätte die Analyse der
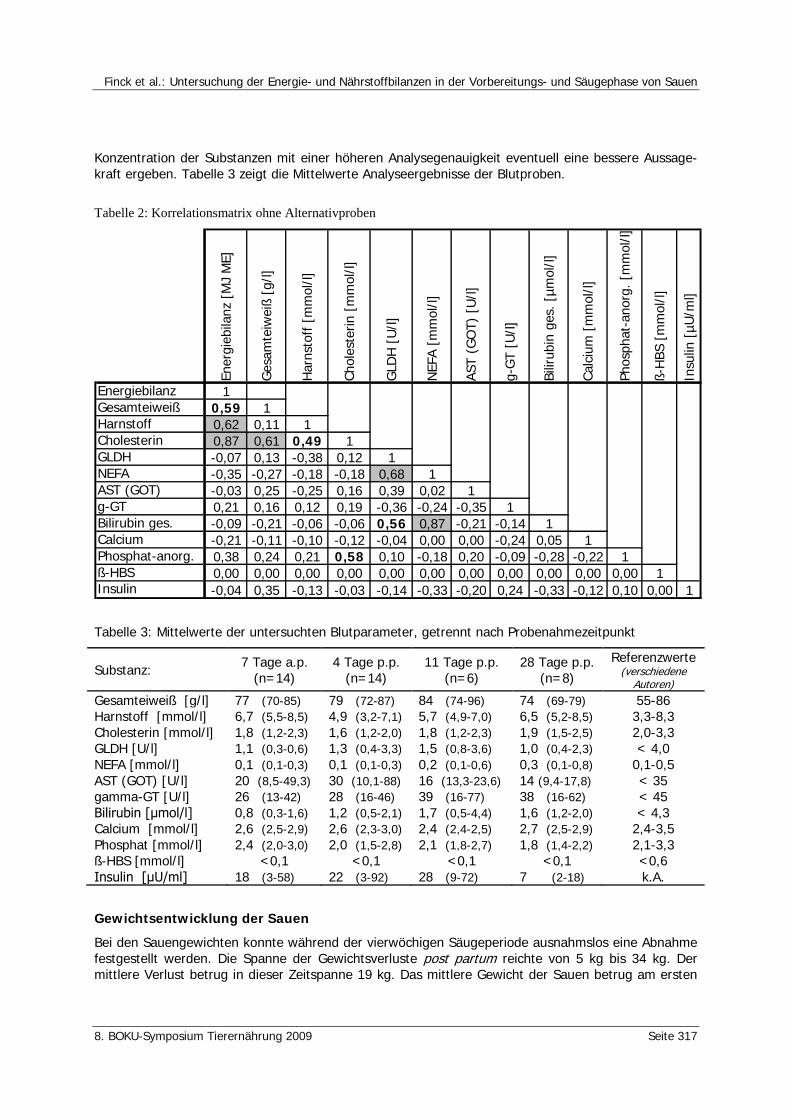
Finck et al.: Untersuchung der Energie- und Nährstoffbilanzen in der Vorbereitungs- und Säugephase von Sauen
8. BOKU-Symposium Tierernährung 2009 Seite 317
Konzentration der Substanzen mit einer höheren Analysegenauigkeit eventuell eine bessere Aussage-kraft ergeben. Tabelle 3 zeigt die Mittelwerte Analyseergebnisse der Blutproben.
Tabelle 2: Korrelationsmatrix ohne Alternativproben En
ergi
ebila
nz [
MJ
ME]
Ges
amte
iwei
ß [g
/l]
Har
nsto
ff [
mm
ol/l]
Chol
este
rin [
mm
ol/l]
GLD
H [
U/l]
NEF
A [m
mol
/l]
AST
(GO
T) [
U/l]
g-G
T [U
/l]
Bilir
ubin
ges
. [µm
ol/l]
Calc
ium
[m
mol
/l]
Phos
phat
-ano
rg. [
mm
ol/l]
ß-H
BS [
mm
ol/l]
Insu
lin [
µU/m
l]
Energiebilanz 1Gesamteiweiß 0,59 1Harnstoff 0,62 0,11 1Cholesterin 0,87 0,61 0,49 1GLDH -0,07 0,13 -0,38 0,12 1NEFA -0,35 -0,27 -0,18 -0,18 0,68 1AST (GOT) -0,03 0,25 -0,25 0,16 0,39 0,02 1g-GT 0,21 0,16 0,12 0,19 -0,36 -0,24 -0,35 1Bilirubin ges. -0,09 -0,21 -0,06 -0,06 0,56 0,87 -0,21 -0,14 1Calcium -0,21 -0,11 -0,10 -0,12 -0,04 0,00 0,00 -0,24 0,05 1Phosphat-anorg. 0,38 0,24 0,21 0,58 0,10 -0,18 0,20 -0,09 -0,28 -0,22 1ß-HBS 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 1Insulin -0,04 0,35 -0,13 -0,03 -0,14 -0,33 -0,20 0,24 -0,33 -0,12 0,10 0,00 1
Tabelle 3: Mittelwerte der untersuchten Blutparameter, getrennt nach Probenahmezeitpunkt
Substanz: 7 Tage a.p. (n=14)
4 Tage p.p. (n=14)
11 Tage p.p. (n=6)
28 Tage p.p. (n=8)
Referenzwerte (verschiedene
Autoren) Gesamteiweiß [g/l] 77 (70-85) 79 (72-87) 84 (74-96) 74 (69-79) 55-86 Harnstoff [mmol/l] 6,7 (5,5-8,5) 4,9 (3,2-7,1) 5,7 (4,9-7,0) 6,5 (5,2-8,5) 3,3-8,3 Cholesterin [mmol/l] 1,8 (1,2-2,3) 1,6 (1,2-2,0) 1,8 (1,2-2,3) 1,9 (1,5-2,5) 2,0-3,3 GLDH [U/l] 1,1 (0,3-0,6) 1,3 (0,4-3,3) 1,5 (0,8-3,6) 1,0 (0,4-2,3) < 4,0 NEFA [mmol/l] 0,1 (0,1-0,3) 0,1 (0,1-0,3) 0,2 (0,1-0,6) 0,3 (0,1-0,8) 0,1-0,5 AST (GOT) [U/l] 20 (8,5-49,3) 30 (10,1-88) 16 (13,3-23,6) 14 (9,4-17,8) < 35 gamma-GT [U/l] 26 (13-42) 28 (16-46) 39 (16-77) 38 (16-62) < 45 Bilirubin [μmol/l] 0,8 (0,3-1,6) 1,2 (0,5-2,1) 1,7 (0,5-4,4) 1,6 (1,2-2,0) < 4,3 Calcium [mmol/l] 2,6 (2,5-2,9) 2,6 (2,3-3,0) 2,4 (2,4-2,5) 2,7 (2,5-2,9) 2,4-3,5 Phosphat [mmol/l] 2,4 (2,0-3,0) 2,0 (1,5-2,8) 2,1 (1,8-2,7) 1,8 (1,4-2,2) 2,1-3,3 ß-HBS [mmol/l] <0,1 <0,1 <0,1 <0,1 <0,6 Insulin [μU/ml] 18 (3-58) 22 (3-92) 28 (9-72) 7 (2-18) k.A.
Gewichtsentwicklung der Sauen
Bei den Sauengewichten konnte während der vierwöchigen Säugeperiode ausnahmslos eine Abnahme festgestellt werden. Die Spanne der Gewichtsverluste post partum reichte von 5 kg bis 34 kg. Der mittlere Verlust betrug in dieser Zeitspanne 19 kg. Das mittlere Gewicht der Sauen betrug am ersten

Finck et al.: Untersuchung der Energie- und Nährstoffbilanzen in der Vorbereitungs- und Säugephase von Sauen
Seite 318 8. BOKU-Symposium Tierernährung 2009
Tag post partum 264 kg (208 kg bis 321 kg). Die mittlere Rückenfettdicke (RFD) zu Laktationsbeginn betrug 20,4 mm. Die Spanne reichte von 14,7 bis 28,2 mm Fettauflage. Diese reduzierte sich im Mittel um 6,3 mm. Diese Reduktion der RFD bei den 14 untersuchten Sauen beinhaltet Werte von 1,9 bis 10,4 mm Abnahme. Tabelle 4 beinhaltet die Mittelwerte aller Sauen, sowie die Spanne der relativen Entwicklung der Einzeltiere.
Tabelle 4: Entwicklung des Gewichtes und der Rückenfettdicke der Sauen (Mittelwerte, n=14):
Gewicht Rückenfettdicke Mittelwerte Einzeltiere Mittelwerte Einzeltiere [kg] relativ [%] [mm] relativ [%] 7. Tag a.p. 288 109,1 104,8-114,5 20,5 100,5 93,7-108,4 1. Tag p.p. 264 100,0 --- 20,4 100,0 --- 8. Tag p.p. 258 97,7 93,2-100,4 17,6 86,4 76,9-96,9 15. Tag p.p. 255 96,6 92,5-101,8 16,3 79,8 68,2-95,6 22.Tag p.p. 250 94,4 88,8-100,4 15,1 74,2 60,7-92,5 28. Tag p.p. 246 92,7 86,3-97,7 14,1 69,4 58,6-90,9
Energiebilanzen
Es war festzustellen, dass jede Sau, bis auf eine Ausnahme, eine negative (kalkulierte) Energiebilanz während der vierwöchigen Säugephase hatte. Im Mittel trat ein Energiedefizit von 690 MJ ME auf. Die aufgenommene Futtermenge reichte somit nicht aus, um den Energiebedarf zu decken. Das höchste Defizit wurde mit -1.375 MJ ME ermittelt. Abbildung 1 zeigt den Verlauf der mittleren Energieaufnah-me und des mittlere Energiebedarfes.
0102030405060708090
100110120
1 2 3 4 5 6 7 8 9 10 11 12 13 14Laktationstag
Ener
gie
[MJ M
E]
Energieaufnahme Energiebedarf (kalkuliert)
Abbildung 1: Mittlerer Energiestatus der Sauen (n=14) vom 1. bis 14.-Laktationstag
Lysin/Ca-Versorgung
Bei einer Annahme einer mittleren pc VQ des Lysin von 85% beträgt die intermediäre Verwertung des pcv Lys 74%. Der Bedarf an pcv Lys je kg Sauenmilch errechnet sich somit zu 5,1 g/kg. Bei einem Milchaufwand von 4.1 kg/kg Ferkelmassezuwachs ergibt sich ein Bedarf an pcv Lys von 20,9 g/kg Ferkelmassezuwachs. (GfE, 2006)

Finck et al.: Untersuchung der Energie- und Nährstoffbilanzen in der Vorbereitungs- und Säugephase von Sauen
8. BOKU-Symposium Tierernährung 2009 Seite 319
05
1015202530354045505560
1 2 3 4 5 6 7 8 9 10 11 12 13 14Laktationstag
pcv
Lysi
n (g
/Tag
)
pcv Lysin - Aufnahme pcv Lysin - Bedarf (kalkuliert)
Abbildung 2: Mittlere pcv Lysin-Aufnahme / Bedarf der Sauen (n=14) vom 1. bis 14. Laktationstag
Durch Körpermassenverluste erhält man eine weitgehend konstante Proteinmobilisierung von 150 g je kg Lebendmasse. Bei einem Gehalt an Lysin im Körperprotein von 7,1 g/16 g N und einer Verwertung des mobilisierten Lys für Milchbildung von 85%, liefert 1 kg Körpermassenabbau 9 g Lys in der Milch. Bezogen auf das Futter können je 1 kg Körpermasseabbau 12 g pcv Lys eingespart werden. (GfE, 2006)
05
1015202530354045
1 2 3 4 5 6 7 8 9 10 11 12 13 14Laktationstag
Ca
(g/T
ag)
Ca - Aufnahme Ca - Bedarf (kalkuliert)
Abbildung 3: Mittlere Ca-Aufnahme / Bedarf der Sauen (n=14) vom 1. bis 14. Laktationstag
Wie aus Abb.2 zu erkennen ist, besteht insbesondere in der ersten Laktationswoche ein deutliches pcv Lysin-Defizit. Dieses kann auch durch den Körpermasseabbau nicht abgedeckt werden. Im Mittel hat-ten die Sauen einen täglichen Körpermasseabbau von 0,650 kg. Somit würde dieses im Mittel lediglich 7,8 g pcv Lys je Tag kompensieren können.
Die Kalkulation des Ca-Bedarfes von Sauen basiert auf die Faktoren für die Ableitung des Nettobedar-fes an Calcium der GfE (2006). - Calcium-Gehalt der Milch von 2,2 g/ kg - unvermeidliche Verluste von 20 mg/ kg Lebendmasse und Tag - Gesamtverwertbarkeit von Calcium von 70 %

Finck et al.: Untersuchung der Energie- und Nährstoffbilanzen in der Vorbereitungs- und Säugephase von Sauen
Seite 320 8. BOKU-Symposium Tierernährung 2009
Das kalkulierte Ca-Defizit wurde ab dem 6. Laktationstag durch die Steigerung der Futteraufnahme und somit Ca-Aufnahme, ausgeglichen. Es ist zu erwarten, dass dieses Defizit durch die Homöostase ausgeglichen werden kann. Abbildung 3 zeigt den Verlauf von Ca-Aufnahme und Ca-Bedarf.
Schlussfolgerung Die errechneten Energiebilanzen, die Reduktion der Rückenfettdicke und des Körpergewichts während der Säugephase zeigen, dass in dieser Phase katabole Prozesse ablaufen. Der Abbau von Körpersub-stanz erfolgt zur Kompensation energetischer Unterversorgung sowie der Nährstoffversorgung. In welcher Höhe und Intensität dieser Abbau erfolgen muss, damit das Krankheitsbild einer Ketose auf-tritt, beziehungsweise Blutparameter diesen Zustand anzeigen, konnte in vorliegender Untersuchung nicht ausreichend ermittelt werden. Eine Indikator-Substanz im Blut und ein dazu gehörender Schwel-lenwert, der eine eindeutige Diagnose einer Ketose bei Sauen zulässt, ist bisher aus der Literatur nicht bekannt. Eine zuverlässige Ermittlung des Energieversorgungsstatus von Zuchtsauen anhand von Blutproben wäre eine Möglichkeit, frühzeitig Maßnahmen zu ergreifen, die das Auftreten klinischer Symptome einer Ketose verhindern. In unserer Untersuchung wurden Nährstoff- und Energiedefizite während der Säugephase festgestellt (kalkuliert), aber es konnten keine Blutparameter herausgestellt werden, die in einer signifikanten Beziehung zu dem ermittelten Defizit stehen. ALSOP et al. (1994) halten es für sehr wahrscheinlich, dass in Schweine haltenden Betrieben Ketose häufig auftritt, aber aufgrund mangelnder, unspezifischer Diagnosen nicht erkannt wird. Ein Blutparameter, der diese Di-agnose ermöglicht wäre hier hilfreich.
Literatur
ALSOP, J. E., HURNIK, D., BILDFELD, R. J. (1994): Porcine ketosis: A case report and literature summary, Swine health and production, Journal, S. 5-8
DRACKLEY, J. K., RICHARD, M. J., BEITZ, D. C., YOUNG, J.W. (1992): METABOLIC CHANGES IN DAIRY COWS WITH KETONEMIA IN RESPONSE TO FEED RESTRICTION AND DIETARY 1,3-BUTANEDIOL, J. DAIRY SCI. 75: 1622-1634
GESELLSCHAFT FÜR ERNÄHRUNGSPHYSIOLOGIE GfE (2006): Empfehlungen zur Energie- und Nährstoffver-sorgung von Schweinen, DLG-Verlag Frankfurt
IBEN, B. und SCHNURRBUSCH, U. (1999): Sauenbesamung: eine praktische Anleitung, Verlag für Agrarwissenschaften und Veterinärmedizin, Aus der Schriftenreihe des Arbeitskreises Großtierpraxis, Dannenberg
NEWCOMB, M. D., HARMON, D. L., NELSSEN, J. L., THULIN, A. J., ALLEE, G. L. (1991): Effect of energy source fed to sows during late gestation on neonatal blood metabolite homeostasis, energy stores and composition, J. Anim Sci. 69: 230-236
REVELL, D. K., WILLIAMS, I. H., MULLAN, B. P., RANFORD, J. L., SMITS, R. J. (1998): Body composition at farrowing and nutrition during lactation affect the performance of primiparous sows: I. Voluntary feed intake, weight loss, and plasma metabolites, J. Anim Sci. 76: 1729-1737
SIEVERDING, E. (2000): Handbuch - Gesunde Schweine, Kamlag Verlag
WELDON, W. C., LEWIS, A. J., LOUIS, G. F., KOVAR, J. L., GIESEMANN, M. A., MILLER, P. S. (1994): Postpartum hypophagia in primipar-ous sows: I. Effects of gestation feeding level on feed intake, feeding behavior, and plasma metabolite concentrations during lactation, J. Anim Sci. 72: 387-394
Autorenanschrift
M. Sc. Elmar Finck Fachbereich 1, Life Scienses, Agrarwirtschaft/Agriculture Fachhochschule Bingen Berlinstrasse 109, 55411 Bingen/Rhein E-Mail: [email protected] Tel.: 06721/409-189

Novák et al.: Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period
8. BOKU-Symposium Tierernährung 2009 Seite 321
Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period
L. Novák, L. Zeman and P. Mares
Mendel University of Agriculture and Forestry Brno, Czech Republic
Introduction
As in the farmers practice so in the research practice the selected breeds of pigs or their hybride com-bination are used. However each group of animals is always composed of individuals. Individual ani-mal in the group have always to be considered as biologic individuals regulated by the genotype. Body mass growth so as growth of their bodies metric dimensions are components of their individual phe-notype. That means tat these growing values do represent expression of genotype formed after syl-lepsis, fertilization of the ova by the spermatozoid. In the arisen new cell embryo, the genom of the ova and the genom of the spermatozoid, do form the genotype that regulates the growth of the started biological individual. The instructions coded in the genotype during the intrauterine develop-ment have formed the autonomous self-regulating system of the body. The basic characteristic of the genotype after the birth expresses in the growth of body mass, one of the phenotype most pro-nounced marks of genotype expression influenced by the environment. The basic form of the body mass growth curve is coded in the genotype and can be considered as specific feature of that particu-lar individual. In the common practice farmers and scientists exhibit the genotype expression by the form and course of the growth curve formed from the average values of the group. In other words the value that depends more on the rules of probability expressed by the chance which individual have been at disposal at formation of the group for fattening or experiment. Therefore according to Schinckel et al (2003) are again searched methods to predict the growth of individual pigs from birth to approximately 60 days of age. The individual growth curves represented in the group do form the resulting average growth curve. The individuality of single pigs are nevertheless lost.
Calculation of the body mass growth curve by the use of the dynamic phenotype method by means of the three biological parameters G0 – the body mass at the origin of the growth curve, GLi – the asymptote limiting the highest value of body mass appropriate the gender, race, line, or hybrid com-bination. The third parameter represented by the highest daily body mass increase dGmax, is located to the age when the body mass growth curve passes the inflection point region. Parameters G0 and GLi can be considered as more dependent on the genotype. The maximum daily body mass increase dGmax reflect in greater measure the modifying effect of the environment on expression of genotype into the phenotype finally expressed by the growth curve development. Diminution of the daily body mass increases (dG/dt) reflects the impact of ominous stressing effect of the environment. This phe-nomenon is based on the distribution of the net energy gained from the food between satisfaction of vital needs and the use for synthesis of proteins, lipids and saccharides retained during the growth in the body. After the unfavorable conditions vanish, the growth curve returns into the corridor represented by the space demarcated at the distance of ± 5 to 10% around the growth curve.
In this presentation we would like to demonstrate that the dynamic phenotype method is able to ex-press the growth curves by means the three biologic parameters G0, GLi and dGmax as in the evalua-tion of the growth in individual pigs so in growth of pigs group

Novák et al.: Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period
Seite 322 8. BOKU-Symposium Tierernährung 2009
Material and methods The first group represents fattening of pigs of both sexes, hybrid combination “bile uslechtile breed x landrase breed”. The experiment was curried in the year 1987 in Pohorelice. The pigs were stabled in standard conditions and fed with the balanced mixture with content of 11,7 MJ/kg in form of the food consumed. The body mass growth and the feed consumption are presented in the Table 1.
Table 1 Body mass growth and feed conversion ration in pigs group “Zeman Simecek Siske Hap Poho-relice 1987” pigs hybrid combination bile uslechtile breed x landrase breed
Body mass Increses Daily feed Feed convAge, days G, kg dG, kg/d intake kg/d kg/kg
28 6,835 7,6 0,31 0,50 1,90942 9,8 0,33 0,60 1,97849 12,1 0,37 0,65 1,75056 14,7 0,40 0,65 2,12563 17,5 0,43 0,85 2,14770 20,5 0,49 0,92 2,30677 23,9 0,51 1,12 2,35384 27,5 0,54 1,21 2,48791 31,3 0,57 1,35 2,62598 35,3 0,60 1,50 2,750105 39,5 0,63 1,65 2,927112 43,9 0,66 1,84 3,043119 48,5 0,66 2,00 3,272126 53,1 0,69 2,15 3,354133 57,9 0,69 2,30 3,500140 62,7 0,71 2,40 3,570147 67,7 0,71 2,55 3,710154 72,7 0,74 2,65 3,729161 77,9 0,74 2,77 3,904168 83,1 0,76 2,90 3,923175 88,4 0,77 2,97 3,954182 93,8 0,76 3,05 4,094189 99,1 0,74 3,10 4,200196 104,3 0,74 3,12 4,213203 109,5 0,73 3,13 4,324210 114,6 0,71 3,15 4,438217 119,6 0,70 3,17 4,557224 124,5 3,19
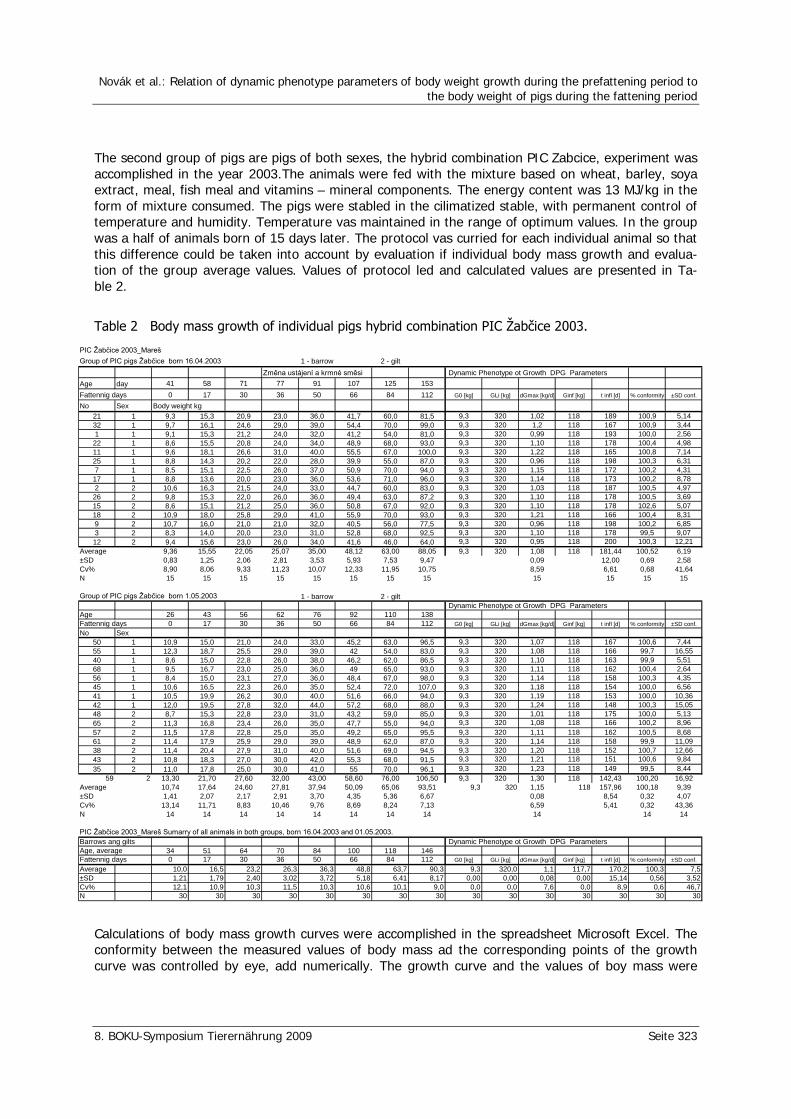
Novák et al.: Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period
8. BOKU-Symposium Tierernährung 2009 Seite 323
The second group of pigs are pigs of both sexes, the hybrid combination PIC Zabcice, experiment was accomplished in the year 2003.The animals were fed with the mixture based on wheat, barley, soya extract, meal, fish meal and vitamins – mineral components. The energy content was 13 MJ/kg in the form of mixture consumed. The pigs were stabled in the cilimatized stable, with permanent control of temperature and humidity. Temperature vas maintained in the range of optimum values. In the group was a half of animals born of 15 days later. The protocol vas curried for each individual animal so that this difference could be taken into account by evaluation if individual body mass growth and evalua-tion of the group average values. Values of protocol led and calculated values are presented in Ta-ble 2.
Table 2 Body mass growth of individual pigs hybrid combination PIC Žabčice 2003.
Calculations of body mass growth curves were accomplished in the spreadsheet Microsoft Excel. The conformity between the measured values of body mass ad the corresponding points of the growth curve was controlled by eye, add numerically. The growth curve and the values of boy mass were
PIC Žabčice 2003_MarešGroup of PIC pigs Žabčice born 16.04.2003 1 - barrow 2 - gilt
Změna ustájení a krmné směsi Dynamic Phenotype ot Growth DPG Parameters Age day 41 58 71 77 91 107 125 153
0 17 30 36 50 66 84 112 G0 [kg] GLi [kg] dGmax [kg/d] Ginf [kg] t infl [d] % conformity ±SD conf.
No Sex Body weight kg21 1 9,3 15,3 20,9 23,0 36,0 41,7 60,0 81,5 9,3 320 1,02 118 189 100,9 5,1432 1 9,7 16,1 24,6 29,0 39,0 54,4 70,0 99,0 9,3 320 1,2 118 167 100,9 3,441 1 9,1 15,3 21,2 24,0 32,0 41,2 54,0 81,0 9,3 320 0,99 118 193 100,0 2,56
22 1 8,6 15,5 20,8 24,0 34,0 48,9 68,0 93,0 9,3 320 1,10 118 178 100,4 4,9811 1 9,6 18,1 26,6 31,0 40,0 55,5 67,0 100,0 9,3 320 1,22 118 165 100,8 7,1425 1 8,8 14,3 20,2 22,0 28,0 39,9 55,0 87,0 9,3 320 0,96 118 198 100,3 6,317 1 8,5 15,1 22,5 26,0 37,0 50,9 70,0 94,0 9,3 320 1,15 118 172 100,2 4,31
17 1 8,8 13,6 20,0 23,0 36,0 53,6 71,0 96,0 9,3 320 1,14 118 173 100,2 8,782 2 10,6 16,3 21,5 24,0 33,0 44,7 60,0 83,0 9,3 320 1,03 118 187 100,5 4,97
26 2 9,8 15,3 22,0 26,0 36,0 49,4 63,0 87,2 9,3 320 1,10 118 178 100,5 3,6915 2 8,6 15,1 21,2 25,0 36,0 50,8 67,0 92,0 9,3 320 1,10 118 178 102,6 5,0718 2 10,9 18,0 25,8 29,0 41,0 55,9 70,0 93,0 9,3 320 1,21 118 166 100,4 8,319 2 10,7 16,0 21,0 21,0 32,0 40,5 56,0 77,5 9,3 320 0,96 118 198 100,2 6,853 2 8,3 14,0 20,0 23,0 31,0 52,8 68,0 92,5 9,3 320 1,10 118 178 99,5 9,0712 2 9,4 15,6 23,0 26,0 34,0 41,6 46,0 64,0 9,3 320 0,95 118 200 100,3 12,21
Average 9,36 15,55 22,05 25,07 35,00 48,12 63,00 88,05 9,3 320 1,08 118 181,44 100,52 6,19±SD 0,83 1,25 2,06 2,81 3,53 5,93 7,53 9,47 0,09 12,00 0,69 2,58Cv% 8,90 8,06 9,33 11,23 10,07 12,33 11,95 10,75 8,59 6,61 0,68 41,64N 15 15 15 15 15 15 15 15 15 15 15 15
Group of PIC pigs Žabčice born 1.05.2003 1 - barrow 2 - gilt Dynamic Phenotype ot Growth DPG Parameters
Age 26 43 56 62 76 92 110 1380 17 30 36 50 66 84 112 G0 [kg] GLi [kg] dGmax [kg/d] Ginf [kg] t infl [d] % conformity ±SD conf.
No Sex50 1 10,9 15,0 21,0 24,0 33,0 45,2 63,0 96,5 9,3 320 1,07 118 167 100,6 7,4455 1 12,3 18,7 25,5 29,0 39,0 42 54,0 83,0 9,3 320 1,08 118 166 99,7 16,5540 1 8,6 15,0 22,8 26,0 38,0 46,2 62,0 86,5 9,3 320 1,10 118 163 99,9 5,5168 1 9,5 16,7 23,0 25,0 36,0 49 65,0 93,0 9,3 320 1,11 118 162 100,4 2,6456 1 8,4 15,0 23,1 27,0 36,0 48,4 67,0 98,0 9,3 320 1,14 118 158 100,3 4,3545 1 10,6 16,5 22,3 26,0 35,0 52,4 72,0 107,0 9,3 320 1,18 118 154 100,0 6,5641 1 10,5 19,9 26,2 30,0 40,0 51,6 66,0 94,0 9,3 320 1,19 118 153 100,0 10,3642 1 12,0 19,5 27,8 32,0 44,0 57,2 68,0 88,0 9,3 320 1,24 118 148 100,3 15,0548 2 8,7 15,3 22,8 23,0 31,0 43,2 59,0 85,0 9,3 320 1,01 118 175 100,0 5,1365 2 11,3 16,8 23,4 26,0 35,0 47,7 55,0 94,0 9,3 320 1,08 118 166 100,2 8,9657 2 11,5 17,8 22,8 25,0 35,0 49,2 65,0 95,5 9,3 320 1,11 118 162 100,5 8,6861 2 11,4 17,9 25,9 29,0 39,0 48,9 62,0 87,0 9,3 320 1,14 118 158 99,9 11,0938 2 11,4 20,4 27,9 31,0 40,0 51,6 69,0 94,5 9,3 320 1,20 118 152 100,7 12,6643 2 10,8 18,3 27,0 30,0 42,0 55,3 68,0 91,5 9,3 320 1,21 118 151 100,6 9,8435 2 11,0 17,8 25,0 30,0 41,0 55 70,0 96,1 9,3 320 1,23 118 149 99,5 8,44
59 2 13,30 21,70 27,60 32,00 43,00 58,60 76,00 106,50 9,3 320 1,30 118 142,43 100,20 16,92Average 10,74 17,64 24,60 27,81 37,94 50,09 65,06 93,51 9,3 320 1,15 118 157,96 100,18 9,39±SD 1,41 2,07 2,17 2,91 3,70 4,35 5,36 6,67 0,08 8,54 0,32 4,07Cv% 13,14 11,71 8,83 10,46 9,76 8,69 8,24 7,13 6,59 5,41 0,32 43,36N 14 14 14 14 14 14 14 14 14 14 14
PIC Žabčice 2003_Mareš Sumarry of all animals in both groups, born 16.04.2003 and 01.05.2003. Barrows ang gilts Dynamic Phenotype ot Growth DPG Parameters Age, average 34 51 64 70 84 100 118 146
0 17 30 36 50 66 84 112 G0 [kg] GLi [kg] dGmax [kg/d] Ginf [kg] t infl [d] % conformity ±SD conf.Average 10,0 16,5 23,2 26,3 36,3 48,8 63,7 90,3 9,3 320,0 1,1 117,7 170,2 100,3 7,5±SD 1,21 1,79 2,40 3,02 3,72 5,18 6,41 8,17 0,00 0,00 0,08 0,00 15,14 0,56 3,52Cv% 12,1 10,9 10,3 11,5 10,3 10,6 10,1 9,0 0,0 0,0 7,6 0,0 8,9 0,6 46,7N 30 30 30 30 30 30 30 30 30 30 30 30 30 30 30
Fattennig days
Fattennig days
Fattennig days

Novák et al.: Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period
Seite 324 8. BOKU-Symposium Tierernährung 2009
presented on the monitor in the Cartesian coordinates arranged according the age of the animals. The growth curves trait was formatted by the biological parameters of the dynamic phenotype (DFG; G0, Dli, dGmax) set in the following equations.
The body mass growth
Gt = Gli . exp(-ln(Gli/G0). exp((-e.dGmax/Gli).(t-t0))) [kg] (1)
Velocity of the body mass growth, the daily body mass increase
dG/dt = (e.dGmax.ln(Gli)/Gli) – (e.dGmax /Gli.Gt.lnGt)) [kg] (2)
Conformity of measured body mass values and d of the corresponding points on the growth curve was numerically evaluated by the formula
%Conformity = 100(Gmeasured / Gt calculated) [%] (3)
It is evident that the mathematic coefficients of anabolism and catabolism are expressed directly by the parameters of the dynamic phenotype of the growth curve (DFG; G0, Gli, dGmax). The assess-ment of the conformity between the measured values and the corresponding points of the growth curve were assessed by eye ad numerically by the formula of %Conformity. The conformity is the best if course of the growth curve formatted by the parameters of dynamic phenotype evokes in the aver-age value of %Conformity index as close as possible to 100%. At that moment, the parameters of dynamic phenotype indicate the right values of dynamic phenotype of the evaluated individual. The estimated parameter values are set in the table under the graph and represent the individuality joined with the individual’s phenotype.
Results and discussion The curves of dynamic phenotype body mass growth on the figure 1 indicate the difference in the fattening the pig in offset of 16 year. It reflex the differences in standards of the feed used, the quali-ty of stabling and the used hybrid combination of pigs. The phenotype growth curves however optical-ly exhibit that the measured averages of body mass grows as in the group Zeman Mares 2003 so in the group Zeman Simecek et all 1987 are distributed in the narrow corridor around the calculated curves. The width of the corridor is about 5% and in the graphs do the perpendicular lines mark corri-dor width. Corresponding parameters of dynamic phenotypes are shown for each experimental group in the table below the graph. Special attention deserves the graph on the right side of figure 1. The velocity of the body mass growth reflects the effect of the actual environment on the on the pheno-type value. The calculated values are distributed in the wider corridor around the parabolic curves. Also these parabolic curves are calculated according the equation (2) using the same parameters as the equation (1) used for calculation of body mass growth. The difference between the curve of the group Zeman Simecek 1987 and the group Zeman Mares 2003 exhibits the difference joined with the results in the breeding work together with the new breeding technologies, nutrition and the mainten-ance care of animals.
Estimation of phenotype growth curves in individual pigs and their comparison with the measured body mass for the hybrid combination PIC Zabcice declared as group Zeman Mares 2003 are pre-sented in the Table 2 and the Figure 2. The difference in the description of individuality of single pigs by the parameters of dynamic phenotype shown in the right part of the Table 2 indicate clearly that dGmax parameter at age when the inflexion point of the growth curve is reached are interesting indi-vidual markers. The relation of the experimental body weight to the body growth curve is shown in three barrows and three gilts shown in Figure 2. The presented graphs indicate the relation between

Novák et al.: Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period
8. BOKU-Symposium Tierernährung 2009 Seite 325
the steepness of the body mass growth curves and the dGmax parameter of dynamic phenotype as shown in the tables below the graphs. The distribution of measured body weights in the corridor around the growth is similar as in the relation of average body mass values to the dynamic phenotype values shown in the Figure 1. In the graphs of individual pigs are illustrated the differences between the daily body mass increases and the average body mass increase calculated as average for the whole fattening period. This value is in the graphs marked by the horizontal lines. All these indicators are calculated for each single pig and are formed by the three parameters of dynamic phenotype (DFG; G0, Gli, dGmax).
Figure 1 Differences in body mass growth of pigs hybrid combination “bile uslechtile x landrase” and the body mass growth of pigs hybrid combination PIC Žabčice curried in experiments with delay of 16 year.
Calculation of the body mass growth curve by the method of dynamic phenotype parameters on con-trary to the statistic methods currently used for modeling the body mass growth curve needs only the three parameters. The DFG parameter method uses only directly measurable parameters. It is inter-esting to note, that the real growth values are distributed in the relatively narrow corridor build around the calculated growth curve. The results are compatible with individual characteristic of single pigs. The fact that it is possible to express the trail of the body mass growth by three biological values for the interval from birth to the sexual and physical maturity deserves attention for its possible rela-tion between the descriptions of phenotype in its relation to genotype’s expression under the actual conditions of the environment.
DPG G0 GLi dGmax Ginfl t infl DPG G0 GLi dGmax Ginfl t inflZeman Šimeček 1987 2 220 0,76 80,9 164,8 Zeman Šimeček 1987 2 220 0,76 80,9 164,8Zeman Mareš 2003 3 260 1,05 118 142 Zeman Mareš 2003 3 260 1,05 118 142
Body mass growth of fattened pigs
0
10
20
30
40
50
60
70
80
90
100
110
120
130
140
150
0 20 40 60 80 100 120 140 160 180 200
Age [d]
Bod
y m
ass
[kg]
dG max 0,76 G DFG Zeman Šimeček 1987 G kgdG max 1,05 G DFG Zeman Mareš 2003
Velocity of body mass growth dG/dt in fattened pigs
0,00
0,19
0,39
0,58
0,78
0,97
1,17
0 20 40 60 80 100 120 140 160 180 200
Age [d]
dG/d
t [k
gd]
dGmax 0,76 dGmax 1,05dG/dt Zeman Šimeček 1987 dG/dt Zeman Mareš 2003

Novák et al.: Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period
Seite 326 8. BOKU-Symposium Tierernährung 2009
G0, kg GLi, kg dGmaxkg/d Ginfl, kg t infl days G0, kg GLi, kg dGmaxkg/d Ginfl, kg t infl days9,1 320 0,970 117,7 154,1 9,4 320 1,040 117,7 142,9
G0, kg GLi, kg dGmaxkg/d Ginfl, kg t infl days G0, kg GLi, kg dGmaxkg/d Ginfl, kg t infl days8,6 320 1,150 117,7 131,6 9,8 320 1,100 117,7 133,6
Slow growing barrow No. 1-1 (data Zeman Mareš, calculations Novák)
0,000,100,200,300,400,500,600,700,800,901,001,101,201,301,401,501,601,701,801,902,00
0 28 56 84 112 140 168 196 224 252 280
Age [d]
Gro
wth
vel
ocity
[kg/
day]
0102030405060708090100110120130140150160170180190200
Bod
y w
eigh
tt [k
g]
dG/dt gomp 1 Gain in weight Gain in weight
DFGgomp Inflexion G* [kg] Body weight
Average growing barrow No. 22-1 (data Zeman Mareš, calculations Novák)
0,000,100,200,300,400,500,600,700,800,901,001,101,201,301,401,501,601,701,801,902,00
0 28 56 84 112 140 168 196 224 252 280
Age [d]
Gro
wth
vel
ocity
[kg/
d]
0102030405060708090100110120130140150160170180190200
Bod
y w
eigh
tt [k
g]
dG/dt gomp 22 Gain in weight Growth velocity
DFGgomp Body weight Inflexion G* [kg]
Fast growing barrow No. 11-1 (data Zeman Mareš, calculations Novák)
0,000,100,200,300,400,500,600,700,800,901,001,101,201,301,401,501,601,701,801,902,00
0 28 56 84 112 140 168 196 224 252 280
Age [d]
Gro
wth
vel
ocity
[kg/
day]
0102030405060708090100110120130140150160170180190200
Bod
y w
eigh
tt [k
g]
dG/dt gomp Gain in weight Growth velocity
DFGgomp Inflexion G* [kg] Body weight
Fast growing gilt No. 18-2 (data Zeman Mareš, calculations Novák)
0,000,100,200,300,400,500,600,700,800,901,001,101,201,301,401,501,601,701,801,902,00
0 28 56 84 112 140 168 196 224 252 280
Age [d]
Gro
wth
vel
ocity
[kg/
d]
0102030405060708090100110120130140150160170180190200
Bod
y w
eigh
tt [k
g]
dG/dt gomp Gain in weight Growth velocity
DFGgomp Inflexion G* [kg] Body weight
Slow growing gilt No. 12-2 (data Zeman Mareš, calculations Novák)
0,000,100,200,300,400,500,600,700,800,901,001,101,201,301,401,501,601,701,801,902,00
0 28 56 84 112 140 168 196 224 252 280
Age [d]
Gro
wth
vel
ocity
[kg/
d]
0102030405060708090100110120130140150160170180190200
Bod
y w
eigh
tt [k
g]
dG/dt gomp Gain in weight Growth velocity
Inflexion G* [kg] DFGgomp Body weight
Average growing gilt No. 26-2 (data Zeman Mareš, calculations Novák)
0,000,100,200,300,400,500,600,700,800,901,001,101,201,301,401,501,601,701,801,902,00
0 28 56 84 112 140 168 196 224 252 280
Age [d]
Gro
wth
vel
ocity
[kg/
d]
0102030405060708090100110120130140150160170180190200
Bod
y w
eigh
tt [k
g]
dG/dt gomp Gain in weight Body weight
Inflexion G* [kg] DFGgomp Body weight
Figure 2: Growth curves and velocity curves of individual pigs PIC Zabcice 2003. Coincidence of calculated growth curves with measured values. Differences between slow growing medium growing and fast growing barrows and gilts

Novák et al.: Relation of dynamic phenotype parameters of body weight growth during the prefattening period to the body weight of pigs during the fattening period
8. BOKU-Symposium Tierernährung 2009 Seite 327
Conclusion On calculated growth curves of dynamic phenotype used for interpretation of body mass growth curves was confirmed the possible use of dynamic phenotype to the assessment of individual differ-ences in the phenotype expression of the genotype. Phenotype growth curve is not dependent on the regular age intervals of body mass measurement. It is remarkable that estimating the parameters of dynamic phenotype at the begin of the growth curve we can forecast the probable course of the indi-viduals growth during the fattening at least to the age when the growth of pig reaches the inflexion point of the growth curve.
Acknowledgement
Research project was supported by grant of Ministry of Agriculture Czech Republic under notification No. NAZV QG 60118
References
Novak, L. 2003:The growth of farm animals simulated by the biologic model of growth.Tagungsband 2. BOKU-Symposium Tierernährung, 02. October 2003, Wien. pp. 87-89.
Novák, L. Zeman, L., Novák, P., Mareš, P.: 2004 Homogenita fenotypu vykrmovaných prasat a metodika jejího stanovení Acta univ. agric. et silvic. Mendel. Brun. 2004, LII, No. 2, 53-64. ISSN 1211 – 8516 (Homogenity of phenotype in fattened pigs and he method of its estimation, in Czech, abstract in English)
Novák L. Zeman L. Novák P.:2006 Simulation of body mass growth of livestock using the method of the phenotype values Tagungsband: 5. Boku-Symposium Tierernährung. Qualitätsmindernde Futterinhaltstoffe. 02. November- 2006, pp. 174-178. ISBN-10: 900962-66-9; ISBN-13: 978-3900962-66-1.
Novák L. Zeman L. and Mareš P. 2008: The velocity of body weight growth and dynamic phenotype expression in pigs. Ta-gungsband 7.BOKU-Symposium Tierernährung 4. Dezember 2008 Wien 355 –362.
Schinckel A.P., J. Ferrell, M.E. Einstein, S.A. Pearce, and R.D. Boyd 2003: Analysis of Pig Growth From Birth to Sixty Days of Age 2003 Swine Research Report Purdue University 57
Corresponding author
Prof. Ing. Ladislav Zeman, CSc Mendel University of Agriculture and Forestry Brno Department of Animal Nutrition and Forage Production Zemedelska street No. 1 613 00 BRNO phone: 00420545133160 fax: 00420545133199 E-mail: [email protected]

Trautwein et al.: Einsatz einer Trocken-TMR in der Mastrinderfütterung
Seite 328 8. BOKU-Symposium Tierernährung 2009
Einsatz einer Trocken-TMR in der Mastrinderfütterung
Julia Trautwein1, Karl Landfried2, Beate Hlawitschka2 und Georg Dusel1 1 Fachhochschule Bingen, Fachbereich 1 - Life Sciences – Agrarwirtschaft, Bingen 2 Lehr- und Versuchsanstalt für Viehhaltung Hofgut Neumühle, Münchweiler/Alsenz
Einleitung In den letzten Jahren wurde in vielen landwirtschaftlichen Betrieben durch die Investition in Biogas-anlagen ein neuer Betriebszweig eingeführt. In den meisten Biogasanlagen dienen Mais- bzw. Grassi-lage als Basis-Substrate für die Fermentation. Dadurch entsteht in diesen Betrieben oftmals eine Kon-kurrenz zwischen den Betriebszweigen Bullenmast und Biogasanlagen um das Substrat Mais bzw. Gras. Des Weiteren haben sich in der BRD mit der Einführung der Betriebsprämie ab 2005 und der damit verbundenen Entkopplung der Direktzahlungen die Rahmenbedingungen für das Produktions-verfahren „Bullenmast mit Stroh-Kraftfutter-Ration“ wesentlich verändert. Durch die entkoppelte Be-triebsprämie hat sich die relative Vorzüglichkeit der Bullenmast dieser „Trocken-TMR“ im Vergleich zur Mast mit Silomais verbessert. Vorteilhaft ist zudem die gute Lagerstabilität, wodurch die Trocken-TMR auf Vorrat angemischt werden kann.
Neben den Fragen der Futter- und Futterproduktionskosten sind tiergesundheitliche und pansenphy-siologische Aspekte bei diesem Fütterungsregime ebenso von großem Interesse wie die Entwicklung der zootechnischen Leistungsparameter.
In zwei Fütterungsversuchen mit Mastbullen der Rasse Fleckvieh wurde das Fütterungsregime einer Trocken-TMR auf Stroh/Getreide-Basis der klassischen Silage/Kraftfutter-Mast in einem Gewichtsbe-reich von 280 bis ca. 650kg LM gegenübergestellt. Neben den zootechnischen Leistungsparametern Futteraufnahme, Lebendmassezuwachs und Futteraufwand wurden auch pansenphysiologische Para-meter sowie Stoffwechselparameter erfasst. Als weiteres Untersuchungs-kriterium wurden zwei ver-schieden Methoden der Pansensaftgewinnung (Schlundsonde und Pansenpunktion von der Hunger-grube aus) auf ihre Praxistauglichkeit hin überprüft.
Material und Methoden
Tiermaterial und Aufstallung
Insgesamt wurden 60 Fleckviehbullen (30 Tiere je Versuchsdurchgang) gleichmäßig auf zwei Fütte-rungsgruppen aufgeteilt. Das mittlere Anfangsgewicht der Trocken-TMR-Gruppen lag bei 284 kg LM, das der Kontrollgruppe Versuch I bei 288 kg sowie 285 kg in Versuch II. Die Tiere wurden auf Voll-spalten in jeweils drei 4-er Gruppen und einer 3-er Gruppe aufgestallt.
Versuchsdesign
Die Versuche erstreckten sich über etwa ca. 300 Masttage. Die beiden Versuchsvarianten in den Durchgängen I und II unterschieden sich lediglich im Hinblick auf ihre Versuchsdiät (siehe Tab. 1). Die Tiere der Versuchsgruppe erhielten ca. 2-2,5 kg Stroh und Heu, ca. 500 g Melasse je Tier und Tag sowie Kraftfutter an Hand einer Futterkurve (vgl. Abb. 1). Diese Komponenten wurden im Futtermi-scher vermengt und den Tieren ad libitum vorgelegt. Während in Versuch I die Kraftfuttermenge in der Versuchsgruppe „moderat“ gesteigert wurde, erfolgte im 2. Versuch eine intensivere Kraftfutterzu-

Trautwein et al.: Einsatz einer Trocken-TMR in der Mastrinderfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 329
teilung (vgl. Abb. 1). Die Kontrollgruppe erhielt Mais- und Grassilage (Verhältnis 1:1) ad libitum sowie 2,0-3,0 kg Kraftfutter je Tier und Tag. Die eingesetzt KF-Mischung war für beide Gruppen identisch.
Tabelle 1: Versuchsdesign (Versuch I und II)
Trocken-TMR
Maissilage / Grassilage (1:1) + Kraftfutterkonzentrat
ad libitum 2,0-3,0 kg
getrennte Vorlage
Kontrolle Stroh/Heu (VI=1:1/VII=2:1) + Melasse + Kraftfutterkonzentrat
ad libitum bis max. 9 bzw. 11 kg/Tier/Tag
Trocken-TMR (KF über Melasse an Stroh gebunden)
Tabelle 2: Energie- und Proteingehalt der Futtermittel (100% TS)
Mais-/Grassilage Heu Stroh Kraftfutter
XP (g/kg) MJ ME XP (g/kg) MJ ME XP (g/kg) MJ ME XP (g/kg) MJ ME
Versuch I 149 10,0 44 8,6 56 6,8 189 12,0
Versuch II 107 9,1 111 8,4 33 6,6 191 11,9
Abb. 1: Gegenüberstellung der Kraftfutter- und Energieaufnahme pro Tier und Tag in der Tro-cken-TMR-Gruppe beider Versuche
Datenerfassung Leistungsparameter Analysen /Auswertung
Die Analyse der Rohnährstoffe erfolgte nach Weender-Analyse im Labor der FH Bingen. Die MJ ME errechnete sich nach der Mischfutterformel (DLG, 1997) aus den Rohnährstoffgehalten.
Vorperiode

Trautwein et al.: Einsatz einer Trocken-TMR in der Mastrinderfütterung
Seite 330 8. BOKU-Symposium Tierernährung 2009
Die Tiergewichte wurden monatlich tierindividuell erfasst. Die Futteraufnahme errechnete sich durch Ein- und Rückwaage täglich je Bucht. Somit konnte auch der Futteraufwand pro Bucht errechnet wer-den.
An Versuchstag 205 (Versuch I) bei einer mittleren LM von ca. 500 kg wurden 5 repräsentativen Bul-len je Variante Pansensaft sowohl über Schlundsonde als auch durch Rumenozentese sowie Blut ent-nommen. In Versuch II erfolgte die Pansensaftentnahme am 92 Versuchstag bei mittlerer Lebendmas-se der Bullen von 435 kg durch Rumenozentese.
Die statistische Auswertung erfolgte mit einem zweiseitigen T-Test (p<0,05) (Microsoft Excel).
Pansensaftgewinnung
Anlässlich der Pansensaftentnahme im Rahmen des Fütterungsversuchs I, wurde die Punktionstechnik im Vergleich zur Schlundsonde (Typ Select) getestet. Die Tiere wurden dazu linksseitig eine handbreit hinter der letzten Rippe und den lateralen Querfortsätzen rasiert, desinfiziert und lokal anästhesiert. Danach wurde mit einem Skalpell eine Stichinzesion durchgeführt und eine 25cm lange Punktionsna-del in Richtung gegenüber liegendem Ellenbogenhöcker tief eingestochen. Nach Entnehmen des Mandrin konnte ca. 2x20 ml Pansensaft mit einer Spritze aufgesogen werden.
Ergebnisse und Diskussion
Leistungsparameter
VERSUCH I
Die Ergebnisse der ersten 70 Versuchstage zeigen sehr deutlich die Auswirkungen der moderaten Kraftfuttersteigerung im Versuchsdurchgang I. Die verhaltene Anhebung der KF-Menge zu Versuchs-beginn führte zu einer geringen Futter-TM-Aufnahme, obwohl die Menge an Heu und Stroh mit ca. 2,5 kg TM (1:1) je Tier und Tag auch im weiteren Versuchsverlauf konstant blieb. Es war aber nicht mög-lich, durch die geringe KF-Gabe hier eine Steigerung der Strukturfutteraufnahme zu erreichen. So blieb v.a. die Proteinaufnahme der Versuchsgruppe in den ersten 70 Tagen deutlich hinter der Kon-trollgruppe zurück.
Die rasche KF-Steigerung nach dem 70. Tag (VERSUCH I) führte dann zu einer verbesserten Futter-aufnahme sowie LM-Entwicklung. Über den Versuch gesehen konnte aber die Versuchsgruppe das Leistungsdefizit der ersten 70 Tage nicht mehr kompensieren. Der höhere Protein- und Energieauf-wand je kg Lebendmassezuwachs der Trocken-TMR-Gruppe über den Versuchszeitraum hinweg ist auch hier auf die ersten 70 Versuchstage zurückzuführen.
VERSUCH II
Auf Grund der Beobachtung aus Versuchsansatz I wurde im Versuch II eine intensive Kraftfuttergabe ab der Einstallung der Tiere gewählt. Nach kurzer Adaptionsphase wurden bereits in den ersten Ver-suchswochen Kraftfuttermengen von ca. 6 kg pro Tier und Tag eingesetzt. Die intensive KF-Kurve wurde bis zum 80. Versuchstag auf 10 kg / Tier und Tag gesteigert. Somit lag der Einsatz des Kraft-futters in dieser Phase mit ca. 3-4 kg pro Tier und Tag über der Kraftfuttermenge von Versuch I (sie-he Abb. 1). Durch den hohen KF-Einsatz in der ersten Versuchsphase lagen die tägliche Proteinauf-nahmen (1.273 g je Tier) und Energieaufnahme (88,6 MJ ME/Tier) auf sehr hohen Niveau gegenüber der Kontrollvariante (569 g Protein und 72,4 MJ ME). Auch gegenüber des ersten Versuchsdurchgan-ges lagen auf Grund der intensiven Kraftfuttergaben in Versuchsdurchgang II der Trocken-TMR-Fütterung die täglichen Aufnahmen von Protein (+438g) und ME (+17,4 MJ) deutlich höher. Die Kont-rollvariante zeigt in der ersten Versuchsphase zwischen den beiden Versuchen I und II eine vergleich-bare tägliche Energieaufnahme von 77,3 MJ bzw. 72,4 MJ bei einem deutlichen Unterschied in der

Trautwein et al.: Einsatz einer Trocken-TMR in der Mastrinderfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 331
täglichen Proteinaufnahme (-605 g/Tier). Dies ist auf die niedrigen Proteingehalte der Mais-/Grassilage in Versuch II zurückzuführen (Tab. 2). Diese niedrige Proteinaufnahme der Kontrollvariante zeigt sich über die gesamte Mastphase und liegt somit nicht im Bereich der Empfehlungen für die Versorgung von Fleckviehbullen (GfE, 1997). Trotz der Proteinunterversorgung erreichten die Kontrolltiere eine mittlere tägl. Zunahme über die gesamte Mastperiode von 1.389g. Die Trocken-TMR-Tiere lagen mit 1.583g TZ um ca. 200 g signifikant über den täglichen Zunahmen der Kontrollvariante.
Tabelle 3: Mittlere tägl. Aufnahme an Trockenmasse, Protein, Energie sowie Tageszunahme (TZ) und Aufwand an Protein und Energie über Versuchszeiträume
-VERSUCH I-
Kontrolle Trocken-TMR
Parameter I 285-380kg LG
II 380-600kg LG
III 285-660kg LG
I 285-380kg LG
II 380-600kg LG
III 285-660kg LG
AUFNAHME (je Tier und Tag)
kg Futter- TM
7,6* + 0,3 9,2* + 0,4 8,9* + 0,2 7,0* + 0,1 9,9* + 0,1 9,4* + 0,2
g Protein 1174* + 42 1403* + 54 1370* + 31 835* + 14 1448* + 14 1333* + 39
MJ ME 77,3* + 2,8 92,4* + 3,6 97,6* + 2,7 71,2* + 1,2 107,0* + 1,0 106,9* + 2,1
g TZ 1755* + 159 1378 + 165 1463* + 144 985* + 179 1441 + 223 1256* + 162
AUFWAND (pro kg Lebendmasse)
g Protein 673* + 56 1030 + 117 932* + 97 882* + 205 1026 + 161 1079* + 176
MJ ME 44,3* + 3,7 67,8 + 7,7 61,3* + 6,4 75,2* + 17,5 75,8 + 11,9 81,3* + 13,1
-VERSUCH II-
Kontrolle Trocken-TMR
Parameter I 285-440kg LG
II 440-640kg LG
III 285-640kg LG
I 285-440kg LG
II 440-640kg LG
III 285-640kg LG
AUFNAHME (je Tier und Tag)
kg Futter- TM
7,3* + 0,4 8,8* + 0,5 8,6* + 0,4 9,0* + 0,5 11,8* + 0,7 10,8* + 0,5
g Protein 569* + 38 710* + 57 766* + 44 1273* + 67 1667* + 101 1520* + 68
MJ ME 72,4* + 3,3 87,5* + 4,8 85,0* + 3,7 88,6* + 4,6 116,2* + 4,7 106* + 5,0
g TZ 1409* + 128 1376 + 198 1389* + 129 1875* + 222 1393 + 188 1583* + 161
AUFWAND (pro kg Lebendmasse)
g Protein 413* + 30 574 + 38 452* + 20 756* + 84 1208 + 97 931* + 73
MJ ME 52,5 + 3,0 70,8 + 6,2 56,3* + 3,5 52,6 + 5,8 84,6 + 6,8 65,2* + 5,1
Pansenphysiologische Parameter und Technik der Pansensaftgewinnung

Trautwein et al.: Einsatz einer Trocken-TMR in der Mastrinderfütterung
Seite 332 8. BOKU-Symposium Tierernährung 2009
Abb. 2: Einfluss der Versuchsdiät auf den pH-Wert des Pansensaftes
Abb. 4: Einfluss der Versuchsdiät auf die Säurekennzahlen des Pansensaftes
In Abb. 2/3 sind die Ergebnisse der Pansensaftuntersuchung im Hinblick auf die Fütterungsstrategie sowie der Pansensaftentnahmetechnik dargestellt.
Der Pansen-pH-Wert der Stroh-KF-Gruppe lag bei beiden Versuchen signifikant um ca. 0,5 pH-Einheiten unter dem pH-Wert der Kontrollgruppe.
Ebenso ergab sich ein signifikanter Unterschied (p<0,037) zwischen den Entnahmetechniken Schlund-sonde und Rumenozentese. Hier wurde in Versuch I unabhängig von der Fütterungsstrategie ein pH-Wert nach Sondenentnahme von 7,1 ermittelt, nach Pansensaftentnahme mittels Rumenozentese von 6,7. Die Differenz von 0,4 pH-Wert-Einheiten deckt sich weitgehend mit den Ergebnissen von STRABEL
Abb. 3: pH-Werte in Abhängig-keit von der Pansensaft-entnahmetechnik

Trautwein et al.: Einsatz einer Trocken-TMR in der Mastrinderfütterung
8. BOKU-Symposium Tierernährung 2009 Seite 333
et al. 2007, die eine Differenz von 0,5 pH-Einheiten feststellten. Da bei der Entnahme von Pansensaft über die Schlundsonde unvermeidlich Speichel mit eingebracht wird kommt es zu einer Erhöhung des Pansen-pH-Wertes. Dies kann bei der Gewinnung von Pansensaft durch Punktion des Pansens ver-mieden werden. Dieses Vorgehen ist in der internationalen Veterinär-Praxis als Diagnostikmethode verbreitet, birgt aber die Gefahr pathologischer Veränderungen. STRABEL et al. 2007 lehnten nach ihrer Untersuchung der Methode das Vorgehen auf Grund von gesundheitlichen Risiken zur Diagnostik ab. Es wurde in dieser Studie die Methode nach NORDLUND et al. 1994 angewandt. Auf Grund heftiger Ab-wehrreaktion seitens der Tiere sowie schmerzhafte Punktionsstellen bis hin zu Fieber wurde diese Methode zur Diagnostik nicht empfohlen. Die hier angewandte Technik der Pansenpunktion verlief komplikationslos. Die Tiere zeigten kaum Abwehrreaktionen (im Gegensatz dazu zeigten die Tiere bei Einführen der Schlundsonde starke Gegenwehr) und der Eingriff konnte nach ca. 2-3 Minuten beendet werden. Die Tiere wurden in den nachfolgenden Tagen verstärkt beobachten und die Körpertempera-tur wurde kontrolliert. Bei keinem Tier zeigten sich Auffälligkeiten im Gesundheitszustand und die Einstichstellen verheilten ohne Komplikationen. Auch im weiteren Versuchsverlauf zeigten sich hin-sichtlich der Futteraufnahme sowie der Gewichtsentwicklung keine Auffälligkeiten der punktierten Tiere. Unterschiede im Hinblick auf die Entnahmetechnik bei den Säurekennzahlen waren erwartungs-gemäß nicht abzusichern.
Bezogen auf die beiden Fütterungsstrategien konnten signifikante Unterschiede in der Buttersäure-konzentration ermittelt werden. Eine Erhöhung der Propionsäure durch den erhöhten KF-Einsatz in der Versuchsgruppe konnte in Versuch I nicht eindeutig nachgewiesen werden. Dagegen zeigen sich signi-fikant höhere Konzentrationen von Essigsäure, Buttersäure und Propionsäure auf Grund der intensiven Kraftfuttergabe in Versuch II. Insbesondere die hohen Gehalte von Essigsäure (135 mmol/l) in der Trocken-TMR in Versuch II sind zu bemerken. Dies könnte durch eine hohe Aufnahme von strukturier-tem Raufutter (ca. 2,5 kg Stroh/Heu-Gemisch) in dieser Versuchsphase zu begründen sein. Insgesamt befinden sich die Säurekonzentrationen über den Versuchen im physiologischen Bereich (Abb. 4)
Weitere Untersuchungsparameter
Tabelle 4: Ausgewählte Blutparameter (n=5 Tiere / Behandlung), * = p < 0,05
Eiweiß
(g/l)
Harnstoff (mmol/l)
Cholesterin (mmol/l)
Bilirubin (µmol/l)
ß-HBS (mmol/l)
NEFA (mmol/l)
VERSUCH I
Kontrolle 71,5 + 3,4 3,8 + 0,6 2,2 + 0,38 1,4* + 0,22 0,30 + 0,10 0,22 + 0,11
Tr.-TMR 69,5 + 3,6 4,3 + 0,8 1,8 + 0,28 0,7* + 0,22 0,44 + 0,13 0,18 + 0,08
VERSUCH II
Kontrolle 63,1 + 3,1 2,4* + 0,3 2,1 + 0,23 1,3 + 0,40 0,66 + 0,13 0,14 + 0,06
Tr.-TMR 65,8 + 2,0 4,7* + 0,9 2,1 + 0,34 0,9 + 0,38 0,62 + 0,13 0,12 + 0,05
Alle untersuchten Blutparameter lagen im physiologischen Bereich. Die signifikanten Unterschiede im Harnstoffgehalt der Kontroll- und Trocken-TMR-Variante (Versuch II) sind auf die Proteinunter- bzw. Proteinüberversorgung der beiden Varianten zurückzuführen. Eine Leberbelastung wie sie in der Stu-die von LÖHNERT et al., 2004 mit ad lib. KF-Aufnahme beobachtet wurde war nicht feststellbar.
In der Schlachtkörperauswertung konnten keine Behandlungsunterschiede festgehalten werden.

Trautwein et al.: Einsatz einer Trocken-TMR in der Mastrinderfütterung
Seite 334 8. BOKU-Symposium Tierernährung 2009
Die Ausschlachtung lag bei 54,1% in der Versuchsgruppe und 53,7% in der Kontrollgruppe in Versuch I und bei 54,6% bei beiden Gruppen in Versuch II. Hinsichtlich Klassifizierung und daraus resultieren-der monetärer Bewertung ergaben sich ebenfalls keine Unterschiede.
Schlussfolgerung In zwei Bullenmastversuchen mit insgesamt 60 Fleckviehbullen im Gewichtsbereich zwischen 285kg LM bis zum Mastende wurde der Einsatz einer Trocken-TMR (Stroh/Heu über Melasse an Kraftfutter gebunden) der konventionellen Fütterungsstrategie mit Mais-/Grassilage und rationiertem KF gegen-übergestellt. Neben der Erfassung der zootechnischen Parameter wurden Untersuchungen zum pan-senphysiologischen Status sowie der Stoffwechsellage durchgeführt.
Es zeigte sich, dass durch intensive Kraftfuttergaben eine Verbesserung der zootechnischen Leis-tungsparameter zu realisieren war. Eine Anpassung der Proteinversorgung in den verschiedenen Mastphasen wäre durch die Staffelung von Proteingehalten im Kraftfutter zu empfehlen. Die Untersu-chungen der Pansen- und Blutparameter ergaben trotz hoher Kraftfuttergaben von bis zu 11 kg pro Tier und Tag in der Trocken-TMR-Variante keine kritische pansenphysiologische Werte (pH-Werte, Fettsäurenkonzentrationen).
Neben der Versuchsfrage der Fütterungsstrategie wurde die Rumenozentesetechnik von der linken Hungergrube aus zur Pansensaftgewinnung auf ihre Praxistauglichkeit überprüft. Es wurden 10 Tiere auf diese Weise Pansensaft entnommen. Die Technik zeigte sich als komplikationslos im Hinblick auf die Durchführung, die Duldung durch die Tiere sowie den Heilungsprozess. Gegenüber dem mittels Schlundsonde gezogenen Pansensaft wurde ein um 0,4 Einheiten niedrigerer pH-Wert gemessen (p<0,037).
Literatur
1. DLG, 1997: DLG-Futterwerttabelle für Wiederkäuer, DLG-Verlag, 7., erw. und überarb. Auflage, Frankfurt am Main,
2. LÖHNERT, H.-J., W.I. OCHRIMENKO, S. DUNKEL, H. LÜDKE, 2004: Untersuchungen zu Stroh-Konzentrat-Rationen in der Mastrinderfütterung, Tagungsband Forum angew. Forschung 2004, 25-28
3. NORDLUND, K.V. AND E.F. GARRET, 1994: Rumenocentesis: A technique for the diagnosis of subacute rumen acidosis in dairy herds. Bovine Pract. 28:109–112
4. STRABEL, D., A. EWY, T. KAUFMANN, A. STEINER, M. KIRCHHOFER, 2007: Rumenozentese: Eine geiignete Methode zur pH-Bestimmung im Pansensaft? Schweiz. Arch. Tierheilk. 149, Heft 7, Juli 2007, 301-307
Autorenanschrift
M. Sc. Julia Trautwein Fachhochschule Bingen/University of Applied Sciences Fachbereich 1 – Life Sciences and Engineering Berlinstrasse 109, D- 55411 Bingen, Germany E-Mail: [email protected] Tel. +49 6721 409 189

Autorenverzeichnis
8. BOKU-Symposium Tierernährung 2009 Seite 335
Autorenverzeichnis
Ahn, B.K. ................................................. 165
Aigner, S. ................................................. 287
Applegate, T.J. .................................. 174, 303
Asamer, A. ........................................ 153, 157
Asmenskaite, L. ........................................ 208
Balabánová, M. ................................. 264, 308
Bartelt, J. ................................................. 250
Baulez, M. ............................................ 51, 54
Boulianne, M. ............................................. 54
Bourdonnais, A. .......................................... 57
Braunsberger, F. ...................................... 219
Bravo, D. .......................................... 129, 148
Brestenský, M. ........................................... 94
Brož, J. ...................................................... 94
Bruininx, E. .............................................. 116
Brunauer, C. ............................................ 268
Čerešňáková, Z. ......................................... 62
Chrenková, M. ............................................ 62
Cypriano, L. ............................................. 157
der Kinderen, L. ....................................... 116
Doležal, P. ................................................. 66
Dombazyan, V. ......................................... 292
Domig, K.J. ....................................... 287, 292
Durosoy, S. ........................................ 70, 116
Dusel, G. .................................. 212, 314, 328
Erben, R.G. .................................................. 1
Ettle, T. ............................................ 255, 259
Filho, J.B.P. .............................................. 157
Finck, E. .................................................. 314
Flachowsky, G. ........................................... 28
Fľak, P. ...................................................... 62
Flocke, F. ................................................. 102
Franke, K. ................................................. 28
Fry, R.S. .................................................... 70
Gallnböck, M. ........................................... 239
Gärtner, S. ............................................... 160
Germ, T. .................................................. 188
Gövert, J. ................................................. 102
Haese, E. .................................................... 5
Hagmüller, W. .................................. 239, 292
Hammerl, S. .............................................. 76
Heger, J. ................................................... 94
Heseker, A. .............................................. 102
Hlawitschka, B. ......................................... 328
Hong, J.-C. ............................................... 179
Hopp, J. ................................................... 102
Hošek, M. ................................................. 264
Hošková, Š. ...................................... 264, 308
Iben, C. .................................................... 121
Jancikova, P. ............................................ 137
Jankowski, J. ............................................. 82
Jerešiūnas, A. ........................................... 184
Jerešiūnienė, O. ........................................ 184
Jeroch, H. ................................................. 82
Kang, C.W. ............................................... 165
Kern, C. ................................................... 297
Kickinger, T. .............................................. 37
Kienzle, E. ................................................. 24
Kim, J.S. .................................................. 165
Klaßen, M. ................................................ 314
Klimitsch, A. ............................................. 188
Klose, V. .......................................... 297, 303
Kluge, H. .................................................. 250
Kneifel, W. ....................................... 287, 292

Autorenverzeichnis
Seite 336 8. BOKU-Symposium Tierernährung 2009
Koch, C. ................................................... 232
Kocsis, L. ................................................. 281
Kozłowski, K............................................... 82
Kroismayr, A. ........................................... 219
Kühl, J. .................................................... 232
Kulpys, J. ........................................... 86, 184
Laguna Paredes, C. .................................. 110
Landfried, K. ............................. 232, 314, 328
Li, Q. ......................................................... 76
Liebhard, P. ............................................. 281
Lien, T.-F. ................................................ 179
Liu, J. ...................................................... 212
Lobinski, R. .............................................. 129
Mader, A. .......................................... 153, 157
Maevis, G. ................................................ 232
Mair, C..................................................... 268
Manawadu, A. .......................................... 121
Mandl, J. .................................................. 274
Mareš, P. .................................. 137, 171, 321
Martens, H. ................................................ 11
Matusevicius, P. ....................................... 208
Mayrhofer, S. ........................................... 287
Meyer, U. ................................................... 28
Mul, A. ..................................................... 116
Müller, A. ................................................. 297
Nagel, P. .................................................. 292
Neufeld, K. ............................................... 219
Nitrayová, S. .............................................. 94
Nitsch, S. .......................................... 174, 188
Novák, L. ................................................. 321
Obermaier, A. ................................... 255, 259
Oguey, S. ................................................ 129
Padoan, D. ............................................... 203
Parsons, C.M. ........................................... 148
Pastore, N.S. ............................................ 157
Patráš, P. .................................................. 94
Petersson, A. ............................................ 292
Piccini, I. .................................................. 157
Pietsch, M. ............................................... 212
Poláčiková, M. ........................................... 62
Poštulka, R. ............................................... 66
Rapp, C. ................................................... 102
Reisinger, N. ............................................. 174
Roch, G. .................................................... 54
Rodehutscord, M.......................................... 5
Rohrer, E. ................................................ 194
Romberg, F.-J. .......................................... 232
Roth, N. ........................................... 188, 203
Röttger, A. ................................................ 28
Rutzmoser, K. ................................... 255, 259
Sager, M. ......................................... 110, 114
Sattler, V. A. ............................................. 297
Schatzmayr, G. .......................... 174, 297, 303
Schedle, K. ................................................ 76
Schlegel, P. ........................................ 70, 116
Schreiner, M. ............................................ 268
Seresinhe, T. ............................................ 121
Skládanka, J. ............................................. 66
Spears, J.W. .............................................. 70
Stangl, G. ................................................. 250
Stankevičius, R. ......................................... 86
Steiner, T. ........................................ 174, 179
Steingaß, H. ............................................. 232
Stückler, R. .............................................. 274
Stumpff, F. ................................................ 11
Südekum, K-H. ......................................... 232
Sudikas, G. ............................................... 184
Tichy, A. .................................................. 239

Autorenverzeichnis
8. BOKU-Symposium Tierernährung 2009 Seite 337
Trautwein, J. ..................................... 212, 328
Tschirner, K. ............................................ 165
Ubbink-Blanksma, M. ................................ 116
Urbaityte, R. ..................................... 188, 203
Urbonas, R. ................................................ 86
Utterback, P. ............................................ 148
Vacchina, V. ............................................. 129
van Hoek, A.H.A.M. .................................. 287
Vašátková, A. .................................... 264, 308
Večerek, M. ............................... 137, 171, 308
Vielhaber, B. ............................................ 239
Vikari, A. .................................................. 148
von Rosenberg, S. .................................... 246
Wegl, G. ........................................... 297, 303
Wehr, U. .................................................. 246
Wendler, K.R. ........................................... 153
Werteker, M. ............................................ 114
Wetscherek, W. .......... 142, 194, 226, 274, 281
Wetscherek-Seipelt, G. ....... 142, 194, 226, 274
Windisch, W. ................................ 37, 76, 268
Würzner, H. .......................................... 37, 45
Zeman, L. ............. 66, 137, 171, 264, 308, 321
Zentek, J. ................................................. 160
Zitterl-Eglseer, K. ...................................... 239
Zollitsch, W. ............................................. 292

Sponsoren
Seite 338 8. BOKU-Symposium Tierernährung 2009
Sponsoren
Wir danken folgenden Firmen für ihre großzügige Unterstüt-zung: Alphabetische Reihenfolge
Special Sponsoring
Pancosma S.A. Voie-des-Traz 6 CH-1218 Le Grand-Saconnex
Agentur für Gesundheit und Ernährungssicherheit GmbH Spargelfeldstraße 191 A-1226 Wien
AGRANA Bioethanol GmbH Industriegelände Pischels-dorf A-3435 Pischelsdorf
Agrarmarkt Austria Marketing GesmbH Dresdner Straße 68a A-1200 Wien
agromed Austria GmbH Bad Haller Straße 23 A-4550 Kremsmünster
Albion Animal Nutrition International 7 Rue Jean Macé FR-35000 Rennes
Alltech Deutschland GmbH Wiesenstraße 21 (W21) D-40549 Düsseldorf

Sponsoren
8. BOKU-Symposium Tierernährung 2009 Seite 339
BASF ChemTrade GmbH Industriestraße 20 D-40549 Düsseldorf
BIOMIN GmbH Industriestraße 21 A-3130 Herzogenburg
BOKU-Wien Gregor Mendel-Straße 33 A-1180 Wien
Daem eCo bvba Nerumstraat 7 B-9340 Lede
Delacon Biotechnik Ges.m.b.H. Weissenwolffstraße 14 A-4221 Steyregg
DSM Nutritional Products Ltd P.O. Box 3255 CH-4002 Basel
Dr. Eckel GmbH Im Stiefelfeld 10 D-56651 Niederzissen
Inntaler Mischfutter GmbH & Co. KG Klötzlmüllerstraße 140 D-84034 Landshut

Sponsoren
Seite 340 8. BOKU-Symposium Tierernährung 2009
Lohmann Animal Health GmbH & Co.KG Heinz-Lohmann-Strasse 4 D-27472 Cuxhaven
Metall und Farben Ges.m.b.H. Ottakringer Straße 89 A-1160 Wien
Reisenberger GmbH Stuttgarterstraße 2 A-2380 Pertoldsdorf
JRS J. Rettenmaier & Söhne GmbH+Co.KG Holzmühle 1 D-73494 Rosenberg
H. Willhelm SCHAUMANN GmbH & Co. KG Jakob-Fuchs-Gasse 25-27 A-2345 Brunn am Gebirge
Trouw Nutrition Deutschland GmbH Gempfinger Str. 15 D-86666 Burgheim
Zinpro Animal Nutrition Gerard Doustraat 4a NL-5831 CC Boxmeer




















