
Systematic identification of factors involved in post-transcriptional processes in wheat grain
17
Abstract Post-transcriptional processing of primary transcripts can significantly affect both the quantity and the structure of mature mRNAs and the corre- sponding protein products. It is an important mecha- nism of gene regulation in animals, yeast and plants. Here we have investigated the interactive networks of pre-mRNA processing factors in the developing grain of wheat (Triticum aestivum), one of the world’s major food staples. As a first step we isolated a homologue of the plant specific AtRSZ33 splicing factor, which has been shown to be involved in the early stages of em- bryo development in Arabidopsis. Real-time PCR showed that the wheat gene, designated TaRSZ38, is expressed mainly in young, developing organs (flowers, root, stem), and expression peaks in immature grain. In situ hybridization and immunodetection revealed preferential abundance of TaRSZ38 in mitotically ac- tive tissues of the major storage organ of the grain, the endosperm. The protein encoded by TaRSZ38 was subsequently used as a starting bait in a two-hybrid screen to identify additional factors in grain that are involved in pre-mRNA processing. Most of the iden- tified proteins showed high homology to known splic- ing factors and splicing related proteins, supporting a role for TaRSZ38 in spliceosome formation and 5¢ site selection. Several clones were selected as baits in fur- ther yeast two-hybrid screens. In total, cDNAs for 16 proteins were isolated. Among these proteins, TaRSZ22, TaSRp30, TaU1-70K, and the large and small subunits of TaU2AF, are wheat homologues of known plant splicing factors. Several, additional pro- teins are novel for plants and show homology to known pre-mRNA splicing, splicing related and mRNA export factors from yeast and mammals. Keywords Grain development Pre-mRNA processing factors Protein–protein interaction Splicing Yeast two-hybrid system Abbreviations AD Activation domain BD Binding domain DAP Days after pollination dbEST GenBank expressed sequence tag database GUS b-Glucuronidase NEB Nuclei extraction buffer NLS Nuclear localization signal NMD Nonsense mediated decay RRM RNA recognition motif RSZproteins Arginine/serine proteins with one or two zinc fingers SCL proteins SC35-like proteins SD domains Serine/aspartic acid rich domains snRNPs Small nuclear ribonucleoprotein particles Electronic Supplementary Material Supplementary material is available to authorised users in the online version of this article at http://dx.doi.org/10.1007/s11103-006-9046-6. S. Lopato (&) A. S. Milligan N. Shirley N. Bazanova P. Langridge Australian Centre for Plant Functional Genomics, The University of Adelaide, PMB1, Glen Osmond, SA 5064, Australia e-mail: [email protected] L. Borisjuk Institut fur Pflanzengenetik und Kulturpflanzenforschung, D-06466 Gatersleben, Germany Plant Mol Biol (2006) 62:637–653 DOI 10.1007/s11103-006-9046-6 123 Systematic identification of factors involved in post- transcriptional processes in wheat grain Sergiy Lopato Ljudmilla Borisjuk Andrew S. Milligan Neil Shirley Natalia Bazanova Peter Langridge Received: 8 March 2006 / Accepted: 6 July 2006 / Published online: 29 August 2006 ȑ Springer Science+Business Media B.V. 2006
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Systematic identification of factors involved in post-transcriptional processes in wheat grain

Abstract Post-transcriptional processing of primary
transcripts can significantly affect both the quantity
and the structure of mature mRNAs and the corre-
sponding protein products. It is an important mecha-
nism of gene regulation in animals, yeast and plants.
Here we have investigated the interactive networks of
pre-mRNA processing factors in the developing grain
of wheat (Triticum aestivum), one of the world’s major
food staples. As a first step we isolated a homologue of
the plant specific AtRSZ33 splicing factor, which has
been shown to be involved in the early stages of em-
bryo development in Arabidopsis. Real-time PCR
showed that the wheat gene, designated TaRSZ38, is
expressed mainly in young, developing organs (flowers,
root, stem), and expression peaks in immature grain. In
situ hybridization and immunodetection revealed
preferential abundance of TaRSZ38 in mitotically ac-
tive tissues of the major storage organ of the grain, the
endosperm. The protein encoded by TaRSZ38 was
subsequently used as a starting bait in a two-hybrid
screen to identify additional factors in grain that are
involved in pre-mRNA processing. Most of the iden-
tified proteins showed high homology to known splic-
ing factors and splicing related proteins, supporting a
role for TaRSZ38 in spliceosome formation and 5¢ site
selection. Several clones were selected as baits in fur-
ther yeast two-hybrid screens. In total, cDNAs for 16
proteins were isolated. Among these proteins,
TaRSZ22, TaSRp30, TaU1-70K, and the large and
small subunits of TaU2AF, are wheat homologues of
known plant splicing factors. Several, additional pro-
teins are novel for plants and show homology to known
pre-mRNA splicing, splicing related and mRNA
export factors from yeast and mammals.
Keywords Grain development Æ Pre-mRNA
processing factors Æ Protein–protein interaction ÆSplicing Æ Yeast two-hybrid system
Abbreviations
AD Activation domain
BD Binding domain
DAP Days after pollination
dbEST GenBank expressed sequence tag
database
GUS b-Glucuronidase
NEB Nuclei extraction buffer
NLS Nuclear localization signal
NMD Nonsense mediated decay
RRM RNA recognition motif
RSZproteins Arginine/serine proteins with one or
two zinc fingers
SCL proteins SC35-like proteins
SD domains Serine/aspartic acid rich domains
snRNPs Small nuclear ribonucleoprotein
particles
Electronic Supplementary Material Supplementary materialis available to authorised users in the online version of thisarticle at http://dx.doi.org/10.1007/s11103-006-9046-6.
S. Lopato (&) Æ A. S. Milligan Æ N. Shirley ÆN. Bazanova Æ P. LangridgeAustralian Centre for Plant Functional Genomics,The University of Adelaide, PMB1, Glen Osmond,SA 5064, Australiae-mail: [email protected]
L. BorisjukInstitut fur Pflanzengenetik und Kulturpflanzenforschung,D-06466 Gatersleben, Germany
Plant Mol Biol (2006) 62:637–653
DOI 10.1007/s11103-006-9046-6
123
Systematic identification of factors involved in post-transcriptional processes in wheat grain
Sergiy Lopato Æ Ljudmilla Borisjuk ÆAndrew S. Milligan Æ Neil Shirley ÆNatalia Bazanova Æ Peter Langridge
Received: 8 March 2006 / Accepted: 6 July 2006 / Published online: 29 August 2006� Springer Science+Business Media B.V. 2006

SR domains Serine/arginine rich domains
TRN-SR Transportin-SR
Introduction
Comparative genome analyses indicate that increases
in gene number do not necessarily correlate with in-
creases in morphological and behavioral complexity
(Ewing and Green 2000). Pre-mRNA processing
(Reddy 2001; Quesada et al. 2003), transport (Lei and
Silver 2002), turnover (Abler and Green 1996) and
translational initiation (Kawaguchi and Bailey-Serres
2002) contribute additional sources of complexity.
Over the last two decades, numerous studies have
shown the importance in many organisms of these post-
transcriptional mechanisms of regulation, particularly
during embryogenesis (Sun et al. 1997; Lopez 1998;
Groth and Lardelli 2002; Kalyna et al. 2003; Lalli et al.
2003). Both the quantity and the structure of mature
mRNAs, and the corresponding protein products, can
be significantly affected by the post-transcriptional
processing of primary transcripts (Barrass and Beggs
2003; Kriventseva et al. 2003). Post-transcriptional
control allows a quicker response to environmental
stimuli compared with de novo transcription and in
some cases permits regulation of gene expression even
in the absence of transcription.
Splicing takes place in a large complex, called the
spliceosome, which comprises five small nuclear ribo-
nucleoprotein particles (snRNPs) and about two hun-
dred non-snRNP proteins (Zhou et al. 2002). Protein
factors essential for pre-mRNA processing in eukary-
otes have been identified and characterized primarily
through the study of nuclear extracts derived from
mammalian cells, Saccharomyces cerevisiae genetics
(Mount and Salz 2000), or through homology search-
ing. They include SR-rich RNA binding proteins
(Blencowe et al. 1999; Reddy 2004), protein kinases
(Kuroyanagi et al. 1998; Wang et al. 1998b; Savaldi-
Goldstein et al. 2003), phosphatases (Murray et al.
1999), RNA helicases (Tuteja 2000), peptidylprolyl cis–
trans-isomerases (Mortillaro and Berezney 1998;
Zhang et al. 2002; Lorkovic et al. 2004) and other
proteins with still unknown function. However, there
remain obstacles to the characterization of pre-mRNA
processing in plants. Despite attempts by several
groups to obtain active plant extracts for splicing in
vitro, a plant in vitro system is still not available, and
genetic approaches are more complex and less efficient
than in yeast or nematodes. Homology searching has
been used to identify RNA recognition motif (RRM)
and KH domain-containing RNA binding proteins in
Arabidopsis (Lorkovic and Barta 2002), but this ap-
proach may overlook factors with low sequence con-
servation, plant specific factors and factors connecting
pre-mRNA processing to other cellular processes. Gi-
ven these limitations, the yeast two-hybrid system
emerges as a useful additional tool for the character-
ization of RNA splicing, allowing the identification of
networks of protein–protein interactions in the
spliceosome, as has been demonstrated for yeast
(Fromont-Racine et al. 1997; Ben-Yehuda et al. 2000),
nematodes (Li et al. 2004), Drosophila (Stanyon and
Finley 2000) and plants (Golovkin and Reddy 1998,
1999; Lopato et al. 2002; Lorkovic et al. 2004). This
approach facilitates not only the cloning of cDNAs
encoding novel factors, but also the assignment of
possible function, inferred through the function of
interacting partners, and the grouping of proteins into
functional protein complexes.
Many splicing factors have been shown to be tissue
specific and are themselves developmentally regulated
both at the transcriptional and post-transcriptional
levels (Fu and Maniatis 1992; Screaton et al. 1995;
Hanamura et al. 1998; Lopato et al. 1999b, 2002; Ka-
lyna et al. 2003; Fang et al. 2004). Thus the spliceo-
some structure can be dynamic and might be specified
in different tissues or even groups of cells, regulating
organism development, by participation of tissue spe-
cific splicing factors. Several animal splicing factors
have been demonstrated to be involved in develop-
ment and, in particular, embryo development (for a
review, see Lopez 1998; Lalli et al. 2003), while gain-
and loss-of-function studies in plants indicate the cru-
cial role of alternative splicing in plant development
(Lopato et al. 1999b; Golovkin and Reddy 2003; Ka-
lyna et al. 2003; Savaldi-Goldstein et al. 2003).
Direct involvement in the early stages of embryo
development has recently been demonstrated for the
plant splicing factor AtRSZ33 (Kalyna et al. 2003).
AtRSZ33 is a nuclear localized plant specific RNA-
binding protein from Arabidopsis which is involved in
splicing, as demonstrated by its interaction with several
known splicing factors in the yeast two-hybrid system
and in pull-down assays (Lopato et al. 2002). Regula-
tion of alternative splicing of several genes has been
demonstrated in transgenic plants ectopically overex-
pressing AtRSZ33 (Kalyna et al. 2003). Moderate
levels of AtRSZ33 overexpression changed the cell
shape, and the plane of cell division in several differ-
entiating tissues became misorientated. The most
striking changes were found during early embryo
development, leading to either seed abortion or to the
638 Plant Mol Biol (2006) 62:637–653
123

appearance of monocotyledonous or twin plants (Ka-
lyna et al. 2003).
In monocotyledonous species, several examples of
alternative splicing regulation have been described
(Nash and Walbot 1992; Montag et al. 1995; Maga-
raggia et al. 1997; Locatelli et al. 2000; Halterman
et al. 2003). However, little is known about the
protein factors involved in pre-mRNA processing in
grasses and, particularly, about the role of such fac-
tors in grain development in crop plants. This
knowledge will provide us with the tools for manip-
ulating aspects of grain development for improved
yield or quality.
Here, the wheat (Triticum aestivum) homologue of
AtRSZ33 is described and shown to be expressed in the
embryo and in the mitotically active part of the syn-
cytial and cellularizing endosperm. This protein was
used as a starting bait to identify factors involved in
pre-mRNA processing in the early grain. A network of
their interactions are described for early wheat grain
using further yeast two-hybrid screens, in which preys
from previous screens were used sequentially as baits.
In addition to homologues of known factors involved in
pre-mRNA processing and transport in plants, animals
and yeast, cDNAs encoding several novel proteins
were identified.
Materials and methods
Plant material
Bread wheat (Triticum aestivum L. cv. Chinese Spring)
plants were grown in glasshouses with day tempera-
tures of 18–25�C and night temperatures of 18–21�C,
with a 13 h photoperiod. Spikes were labeled at
anthesis. Whole immature caryopses were harvested
daily at 0–6 days after pollination (DAP), and pooled
samples of liquid endosperm were collected at 3–7
DAP by pipette aspiration from the embryo sacs of
dissected micropylar tips of the grain.
mRNA Isolation and hybridization techniques
Total RNA was isolated from wheat samples using TRI
REAGENT (Molecular Research Centre, Inc., Cin-
cinnati, OH), and poly(A)+ RNA was then obtained
using an mRNA Direct Kit (Dynal, Norway), and used
for the preparation of MATCHMAKER Two-Hybrid
System 3 (Clontech, Palo Alta, CA) cDNA libraries.
In situ hybridization was performed according to
Weber et al. (1995) using [a-33P] dCTP-labeled cDNA
as a probe. To confirm the specificity of the in situ
hybridization several controls were performed. RNase
treatment before hybridization abolished any labeling
when using the same probe, indicating that the signal is
RNA specific. To check unspecific labeling we used as
a probe a 33P-labelled cDNA fragment encoding a
human methyltransferase gene with no known homol-
ogy to genes in higher plants (Rydberg et al. 1990) and
did not observe any labeling of our plant tissues with
this probe.
Plasmid construction and yeast two-hybrid
screening
The full-length coding region of the TaRSZ38 cDNA
was identified by constructing a contig of ESTs, and
was then amplified by PCR from the syncytial endo-
sperm cDNA library. The PCR fragment was subcl-
oned into the EcoRI–BamHI sites of the plasmid
pGBKT7 (Clontech) and used as a bait to screen the
above MATCHMAKER cDNA libraries. Two preys
of TaRSZ38, TaTra2 and TaTRN-SR were themselves
cloned into pGBKT7, as full-length and N-terminally
truncated sequences, respectively, either by PCR or by
direct subcloning of inserts from the prey construct
pGADT7-Rec. TaTra2 and TaTRN-SR were then used
as baits in subsequent two-hybrid screens of the same
cDNA libraries. Yeast transformation and screening
were performed essentially as described in the Yeast
MATCHMAKER System 3 manual. Putative positive
clones were verified by co-transformation with the bait
plasmid into yeast, and their sequences were analyzed
by HaeIII digestion, to identify redundant clones, and
direct sequencing. Full-length sequences of partial
clones identified in the two-hybrid screen, except
TaDRH1, were obtained using PCR of cDNA pre-
pared from wheat (cv. Chinese Spring) zygotic em-
bryos isolated at 5 DAP. The embryo-derived cDNA
had been synthesized and cloned into the phage vector
kTriplex2, as described in the SMARTTM cDNA Li-
brary Construction Kit (Clontech).
Quantitative PCR analysis
The primer pairs for TaRSZ38, TaPrp38 and
TaRSZBP1 were designed for the wheat variety Chi-
nese Spring using 3¢UTR sequences of cDNAs as a
template. The Q-PCR amplification was performed in
the RG 2000 Rotor-Gene Real Time Thermal Cycler
(Corbett Research, NSW, Australia) using QuantiTect
SYBR Green PCR reagent (Qiagen, VIC, Australia),
as described in Burton et al. (2004). The Rotor-Gene
V4.6 software (Corbett Research) was used to deter-
mine the optimal cycle threshold (CT) from dilution
Plant Mol Biol (2006) 62:637–653 639
123

series, and the mean expression level and standard
deviations for each set of four replicates for each
cDNA were calculated.
Protein expression
A PCR product containing the bacteriophage T7 pro-
moter, an N-terminal c-Myc epitope and the full-length
coding region of TaRSZ38 was used as a template to
express TaRSZ38 in vitro in the TnT Coupled Wheat
Germ Extract System Kit (Promega, NSW, Australia).
Five microliters of the reaction mix were analyzed by
SDS-PAGE, blotted onto nitrocellulose membrane,
and TaRSZ38 was immunodetected either with anti-
c-Myc monoclonal antibodies (Clontech) or with
antibodies against AtRSZ33 (Lopato et al. 2002). Pre-
immune preparations of antibodies were used as neg-
ative controls.
For expression in yeast (Saccharomyces cerevisiae),
the coding region of TaRSZ38 with an N-terminal c-
Myc epitope was subcloned into the pYES2.1 expres-
sional vector using the pYES2.1 TOPO TA Expression
Kit (Invitrogen, VIC, Australia). TaRSZ38 protein was
expressed in the EGY48 yeast strain and visualized by
western blots using an anti-c-Myc antibody (Clontech).
Fractionation of the syncytial endosperm and
immunodetection of RSZ proteins
Liquid endosperm from the caryopses of two spikes
was collected into 0.5 ml nuclei extraction buffer
(NEB) [25 mM Tris–HCl, pH 8.0, 0.44 M sucrose,
5 mM MgCl2, 2.5% Ficoll 400, 5% dextran, 0.5%
Triton X-100, 2 mM spermine, 2.5 mM dithiothreitol,
0.5 mM phenylmethylsulfonyl fluoride, protease
inhibitor cocktail (Sigma, concentration as recom-
mended in manual)]. The nuclei were isolated using a
protocol adapted from Chua et al. (2001). The liquid
part of the endosperm was centrifuged at 500g for
10 min at 4�C. The supernatant was collected and
used as a nuclei-free cytoplasmic fraction. The pellet
(partially purified nuclear fraction) was resuspended
in 20–100 ll NEB (without spermine) by gentle tap-
ping. The suspension of nuclei was loaded on a Per-
coll gradient (0.5 ml 80% and 0.5 ml 30% Percoll)
and centrifuged at 1,000g for 30 min at 4�C. The
intact nuclei were collected from the interface
between the 30 and 80% Percoll solutions. All frac-
tions were stained with 4¢,6-diamidino-2-phenylindole
and assessed by visualization under a fluorescence
phase microscope (Zeiss Axioskop; Carl Zeiss Pty.
Ltd., NSW, Australia) with the Zeiss number 2 filter
set (excitation filter 365 nm, barrier filter 420 nm) to
determine the fraction with the majority of nuclei and
minimum contamination.
The layer of nuclei was carefully removed and 10- to
20-fold diluted with NEB (without spermine). Nuclei
were pelleted by centrifugation at 1,500g at 4�C and
used for SDS-PAGE as a fraction of pure nuclei.
Proteins were loaded onto SDS-PAGE gels on an
equal protein basis, blotted onto nitrocellulose mem-
branes (Hybond C-extra; Amersham Biosciences,
NSW, Australia) and stained with Ponceau S. Wheat
RSZ proteins were immunodetected on western blots
using polyclonal antibodies raised against AtRSZ33
and AtRSZp22 (Lopato et al. 1999a, 2002). Pre-im-
mune antiserum was used as a negative control.
Sequence data from this article have been deposited
with the EMBL/GenBank data libraries under acces-
sion numbers DQ019624–DQ019639.
Results
Structural organization and expression of TaRSZ38
A tblastn computer search (Altschul et al. 1997) of the
GenBank expressed sequence tag database (dbEST),
using as a query the translated full-length cDNA of the
AtRSZ33 gene from Arabidopsis (Accession
AJ293801), returned several wheat ESTs with a high
level of identity to the RNA recognition motif (RRM)
and zinc finger (ZF) domains of AtRSZ33. The contigs
of the ESTs resulted in two cDNAs, both with open
reading frames encoding proteins of 333 amino acids
and a calculated molecular mass of 38 kDa, designated
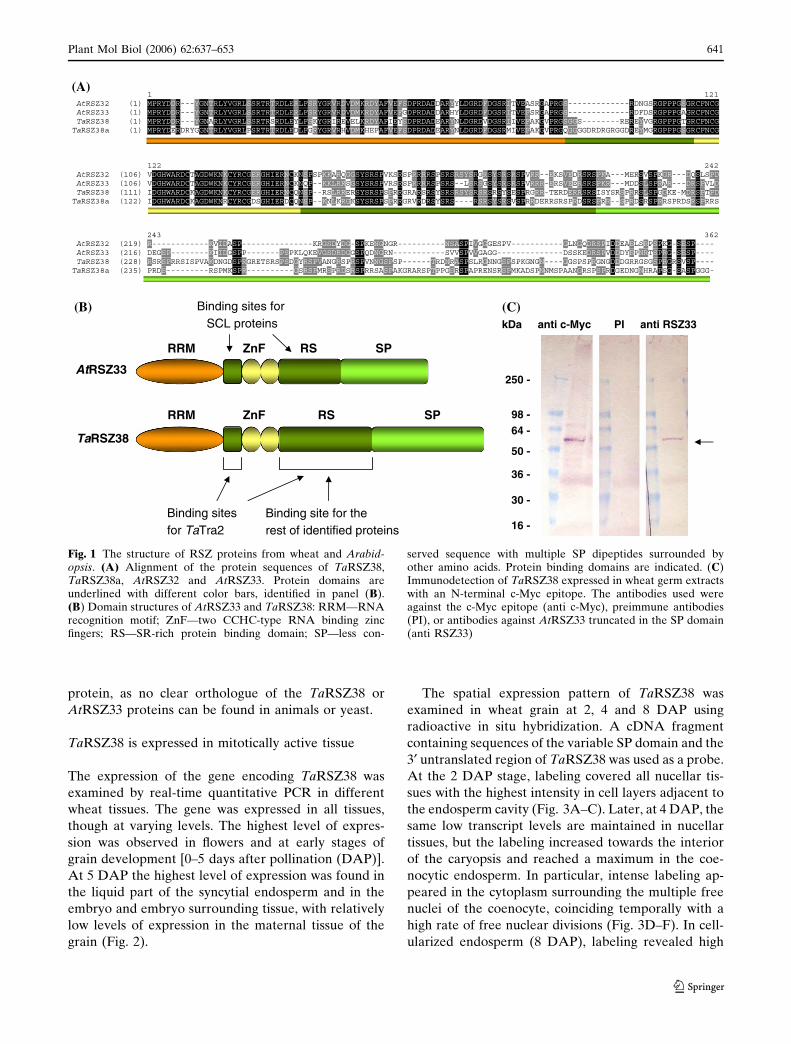
TaRSZ38 and TaRSZ38a (Fig. 1A). The TaRSZ38 and
TaRSZ38a proteins have the same well-defined mod-
ular domains as AtRSZ33 (Fig. 1A, B). Alignment of
the deduced protein sequences of TaRSZ38 and
TaRSZ38a to AtRSZ33 and the related AtRSZ32
(Lopato et al. 2002) demonstrated a high level of
conservation in the RRM domain, the two CCHC-type
zinc fingers and the adjacent short RS-rich domain.
The identity across the RNA-binding domains is
slightly higher between TaRSZ38 and AtRSZ33 (68%
identity) than between TaRSZ38a and AtRSZ33 (66%
identity). In contrast, the SP domain of the wheat
proteins is longer and has no sequence similarity to the
SP domain of AtRSZ33. The level of conservation
between the two wheat proteins in the C-terminal part
is also low. No genes or ESTs were found in the
databases of Arabidopsis using the variable C-terminal
part of the wheat proteins. Therefore, TaRSZ38 has
the highest level of identity to AtRSZ33 of any known
or deduced wheat protein. TaRSZ38 is a plant specific
640 Plant Mol Biol (2006) 62:637–653
123
protein, as no clear orthologue of the TaRSZ38 or
AtRSZ33 proteins can be found in animals or yeast.
TaRSZ38 is expressed in mitotically active tissue
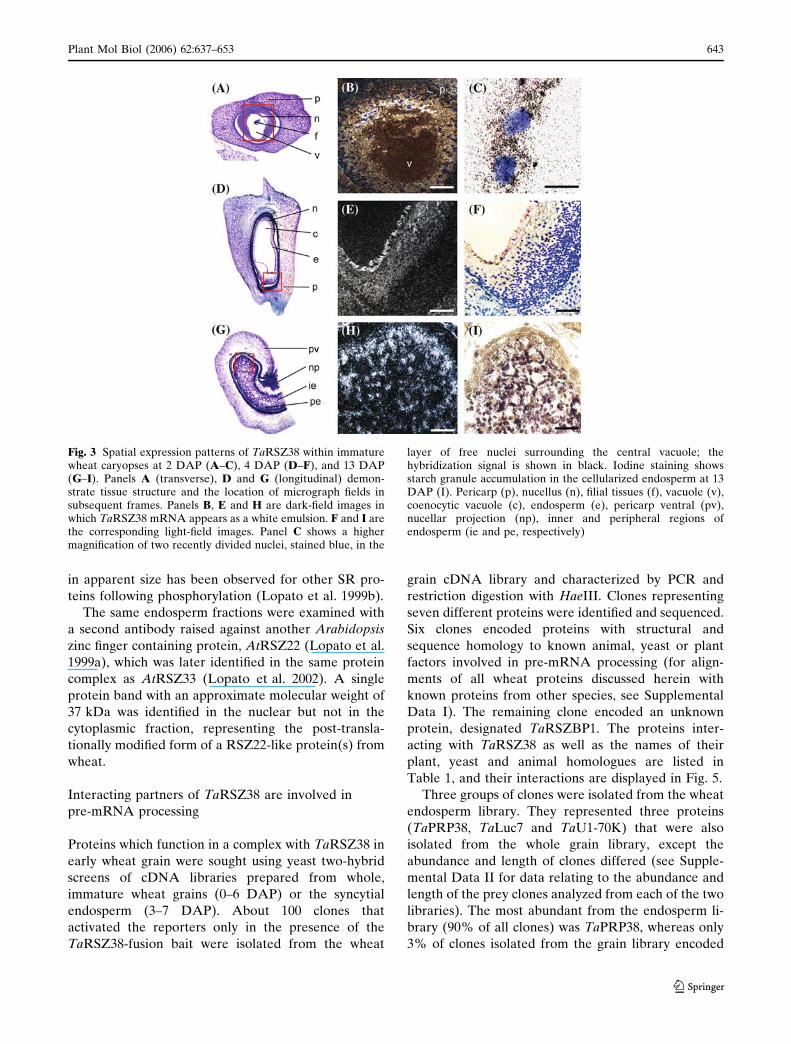
The expression of the gene encoding TaRSZ38 was
examined by real-time quantitative PCR in different
wheat tissues. The gene was expressed in all tissues,
though at varying levels. The highest level of expres-
sion was observed in flowers and at early stages of
grain development [0–5 days after pollination (DAP)].
At 5 DAP the highest level of expression was found in
the liquid part of the syncytial endosperm and in the
embryo and embryo surrounding tissue, with relatively
low levels of expression in the maternal tissue of the
grain (Fig. 2).
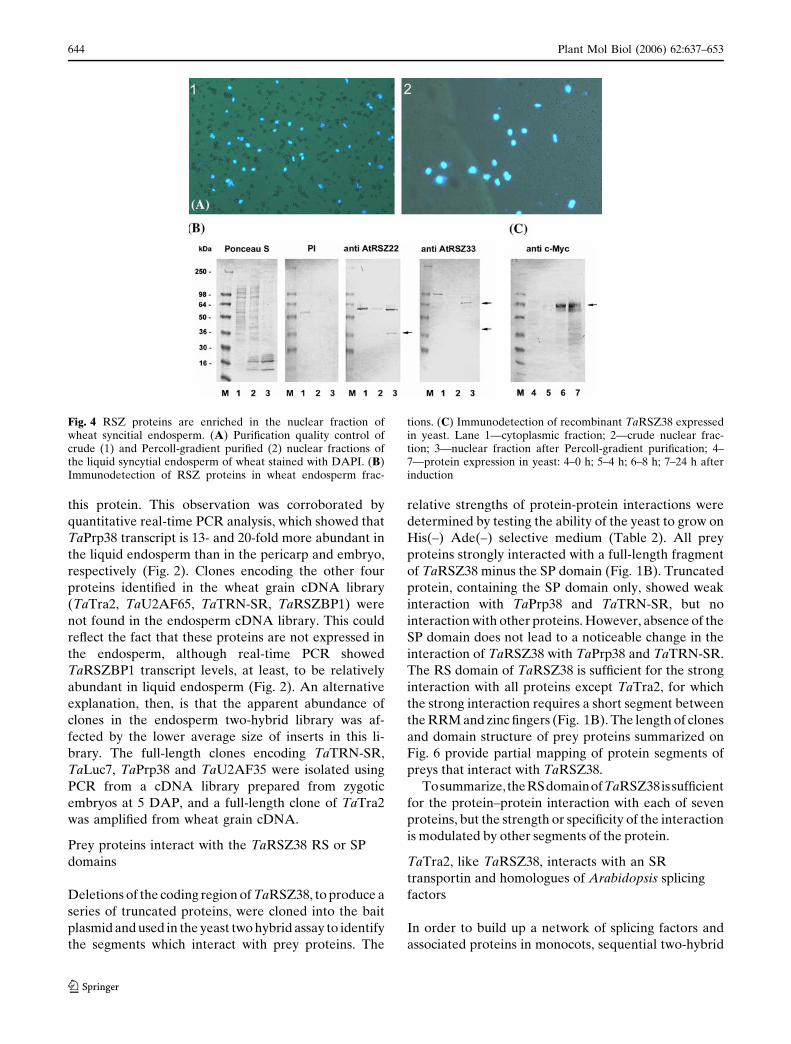
The spatial expression pattern of TaRSZ38 was
examined in wheat grain at 2, 4 and 8 DAP using
radioactive in situ hybridization. A cDNA fragment
containing sequences of the variable SP domain and the
3¢ untranslated region of TaRSZ38 was used as a probe.
At the 2 DAP stage, labeling covered all nucellar tis-
sues with the highest intensity in cell layers adjacent to
the endosperm cavity (Fig. 3A–C). Later, at 4 DAP, the
same low transcript levels are maintained in nucellar
tissues, but the labeling increased towards the interior
of the caryopsis and reached a maximum in the coe-
nocytic endosperm. In particular, intense labeling ap-
peared in the cytoplasm surrounding the multiple free
nuclei of the coenocyte, coinciding temporally with a
high rate of free nuclear divisions (Fig. 3D–F). In cell-
ularized endosperm (8 DAP), labeling revealed high
1 121AtRSZ32 (1) MPRYDDR---YGNTRLYVGRLSSRTRTRDLERLFSRYGRVRDVDMKRDYAFVEFSDPRDADDARYYLDGRDFDGSRITVEASRGAPRGS-------------RDNGSRGPPPGSGRCFNCGAtRSZ33 (1) MPRYDDR---YGNTRLYVGRLSSRTRTRDLERLFSRYGRVRDVDMKRDYAFVEFGDPRDADDARHYLDGRDFDGSRITVEFSRGAPRGS-------------RDFDSRGPPPGAGRCFNCGTaRSZ38 (1) MPRYDDR---YGNARLYVGRLSSRTRSRDLEYLFSKYGRIREVELKRDYAFIEYSDPRDADEARYNLDGRDVDGSRIIVEFAKGVPRGSGGS--------REREYVGRGPPPGTGRCFNCG
TaRSZ38a (1) MPRYDERDRYGGNTRLYVGRLPSRTRTRDLEDLFGRYGRVRHVDMKHEFAFVEFSDPRDADEARYNLDGRDFDGSRMIVEFAKGVPRGQGGGGDRDRGRGGDREYMGRGPPPGSGRCFNCG
122 242AtRSZ32 (106) VDGHWARDCTAGDWKNKCYRCGERGHIERNCKNSPSPKKARQGGSYSRSPVKSRSPRRRRSPSRSRSYSRGRSYSRSRSPVRR--EKSVEDRSRSPKA---MERSVSPKGR---DQSLSPDAtRSZ33 (106) VDGHWARDCTAGDWKNKCYRCGERGHIERNCKNQP--KKLRRSGSYSRSPVRSRSPRRRRSPSRS--LSRSGSYSRSRSPVRRR-ERSVEERSRSPKR---MDDSLSPRAR---DRSPVLDTaRSZ38 (111) IDGHWARDCKAGDWKNKCYRCGERGHIERNCQNSP--RSLRRERSYSRSPSPRRGRARSRSYSRSRSYSRSRSRSYSESPRGRR-TERDERRSRSISYSRSPRRSLSPGGKE-MDRSPTPD
TaRSZ38a (122) IDGHWARDCKAGDWKNRCYRCGDSGHIERDCQNSP--KNLKRGKSYSRSPSPRRGRVRDRSYSRS----RSRSYSRSVSPRRDERRSRSPRDSRSPRR--SPRDSRSPRRSPRDSRSPRRS
243 362AtRSZ32 (219) R------------KVIDASP---------------KRGSDYDG-SPKENGNGR----------NSASPIVGGGESPV-----------GLNGQDRSPIDDEAELSRPSPKG-SESP---- AtRSZ33 (216) DEGSP--------KIIDGSPP-------PSPKLQKEVGSDRDGGSPQDNGRN-----------SVVSPVVGAGG--------------DSSKEDRSPVDDDYEPNRTSPRG-SESP---- TaRSZ38 (228) RSRSPRRSISPVAKDNGDSPRGRETSRSPSDGYRSPVANGRSPRSPVNNGSPSP------TRDNRASPSLRGNNGSPSPKGNGN----GGSPSPRGNGDDDGRRGSGSPRGRSVSP----
TaRSZ38a (235) PRDS---------RSPMKSPR----------GSRSPMRSPRDSRSPRRSASPAKGRARSPTPPGSRSPAPRENSRSPMKADSPNNMSPAANGRSPSPRDGEDNGNHRAPSG-SASPGGG-
(A)
Binding site for the rest of identified proteins
ZnF RS SP
AtRSZ33
TaRSZ38
Binding sites forSCL proteins
Binding sites for TaTra2
RRM
ZnF RS SPRRM
(B)kDa
250 -
98 -64 -
50 -
36 -
30 -
16 -
anti c-Myc PI anti RSZ33
(C)
Fig. 1 The structure of RSZ proteins from wheat and Arabid-opsis. (A) Alignment of the protein sequences of TaRSZ38,TaRSZ38a, AtRSZ32 and AtRSZ33. Protein domains areunderlined with different color bars, identified in panel (B).(B) Domain structures of AtRSZ33 and TaRSZ38: RRM—RNArecognition motif; ZnF—two CCHC-type RNA binding zincfingers; RS—SR-rich protein binding domain; SP—less con-
served sequence with multiple SP dipeptides surrounded byother amino acids. Protein binding domains are indicated. (C)Immunodetection of TaRSZ38 expressed in wheat germ extractswith an N-terminal c-Myc epitope. The antibodies used wereagainst the c-Myc epitope (anti c-Myc), preimmune antibodies(PI), or antibodies against AtRSZ33 truncated in the SP domain(anti RSZ33)
Plant Mol Biol (2006) 62:637–653 641
123

expression of TaRSZ38 at the periphery of the endo-
sperm, in mitotically active cells with dense cytoplasm
and a low content of starch grains. The level of
expression decreases toward the inner region of the
endosperm, where the major storage tissue of the grain
at this time is characterized by cells of different size
with large vacuoles and large starch granules (Fig. 3G–
I). The labeling pattern described here therefore im-
plies preferential expression of TaRSZ38 in mitotically
active tissues of the developing caryopsis.
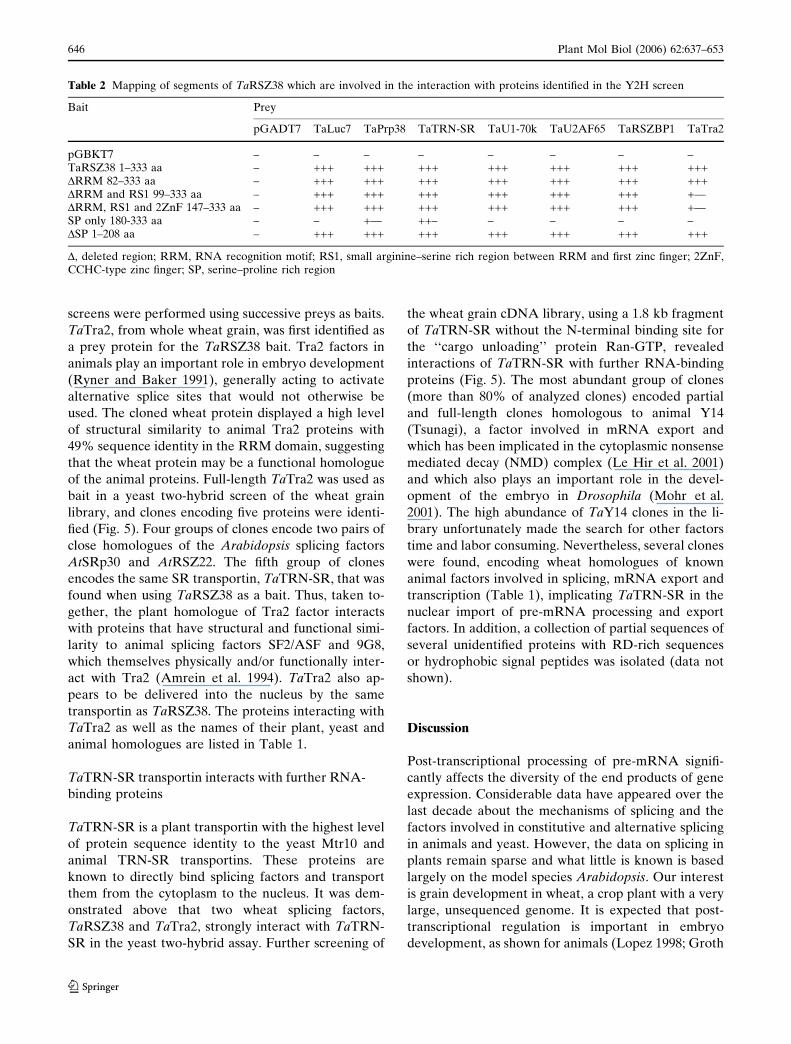
Immunodetection of TaRSZ38 in the nuclear
fraction of the syncytial endosperm
The large syncytial endosperm of wheat provided a
unique opportunity to purify intact nuclei and nuclei-
free cytoplasmic fractions from a single multinucleate
cell. The high level of sequence similarity between
TaRSZ38 and AtRSZ33 suggested also that the wheat
protein might be recognized by antibodies raised
against AtRSZ33 from Arabidopsis (Lopato et al.
2002). To test this, TaRSZ38 was expressed with an
N-terminal c-Myc epitope in wheat germ extracts, and
found to cross-react with the anti c-Myc and anti-
AtRSZ33 antibodies (Fig. 1C). Analyses of the
cytoplasmic and nuclear fractions of the syncytial
endosperm using antibodies against AtRSZ33 revealed
one band in the nuclei-free cytoplasm and several
protein bands in the nuclear fraction with a major
protein band at about 64 kDa (Fig. 4). Expression of
recombinant TaRSZ38 in eukaryotic yeast cells, which
contain homologues of specific SR protein kinases
(Siebel et al. 1999), resulted in a single protein band of
the same size (Fig. 4). Therefore it appears that the
64 kDa protein band represented a form of TaRSZ38
that had been modified post-translationally, running
more slowly in SDS-PAGE than the unmodified form
(deduced molecular weight of 38 kDa). A similar shift
TaPrp38
0
5000
10000
15000
20000
Co
pie
s/m
icro
gra
m o
f R
NA
0
15000
30000
45000
60000
Inflo
resc
ence
Flowers
, meio
sis
Flowers
1 d pre
-pol
Grain
0-1 D
AP
Grain
2-3 D
AP
Grain
4-5
DAP
Grain
6-7 D
AP
Grain
8-1
0 DAP
Grain
11-15
DAP
Grain
16-20
DAP
Embryo (e
nrich
ed) 5 D
AP
Endo
sper
m (liqu
id) 5
DAP
Pericarp
5 D
AP
Roots (s
eedling)
Shoots
(seed
ling)
Wou
nded
LeafSte
m
Cop
ies/
mic
rogr
am o
f RN
A
TaRSZBP1
Flowers Developing grain Grain fractions Vegetative tissue
0
5000
10000
15000
20000
25000
Co
pie
s/m
icro
gra
m o
f R
NA
TaRSZ38
Fig. 2 Expression of TaRSZ38, TaPrp38 and TaRSZBP1 at different stages of flower and grain development, in grain fractions andvegetative tissue, as determined by quantitative real-time PCR. Data are normalized against the input amount of RNA
642 Plant Mol Biol (2006) 62:637–653
123
in apparent size has been observed for other SR pro-
teins following phosphorylation (Lopato et al. 1999b).
The same endosperm fractions were examined with
a second antibody raised against another Arabidopsis
zinc finger containing protein, AtRSZ22 (Lopato et al.
1999a), which was later identified in the same protein
complex as AtRSZ33 (Lopato et al. 2002). A single
protein band with an approximate molecular weight of
37 kDa was identified in the nuclear but not in the
cytoplasmic fraction, representing the post-transla-
tionally modified form of a RSZ22-like protein(s) from
wheat.
Interacting partners of TaRSZ38 are involved in
pre-mRNA processing
Proteins which function in a complex with TaRSZ38 in
early wheat grain were sought using yeast two-hybrid
screens of cDNA libraries prepared from whole,
immature wheat grains (0–6 DAP) or the syncytial
endosperm (3–7 DAP). About 100 clones that
activated the reporters only in the presence of the
TaRSZ38-fusion bait were isolated from the wheat
grain cDNA library and characterized by PCR and
restriction digestion with HaeIII. Clones representing
seven different proteins were identified and sequenced.
Six clones encoded proteins with structural and
sequence homology to known animal, yeast or plant
factors involved in pre-mRNA processing (for align-
ments of all wheat proteins discussed herein with
known proteins from other species, see Supplemental
Data I). The remaining clone encoded an unknown
protein, designated TaRSZBP1. The proteins inter-
acting with TaRSZ38 as well as the names of their
plant, yeast and animal homologues are listed in
Table 1, and their interactions are displayed in Fig. 5.
Three groups of clones were isolated from the wheat
endosperm library. They represented three proteins
(TaPRP38, TaLuc7 and TaU1-70K) that were also
isolated from the whole grain library, except the
abundance and length of clones differed (see Supple-
mental Data II for data relating to the abundance and
length of the prey clones analyzed from each of the two
libraries). The most abundant from the endosperm li-
brary (90% of all clones) was TaPRP38, whereas only
3% of clones isolated from the grain library encoded
Fig. 3 Spatial expression patterns of TaRSZ38 within immaturewheat caryopses at 2 DAP (A–C), 4 DAP (D–F), and 13 DAP(G–I). Panels A (transverse), D and G (longitudinal) demon-strate tissue structure and the location of micrograph fields insubsequent frames. Panels B, E and H are dark-field images inwhich TaRSZ38 mRNA appears as a white emulsion. F and I arethe corresponding light-field images. Panel C shows a highermagnification of two recently divided nuclei, stained blue, in the
layer of free nuclei surrounding the central vacuole; thehybridization signal is shown in black. Iodine staining showsstarch granule accumulation in the cellularized endosperm at 13DAP (I). Pericarp (p), nucellus (n), filial tissues (f), vacuole (v),coenocytic vacuole (c), endosperm (e), pericarp ventral (pv),nucellar projection (np), inner and peripheral regions ofendosperm (ie and pe, respectively)
Plant Mol Biol (2006) 62:637–653 643
123
this protein. This observation was corroborated by
quantitative real-time PCR analysis, which showed that
TaPrp38 transcript is 13- and 20-fold more abundant in
the liquid endosperm than in the pericarp and embryo,
respectively (Fig. 2). Clones encoding the other four
proteins identified in the wheat grain cDNA library
(TaTra2, TaU2AF65, TaTRN-SR, TaRSZBP1) were
not found in the endosperm cDNA library. This could
reflect the fact that these proteins are not expressed in
the endosperm, although real-time PCR showed
TaRSZBP1 transcript levels, at least, to be relatively
abundant in liquid endosperm (Fig. 2). An alternative
explanation, then, is that the apparent abundance of
clones in the endosperm two-hybrid library was af-
fected by the lower average size of inserts in this li-
brary. The full-length clones encoding TaTRN-SR,
TaLuc7, TaPrp38 and TaU2AF35 were isolated using
PCR from a cDNA library prepared from zygotic
embryos at 5 DAP, and a full-length clone of TaTra2
was amplified from wheat grain cDNA.
Prey proteins interact with the TaRSZ38 RS or SP
domains
Deletions of the coding region of TaRSZ38, to produce a
series of truncated proteins, were cloned into the bait
plasmid and used in the yeast two hybrid assay to identify
the segments which interact with prey proteins. The
relative strengths of protein-protein interactions were
determined by testing the ability of the yeast to grow on
His(–) Ade(–) selective medium (Table 2). All prey
proteins strongly interacted with a full-length fragment
of TaRSZ38 minus the SP domain (Fig. 1B). Truncated
protein, containing the SP domain only, showed weak
interaction with TaPrp38 and TaTRN-SR, but no
interaction with other proteins. However, absence of the
SP domain does not lead to a noticeable change in the
interaction of TaRSZ38 with TaPrp38 and TaTRN-SR.
The RS domain of TaRSZ38 is sufficient for the strong
interaction with all proteins except TaTra2, for which
the strong interaction requires a short segment between
the RRM and zinc fingers (Fig. 1B). The length of clones
and domain structure of prey proteins summarized on
Fig. 6 provide partial mapping of protein segments of
preys that interact with TaRSZ38.
Tosummarize, theRSdomainofTaRSZ38issufficient
for the protein–protein interaction with each of seven
proteins, but the strength or specificity of the interaction
is modulated by other segments of the protein.
TaTra2, like TaRSZ38, interacts with an SR
transportin and homologues of Arabidopsis splicing
factors
In order to build up a network of splicing factors and
associated proteins in monocots, sequential two-hybrid
Fig. 4 RSZ proteins are enriched in the nuclear fraction ofwheat syncitial endosperm. (A) Purification quality control ofcrude (1) and Percoll-gradient purified (2) nuclear fractions ofthe liquid syncytial endosperm of wheat stained with DAPI. (B)Immunodetection of RSZ proteins in wheat endosperm frac-
tions. (C) Immunodetection of recombinant TaRSZ38 expressedin yeast. Lane 1—cytoplasmic fraction; 2—crude nuclear frac-tion; 3—nuclear fraction after Percoll-gradient purification; 4–7—protein expression in yeast: 4–0 h; 5–4 h; 6–8 h; 7–24 h afterinduction
644 Plant Mol Biol (2006) 62:637–653
123

Ta
ble
1P
rote
incl
ass
an
dfu
nct
ion
al
info
rma
tio
nfo
rcl
on
es
ide
nti
fie
din
ye
ast
two
-hy
bri
dsc
ree
ns
Wh
ea
tp
rote
inP
rote
incl
ass
Fu
nct
ion
Ma
inco
mp
lex
ass
oci
ati
on
RN
A-b
ind
ing
site
sL
oca
liza
tio
nA
nim
al/
ye
ast
or
pla
nt
ho
mo
log
ue
sR
efe
ren
ces
TaR
SZ
38
Pla
nt-
spe
cifi
cS
Rp
rote
inw
ith
two
zin
cfi
ng
ers
Sp
lice
oso
me
form
ati
on
,a
lte
rna
tiv
esp
lici
ng
reg
ula
tio
n,
de
ve
lop
me
nta
lsi
gn
al
tra
nsd
uct
ion
(?)
Sp
lice
oso
me
Un
kn
ow
nN
ucl
ea
r‘‘
RS
Z’’
spe
ckle
s,cy
top
lasm
(?)
AtR
SZ
33
Lo
pa
toe
ta
l.(2
00
2)
TaT
ra2
RN
A-b
ind
ing
pro
tein
wit
htw
oR
Sd
om
ain
s
Alt
ern
ati
ve
spli
cin
gre
gu
lato
rS
pli
ceo
som
eG
C-r
ich
ex
on
ice
nh
an
cers
Nu
cle
ar
spe
ckle
sT
ra2
Ry
ne
ra
nd
Ba
ke
r(1
99
1)
TaL
uc7
U1
snR
NP
-b
ou
nd
pro
tein
Co
nn
ect
sca
pp
ing
pro
tein
com
ple
xw
ith
U1
snR
NP
U1
snR
NP
-bo
un
dU
nk
no
wn
Sp
lice
oso
me
,U
1sn
RN
PL
uc7
/CR
OP
Fo
rte
se
ta
l.(1
99
9)
TaU
1-
70
kD
RS
-co
nta
inin
gp
rote
inR
eco
gn
itio
no
fth
e5
¢-sp
lice
site
Re
cru
its
U1
snR
NP
toth
e5
¢-sp
lice
site
5¢-s
pli
cesi
teS
pli
ceo
som
eU
1-7
0k
D,
AtU
1-7
0K
Go
lov
kin
an
dR
ed
dy
(19
96
)
TaU
2A
F6
5
TaU
2A
F6
5a
RS
-co
nta
inin
gp
rote
inR
eco
gn
itio
no
fth
eb
ran
chp
oin
t;re
cru
its
U2
snR
NP
La
rge
sub
un
ito
fU
2A
Ffa
cto
rIn
tro
nic
U-r
ich
seq
ue
nce
Nu
cle
ar
spe
ckle
sU
2A
F6
5,
Np
U2
AF
65
Do
mo
ne
ta
l.(1
99
8)
TaP
rp3
8D
Ex
D/H
-bo
xR
NA
-he
lica
se-
lik
ep
rote
in
Un
win
din
go
fth
eU
4/U
6sn
RN
AU
6/U
4.U
5tr
i-sn
RN
PU
nk
no
wn
Sp
lise
oso
me
,e
mb
ryo
-sp
eci
fic
inw
orm
s
Prp
38
Bla
nto
ne
ta
l.(1
99
2)
TaT
RN
-SR
Nu
cle
ar
tra
nsp
ort
rece
pto
rT
ran
spo
rto
fp
re-m
RN
Ap
roce
ssin
gfa
cto
rsto
the
nu
cle
us
Nu
cle
ar
tra
nsp
ort
pro
tein
com
ple
xN
oN
ucl
ea
rsp
eck
les,
cyto
pla
smT
RN
-SR
,M
tr1
0S
en
ge
re
ta
l.(1
99
8);
Ka
tao
ka
et
al.
(19
99
)T
aR
SZ
BP
1P
lan
t(m
on
oco
t?)-
spe
cifi
cU
nk
no
wn
Un
kn
ow
nU
nk
no
wn
Un
kn
ow
nN
ot
ide
nti
fie
d
TaS
Rp
30
SR
pro
tein
Sp
lice
oso
me
form
ati
on
,re
gu
lati
on
of
5¢
alt
ern
ati
ve
spli
cin
g
Sp
lice
oso
me
Ex
on
ica
nd
intr
on
icsp
lici
ng
en
ha
nce
rsN
ucl
ea
rsp
eck
les,
nu
cle
ar-
cyto
pla
smic
shu
ttli
ng
pro
tein
SF
2/A
SF
,A
tSR
p3
0,
SR
1L
aza
re
ta
l.(1
99
5);
Lo
pa
toe
ta
l.(1
99
9b
)
TaS
Rp
30
a
TaR
SZ
22
SR
pro
tein
wit
ha
sin
gle
zin
cfi
ng
er
Sp
lici
ng
fact
or
Sp
lice
oso
me
Sp
lici
ng
en
ha
nce
rssi
mil
ar
toe
nh
an
cer
reco
gn
ize
db
yh
um
an
9G
8
Nu
cle
ar
‘‘R
SZ
’’sp
eck
les
Pla
nt-
spe
cifi
cp
rote
in,
com
bin
es
fea
ture
so
fh
um
an
SR
p2
0a
nd
9G
8
Go
lov
kin
an
dR
ed
dy
(19
98
);L
op
ato
et
al.
(19
99
a)
TaR
SZ
22
a
TaU
2A
F3
5R
S-c
on
tain
ing
pro
tein
Re
cog
nit
ion
of
the
3¢-s
pli
cesi
teS
ma
llsu
bu
nit
of
U2
AF
fact
or
Re
cog
nit
ion
of
the
3-s
pli
cesi
teN
ucl
ea
rsp
eck
les
U2
AF
35
Do
mo
ne
ta
l.(1
99
8)
TaY
14
RN
A-b
ind
ing
pro
tein
mR
NA
ex
po
rta
nd
turn
ov
er
mR
NA
ex
po
rtco
mp
lex
,N
MD
com
ple
xin
cyto
pla
sm
Re
cog
nit
ion
of
ex
on
–e
xo
nju
nct
ion
site
inm
RN
A
Nu
cle
ar-
cyto
pla
smic
shu
ttli
ng
pro
tein
Y1
4,
Tsu
na
gi
Ka
tao
ka
et
al.
(20
00
);M
oh
re
ta
l.(2
00
1)
TaD
RH
1D
EA
D-b
ox
AT
Pa
se-
RN
A-h
eli
case
AT
Pd
ep
en
de
nt
dsR
NA
un
win
din
gS
pli
cin
g,
tra
nsl
ati
on
,ri
bo
som
ea
sse
mb
ly,
RN
Ad
eg
rad
ati
on
Un
kn
ow
nN
ucl
ea
r-cy
top
lasm
icsh
utt
lin
gp
rote
in
DR
H1
,p
68
Igg
oe
ta
l.(1
99
1);
Ok
an
am
ie
ta
l.(1
99
8)
Plant Mol Biol (2006) 62:637–653 645
123
screens were performed using successive preys as baits.
TaTra2, from whole wheat grain, was first identified as
a prey protein for the TaRSZ38 bait. Tra2 factors in
animals play an important role in embryo development
(Ryner and Baker 1991), generally acting to activate
alternative splice sites that would not otherwise be
used. The cloned wheat protein displayed a high level
of structural similarity to animal Tra2 proteins with
49% sequence identity in the RRM domain, suggesting
that the wheat protein may be a functional homologue
of the animal proteins. Full-length TaTra2 was used as
bait in a yeast two-hybrid screen of the wheat grain
library, and clones encoding five proteins were identi-
fied (Fig. 5). Four groups of clones encode two pairs of
close homologues of the Arabidopsis splicing factors
AtSRp30 and AtRSZ22. The fifth group of clones
encodes the same SR transportin, TaTRN-SR, that was
found when using TaRSZ38 as a bait. Thus, taken to-
gether, the plant homologue of Tra2 factor interacts
with proteins that have structural and functional simi-
larity to animal splicing factors SF2/ASF and 9G8,
which themselves physically and/or functionally inter-
act with Tra2 (Amrein et al. 1994). TaTra2 also ap-
pears to be delivered into the nucleus by the same
transportin as TaRSZ38. The proteins interacting with
TaTra2 as well as the names of their plant, yeast and
animal homologues are listed in Table 1.
TaTRN-SR transportin interacts with further RNA-
binding proteins
TaTRN-SR is a plant transportin with the highest level
of protein sequence identity to the yeast Mtr10 and
animal TRN-SR transportins. These proteins are
known to directly bind splicing factors and transport
them from the cytoplasm to the nucleus. It was dem-
onstrated above that two wheat splicing factors,
TaRSZ38 and TaTra2, strongly interact with TaTRN-
SR in the yeast two-hybrid assay. Further screening of
the wheat grain cDNA library, using a 1.8 kb fragment
of TaTRN-SR without the N-terminal binding site for
the ‘‘cargo unloading’’ protein Ran-GTP, revealed
interactions of TaTRN-SR with further RNA-binding
proteins (Fig. 5). The most abundant group of clones
(more than 80% of analyzed clones) encoded partial
and full-length clones homologous to animal Y14
(Tsunagi), a factor involved in mRNA export and
which has been implicated in the cytoplasmic nonsense
mediated decay (NMD) complex (Le Hir et al. 2001)
and which also plays an important role in the devel-
opment of the embryo in Drosophila (Mohr et al.
2001). The high abundance of TaY14 clones in the li-
brary unfortunately made the search for other factors
time and labor consuming. Nevertheless, several clones
were found, encoding wheat homologues of known
animal factors involved in splicing, mRNA export and
transcription (Table 1), implicating TaTRN-SR in the
nuclear import of pre-mRNA processing and export
factors. In addition, a collection of partial sequences of
several unidentified proteins with RD-rich sequences
or hydrophobic signal peptides was isolated (data not
shown).
Discussion
Post-transcriptional processing of pre-mRNA signifi-
cantly affects the diversity of the end products of gene
expression. Considerable data have appeared over the
last decade about the mechanisms of splicing and the
factors involved in constitutive and alternative splicing
in animals and yeast. However, the data on splicing in
plants remain sparse and what little is known is based
largely on the model species Arabidopsis. Our interest
is grain development in wheat, a crop plant with a very
large, unsequenced genome. It is expected that post-
transcriptional regulation is important in embryo
development, as shown for animals (Lopez 1998; Groth
Table 2 Mapping of segments of TaRSZ38 which are involved in the interaction with proteins identified in the Y2H screen
Bait Prey
pGADT7 TaLuc7 TaPrp38 TaTRN-SR TaU1-70k TaU2AF65 TaRSZBP1 TaTra2
pGBKT7 – – – – – – – –TaRSZ38 1–333 aa – +++ +++ +++ +++ +++ +++ +++DRRM 82–333 aa – +++ +++ +++ +++ +++ +++ +++DRRM and RS1 99–333 aa – +++ +++ +++ +++ +++ +++ +––DRRM, RS1 and 2ZnF 147–333 aa – +++ +++ +++ +++ +++ +++ +––SP only 180-333 aa – – +–– ++– – – – –DSP 1–208 aa – +++ +++ +++ +++ +++ +++ +++
D, deleted region; RRM, RNA recognition motif; RS1, small arginine–serine rich region between RRM and first zinc finger; 2ZnF,CCHC-type zinc finger; SP, serine–proline rich region
646 Plant Mol Biol (2006) 62:637–653
123

AD-TaLuc7
AD-TaU1-70K
pGADT7
AD-TaPrp38
AD-TaTra2
AD-TaTRN-SR
AD-TaU2AF65
AD-TaRSZBP1
pGBKT7 BD-TaRSZ38 BD-TaRSZ38pGBKT7
-Leu, -Trp -Leu, -Trp, -His, -Ade, + X-GAL
pGADT7
AD-TaSRp30
AD-TaSRp30a
AD-TaRSZ22
AD-TaRSZ22a
AD-TaTRN-SR
pGBKT7 BD-TaTra2 pGBKT7 BD-TaTra2
pGADT7
AD-DRH1
AD-U2AF35
AD-TaY14
AD-TaU2AF65
pGBKT7 BD-TaTRN-SR pGBKT7BD-TaTRN-SR
TaRSZBP1
TaU2AF65
TaTra2
TaPrp38
TaLuc7
TaU1-70K
TaTRN-SR
TaSRp30
TaRSZ22
TaU2AF35TaRSZ38
TaSRp30aTaRSZ22a
TaDRH1
TaY14
Unknown proteinswith SD-rich sequences
TaU2AF65a
Data obtained for wheat grain
Published data for other organisms
(A)
(B)
(C)
(D)
Plant Mol Biol (2006) 62:637–653 647
123

and Lardelli 2002; Lalli et al. 2003) and Arabidopsis
(Kalyna et al. 2003), and exerts some control over such
factors as embryo size and composition. Equally, an
understanding of the specific features of splicing reg-
ulation in the developing endosperm may provide tools
for manipulating endosperm size, composition or
morphology, and consequently grain yield and quality.
As a starting bait in yeast two-hybrid screens of
cDNA libraries prepared from early wheat grain (0–6
DAP) and the liquid part of the syncytial endosperm
(3–7 DAP), the wheat homologue of AtRSZ33 was
chosen as this is the only plant splicing factor with a
demonstrated role in early seed development (Kalyna
et al. 2003). This protein is expressed in different plant
tissues, but only in cells undergoing active division and
elongation. Overexpression of AtRSZ33 in transgenic
Arabidopsis resulted in several pronounced changes in
plant development (Kalyna et al. 2003). The most
interesting changes took place in the early stages of
embryo development, with the appearance of either
twin or monocotyledonous plants after germination.
The wheat homologue of AtRSZ33 was expressed in all
tissues examined, with the highest level of expression
in developing grain. These data correlate with expres-
sion data obtained earlier for AtRSZ33 using pro-
moter:GUS constructs (Lopato et al. 2002; Lorkovic
and Barta 2002; Kalyna et al. 2003). AtRSZ33 is
expressed in restricted groups of cells in different
organs of Arabidopsis including seeds. The large size of
the wheat grain relative to Arabidopsis was exploited
to demonstrate that TaRSZ38 is expressed mainly in
cells which are undergoing intense mitotic division.
The highest expression was found in the syncytium
around the dividing free nuclei. Expression of
TaRSZ38 in the dividing nuclei of the coenocyte was
further supported by immunodetection of the
TaRSZ38 protein in the nuclear fraction purified from
syncytial endosperm. These data suggest that the role
of TaRSZ38 is not restricted to the regulation of em-
bryo development, but might be extended to endo-
sperm development. Although it was demonstrated
earlier that AtRSZ33 is expressed in Arabidopsis
endosperm (Kalyna et al. 2003), changes in endosperm
in transgenic plants overexpressing AtRSZ33 were not
studied.
A previous two-hybrid screen with AtRSZ33 from
Arabidopsis did not include the SP domain in the bait
construct, and clones from organs such as flowers and
siliques were under-represented in the prey library
(Lopato et al. 2002). In contrast, the full-length
TaRSZ38 could be expressed successfully in yeast,
providing an ideal bait. Two cDNA libraries were
screened, prepared from the whole grain and liquid
coenocyte at overlapping stages of development. Six
from seven identified proteins were either known
splicing factors or splicing related proteins. In a study
of this size, the usual method for confirming yeast two-
hybrid data, pull-down assays, is extremely difficult to
achieve. Protein phosphorylation is a likely pre-
requisite for correct protein interactions, particularly
involving RS domains, precluding the use of re-
combinant proteins expressed in bacteria, while pull-
down assays using plant material would require the
generation of antibodies specific to each protein. In the
circumstances, we have corroborated the observed
yeast two-hybrid data by comparison to protein inter-
action databases that include homologues of the cloned
wheat genes.
No members of the SC35-like (SCL) family of pro-
teins, identified in a previous study (Lopato et al.
2002), were found in these screens. One prey protein
(TaTra2) resembles SCL proteins, having a single
RRM domain in the central part of the protein (Fig. 6),
but the wheat protein is in fact most closely related to
animal Tra2 splicing factor (49% identity in the RRM
domain). Animal Tra2 factors are important for
development, blocking splice sites in specific
pre-mRNAs during the early stages of Drosophila
embryo development (Ryner and Baker 1991) and
likely participating in the control of human cell specific
splicing patterns (Tacke et al. 1998). TaTra2 interacts
in the grain with other splicing factors and may have a
similar role in regulating splicing events during devel-
opment. Despite the fact that TaTra2 and SCL proteins
are structurally distinct, mapping of protein–protein
interactions demonstrated that they interact with the
same segments of RSZ proteins.
Homologues of two further proteins identified in the
screen with RSZ33 from Arabidopsis, AtSRp34 and
AtRSZ22, were not found in the screen with wheat
TaRSZ38. Surprisingly, several homologues of each of
Fig. 5 Protein–protein interaction data. (A–C) reconstitution oftwo-hybrid interactions found in yeast two-hybrid screening.Yeast cells were co-transformed either with prey plasmidsisolated in a yeast two-hybrid screening (indicated at the leftside) and empty bait vector (pGBKT7), or with the preyplasmids and the bait plasmid used in the respective screens(A, BD-TaRSZ38; B, BD-TaTra2; and C, BD-TaTRN-SR).Yeast cells were maintained on selective media without leucineand tryptophan (-Leu, -Trp) or without leucine, tryptophan,histidine and adenine, but with X-GAL to test for galactosidaseactivity (-Leu, -Trp, -His, -Ade, + X-GAL). (D) Schematicrepresentation of protein–protein interaction data obtained fromthe yeast two-hybrid screens and partially confirmed by datapublished for other organisms. AD, activation domain; BD,binding domain
m648 Plant Mol Biol (2006) 62:637–653
123

these proteins were found in a subsequent two-hybrid
screen with TaTra2 as bait. This supports the idea that
in wheat and Arabidopsis small 9G8-like RSZ proteins
and the more evolutionary conserved SF2/ASF-like
SRp proteins function in the same complex together
with either Tra2-like and/or SCL proteins. Interaction
of the Drosophila splicing regulator Tra2 with the SF2/
ASF splicing factor has been demonstrated in vitro and
in the yeast two-hybrid assay (Wu and Maniatis 1993;
Amrein et al. 1994). It was also demonstrated in
human cell extracts that Tra2 recognizes purine-rich
enhancer elements and recruits to these elements 9G8
and SF2/ASF proteins (Lynch and Maniatis 1996).
TaRSZ38 appears to be a plant specific member of a
similar protein complex in the plant spliceosome,
which might be responsible for plant specific regulation
of constitutive and/or alternative splicing.
It was also found that TaRSZ38 can directly and
strongly interact with TaU1-70K and TaLuc7 proteins.
U1-70K from metazoans has been shown to interact
with SR-splicing factors SC35, ASF/SF2 and Sip1, and is
involved in both basic and alternative splicing (Wu and
RDE
TaU2AF65
RS RSTaTra2 P
PRP38 RDH RSKTaPrp38
LUC7 RDRSTaLuc7
TaU1-70K
RDH
TaTRN-SR
TaRSZBP1
RSTaSRp30
7X(-GRRRSSRREDHSSDG-)
RSTaRSZ22 ZFRRM
RRM
RRM ψRRM
RRM
RRM RRM RRM
RSDETaU2AF35 ψRRM
RSPTaY14 RRM
C2H2C3H
Ran-BD RS-BD
RG RG
C3H
TaDRH1 RGRSHELICDEAD
Fig. 6 Domain structure of the proteins identified in yeast two-hybrid screens: RRM, RNA recognition motif; YRRM, stronglydegenerated version of RRM; TD, transmembrane domain;Kinase domain, S/T kinase active center; Ran-BD, Ran-GTP-binding domain; RS-BD, SR-protein binding domain; ZF, CCHC-type zinc fingers; C3H and C2H2, CCCH and CCHH zinc fingers;HELIC, RNA helicase domain; DEAD, signature sequence ofDEAD box RNA helicases; LUC7 and PRP38, conserved
domains within the families of proteins with unknown function;P, proline-rich sequence usually with proline stretches; RS, RSK,RDRS, RDE, RSP and RGRS, usually protein binding domains(sequences) enriched in particular amino acids with repeateddipeptides containing arginines; 7X(-GRRRSSRREDHSSDG-),seven near perfect repeats of 15 amino acids. The length ofproteins deduced from clones obtained in the yeast two-hybridscreen is indicated with arrows
Plant Mol Biol (2006) 62:637–653 649
123

Maniatis 1993; Kohtz et al. 1994; Manley and Tacke
1996). The homologue of U1-70K from Arabidopsis can
interact in vivo and in vitro with small RSZ22-like
proteins and SC35-like proteins (Golovkin and Reddy
1998). Luc7p is a component of the yeast U1 small
nuclear ribonucleoprotein particle (snRNP) and is
essential for the formation of the spliceosome com-
mitment complex in vitro (Fortes et al. 1999). In the
absence of Luc7p the protein composition of the U1
snRNP is changed, splicing efficiency is reduced and 5¢splice site selection is altered. Isolation of plant ho-
mologues of Luc7p had not been reported previously.
Interaction of TaRSZ38 with two proteins bound to
the U1 snRNP suggests involvement of this protein in
the early steps of spliceosome assembly and in the
recognition of the 5¢ splice site. However, interaction
of TaRSZ38 with another protein, designated TaPrp38,
suggests that TaRSZ38 participates in splicing at least
until the end of cleavage of the first exon. Prp38p is an
essential splicing factor in yeast, as a mutation in the
PRP38 gene is lethal. In the temperature-sensitive
mutant prp38, high temperature prevents the first
cleavage-ligation event in excision of introns from
mRNA precursors (Blanton et al. 1992). In the ab-
sence of Prp38p activity, spliceosomes form but they
arrest in a catalytically impaired state (Xie et al. 1998).
Nothing is known about homologues of Prp38p in
multicellular organisms, although database searches
for nematodes indicate embryo specific expression of a
related protein, for which RNAi silencing caused early
embryo defects, reabsorbed polar bodies, and embry-
onic lethality (see annotation to the gene product
Accession NP_505762). TaPrp38 is also expressed in
wheat embryos, as we could amplify the full-length
cDNA using a library prepared from zygotic embryos
isolated at 5 DAP. An important role for TaPrp38 in
endosperm development is supported by the relative
abundance of TaPrp38 among clones isolated in the
yeast two-hybrid screen from the wheat endosperm
cDNA library (90% of all clones) and 13- to 20-fold
higher abundance of TaPrp38 mRNA in the liquid
endosperm in comparison with the rest of the grain.
An additional involvement of TaRSZ38 in 3¢ splice
site regulation cannot be excluded, since it interacts
U2AF35
A
E1 E2Branchpoint
Polypyrimidine tract
5’ splicesite
3’ splicesite
A UUUUU
U1 snRNP U2 snRNPTra2U2AF35
SRpU1-70K
Luc7
Capping complex recruitment
Enhancer
U5 snRNP
U6 snRNP
U4 snRNP
Prp38
UUUUUA
U1 snRNP
U4 snRNP
U2 snRNPU6 snRNP
U5 snRNPPrp38U6/U4 unwinding
UUUUU
UUUUUA
Pre-mRNA
Pre-spliceosome
Spliceosome
Spliced mRNA Lariat intron
+
U2 snRNP recruitment andalternative splice site selection
U1 snRNPrecruitment andalternative splice site selection
RSZ22
Y14
Luc7
U1-70K
Tra2
Prp38
TaLuc7
TaU1-70K
TaU2AF65
TaTra2
TaPrp38
U2AF35
U2AF65
TaU2AF35
SRp TaSF2/ASF-like(TaSRp30, TaSRp30a)
RSZ22 TaRSZ22-like(TaRSZ22, TaRSZ22a)
Y14 TaY14
Proteins isolated with TaRSZ38
Proteins isolated with TaTra2 andTaTRN-SR
mRNA export
U2AF35
Fig. 7 Schematic representation of a typical splicing reaction, showing the deduced positions and functions of proteins identified in theyeast two-hybrid screen
650 Plant Mol Biol (2006) 62:637–653
123

with U2AF65, the large subunit of the U2 auxiliary
factor. Animal U2AF65 recruits U2 snRNPs by inter-
action with U2 snRNP components (Gozani et al.
1998; Wang et al. 1998a). The polypyrimidine tract,
which is located on the intron sequence between the
branch point and 3¢ splice site, is recognized by the
large subunit of U2AF (Zamore et al. 1992).
In the two screens with TaRSZ38 and TaTra2 as
baits the same partial clones were isolated that encode
a protein with a high identity to Mtr10 from yeast and
animal TRN-SR. It has been demonstrated that nu-
clear import of splicing factors with SR domains is
mediated by a specific import receptor, termed trans-
portin-SR (TRN-SR). TRN-SR is a member of the
importin b family and it binds directly, without the help
of any a subunit, to nuclear localization signals (NLSs)
within SR domains (Kataoka et al. 1999; Lai et al.
2000). Using a partial sequence of TaTRN-SR as bait it
was found that this protein identifies several other
putative splicing factors and factors involved in mRNA
export. These include cDNA clones encoding both
subunits of U2AF splicing factor, an RNA-helicase, a
homologue of the animal Y14 protein and several
partial clones encoding unidentified proteins contain-
ing SD-rich sequences.
Mtr10p is the closest yeast homologue to TRN-SR.
This is a nuclear import receptor specialized in the
transport of RNA-binding proteins. A mutation in
MTR10 also causes accumulation of poly(A)+ RNA in
the nucleus (Kadowaki et al. 1994) and the question
arises if Mtr10p can be either directly or indirectly (by
delivery of export factors to the nucleus) involved in
mRNA export. The participation also of TaTRN-SR in
mRNA export cannot be excluded, as it interacted with
both Y14 and subunits of U2AF, known to be involved
in mRNA export (Le Hir et al. 2001; Zolotukhin et al.
2002; Singh and Lykke-Andersen, 2003; Blanchette
et al. 2004) and the cytoplasmic nonsense mediated
decay complex (Gatfield et al. 2003; Gehring et al.
2003).
This study has identified a number of proteins with
putative roles in RNA splicing during early grain
development in wheat. Figure 7 shows a schematic
representation of a typical splicing reaction, showing
the deduced positions and functions of the wheat
proteins identified in the yeast two-hybrid experiments.
The developing wheat grain is an interesting model for
this type of study. The transition from liquid to cellu-
larized endosperm is a key process in grain develop-
ment and modification of this may lead to alterations in
grain shape, size and composition. It seems probable
that RNA splicing is one component of the regulatory
machinery used for this transition. The size of the
wheat grain makes the analysis of endosperm devel-
opment far simpler than for Arabidopsis and, impor-
tantly, the uncellularized, liquid endosperm or
syncytium can be isolated and used as a source for
components of the splicing apparatus. Ultimately the
liquid endosperm may allow the development of an in
vitro splicing assay.
Acknowledgments We thank Ursula Langridge for assistancewith growing plants in the glasshouse. This work was supportedby the Australian Grain Research and Development Corpora-tion (grant no. UA00083 to P.L.).
References
Abler ML, Green PJ (1996) Control of mRNA stability in higherplants. Plant Mol Biol 32:63–78
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z,Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search pro-grams. Nucleic Acids Res 25:3389–3402
Amrein H, Hedley ML, Maniatis T (1994) The role of specificprotein-RNA and protein–protein interactions in positiveand negative control of pre-messenger-RNA splicing bytransformer-2. Cell 76:735–746
Barrass JD, Beggs JD (2003) Splicing goes global. Trends Genet19:295–298
Ben-Yehuda S, Dix I, Russell CS, McGarvey M, Beggs JD,Kupiec M (2000) Genetic and physical interactions betweenfactors involved in both cell cycle progression and pre-mRNA splicing in Saccharomyces cerevisiae. Genetics156:1503–1517
Blanchette M, Labourier E, Green RE, Brenner SE, Rio DC(2004) Genome-wide analysis reveals an unexpected func-tion for the Drosophila splicing factor U2AF(50) in thenuclear export of intronless mRNAs. Mol Cell 14:775–786
Blanton S, Srinivasan A, Rymond BC (1992) PRP38 encodes ayeast protein required for pre-mRNA splicing and mainte-nance of stable U6 small nuclear RNA levels. Mol Cell Biol12:3939–3947
Blencowe BJ, Bowman JAL, McCracken S, Rosonina E (1999)SR-related proteins and the processing of messenger RNAprecursors. Biochem Cell Biol 77:277–291
Burton RA, Shirley NJ, King BJ, Harvey AJ, Fincher GB (2004)The CesA gene family of barley. Quantitative analysis oftranscripts reveals two groups of co-expressed genes. PlantPhysiol 134:224–236
Chua YL, Brown AP, Gray JC (2001) Targeted histone acety-lation and altered nuclease accessibility over short regionsof the pea plastocyanin gene. Plant Cell 13:599–612
Domon C, Lorkovic ZJ, Valcarcel J, Filipowicz W (1998) Mul-tiple forms of the U2 small nuclear ribonucleoprotein aux-iliary factor U2AF subunits expressed in higher plants. JBiol Chem 273:34603–34610
Ewing B, Green P (2000) Analysis of expressed sequence tagsindicates 35,000 human genes. Nat Genet 25:232–234
Fang Y, Hearn S, Spector DL (2004) Tissue-specific expressionand dynamic organization of SR splicing factors in Arabid-opsis. Mol Biol Cell 15:2664–2673
Plant Mol Biol (2006) 62:637–653 651
123

Fortes P, Bilbao-Cortes D, Fornerod M, Rigaut G, Raymond W,Seraphin B, Mattaj IW (1999) Luc7p, a novel yeast U1snRNP protein with role in 5¢ splice site recognition. GeneDev 13:2425–2438
Fromont-Racine M, Rain JC, Legrain P (1997) Toward a func-tional analysis of the yeast genome through exhaustive two-hybrid screens. Nat Genet 16:277–282
Fu XD, Maniatis T (1992) Isolation of a complementary-DNAthat encodes the mammalian splicing factor SC35. Science256:535–538
Gatfield D, Unterholzner L, Ciccarelli FD, Bork P, Izaurralde E(2003) Nonsense-mediated mRNA decay in Drosophila: atthe intersection of the yeast and mammalian pathways.EMBO J 22:3960–3970
Gehring NH, Neu-Yilik G, Schell T, Hentze MW, Kulozik AE(2003) Y14 and hUpf3b form an NMD-activating complex.Mol Cell 11:939–949
Golovkin M, Reddy ASN (1996) Structure and expression of aplant U1 snRNP 70K gene: alternative splicing of U1snRNP 70K pre-mRNAs produces two different transcripts.Plant Cell 8:1421–1435
Golovkin M, Reddy ASN (1998) The plant U1 small nuclearribonucleoprotein particle 70K protein interacts with twonovel serine/arginine-rich proteins. Plant Cell 10:1637–1647
Golovkin M, Reddy ASN (1999) An SC35-like protein and anovel serine/arginine-rich protein interact with ArabidopsisU1-70K protein. J Biol Chem 274:36428–36438
Golovkin M, Reddy ASN (2003) Expression of U1 small nuclearribonucleoprotein 70K antisense transcript using APET-ALA3 promoter suppresses the development of sepals andpetals. Plant Physiol 132:1884–1891
Gozani O, Potashkin J, Reed R (1998) A potential role forU2AF-SAP 155 interactions in recruiting U2 snRNP to thebranch site. Mol Cell Biol 18:4752–4760
Groth C, Lardelli M (2002) The structure and function of ver-tebrate fibroblast growth factor receptor 1. Int J Dev Biol46:393–400
Halterman DA, Wei F, Wise RP (2003) Powdery mildew-in-duced Mla mRNAs are alternatively spliced and containmultiple upstream open reading frames. Plant Physiol131:558–567
Hanamura A, Caceres JF, Mayeda A, Franza BR, Krainer AR(1998) Regulated tissue-specific expression of antagonisticpre-mRNA splicing factors. RNA 4:430–444
Iggo RD, Jamieson DJ, Macneill SA, Southgate J, McPheat J,Lane DP (1991) P68 RNA helicase – identification of anucleolar form and cloning of related genes containing aconserved intron in yeasts. Mol Cell Biol 11:1326–1333
Kadowaki T, Hitomi M, Chen S, Tartakoff AM (1994) NuclearmRNA accumulation causes nucleolar fragmentation inyeast mtr2 mutant. Mol Biol Cell 5:1253–1263
Kalyna M, Lopato S, Barta A (2003) Ectopic expression ofatRSZ33 reveals its function in splicing and causespleiotropic changes in development. Mol Biol Cell14:3565–3577
Kataoka N, Bachorik JL, Dreyfuss G (1999) Transportin-SR, anuclear import receptor for SR proteins. J Cell Biol145:1145–1152
Kataoka N, Yong J, Kim VN, Velazquez F, Perkinson RA, WangF, Dreyfuss G (2000) Pre-mRNA splicing imprints mRNAin the nucleus with a novel RNA-binding protein that per-sists in the cytoplasm. Mol Cell 6:673–682
Kawaguchi R, Bailey-Serres J (2002) Regulation of translationalinitiation in plants. Curr Opin Plant Biol 5:460–465
Kohtz JD, Jamison SF, Will CL, Zuo P, Luhrmann R, Garcia-blanco MA, Manley JL (1994) Protein–protein interactions
and 5¢-splice-site recognition in mammalian messenger-RNA precursors. Nature 368:119–124
Kriventseva EV, Koch I, Apweiler R, Vingron M, Bork P,Gelfand MS, Sunyaev S (2003) Increase of functionaldiversity by alternative splicing. Trends Genet 19:124–128
Kuroyanagi N, Onogi H, Wakabayashi T, Hagiwara M (1998)Novel SR-protein-specific kinase, SRPK2, disassembles nu-clear speckles. Biochem Bioph Res Co 242:357–364
Lai MC, Lin RI, Huang SY, Tsai CW, Tarn WY (2000) A humanimportin-beta family protein, transportin-SR2, interactswith the phosphorylated RS domain of SR proteins. J BiolChem 275:7950–7957
Lalli E, Ohe K, Latorre E, Bianchi ME, Sassone-Corsi P (2003)Sexy splicing: regulatory interplays governing sex determi-nation from Drosophila to mammals. J Cell Sci 116:441–445
Lazar G, Schaal T, Maniatis T, Goodman HM (1995) Identifi-cation of a plant serine-arginine-rich protein similar to themammalian splicing factor SF2/ASF. Proc Natl Acad SciUSA 92:7672–7676
Le Hir H, Gatfield D, Izaurralde E, Moore MJ (2001) The exon-exon junction complex provides a binding platform forfactors involved in mRNA export and nonsense-mediatedmRNA decay. EMBO J 20: 4987–4997
Lei EP, Silver PA (2002) Protein and RNA export from thenucleus. Dev Cell 2:261–272
Li S, Armstrong CM, Bertin N, Ge H, Milstein S, Boxem M,Vidalain PO, Han JD, Chesneau A, Hao T, Goldberg DS, LiN, Martinez M, Rual JF, Lamesch P, Xu L, Tewari M, WongSL, Zhang LV, Berriz GF, Jacotot L, Vaglio P, Reboul J,Hirozane-Kishikawa T, Li Q, Gabel HW, Elewa A, Baum-gartner B, Rose DJ, Yu H, Bosak S, Sequerra R, Fraser A,Mango SE, Saxton WM, Strome S, Van Den Heuvel S,Piano F, Vandenhaute J, Sardet C, Gerstein M, Doucette-Stamm L, Gunsalus KC, Harper JW, Cusick ME, Roth FP,Hill DE, Vidal M (2004) A map of the interactome networkof the metazoan C. elegans. Science 303:540–543
Locatelli F, Bracale M, Magaraggia F, Faoro F, Manzocchi LA,Coraggio I (2000) The product of the rice myb7 unsplicedmRNA dimerizes with the maize leucine zipper Opaque2and stimulates its activity in a transient expression assay.J Biol Chem 275:17619–17625
Lopato S, Gattoni R, Fabini G, Stevenin J, Barta A (1999a) Anovel family of plant splicing factors with a Zn knucklemotif: examination of RNA binding and splicing activities.Plant Mol Biol 39:761–773
Lopato S, Kalyna M, Dorner S, Kobayashi R, Krainer AR, BartaA (1999b) atSRp30, one of two SF2/ASF-like proteins fromArabidopsis thaliana, regulates splicing of specific plantgenes. Gene Dev 13:987–1001
Lopato S, Forstner C, Kalyna M, Hilscher J, Langhammer U,Indrapichate K, Lorkovic ZJ, Barta A (2002) Network ofinteractions of a novel plant-specific Arg/Ser-rich protein,atRSZ33, with atSC35-like splicing factors. J Biol Chem277:39989–39998
Lopez AJ (1998) Alternative splicing of pre-mRNA: develop-mental consequences and mechanisms of regulation. AnnuRev Genet 32:279–305
Lorkovic ZJ, Barta A (2002) Genome analysis: RNA recognitionmotif (RRM) and K homology (KH) domain RNA-bindingproteins from the flowering plant Arabidopsis thaliana.Nucleic Acids Res 30:623–635
Lorkovic ZJ, Lopato S, Pexa M, Lehner R, Barta A (2004)Interactions of Arabidopsis RS domain containing cyclo-philins with SR proteins and U1 and U11 snRNP-specificproteins suggest their involvement in pre-mRNA splicing. JBiol Chem 279:33890–33898
652 Plant Mol Biol (2006) 62:637–653
123

Lynch KW, Maniatis T (1996) Assembly of specific SR proteincomplexes on distinct regulatory elements of the Drosophiladoublesex splicing enhancer. Gene Dev 10:2089–2101
Magaraggia F, Solinas G, Valle G, Giovinazzo G, Coraggio I(1997) Maturation and translation mechanisms involved inthe expression of a myb gene of rice. Plant Mol Biol35:1003–1008
Manley JL, Tacke R (1996) SR proteins and splicing control.Gene Dev 10:1569–1579
Mohr SE, Dillon ST, Boswell RE (2001) The RNA-bindingprotein Tsunagi interacts with Mago Nashi to establishpolarity and localize oskar mRNA during Drosophilaoogenesis. Gene Dev 15:2886–2899
Montag K, Salamini F, Thompson RD (1995) ZEMa, a memberof a novel group of MADS box genes, is alternatively splicedin maize endosperm. Nucleic Acids Res 23:2168–2177
Mortillaro MJ, Berezney R (1998) Matrin CYP, an SR-rich cy-clophilin that associates with the nuclear matrix and splicingfactors. J Biol Chem 273:8183–8192
Mount SM, Salz HK (2000) Pre-messenger RNA processingfactors in the Drosophila genome. J Cell Biol 150:F37-F43
Murray MV, Kobayashi R, Krainer AR (1999) The type 2C Ser/Thr phosphatase PP2C gamma is a pre-mRNA splicingfactor. Gene Dev 13:87–97
Nash J, Walbot V (1992) Bronze-2 gene-expression and intronsplicing patterns in cells and tissues of Zea-Mays L. PlantPhysiol 100:464–471
Okanami M, Meshi T, Iwabuchi M (1998) Characterization of aDEAD box ATPase/RNA helicase protein of Arabidopsisthaliana. Nucleic Acids Res 26:2638–2643
Quesada V, Macknight R, Dean C, Simpson GG (2003) Auto-regulation of FCA pre-mRNA processing controls Arabid-opsis flowering time. EMBO J 22:3142–3152
Reddy ASN (2001) Nuclear pre-mRNA splicing in plants. CritRev Plant Sci 20:523–571
Reddy ASN (2004) Plant serine/arginine-rich proteins and theirrole in pre-mRNA splicing. Trends Plant Sci 9:541–547
Rydberg B, Spur B, Karran B (1990) cDNA cloning and chro-mosomal assignment of the human O6-methylguanine-DNAmethyltransferase. cDNA expression in Escherichia coli andgene expression in human cells. J Biol Chem 265:9563–9569
Ryner LC, Baker BS (1991) Regulation of Doublesex pre-mes-senger-RNA processing occurs by 3¢-splice site activation.Gene Dev 5:2071–2085
Savaldi-Goldstein S, Aviv D, Davydov O, Fluhr R (2003)Alternative splicing modulation by a LAMMER kinaseimpinges on developmental and transcriptome expression.Plant Cell 15:926–938
Screaton GR, Caceres JF, Mayeda A, Bel MV, Plebanski M,Jackson DG, Bell JI, Krainer AR (1995) Identification andcharacterization of 3 members of the human SR family ofpre-messenger-RNA splicing factors. EMBO J 14:4336–4349
Senger B, Simos G, Bischoff FR, Podtelejnikov A, Mann M,Hurt E (1998) Mtr10p functions as a nuclear import receptorfor the mRNA-binding protein Np13p. EMBO J 17:2196–2207
Siebel CW, Feng LN, Guthrie C, Fu XD (1999) Conservation inbudding yeast of a kinase specific for SR splicing factors.Proc Natl Acad Sci USA 96:5440–5445
Singh G, Lykke-Andersen J (2003) New insights into the for-mation of active nonsense-mediated decay complexes.Trends Biochem Sci 28:464–466
Stanyon CA, Finley RL Jr (2000) Progress and potential ofDrosophila protein interaction maps. Pharmacogenomics1:417–431
Sun YJ, Flannigan BA, Madison JT, Setter TL (1997) Alterna-tive splicing of cyclin transcripts in maize endosperm. Gene195:167–175
Tacke R, Tohyama M, Ogawa S, Manley JL (1998) Human Tra2proteins are sequence-specific activators of pre-mRNAsplicing. Cell 93:139–148
Tuteja N (2000) Plant cell and viral helicases: essential enzymesfor nucleic acid transactions. Crit Rev Plant Sci 19:449–478
Wang CY, Chua K, Seghezzi W, Lees E, Gozani O, Reed R(1998a) Phosphorylation of spliceosomal protein SAP 155coupled with splicing catalysis. Gene Dev 12:1409–1414
Wang HY, Lin W, Dyck JA, Yeakley JM, Zhou SY, Cantley LC,Fu XD (1998b) SRPK2: a differentially expressed SR pro-tein-specific kinase involved in mediating the interactionand localization of pre-mRNA splicing factors in mamma-lian cells. J Cell Biol 140:737–750
Weber H, Borisjuk L, Heim U, Buchner P, Wobus U (1995) Seedcoat-associated invertases of fava bean control bothunloading and storage functions: cloning of cDNAs and celltype-specific expression. Plant Cell 7:1835–1846
Wu JY, Maniatis T (1993) Specific interactions between proteinsimplicated in splice-site selection and regulated alternativesplicing. Cell 75:1061–1070
Xie J, Beickman K, Otte E, Rymond BC (1998) Progressionthrough the spliceosome cycle requires Prp38p function forU4/U6 snRNA dissociation. EMBO J 17:2938–2946
Zamore PD, Patton JG, Green MR (1992) Cloning and domain-structure of the mammalian splicing factor U2AF. Nature355:609–614
Zhang WQ, Yuan YF, Wang YN, Yuan Z, Mi HF, He BL (2002)Human cyclophilin 33 (hCyP33) in T-cell binds specificallyto poly(A)(+)RNA (mRNA). Sci China Ser B 45:275–281
Zhou Z, Licklider LJ, Gygi SP, Reed R, Sim J, Griffith J (2002)Comprehensive proteomic analysis of the human spliceo-some: purification and electron microscopic visualization offunctional human spliceosomes. Nature 419:182–185
Zolotukhin AS, Tan W, Bear J, Smulevitch S, Felber BK (2002)U2AF participates in the binding of TAP (NXF1) tomRNA. J Biol Chem 277:3935–3942
Plant Mol Biol (2006) 62:637–653 653
123




















