
Radiation fields, dosimetry, biokinetics and biophysical ...

Biochemical and Biophysical Research Communications 399 (2010) 505–511
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
Substrate cleavage pattern, biophysical characterization and low-resolutionstructure of a novel hyperthermostable arabinanase from Thermotoga petrophila
Fabio M. Squina a,*, Camila R. Santos b, Daniela A. Ribeiro a, Júnio Cota a,c, Renata R. de Oliveira b,Roberto Ruller a, Andrew Mort d, Mario T. Murakami b, Rolf A. Prade e,**
a Laboratório Nacional de Ciência e Tecnologia do Bioetanol (CTBE), do Centro Nacional de Pesquisa em Energia e Materiais (CNPEM), Campinas, SP, Brazilb Laboratório Nacional de Biociências (LNBio), do Centro Nacional de Pesquisa em Energia e Materiais (CNPEM), Campinas, SP, Brazilc Departamento de Ciências de Alimentos da Universidade Estadual de Campinas, SP, Brazild Department of Biochemistry, Oklahoma State University, Stillwater, OK 74078, USAe Department of Microbiology and Molecular Genetics, Oklahoma State University, Stillwater, OK 74078, USA
a r t i c l e i n f o
Article history:Received 25 June 2010Available online 1 August 2010
Keywords:ArabinanaseCapillary zone electrophoresisSmall angle X-ray scattering (SAXS)Hyperthermostable enzymeArabinan
0006-291X/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.bbrc.2010.07.097
* Corresponding author. Address: Rua Giuseppe Máde Alta Tecnologia, Caixa Postal 6170 – Campinas-SP, CBrazil. Tel.: +55 19 3518 3104.** Corresponding author.
E-mail address: [email protected] (F.M.
a b s t r a c t
Arabinan is a plant structural polysaccharide degraded by two enzymes; a-L-arabinofuranosidase andendo-1,5-a-L-arabinanase. These enzymes are highly diversified in nature, however, little is known abouttheir biochemical and biophysical properties. We have characterized a novel arabinanase (AbnA) isolatedfrom Thermotoga petrophila with unique thermostable properties such as the insignificant decrease ofresidual activity after incubation up to 90 �C. We determined the AbnA mode of operation through cap-illary zone electrophoresis, which accumulates arabinotriose and arabinobiose as end products afterhydrolysis of arabinan-containing polysaccharides. Spectroscopic analyses by Far-UV circular dichroismand intrinsic tryptophan fluorescence emission demonstrated that AbnA is folded and formed mainly byb-sheet structural elements. In silico molecular modeling showed that the AbnA structure encompasses afive-bladed b-propeller catalytic core juxtaposed by distorted up-and-down b-barrel domain. The low-resolution structure determined by small angle X-ray scattering indicated that AbnA is monomeric insolution and its molecular shape is in full agreement with the model.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
Plant structural polysaccharides are the most abundant sourceof renewable carbon in the biosphere and represent a valuableindustrial substrate in several applications, such as bio-energy pro-duction, pulp and paper, food technology, detergent, textile, nutri-tional or medical research and organic synthesis [7]. Aiming at thedevelopment of sugar-platform based chemicals; plant biomasshydrolytic enzymes are the key technological component for effi-cient use of renewable feedstocks through an environmentalfriendly bioconversion route.
Arabinan is composed of an a-1–5 linked L-arabinofuranosylresidue backbone, substituted with a-1,3 and a-1,2 linked chainsof L-arabinofuranosyl units. They occur as either homoglycans, ara-binans, or as heteroglycans such as arabinoxylans and arabinoga-lactans [25]. Two major enzyme families are involved in arabinanhydrolysis, the a-L-arabinofuranosidases (ABFs) (EC 3.2.1.55),
ll rights reserved.
ximo Scolfaro, 10.000 Polo IIEP: 13083-970 Campinas, SP,
Squina).
which remove arabinose side chains, allowing the access of endo-1,5-a-L-arabinanase (ABNs) (EC 3.2.1.99) that releases arabino-oli-gosaccharides and L-arabinose as hydrolysis products. Arabinasesand arabinofuranosidases are classified by CAZy [5,6] as beingmembers of glycoside hydrolase families GH3, GH43, GH51,GH54 and GH62, which display a wide range of enzymatic activi-ties. Only little information regarding biochemical and biophysicalproperties is available.
To date, crystal structures from bacterial family GH51 arabino-furanosidases (EC 3.2.1.55) have been published from Geobacillusstearothermophilus [9], Clostridium thermocellum [23] and Thermo-bacillus xylanilyticus [15], and crystal structures from arabinasesof the related family GH43 (EC 3.2.1.99) have been determinedfrom Cellvibrio japonicus [13], Bacillus subtillis [17] and G. stearo-thermophilus [1].
Members from CAZy GH43 family shows three conserved acidicresidues, two aspartic acids and one glutamic acid, located at theactive site providing the general acid and base components forglycosidic bond hydrolysis, with inversion of the anomeric config-uration [16]. These catalytic residues are close in space in the five-bladed beta-propeller fold, which was structurally determined forABN from C. japonicus [13] and B. subtilis [17]. A long V-shaped sur-face groove, partially enclosed at one end, forms a single extended

506 F.M. Squina et al. / Biochemical and Biophysical Research Communications 399 (2010) 505–511
substrate-binding surface across the face of the propeller, whichcontributes to the arabinan exo-activity [13], whereas the bindingsite is open at both ends in endo acting polysaccharidases [3].
In this study, we report the characterization of a novel high-temperature operating and thermostable endo-1–5-a-arabinanasewith potential biotechnological applications [14,18]. The protein-coding portion of the ABN gene was isolated from Thermotoga pet-rophila, a hyper thermophilic bacterium that grows optimally at80 �C [21], and introduced into an Escherichia coli for over-expres-sion of the recombinant protein. Here, we show a detailed descrip-tion of the enzymatic specificity and the mode of operation of therecombinant enzyme along with biophysical and structural in-sights over enzyme hyperthermophilicity.
2. Material and methods
2.1. Cloning of the endo-1–4-a-arabinanase gene from T. petrophila
The full-length coding GH43 gene (Tpet_0627) from T. petrophil-a (AbnA), cloned into pBAD/Myc-His B vector (Invitrogen) [20], wasused as template for standard PCR methods for cloning the matureenzyme without signal peptide. The primer sets used for amplifica-tion was 50- GGCCGGGGATCCGAACAACCCACCTTTCGATG -30 and 50
GGCCGGGGTACCTTATTCTTCCACTCTTATTCCCC 30. This set of prim-ers generated the gene segment for cloning into the pQTEV expres-sion vector, which carries a fusion sequence encoding 6 � Hisresidues at the N-terminus of the expressed protein.
2.2. Over-expression and purification
E. coli BL21 (Novagen) cells containing the fusion protein vectorwas inoculated into LB medium at 37 �C and induced for 4 h byadding 1 mM of IPTG when the OD600 reached 0.5–0.6. The cellswere harvested by centrifugation and resuspended in lysis buffer(50 mM Tris–HCl pH 7.5 100 mM NaCl). The suspension was soni-cated for 30 s at 4 �C after treatment with 0.5 mg ml�1 lysozymeand 50 lg ml�1 DNase I. The solution was centrifuged at 10,000gfor 30 min and the supernatant was loaded onto a Ni2+-chelatingaffinity column (GE Healthcare). The elution was carried out by anon-linear gradient of 0.0–0.5 M imidazole in lysis buffer and thetarget protein was eluted with 100 mM imidazole. To attain ahomogenous sample, the sample was further loaded on a gel filtra-tion Superdex 75 column (GE Healthcare), which had been pre-equilibrated with a 25 mM Tris–HCl buffer (pH 7.5). The purifiedAbnA was further analyzed by SDS–PAGE. The protein contentwas assessed using the method of Bradford [26].
2.3. Enzyme characterization
The enzymatic assays for ABN were performed following Squinaet al. [19], where 50 ll of substrate solution (0.5% polysaccharidecontent) in 50 mM citrate buffer (pH 6) was incubated with 25 llof diluted enzyme for 30 min at 75 �C. The enzymatic activityand substrate specificity were determined from the amount ofreducing sugar liberated from different polysaccharide substrates(purchased from Megazyme, Ireland and Sigma–Aldrich, USA) asdetermined by the DNS method [12]. One unit of enzyme was de-fined as the quantity of enzyme that liberated reducing sugar atrate of 1 lmol/min. To determine the optimum pH and tempera-ture profiles, the enzymatic reaction was carried out at differentpHs in several buffer systems and various temperatures as indi-cated in the figure legends. For thermostability evaluation, enzymewas incubated at 70, 80, 90 and 95 �C. After, an aliquot of enzymewas taken and the residual activity was measured with the specificpolysaccharide substrate. The kinetic parameters for ABN activity
were calculated from initial velocities at eight substrate concentra-tions in the 2.5–25 mg/ml range in citrate buffer at pH 6 and 75 �C.The assays were performed in triplicate and at least three indepen-dent experiments were carried out.
2.4. Capillary zone electrophoresis of oligosaccharides
Arabinoheptaose (purchase form Megazyme) was derivatizedwith 8-aminopyreno-1,3,6-trisulfonic acid (APTS) by reductiveamination as described by Naran et al. [2,27] Enzymatic reactionswere performed as described for standard enzymatic activity,except that 1 pM of APTS-labeled arabinoheptaose was used as asubstrate. Capillary zone electrophoresis of arabinoheptaosebreakdown products was performed on a BioFocus 2000 (Bio-RadLaboratories,) with laser-induced fluorescence detection. A fused-silica capillary (TSP050375, Polymicro Technologies) of internaldiameter 50 lM and length 31 cm was used as the separation col-umn for oligosaccharides. Samples were injected by application of4.5 lb/in2 20 of helium pressure for 0.22 s. Electrophoresis condi-tions were 15 kV/70–100 lA with the cathode at the inlet, 0.1 Msodium phosphate, pH 2.5, as running buffer, and a controlled tem-perature of 20 �C. The capillary was rinsed with 1 M NaOH fol-lowed by running buffer to prevent carryovers. APTS-labeledoligomers were excited at 488 nm and emission was collectedthrough a 520-nm band pass filter. Because of the small volumesof capillary electrophoresis combined small variations in bufferstrength, retention times vary slightly when comparing separateelectrophoresis runs. Since, we used a pure and size defined sub-strate, arabinoheptaose, retention time variations were not cor-rected in Fig. 2.
2.5. Spectroscopic methods
Dynamic light-scattering experiments were carried out using aDynaPro 810 (Protein Solutions) apparatus equipped with a Peltiermodule for temperature control. The wavelength of the laser lightand the output power were set at 830 nm and 30 mW, respectively.Hydrodynamic parameters were determined using the softwareDYNAMICS v.6.3.40.
Far-UV circular dichroism (CD) measurements (190–260 nm)were carried out using a JASCO 810 spectropolarimeter (JASCOInc., Tokyo, Japan) with a 2.0-nm bandwidth, 1 nm resolution,and 1 s response time. A 1 mm pathlength cuvette was used. Re-sults are expressed as mean residual molar ellipticity indeg cm2 dmol�1. The prediction of secondary structure from thedichroism circular spectrum of recombinant AbnA was obtainedusing DICHROWEB server [28].
Intrinsic tryptophan fluorescence emission (IFTE) spectra weremeasured using a Cary Varian spectrofluorimeter (Varian, USA) be-tween 300 and 450 nm using an excitation wavelength of 295 nm.The excitation and emission slit widths fixed at 4 nm and the pho-tomultiplier tube voltage was 700 V.
Blank corrections were made in all spectra. All spectroscopicmeasurements were carried out with a sample prepared in 20 mMphosphate buffer pH 6.0 at the final concentration of 0.1 mg/ml.
2.6. Small angle X-ray scattering
Small angle X-ray scattering (SAXS) data were collected on theSAXS2 beamline at the Brazilian Synchrotron Light Laboratory. Theradiation wavelength was set to 1.48 Å and a 165 mm MarCCDdetector was used to record the scattering patterns. The sample-to-detector distance was set to 1559.13 mm to give a scatteringvector range from 0.2 to 2.2 nm�1. Protein samples were preparedin 20 mM phosphate buffer (pH 6.0) at 4 mg/ml�1. Fitting of theexperimental data and evaluation of the pair–distance distribution

100
(%)
0
25
50
75
100
30 40 50 60 70 80 90 100 110
Rel
ativ
e A
ctiv
ity (%
)
Temperature (ºC)
A
B
F.M. Squina et al. / Biochemical and Biophysical Research Communications 399 (2010) 505–511 507
function p(r) were performed using the program GNOM [35]. Thelow-resolution envelope of the AbnA was determined using ab ini-tio modeling as implemented in the program DAMMIN [36]. Anaverage model was generated from several runs using the suiteof programs DAMAVER [29]. The low-resolution model and themodeled structure were superimposed using the program SUP-COMB [29].
2.7. Homology molecular modeling
The crystal structure of a GH43 arabinanase from B. lichenifor-mis (PDB entry code: 3LV4) was used as 3D-template for re-straint-based modeling as implemented in the MODELLERprogram [30]. The overall and local quality analyses of the finalmodel were assessed by VERIFY3D [31], PROSA [37] and PRO-CHECK [32]. Three-dimensional structures were displayed, ana-lyzed and compared using the program COOT [33].
0
25
50
75
1 2 3 4 5 6 7 8 9 10 11 12
Rel
ativ
e Ac
tivity
pH
0
25
50
75
100
0 100 200 300 400 500 600Time (min)
80ºC90ºC95ºC
Rel
ativ
e Ac
tivity
(%)
C
Fig. 1. Influence of temperature on the AbnA activity. (A) The enzyme activity wasassayed in different buffer systems (B) indicted as solid line, in the range of 2.2–8.0using the McIlvaine buffer, and dashed line for the relative activity in phosphatebuffer (pH 2.0), citric acid buffer (pH 3 and 4), MES buffer (pH 5.5 and 6), MOPSbuffer (pH 7), arginine buffer (pH 9 and 12) and CAPS buffer (pH 10) Thermalinactivation (C) of the ABN was carried out at 95 (triangle), 90 (circle) and 80 �C(square).
3. Results and discussion
3.1. Expression and purification of the recombinant AbnA
The study herein introduces a novel arabinan hydrolytic enzymewith distinct thermostable properties compared to other charac-terized arabinases (ABNs). According to the genome sequenceinformation from T. petrophila, Accession number ABQ46657, ourtarget protein is annotated as a CAZy GH43 family member. TheDNA sequence was 1416 bp long and putatively encodes a 471amino acid protein, bearing a cleavage site for a signal peptide atamino acid position 19 and 20. The mature protein without thesignal peptide, containing a 6 � His-Tag at the N-terminus, wassuccessfully overexpressed in the cytoplasmatic fraction of E. coliBL21 (DE3), after induction by IPTG for 4 h at 37 �C. The proteinpurification steps included Ni2+-chelating affinity and size-exclusion chromatography and resulted in a highly purified enzymesuitable for biochemical and biophysical assays.
3.2. Functional characterization
Cellulases and xylanases have been subjected to extensive anal-ysis over their structure and biochemistry, however, there is littleinformation about arabinan degrading enzymes, which also con-tribute and are important components of the enzymatic repertoryinvolved in plant structural polysaccharides degradation. The puri-fied AbnA showed optimum activity at 73 �C with a relative activityranging from 77% to 78% at temperatures of 62 and 80 �C, respec-tively (Fig. 1A). The optimal pH of the recombinant enzyme was 6,although it retained more than 75% activity over a pH range from 4to 7 (Fig. 1B). Aiming at the investigation of heat stability, AbnAwas pre-incubated in buffer without substrate for the indicatedtime-periods, followed by arabinase activity assay (Fig. 1C). AbnAwas fully heat-stable (up to 10 h) and retained activity at temper-atures up to 90 �C. The enzyme lost its activity only after 90 minwhen incubated at 95 �C (Fig. 1C). The optimum temperaturereported for the AbnA is comparable to other bacterial ABNsfrom B. thermodenitrificans and Caldicellulorsiruptor saccharolyticus,which display temperature optima at 75 and 70 �C, respectively[8,22]. Although, AbnA shows a higher optimum temperature ofoperation compared to other structurally related ABNs from B. sub-tilis and A. niger, 60 and 55 �C, respectively [4,10], this enzyme cer-tainly differentiates from other ABNs regarding to itsthermostability, whereas the residual activity insignificantly de-creases after incubation at 90 �C for 10 h. On the other hand, AbnAresembles other bacterial ABNs with respect to the pH profile,exhibiting activity in the neutral to slightly acidic range [10,8,24].
Kinetic studies using AbnA in the presence of debranched arab-inan as substrate at optimum pH and temperature allowed thedetermination of the Michaelis–Menten parameters. The specificactivity of the purified protein, against debranched arabinan asthe substrate, was 152.1 ± 10.3 U/mg, with Km and Vmax values of19.9 ± 4.7 mg/ml and 478.2 ± 63.6 mg/ml/min, respectively. Theapparent Km indicates that the affinity of AbnA for debranchedarabinan is lower than from those ABNs from Bacillus spp.[10,22], but similar to the ABN from C. saccharolyticus [8] and anuncultured microorganism recently described [24].
We studied the mode of operation of AbnA, which specificallyhydrolyzes arabinan-containing polysaccharides. The enzyme actson branched arabinan (from sugar beet), but more slowly whencompared to linear or debranched arabinan. This result indicatesthat the enzyme displays internal cleavage of (a 1–5)-linked L-arabinofuranose residues along the polysaccharide chain [34].Fig. 2 describes our attempt to define the mode of action for AbnAon APTS-labeled arabinoheptaose through capillary zone electro-phoresis. During the course of hydrolysis (partial), the enzyme
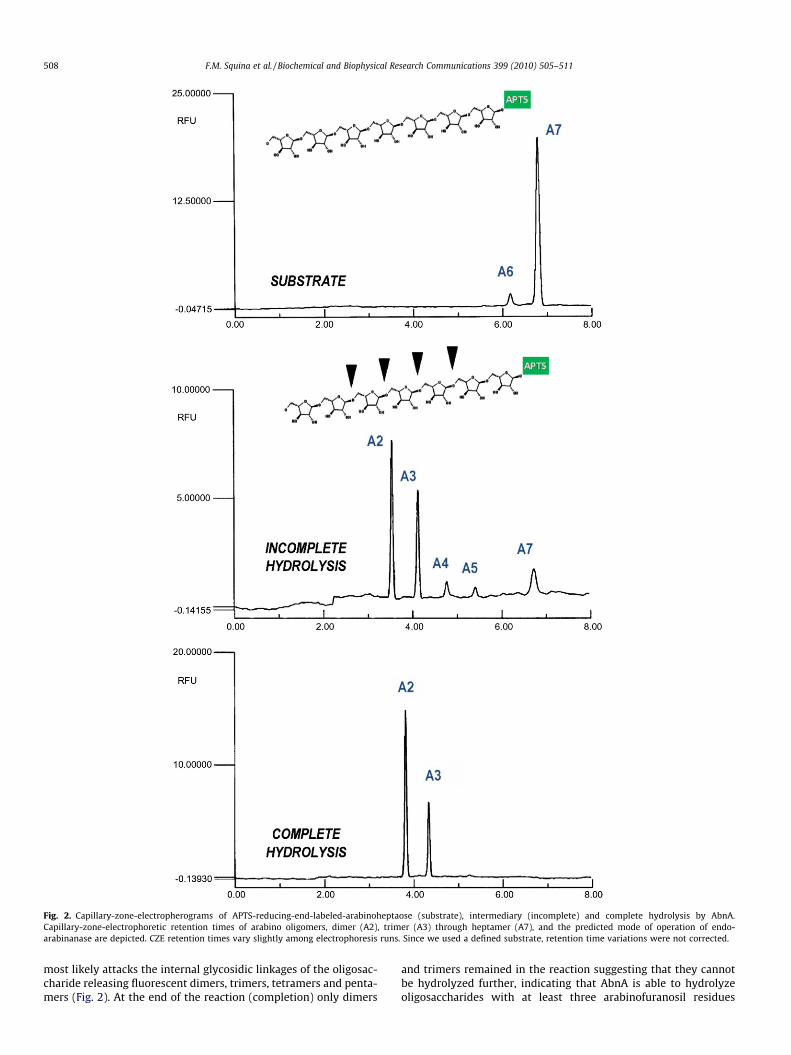
Fig. 2. Capillary-zone-electropherograms of APTS-reducing-end-labeled-arabinoheptaose (substrate), intermediary (incomplete) and complete hydrolysis by AbnA.Capillary-zone-electrophoretic retention times of arabino oligomers, dimer (A2), trimer (A3) through heptamer (A7), and the predicted mode of operation of endo-arabinanase are depicted. CZE retention times vary slightly among electrophoresis runs. Since we used a defined substrate, retention time variations were not corrected.
508 F.M. Squina et al. / Biochemical and Biophysical Research Communications 399 (2010) 505–511
most likely attacks the internal glycosidic linkages of the oligosac-charide releasing fluorescent dimers, trimers, tetramers and penta-mers (Fig. 2). At the end of the reaction (completion) only dimers
and trimers remained in the reaction suggesting that they cannotbe hydrolyzed further, indicating that AbnA is able to hydrolyzeoligosaccharides with at least three arabinofuranosil residues

F.M. Squina et al. / Biochemical and Biophysical Research Communications 399 (2010) 505–511 509
(Fig. 2). The predominant release of arabinobiose and arabinotrioseafter hydrolysis suggests that the enzyme most frequently attacks4th or 5th residues in arabinoheptaose from the non-reducing end.With respect to product formation, the accumulation of fluorescentarabinotriose and arabinobiose as end product resembles otherCAZy GH43 family members [4,11].
3.3. Biophysical characterization
The oligomeric state and polydispersity of purified AbnA wereassessed by DLS, which displayed a single peak with a hydrody-namic radius of 3.0 nm (�molecular weight of 49 kDa) correspond-ing to a monomer. The polydispersity was estimated ofapproximate 14% indicating a monomodal distribution adequatefor further spectroscopic and SAXS studies.
Fig. 3. Far UV-CD (A) and ITFE (B) spectra
Fig. 4. Ab initio shape determination of AbnA. (A) Experimental SAXS curve and the inlet slow-resolution DAMMIN envelope model of AbnA in solution.
The recombinant AbnA is formed mainly by b-sheet secondarystructure elements. Fig. 3 shows the FAR-UV CD spectrum of therecombinant AbnA with a negative minimum at 212 nm, which istypical of proteins mainly containing b-sheet structures, and an in-tense positive band maximum at 230 nm, which is expected, sinceAbnA is rich in tyrosine (7.4%) and tryptophan (2.3%) residues datadeconvolution using the K2D algorithm as implemented in theDichroweb server at http://www.cryst.bbk.ac.uk/cdweb/html [28]resulted in secondary structure contents of 9% a-helix, 47% b-sheetand 44% random coil, which are in reasonable agreement withthose calculated from the in silico model.
ITFE analysis by excitation at 295 nm resulted in an emissionspectrum with a maximum fluorescence peak at 341 nm (Fig. 3),which is typical of the situation for most native proteins. Folded pro-teins usually have emission maxima in the range of 331–343 nm.
of AbnA recorded at pH 6.0 and 20 �C.
hows the pair distances distribution function p(r). (B) Different views of the ab initio

510 F.M. Squina et al. / Biochemical and Biophysical Research Communications 399 (2010) 505–511
3.4. In silico structural model
The AbnA model displayed good overall stereochemistry, with95.5% of the residues in the allowed regions of the Ramachandranplot, as calculated using PROCHECK [32]. The molecular structureis organized into two domains: the N-terminal five-bladed b-pro-peller, which is the catalytic module, and a C-terminal b-barrel.Each blade of the propeller is a b-sheet formed by four antiparallelb-strands, with the first strand being the innermost. The b-sheetsare arranged around a central axis and they pack face-to-face(Fig. 4). The loops connecting strands two and three compose theactive site. The conventional propellers are closed by a molecularVelcro, which is made by incorporating the N- and C-termini inthe same blade; however, this is not observed in AbnA, due tothe fact that N- and C-termini are found in distinct domains. Theb-barrel consists of eight tandem b-strands forming a distortedup-and-down barrel. Despite this domain does not share sequencesimilarity with carbohydrate-binding modules (CBM), its presenceas an accessory domain to a glycoside hydrolase suggests that itcould be derived from an ancient CBM. The CAZy GH43 arabinan-ase from B. subitilis has an active CBM6 at its C-terminus, support-ing our hypothesis.
3.5. Low-resolution molecular structure
Complementing the detailed functional and in silico structuralstudies, we have also determined its low-resolution structure bySAXS. The X-ray scattering and the pair–distance distributioncurves are shown in Fig. 4. Guinier analysis at very low angles indi-cated that the AbnA sample was monodisperse and resulted in agyration radius of 2.68 ± 0.01 nm. Analysis of the p(r) functionshows that AbnA has an elongated shape with a Dmax of7.5 ± 0.4 nm. These values are in agreement with those obtainedfrom the in silico structural model and indicated that AbnA is amonomer in solution which is corroborated with DLS and SEC.SAXS measurements were also carried out at protein concentra-tions of 2, 4 and 10 mg/ml to check for the effect of concentrationon the oligomeric state; however, the protein behaved as monomereven at high protein concentrations, without evidence of oligomer-ization or aggregation. The ab initio low-resolution molecularshape of AbnA was derived from the SAXS data using the programDAMMIN without imposing any symmetry constraints. The SAXSenvelope superposed well on the in silico structural model(Fig. 4) showing that the elongated shape comes from the presenceof the C-terminal b-sandwich domain juxtaposed to the catalyticb-propeller domain.
In conclusion, we have successfully over expressed a novel ABNfrom T. petrophila with remarkable thermostable properties. Weevaluated the AbnA mode of operation, performed comprehensivespectroscopic analyses and determined the low-resolution struc-ture of this novel enzyme. This study provides biochemical andstructural basis for further studies on arabinan hydrolytic en-zymes, which are important components of the enzymatic reper-tory involved in plant structural polysaccharides degradation.
Acknowledgments
This research was supported by grants from FAPESP (08/58037-9) to FMS and CNPq (478059/2009-4) to MTM. RAP has fundingfrom the Department of Energy, awards 06103-OKL and ZDJ-7-77608-01.
References
[1] A. Alhassid, A. Ben-David, O. Tabachnikov, D. Libster, E. Naveh, G. Zolotnitsky,Y. Shoham, G. Shoham, Crystal structure of an inverting GH 43 1,5-alpha-L-
arabinanase from Geobacillus stearothermophilus complexed with its substrate,Biochem. J. 422 (2009) 73–82.
[2] F.T. Chen, R.A. Evangelista, Analysis of mono- and oligosaccharide isomersderivatized with 9-aminopyrene-1,4,6-trisulfonate by capillary electro-phoresis with laser-induced fluorescence, Anal. Biochem. 230 (1995) 273–280.
[3] G. Davies, B. Henrissat, Structures and mechanisms of glycosyl hydrolases,Structure 3 (1995) 853–859.
[4] M.P. Dunkel, R. Amado, Analysis of endo-(1?5)-alpha-L-arabinanase degradationpatterns of linear (1?5)-alpha-L-arabino-oligosaccharides by high-performanceanion-exchange chromatography with pulsed amperometric detection,Carbohydr. Res. 268 (1995) 151–158.
[5] B. Henrissat, A classification of glycosyl hydrolases based on amino acidsequence similarities, Biochem. J. 280 (Pt. 2) (1991) 309–316.
[6] B. Henrissat, G. Davies, Structural and sequence-based classification ofglycoside hydrolases, Curr. Opin. Struct. Biol. 7 (1997) 637–644.
[7] M.E. Himmel, S.Y. Ding, D.K. Johnson, W.S. Adney, M.R. Nimlos, J.W. Brady, T.D.Foust, Biomass recalcitrance: engineering plants and enzymes for biofuelsproduction, Science 315 (2007) 804–807.
[8] M.R. Hong, C.S. Park, D.K. Oh, Characterization of a thermostable endo-1,5-alpha-L-arabinanase from Caldicellulorsiruptor saccharolyticus, Biotechnol. Lett.31 (2009) 1439–1443.
[9] K. Hovel, D. Shallom, K. Niefind, V. Belakhov, G. Shoham, T. Baasov, Y. Shoham,D. Schomburg, Crystal structure and snapshots along the reaction pathway of afamily 51 alpha-L-arabinofuranosidase, EMBO J. 22 (2003) 4922–4932.
[10] T.F. Leal, I. de Sa-Nogueira, Purification characterization and functionalanalysis of an endo-arabinanase (AbnA) from Bacillus subtilis, FEMSMicrobiol. Lett. 241 (2004) 41–48.
[11] V.A. McKie, G.W. Black, S.J. Millward-Sadler, G.P. Hazlewood, J.I. Laurie, H.J.Gilbert, Arabinanase A from Pseudomonas fluorescens subsp. cellulosa exhibitsboth an endo- and an exo-mode of action, Biochem. J. 323 (Pt. 2) (1997) 547–555.
[12] G.L. Miller, Use of dinitrosalicilic acid reagent for determination of reducingsugar, Anal. Chem. 31 (1959) 426–428.
[13] D. Nurizzo, J.P. Turkenburg, S.J. Charnock, S.M. Roberts, E.J. Dodson, V.A. McKie,E.J. Taylor, H.J. Gilbert, G.J. Davies, Cellvibrio japonicus alpha-L-arabinanase 43Ahas a novel five-blade beta-propeller fold, Nat. Struct. Biol. 9 (2002) 665–668.
[14] S. Osaki, T. Kimura, T. Sugimoto, S. Hizukuri, N. Iritani, L-Arabinose feedingprevents increases due to dietary sucrose in lipogenic enzymes andtriacylglycerol levels in rats, J. Nutr. 131 (2001) 796–799.
[15] G. Paes, L.K. Skov, M.J. O’Donohue, C. Remond, J.S. Kastrup, M. Gajhede, O.Mirza, The structure of the complex between a branched pentasaccharide andThermobacillus xylanilyticus GH-51 arabinofuranosidase reveals xylan-bindingdeterminants and induced fit, Biochemistry 47 (2008) 7441–7451.
[16] T. Pons, D.G. Naumoff, C. Martinez-Fleites, L. Hernandez, Three acidic residuesare at the active site of a beta-propeller architecture in glycoside hydrolasefamilies 32, 43, 62, and 68, Proteins 54 (2004) 424–432.
[17] M.R. Proctor, E.J. Taylor, D. Nurizzo, J.P. Turkenburg, R.M. Lloyd, M. Vardakou,G.J. Davies, H.J. Gilbert, Tailored catalysts for plant cell-wall degradation:redesigning the exo/endo preference of Cellvibrio japonicus arabinanase 43A,Proc. Natl. Acad. Sci. USA 102 (2005) 2697–2702.
[18] K. Seri, K. Sanai, N. Matsuo, K. Kawakubo, C. Xue, S. Inoue, L-Arabinoseselectively inhibits intestinal sucrase in an uncompetitive manner andsuppresses glycemic response after sucrose ingestion in animals, Metabolism45 (1996) 1368–1374.
[19] F.M. Squina, A.J. Mort, S.R. Decker, R.A. Prade, Xylan decomposition byAspergillus clavatus endo-xylanase, Protein Expr. Purif. 68 (2009) 65–71.
[20] F.M. Squina, R.A. Prade, H. Wang, M.T. Murakami, Expression, purificationcrystallization and preliminary crystallographic analysis of an endo-1,5-alpha-L-arabinanase from hyperthermophilic Thermotoga petrophila, Acta Crystallogr.Sect. F Struct. Biol. Cryst. Commun. 65 (2009) 902–905.
[21] Y. Takahata, M. Nishijima, T. Hoaki, T. Maruyama, Thermotoga petrophila sp.nov. and Thermotoga naphthophila sp. nov., two hyperthermophilic bacteriafrom the Kubiki oil reservoir in Niigata, Japan, Int. J. Syst. Evol. Microbiol. 51(2001) 1901–1909.
[22] M. Takao, K. Akiyama, T. Sakai, Purification and characterization ofthermostable endo-1,5-alpha-L-arabinase from a strain of Bacillusthermodenitrificans, Appl. Environ. Microbiol. 68 (2002) 1639–1646.
[23] E.J. Taylor, N.L. Smith, J.P. Turkenburg, S. D’Souza, H.J. Gilbert, G.J. Davies,Structural insight into the ligand specificity of a thermostable family 51arabinofuranosidase, Araf51, from Clostridium thermocellum, Biochem. J. 395(2006) 31–37.
[24] D.W. Wong, V.J. Chan, A.A. McCormack, Functional cloning and expression of anovel endo-alpha-1,5-L-arabinanase from a metagenomic library, Protein.Pept. Lett. 16 (2009) 1435–1441.
[25] G. Beldman, H.A. Schols, S.M. Pitson, M.J.F. Seale-van Leeuwen, A.G.J. Voragen,Advances in Macromolecular Carbohydrate Research, JAI Press, Stanford, 1997.
[26] M. Bradford, A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein–dye binding, Anal.Biochem. 72 (1976) 248–252.
[27] R. Naran, M.L. Pierce, A.J. Mort, Detection and identification ofrhamnogalacturonan lyase activity in intercellular spaces of expandingcotton cotyledons, Plant J. 50 (2007) 95–107.
[28] A. Lobley, L. Whitmore, B.A. Wallace, DICHROWEB: an interactive website forthe analysis of protein secondary structure from circular dichroism spectra,Bioinformatics 18 (2002) 211–212.

F.M. Squina et al. / Biochemical and Biophysical Research Communications 399 (2010) 505–511 511
[29] M.B. Kozin, D.I. Svergun, Automated matching of high- and low-resolutionstructural models, J. Appl. Crystallogr. 34 (2001) 33–41.
[30] A. Sali, T.L. Blundell, Comparative protein modelling by satisfaction of spatialrestraints, J. Mol. Biol. 234 (1993) 779–815.
[31] R. Lüthy, J.U. Bowie, D. Eisenberg, Assessment of protein models with three-dimensional profiles, Nature 356 (1992) 83–85.
[32] R.A. Laskowski, M.W. MacArthur, D.S. Moss, J.M. Thornton, PROCHECK – aprogram to check the stereochemical quality of protein structures, J. Appl.Crystallogr. 26 (1993).
[33] P. Emsley, K. Cowtan, Coot: model-building tools for molecular graphics, ActaCrystallogr. Sect. D: Biol. Crystallogr. 60 (2004) 2126–2132.
[34] G. Beldman, M.J.F. Searle-van Leeuwen, G.A. De Ruiter, H.A. Siliha, A.G.J.Voragen, Degradation of arabinans by arabinanases from Aspergillus aculeatusand Aspergillus niger, Carbohydr. Polym. 20 (1993) 159–168.
[35] D.I. Svergun, Determination of the regularization parameter in indirect-transform methods using perceptual criteria, J. Appl. Crystallogr. 25 (1992).
[36] D.I. Svergun, Restoring low resolution structure of biological macromoleculesfrom solution scattering using simulated annealing, Biophys. J. 76 (1999)2879–2886.
[37] M. Wiederstein, M.J. Sippl, ProSA-web: interactive web service for therecognition of errors in three-dimensional structures of proteins, NucleicAcids Res. 35 (2007) 407–410.
Copyright © 2022 FDOKUMEN




















