Subchronic administration of Trichilia catigua ethyl-acetate fraction promotes antidepressant-like...
6
Subchronic administration of Trichilia catigua ethyl-acetate fraction promotes antidepressant-like effects and increases hippocampal cell proliferation in mice Vivian Taciany Bonassoli a , Janaine Micheli Chassot a , Renata Longhini b , Humberto Milani a , Jo ~ ao Carlos P. Mello b , Ru ´ bia Maria Weffort de Oliveira a,n a Laboratory of Neuropsychopharmacology, Department of Pharmacology and Therapeutics, State University of Maringa ´, Av. Colombo n. 5790, 87020-900, Maringa ´, Parana ´, Brazil b Laboratory of Pharmacognosy, Department of Pharmacy, State University of Maringa ´, Av. Colombo n. 5790, 87020-900 Maringa ´, Parana ´, Brazil article info Article history: Received 20 March 2012 Received in revised form 11 May 2012 Accepted 12 June 2012 Available online 19 June 2012 Keywords: Trichillia catigua Antidepressant-like effect Hippocampal neurogenesis Mice abstract Ethnopharmacological relevance: Trichilia catigua preparations have been popularly used in Brazil as a tonic for the treatment of fatigue, stress, impotence, and memory deficits. We recently demonstrated an antidepressant-like effect of acute administration of the Trichilia catigua ethyl-acetate fraction (EAF) in mice. The aim of the present study was to evaluate whether subchronic Trichilia catigua EAF administration maintains its antidepressant-like effects and whether these effects are related to hippocampal neurogenesis. Material and methods: Trichilia catigua EAF (200 and 400 mg/kg) was orally administered to mice for 14 day. The animals were tested in the forced swim test (FST) or tail suspension test (TST). After behavioral testing, the animals received bromodeoxyuridine (BrdU; 200 mg/kg, i.p.) and were euthanized 24 h, 7 day, or 15 day later. The brains were assayed for BrdU and doublecortin (DCX) immunohistochemistry to detect cell proliferation/survival and neurogenesis, respectively. Results: Subchronic administration of 400 mg/kg Trichilia catigua EAF promoted antidepressant-like effects in mice in both the FST and TST. The antidepressant-like effect was accompanied by an increase in cell proliferation in the dentate gyrus (DG) of the hippocampus 24 h after the treatments were discontinued. This proliferative effect, however, did not influence cell survival or neurogenesis because no change in the number of BrdU- or DCX-positive cells was detected 7 or 15 day after the last EAF administration compared with controls. Conclusions: Trichilia catigua EAF produced antidepressant-like effects and induced hippocampal cell proliferation in mice. The results contribute information on the pharmacological and molecular mechanisms involved in the antidepressant-like effect of Trichilia catigua EAF. & 2012 Elsevier Ireland Ltd. All rights reserved. 1. Introduction For many decades, the main hypothesis to explain the cause and treatment of depression has been based on the pharmacolo- gical effects of antidepressants on monoamine neurotransmission (i.e., blockade of serotonin, norepinephrine, and dopamine reup- take or breakdown; Lanni et al., 2009). However, the time lag for a therapeutic response has led to the hypothesis that cellular and molecular adaptations to the actions of antidepressants occur and are necessary for their antidepressant effects. Alterations in cell morphology, the length and complexity of neuronal processes, and the birth of new neurons in the hippocampus (i.e., neurogenesis) have been associated with the action of antidepressants. Various antidepressants, including tricyclic anti- depressants and selective serotonin reuptake inhibitors, increase neuronal proliferation in the hippocampal dentate gyrus (DG) in adult rodents that receive chronic treatment (Malberg et al., 2000; Malberg, 2004). Antidepressant-induced neurogenesis occurs over 2–4 weeks, a time scale that corresponds to the therapeutic delay typically seen in clinical treatment (Malberg et al., 2000). Therefore, demonstrating increased hippocampal neurogenesis induced by a novel compound has been considered one way to validate the compound’s antidepressant effects (Malberg, 2004). Trichilia catigua Juss (Meliaceae) is a medium-sized flowering tree widely distributed in South American countries. In Brazil, it is known as ‘‘catuaba’’, ‘‘catuama,’’ and ‘‘catigua ´ ’’ and has been used in folk medicine as a tonic for the treatment of fatigue, stress, Contents lists available at SciVerse ScienceDirect journal homepage: www.elsevier.com/locate/jep Journal of Ethnopharmacology 0378-8741/$ - see front matter & 2012 Elsevier Ireland Ltd. All rights reserved. http://dx.doi.org/10.1016/j.jep.2012.06.021 n Corresponding author. Tel.: þ55 44 3011 5165; fax: þ55 44 3011 4999. E-mail address: [email protected] (R.M.W. de Oliveira). Journal of Ethnopharmacology 143 (2012) 179–184
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Subchronic administration of Trichilia catigua ethyl-acetate fraction promotes antidepressant-like...

Journal of Ethnopharmacology 143 (2012) 179–184
Contents lists available at SciVerse ScienceDirect
Journal of Ethnopharmacology
0378-87
http://d
n Corr
E-m
journal homepage: www.elsevier.com/locate/jep
Subchronic administration of Trichilia catigua ethyl-acetate fraction promotesantidepressant-like effects and increases hippocampal cell proliferationin mice
Vivian Taciany Bonassoli a, Janaine Micheli Chassot a, Renata Longhini b, Humberto Milani a,Jo~ao Carlos P. Mello b, Rubia Maria Weffort de Oliveira a,n
a Laboratory of Neuropsychopharmacology, Department of Pharmacology and Therapeutics, State University of Maringa, Av. Colombo n. 5790, 87020-900, Maringa, Parana, Brazilb Laboratory of Pharmacognosy, Department of Pharmacy, State University of Maringa, Av. Colombo n. 5790, 87020-900 Maringa, Parana, Brazil
a r t i c l e i n f o
Article history:
Received 20 March 2012
Received in revised form
11 May 2012
Accepted 12 June 2012Available online 19 June 2012
Keywords:
Trichillia catigua
Antidepressant-like effect
Hippocampal neurogenesis
Mice
41/$ - see front matter & 2012 Elsevier Irelan
x.doi.org/10.1016/j.jep.2012.06.021
esponding author. Tel.: þ55 44 3011 5165; fa
ail address: [email protected] (R.M.W. d
a b s t r a c t
Ethnopharmacological relevance: Trichilia catigua preparations have been popularly used in Brazil as a
tonic for the treatment of fatigue, stress, impotence, and memory deficits. We recently demonstrated an
antidepressant-like effect of acute administration of the Trichilia catigua ethyl-acetate fraction (EAF) in
mice. The aim of the present study was to evaluate whether subchronic Trichilia catigua EAF
administration maintains its antidepressant-like effects and whether these effects are related to
hippocampal neurogenesis.
Material and methods: Trichilia catigua EAF (200 and 400 mg/kg) was orally administered to mice for 14
day. The animals were tested in the forced swim test (FST) or tail suspension test (TST). After behavioral
testing, the animals received bromodeoxyuridine (BrdU; 200 mg/kg, i.p.) and were euthanized 24 h,
7 day, or 15 day later. The brains were assayed for BrdU and doublecortin (DCX) immunohistochemistry
to detect cell proliferation/survival and neurogenesis, respectively.
Results: Subchronic administration of 400 mg/kg Trichilia catigua EAF promoted antidepressant-like
effects in mice in both the FST and TST. The antidepressant-like effect was accompanied by an increase
in cell proliferation in the dentate gyrus (DG) of the hippocampus 24 h after the treatments were
discontinued. This proliferative effect, however, did not influence cell survival or neurogenesis because
no change in the number of BrdU- or DCX-positive cells was detected 7 or 15 day after the last EAF
administration compared with controls.
Conclusions: Trichilia catigua EAF produced antidepressant-like effects and induced hippocampal cell
proliferation in mice. The results contribute information on the pharmacological and molecular
mechanisms involved in the antidepressant-like effect of Trichilia catigua EAF.
& 2012 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
For many decades, the main hypothesis to explain the causeand treatment of depression has been based on the pharmacolo-gical effects of antidepressants on monoamine neurotransmission(i.e., blockade of serotonin, norepinephrine, and dopamine reup-take or breakdown; Lanni et al., 2009). However, the time lag for atherapeutic response has led to the hypothesis that cellularand molecular adaptations to the actions of antidepressants occurand are necessary for their antidepressant effects. Alterationsin cell morphology, the length and complexity of neuronalprocesses, and the birth of new neurons in the hippocampus
d Ltd. All rights reserved.
x: þ55 44 3011 4999.
e Oliveira).
(i.e., neurogenesis) have been associated with the action ofantidepressants. Various antidepressants, including tricyclic anti-depressants and selective serotonin reuptake inhibitors, increaseneuronal proliferation in the hippocampal dentate gyrus (DG) inadult rodents that receive chronic treatment (Malberg et al.,2000; Malberg, 2004). Antidepressant-induced neurogenesisoccurs over 2–4 weeks, a time scale that corresponds to thetherapeutic delay typically seen in clinical treatment (Malberget al., 2000). Therefore, demonstrating increased hippocampalneurogenesis induced by a novel compound has been consideredone way to validate the compound’s antidepressant effects(Malberg, 2004).
Trichilia catigua Juss (Meliaceae) is a medium-sized floweringtree widely distributed in South American countries. In Brazil, it isknown as ‘‘catuaba’’, ‘‘catuama,’’ and ‘‘catigua’’ and has been usedin folk medicine as a tonic for the treatment of fatigue, stress,

V. Taciany Bonassoli et al. / Journal of Ethnopharmacology 143 (2012) 179–184180
impotence, and memory deficits (Pizzolatti et al., 2002). InCampos et al. (2005). demonstrated the antidepressant-likeeffects of a Trichilia catigua hydroethanolic extract in rodentssubjected to the forced swim test (FST). The antidepressant-likeeffects of Trichilia catigua were mainly associated with themodulation of the dopaminergic system, based on the fact thattwo dopamine receptor antagonists, haloperidol and chlorproma-zine, completely reversed the effects of Trichilia catigua in the FSTin rats. We recently demonstrated an antidepressant-like effect ofacute administration of the Trichilia catigua ethyl-acetate fraction(EAF) in mice. In addition to its antidepressant-like effects,Trichilia catigua EAF improved cognition in an inhibitory avoid-ance task (i.e., the step-down task; Chassot et al., 2011).
Given that hippocampal neurogenesis has been associatedwith memory improvements (Deng et al., 2010) and antidepres-sant-like effects (Sahay et al., 2007), the aim of the present studywas to test whether subchronic Trichilia catigua EAF administra-tion has antidepressant-like effects and whether these effects arerelated to hippocampal neurogenesis.
2. Material and methods
2.1. Plant material and extract preparation
The bark of Trichilia catigua was acquired in Caetite, Bahia,Brazil, in 2008, and a voucher specimen was identified byDr. Cassia Monica Sakuragui, Universidade Federal do Rio deJaneiro, RJ, Brazil, and deposited at the Herbarium of UniversidadeEstadual de Maringa (no. 19.434), Maringa, PR, Brazil. The extractswere produced as described by Chassot et al. (2011). Briefly, thecrude extract (CE) was produced with acetone–water (7:3; v/v) toyield 101 g from 450 g of bark. The semipurified EAF was obtainedafter partition with ethyl acetate to yield 13 g from 50 g of CE.
2.2. Animals
Male Swiss mice, weighing 30–40 g, were housed in groups(n¼5 per group) with food and water freely available andmaintained at 2272 1C under a 12 h/12 h light/dark cycle. All ofthe procedures were conducted in accordance with the BrazilianCollege of Animal Experimentation (COBEA), which comply withinternational laws, and were approved by the Ethics Committeeon Animal Experimentation of the State University of Maringa(CEEA 042/2007).
2.3. Treatments
Saline, 20 mg/kg imipramine (Sigma-Aldrich, St. Louis, MO,USA), and 200 and 400 mg/kg EAF were administered orally bygavage in a volume of 10 ml/kg using a tuberculin syringe fittedwith an oral cannula (0.1 cm�4 cm). The treatments wereadministered once per day for 14 consecutive days. The doses ofEAF were based on a previous study that found antidepressant-like effects of a single EAF administration in mice (Chassot et al.,2011).
2.4. Behavioral testing
One hour after the last administration, the animals weresubjected to the FST. A separate group of animals was evaluatedin the tail suspension test (TST). Behavioral testing was performedin a dark room with minimal background noise. Hand-operatedcounters and stopwatches were used by a blind observer to scorethe behavioral parameters.
Forced swim test. The FST was performed according to themethods described by Porsolt et al. (1977). Briefly, the mice weregently and individually placed in a glass beaker (25 cmheight�10 cm diameter) that contained 2372 1C water thatwas deep enough so that the mice could not touch the bottomof the container or escape. The amount of time the mouse spentactive until its first immobility episode (i.e., latency) and immo-bility time, during which the mouse did not struggle and madeonly movements necessary to keep its head above water, weremeasured during a 6 min swimming session. Immobility time wasmeasured during the last 4 min of the session.
Tail suspension test. The TST was conducted as previouslydescribed by Steru et al. (1985) with slight modifications. Amouse was individually suspended by its tail with adhesive tape(1 cm from the tip of the tail) in a black wooden box(45 cm�45 cm�30 cm) for 6 min. The mouse was consideredimmobile only when it hung passively and completely motion-less. Immobility time was measured during the last 4 min of thesession.
2.5. Histological procedures
Immediately after behavioral testing in the FST, all of the micereceived the thymidine analog 5-bromo-2-deoxyuridine (BrdU;Sigma-Aldrich, St. Louis, MO, USA) intraperitoneally at 200 mg/kg,divided into four injections 2 h apart. To observe the effects of thetreatments on neuronal proliferation, a group of animals (n¼4–6)was sacrificed 24 h after BrdU administration. To verify thesurvival of proliferating cells (BrdU), subgroups of animals(n¼4–7) were sacrificed 7 or 15 day after the last BrdU admin-istration. Neurogenesis was confirmed by immunohistochemistryusing the antibody anti-microtubule-associated protein double-cortin (DCX), which is expressed transiently in newly generatedneurons, 24 h and, 7 and 15 day after the treatments.
Twenty-four hours, 7 day, or 15 day after the last BrdUadministration, the mice were deeply anesthetized with an over-dose of sodium thiopental (Thiopentax, Cristalia, SP, Brazil) andtranscardially perfused with saline followed by 4% paraformalde-hyde in 0.01 M phosphate-buffered saline (PBS). The brains wereremoved, post-fixed for 2 h in the same fixative, and cryopro-tected by immersion in 30% sucrose for 48 h. Thereafter, frozentissue was serially sectioned on a cryostat (Criocut 1800, Reich-ert-Jung, Heidelberg, Germany) into 40 mm coronal sections andcollected in quadruplicate in Eppendorf tubes that contained anti-freeze solution. The sections were then processed for BrdU or DCXimmunohistochemistry to evaluate cell proliferation and neuro-genesis, respectively.
Immunohistochemistry. For BrdU immunohistochemistry, thesections were first digested in 0.005% trypsin for 5 min at 37 1C,incubated in 1 N HCl for 75 min at 40 1C for DNA denaturation,and neutralized with 0.1 M borate buffer (pH 8.5). Afterward, allof the sections were quenched in 1% H2O2 for 30 min and thenblocked with 2% bovine serum albumin in PBS for 60 min. Thesections were incubated with rat monoclonal anti-BrdU antibody(1:100; Santa Cruz Biotechnology, Santa Cruz, CA, USA) or goatpolyclonal anti-DCX antibody (1:400; AbCam, St. Louis, MO, USA)in PBS that contained 0.3% Triton X-100 for 48 h at roomtemperature. The sections were then incubated with respectivebiotinylated secondary antibodies (1:500) for 2 h at room tem-perature. The sections were further incubated in ABC solution(Vectastain Elite ABC Kit, Vector Laboratories, Burlingame, CA,USA) for 2 h. The peroxidase reaction was performed by incubat-ing the sections with 3-30-diaminobenzidine (DAB; Sigma) and0.05% H2O2. The sections were mounted onto gelatin-coatedslides, air dried, cleared with xylene, and coverslipped withPermount.
V. Taciany Bonassoli et al. / Journal of Ethnopharmacology 143 (2012) 179–184 181
Cell counting. The slides that contained hippocampal sections wererandomized and coded prior to quantitative analysis. BrdU- and DCX-positive cells were scored in the DG under 40� or 100� magnifica-tion, respectively, in both brain hemispheres using an optical micro-scope (Olympus Optical, BX50, Center Valley, PA, USA).
For BrdU counting, six to eight coronal sections, spaced160 mm apart and corresponding to the whole hippocampalextension, �1.06 to �3.64 mm from bregma (Franklin andPaxinos, 1997), were analyzed. Doublecortin-positive cells in theDG were counted in three brain sections per animal, correspond-ing to the coronal coordinates �1.58 to �2.30 mm from bregma(Schiavon et al., 2010). The data are expressed as the meannumber of positive cells in both cerebral hemispheres.
2.6. Statistical analysis
The data are expressed as the mean7SEM of the experimentalgroups. Differences among means were analyzed by one-way analysisof variance (ANOVA), followed by the Newman Keuls post-hoc test.Values of po0.05 were considered statistically significant.
3. Results
3.1. Subchronic administration of 400 mg/kg Trichilia catigua EAF
exerts antidepressant-like effects
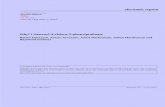
Subchronic administration of 20 mg/kg imipramine and400 mg/kg Trichilia catigua EAF decreased immobility time inthe FST compared with the saline group (F3,42¼12.1, po0.0001).No significant effect was observed in mice that received 200 mg/kgTrichilia catigua EAF (p40.05). No difference was detected amongthe experimental groups in the latency to the first immobilityepisode (F3,42¼1.99, p¼0.13; Fig. 1A).
The antidepressant-like effects of 20 mg/kg imipramine and400 mg/kg Trichilia catigua EAF were confirmed in the TST(F2,29¼20.10, po0.0001), reflected by a significant decrease inimmobility time in the EAF group compared with the saline group(Fig. 1B). No significant effect on latency was observed in any ofthe experimental groups (F2,26¼0.54, p¼0.59).
3.2. Subchronic administration of 400 mg/kg Trichilia catigua EAF
increased hippocampal cell proliferation
An enhanced number of BrdU-positive cells was observed24 h after BrdU administration in both the imipramine- and
Fig. 1. Imipramine and Trichilia catigua EAF reduced immobility time in mice subjecte
(20 mg/kg), Trichilia catigua EAF (200 and 400 mg/kg), or saline were orally administer
in the FST or TST. The latency and immobility time were recorded during the last 4 m
(ANOVA followed by Newman Keuls post-hoc test).
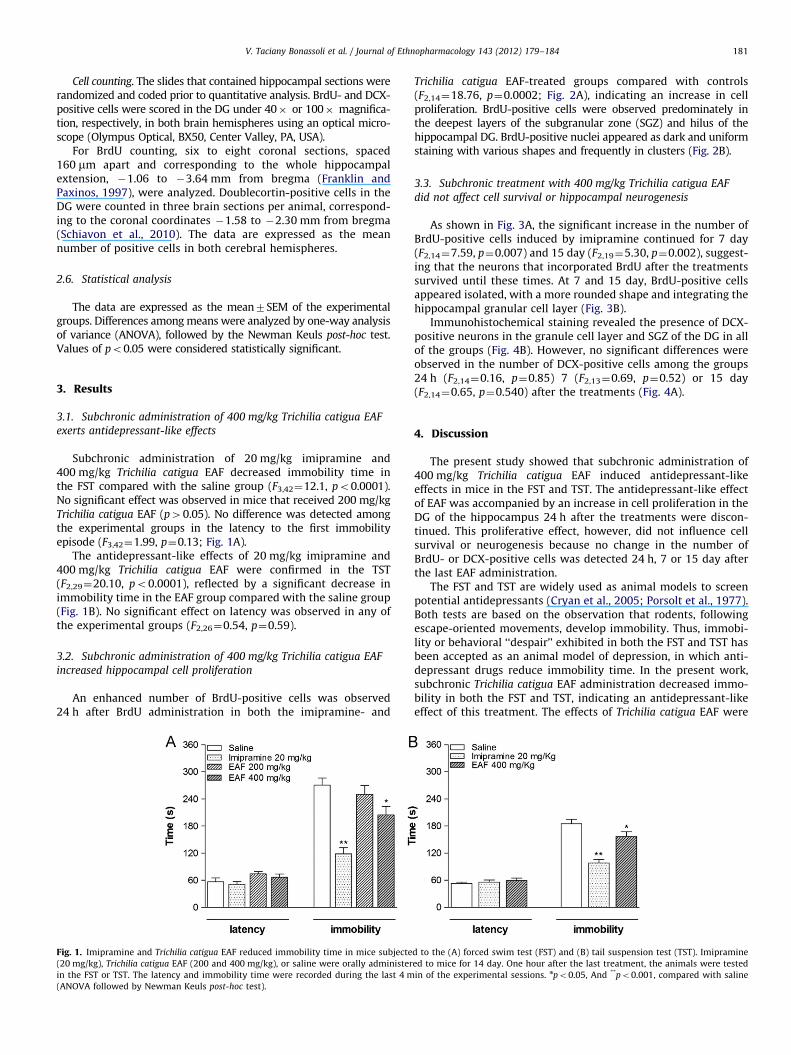
Trichilia catigua EAF-treated groups compared with controls(F2,14¼18.76, p¼0.0002; Fig. 2A), indicating an increase in cellproliferation. BrdU-positive cells were observed predominately inthe deepest layers of the subgranular zone (SGZ) and hilus of thehippocampal DG. BrdU-positive nuclei appeared as dark and uniformstaining with various shapes and frequently in clusters (Fig. 2B).
3.3. Subchronic treatment with 400 mg/kg Trichilia catigua EAF
did not affect cell survival or hippocampal neurogenesis
As shown in Fig. 3A, the significant increase in the number ofBrdU-positive cells induced by imipramine continued for 7 day(F2,14¼7.59, p¼0.007) and 15 day (F2,19¼5.30, p¼0.002), suggest-ing that the neurons that incorporated BrdU after the treatmentssurvived until these times. At 7 and 15 day, BrdU-positive cellsappeared isolated, with a more rounded shape and integrating thehippocampal granular cell layer (Fig. 3B).
Immunohistochemical staining revealed the presence of DCX-positive neurons in the granule cell layer and SGZ of the DG in allof the groups (Fig. 4B). However, no significant differences wereobserved in the number of DCX-positive cells among the groups24 h (F2,14¼0.16, p¼0.85) 7 (F2,13¼0.69, p¼0.52) or 15 day(F2,14¼0.65, p¼0.540) after the treatments (Fig. 4A).
4. Discussion
The present study showed that subchronic administration of400 mg/kg Trichilia catigua EAF induced antidepressant-likeeffects in mice in the FST and TST. The antidepressant-like effectof EAF was accompanied by an increase in cell proliferation in theDG of the hippocampus 24 h after the treatments were discon-tinued. This proliferative effect, however, did not influence cellsurvival or neurogenesis because no change in the number ofBrdU- or DCX-positive cells was detected 24 h, 7 or 15 day afterthe last EAF administration.
The FST and TST are widely used as animal models to screenpotential antidepressants (Cryan et al., 2005; Porsolt et al., 1977).Both tests are based on the observation that rodents, followingescape-oriented movements, develop immobility. Thus, immobi-lity or behavioral ‘‘despair’’ exhibited in both the FST and TST hasbeen accepted as an animal model of depression, in which anti-depressant drugs reduce immobility time. In the present work,subchronic Trichilia catigua EAF administration decreased immo-bility in both the FST and TST, indicating an antidepressant-likeeffect of this treatment. The effects of Trichilia catigua EAF were
d to the (A) forced swim test (FST) and (B) tail suspension test (TST). Imipramine
ed to mice for 14 day. One hour after the last treatment, the animals were tested
in of the experimental sessions. npo0.05, And **po0.001, compared with saline
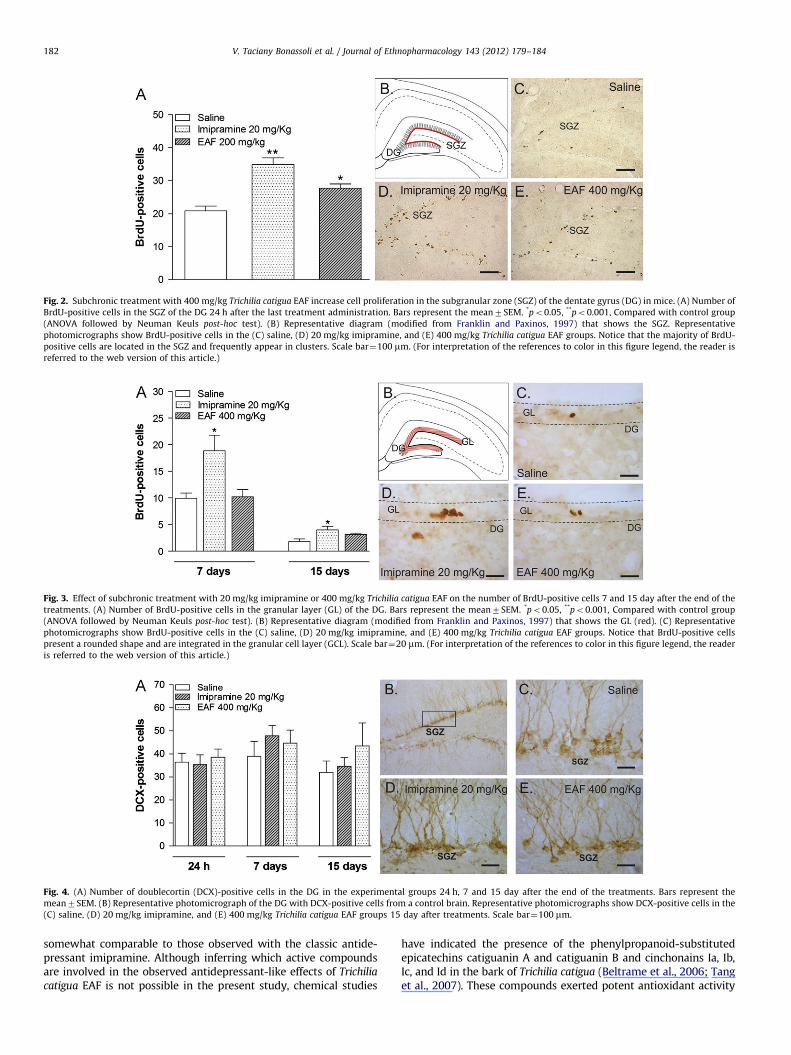
Fig. 3. Effect of subchronic treatment with 20 mg/kg imipramine or 400 mg/kg Trichilia catigua EAF on the number of BrdU-positive cells 7 and 15 day after the end of the
treatments. (A) Number of BrdU-positive cells in the granular layer (GL) of the DG. Bars represent the mean7SEM. *po0.05, **po0.001, Compared with control group
(ANOVA followed by Neuman Keuls post-hoc test). (B) Representative diagram (modified from Franklin and Paxinos, 1997) that shows the GL (red). (C) Representative
photomicrographs show BrdU-positive cells in the (C) saline, (D) 20 mg/kg imipramine, and (E) 400 mg/kg Trichilia catigua EAF groups. Notice that BrdU-positive cells
present a rounded shape and are integrated in the granular cell layer (GCL). Scale bar¼20 mm. (For interpretation of the references to color in this figure legend, the reader
is referred to the web version of this article.)
Fig. 4. (A) Number of doublecortin (DCX)-positive cells in the DG in the experimental groups 24 h, 7 and 15 day after the end of the treatments. Bars represent the
mean7SEM. (B) Representative photomicrograph of the DG with DCX-positive cells from a control brain. Representative photomicrographs show DCX-positive cells in the
(C) saline, (D) 20 mg/kg imipramine, and (E) 400 mg/kg Trichilia catigua EAF groups 15 day after treatments. Scale bar¼100 mm.
Fig. 2. Subchronic treatment with 400 mg/kg Trichilia catigua EAF increase cell proliferation in the subgranular zone (SGZ) of the dentate gyrus (DG) in mice. (A) Number of
BrdU-positive cells in the SGZ of the DG 24 h after the last treatment administration. Bars represent the mean7SEM. *po0.05, **po0.001, Compared with control group
(ANOVA followed by Neuman Keuls post-hoc test). (B) Representative diagram (modified from Franklin and Paxinos, 1997) that shows the SGZ. Representative
photomicrographs show BrdU-positive cells in the (C) saline, (D) 20 mg/kg imipramine, and (E) 400 mg/kg Trichilia catigua EAF groups. Notice that the majority of BrdU-
positive cells are located in the SGZ and frequently appear in clusters. Scale bar¼100 mm. (For interpretation of the references to color in this figure legend, the reader is
referred to the web version of this article.)
V. Taciany Bonassoli et al. / Journal of Ethnopharmacology 143 (2012) 179–184182
somewhat comparable to those observed with the classic antide-pressant imipramine. Although inferring which active compoundsare involved in the observed antidepressant-like effects of Trichilia
catigua EAF is not possible in the present study, chemical studies
have indicated the presence of the phenylpropanoid-substitutedepicatechins catiguanin A and catiguanin B and cinchonains Ia, Ib,Ic, and Id in the bark of Trichilia catigua (Beltrame et al., 2006; Tanget al., 2007). These compounds exerted potent antioxidant activity

V. Taciany Bonassoli et al. / Journal of Ethnopharmacology 143 (2012) 179–184 183
that was related to the neuroprotective effects and neuritogenicproperties of catechins and cinchonains (Weinreb et al., 2009). It ispossible that the antidepressant-like effect of Trichilia catigua EAFmay be related to antioxidant and neuroplastic activity.
The behavioral effects of Trichilia catigua EAF were furthercharacterized by immunohistochemistry, showing that Trichilia
catigua EAF also enhanced cell proliferation in the hippocampus,an index of antidepressant-like activity. We compared the pro-liferation and survival of newborn cells in the DG in mice treatedwith imipramine and Trichilia catigua EAF using immunohisto-chemistry to detect BrdU, a thymidine analog that can be retainedwithin cells during division (Huang and Herbert, 2006). BrdU wasadministered after Trichilia catigua EAF, and the number of BrdU-positive cells was measured 24 h, 7 day, and 15 day later.Imipramine and Trichilia catigua EAF increased the number ofBrdU-positive cells in the DG 24 h after the last administration.This finding indicates an increase in cell proliferation induced bythese treatments. However, tracking BrdU-positive cells 7 and15 day post-treatment, we observed no significant differences inthe number of BrdU-positive cells between the Trichilia catigua
EAF- and saline-treated groups, indicating that subchronic Trichi-
lia catigua EAF treatment did not influence cell survival. Increasedgranule cell proliferation induced by antidepressant treatmentsindicates that more competing neurons are available for selection,which may improve the ability of the hippocampus to adapt toemerging environmental challenges. This phenomenon mightplay a role in the beneficial clinical effects of antidepressanttreatments (Von Bohlen and Halbach, 2011).
Concerning the experiment that focused on cell survival, adecrease in the number of BrdU-positive cells was observed overtime in all of the experimental groups, including the controlgroup. This finding is consistent with loss caused by cell death orBrdU dilution through repeated rounds of mitosis (Bingham et al.,2005). Regardless of the reason for the decrease in the number ofBrdU-labeled cells, the cells that remained beyond 7 and 15 daypost-treatment were localized to the granular cell layer of the DG,indicating that these cells survived after BrdU administration. Thedetection and specific localization of BrdU-positive cells in thegranular cell layer of the DG is a strong indicator of the neuronalphenotype of newborn cells. As expected, in addition to increas-ing cell proliferation, imipramine also enhanced the survival ofpostmitotic cells, confirming its antidepressant-like properties.
To confirm the effects of Trichilia catigua EAF treatment onhippocampal neurogenesis, we also performed immunohisto-chemistry to detect DCX-positive neurons. Doublecortin is aprotein that promotes microtubule polymerization (Franciset al., 1999). Its expression is thought to be specific to newlygenerated neurons because virtually all DCX-positive cells expressearly neuronal antigen but lack antigens specific to glia, undiffer-entiated cells, and apoptotic cells (Rao and Shetty, 2004). Dou-blecortin expression begins shortly after the neurons are born andpersists for approximately 2 weeks in rats and 3 weeks in mice.Doublecortin expression then gradually declines as the neuronscontinue to mature (Rao and Shetty, 2004; Snyder et al., 2009). Inthe present study, no significant difference in the number ofDCX-positive cells was observed in mice treated with imipramineor Trichilia catigua EAF. Unexpectedly, imipramine treatment for14 day did not change the number of DCX-positive neuronscompared with controls, although the treatment regimen usedhere was similar to previous studies showing positive effects ofimipramine on DCX expression (Murray et al., 2008; Schiavonet al., 2010). The reason for the lack of effect of imipramine onthe survival of newborn neurons is unknown. However, previousresearch showed that imipramine may induce neurogenesis onlyafter 3 weeks of administration (Pechnick et al., 2008; Sairanenet al., 2005). Therefore, the imipramine and Trichilia catigua
EAF treatment duration may not have been sufficient to promotea significant increase in neurogenesis in the DG of the hippocam-pus. Further studies with longer administration periods and otherdoses of imipramine and Trichilia catigua EAF need to be con-ducted to resolve this issue.
In conclusion, the present results demonstrated that 400 mg/kg Trichilia catigua EAF produced antidepressant-like effects in theFST and TST and increased cell proliferation in the hippocampalDG. The results provide information on the pharmacological andmolecular mechanisms involved in the antidepressant-like effectof Trichilia catigua EAF and implicate its potential as a coadjuvanttreatment for mood disorders.
Acknowledgments
The authors thank Marco Alberto Trombelli for his technicalsupport. This study was supported by Conselho Nacional deDesenvolvimento Cientıfico e Tecnologico (CNPq), Coordenac- ~aode Aperfeic-oamento de Pessoal de Nıvel Superior (CAPES), andFundac- ~ao Araucaria.
References
Beltrame, F.L., Rodrigues Filho, E., Barros, F.A.P., Cortez, D.A.G., Cass, Q.B.A., 2006.Validated higher-performance liquid chromatography method for quantifica-tion of cinchonain ib in bark and phytopharmaceuticals of Trichilia catiguaused as catuaba. Journal of Chromatography 25, 257–263.
Bingham, B., Liu, D., Wood, A., Cho, S., 2005. Ischemia-stimulated neurogenesis isregulated by proliferation, migration, differentiation and caspase activation ofhippocampal precursor cells. Brain Research 1058, 167–177.
Campos, M.M., Fernandes, E.S., Ferreira, J., Santos, A.R., Calixto, J.B., 2005.Antidepressant-like effects of Trichilia catigua (Catuaba) extract: evidence fordopaminergic-mediated mechanisms. Psychopharmacology (Berlin) 182,45–53.
Chassot, J.M., Longhini, R., Gazarini, L., Mello, J.C., de Oliveira, R.M., 2011.Preclinical evaluation of Trichilia catigua extracts on the central nervoussystem of mice. Journal of Ethnopharmacolology 137, 1143–1148.
Cryan, J.F., Mombereau, C., Vassout, A., 2005. The tail suspension test as a modelfor assessing antidepressant activity: review of pharmacological and geneticstudies in mice. Neuroscience and Biobehavioral Reviews 29, 571–625.
Deng, W., Aimone, J.B., Gage, F.H., 2010. New neurons and new memories: howdoes adult hippocampal neurogenesis affect learning and memory? NatureReviews Neuroscience 11, 339–350.
Francis, F., Koulakoff, A., Boucher, D., Chafey, P., Schaar, B., Vinet, M.C., Friocourt,G., McDonnell, N., Reiner, O., Kahn, A., McConnell, S.K., Berwald-Netter, Y.,Denoulet, P., Chelly, J., 1999. Doublecortin is a developmentally regulated,microtubule-associated protein expressed in migrating and differentiatingneurons. Neuron 23, 247–256.
Franklin, B.J., Paxinos, G., 1997. The mouse brain in stereotaxic coordinates.Academic Press, London.
Huang, G.J., Herbert, J., 2006. Stimulation of neurogenesis in the hippocampus ofthe adult rat by fluoxetine requires rhythmic change in corticosterone.Biological Psychiatry 59, 619–624.
Lanni, C., Govoni, S., Lucchelli, A., Boselli, C., 2009. Depression and antidepressants:molecular and cellular aspects. Cellular and Molecular Life Sciences. 66,2985–3008.
Malberg, J.E., 2004. Implications of adult hippocampal neurogenesis in antide-pressant action. Journal of Psychiatry and Neuroscience 29, 196–205.
Malberg, J.E., Eisch, A.J., Nestler, E.J., Duman, R.S., 2000. Chronic antidepressanttreatment increases neurogenesis in adult rat hippocampus. Journal ofNeuroscience 20, 9104–9110.
Murray, F., Smith, D.W., Hutson, P.H., 2008. Chronic low dose corticosteroneexposure decreased hippocampal cell proliferation, volume and inducedanxiety and depression like behaviours in mice. European Journal of Pharma-cology 31, 115–127.
Pechnick, R.N., Bresee, C.J., Manalo, C.M., Poland, R.E., 2008. Comparison of theeffects of desmethylimipramine on behavior in the forced swim test inperipubertal and adult rats. Behavioral Pharmacology 19, 81–84.
Pizzolatti, M.G., Venson, A.F., Smania, A.J., Smania, E.F.A., Braz-Filho, R., 2002. Twoepimeric flavalignans from Trichilia catigua (Meliaceae) with antimicrobialactivity. Zeitschrift der Naturforschung 57, 483–488.
Porsolt, R.D., Bertin, A., Jalfre, M., 1977. Behavioral despair in mice: a primaryscreening test for antidepressants. Archieves of Internal Pharmacodynamicsand Therapeutics 229, 327–336.
Rao, M.S., Shetty, A.K., 2004. Efficacy of doublecortin as a marker to analyse theabsolute number and dendritic growth of newly generated neurons in theadult dentate gyrus. European Journal of Neuroscience 19, 234–246.

V. Taciany Bonassoli et al. / Journal of Ethnopharmacology 143 (2012) 179–184184
Sahay, A., Drewm, M.R., Hen, R., 2007. Dentate gyrus neurogenesis and depression.Progress in Brain Research 163, 697–722.
Sairanen, M., Lucas, G., Ernfors, P., Castren, M., Castren, E., 2005. Brain-derivedneurotrophic factor and antidepressant drugs have different but coordinatedeffects on neuronal turnover, proliferation, and survival in the adult dentategyrus. Journal of Neuroscience 25, 1089–1094.
Schiavon, A.P., Milani, H., Romanini, C.V., Foresti, M.L., Castro, O.W., Garcia-Cairasco, N., de Oliveira, R.M., 2010. Imipramine enhances cell proliferationand decreases neurodegeneration in the hippocampus after transient globalcerebral ischemia in rats. Neuroscience Letters. 470, 43–48.
Snyder, J.S., Glover, L.R., Sanzone, K.M., Kamhi, J.F., Cameron, H.A., 2009. The effectsof exercise and stress on the survival and maturation of adult-generatedgranule cells. Hippocampus 19, 898–906.
Steru, L., Chermat, R., Thierry, B., Simon, P., 1985. The tail suspension test: a newmethod for screening antidepressants in mice. Psychopharmacology (Berlin)85, 367–370.
Tang, W., Hioki, H., Harada, K., Kubo, M., Fukuyama, Y., 2007. Antioxidantphenylpropanoid-substituted epicatechins from Trichilia catigua. Journal ofNatural Products 70, 2010–2013.
Von Bohlen, O., Halbach, O., 2011. Immunohistological markers for proliferativeevents, gliogenesis, and neurogenesis within the adult hippocampus. Cellularand Tissue Research 345, 1–19.
Weinreb, O., Amit, T., Mandel, S., Youdim, M.B., 2009. Neuroprotective molecularmechanisms of (–)-epigallocatechin-3-gallate: a reflective outcome of itsantioxidant, iron chelating and neuritogenic properties. Genes and Nutrition4, 283–296.













![Synthesis, Single Crystal X-Ray Structure, and Antimicrobial Activity of 6-(1,3-Benzodioxol-5-ylmethyl)-5-ethyl-2-{[2-(morpholin-4-yl)ethyl]sulfanyl}pyrimidin-4(3H)-one](https://static.fdokumen.com/doc/165x107/6324d4c2c9c7f5721c01c4b5/synthesis-single-crystal-x-ray-structure-and-antimicrobial-activity-of-6-13-benzodioxol-5-ylmethyl-5-ethyl-2-2-morpholin-4-ylethylsulfanylpyrimidin-43h-one.jpg)
![1-[2-(2,6-Dichlorobenzyloxy)-2-(2-furyl)ethyl]-1 H -benzimidazole](https://static.fdokumen.com/doc/165x107/63152ec4fc260b71020fe0ce/1-2-26-dichlorobenzyloxy-2-2-furylethyl-1-h-benzimidazole.jpg)