
Chapter 22. Genetic diseases reveal DNA nucleotide excision ...

Study of an Isolated Population at the Nucleotide Level: up49 Region of a Canarian Population of Drosophila subobscura 1
Julio Rozas and Montserrat Aguade’ Departament de Genetica, Facultat de Biologia, Universitat de Barcelona
Restriction-map variation in a 1.6-kb region that includes the rp49 gene has been analyzed in 54 lines from a natural population of Drosophila subobscura from Tenerife (Canary Islands). The sample was monomorphic for the 03* chromosomal gene arrangement. By use of seven 4-cutter restriction enzymes, 12 polymorphisms (eight restriction-site polymorphisms and four insertion/deletions of 1 - 15 bp) and 23 different haplotypes have been identified. Estimated haplotype and nucleotide diversity-O.870 and 0.0028, respectively-are lower than those in continental populations. The results suggest that, although within the chromosomal class OS+~ the continental populations are homogeneous, the Tenerife population is genetically different. In agreement with previous results, this indicates that it is an old popu- lation, long isolated from the mainland.
Introduction
Populations of Drosophila subobscura from the Canary Islands have been studied at the chromosomal (Prevosti 197 1, 1974; Prevosti et al. 1975 ), enzymatic (Cabrera et al. 1980, 1983; Larruga et al. 1983), and mitochondrial DNA levels (Latorre et al. 1986; Afonso et al. 1990). Prevosti ( 1974) observed that the level of chromosomal polymorphism in the Canary Islands is lower than that on the continent and that the polymorphism in the islands is also qualitatively different from that on the adjacent mainland (South Morocco), suggesting that Canarian populations have been long isolated. Studies at the enzymatic (Larruga et al. 1983 ) and mitochondrial DNA levels (Afonso et al. 1990) have also shown that those populations differ from both European and African populations. In the present paper we analyze both ( 1) nucleotide variation in a region that includes the rp49 gene (ribosomal protein 49) and ( 2) chromosomal polymorphism for a population of D. subobscura from the island of Tenerife (Canary Islands). The rp49 gene is a nuclear single-copy gene located in segment I of the 0 chromosome (band 9 1C) of D. subobscura ( AguadC 1988). Variation in this region has been previously assayed in two European populations of this species (Barcelona and Ter Apel, The Netherlands) (Rozas and AguadC 1990).
Material and Methods Fly Samples
Fiftylfour isofemale lines of Drosophila subobscura were established from wild- caught flies collected at Las Raices (Tenerife) in June 1987. The balancer stock for the 0 chromosome Va/Ba (Sperlich et al. 1977) was used to obtain heterozygotes of
1. Key words: rp49 region, nucleotide polymorphism, Drosophila subobscura.
Address for correspondence and reprints: Julio Rozas, Departament de Genetica, Facuhat de Biologia, Universitat de Barcelona, Diagonal 645,0807 I Barcelona, Spain
Mol. Biol. Evol. 8(2):202-2 I I. 199 I. 0 1991 by The University of Chicago. All rights reserved. 0737-4030/91/0802-0003$02.00
202
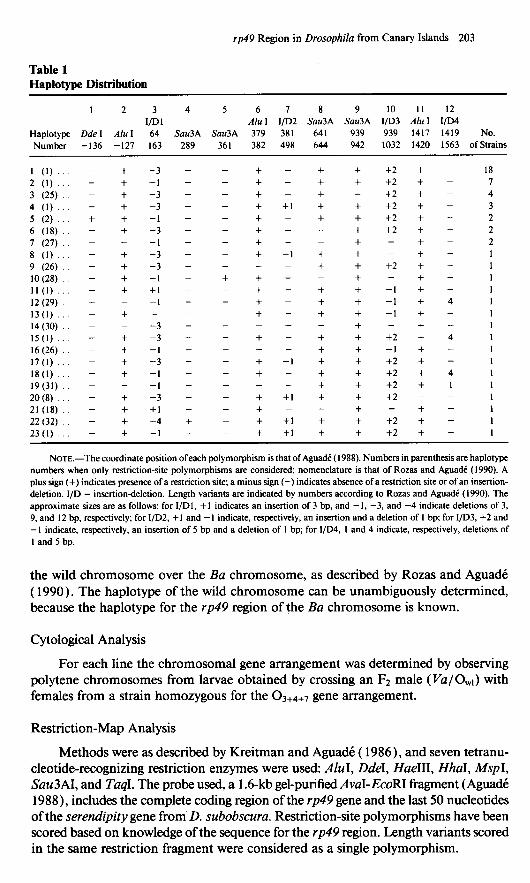
Table 1
rp49 Region in Drosophila from Canary Islands 203
Hadotvue Distribution
1 2 3 4 5 6 7 8 9 10 II 12 l/D1 AIuI I/D2 Sau3A SadA I/D3 Ah I I/D4
Haplotype Dde I Ah I 64 Sau3A Sau3A 319 381 641 939 939 1417 1419 NO. Number -136 -127 163 289 361 382 498 644 942 1032 1420 1563 ofStrains
1 (1) _ 2 (1) - 3 (25) _ 4 (1) - 5 (2) + 6 (18) - 7 (27) _ 8 (1) _ 9 (26) - lO(28) - 11 (1) _ 12 (29) - 13(l) - 14(30) _ 15(l) - 16(26) _ 17(l) _ 18(l) - 19(31) - 20 (8) _ 21 (18) - 22 (32) - 23(l) -
+ + + + t t - + t t t _ t _ + t t t - t t t t
-3 -1 -3 -3 -1 -3 -1 -3 -3 -1 t1 -1 -
-3 -3 -1 -3 -1 -1 -3 t1 -4 -1
- _ t - _ t _ - t - - t _ - t - _ t _ _ t - _ t - - - _ t t _ _ t - - + _ _ t - - _ - - + - - _ + _ - t _ - - + _ - t + - t - _ t
- - _
t1 _ - -
-1 _ - - _ - - _ -
-1 _ -
t1 _
+I +I
+ t + t t - t + t t - + _ t t + t t _ t t t + t + t - + t t t + t t t t + t t t - t + t t t
t2 +2 t2 t2 i-2 +2 - -
+2 - -1 -1 -1 _
t2 -1 +2 t2 t2 t2 -
+2 +2
t _ 18 t _ 7 + _ 4 t - 3 + - 2 t - 2 t _ 2 t _ 1 t - 1 + _ 1 t - 1 t 4 1 t _ 1 t - 1 t 4 1 t - 1 t - 1 + 4 1 t 1 1 _ _ 1 t - 1 t - 1 t - 1
NOTE.-The coordinate position of each polymorphism is that of Aguadk ( 1988). Numbers in parenthesis are haplotype numbers when only restriction-site polymorphisms are considered; nomenclature is that of Rozas and Aguadk (1990). A plus sign (+) indicates presence of a restriction site; a minus sign (-) indicates absence of a restriction site or of an insertion- deletion. I/D = insertion-deletion. Length variants are indicated by numbers according to Rozas and AguadC (1990). The approximate sizes are as follows: for I/Dl, tl indicates an insertion of 3 bp, and - 1, -3, and -4 indicate deletions of 3, 9, and I2 bp, respectively; for 1/D2, t 1 and - 1 indicate, respectively, an insertion and a deletion of 1 bp; for I/D3, t2 and - 1 indicate, respectively, an insertion of 5 bp and a deletion of I bp; for I/D4, I and 4 indicate, respectively, deletions of I and 5 bp.
the wild chromosome over the Ba chromosome, as described by Rozas and AguadC ( 1990). The haplotype of the wild chromosome can be unambiguously determined, because the haplotype for the rp49 region of the Ba chromosome is known.
Cytological Analysis
For each line the chromosomal gene arrangement was determined by observing polytene chromosomes from larvae obtained by crossing an F2 male (Va/O,,) with females from a strain homozygous for the 03+4+7 gene arrangement.
Restriction-Map Analysis
Methods were as described by Kreitman and Aguadt! ( 1986)) and seven tetranu- cleotide-recognizing restriction enzymes were used: &I, D&I, HaeIII, HhaI, MspI, Sau 3A1, and TaqI. The probe used, a 1.6-kb gel-purified AvaI-EcoRI fragment ( Aguadk 1988), includes the complete coding region of the rp49 gene and the last 50 nucleotides of the serendipity gene from D. subobscura. Restriction-site polymorphisms have been scored based on knowledge of the sequence for the rp49 region. Length variants scored in the same restriction fragment were considered as a single polymorphism.
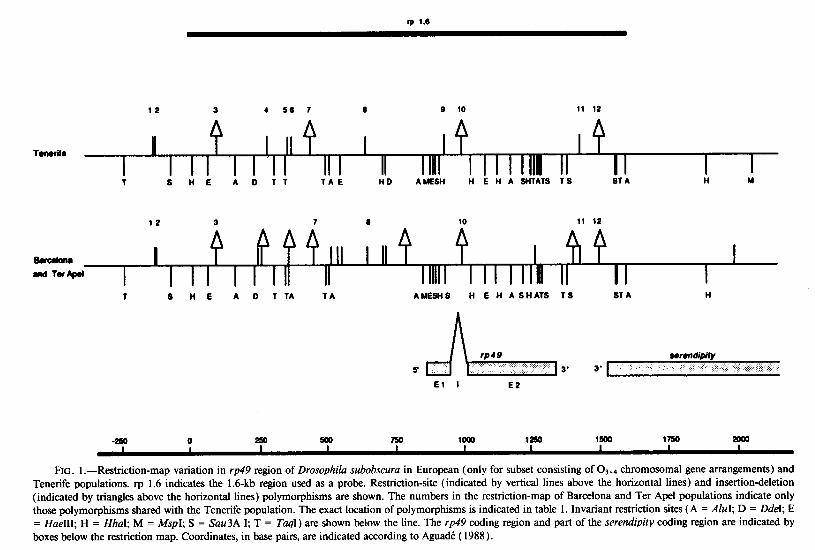
rp 1.0
12 3 4 50 7 2 0 10 11 12
TOIWIO H
I I I
! IIf I
I I II I Ill1 I I I III II I I 1 8 HE AD11 TA E HD AMESH H E H A SWATS 1 S STA H u
12 3 7 2 10 11 12
I III Ill4 I I md Tar&d I I I I I II II II 1111111 II II I 1 S HE A D TTA TA AMESHS H E H ASHATS 18 ST A H
El I E2
-224 0 250 500 750 1002 1252 1500 1750 2002 I I I I I I I I I
FIG. 1 .-Restriction-map variation in rp49 region of Drosophila subobscuru in European (only for subset consisting of 0 3+4 chromosomal gene arrangements) and Tenerife populations. rp 1.6 indicates the 1.6-kb region used as a probe. Restriction-site (indicated by vertical lines above the horizontal lines) and insertion-deletion (indicated by triangles above the horizontal lines) polymorphisms are shown. The numbers in the restriction-map of Barcelona and Ter Ape1 populations indicate only those polymorphisms shared with the Tenerife population. The exact location of polymorphisms is indicated in table 1. Invariant restriction sites (A = AhI; D = D&I; E = HueIII; H = HhaI; M = MspI; S = Suu3A I; T = Tuql) are shown below the line. The rp49 coding region and part of the serendipity coding region are indicated by boxes below the restriction map. Coordinates, in base pairs, are indicated according to AguadC ( 1988).

206 Rozas and Aguadb
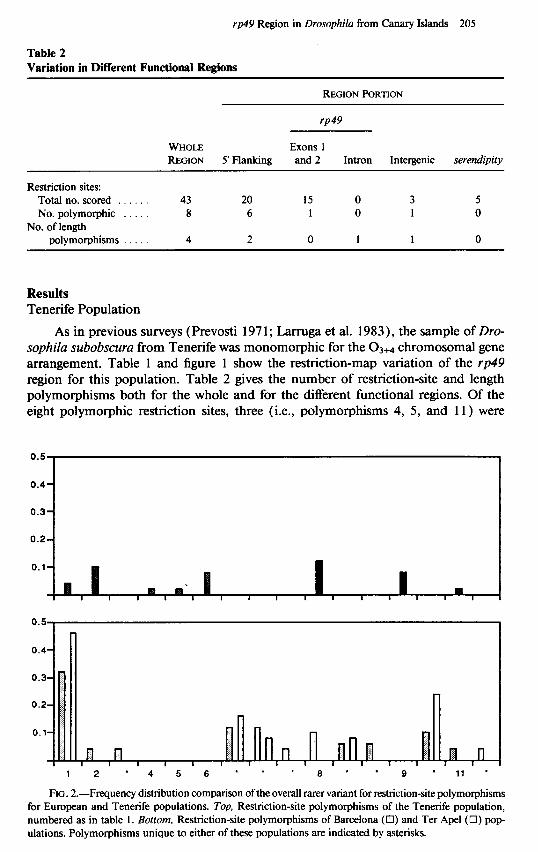
Table 3 Linkage Disequilibrium Expressed as Correlation Coefficient r
POLYMORPHIC SITES POLYMORPHIC
SITEa 1 2 6 8
2 .......... 0.063 6 .......... 0.056 0.397* 8 .......... 0.076 0.447* 0.101 9 .......... 0.056 -0.090 -0.080 -0.109
NOTE.-Statistical significance was determined by a tw&ailed Fisher’s exact test of indepen- dence.
* Numbered as in table I. * P c 0.05.
present only once in the sample, the rest ranging in frequency between 0.037 and 0.130 (fig. 2).
All four insertion/deletion polymorphisms detected (i.e., 3, 7, 10, and 12) show more than two length variants, varying up to 15 t- 1 bp in length. Length polymor- phisms 3, 10, and 12 are located in regions with either different repeated short motifs or polyA / polyT runs, suggesting that they might be generated by mispairing (Levinson and Gutman 1987 ) .
Twenty-three different haplotypes have been identified ( 16 of them being unique) when all variation is considered (table 1) . Only 12 have been identified when only restriction-site polymorphisms are considered.
Nucleotide diversity was estimated by three different methods (Engels 198 1; Hudson 1982; Nei and Tajima 1983). Engels’ and Nei and Tajima’s methods gave similar results (0.00272 and 0.00278, respectively), but these estimates were lower than Hudson’s (0.00642 ). Hudson’s estimator differs from the others because it takes into account the number of polymorphisms but not their frequency. Although its higher value might be explained by the frequency spectrum of restriction-site poly- morphisms (frequencies range 0.02-o. 13)) no deviation from neutrality could be de- tected when applying the test proposed by Tajima ( 1989): D = -1.365, P > 0.1.
Linkage disequilibrium between restriction-site polymorphisms has been analyzed when the rarer variant is not unique in the population. Only two of the 10 pairwise comparisons show significant linkage disequilibrium at the 5% level (table 3). See Hill and Robertson (1968) for discussion of r in linkage disequilibrium.
Comparison between Tenerife and European Populations
Rozas and AguadC ( 1990) have analyzed the rp49 region in two European pop- ulations of D. subobscuru. There was heterogeneity in the distribution of haplotypes and of restriction-site polymorphisms among but not within chromosomal classes. The data from the Tenerife population have therefore only been compared with the subset of O3+4 chromosomal gene arrangements from both European populations.
Figure 2 shows the frequency of the rarer variant for each restriction-site poly- morphism for Tenerife and European populations. Of eight restriction-site polymor- phisms, four (i.e., 4,5,6, and 9) detected in Tenerife are not observed in the European populations, and two of these four (i.e., 6 and 9) are not unique.
All four length polymorphisms present in Tenerife are also present in the European populations. Nevertheless, for two of these four shared polymorphisms there are the following differences: (a) for length polymorphism 3, the most common variant in
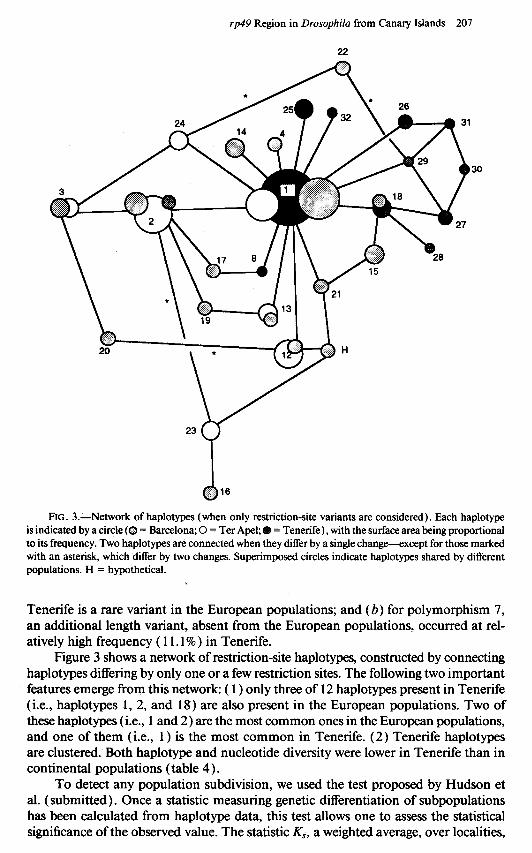
rp49 Region in Drosophila from Canary Islands 207
FIG. 3.-Network of haplotypes (when only restriction-site variants are considered). Each haplotype is indicated by a circle (0 = Barcelona; 0 = Ter Apel; 8 = Tenerife) , with the surface area being proportional to its frequency. Two haplotypes are connected when they differ by a single change-except for those marked with an asterisk, which differ by two changes. Superimposed circles indicate haplotypes shared by different populations. H = hypothetical.
Tenerife is a rare variant in the European populations; and (b) for polymorphism 7, an additional length variant, absent from the European populations, occurred at rel- atively high frequency ( 11.1% ) in Tenerife.
Figure 3 shows a network of restriction-site haplotypes, constructed by connecting haplotypes differing by only one or a few restriction sites. The following two important features emerge from this network: ( 1) only three of 12 haplotypes present in Tenerife (i.e., haplotypes 1, 2, and 18) are also present in the European populations. Two of these haplotypes (i.e., 1 and 2) are the most common ones in the European populations, and one of them (i.e., 1) is the most common in Tenerife. (2) Tenerife haplotypes are clustered. Both haplotype and nucleotide diversity were lower in Tenerife than in continental populations (table 4).
To detect any population subdivision, we used the test proposed by Hudson et al. (submitted). Once a statistic measuring genetic differentiation of subpopulations has been calculated from haplotype data, this test allows one to assess the statistical significance of the observed value. The statistic K,, a weighted average, over localities,

208 Rozas and AguadC
Table 4 Haplotype Diversity and Nucleotide Diversity
Tenerife Barcelona and Ter Apel’
Haplotype diversity:b Restriction site Length
Total Nucleotide diversity’
0.5759 + 0.0792 0.8647 + 0.0387 0.7589 + 0.0548 0.9757 f 0.0106 0.8700 + 0.0380 0.9937 + 0.0067 0.00278 + 0.00192 0.00562 + 0.00290
’ Both populations pooled. For these populations only the subset of Os+, chromosomes has been considered.
b Estimated according to the method of Nei and Tajima (198 I). ’ Estimated according to the method of Nei and Tajima (1983). Sampling variance was calculated
according to the method of Lynch and Crease (1990).
of the average number of differences between sequences from within a locality (IQ, has been used as a measure of genetic differentiation of localities. To test for departure of the hypothesis of structural homogeneity of two localities, we obtained permutations of the total number of observed haplotypes (n, + nz) and then randomly assigned them to subsamples of size 12, and n2. For each permutation the statistic KS is estimated. The associated probability (p) is the proportion of those estimated values that are less than the observed value. Table 5 gives the KS values and associated prob- abilities (for 1,000 permutations) for pairwise comparisons, both when all polymor- phisms are considered and when only restriction-site polymorphisms are considered. The results suggest (a) that within the chromosomal class 03+4, Barcelona and Ter Ape1 are homogeneous but (b) that the Tenerife population is structurally different. T
Discussion
The present study, using high-resolution restriction mapping (Kreitman and AguadC 1986 ), suggests that for the rp49 region there is genetic differentiation between Canarian and European populations. This corroborates previous chromosomal (Pre- vosti 197 1, 1974; Prevosti et al. 1975), enzymatic (Cabrera et al. 1980; Larruga et al. 1983), and mitochondrial DNA results ( Afonso et al. 1990).
Analysis of,both common and unique polymorphisms and haplotypes of the rp49 region indicate that Canarian populations have been long isolated from the mainland: first, several restriction-site and insertion/deletion polymorphisms found in relatively high frequency in Tenerife are not present at appreciable frequencies in
Table 5 K, Values for Pairwise Comparisons between Localities
Ks (P)
All Polymorphisms
Restriction-Site Polymorphisms
Only
Barcelona vs. Ter Ape1 (n, = 31, nz = 13) Barcelonavs.Tenerife(n,=31,nt=54) _....,...,..,. Ter Ape.1 vs. Tenerife (n, = 13, n2 = 54) Barcelona + Ter Ape1 vs. Tenerife (n, = 44, n2 = 54)
4.0182 (0.185) 1.7500 (0.680) 2.7870 (0.000) 1.2237 (0.000) 2.5387 (0.000) 0.9902 (0.000) 2.9903 (0.000) 1.3178 (0.000)

rp49 Region in Drosophila from Canary Islands 209
the European populations; second (and conversely), when only restriction-site poly- morphisms are considered, nine of 12 haplotypes detected in Tenerife are endemic; and, third, in the network these endemic haplotypes cluster.
In Drosophila subobxuru the frequencies of some chromosomal arrangements vary clinally with latitude (Krimbas and Loukas 1980). However, the frequencies of allozyme polymorphisms within a chromosomal arrangement are in general homo- geneous throughout Europe (Larruga and Pinsker 1984; Pinsker and Biihm 1989; for an exception, see Prevosti et al. 1983), as are the frequencies of haplotypes and of restriction-site polymorphisms (Rozas and Aguade 1990). For the rp49 region there is no evidence for population subdivision within chromosomal classes in continental populations; the population from Tenerife seems, however, to be genetically distinct. Such homogeneity within Europe may arise from the high dispersal capacity of D. subobscuru (Prevosti et al. 1987, 1988; Serra et al. 1987). However, barriers to gene flow result in independent evolution on oceanic islands.
In the present study, estimated nucleotide diversity in the rp49 region is lower in Tenerife than in the European populations (table 4). This could be due to a smaller effective population size on the islands. According to the infinite-site model of the neutral theory (Kimura 1983)) where the expected heterozygosity is approximately equal to 4N,p, a higher level of heterozygosity would indicate a larger effective size, if the same mutation rate is assumed. Anomalously, mitochondrial DNA heterozygosity was actually higher in the Canarian as compared with European populations ( Afonso et al. 1990; Rozas et al. 1990). Since there are no good methods available yet to test for differences in nucleotide diversity estimates, and because the variance of these estimates that is due to nucleotide and individual sampling is large (Lynch and Crease 1990)) more data from both European and Canarian populations’are necessary before this discrepancy can be properly evaluated.
Acknowledgments
We thank the Departamento de Genetica of Universidad de La Laguna (Tenerife, Spain) for allowing us to use their facilities during our collection of Drosophila sub- obscura. We thank A. Berry, M. Kreitman, N. Miyashita, and W. Stephan for com- ments; M. Lynch for programs; and R. R. Hudson and N. Kaplan for sharing their manuscript and programs. This work was supported by CAICYT grant PB85-0157 to M.A. J.R. was supported by a predoctoral fellowship (F.P.I.) from Ministerio de Education y Ciencia, Spain.
LITERATURE CITED
AFONSO, J. M., A. VOLZ, M. HERNANDEZ, H. RUTTKAY, M. GONZALEZ, J. M. LARRUGA, V. M. CABRERA, and D. SPERLICH. 1990. Mitochondrial DNA variation and genetic structure in Old-World populations of Drosophila subobscura. Mol. Biol. Evol. 7: 123- 142.
AGUADB, M. 1988. Nucleotide sequence comparison of the rp49 gene region between Drosophila subobscura and D. melanogaster. Mol. Biol. Evol. 5:433-44 1.
CABRERA, V. M., A. M. GONZALEZ, and A. GULL~N. 1980. Enzymatic polymorphism in Dro- sophila subobscura populations from the Canary islands. Evolution 34:875-887.
CABRERA, V. M., A. M. GONZALEZ, J. M. LARRUGA, and C. VEGA. 1983. Linkage disequilibrium in chromosome A of Drosophila subobscura. Genetica 61:3-8.
ENGELS, W. R. 198 1. Estimating genetic divergence and genetic variability with restriction endonucleases. Proc. Natl. Acad. Sci. USA 78:6329-6333.
HILL, W. G., and A. ROBERTSON. 1968. Linkage disequilibrium in finite populations. Theor. Appl. Genet. 38:226-23 1.

2 10 Rozas and Aguadb
HUDSON, R. R. 1982. Estimating genetic variability with restriction endonucleases. Genetics loo:71 1-719.
HUDSON, R. R., D. D. BOOST, and N. L. KAPLAN. ,A statistical test for detecting population subdivision. (submitted).
KIMURA, M. 1983. The neutral theory of molecular evolution. Cambridge University Press, Cambridge.
KREITMAN, M., and M. AGUAD~. 1986. Genetic uniformity in two populations of Drosophila melanogaster as revealed by filter hybridization of four-nucleotide-recognizing restriction enzyme digests. Proc. Natl. Acad. Sci. USA 83:3562-3566.
RRIMBAS, C. B., and M. LOUKAS. 1980. The inversion polymorphism of Drosophila subobscura. Evol. Biol. 12: 163-234.
LARRUGA, J. M., V. M. CABRERA, A. M. GONZALEZ, and A. GULL~N. 1983. Molecular and chromosomal polymorphism in continental and insular populations from the southwestern range of Drosophila subobscura. Genetica 60: 19 l-205.
LARRUGA, J. M., and W. PINSKER. 1984. The importance of geographic isolation, structural rearrangement and speciation for the genetic divergence of chromosome 0 in Drosophila subobscura, Drosophila guanche, and Drosophila madeirensis. Z. Zool. Syst. Evol.-forsch. 22:103-l 13.
LATORRE, A., A. MOYA, and F. AYALA. 1986. Evolution of mitochondrial DNA in Drosophila subobscura. Proc. Natl. Acad. Sci. USA 83:8649-8653.
LEVINSON, G., and G. A. GUTMAN. 1987. Slipped-strand mispairing: a major mechanism for DNA sequence evolution. Mol. Biol. Evol. 4:203-22 1.
LYNCH, M., and T. J. CREASE. 1990. The analysis of population survey data on DNA sequence variation. Mol. Biol. Evol. 7~377-394.
NEI, M., and F. TAJIMA. 1981. DNA polymorphism detectable by restriction endonucleases. Genetics 97~145-163.
-. 1983. Maximum likelihood estimation of the number of nucleotide substitutions from restriction sites data. Genetics 105:207-2 17.
PINSKER, W., and I. B~HM. 1989. Allozyme divergence between different gene arrangements in Drosophila subobscura. J. Evol. Biol. 2:353-366.
PREVOSTI, A. 197 1. Chromosomal polymorphism in Drosophila subobscura Coll. populations from the Canary islands. Genet. Iber. 23:69-84.
-. 1974. Chromosomal inversion polymorphism in the southwestern range of Drosophila subobscura distribution area. Genetica 45: 1 1 1 - 124.
PREVOSTI, A., M. P. GARCIA, L. SERRA, M. AGUADB, G. RIBS, and E. SAGARRA. 1983. As- sociation between allelic isozyme alleles and chromosomal arrangements in European pop ulations and Chilean colonizers of Drosophila subobscura. Pp. 17 1 - 19 1 in M. C. RATAZZI, J. G. SCANDALIOS, and G. S. WHITT, eds. Isozymes: current topics in biological and medical research, vol. 10: Genetics and evolution. Alan R. Liss, New York.
PREVOSTI, A., J. OCARA, and G. ALONSO. 1975. Distances between populations of Drosophila subobscura, based on chromosome arrangement frequencies. Theor. Appl. Genet. 45:231- 241.
PREVOSTI, A., G. RIBS, L. SERRA, M. AGUAD~, J. BALA~QA, M. MONCL& and F. MESTRES. 1988. Colonization of America by Drosophila subobscura: experiment in natural populations that supports the adaptive role of chromosomal-inversion polymorphism. Proc. Natl. Acad. Sci. USA 85:5597-5600.
PREVOSTI, A., L. SERRA, M. MONCL~JS, F. MESTRES, A. LATORRE, G. RIB& and M. AGUAD~. 1987. Colonization de America por Drosophila subobscura. Evol. Biol. (Bogata) l:l-24.
ROZAS, J., and M. AGUADB. 1990. Evidence of extensive genetic exchange in the rp49 region among polymorphic chromosome inversions in Drosophila subobscura. Genetics 126~4 17- 426.
ROZAS, J., M. HERNANDEZ, V. M. CABRERA, and A. PREVOSTI. 1990. Colonization of America

rp49 Region in Drosophila from Canary Islands 2 11
by Drosophila subobscura: effect of the founder event on the mitochondrial DNA polymor- phism. Mol. Biol. Evol. 7: 103-109.
SERRA, L., G. PEGUEROLES, and F. MESTRES. 1987. Capacity of dispersal of a colonizing species: Drosophila subobscura. Genetica 73:223-235.
SPERLICH, D., H. FEUERBACH-MRAVLAG, P. LANGE, A. MICHAELIDIS, and A. PENTZOS-DA- PONTE. 1977. Genetic load and viability distribution in central and marginal populations of Drosophila subobscura. Genetics 86~835-848.
TAJIMA, F. 1989. Statistical method for testing the neutral mutation hypothesis by DNA poly- morphism. Genetics 123:585-595.
BRIAN CHARLESWORTH, reviewing editor
Received April 6, 1990; revision received October 3 1, 1990
Accepted October 3 1, 1990
Copyright © 2022 FDOKUMEN




















