
Studies on cellular differentiation of Magnaporthe grisea. Physicochemical aspects of substratum...
10
Plysiologicnl a’nd Molecular Plmr~ Pafholou (1994) 44, 227-236 227 Studies on cellular differentiation of Magnaporthe grisea. Physicochemical aspects of substratum surfaces in relation to appressorium formation J.-Z. XIAO, T. WATANABE, T. KAMAKURA, A. OHSHIMA* and I. YAMAGUCHI Microbial Toxicology Labornloy, The hslilule of Plysical and Chemical Research (RIKEN), IV&o, Sailomn 351-01, Japaa (hepled for publication Febrtql 1994) Pathogenesis by Magtmporthe g&en, the causal fungus of rice blast disease, involves formation of a special infection structure termed an appressorium. Formation of the appressorium occurs apart from the host, but the regulation mechanism is not completely understood. Studies were performed on various liquids and solids to elucidate the physicochcmical aspects involved in appressorium formation. Conidia started to germinate when they contacted either a solid or a liquid surface; nutrient was not required. Elongated germ-tubes differentiated to form appressoria on solid surfaces but not on liquid or agar gel surfaces. Both hydrophobic and hydrophilic solid surfaces were highly inductive for appressorium formation. Topographic characteristics did not appear to be important because appressoria were formed on both rough and apparently smooth surfaces. When germinated on freshly prepared agar gel surfaces, elongated germ-tubes penetrated directly into the soft surfaces without forming apprcssorium. On partially dried agar gel surfaces, however, they differentiated to form appressoria and then penetrated the surface with penetration hyphae elaborated from the appressoria. From these results, we inferred that a hard solid surface triggered apprcssorium differentiation. INTRODUCTION Many plant pathogens form a specialized infection structure, the appressorium which facilitates adhesion and penetration of the host [4]. The formation of an appressorium involves cellular differentiation. Considerable research has been devoted to the biochemistry and ultrastructure of appressorium formation [for review see 4, 71. Formation of appressoria by rust fungi, especially by Uromyces spp. has been extensively studied to date. Staples and colleagues reported changes in protein and RNA synthesis during appressorium formation by Uromyces appendiculatus, and they described the substratum surface parameters necessary to trigger the formation of infection structures [7,8, 12, 141. Recently, two genes specifically expressed during infection structure formation were cloned and characterized in this fungus [I, 171. Magnaporthe grisea Bar (imperfect stage Pyricularia oryzae Car.) is a pathogen of rice causing a devastating disease called rice blast. This fungus produces darkly melanized appressoria which are a prerequisite for penetration into host tissues. An essential role of * Present address: Yuki Research Center, Nihon Bayer Agrochcm K.K., Yuki 951 l-4, Yuki, Ibaraki 307, Japan. 0885-5765/94/030227 + IO $08.00/O 0 1994 Academic Press Limited
-
Upload
harwajdatvjhdvckh -
Category
Documents
-
view
4 -
download
0
Transcript of Studies on cellular differentiation of Magnaporthe grisea. Physicochemical aspects of substratum...

Plysiologicnl a’nd Molecular Plmr~ Pafholou (1994) 44, 227-236 227
Studies on cellular differentiation of Magnaporthe grisea. Physicochemical aspects of substratum surfaces in relation to appressorium formation
J.-Z. XIAO, T. WATANABE, T. KAMAKURA, A. OHSHIMA* and I. YAMAGUCHI
Microbial Toxicology Labornloy, The hslilule of Plysical and Chemical Research (RIKEN), IV&o, Sailomn 351-01, Japaa
(hepled for publication Febrtql 1994)
Pathogenesis by Magtmporthe g&en, the causal fungus of rice blast disease, involves formation of a special infection structure termed an appressorium. Formation of the appressorium occurs apart from the host, but the regulation mechanism is not completely understood. Studies were performed on various liquids and solids to elucidate the physicochcmical aspects involved in appressorium formation. Conidia started to germinate when they contacted either a solid or a liquid surface; nutrient was not required. Elongated germ-tubes differentiated to form appressoria on solid surfaces but not on liquid or agar gel surfaces. Both hydrophobic and hydrophilic solid surfaces were highly inductive for appressorium formation. Topographic characteristics did not appear to be important because appressoria were formed on both rough and apparently smooth surfaces. When germinated on freshly prepared agar gel surfaces, elongated germ-tubes penetrated directly into the soft surfaces without forming apprcssorium. On partially dried agar gel surfaces, however, they differentiated to form appressoria and then penetrated the surface with penetration hyphae elaborated from the appressoria. From these results, we inferred that a hard solid surface triggered apprcssorium differentiation.
INTRODUCTION
Many plant pathogens form a specialized infection structure, the appressorium which facilitates adhesion and penetration of the host [4]. The formation of an appressorium involves cellular differentiation. Considerable research has been devoted to the biochemistry and ultrastructure of appressorium formation [for review see 4, 71. Formation of appressoria by rust fungi, especially by Uromyces spp. has been extensively studied to date. Staples and colleagues reported changes in protein and RNA synthesis during appressorium formation by Uromyces appendiculatus, and they described the substratum surface parameters necessary to trigger the formation of infection structures [7,8, 12, 141. Recently, two genes specifically expressed during infection structure formation were cloned and characterized in this fungus [I, 171.
Magnaporthe grisea Bar (imperfect stage Pyricularia oryzae Car.) is a pathogen of rice causing a devastating disease called rice blast. This fungus produces darkly melanized appressoria which are a prerequisite for penetration into host tissues. An essential role of
* Present address: Yuki Research Center, Nihon Bayer Agrochcm K.K., Yuki 951 l-4, Yuki, Ibaraki 307, Japan.
0885-5765/94/030227 + IO $08.00/O 0 1994 Academic Press Limited

. 228 J.-Z. Xiao et al.
melanization has been indicated using.melanin biosynthesis inhibitors and mutants [3, 18,191. Melanization was shown to empower the pathogen to penetrate the surface of the host by keeping a high internal hydrostatic pressure in the sealed appressorium [9, IO].
M. grisea appressoria can be induced apart from the host plant. Germinating conidia formed appressoria on hydrophobic surfaces, such as Teflon film and Mylar film [2,&S, 101 as well as on hydrophilic surfaces such as those of glass and cellophane membranes [1.5, 161. Uchiyama and Okuyama [15J also reported that rice leaf wax promoted appressorium formation in vitro. However, the mechanism and physiology of appressorium formation by A4. grisea are not fully understood.
In this paper, we report some physicochemical aspects of substratum surfaces involved in appressorium formation, and will discuss the probable mechanism regulating appressorium differentiation.
MATERIALS AND METHODS
Fungus and culture Magnaporthe grisea Bar isolate P2, pathogenic on rice, was used throughout this study. The fungus was grown on oat-meal agar medium at 27 “C for 2 weeks in the dark, followed by incubation under a black light to induce sporulation after removing aerial mycelia with a sterilized brush. Abundant conidia were produced within 3-4 days, which were then brushed off, suspended in sterile water and collected by centrifugation.
Preparation of germination substrata The following materials were tested as germination substrates: cellophane membrane (Sanwa Pure Chemical Co. Ltd, Tokyo 101, Japan), Parafilm “M” (American National Can,,,, CT 06836, U.S.A.) sheets of nylon 66 (Sanplatec Corp., Tokyo 101, Japan), polyalkylbenzenestyrene (Sanplatec), polycarbonate (Sanplatec), polyethylene (Sanplatec), polypropylene (Sanplatec) and polytetrafluoroethylene (Teflon, San- platec), and slide glass (Toshinriko Co. Ltd, Tokyo, Japan). Before use, the cellophane films were washed by boiling them in distilled water three times for 10 min. Newly purchased slide glasses were well washed by scrubbing with a sponge mat or with a brush. The others were washed carefully with distilled water avoiding abrasion to the surfaces.
Liquid paraffin, composed of alkanes, were purchased from Junsei Chemical Co. Ltd, Tokyo, Japan. Nissan polybutenes ON, 5N, 30N and 200N polymers of butyl and n-butyl units were the products of Nippon Oil and Fat Corp. Ltd, Tokyo, Japan. Their average molecular weights and viscosity are shown in Table 1.
Agar gel layers (5 %, w/v, 2 mm in thickness) deposited on slide glasses were used fresh or after drying in a sterilizing oven at 45 “C for times given in Results.
Characterization of germination substrates
The hydrophobicity of the various substrates was determined by assessing the contact angle formed by a 153 pl droplet of distilled water placed on each substrate, using a Contact-Angle Metre (Model CA-DT, Kyowa Interface Science Co. Ltd, Tokyo, Japan). The greater the contact angle the greater the hydrophobicity of the surface.
Surface hardness was determined by Vicker’s indentation technique [13] using an

Appressorium formation by Magnaporthe grisea 229
TABLE 1
Appressoriutn fomlation by Magnaporthe grisea on liquid substrala
Average Conidium Appressorium
mol. wt* Viscosity* germination (o/b)t rormation (%)
Nissan polybutcne 200N 2650 4400 93.2 0 Nissan polybutcne 30N 1350 700 91.6 0 Nissan polybutene 5N 780 120 92.3 0 Nissan polybutene ON 370 48 94.5 0 Liquid paraffin - - 896 0
*The values for average mol.wt. and viscosity (cSt, at 98.9 “C) ofNissan polybutenes shown were provided by Nippon Oil and Fat Corp. Ltd, Japan.
tDroplets (60 pi) or conidium suspensions (2 x 10” conidia m-r) in distilled water were incubated on each substrate for 8-10 h. Conidium germination was assessed using the microscope.
Akashi AK-A Hardness Tester (Akashi Co. Ltd, Japan) at loads of l-50 g. Hardness (Hv), expressed in kg mm-‘, was calculated as Hv = 1.8537 P-l d-‘, where P is the applied load (kg) and d is the diagonal of indentation (mm).
Determination of conidium germination and appressorium formation
Droplets (60 pl) of conidium suspensions (2 x lo4 conidia ml-‘) were placed on the surfaces of different substrates, which were then kept in a closed plastic chamber at 25 “C with high humidity for an appropriate time. To facilitate observation, fungal structures were stained with a solution consisting of0.2 o/0 trypan blue, 20 o/0 v/v melted phenol, 20 o/o v/v lactic acid, 40 o/o v/v glycerol and 20% v/v water. Four to five hundred conidia were evaluated for each replicate using a light microscope and germination and/or appressorium formation were determined.
Scanning electron microscopic observation of appressorium formation Conidia which germinated and formed appressoria on different substrates were fixed with 5 o/0 glutaraldehyde containing 0.2% tannic acid for 12 h, and then fixed successively for another 12 h by increasing tannic acid content to 2 %. After washing them with distilled water for 2 h, the conidia were further fixed with 1 o/o osmic acid for approximately 12-14 h. The fixed conidia were washed with distilled water for 1 h, then dehydrated stepwise with 30, 50, 70, 90, 99 and lOOo/o ethanol, and finally with 100% 2-methyl-2-propanol each for 20 min. The samples were dried in vacuum and then sputter-coated with gold. Appressoria were examined with a Hitachi S-2350 scanning electron microscope.
RESULTS
Solidity is important for induction of appressorium formation When placing a droplet of conidium suspension on a substrate, most of the conidia settled onto the contact surface within a few minutes and then started to germinate. On some surfaces, the germinated conidia formed appressoria on the tips of germ-tubes. The substrates listed in Tables 1 and 2 were employed. To elucidate the probable

’ 230 J.-Z. Xiao et al.
trigger mechanisms involved in appressorium differentiation, i.e. hydrophobicity, topographic parameters or hardness. Table 2 shows the hardness and hydrophobicity of the solid materials and the rates of appressorium formation on them. Among these, cellophane and nylon 66 are highly hydrophilic with contact angles of approximately 25 and 64 ‘, respectively, while Teflon and parafilm “M” were the most hydrophobic ones with contact angles of more than 100 ‘. On all substrates, conidia produced germ- tubes within 2 h. Continued incubation resulted in appressorium differentiation; small swell,ings appeared on the tips of germ-tubes within 3 h, and mature/melanized appressoria were formed after 6 h incubation. More than 90% of the conidia germinated and more than 85% of germinated conidia formed appressoria without significant difference among the substrates (Table 2).
TABLE 2
Hardness (kg mm-‘) *
contact angle
(degree) t
Appressorium formation (Ob):
Teflon 2.92 Parafilm “A~” nd Polypropylene 8.29 Alkylbenzenstyrene 12.2 Polyethylene 5.59 Polycarbonatc 13.5 Nylon 66 6.94 Cellophane nd Slide glass nd
104-106 87.8k8.5 100 887&11.0
96-98 97.0 * 10.3 94-96 860 f 7.9 92-96 895& IO.5 88-90 8+5f 10.6 64 9@5f6G 24-26 90.1* 13.1 20-26 93.2f 10.1
*Hardness was expressed by Vickcr’s hardness numbers measured as described in Materials and Methods.
tContact angles were measured as described in Materials and Methods. A greater angle indicates a greater hydrophobicity.
:Droplets (GO ~1) of conidium suspensions (2 x IO” conidia ml-‘) in distilled water were placed on each substrate and incubated for 8-10 h. Percentage appressorium formation was determined by assessing number of apprcssoria formed per approximately 400 germ-tubes. Results are the combined data from two separated experiments each with three replications. A similar level of conidium germination and germ-tube elongation was observed for all substrates.
nd, not determined.
Liquid surfaces do not induce appressorium formation Liquid materials of Nissan polybutenes 200N (highly viscous liquid), 30N (viscous liquid), 5N (liquid) and ON (liquid) and liquid paraffin were tested as substrates for conidium germination and appressorium formation (Fig. 1). On contact with these highly hydrophobic surfaces, the conidia germinated, but, the germ-tubes grew into the hydrophobic phase without forming appressoria (Table 1).
Surface roughness is not a prerequisite for appressorium formation Solid substrates listed in Table 2 which were highly inducive of appressorium formation were observed under a scanning electronic microscope (Fig. 2). Among these, the surfaces of polycarbonate, polypropylene and slide glass were the smoothest ones

Appressorium formation by Magnaporthe grisea 231
FIG. I. Diagrammatic cross-section showing conidium germination by ~\4~gnnpor//te grisea at the interface of water and a hydrophobic liquid such as Nissan polybutene and liquid paraffin. A droplet of conidium suspension in distilled water was placed on the surface of the hydrophobic liquid and then incubated for germination. The droplet sank into the middle of hydrophobic phase due to its higher specific gravity and the conidia settled at the interface between the water and the hydrophobic liquid where the conidia germinated.
without any detectable surface roughness or ridging [Fig. 2 (A, D, E)]. Polyethylene, nylon 66 and polalkylbenzenestyrene possessed slightly irregular surfaces but no obvious surface ridges [Fig. 2 (B, C, F)]. No significant topographic features were found on the other substrates (data not shown).
FIG. 2. Scanning electron micrographs of appressoria formed by Mag~taporfhe grisea on different substrates. (A) Polycarbonate, (B) polyethylene, (C) nylon 66, (D) polypropylene, (E) slide glass, (F) alkylbcnzcncstyrcne. Bar = 2 pm.

232 J.-Z. Xiao et al.
* TABLE 3 Appressorirtm formation by Magnaporthe grisea ON agnr membranes*
Drying time Thickness Appressorium
O-4 (mm) formation (%)
0 2.15f01 0 3 1.3e+o.1 2.3k2.0 6 0.78kO.2 1+9*&I 9 CO.2 23.1 f 7.5
12 CO.2 35.8 f 10.5 24 <@2 Sl.Sk3.5
*Droplets (60 11) of conidium suspensions (2 x 10’ conidia ml-‘) were incubated on agar membranes for 8 h. Preparation of agar membranes was done as dcscribcd in Material and Methods. The agar membranes were dried at 45 ‘C for the drying times indicated. Percentage appressorium formation was determined by assessing number of appressoria formed per approximately 400 germ-tubes. High percentages (> 85 o/o) of conidium germination were obtained for each treatment. Results are the combined data from two separate experiments each with three replications per treatment.
FIG. 3. Photographs and diagrams of conidium germination and appressorium formation by Magnaporlhe g&a on different substrates. (A and D) On fresh agar membrane. Germ-tube penetrated the agar gel surface directly without the formation of appressoria. (B and E) On dried agar membrane. Germ-tube differentiated to form a lightly melanized appressorium from which penetration hypha elaborated. (C and F) On other membranes with hard surfaces, such as polycarbonate. Germ-tube differentiated to form a much larger and darkly melanized appressorium but failed to penetrate. Co, conidium; Gt, germ-tube; Ap, appressbrium; Ph, penetration hypha.
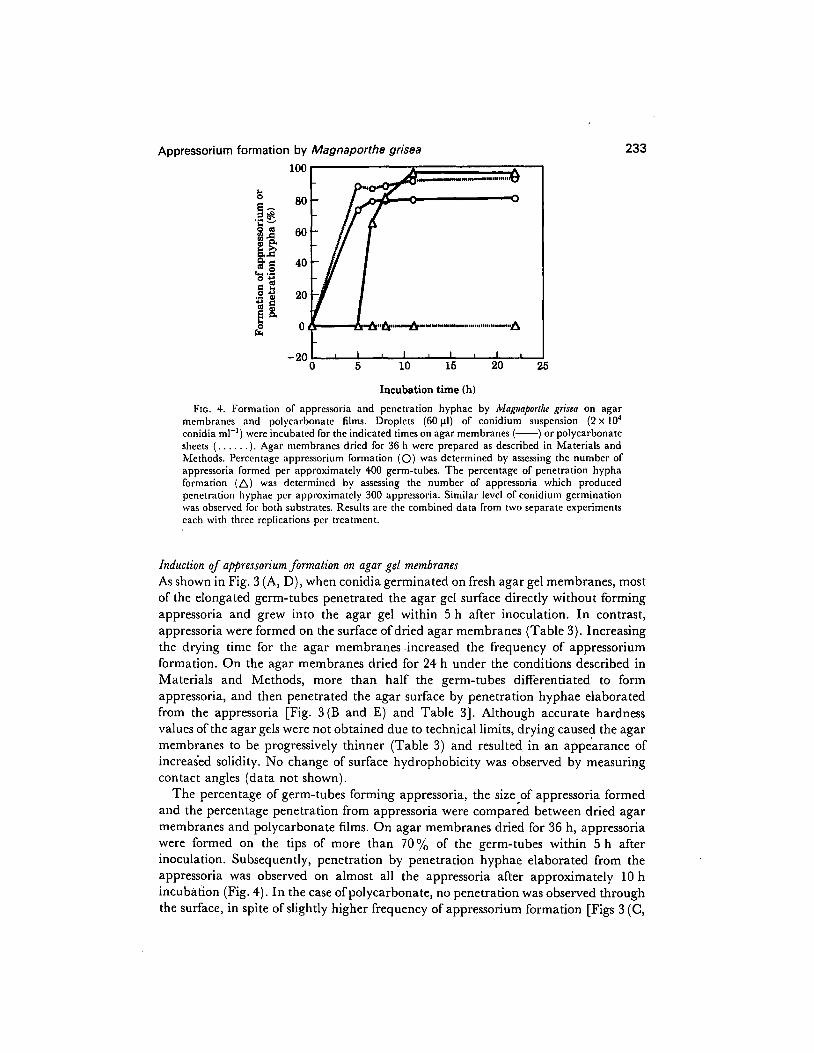
Appressorium formation by Magnaporthe grisea 233
-201 " " " " ' J 0 6 10 15 20 25
Incubation time (h)
FIG. 4. Formation of appressoria and penetration hyphac by ~Lfagttaporfl~ grisea on agar membranes and polycarbonate films. Droplets (60 pl) of conidium suspension (2 x IO4 conidia ml-‘) were incubated for the indicated times on agar membranes (-) or polycarbonate sheets (. . . . .). Agar membranes dried for 36 h were prepared as described in Materials and Methods. Percentage appressorium formation (0) was determined by assessing the number of appressoria formed per approximately 400 germ-tubes. The percentage of penetration hypha formation (A) was determined by assessing the number of appressoria which produced penetration hyphae per approximately 300 appressoria. Similar level of conidium germination was observed for both substrates. Results are the combined data from two separate experiments each with three replications per treatment.
Induction of appressorium formation on agar gel membranes As shown in Fig. 3 (A, D), when conidia germinated on fresh agar gel membranes, most of the elongated germ-tubes penetrated the agar gel surface directly without forming appressoria and grew into the agar gel within 5 h after inoculation. In contrast, appressoria were formed on the surface of dried agar membranes (Table 3). Increasing the drying time for the agar membranes -increased the frequency of appressorium formation. On the agar membranes dried for 24 h under the conditions described in Materials and Methods, more than half the germ-tubes differentiated to form appressoria, and then penetrated the agar surface by penetration hyphae elaborated from the appressoria [Fig. 3 (B and E) and Table 31. Although accurate hardness values of the agar gels were not obtained due to technical limits, drying caused the agar membranes to be progressively thinner (Table 3) and resulted in an appearance of increased solidity. No change of surface hydrophobicity was observed by measuring contact angles (data not shown).
The percentage of germ-tubes forming appressoria, the size of appressoria formed and the percentage penetration from appressoria were compared between dried agar membranes and polycarbonate films. On agar membranes dried for 36 h, appressoria were formed on the tips of more than 70% of the germ-tubes within 5 h after inoculation. Subsequently, penetration by penetration hyphae elaborated from the appressoria was observed on almost all the appressoria after approximately 10 h incubation (Fig. 4). In the case of polycarbonate, no penetration was observed through the surface, in spite of slightly higher frequency of appressorium formation [Figs 3 (C,

234 J.-Z. Xiao et al.
F) and 41. However, the appressoria formed on polycarbonate sheets were larger than those formed on the dried agar membranes (average size in diameter: 8.5 +0*5 u. 49+0.9 mm), and were much more darkly pigmented.
DISCUSSION
Infection of rice plants by conidial germlings of Ad. g&~ involves germination, formation of an appressorium at the tip of the germ-tube and penetration through the rigid plant cuticle and cell walls by an infection hypha elaborated from the appressorium. Similar processes have been observed apart from the host, even on substrates which provide no nutrients [Z, 101. Melanized appressoria are known to be structures with a high internal hydrostatic pressure which enables the pathogen to penetrate the hard surface by a thin penetration hypha [9,10]. Thus, formation of the appressorium by M. grisea might be considered as a tool for elective penetration of a hard solid.
Our results indicated that the recognition of solidity of the surface could trigger appressorium formation by h/l. grisea. The results also suggest that the hydrophobicity and topographic features of the surface are not of importance. Appressoria formed at high frequency on all kinds of solid surfaces tested. Hydrophilic and hydrophobic surfaces induced appressorium formation to a similar extent and it made no difference whether the surface was smooth or rough. Appressoria were not formed on liquid surfaces, such as liquid paraffin and Nissan polybutenes. On these surfaces, the germ- tubes grew inside the hydrophobic liquids. Since appressorium formation is presumably an energy-consuming process to the pathogen, it may be triggered only when necessary, i.e., when the germ-tube fails to penetrate the surface.
Recognition of solidity was further implicated by the formation of appressoria on dried agar membranes. Fresh agar gel surfaces were shown not to induce appressorium formation. In fact, the germ-tubes of M. grisea penetrated the soft agar surface directly. Because preparation of a sufficiently hard agar membrane by increasing the agar content was technically impractical, we did it by drying the soft agar membrane. Drying caused the agar gel to be progressively thinner and harder without altering surface hydrophilicity. AS expected, the germ-tubes failed to penetrate the harder agar surface. Instead, the germ-tubes differentiated to form appressoria and then penetrated the surface by penetration hyphae elaborated from the appressoria. However, agar gels are not suitable materials for the elucidation of any correlation between degrees of hardness and effectiveness in induction of appressoria because the dried agar gels reabsorb a considerable amount of water during incubation. We are employing other materials for further studies on this phenomenon.
Plant pathogens seem to employ different mechanisms to regulate appressorium formation. n/r. grisea infects host plants by directly penetrating through the cuticle layers and entering epidermal cells. Penetration of the rigid plant surface is a prerequisite for a successful infection. For U. appendiculatus, on the other hand, which infects bean leaves by penetration through stomata, recognition of the infection sites may be the most important event. U. appendiculatus was shown to form appressoria in response to a specific topographic signal, namely, a ridge of approximately 0.5 l.tm high, which equates to the height of bean leaf stomata1 guard cells [8]. Clearly, these

Appressorium formation by Magnaporthe grisea 235
two fungi differ in requirements for appressorium formation in line with their different modes of entering plant tissues.
Lee and Dean [II] discussed the endogenous and exogenous signals controlling the development process of appressorium formation by M. grisea. They inferred that appressorium formation is induced by the hydrophobicity of the surfaces of rice leaves or artificial substrata. This appeared to be based mainly on observations of rare appressdrium formation on the surfaces of slide glasses and GelBond (polyester sheet) coated with Agarose (hydrophilic). Unused clean slide glass often gives poor appressorium formation. The reason for this is not clear but well-washed or used slide glasses are known to be more favourable for induction of appressoria (H. Yaegashi, National Institute ofAgro-Environmental Sciences, Ibaraki 305, Japan, pers. comm.). We found no significant difference in the hydrophobicity between well-washed and new, unwashed slide glasses. Further, an Agarose film formed on GelBond sheets might have interfered with appressorium formation as was the case on the soft surface of fresh agar used in the present study. Other workers have also shown appressorium formation on hydrophilic surfaces, such as cover glass and cellophane membrane [Z, 9, 15, 161.
The authors are grateful to Dr T. Arie and MS S. Sekido, Microbial Toxicology Laboratory, RIKEN, for their valuable discussions. This research was supported in part by a grant for “Biodesign Research Program ” from The Institute of Physical and Chemical Research (RIKEN).
REFERENCES
I, Bhairi SM, Staples RC, Freve P, Yoder OC. 1989. Characterization of an inrection structure-specific gent from the rust fungus Urongses apperrdiculalrrs. Cm 81 : 237-243.
2. Bourett TM, Howard RJ. 1990. In vitro development ofpenetration structure in the rice blast fungus Magnapor~he grisea. Canodinn Jorrrnnl of Bofagl 68: 239-342.
3. Chumley FG, Valent B. 1990. Genetic analysis of melanin-deficient, nonpathogenic mutants or Magnoporthe grisea. Molecular Plan-Microbe Inferaclions 3 : I53- 143.
4. Emmett RW, Parbery DG. 1975. Apprcssoria. cbt~tral Ret&v ~JPh~~~opnll~olog~~ 13: 147-167. 5. Hamer JE, Howard RJ, Valent B, Chumley FG. 1989. A mechanism for surface attachment in spores
of a plant pathogenic fungus. Sciettce ‘239: 288-290. 6. Hamer JE, Valent B, Chumley FG. 1989. Mutations at the SMO genetic locus affect the shape of
diverse cell types in the rice blast fungus. &relics 122: 35 l-361. 7. Hoch HC, Staples RC. 1987. Structural and chemical changes among the rust fungi during
appressorium development. Atmual Review of Pi!,topnlholo~ 25: 23 l-247. 8. Hoch HC, Staples RC, Whitehead B, Comeau J, Wolf ED. 1987. Signalling for growth orientation
and ccl1 differentiation by surface topography in Lirog~ces. Sciertce 235: 1659%IGG2. ’ 9. Howard R, Ferrari M. 1989. Role of melanin in appressorium function. Experimental A(~cologv 13:
403-418. 10. Howard R, Ferrari MA, Roach DH, Money NP. 1991. Penetration of hard substrates by a fungus
employing enormous turgor pressures. Proceeding oJ.Nafionol Academic Science U.S.A. 88: I1 281-1 1284. 1 I. Lee Y-H, Dean RA. 1993. CAMP regulates infection structure formation in-the plant pathogenic fungus
Mngnaporlhe grisea. The Plant Cell 5: 693-700. 12. Shaw MC. 1966. Mechanical Be/r&or of Moferials. McClintock PA, Argon A, eds. Reading, MA: Addison-
\‘\‘esley, 443448. 13. Staples RC, Hoch HC. 1987. Infection structures-form and function. Experimental ~Mycolosy 11:
163-169. 14. Staples RC, Hoch HC, Epstein L, Laccetti L, Hassouna S. 1985. Recognition of host morphology
by rust fungi : responses and mechanisms. Canadian Journal oJ Plan1 Pathology 7: 3 14-322. 15. Uchiyama T, Okuyama K. 1990. Participation of Oryra s&n leaf wax in appressorium formation
by Qricttloria oryzae. Plytochemis!rJ~ 29 : 9 I-92.

236 J.-Z. Xiao et al.
16. Uchiyama T, Ogasawara N, Nanba Y, Ito H. 1979. Conidial germination and appressorial formation of the plant pathogenic fungi on the cover glass or cellophane coated with various lipid components of plant waxes. rlgricultural and Biological Chemistry 43: 383-384.
17. Xuei X, Bhairi SM, Staples RC, Yoder OC. 1992. Characterization of INF56, a gene expressed during infection structure development oT Chornyces a),bendiculatus. Gene 110: 49-55.
18. Yamaguchi I, Sekido S, Misato T. 1982. The effect ofnon-fungicidal anti-blast chemicals on melanin biosynthesis and infection by Pyricslaria opus. j’orwnal of Peslicide Science 7: 523-529.
19. Yamaguchi I, Sekido S, Misato T. 1983. Inhibition of appressorial melanization in Pyricelaria o<~oe by non-fungicidal anti-blast chemicals. journal of Pesticide Science 8: 545-550.




















