
Stone tool use by adult wild bearded capuchin monkeys ( Cebus libidinosus). Frequency, efficiency...
11
Author's personal copy Stone tool use by adult wild bearded capuchin monkeys (Cebus libidinosus). Frequency, efficiency and tool selectivity Noemi Spagnoletti a, b , Elisabetta Visalberghi a, * , Eduardo Ottoni c , Patricia Izar c , Dorothy Fragaszy d a Istituto di Scienze e Tecnologie della Cognizione, Consiglio Nazionale delle Ricerche, Via Aldrovandi 16B, 00161 Rome, Italy b Dipartimento di Biologia Animale e dell’Uomo, Università La Sapienza di Roma, Rome, Italy c Institute of Psychology, Department of Experimental Psychology, University of São Paulo, São Paulo, Brazil d Psychology Department, University of Georgia, Athens, GA 30602, USA article info Article history: Received 22 October 2009 Accepted 9 February 2011 Keywords: Nut cracking Body mass Energetic costs Hammer selection Sexual differences abstract Chimpanzees have been the traditional referential models for investigating human evolution and stone tool use by hominins. We enlarge this comparative scenario by describing normative use of hammer stones and anvils in two wild groups of bearded capuchin monkeys (Cebus libidinosus) over one year. We found that most of the individuals habitually use stones and anvils to crack nuts and other encased food items. Further, we found that in adults (1) males use stone tools more frequently than females, (2) males crack high resistance nuts more frequently than females, (3) efficiency at opening a food by percussive tool use varies according to the resistance of the encased food, (4) heavier individuals are more efficient at cracking high resistant nuts than smaller individuals, and (5) to crack open encased foods, both sexes select hammer stones on the basis of material and weight. These findings confirm and extend previous experimental evidence concerning tool selectivity in wild capuchin monkeys (Visalberghi et al., 2009b; Fragaszy et al., 2010b). Male capuchins use tools more frequently than females and body mass is the best predictor of effi- ciency, but the sexes do not differ in terms of efficiency. We argue that the contrasting pattern of sex differences in capuchins compared with chimpanzees, in which females use tools more frequently and more skillfully than males, may have arisen from the degree of sexual dimorphism in body size of the two species, which is larger in capuchins than in chimpanzees. Our findings show the importance of taking sex and body mass into account as separate variables to assess their role in tool use. Ó 2011 Elsevier Ltd. All rights reserved. Introduction Stone pounding was a likely precursor of tool making by hom- inins, since the actions of pounding “can lend themselves to the application of the same bodily motor actions to different raw materials and with novel purposes” (Davidson and McGrew, 2005: 796). Among nonhuman primates living in natural settings, only a few populations of chimpanzees routinely use stone tools to crack nuts (Whiten et al., 1999; Boesch and Boesch-Achermann, 2000). Chimpanzees have been the traditional referential models for those investigating human evolution and the features characterizing hominins’ use of stone tools, including the origins of sex differences in tool use and in the division of labor (McGrew, 1979; Tooby and DeVore, 1987). Our goal here is to expand this comparative scenario to include Cebus libidinosus, a South American monkey species that last shared a common ancestor with the Catarrhini about 35 million years ago (Schrago and Russo, 2003). A few populations of this species have been recently discovered to use tools in the wild (Fragaszy et al., 2004a; Moura and Lee, 2004; Waga et al., 2006; Canale et al., 2009; see Ottoni and Izar, 2008 for a review). As argued by Haslam et al. (2009), these findings may (a) challenge the role of continuity in primate tool use, (b) suggest that convergence is plausible (probably owing to similar adaptive pressures, foraging requirements and/or physiological constraints), and (c) open the door to the possibility of repeated gain and loss of tool use among extinct primates in multiple lineages over millions of years. Specifically, on the basis of one year of systematic observations of stone tool activities by adult capuchins, we examine the frequency of tool use, success and stone selectivity in relation to the resistance of food targets. These findings on the spontaneous behavior of wild capuchin monkeys, together with those collected in a series of parallel field experiments (for a review Visalberghi and Fragaszy, in * Corresponding author. E-mail address: [email protected] (E. Visalberghi). Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol 0047-2484/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.jhevol.2011.02.010 Journal of Human Evolution 61 (2011) 97e107
Transcript of Stone tool use by adult wild bearded capuchin monkeys ( Cebus libidinosus). Frequency, efficiency...

Author's personal copy
Stone tool use by adult wild bearded capuchin monkeys (Cebus libidinosus).Frequency, efficiency and tool selectivity
Noemi Spagnoletti a,b, Elisabetta Visalberghi a,*, Eduardo Ottoni c, Patricia Izar c, Dorothy Fragaszy d
a Istituto di Scienze e Tecnologie della Cognizione, Consiglio Nazionale delle Ricerche, Via Aldrovandi 16B, 00161 Rome, ItalybDipartimento di Biologia Animale e dell’Uomo, Università La Sapienza di Roma, Rome, Italyc Institute of Psychology, Department of Experimental Psychology, University of São Paulo, São Paulo, Brazild Psychology Department, University of Georgia, Athens, GA 30602, USA
a r t i c l e i n f o
Article history:Received 22 October 2009Accepted 9 February 2011
Keywords:Nut crackingBody massEnergetic costsHammer selectionSexual differences
a b s t r a c t
Chimpanzees have been the traditional referential models for investigating human evolution and stonetool use by hominins. We enlarge this comparative scenario by describing normative use of hammerstones and anvils in two wild groups of bearded capuchin monkeys (Cebus libidinosus) over one year. Wefound that most of the individuals habitually use stones and anvils to crack nuts and other encased fooditems. Further, we found that in adults (1) males use stone tools more frequently than females, (2) malescrack high resistance nuts more frequently than females, (3) efficiency at opening a food by percussivetool use varies according to the resistance of the encased food, (4) heavier individuals are more efficientat cracking high resistant nuts than smaller individuals, and (5) to crack open encased foods, both sexesselect hammer stones on the basis of material and weight. These findings confirm and extend previousexperimental evidence concerning tool selectivity in wild capuchin monkeys (Visalberghi et al., 2009b;Fragaszy et al., 2010b).
Male capuchins use tools more frequently than females and body mass is the best predictor of effi-ciency, but the sexes do not differ in terms of efficiency. We argue that the contrasting pattern of sexdifferences in capuchins compared with chimpanzees, in which females use tools more frequently andmore skillfully than males, may have arisen from the degree of sexual dimorphism in body size of thetwo species, which is larger in capuchins than in chimpanzees. Our findings show the importance oftaking sex and body mass into account as separate variables to assess their role in tool use.
� 2011 Elsevier Ltd. All rights reserved.
Introduction
Stone pounding was a likely precursor of tool making by hom-inins, since the actions of pounding “can lend themselves to theapplication of the same bodily motor actions to different rawmaterials and with novel purposes” (Davidson and McGrew, 2005:796). Among nonhuman primates living in natural settings, onlya few populations of chimpanzees routinely use stone tools to cracknuts (Whiten et al., 1999; Boesch and Boesch-Achermann, 2000).Chimpanzees have been the traditional referential models for thoseinvestigating human evolution and the features characterizinghominins’ use of stone tools, including the origins of sex differencesin tool use and in the division of labor (McGrew, 1979; Tooby andDeVore, 1987).
Our goal here is to expand this comparative scenario to includeCebus libidinosus, a South Americanmonkey species that last shareda common ancestor with the Catarrhini about 35 million years ago(Schrago and Russo, 2003). A few populations of this species havebeen recently discovered to use tools in the wild (Fragaszy et al.,2004a; Moura and Lee, 2004; Waga et al., 2006; Canale et al.,2009; see Ottoni and Izar, 2008 for a review). As argued byHaslam et al. (2009), these findings may (a) challenge the role ofcontinuity in primate tool use, (b) suggest that convergence isplausible (probably owing to similar adaptive pressures, foragingrequirements and/or physiological constraints), and (c) open thedoor to the possibility of repeated gain and loss of tool use amongextinct primates in multiple lineages over millions of years.Specifically, on the basis of one year of systematic observations ofstone tool activities by adult capuchins, we examine the frequencyof tool use, success and stone selectivity in relation to the resistanceof food targets. These findings on the spontaneous behavior of wildcapuchin monkeys, together with those collected in a series ofparallel field experiments (for a review Visalberghi and Fragaszy, in
* Corresponding author.E-mail address: [email protected] (E. Visalberghi).
Contents lists available at ScienceDirect
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
0047-2484/$ e see front matter � 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.jhevol.2011.02.010
Journal of Human Evolution 61 (2011) 97e107

Author's personal copy
press-a), constitute an ample dataset on which to build compari-sons with chimpanzees and to interpret the behavior of homininsbefore the occurrence of flaking technology.
In many chimpanzee populations, several members of a groupuse tools throughout the year (Whiten et al., 1999), thus earning thelabel of “habitual” tool users (sensu McGrew, 1992). Habitual tooluse suggests that technology plays a significant role in the ecologyof the population, as opposed to an incidental activity. Here, weprovide longitudinal data for a population of wild capuchins,showing that they are also habitual tool users.
In chimpanzees, the use of tools differs markedly between thetwo sexes. Females use both probing tools and stone tools morefrequently than males (see McGrew, 1992 for a review). Forexample, in Gombe, female chimpanzees engage in termite fishingthree times more often than males (McGrew, 1979; see alsoPandolfi et al., 2003 for an analysis of the data collected in Gombeover 17 years). Females start to fish for termites at a younger agethan males (31 vs. 50 months) and achieve higher levels of profi-ciency. Moreover, females use a technique similar to their mother’s,whereas males do not (Lonsdorf et al., 2004). Females outperformmales in the two most difficult forms of stone tool use that chim-panzees practice, namely cracking Coula nuts aloft in the tree andcracking Panda nuts (which are the most resistant nuts thatchimpanzees crack) (Boesch and Boesch, 1984). Finally, femalescrack Coula nuts regardless of whether the nuts are fresh and moredifficult to crack, or dry and less difficult to crack, whereas malescrack mostly dry, brittle nuts (Boesch and Boesch, 1984). Thus,female chimpanzees, across many regions and foraging tasks, usetools more often and in some circumstances more skillfully thanmales. This difference has been interpreted as a strategy by whichfemales obtain reliable foods, whereas males devote more time tosocial activities and to other forms of foraging, such as hunting(Boesch and Boesch, 1984).
Sex differences in tool use in wild capuchins have been littleinvestigated. The few studies on tool use carried out so far withcapuchins provide mixed evidence (see Moura and Lee, 2010 ona group of C. libidinosus partially composed of reintroduced indi-viduals; Ottoni andMannu, 2001 on a semi-free ranging group; andFragaszy et al., 2004b for review of studies with captive capuchins).Similarly, the results from the studies examining precursors of tooluse, namely actions combining objects and surfaces (for a review,see Fragaszy et al., 2004b) are contradictory. For example, inFragaszy and Adams-Curtis (1991) and in Byrne and Suomi (1996),male and female capuchins interact with objects with similardegrees of interest, whereas in Visalberghi’s (1988) study malesscore significantly higher than females for the pooled category ofactions, which include pounding, throwing, and rubbing. Boinskiet al. (2000) report that wild Cebus apella males process huskedfruit by pounding them on hard substrates more often thanfemales, although no sex difference in skill and coordination isapparent. In contrast, wild female wedge-capped capuchins (Cebusolivaceus) devote absolutely more time than males to foraging and,while foraging, to banging food items (Fragaszy, 1990 and unpub-lished results). Finally, males are prominent when percussion isused as an aggressive display and as a sexual display (Moura, 2007).Boinski (2004) argues that percussion by males may attractfemales’ attention, and serve as a signal of male fitness.
Although the published findings concerning sex differences intool use and object manipulation in capuchins are contradictory,energetic and morphological considerations lead to clear predic-tions about sex differences in frequency and performance of tooluse, and stone selectivity by adult wild capuchins.
Other factors being equal, the nutritional needs of an animal areaffected both by reproduction (during the present study seven outof eight of our adult females were either pregnant and/or lactating)
and body mass (Key and Ross, 1999). In primates, the degree ofsexual dimorphism in body mass varies greatly among species(Plavcan, 2004) and capuchins are more dimorphic in body massthan chimpanzees (Smith and Jungers, 1997; Key and Ross, 1999;Fragaszy et al., 2010a; see Discussion for details). In particular, thebody mass of the adult males observed in the present study was64% larger than the body mass of the adult females (see Table 1).Key and Ross (1999) examined the relationship between energeticcosts for males and females in 19 nonhuman primate species (notincluding capuchins) and found that when the males’ body massexceeds females’ bodymass by 60% or more, themajor determinantof sex differences in energetic costs is body mass dimorphism,whereas whenmales are between 30% and 50% larger than females,the costs due to reproduction almost balance those of the males’larger body mass.
Since male capuchins needmore energy than females due to themales’ larger body mass, our first prediction is that adult malecapuchins will use tools to crack nuts as or more frequently thanadult females. Second, we predict that heavier (and thus presum-ably stronger) individuals (i.e., adult males) will be more efficient atcracking nuts (in terms of rate of success, average number of strikesper successful nut cracking episode, and average duration persuccessful episode) than smaller individuals (i.e., adult females andjuveniles of both sexes). However, consideration of the actioninvolved in cracking leads to amore refined set of predictions. Somespecies of palm nuts are extremely resistant to cracking(Visalberghi et al., 2008), and thus the monkeys must strike eachnut several times to crack it (Fragaszy et al., 2010a). Body mass isa strong predictor of efficiency in cracking (measured as thenumber of strikes needed to crack a piassava nut, the most resistantspecies of nuts; Fragaszy et al., 2010a) and monkeys of both sexesprefer to use heavier stones to crack nuts (Visalberghi et al., 2009b;Fragaszy et al., 2010b).
Therefore, on the basis of dimorphism in bodymass in capuchinsand energetic considerations, we make three further predictions.One is that the resistance of the nut (Visalberghi et al., 2008) willaffect the rate of success, the average number of strikes per episode,and the duration of successful episodes (third prediction). Theothers are that females, to maximize reliability of energetic return,will exploit high resistance nuts proportionally less often thanmales(fourth prediction), but will exploit low resistance nuts and otherencased foods at the same frequency as males (fifth prediction).
Inwild chimpanzees, deliberate selection of specific materials touse as tools has been inferred by comparing the features ofhammers and anvils used to crack open nuts of different resistance.Boesch and Boesch (1984) proceeded in this way at Taï and inferredthat high resistance Panda oleosa nuts were more often crackedopen on stone anvils and stone hammers than with wooden anvilsand hammers, compared with the less resistant Coula edulis nuts.Furthermore, chimpanzees use heavier stone hammers to crackopen Panda nuts than Coula nuts. Therefore, chimpanzees seem totake into account the resistance of nuts when using percussivetools. Subsequent direct observations confirmed this pattern of tooluse in Taï (Boesch and Boesch-Achermann, 2000) as well as inBossou (Matsuzawa, 1994), indicating that chimpanzees “canrecognize the functions of the tools and which stone or combina-tion of stones function best” (Matsuzawa, 1994: 361). Recently,Visalberghi et al. (2009b) experimentally demonstrated that wildcapuchins, belonging to the same population investigated in thepresent study, when presented with a concurrent choice of two ormore stones, selectively used stones of particular material andweight to crack low resistance nuts (see also Fragaszy et al., 2010b).Therefore, our sixth and last prediction is that capuchins will selectstone tools based on the material and weight that are adequate toovercome the resistance of the food resource at hand.
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e10798

Author's personal copy
Methods
Site
The study area is located at Fazenda Boa Vista (FBV) in thenortheastern Brazilian state of Piauí (9�390S, 45�250W), 21 kmnorthwest of the town of Gilbués. The physical geography of thefield site is a sandy plain at approximately 420 m above sea level,punctuated by sandstone ridges, pinnacles and plateaus, and sur-rounded by cliffs composed of sedimentary rock rising steeply from20 to 100 m above the plain. The cliff and plateau consist of inter-bedded sandstone, siltstone and shale. Boulders often break off ofthese formations and fall to the base of the cliff close to the plain(for further information about the geology of FBV, see Visalberghiet al., 2007). The sandstone cliffs and plateaus are heavily erodedand there are ephemeral watercourses that have running wateronly after rainfall. The climate is seasonally dry (average annualrainfall 1156 mm, total rainfall during dry season, April toSeptember 230 mm, data from 1971 to 1990, source: Embrapa,Brazilian Agricultural Research Corporation).
FBV presents four types of vegetation physiognomies accordingto the terrain and the proximity towater sources. The sandy plain ischaracterized by a high abundance of palms with subterraneanstems and trees of medium height such as Eschweilera nana andHymaenaea courbaril. The vegetation by the lowest area of theplain, a marsh, is characterized by a higher diversity of treesforming gallery forests and a high density of the tall palm tree,Mauritia flexuosa. Shrubs and small trees dominate the cliff and thetalus, whereas herbaceous vegetation dominates on the plateau,especially bromeliads and cactus.
Subjects
From June 2006 to May 2007, we observed the behavior of 28bearded capuchins living in two groups, the Chicão group (CH) andthe Zangado group (ZA) (eight adult females, seven adult males,
nine juveniles and four infants). Adult status was conferred on thebasis of size and behavior for animals of unknown age. At thebeginning of the study, all adult females except one were eitherpregnant or lactating.
Behaviors and other measures scored
Each group was followed from dawn to dusk for seven to tendays per month by Noemi Spagnoletti (hereafter N.S.) or Eduardo D.Ramos da Silva (hereafter E.D.R.S.), aided by a field assistant (seebelow). For 1709 hours, we sampled all episodes of tool use (bothwhen in progress or about to begin) until they ended using adlibitum sampling. A tool use episode started when the subject (S)approached the anvil site (by definition, an anvil site is a boulder oran exposed stone with a hammer stone on it or within threemetres; for details see Visalberghi et al., 2007) with an encasedfood item and ended when the item was cracked open, or when Sleft the anvil without the hammer stone. When S began to cracka new food item, a new episode was recorded. If wewere observinga tool use episode and another began, we continued to observe thefirst episode. During concurrent episodes, the field assistant notedthe identity of the tool user in the second episode and collected thesame data about it. This led to a total of 1565 tool use episodes thatwere used to analyze tool use frequencies in the two groups. Theanalyses concerning sex differences in frequency of tool use and thefood exploitedwere based on the 1310 episodes performed by adultcapuchins. For the analyses on sex differences in tool use efficiency,we included an additional 47 episodes, which were observed whilethe subjects cracked nuts provided by the observers for purposes ofphotography and videography. N.S. collected detailed data on 844episodes that were used for all of the analyses (frequency, efficiencyand selectivity of tool use, see below). E.D.R.S. contributed 513episodes collected with ad libitum sampling, while conductinga concurrent study using the same behavioral definitions (Ramosda Silva, 2008).
Table 1Adult subjects, group towhich they belong (CH¼ Chicão group; ZA¼ Zangado group), body mass, rank, total number of tool use episodes observed, number of incomplete andcomplete episodes involving palm nuts and other food items.
Group Subject Body mass (kg) Rank # of episodes of tool use
Total Incomplete Complete Complete and identified
Nuts Other food items Nuts Other food items Low resistance nuts High resistance nuts
Males CH Chicão 4.4 1 142 130 12 89 12 52 21ZA Zangado 4.4 e 1 296 276 20 77 4 29 28ZAeCH Mansinho 3.5 4 196 193 3 108 2 65 20ZAeCH Dengoso 3.3 3 159 147 12 64 9 21 19ZAeCH Jatoba 2.7 6 103 93 10 40 7 13 5ZAeCH Teimoso 2.7 3 84 72 12 28 7 10 7CH Segundo* 3.6 e 3 14 13 1 7 1 e
Subtotal 994 924 70 413 41 190 100
Females CH Dendé 1.9 5 18 15 3 11 3 19 4CH Teninha 2.2 3 25 24 1 20 1 11 11CH Piassava 1.9 2 55 40 15 24 20 16 1CH Chuchu 2.2 3 60 57 3 39 3 19 11ZA Moça-Br 1.9 e 2 44 27 17 6 8 1 2ZA Moça-Ch 2.2 e 3 141 100 41 29 13 16 6ZA Emilia* 2.4 e 4 20 6 14 0 6 e e
CH Chiquinha* 2.4 3 0 0 0 0 0 e e
Subtotal 363 269 94 129 54 68 29
Total 1357 1193 164 542 95 258 129
The complete episodes were those recorded from the beginning to the end for which success, latency to solution and number of strikes were scored. The complete andidentified episodes were those in which the species of the nut was identified.ZAeCH indicates individuals that during data collection migrated from ZA group to CH group. The weight of each subject was either assessed with a scale (see Fragaszy et al.,2010a) or estimated (e). (*) individuals excluded from the statistical analyses concerning success and tool selectivity.
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107 99

Author's personal copy
As in Resende et al. (2008), N.S. noted per episode whether the(i) food itemwas cracked (effective nut cracking), (ii) food itemwasnot cracked despite the correctness of both the positioning of thetool in relation to the food item and the strikes (non-effective nutcracking), and (iii) food itemwas not cracked because the behaviorwas inadequate (simple pounding, i.e., episodes in which themonkey pounded the nut on the anvil or on the hammer). Whenpossible, N.S. observed the episode from the beginning to the end,noted its duration, counted the number of strikes (i.e., the action ofpounding the food item positioned on the anvil with the hammer)performed by the monkey, and the outcome of the episode.Episodes for which all of these variables were recorded are labeledcomplete episodes. For the complete episodes resulting in a crackednut and access to the kernel, we scored the number of strikes andthe time elapsed (in seconds) between the beginning of the episodeand access to the kernel (duration of episode). When possible, N.S.also determined the food exploited and its state at the beginning ofthe episode (intact or partially cracked) on the basis of directobservation.
Finally, N.S. examined the hammer stone used by the subjectand noted its weight and lithology at the end of the episode, whenpossible. Lithology was assessed on the basis of experience andresemblance to the stones available in our collection in FBV.Weightwas assessed to the nearest 10 g (stones up to 0.5 kg) or the nearest20 g (stones from 0.5 kg up to 2.5 kg) using spring scales (Pesola,Switzerland). N.S. also noted whether the subject had used a stoneor an intact palm nut as a “hammer.” Hammer stones were cate-gorized as “friable” or “durable.” Visalberghi et al. (2007) reportthat the durable and effective hard stones that capuchins use ashammers to crack open palm nuts are quartzite, siltstone andironstone (all of these stones are relatively harder and more densethan the prevailing friable sandstone).
The items processed belong to three major categories: (a) nutswith relatively high peak-force-at-failure, namely piassava(Orbignya sp.) and catulí (Attalea sp.), hereafter labeled “highresistance” nuts; (b) nuts with relatively low peak-force-at-failure,hereafter labeled “low resistance” nuts, namely catulè (Attaleabarreirensis) and tucum (Astrocaryum campestre) (see Visalberghiet al., 2008 for details), and (c) other encased food items whosepeak-force-at-failure is much lower than that reported for palmnuts (e.g., fruta-danta, family Icacinaceae; caju, family Anacardia-ceae; caroba, family Bignoniaceae; manioca-brava, family Euphor-biaceae). Whereas palm nuts in categories (a) and (b) requirehammers of specific lithology and weight, the other encased fooditems (c) can be cracked open with lighter and more friable stones.
Analyses
Since monthly frequencies of tool use did not differ betweengroups (ManneWhitney U-test¼ 64, n1¼12, n2¼12, p¼ 0.64, seealso Spagnoletti, 2009), we pooled the data from the two groups. Torule out the possibility that the number of observed episodesdepended on the different likelihood of observing adult males,adult females, and juveniles, we determined whether these sex andage classes were similarly “in view” to the observer. When themonkeys were not using tools, N.S. sampled their activity every20 min using group scan sampling (Martin and Bateson, 1993; seealso Spagnoletti, 2009). An index of individual visibility is indicatedby the number of scans in which each of the 22 tool users wasobserved/total number of scans recorded. These values were notsignificantly different among adult males, adult females and juve-niles (KruskaleWallis ANOVA, H2,22¼ 4.48, p¼ 0.11). Moreover, thenumber of tool use episodes and visibility were not significantlycorrelated (Spearman, rs¼ 0.03, N¼ 22, p¼ 0.89). As there was nobias in the visibility of males versus females in scan sampling, our
data can be considered unbiased with respect to the frequencies oftool use.
Analyses concerning frequencies of tool use were run on all theadult subjects (seven males and eight females). Analyses ofperformance and selectivity were carried out on six adult malesand six adult females (see Table 1). We used the ManneWhitneytest to compare sex differences in frequency of tool use and cate-gory of food exploited. The analyses on performance were based onmixed model regressions, either linear or logistic, in which thesubject identities were inserted as random factors (Rabe-Heskethand Skrondal, 2008). We used mixed model logistic regressions totest whether efficiency (i.e., the success to crack open the fooditem) was affected by body mass, sex, individual rank, resistance ofthe nut and weight of the stone. We used mixed model linearregression to test whether the number of strikes and duration ofepisodes to crack open nuts was related to body mass, sex, resis-tance of the nuts and weight of the stone. In order to explore thequestion of whether adults select hardness and weight of the stonehammer, we usedmixed model linear regressions to test separatelythe effects of individual body mass and sex, and food resistance onthe hardness and weight of the hammer stones used. Furthermore,we used mixed model linear regressions to test if individual bodymass and food resistance predict the weight of the hammer stonesused by males and females separately.
The individuals of the CH group were weighed in July 2007(Table 1; for details about the weighing procedure see Fragaszyet al., 2010a). The body mass of the members of ZA group wasestimated by two independent observers very familiar with bothgroups. To do so, each observer independently paired each indi-vidual of ZA group with the individual(s) most similar in body sizebelonging to the CH group. The two estimates per subject obtainedin this way were averaged.
In each group, we tested if the number of tool use episodesperformed by each individual was related to its rank in the groupdominance hierarchy. Individual rank in each group was estab-lished by Verderane (2010) as a part of her study on social rela-tionships of the two groups, based on all occurrences of contestcompetition analyzed through the Dominance Directed Treemethod (Izar et al., 2006).
Since our data did not followanormal distribution (Lilliefors test,p< 0.05) for the analyses concerning sex differences in frequency oftool use, we used non-parametric statistics (using STATISTICA 7Statsoft). The mixed model regressions were run using Stata 10.1(StataCorp, 2007). For all analyses, the significance level was set atp< 0.05 and all reported probabilities are two tailed.
Results
Frequency of tool use
During the study period, 22 capuchins (all individuals in bothgroups with the exception of one adult female, one juvenile femaleand the four infants) used stones to crack opennuts or other encasedfoods.We observed 1565 episodes of tool use in the two groups overthe course of the study (0.92 episodes per hour/per group). Fourteenadults (seven males, seven females) performed 84% of the episodes(N¼ 1310) and eight juveniles performed the remaining 16% of theepisodes (N¼ 255). The seven adult males accounted for 73.5%(N¼ 963) and the seven adult females for 26.5% (N¼ 347) of the tooluse episodes performed by adults. Tool use episodes primarilyoccurred on sandstone anvils (80%) and log anvils (19%); othersubstrates were rarely used (1%) (Fig. 1). The monkeys used a stoneas the percussor in 98.5% of the episodes. A palm nut was used aspercussor in 24 episodes; 15 episodes to crack open another palmnut and nine episodes to crack open other food items.
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107100

Author's personal copy
Of the nine juveniles, the two oldest (Tucum, an approximatelytwo year-oldmale and Dita, an approximately three to four year-oldfemale, at the beginning of the study) succeeded in opening intactnuts. The other juveniles often pounded nuts on hard surfaces andhad success with tools only when cracking food items other thanwhole nuts, such as partially opened nuts. Finally, one juvenilefemale never used a tool (for further information see Spagnoletti,2009). Since juveniles used tools relatively infrequently, we onlycompare the data recorded for adult males and females.
Sex differences in tool use by adults and food exploited
As shown in Table 1, 86% of the episodes of tool use performedby adults involved palm nuts and 14% involved other food items.Adult males accounted for a higher number of nut crackingepisodes than did adult females (median (interquartile range, IQR):130 (87.5) and 27 (29), respectively; ManneWhitney, U¼ 8, nm¼ 7,nf¼ 8, p< 0.05); whereas the number of episodes aimed at crackingfood items other than nuts did not differ between males andfemales (median (IQR): 12 (5.5) and 14 (13), respectively; Man-neWhitney, U¼ 27.5, nm¼ 7, nf¼ 8, p¼ 0.95). For each group, thecorrelation between an individual’s number of episodes of tool useand its social rank was not significant (CH group rs¼ 0.05, n¼ 11,p¼ 0.88; ZA group rs¼�0.8, n¼ 4, p¼ 0.2).
Monkeys used tools more frequently to crack low resistancenuts (66% and 70% of the episodes for males and females, respec-tively) than high resistance nuts (34% and 30% of the episodes formales and females, respectively). Adult males used tools to crackhigh resistance nuts significantly more often than did adult females(ManneWhitney, U¼ 5, nm¼ 6, nf¼ 6, p< 0.04), whereas there wasno sex difference in the frequency of tool use to crack low resistancenuts (ManneWhitney, U¼ 7, nm¼ 6, nf¼ 6, p¼ 0.093).
Efficiency at cracking nuts
Table 2 reports the percentage of episodes resulting in success,number of strikes and duration of episodes for adult males and
females to crack low resistance and high resistance nuts. The mixedmodel regression analyses, reported in Table 3, showed that successin cracking open nuts is predicted by nut resistance (z¼�2.23,p< 0.05, N¼ 423) but not by individual body mass, sex, rank, andweight of the hammer stone. Success at cracking high resistancenuts was predicted by individual body mass (z¼ 2.59, p< 0.01,N¼ 110) and hammer weight (z¼ 2.68, p< 0.01, N¼ 110).
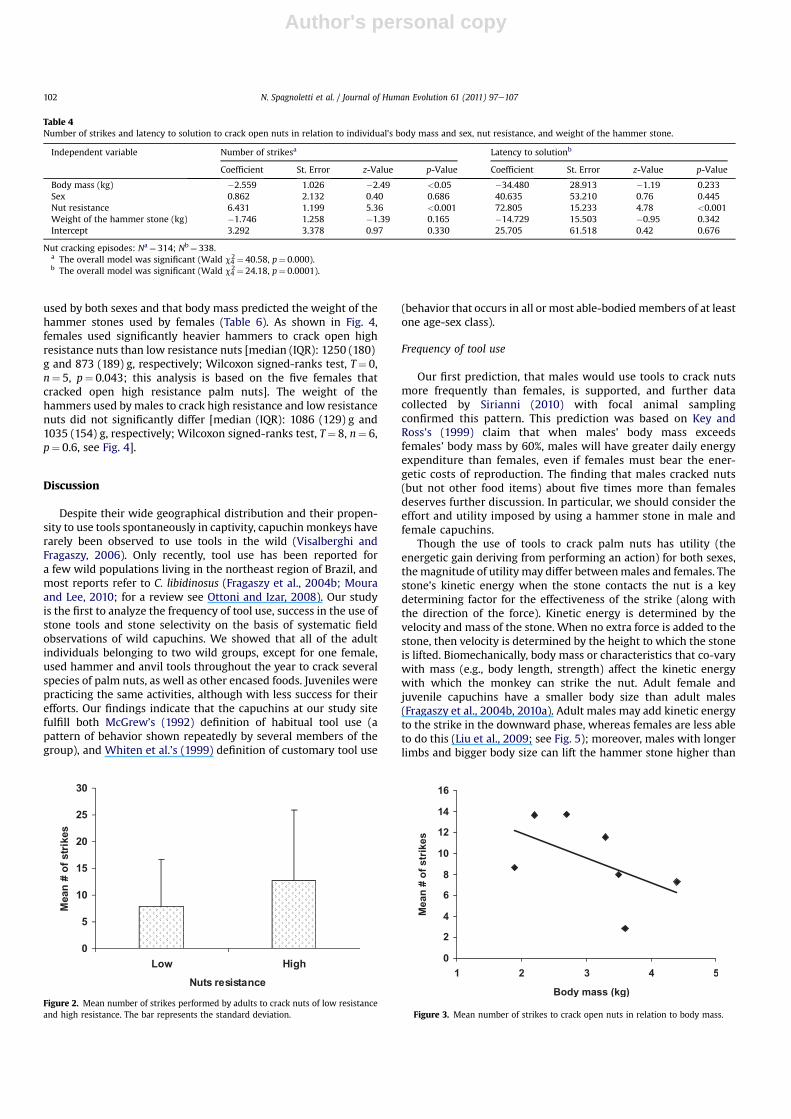
The number of strikes needed to open nuts varied significantly asa function of the individual bodymass and the resistance of the nut,but not to the sex of the individual or the weight of the hammerstone (Table 4; Figs. 2 and 3). The duration of episodes variedsignificantly as a function of the resistance of the nuts, but not to theindividual body mass, sex, and weight of the hammer stone(Table 4). The average duration (�standard deviation) per piassavanut cracking episode, calculated across 37 events for which data areavailable, is 185�159 s, with the maximum of about 13 minutes.
Selection of hammer stones by the two sexes
In most episodes, the monkeys used a hammer stone alreadypresent on the anvil when they arrived. In only 59 cases (3.4% of theepisodes), they transported the hammer to the anvil. To crack palmnuts, the monkeys used hammers with a median weight of aboutone kilogram [males, 100e3000 g (median (IQR)¼ 1059 (23) g);N¼ 373 nut cracking episodes; females 100e2250 g (median(IQR)¼ 991 (188) g); N¼ 133 nut cracking episodes]. In contrast, tocrack other food items, the monkeys used much lighter stones[males, 20e400 g (median (IQR)¼ 127 (64) g); N¼ 18 episodesperformed by five males; females, 40e650 g (median (IQR)¼ 148(154) g); N¼ 19 episodes performed by four females). The mixedmodel linear regression showed that the hardness and weight ofthe hammer stone used to crack open food were predicted by theresistance of the food item, but not by body mass and sex of theindividual (Table 5).
Themixedmodel linear regression split by sex indicated that theresistance of the food predicted the weight of the hammer stones
Figure 1. An adult male (Jatoba) using a log anvil (left) and a sandstone anvil (right) (Photos by E. Visalberghi).
Table 2Percentage of success, number of strikes and duration of the episode for adult malesand females to crack low resistance and high resistance nuts.
Low resistance nuts High resistance nuts
Males Females Males Females
Success % 93 87 90 62Strikes # 8� 1.4 13� 1.4 12� 1.4 15� 5.1Duration (s) 126� 19 146� 24 182� 20 194� 62N 190 68 100 29
N¼ number of episodes.
Table 3Success, measured as number of strikes needed to crack a nut, in relation to indi-vidual’s body mass, sex, and rank, nut resistance, and weight of the hammer stone(nut cracking episodes: N¼ 423).
Independent variable Coefficient St. Error z-value p-value
Body mass (kg) 0.094 0.595 0.16 0.875Sex 0.731 0.952 0.77 0.443Rank �0.314 0.205 �1.54 0.125Nut resistance �0.789 0.354 �2.23 < 0.05Weight of the hammer stone (kg) 0.478 0.387 1.24 0.216Intercept 2.96 1.302 2.27 0.023
The overall model was significant (Wald c52¼ 16, p¼ 0.0068).
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107 101
Author's personal copy
used by both sexes and that body mass predicted the weight of thehammer stones used by females (Table 6). As shown in Fig. 4,females used significantly heavier hammers to crack open highresistance nuts than low resistance nuts [median (IQR): 1250 (180)g and 873 (189) g, respectively; Wilcoxon signed-ranks test, T¼ 0,n¼ 5, p¼ 0.043; this analysis is based on the five females thatcracked open high resistance palm nuts]. The weight of thehammers used bymales to crack high resistance and low resistancenuts did not significantly differ [median (IQR): 1086 (129) g and1035 (154) g, respectively; Wilcoxon signed-ranks test, T¼ 8, n¼ 6,p¼ 0.6, see Fig. 4].
Discussion
Despite their wide geographical distribution and their propen-sity to use tools spontaneously in captivity, capuchin monkeys haverarely been observed to use tools in the wild (Visalberghi andFragaszy, 2006). Only recently, tool use has been reported fora few wild populations living in the northeast region of Brazil, andmost reports refer to C. libidinosus (Fragaszy et al., 2004b; Mouraand Lee, 2010; for a review see Ottoni and Izar, 2008). Our studyis the first to analyze the frequency of tool use, success in the use ofstone tools and stone selectivity on the basis of systematic fieldobservations of wild capuchins. We showed that all of the adultindividuals belonging to two wild groups, except for one female,used hammer and anvil tools throughout the year to crack severalspecies of palm nuts, as well as other encased foods. Juveniles werepracticing the same activities, although with less success for theirefforts. Our findings indicate that the capuchins at our study sitefulfill both McGrew’s (1992) definition of habitual tool use (apattern of behavior shown repeatedly by several members of thegroup), and Whiten et al.’s (1999) definition of customary tool use
(behavior that occurs in all or most able-bodiedmembers of at leastone age-sex class).
Frequency of tool use
Our first prediction, that males would use tools to crack nutsmore frequently than females, is supported, and further datacollected by Sirianni (2010) with focal animal samplingconfirmed this pattern. This prediction was based on Key andRoss’s (1999) claim that when males’ body mass exceedsfemales’ body mass by 60%, males will have greater daily energyexpenditure than females, even if females must bear the ener-getic costs of reproduction. The finding that males cracked nuts(but not other food items) about five times more than femalesdeserves further discussion. In particular, we should consider theeffort and utility imposed by using a hammer stone in male andfemale capuchins.
Though the use of tools to crack palm nuts has utility (theenergetic gain deriving from performing an action) for both sexes,themagnitude of utility may differ betweenmales and females. Thestone’s kinetic energy when the stone contacts the nut is a keydetermining factor for the effectiveness of the strike (along withthe direction of the force). Kinetic energy is determined by thevelocity and mass of the stone. When no extra force is added to thestone, then velocity is determined by the height to which the stoneis lifted. Biomechanically, body mass or characteristics that co-varywith mass (e.g., body length, strength) affect the kinetic energywith which the monkey can strike the nut. Adult female andjuvenile capuchins have a smaller body size than adult males(Fragaszy et al., 2004b, 2010a). Adult males may add kinetic energyto the strike in the downward phase, whereas females are less ableto do this (Liu et al., 2009; see Fig. 5); moreover, males with longerlimbs and bigger body size can lift the hammer stone higher than
Table 4Number of strikes and latency to solution to crack open nuts in relation to individual’s body mass and sex, nut resistance, and weight of the hammer stone.
Independent variable Number of strikesa Latency to solutionb
Coefficient St. Error z-Value p-Value Coefficient St. Error z-Value p-Value
Body mass (kg) �2.559 1.026 �2.49 <0.05 �34.480 28.913 �1.19 0.233Sex 0.862 2.132 0.40 0.686 40.635 53.210 0.76 0.445Nut resistance 6.431 1.199 5.36 <0.001 72.805 15.233 4.78 <0.001Weight of the hammer stone (kg) �1.746 1.258 �1.39 0.165 �14.729 15.503 �0.95 0.342Intercept 3.292 3.378 0.97 0.330 25.705 61.518 0.42 0.676
Nut cracking episodes: Na¼ 314; Nb¼ 338.a The overall model was significant (Wald c4
2¼ 40.58, p¼ 0.000).b The overall model was significant (Wald c4
2¼ 24.18, p¼ 0.0001).
Figure 2. Mean number of strikes performed by adults to crack nuts of low resistanceand high resistance. The bar represents the standard deviation. Figure 3. Mean number of strikes to crack open nuts in relation to body mass.
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107102

Author's personal copy
females can. Because males can add extra force to the stone in thedownward direction and can lift the stone higher, they can producemore effective strikes. Thus it is likely that the utility of crackingpalm nuts with tools differs in the two sexes, being more advan-tageous for adult males than adult females. Analyses nowunderway of the relation between the work (in its technical senseas the kinetic energy added) added to the stone by individual malesand females and the relative effectiveness of their strikes seems tosupport this view (Liu et al., unpublished results). Therefore,differential utilities of nut cracking in the two sexes may well beanother important factor contributing to the higher frequency withwhich wild capuchin males use hammer stones to crack nuts.
Capuchins present a different pattern of sex differences inpercussive tool use than do chimpanzees, which may well berelated to the utility of nut cracking inmales and females of the twospecies. Adult male bearded capuchins in our study groups weigh64% more than adult females (3.5 kg vs. 2.1 kg; although publisheddata indicate a lesser degree of dimorphism (34.8%) in wildC. apella, Ford and Davis, 1992). To crack open nuts, adult femalecapuchins at FBV used stones with a median weight of 991 g (i.e.,44e52% of their body mass, depending on the individual), whereasadult males used stones with a median weight of 1059 g (i.e.,24e40% of their body mass, depending on the individual).However, when experimentally provided with a 3.5 kg stone, bothsexes can use it successfully (Liu, unpublished results). Body massestimates for wild populations of Pan troglodytes verus that usetools to crack nuts are lacking. However, data obtained from wildspecimens present in museums indicate that females of thissubspecies have a median weight of 41.6 kg (N¼ 3) and the onlymale specimen has a weight of 46.3 kg (Smith and Jungers, 1997).Wild female Pan troglodytes schweinfurthii have somewhat smallerbodymass thanmales (Gombe:medianweight of 31.3 kg for femaleand 39 kg for males, Pusey et al., 2005; Mahale: median weight of35 kg for females and 40 kg for males, Uehara and Nishida, 1987),but in all populations the sex difference is proportionally muchsmaller than for our population of capuchins. The heaviest per-cussor used by adult female chimpanzees to crack Panda oleosa nutsweighed 12 kg (Boesch and Boesch, 1984), i.e., less than 29% of theweight reported for wild Pan troglodytes schweinfurthii females.Although an accurate comparison between the effort made by
C. libidinosus and by Pan troglodytes verus to crack nuts should scalebody mass to the 0.75 power (following Kleiber, 1961) and requiresa better dataset for the latter species for body mass and the averageweight of the hammers used by each sex, it still seems likely thatcracking nuts requires more effort for female capuchins at FBV thanfemale chimpanzees from Taï (for an estimate of nut cracking in onemale chimpanzee, see Günther and Boesch, 1993).
A further indication that nut cracking at FBV is more effortful forcapuchins than for chimpanzees at Taï and Bossou comes from thefact that chimpanzees adopt a seated posture to crack nuts andmostly use one hand (Matsuzawa, 1994; Boesch and Boesch-Achermann, 2000), whereas capuchin monkeys adopt a bipedalposture and use both hands (Liu et al., 2009). To crack seeds softerthan palm nuts, capuchinsmay use a small/light stone and hold it inone hand; this occurs both at our site (Spagnoletti, 2009) and in theSerra da Capivara National Park, a site located at a distance of460 km from FBV (Falótico, personal communication). Therefore,the differential frequencies of nut cracking in male and femalecapuchins and chimpanzees can be accounted for by the degree ofsexual dimorphism in body mass, as well as by the differentialutilities of nut cracking (higher for male capuchins than for femalecapuchins; lower for male chimpanzees than for femalechimpanzees).
Differential utilities of cracking account also for the differentpattern with which capuchins of both sexes exploited encasedfoods. As we predicted, females used tools to crack high resistancenuts significantly less often than males, whereas no sex differencewas found for frequencies of cracking low resistance nuts and otherencased foods. Females maximize the reliability of energetic returnby cracking foods that they were virtually certain to be able tocrack.
However, three other factors might also have contributed to sexdifferences in frequency of tool use: male capuchins might havebetter access to anvil site due to their rank, and/or females mighthave a lower propensity to engage in risky activities (nut cracking isnoisy, occurs on the ground and is easy to locate by predators)(Fragaszy, 1986; Rose, 1994), and/or males possess a higherpropensity to perform pounding behaviors than females (asdescribed by Visalberghi, 1988). Let us examine whether contri-butions by these three factors are supported.
Table 5Hardness and weight of the hammer stone used by adults to crack open all food items in relation to individual’s body mass and sex, and food resistance.
Independent variable Hammer stone hardnessa Hammer stone weightb
Coefficient St. Error z-Value p-Value Coefficient St. Error z-Value p-Value
Body mass (kg) �0.026 0.974 �0.27 0.788 14.516 40.005 0.36 0.717Sex �0.229 0.171 �1.34 0.181 138.118 78.928 1.75 0.08Food resistance �0.218 0.030 �7.08 <0.001 332.434 34.645 9.60 <0.001Intercept 2.130 0.175 12.18 0.000 �53.631 96.934 �0.55 0.58
Nut cracking episodes: Na¼ 493; Nb¼ 494.a The overall model was significant (Wald c3
2¼ 62.24, p¼ 0.000).b The overall model was significant (Wald c3
2¼ 125.89, p¼ 0.000).
Table 6Weight of the hammer stone used by adult females and adult males to crack open all food items in relation to individual’s body mass, and food resistance.
Independent variable Adult femalesa Adult malesb
Coefficient St. Error z-Value p-Value Coefficient St. Error z-Value p-Value
Body mass (kg) 681.246 246.332 2.77 <0.05 �1.586 39.806 �0.04 0.968Food resistance 387.587 59.65 6.48 <0.001 295.912 42.136 7.02 <0.001Intercept �1407.314 514.673 �2.73 0.006 362.921 168.887 2.15 0.032
Nut cracking episodes: Na¼ 160; Nb¼ 494.a The overall model was significant (Wald c2
2¼ 54.02, p¼ 0.000).b The overall model was significant (Wald c2
2¼ 49.71, p¼ 0.000).
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107 103

Author's personal copy
In FBV, hammers suitable to crack nuts are rare in the landscape(Visalberghi et al., 2009a) and an anvil site usually has only onehammer stone (Visalberghi et al., 2007). Thus, the anvil sites area valuable and limited resource, which might elicit both repeateduse and competition among tool users. The first point is supportedby our observations showing that (a) the episodes of nut cracking(N¼ 1565) occurred in only 122 different anvils (distributed over anarea of 9 km2), and (b) nearly all of the episodes occurred in anvilswhere the hammer stone was already present (Spagnoletti et al.,2010). Competition also occurred since typically an anvil siteaccommodates use by one individual at a time. Usually, while thenut cracker is at work, the other individuals “wait” nearby for theirturn to use the anvil site. Although infants and juveniles aregenerally tolerated by tool users and often exploit the bits of nutsthat become available on the anvil left from others’ cracking(Visalberghi and Fragaszy, in press-b), adults are less tolerated. Onlyhigh-ranking individuals may try to displace the nut cracker. Thenut cracker seems to signal its “possession” of the hammer (seeKummer and Cords, 1991) by maintaining contact (sometime witha foot) with it during the whole process of extraction. This isparticularly evident with piassava nuts that have several kernelsinside and need repeated bouts of handling with teeth and handsbetween strikes. The exploitation of these nuts lasts severalminutes.
Do the above results imply that rank affects an individual’slikelihood to use tools leading to the observed sex difference in
frequency of tool use? Although in both social groups living in FBVthe frequency of agonistic interactions during nut cracking is higherthan in other feeding contexts (Verderane, 2010), our findingsindicate that social rank and number of tool use episodes are notcorrelated. One dominant male, Zangado, was the most active tooluser of his group, whereas the other dominant male, Chicão, wasnot. Both the third and the fourth ranking males used tools moreoften than Chicão (see Table 1). Thoughmore difficult to investigategiven the small number of subjects, sex did not seem to affectfrequency of tool use in similarly ranking individuals. However, totest whether access to the anvil site, and therefore likelihood to usetools, differs according to an individual’s rank and sex, we need toexamine situations inwhich several tool users (each carrying a nut)are close to the same anvil site with one hammer. This remains tobe done.
Though our study did not assess the propensity to engage inrisky activities, studies exploring the degree of vigilance in the twosexes at FBVmight be relevant to this issue. If females are less pronethanmales to engage in risky activities that might attract predators,such as nut cracking (that predators can easily detect by sound andthat occurs on the ground fromwhere it is difficult to escape), thenthey should be particularly vigilant when engaging in this activity.For a small sample of FBV capuchins (two adult males and two adultfemales), vigilance during nut cracking did not differ between sexes(Cisco, 2006; Liu, unpublished results) and the analysis of activitybudget revealed that in both groups males were more attentive toexternal events (i.e., more likely to give alarm vocalizations) thanfemales (Verderane, 2010). Finally, females are as likely as males tobe on the ground (Spagnoletti et al., 2009). Therefore, females donot seem to be less active nutcrackers as a consequence of higherrisk avoidance. Future studies should address this issue by inves-tigating vigilance in the two sexes during different activities andwhen the monkeys are using different substrates.
Finally, it can be argued that the baseline rate of poundingobjects predicts individual differences in the rate of using tools.Capuchins bring an object into contact with a surface usingpercussive actions within the first year of life and these actions arean indispensable element in the development of nut cracking(Resende et al., 2008). If this is so, and if males are more likely thanfemales to pound objects on surfaces, males would also be morelikely to use percussive tools. In some studies with captive tuftedcapuchins (C. apella spp.), males were more inclined to engage inpounding behavior than females (as described by Visalberghi,1988). Other studies indicate no sex differences in such activities
Figure 4. Average weight of the hammer stones used by females and males whencracking nuts of high resistance and low resistance.
Figure 5. An adult male, 4.2 kg (a) and an adult female, 2.2 kg; (b) crack open a piassava nut using a hammer stone weighing 3.5 kg on a log anvil. This stone is much heavier thanthe average hammer stone used by capuchins at FBV (1.1 kg). The male lifts the stone higher than the female. (Photo by E. Visalberghi).
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107104

Author's personal copy
in captive capuchins (Fragaszy and Adams-Curtis, 1991; Byrne andSuomi, 1996). In any case, we cannot examine this predictionbecause we do not yet have data on the rate of pounding food itemson substrates by the capuchins in FBV. Future studies shouldinvestigate whether an individual’s relative frequency of poundingbehavior in non-tool-using contexts predicts its relative frequencyof tool use.
In sum, since social rank and vigilance can be ruled out, and dataare not yet available to examine the hypothesis linking generalpounding to use of tools, the differential utility of cracking for thetwo sexes deriving from the dimorphism in body size, seems themost plausible explanation for the higher frequency of nut crackingepisodes by male capuchins. If this explanation is correct, then weshould see the same pattern in other populations of beardedcapuchins that use percussive tools, assuming equivalent degrees ofsexual dimorphism and effort to extract the food. In the Serra daCapivara National Park (Piauí), a few hundred kilometers from ourstudy site, wild capuchins use stones to crack fruit and seeds muchless resistant than FBV nuts, and they extract tubers from theground using stones as scraping or excavation tools, in addition tousing sticks as probing tools and to flush prey (Falótico and Ottoni,2009; Mannu and Ottoni, 2009; Moura and Lee, 2010). Moura andLee (2010) report that at this site, male bearded capuchins use toolssignificantly more frequently than females, but due to the limitednumber of cases where the sex of the individual is reported (tool-using episodes N¼ 173, episodes pounding with stone N¼ 26),strong conclusions require replication of this finding. Apparentlythe sexes do not differ in the weight of the stone hammers chosen,nor in their efficiency (Falótico and Ottoni, unpublished results).
Efficiency at cracking nuts
In an experiment carried out in FBV in which 11 capuchinscracked whole and partially opened piassava (hard resistance nuts)with one stone, body mass was the single best predictor of effi-ciency (Fragaszy et al., 2010a). The present study, evaluating effi-ciency at cracking several species of nuts with a variety of stones,provides additional insights. Our second prediction, that heavierindividuals would be more efficient, is supported. As in Fragaszyet al. (2010a), larger individuals were more efficient (with allnuts, regardless of resistance), and they also enjoyed a higher rateof success in cracking high resistant nuts. However, body mass didnot affect the duration of cracking episodes leading to crackinga nut. Apparently the duration of the episodes reflects parametersother than efficiency.
Capuchins were successful in 84% of the episodes with highresistance palm nuts (piassava), 91% of the episodes with lowresistance palm nuts (catulè and tucum), and 99% of the episodesinvolving other encased foods of very low resistance (e.g., caju). Thepiassava nuts have a peak-force-at-failure similar to that reportedfor Panda nuts, the more resistant nuts exploited by the Taï chim-panzees (Boesch and Boesch, 1983). High resistance nuts havea peak-force-at-failure, which is on average 50% higher than lowresistance nuts. Our third prediction, that nut resistance wouldaffect efficiency, was supported. Nut resistance significantlyaffected the proportion of nuts attempted that were cracked, aswell as the number of strikes to crack palm nuts, and duration ofthe episodes. We also found that monkeys were more successfulwhen they used heavier hammers.
Overall, we found that sex did not significantly affect efficiency.This contrasts with what has been described for chimpanzeeswhere females not only perform stone tool use more, but alsooutperform males in the two most difficult forms of stone tool usethat chimpanzees practice (Boesch and Boesch, 1984). This iscertainly due to the different demands of nut cracking for the two
species; cracking nuts is proportionally more strenuous for capu-chins than for chimpanzees (see above) and therefore body size andsometimes hammer weight are crucial factors whereas sex per se isnot.
Tool selectivity
Our sixth predictionwas that capuchins would select stone toolsof suitable material and weight to obtain the food resource at hand.Two aspects of our findings support this prediction. First, monkeysof both sexes used heavier stones to crack nuts than to crack otherencased food items. Second, females (but not males) used heavierstones when cracking high resistance nuts than when cracking lowresistance nuts. The latter finding may indicate that females crackhigh resistance nuts only when the stone is heavy enough to allowsuccess, and/or that only females with heavy bodymass try to crackhigh resistance nuts, since in our sample only larger females (ofabout 2 kg) cracked high resistance nuts. In any case, femalecapuchins seem to take into account both the resistance of the nutand the weight of the stonewhen they decide whether or not to tryto crack a nut. Occasionally, we saw a female arriving with a highlyresistant nut to an anvil site with a light stone and leaving soonafterwards after making only a few strikes.
Because capuchins tend to repeatedly use anvil sites containingstone tools, use of appropriate stones could simply be thebyproduct of using the anvil sites where an appropriate hammer isalready present. However, this hypothesis can be rejected.Visalberghi et al. (2009b) experimentally demonstrated thatcapuchins reliably choose hammer stones that are heavier and lessfriable when given a choice between two or more stones, andFragaszy et al. (2010b) showed that the monkeys selected heavierstones to crack piassava nuts even when the difference in weightbetween the two stones varied in a ratio of 1.35:1. The presentstudy shows differential use of heavier stones to crack nuts versusother encased food and that females use heavier stones to crackmore resistant nuts.
Conclusions
Our findings demonstrate that bearded capuchins at FBV arehabitual tool users and that they select a stone by weight to use asa precursor on the basis of the resistance of the encased food. Thesefindings complement other findings from our team at this site.Capuchins use anvil sites repeatedly, they bring stones to anvils,they discriminate among nuts, anvil sites and stones, selectingspecific combinations that suit current needs, and they positionnuts systematically (Visalberghi et al., 2007, 2009a; Fragaszy et al.,2010b,c). This set of skills is unmatched by other monkey speciesthat have occasionally been reported to use tools (although crab-eating macaques are also promising in this respect, Gumert et al.,2009), and fully comparable with what has been reported forchimpanzees concerning nut cracking. Phylogeny does not predictthis pattern and therefore, nut cracking in capuchins likely aroseindependently of similar behaviors in Hominoidea.
Nut cracking in capuchins offers a valuable opportunity toconsider the behavioral and ecological correlates of stone tool use(Haslam et al., 2009). Further studies on the ecology of chimpanzeeand capuchin populations that use stones as tools with those thatdo not, though living in habitats in which encased foods and per-cussors are available, might clarify the ecological factors andbehavioral factors that contributed to the discovery of percussivetool use in these species, and possibly in the human lineage. In thecase of bearded capuchins, percussive tool use is not restricted topopulations in marginal habitats or to periods of low food avail-ability, as the behavior is present among bearded capuchins across
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107 105

Author's personal copy
their range (Ottoni and Izar, 2008) and occurs throughout the yearat consistent rates in the capuchins at FBV (Spagnoletti et al., 2008;Spagnoletti, 2009; Verderane, 2010; this study). Thus, the data callfor alternative models incorporating, for example, dietary diversityand morphological characteristics (Wright et al., 2009).
Because hominins likely used stone hammers to crack openencased foods before they used stones to flake (even with thesimple techniques evident in the Oldowan material), livingnonhuman primates have been used as models for early hominins.In particular, female supremacy in tool use in chimpanzees hasinspired numerous scenarios of human evolution and of division oflabor between the sexes (e.g., Isaac, 1978; Kuhn and Stiner, 2006).Our study suggests a more nuanced situation in capuchins ratherthan a simple sex difference. Adult male capuchins use stone toolsto crack nuts more frequently than adult female capuchins, butbody size is a better predictor of efficiency of tool use than sex.Therefore, we suggest that the physical requirement of lifting heavystones to crack nuts favors heavier individuals, who are for themostpart, males.
Sexual dimorphism is most probably the primitive conditionretained by humans from the common ancestors of living Africanapes and humans (Lockwood et al., 1996). In Australopithecus afar-ensis and Australopithecus africanus, males seem to be almostdouble the size of females (McHenry, 1992; McHenry and Berger,1998; though Reno et al., 2003 consider skeletal size dimorphismin A. afarensis similar to that of contemporary Homo sapiens).Dimorphism in body size, and consequently muscular mass andstrength, is more marked in our wild capuchin population than inhumans and in chimpanzees (Fleagle,1999; Aiello andWells, 2002).Capuchins illustrate how tool use practices can be impacted bysexual dimorphism in body size, thus contributing to a compre-hensive scenario for the evolution of tool-using skills in the humanlineage.
Acknowledgements
Permission to work in Brazil was granted by IBAMA and CNPq toDF and EV. Thanks to Fundação BioBrasil and to the Familia M forpermission to work at FBV and Jozemar, Arizomar and Junior fortheir assistance in the field; Eduardo D. Ramos da Silva andMicheleP. Verderane for the data and Gabriele Schino for the support instatistical analyses; Qing Liu, Elsa Addessi, Valentina Truppa andGloria Sabbatini for comments on this manuscript. We also thankDavid Begun, Editor of the JHE, Bill McGrew, and the other anony-mous referee. This research was funded by the National GeographicSociety, Leakey Foundation, CNPq, FAPESP, CNR, EU-Analogy (STREPContr. No 029088), La Sapienza University of Rome, Ethoikos srl,European project IM-CLeVeR FP7-ICT-IP-231722.
References
Aiello, L.C., Wells, J.C.K., 2002. Energetics and the evolution of the genus Homo.Annu. Rev. Anthropol. 31, 323e338.
Boesch, C., Boesch, H., 1983. Optimization of nut-cracking with natural hammers bywild chimpanzees. Behaviour 83, 265e286.
Boesch, C., Boesch, H., 1984. Possible causes of sex differences in the use of naturalhammers by wild chimpanzees. J. Hum. Evol. 13, 415e440.
Boesch, C.H., Boesch-Achermann, H., 2000. The Chimpanzees of the Taï Forest.Oxford University Press, Oxford.
Boinski, S., 2004. The beats of different drummers: percussion as sexual signalingamong brown capuchins at Raleighvallen, Suriname. Folia Primatol. 75, 238.
Boinski, S., Quatrone, R.P., Swartz, H., 2000. Substrate and tool use by browncapuchins in Suriname: ecological contexts and cognitive bases. Am. Anthropol.102, 741e761.
Byrne, G., Suomi, S.J., 1996. Individual differences in object manipulation in a colonyof tufted capuchins. J. Hum. Evol. 31, 259e267.
Canale, G.R., Guidorizzi, C.E., Kierulff, M.C.M., Gatto, C., 2009. First record of tool useby wild populations of the yellow-breasted capuchin monkey (Cebus
xanthosternos) and new records for the bearded capuchin (Cebus libidinosus).Am. J. Primatol. 71, 1e7.
Cisco, E., 2006. Analisi del comportamento di uso di strumenti per la rottura di nociin una popolazione di Cebus libidinosus in natura. Facoltà di Scienze matema-tiche, fisiche e naturali. Università “La Sapienza” di Roma.
Davidson, I., McGrew, W.C., 2005. Stone tools and the uniqueness of human culture.J. Roy. Anthropol. Inst. 11, 793e817.
Falótico, T., Ottoni, E.B., 2009. Capuchin monkeys of caatinga: many tools for manythings. Abstracts of the 46th Annual Meeting of the Animal Behavior Society.Pirenópolis, GO, p. 72.
Fleagle, J.G., 1999. Primate Adaptation and Evolution, second ed. Academic Press,San Diego.
Ford, S.M., Davis, L., 1992. Systematics and body size: implications for feedingadaptations in New World monkeys. Am. J. Phys. Anthropol. 88, 415e468.
Fragaszy, D.M., 1986. Time budgets and foraging behavior in wedge-capped capu-chins (Cebus olivaceus): age and sex differences. In: Taub, D.M., King, F.A. (Eds.),Current Perspectives in Primate Social Dynamics. Van Nostrand Reinhold, NewYork, pp. 159e174.
Fragaszy, D.M., 1990. Sex and age differences in the organization of behaviour inwedge-capped capuchins. Behav. Ecol. 1, 81e94.
Fragaszy, D.M., Adams-Curtis, L.E., 1991. Generative aspects of manipulation intufted capuchin monkeys (Cebus apella). J. Comp. Psychol. 105, 387e397.
Fragaszy, D.M., Izar, P., Visalberghi, E., Ottoni, E.B., Oliveira, M.G., 2004a. Wildcapuchin monkeys (Cebus libidinosus) use anvils and stone pounding tools. Am.J. Primatol. 64, 359e366.
Fragaszy, D.M., Fedigan, L., Visalberghi, E., 2004b. The Complete Capuchin: theBiology of the Genus Cebus. Cambridge University Press, Cambridge.
Fragaszy, D.M., Pickering, T., Liu, Q., Izar, P., Ottoni, E., Visalberghi, E., 2010a. Beardedcapuchin monkeys’ and a human’s efficiency at cracking palm nuts with stonetools: field experiments. Anim. Behav. 79, 321e332.
Fragaszy, D.M., Greenberg, R., Visalberghi, E., Ottoni, E., Izar, P., Liu, Q., 2010b. Howwild bearded capuchin monkeys select stones and nuts to minimize thenumber of strikes per nut cracked. Anim. Behav. 80, 205e214.
Fragaszy, D.M., Allen, A., Wright, B., 2010c. Wild bearded capuchin monkeys (Piauí,Brazil) position nuts systematically into pits on anvils before striking the nutswith stone tools. Am. J. Primatol. 72 (Suppl.), 54e55.
Gumert, M.D., Kluck, M., Malaivijitnond, S., 2009. The physical characteristics andusage patterns of stone axe and pounding hammers used by long-tailedmacaques in the Andaman Sea region of Thailand. Am. J. Primatol. 71, 594e608.
Günther, M.M., Boesch, C., 1993. Energetic cost of nut-cracking behaviour in wildchimpanzees. In: Preuschoft, H., Chivers, D.J. (Eds.), Hands of Primates. Springer,New York, pp. 109e129.
Haslam, M., Hernandez-Aguilar, A., Ling, V., Carvalho, S., de la Torre, I., DeStefano, A., Du, A., Foley, R., Hardy, B., Harris, J., Marchant, L., Matsuzawa, T.,McGrew, W., Mercader, J., Mora, R., Petraglia, M., Roche, H., Visalberghi, E.,Warren, R., 2009. Primate archaeology. Nature 460, 339e344.
Isaac, G., 1978. The food-sharing behavior of protohuman hominids. Sci. Am. 238,90e108.
Izar, P., Ferreira, R.G., Sato, T., 2006. Describing the organization of dominancerelationships by dominance directed tree method. Am. J. Primatol. 68, 189e207.
Key, C., Ross, C., 1999. Sex differences in energy expenditure in non-humanprimates. Proc. R. Soc. Lond. B 266, 2479e2485.
Kleiber, M., 1961. The Fire of Life. Wiley, New York.Kuhn, S.L., Stiner, M.C., 2006. What’s a mother to do: a hypothesis about the division
of labor among Neandertals and modern humans. Curr. Anthropol. 47,953e980.
Kummer, H., Cords, M., 1991. Cues of ownership in long-tailed macaques, Macacafascicularis. Anim. Behav. 42, 529e549.
Liu, Q., Simpson, K., Izar, P., Ottoni, E., Visalberghi, E., Fragaszy, E., 2009. Kinematicsand energetics of nut-cracking in wild capuchin monkeys (Cebus libidinosus) inPiauí, Brazil. Am. J. Phys. Anthropol. 138, 210e220.
Lockwood, C.A., Richmond, B.G., Jungers, W.L., Kimbel, W.H., 1996. Randomizationprocedures and sexual dimorphism in Australopithecus afarensis. J. Hum. Evol.31, 537e548.
Lonsdorf, E.V., Pusey, A.E., Eberly, L., 2004. Sex differences in learning in chim-panzees. Nature 428, 715e716.
Mannu, M., Ottoni, E.B., 2009. The enhanced tool-kit of two groups of wild beardedcapuchin monkeys in the caatinga: tool making, associative use, and secondarytools. Am. J. Primatol. 71, 242e251.
Martin, P., Bateson, P., 1993. Measuring Behaviour: an Introductory Guide, seconded. Cambridge University Press, Cambridge.
Matsuzawa, T., 1994. Field experiments on use of stone tools by chimpanzees in thewild. In: Wrangham, R.W., McGrew, W.C., de Waal, F.B.M., Heltne, P.G. (Eds.),Chimpanzee Cultures. Harvard University Press, Cambridge, pp. 351e370.
McGrew, W.C., 1979. Evolutionary implications of sex differences in chimpanzeepredation and tool use. In: Hamburg, D.A., McCown, E. (Eds.), The Great Apes.Benjamin/Cummings, Menlo Park, pp. 441e463.
McGrew, W.C., 1992. Chimpanzee Material Culture. Cambridge University Press,Cambridge.
McHenry, H.M., 1992. Body size and proportions in early hominids. Am. J. Phys.Anthropol. 87, 407e431.
McHenry, H.M., Berger, L.R., 1998. Body proportions in Australopithecus afarensis andA. africanus and the origin of the genus Homo. J. Hum. Evol. 35, 1e22.
Moura, A., 2007. Stone banging by wild capuchin monkeys: an unusual auditorydisplay. Folia Primatol. 78, 36e45.
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107106

Author's personal copy
Moura, A., Lee, P., 2004. Capuchin stone tool use in caatinga dry forest. Science 306,1909e1910.
Moura, A., Lee, P., 2010. Wild capuchins show male-biased feeding tool use. Int. J.Primatol. 31, 457e470.
Ottoni, E.B., Izar, P., 2008. Capuchin monkey tool use: overview and implication.Evol. Anthropol. 17, 171e178.
Ottoni, E.B., Mannu, M., 2001. Semi free-ranging tufted capuchins (Cebus apella)spontaneously use tools to crack open nuts. Int. J. Primatol. 22, 347e358.
Pandolfi, S.S., van Schaik, C.P., Pusey, A.E., 2003. Sex differences in termite fishingamong Gombe chimpanzees. In: de Waal, F.B.M., Tyack, P.L. (Eds.), Animal SocialComplexity: Intelligence, Culture and Individualized Societies. HarvardUniversity Press, Cambridge, pp. 414e418.
Plavcan, J.M., 2004. Sexual selection, measures of sexual selection, and sexualdimorphism in primates. In: Kappeler, P.M., van Schaik, C.P. (Eds.), SexualSelection in Primates, New and Comparative Perspectives. Cambridge Univer-sity Press, Cambridge, pp. 230e252.
Pusey, A.E., Oehlert, G.W., Williams, J.M., Goodall, J., 2005. The influence ofecological and social factors on body mass of wild chimpanzees. Int. J. Primatol.26, 3e31.
Rabe-Hesketh, S., Skrondal, A., 2008. Multilevel and Longitudinal Modeling UsingStata, second ed. Stata Press, College Station.
Ramos da Silva, E.D., 2008. Escolha de alvos coespecificos na observação do uso deferramenta por macaco-prego (Cebus libidinosus) selvagens. Tese de Mestradoem Psicologia Experimental, Universidade de São Paulo.
Reno, P.L., Meindl, R.S., McCollum, M.A., Lovejoy, C.O., 2003. Sexual dimorphism inAustralopithecus afarensis was similar to that of modern humans. Proc. Nat.Acad. Sci. 100, 9404e9409.
Resende, B.D., Ottoni, E.B., Fragaszy, D.M., 2008. Ontogeny of manipulative behaviorand nut-cracking in young tufted capuchin monkeys Cebus apella: a perception-action perspective. Develop. Sci. 11, 828e840.
Rose, L.M., 1994. Sex differences in diet and foraging behavior in white-facedcapuchins (Cebus capucinus). Int. J. Primatol. 15, 95e114.
Schrago, C.G., Russo, C.A., 2003. Timing the origin of NewWorld monkeys. Mol. Biol.Evol. 20, 1620e1625.
Sirianni, G., 2010. Processamento del cibo in una popolazione di Cebus libidinosus(Spix, 1823). Master’s Thesis, Università La Sapienza di Roma.
Smith, R.J., Jungers, W.L., 1997. Body mass in comparative primatology. J. Hum. Evol.32, 523e559.
Spagnoletti, N., 2009. Uso di strumenti in una popolazione di Cebus libidinosus allostato selvatico in Piauí, Brasile. Ph.D. Dissertation, Università La Sapienza di Roma.
Spagnoletti, N., Izar, P., Visalberghi, E., 2008. Effects of food availability on tool use ofwild bearded capuchin monkey (Cebus libidinosus). In: XXII Congress of theInternational Primatological Society (abstract).
Spagnoletti, N., Izar, P., Visalberghi, E., 2009. Tool use and terrestriality in wildbearded capuchin monkey (Cebus libidinosus). 3rd Congress of the EuropeanFederation for Primatology. Folia Primatol. 80, 106e174.
Spagnoletti, N., Visalberghi, E., Presotto, A., Izar, P., 2010. Spatial distribution ofresources for the occurrences of nut cracking with tools in wild bearded
capuchins (Cebus libidinosus). In: XXIII Congress of the International Primato-logical Society (abstract).
StataCorp, 2007. Stata Statistical Software: Release 10. Stata Press, College Station.Tooby, J., DeVore, I., 1987. The reconstruction of hominid behavioral evolution
through strategic modeling. In: Kinzey, W.G. (Ed.), The Evolution of HumanBehavior: Primate Models. SUNY Press, New York, pp. 183e237.
Uehara, S., Nishida, T., 1987. Body weights of wild chimpanzees (Pan troglodytesschweinfurthii) of the Mahale Mountains National Park. Am. J. Phys. Anthropol.72, 315e321.
Verderane, M.P., 2010. Socioecologia de macacos-prego (Cebus libidinosus) em áreade ecótono cerrado/caatinga. Ph.D. dissertation, Universidade de São Paulo.
Visalberghi, E., 1988. Responsiveness to objects in two social groups of tuftedcapuchin monkeys (Cebus apella). Am. J. Primatol. 15, 349e360.
Visalberghi, E., Fragaszy, D., 2006. What is challenging about tool use? The capu-chins’ perspective. In: Wasserman, E.A., Zentall, T.R. (Eds.), ComparativeCognition: Experimental Explorations of Animal Intelligence. Oxford UniversityPress, Oxford, pp. 529e552.
Visalberghi, E., Fragaszy, D. The EthoCebus project. Stone tool use by wild capuchinmonkeys. In: Sanz, C., Call, J., Boesch, C. (Eds.), Multidisciplinary Perspectives onthe Cognition and Ecology of Tool Using Behaviors. Cambridge University Press,Cambridge, in press-a.
Visalberghi, E., Fragaszy, D. Learning how to forage. Socially biased individuallearning and “niche construction” in wild capuchin monkeys. In: de Wall, F.,Ferrari, P.F. (eds.), The Primate Mind. Harvard University Press, in press-b.
Visalberghi, E., Fragaszy, D., Ottoni, E., Izar, P., de Oliveira, M.G., Andrade, F.R.D.,2007. Characteristics of hammer stones and anvils used by wild beardedcapuchin monkeys (Cebus libidinosus) to crack open palm nuts. Am. J. Phys.Anthropol. 132, 426e444.
Visalberghi, E., Sabbatini, G., Spagnoletti, N., Andrade, F.R.D., Ottoni, E., Izar, P.,Fragaszy, D., 2008. Physical properties of palm fruits processed with tools bywild bearded capuchins (Cebus libidinosus). Am. J. Primatol. 70, 884e891.
Visalberghi, E., Spagnoletti, N., Ramos da Silva, E.D., de Andrade, F.R., Ottoni, E.,Izar, P., Fragaszy, D., 2009a. Transport of hammer tools and nuts by wildcapuchin monkeys. Primates 50, 95e104.
Visalberghi, E., Addessi, E., Truppa, V., Spagnoletti, N., Ottoni, E., Izar, P., Fragaszy, D.,2009b. Selection of effective stone tools by wild capuchin monkeys. Curr. Biol.19, 213e217.
Waga, I.C., Dacier, A.K., Pinga, P.S., Tavares, M.C.H., 2006. Spontaneous tool use bywild capuchin monkeys (Cebus libidinosus) in the cerrado. Folia Primatol. 77,337e344.
Whiten, A., Goodall, J., McGrew, W.C., Nishida, T., Reynolds, V., Sugiyama, Y.,Tutin, C.E.G., Wrangham, R.W., Boesch, C., 1999. Cultures in chimpanzees.Nature 399, 682e685.
Wright, B.W., Wright, K.A., Chalk, J., Verderane, M.P., Fragaszy, D., Visalberghi, E.,Izar, P., Ottoni, E.B., Constantino, P., Vinyard, C., 2009. Fallback foraging as a wayof life: using dietary toughness to compare the fallback signal among capuchinsand implications for interpreting morphological variation. Am. J. Phys.Anthropol. 140, 687e699.
N. Spagnoletti et al. / Journal of Human Evolution 61 (2011) 97e107 107




















