Stimulation of Protein Synthesis by Both Insulin and Amino Acids Is Unique to Skeletal Muscle in...
39
R1-E-00517-2001 Stimulation of Protein Synthesis by Both Insulin and Amino Acids Is Unique to Skeletal Muscle in Neonatal Pigs TERESA A. DAVIS, MARTA L. FIOROTTO, DOUGLAS G. BURRIN, PETER J. REEDS, HANH V. NGUYEN, PHILIP R. BECKETT, RHONDA C. VANN, AND PAMELA M. J. O’CONNOR United States Department of Agriculture/Agricultural Research Service, Children's Nutrition Research Center, and Endocrinology and Metabolism Section, Department of Pediatrics, Baylor College of Medicine, Houston, TX 77030 Short title: Protein synthesis in neonates Correspondence and reprint requests: T. A. Davis, Ph.D. USDA/ARS Children's Nutrition Research Center Baylor College of Medicine 1100 Bates Street Houston, TX 77030 Phone: (713) 798-7169 Fax: (713) 798-7171 E-mail: [email protected] Copyright 2002 by the American Physiological Society. AJP-Endo Articles in PresS. Published on January 2, 2002 as DOI 10.1152/ajpendo.00517.2001
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Stimulation of Protein Synthesis by Both Insulin and Amino Acids Is Unique to Skeletal Muscle in...

R1-E-00517-2001
Stimulation of Protein Synthesis by Both Insulin and Amino
Acids Is Unique to Skeletal Muscle in Neonatal Pigs
TERESA A. DAVIS, MARTA L. FIOROTTO, DOUGLAS G. BURRIN, PETER J. REEDS,
HANH V. NGUYEN, PHILIP R. BECKETT, RHONDA C. VANN, AND PAMELA M. J.
O’CONNOR
United States Department of Agriculture/Agricultural Research Service, Children's Nutrition
Research Center, and Endocrinology and Metabolism Section, Department of Pediatrics, Baylor
College of Medicine, Houston, TX 77030
Short title: Protein synthesis in neonates
Correspondence and reprint requests:
T. A. Davis, Ph.D.
USDA/ARS Children's Nutrition Research Center
Baylor College of Medicine
1100 Bates Street
Houston, TX 77030
Phone: (713) 798-7169
Fax: (713) 798-7171
E-mail: [email protected]
Copyright 2002 by the American Physiological Society.
AJP-Endo Articles in PresS. Published on January 2, 2002 as DOI 10.1152/ajpendo.00517.2001

R1-E-00517-2001
ABSTRACT
In neonatal pigs, the feeding-induced stimulation of protein synthesis in skeletal muscle, but
not liver, can be reproduced by insulin infusion when essential amino acids and glucose are
maintained at fasting levels. In the current study, 7- and 26-day-old pigs were studied during: 1)
fasting, 2) hyperinsulinemic-euglycemic-euaminoacidemic clamps, 3) euinsulinemic-euglycemic-
hyperaminoacidemic clamps, and 4) hyperinsulinemic-euglycemic-hyperaminoacidemic clamps.
Amino acids were clamped using a new amino acid mixture enriched in nonessential amino acids.
Tissue protein synthesis was measured using a flooding dose of L-[4-3H]phenylalanine. In 7-day-old
pigs, insulin infusion alone increased protein synthesis in various skeletal muscles (from +35% to
+64%), with equivalent contribution of myofibrillar and sarcoplasmic proteins, as well as cardiac
muscle (+50%), skin (+34%), and spleen (+26%). Amino acid infusion alone increased protein
synthesis in skeletal muscles (from +28% to +50%), also with equivalent contribution of myofibrillar
and sarcoplasmic proteins, as well as liver (+27%), pancreas (+28%), and kidney (+10%). An
elevation of both insulin and amino acids did not have an additive effect. Similar qualitative results
were obtained in 26-day-old pigs, but the magnitude of the stimulation of protein synthesis by insulin
and/or amino acids was lower. The results suggest that, in the neonate, the stimulation of protein
synthesis by feeding is mediated by either amino acids or insulin in most tissues; however, the
feeding-induced stimulation of protein synthesis in skeletal muscle is uniquely regulated by both
insulin and amino acids.
Key Words: Neonate, insulin action, amino acids, protein synthesis, nutrition

R1-E-00517-2001 1
INTRODUCTION
During the neonatal period, the gain in protein mass in skeletal muscle is more rapid than in
other tissues of the body (63). Fractional rates of protein synthesis in skeletal muscle of the newborn
are high, and decline more rapidly than in other tissues of the body during the early postnatal period
(18, 12). This developmental decline in skeletal muscle protein synthesis varies with fiber type, and
is accompanied by a decrease in ribosome number and a specific accumulation of myofibrillar
proteins (18, 25).
The efficiency with which dietary amino acids are utilized for protein deposition is also high
in the neonate, and declines markedly with development (20, 40). This high efficiency is likely due
to the enhanced stimulation of protein synthesis in response to feeding (12, 15, 17, 19). The feeding-
induced stimulation of protein synthesis occurs in virtually all tissues of the neonate; however, the
postprandial rise in protein synthesis and the developmental decline in the response to feeding are
most pronounced in skeletal muscle, particularly in those muscles that are primarily fast-twitch,
glycolytic (8-10, 12, 15, 17, 19). The developmental decline in the response to feeding in muscle
parallels the marked fall in the nutrient-induced activation of translation initiation factors which
regulate the binding of mRNA to the 40S ribosomal subunit (21).
Insulin is likely a central regulatory factor in the elevated rate of muscle protein deposition
in the neonate. Results of studies, using the hyperinsulinemic-euglycemic-euaminoacidemic clamp
technique originally developed in our laboratory, show that insulin stimulates whole-body amino
acid disposal in the neonate, and that both the insulin sensitivity and responsiveness of amino acid
disposal are attenuated with the advancement of development (61). When insulin is infused at doses
reproducing plasma insulin levels observed under fed conditions and essential amino acids and

R1-E-00517-2001 2
glucose are clamped at fasting levels, skeletal muscle protein synthesis increases to values observed
in the fed state (62). The stimulation of protein synthesis by insulin, like the response to feeding,
is greater in muscles that are predominately fast-twitch, glycolytic, and is not specific to myofibrillar
proteins (14). This response to insulin decreases with development (14, 62), in parallel with the
developmental decline in the stimulation of muscle protein synthesis by feeding (12). Recent studies
suggest that the developmental decline in the response of skeletal muscle protein synthesis to food
consumption results from a reduction in the effectiveness of the insulin signaling pathway to
transduce to the translation apparatus the stimulus provided by feeding (36, 37, 53).
A recent study from this laboratory suggests that the feeding-induced stimulation of visceral
tissue protein synthesis in the neonate cannot be attributed to insulin (14). We found that the
infusion of insulin in the neonate stimulates protein synthesis in skeletal muscle, cardiac muscle, and
skin, but insulin did not stimulate, and in fact tended to lower, protein synthesis in liver, intestine,
pancreas, and kidney (14). The ability of amino acids to stimulate liver protein synthesis is
controversial, but limited studies suggest that amino acids increase protein synthesis in the liver of
immature (27, 51) but not mature (2, 44) animals. In contrast, amino acids appear to stimulate
protein synthesis in the skeletal muscle of young, adult, and elderly populations (6, 41, 49, 58, 59).
Therefore, we hypothesized that protein synthesis in all tissues of the neonate is responsive to the
postprandial rise in amino acid concentrations.
During the hyperinsulinemic-euglycemic-euaminoacidemic clamps in previous studies, we
infused Trophamine, an amino acid mixture designed for use in neonates (14, 61, 62). However, in
these studies, detailed examination of amino acid concentrations during “euaminoacidemic”
hyperinsulinemic euglycemic clamps showed that the circulating concentrations of some of the

R1-E-00517-2001 3
nonessential amino acids were lower at the highest insulin concentrations, which could potentially
limit the availability of substrate for protein synthesis. Therefore, we developed a new amino acid
clamp mixture designed specifically to keep the concentrations of all amino acids constant during
insulin stimulation. We determined, in 7- and 26-day-old pigs, the response of tissue protein
synthesis to: 1) the infusion of insulin at a dose that reproduces insulin levels in the fed state, while
circulating amino acids and glucose are maintained at fasting levels; 2) the infusion of amino acids
at a dose that reproduces fed-state amino acid levels, while insulin and glucose are kept at fasting
levels; and 3) the infusion of insulin and amino acids at doses that reproduce insulin and amino acid
levels in the fed state, while glucose is kept at fasting level.
METHODS
Animals and surgery. Multiparous crossbred (Yorkshire x Landrace x Hampshire x Duroc)
sows (n = 8; Agriculture Headquarters, Texas Department of Criminal Justice, Huntsville, TX) were
housed in lactation crates in individual, environmentally controlled rooms, and maintained on a
commercial diet (5084, PMI Feeds Inc., Richmond, IN) throughout a 28-day lactation period. Sows
delivered 8-12 piglets and these remained with the sow for nursing. Piglets were not given
supplemental creep feed. Piglets weighing 2.0 ± 0.1 kg were studied at 7 days of age (n = 35), and
piglets weighing 7.2 ± 0.1 kg were studied at 26 days of age (n = 39). Three to 5 days before the
insulin infusion study, pigs were anesthetized, and catheters were inserted surgically into a jugular
vein and a carotid artery (61). Piglets were returned to the sow until studied. The protocol was
approved by the Animal Care and Use Committee of Baylor College of Medicine. The study was
conducted in accordance with the National Research Council's Guide for the Care and Use of

R1-E-00517-2001 4
Laboratory Animals.
Infusions. Pigs were placed, unanesthetized, in a sling restraint system following a 12-h fast.
Pigs were assigned to one of four treatment groups: 1) fasting (Control); 2) hyperinsulinemic-
euglycemic-euaminoacidemic clamp (Insulin); 3) euinsulinemic-euglycemic-hyperaminoacidemic
clamp (AA); and 4) hyperinsulinemic-euglycemic-hyperaminoacidemic clamp (Insulin+AA). Body
weights were similar in each treatment group at each age studied. During a 30-min basal period,
blood samples were obtained and immediately analyzed for glucose (YSI 2300 STAT Plus, Yellow
Springs Instruments, Yellow Springs, OH) to establish the average basal concentration of blood
glucose to be used in the subsequent euglycemic clamp procedure (22). Plasma samples were
analyzed for total branched-chain amino acids (BCAA), using a rapid enzymatic kinetic assay (5)
to establish the average basal concentration of BCAA to be used in the subsequent euaminoacidemic
or hyperaminoacidemic clamp procedure. The clamps were initiated with a primed, constant (12
mL/h) infusion of insulin (Eli Lilly, Indianapolis, IN) at either 0 or 100 ng � kg-0.66 � min-1. Venous
blood samples (0.2 mL) were acquired every 5 min and immediately analyzed for glucose and BCAA
concentrations. The infusion rate of dextrose (Baxter Healthcare Corp., Deerfield, IL) was adjusted
as necessary to maintain the blood glucose concentration within ±10% of the average basal
concentration. Euaminoacidemia was obtained by adjusting the infusion rate of an amino acid
mixture to maintain the plasma BCAA concentration within 10% of the fasting level.
Hyperaminoacidemia was obtained by infusing an amino acid mixture to raise plasma BCAA
concentrations to twofold the fasting level, in order to reproduce the level of amino acids present in
the fed state (12).
A new amino acid mixture was developed for use as an infusate during the euaminoacidemic

R1-E-00517-2001 5
and hyperaminoacidemic clamp procedures. The composition of the new amino acid mixture was
based on the amino acid composition of body protein (20) and our previous observations on the
changes in plasma amino acid concentrations when TrophAmine 10% (McGaw, Irvine, CA) was
infused during hyperinsulinemic-euglycemic-euaminoacidemic clamp studies (14, 61, 62). The new
mixture utilized two dipeptides, glycyl-tyrosine and alanyl-glutamine, to avoid the problem of poor
solubility of tyrosine and instability of glutamine, and contained higher concentrations of
nonessential amino acids than commercial amino acid mixtures. The new amino acid mixture
contained (mmolar) arginine (20.1), histidine (12.9), isoleucine (28.6), leucine (34.3), lysine (27.4),
methionine (10.1), phenylalanine (12.1), threonine (21.0), tryptophan (4.4), valine (34.1), alanine
(27.3; 38% provided as alanyl-glutamine), aspartate (12.0), cysteine (6.2), glutamate (23.8),
glutamine (17.1; 100% provided as alanyl-glutamine), glycine (54.3; 4% provided as glycyl-
tyrosine), proline (34.8), serine (23.8), taurine (2.0), and tyrosine (7.2; 83% provided as glycyl-
tyrosine).
Tissue protein synthesis in vivo. Fractional rates of protein synthesis were measured with a
flooding dose of L-[4-3H]phenylalanine (29) injected 3.5 h after the initiation of the clamp. Blood
samples were taken at 5, 15, and 30 min after the injection for measurement of the specific
radioactivity of the extracellular free pool of phenylalanine. Pigs were killed at 4 h after the
initiation of the clamp. Samples of longissimus dorsi, gastrocnemius, masseter, and cardiac muscles,
skin, liver, jejunum, pancreas, kidney, and spleen were collected, rapidly frozen in liquid nitrogen,
and stored at -70oC.
Total protein was isolated from all tissues sampled, and sarcoplasmic and myofibrillar
proteins were isolated in longissimus dorsi muscle of 7-day-old pigs as described previously (18, 25).

R1-E-00517-2001 6
The specific radioactivities of the protein hydrolysates, homogenate supernatants, and blood
supernatants were determined as previously described (18).
Plasma insulin and amino acids. Plasma immunoreactive insulin concentrations were
measured using a porcine insulin radioimmunoassay kit (Linco, St. Louis, MO). Plasma amino acid
concentrations were measured with a high-performance liquid chromatography method (PICO-TAG
reverse-phase column, Waters, Milford, MA) as previously described (19).
Calculations and statistics. Fractional rates of protein synthesis (Ks; percent of protein mass
synthesized in a day) were calculated as:
Ks (%/day) = [(Sb/Sa) x (1440/t)] x 100
where Sb is the specific radioactivity of the protein-bound phenylalanine, Sa is the specific
radioactivity of the tissue-free phenylalanine for the labeling period determined from the value of
the animal at the time of the tissue collection, corrected by the linear regression of the blood specific
radioactivity of the animal against time, and t is the time of labeling in minutes. We have
demonstrated that, after a flooding dose of phenylalanine is administered, the specific radioactivity
of tissue free phenylalanine is in equilibrium with the amino-acyl tRNA specific radioactivity, and
therefore, the tissue free phenylalanine is a valid measure of the tissue precursor pool specific
radioactivity (16).
Three-way analysis of variance was used to assess the effect of insulin, amino acids, age, and
their interaction on tissue protein synthesis. When significant interactions were detected, the value
in each treatment group for each age was compared with the control value by use of t-tests. To
determine the effectiveness of the clamp procedure, individual amino acid, glucose, and insulin
concentrations in each treatment group were compared with their basal concentrations by use of t-

R1-E-00517-2001 7
tests. Differences of P < 0.05 were considered statistically significant for all comparisons except
those for plasma amino acid concentrations. Because there was an increased probability that one of
the 22 amino acid comparisons in each of the four treatment groups would be significantly different
among groups by random chance, a more conservative statistical approach for amino acid
comparisons was used; therefore, probability values of < 0.01 were considered statistically different.
Results are presented as means ± SEM.
RESULTS
Infusions. Pigs at 7 and 26 days of age were infused with insulin, glucose, amino acids,
and/or saline to reproduce: 1) fasting insulin, glucose, and amino acid levels (Control group); 2) fed
insulin, fasting glucose, and fasting amino acid levels (hyperinsulinemic-euglycemic-
euaminoacidemic clamp; Insulin group); 3) fasting insulin, fasting glucose, and fed amino acid levels
(euinsulinemic-euglycemic-hyperaminoacidemic clamp; AA group); and 4) fed insulin, fasting
glucose, and fed amino acid levels (hyperinsulinemic-euglycemic-hyperaminoacidemic clamp;
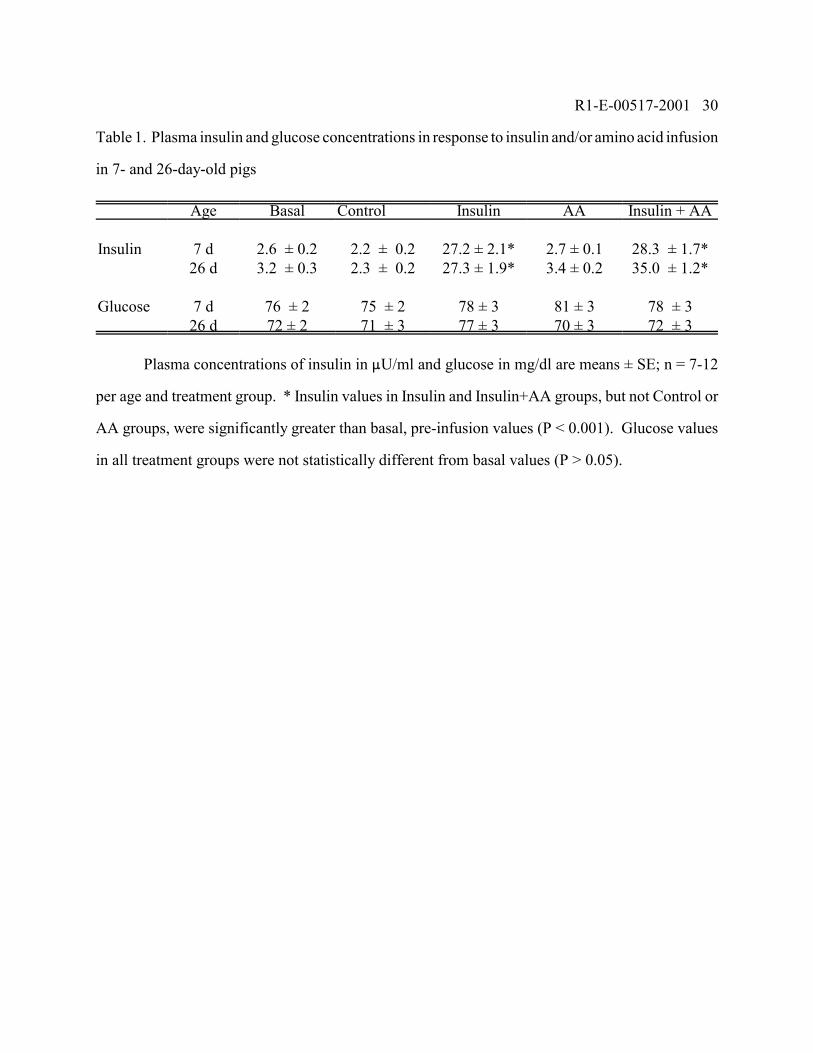
Insulin+AA group). Table 1 shows that the circulating insulin concentrations normally found in the
neonatal pig in the fasting condition and after a meal (3 and 30 �U/ml, respectively ) were largely
reproduced in both 7 and 26-day-old pigs by the infusion of 0 and 100 ng � kg-0.66 � min-1.
Hyperaminoacidemia did not alter circulating insulin concentrations. Plasma glucose concentrations
were maintained at basal fasting levels during the infusion of insulin and/or amino acids in 7- and
26-day-old pigs (Table 1).
Circulating essential and nonessential amino acid concentrations in control and in insulin-
and/or amino acid-infused 7- and 26-day-old pigs are compared to baseline (0 time) values in Figure

R1-E-00517-2001 8
1. During the 4-hr saline infusion period in control pigs, plasma amino acid concentrations remained
stable in 7-day-old pigs, although leucine, isoleucine and lysine increased and alanine and glutamate
decreased in 26-day-old pigs (P < 0.01). Because circulating nonessential amino acid concentrations
decreased in our previous hyperinsulinemic-euglycemic-euaminoacidemic clamp studies which
utilized Trophamine as the amino acid infusate (14, 62), in the current study we developed a new
amino acid mixture which contains higher concentrations of nonessential amino acids than
Trophamine. The circulating concentrations of both nonessential and essential amino acids were
maintained largely at the fasting level during the hyperinsulinemic-euglycemic-euaminoacidemic
clamp using the new amino acid infusate. The exceptions were elevations in histidine and
tryptophan and a reduction in asparagine (P < 0.01). Hyperaminoacidemic clamps, in the presence
of euinsulinemia or hyperinsulinemia, increased the circulating concentrations of essential amino
acids about twofold, and those of nonessential amino acids about 50% (P < 0.01).
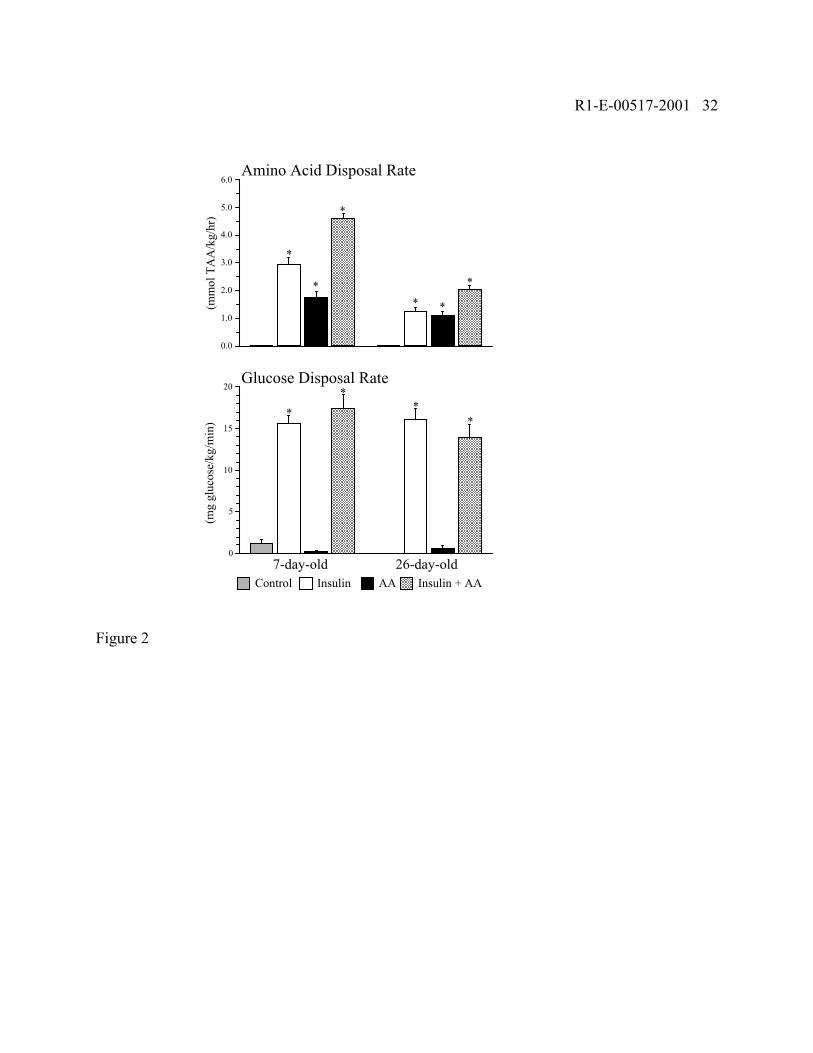
Figure 2 shows the net whole-body amino acid and glucose disposal rates, as indicated by
the amino acid and glucose infusion rates during the last hour of the infusion period. Amino acids
were not infused in the control animals because plasma amino acid concentrations did not fall below
10% of the basal level. During the infusion of either insulin or amino acids, the amino acid disposal
rates were about twofold greater in 7-than in 26-day-old pigs (P < 0.01), but the glucose disposal
rates did not differ significantly with age. The effects of insulin and amino acids on amino acid
disposal were additive.
Tissue protein synthesis. Fractional rates of protein synthesis in skeletal muscles of different
fiber types in 7- and 26-day-old pigs are shown in Fig. 3. Under fasting conditions, the fractional
rates of protein synthesis in longissimus dorsi, gastrocnemius, and masseter muscles were about

R1-E-00517-2001 9
threefold, twofold, and 50% higher in 7- than in 26-day-old pigs, respectively (P < 0.001).
Hyperinsulinemia increased (P < 0.005) protein synthesis in longissimus dorsi (57%), gastrocnemius
(64%), and masseter (36%) muscles, and these responses decreased with age (P < 0.001).
Hyperaminoacidemia also increased (P < 0.01) protein synthesis in longissimus dorsi (48%),
gastrocnemius (50%), and masseter (28%) muscles (P < 0.01). The amino acid response also
decreased with age (P < 0.05). In contrast to whole-body amino acid disposal, the combination of
hyperinsulinemia and hyperaminoacidemia did not exert an additive effect on muscle protein
synthesis (P > 0.05).
To determine whether the stimulation of skeletal muscle protein synthesis by insulin and/or
amino acids is specific to myofibrillar proteins, protein synthesis rates in myofibrillar and
sarcoplasmic proteins were determined in longissimus dorsi muscle of 7-day-old pigs in the four
treatment groups. Hyperinsulinemia and hyperaminoacidemia, either alone or in combination,
increased protein synthesis in both myofibrillar and sarcoplasmic protein fractions to a similar extent
(60-80%; Fig. 4). Rates of protein synthesis did not differ among the two protein fractions in any
treatment group.
Fractional rates of protein synthesis in cardiac muscle, skin, and spleen of 7- and 26-day-old
pigs are shown in Fig. 5. The fractional rates of protein synthesis in cardiac muscle, skin, and spleen
in the basal fasting condition were 15%, 90%, and 30% higher in 7- than in 26-day-old pigs,
respectively (P < 0.01). Hyperinsulinemia increased protein synthesis in cardiac muscle (50%; P <
0.001), skin (34%; P < 0.001), and spleen (26%; P < 0.05), and these responses to insulin decreased
with age (P < 0.05). Hyperaminoacidemia had no effect on protein synthesis in these tissues. The
combination of hyperinsulinemia and hyperaminoacidemia had no greater effect on protein synthesis

R1-E-00517-2001 10
than the effect of hyperinsulinemia alone (P > 0.05).
Fractional rates of protein synthesis in liver, kidney, and pancreas of 7- and 26-day-old pigs
are shown in Fig. 6. The fractional rate of protein synthesis in liver in the basal fasting condition
was 28% higher in 7- than in 26-day-old pigs (P < 0.001). Hyperinsulinemia had no effect on protein
synthesis in any of these organs (P > 0.05). Nevertheless, an elevation in circulating amino acids
increased protein synthesis in liver (27%; P < 0.001), pancreas (28%; P < 0.05), and kidney (10%;
P < 0.005), and the response in liver decreased with age (P < 0.05). The combination of
hyperinsulinemia and hyperaminoacidemia had no greater effect on protein synthesis in these tissues
than the effect of hyperaminoacidemia alone (P > 0.05).
The fractional rates of protein synthesis in the jejunum of 7- and 26-day-old pigs are shown
in Fig. 7. The fractional rate of protein synthesis in jejunum was higher in 7- than in 26-day-old
pigs (P < 0.001). Neither hyperinsulinemia nor hyperaminoacidemia significantly altered the rate
of protein synthesis in the jejunum (P > 0.05).
DISCUSSION
Regulation of skeletal muscle protein synthesis by both insulin and amino acids. Current and
previous studies suggest that insulin plays a key role in the elevated rate of protein deposition and
the increased response of skeletal muscle protein synthesis to feeding in the neonate. First,
postprandial changes in protein synthesis in neonatal pigs are positively correlated with changes in
circulating insulin concentrations (13). Second, insulin infusion, when amino acids and glucose are
maintained near fasting levels, increases amino acid disposal in the neonatal pig and ovine fetus (57,
61). The insulin sensitivity and responsiveness of amino acid disposal decrease with development.

R1-E-00517-2001 11
Third, raising insulin concentrations in the neonatal pig to levels typical of the fed state increases the
rate of skeletal muscle protein synthesis to within the range normally present in the fed state, even
when essential amino acids and glucose are maintained at fasting levels (21, 62). This response to
insulin, like the response to feeding, is attenuated with development. Fourth, insulin infusion
stimulates whole-body and skeletal muscle protein synthesis in the fetal sheep, young lamb, and
weaned rat (28, 38, 60), but has little, if any, effect in the adult human or rat (4, 30, 32, 39, 41).
These findings suggest that the ability of skeletal muscle protein synthesis to respond to the
postprandial rise in insulin wanes with development. Fifth, the developmental decline in the
feeding-induced activation of the insulin signaling pathway leading to translation initiation parallels
the developmental decline in feeding- and insulin-stimulated protein synthesis (21, 37, 53). This
suggests that the high activity of the insulin signaling pathway contributes to the increased response
of muscle protein synthesis rates to feeding in the neonate.
In the present study, we used an amino acid clamp technique to prevent the drop in
circulating amino acid concentrations that occurs when amino acids are not infused concurrently with
insulin. Because the circulating concentrations of many nonessential amino acids decreased at the
highest insulin concentrations in our previous studies (14, 62), in the current study we developed a
new amino acid mixture that contained higher concentrations of nonessential amino acids than the
Trophamine used previously. Using this new amino acid infusate, we maintained almost all
nonessential amino acids, as well as essential amino acids, at the fasting level during the infusion
of insulin. Critically, though, the stimulation of skeletal muscle protein synthesis by insulin was
similar in the current and previous studies (14, 62), and, indeed, was similar to the response to
feeding (12). This suggests that maintenance of nonessential amino acid concentrations is not

R1-E-00517-2001 12
required for the insulin-induced stimulation of skeletal muscle protein synthesis.
To determine the role of amino acids in the postprandial rise in protein synthesis, the new
amino acid mixture was infused to raise circulating amino acid concentrations to near the fed level.
Importantly, circulating concentrations of insulin, measured every 30 min during the 4-hour infusion,
did not increase in response to amino acid infusion. This suggests that the physiological rise in
amino acid concentrations stimulated protein synthesis in skeletal muscle in the absence of a rise in
insulin concentration. Although the combined infusion of amino acids and insulin increased protein
synthesis, the increase was similar to that obtained during amino acid or insulin stimulation alone.
This implies that insulin and amino acids may be interacting with the same signaling pathway within
skeletal muscle. This idea is supported by the observation that the muscle protein synthesis response
to insulin and amino acids decreased with development, in parallel with the developmental decline
in the feeding-induced stimulation of skeletal muscle protein synthesis. In young, adult, and elderly
populations, amino acid infusion, either alone or concurrent with insulin infusion, stimulates protein
synthesis in skeletal muscle (6, 41, 49, 58, 59). This suggests that the ability of amino acids to
stimulate skeletal muscle protein synthesis wanes, but is not lost, with age.
The results of the current study further show that the stimulation of muscle protein synthesis
by amino acids and/or insulin was greater in those muscles containing predominately fast-twitch,
glycolytic muscle fibers. The response to both anabolic agents was greater in longissimus dorsi and
gastrocnemius muscles, which contain fast-twitch fibers that are predominantly glycolytic, than in
the masseter muscle, which is composed entirely of oxidative fibers. These findings are consistent
with our previously reported results (19, 21) and those of others (4, 35), demonstrating that protein
synthesis is more responsive to anabolic agents in muscles composed of predominantly fast fibers

R1-E-00517-2001 13
than in muscles of slow, oxidative fibers. Furthermore, the greater developmental decline in
fractional protein synthesis rates in fast-twitch, glycolytic than in slower oxidative muscles in the
current study is consistent with our previously reported findings in pig and rat muscles (18, 14).
Because the skeletal muscles we studied in the neonatal pig have predominately fast-twitch
contractile characteristics, but different oxidative and glycolytic properties (1, 47, 54), the differences
in protein synthesis rates between muscles which we observed are most likely attributable to the
distinct differences in the muscles metabolic rather than twitch characteristics.
The results of the current study demonstrate that hyperinsulinemia and hyperaminoacidemia,
alone or in combination, stimulate both myofibrillar and sarcoplasmic protein synthesis. In addition,
the degree of stimulation by these agents was proportional. These results are consistent with our
previous studies which showed that the synthesis of both protein fractions is increased similarly by
milk feeding (26) and insulin infusion (14) in neonatal pigs. Furthermore, we have previously shown
in the neonatal pig that myofibrillar and sarcoplasmic protein synthesis rates are similar, as in the
current study, and that the age-associated decline in the synthesis rates of the two protein fractions
is proportional over this phase of development (14).
Regulation of tissue protein synthesis by either insulin or amino acids alone. Feeding
increases protein synthesis in virtually all tissues of the neonatal pig (7, 8, 12, 15). In the current
study, infusing insulin at a dose that reproduced plasma insulin levels in the fed state, while
maintaining glucose, essential amino acids, and nonessential amino acids at the fasting level,
stimulated protein synthesis in cardiac muscle, skin, and spleen. However, in a previous study,
infusing insulin at doses reproducing fed-state plasma insulin levels, while keeping glucose and
essential amino acids, but not nonessential amino acids, at fasting levels, stimulated protein synthesis

R1-E-00517-2001 14
in cardiac muscle and skin, but not spleen (14). This suggests that keeping nonessential amino acid
concentrations at fasting levels is not required for the insulin-induced stimulation of protein synthesis
in cardiac muscle or skin, although it may play a permissive role in spleen. Furthermore, an
elevation in amino acids, in the presence of either euinsulinemia or hyperinsulinemia, had no effect
on protein synthesis in cardiac muscle, skin, or spleen. Thus, the results are consistent with the
hypothesis that the postprandial rise in insulin, but not amino acids, largely mediates the stimulation
of protein synthesis by feeding in the cardiac muscle, skin, and spleen of the neonate.
Insulin also appears to regulate protein synthesis in the cardiac muscle of young (48) but not
adult (41) rats, consistent with the developmental decline in the insulin-induced stimulation of
cardiac muscle protein synthesis observed in the current study. Amino acid infusion has no effect
on cardiac muscle protein synthesis in the adult rat (41), as in the neonatal pigs of the current study,
suggesting that cardiac muscle may be resistant to the anabolic effects of amino acids throughout the
life cycle.
Our previous study showed that insulin infusion does not stimulate, and in some cases,
reduces, protein synthesis in liver, intestine, pancreas, and kidney, despite maintenance of glucose
and essential amino acids at fasting levels (14). In the current study, the infusion of physiological
levels of insulin, when nonessential amino acids, essential amino acids, and glucose were clamped
at fasting levels, did not stimulate, but also did not reduce, protein synthesis in these tissues. This
suggests that the reduction in protein synthesis in liver and intestine during the infusion of insulin
in the previous study (14) was due to a reduction in substrate, i.e., nonessential amino acids or
nitrogen. Moreover, the infusion of amino acids at doses reproducing fed-state amino acid levels
increased protein synthesis in liver, pancreas, and kidney in the current study. These responses to

R1-E-00517-2001 15
amino acid infusion occurred in the presence of either euinsulinemia or hyperinsulinemia. The
results are suggestive of an important role of amino acids, but not insulin, in the regulation of protein
synthesis in the liver, pancreas, and kidney of the neonate.
Many (3, 44, 55), but not all (33, 34), studies report an inability of insulin to stimulate liver
protein synthesis. However, none have reported a reduction in liver protein synthesis with insulin
infusion. The reduction in liver protein synthesis in the previous insulin infusion study (14) may have
been due to the more profound effect of insulin on circulating amino acids in neonatal animals,
which have higher amino acid turnover rates than older animals (23, 61). Indeed, amino acids appear
to stimulate liver protein synthesis in growing (27, 51) but not adult or elderly (2, 44) animals,
consistent with the developmental decline in the amino acid-induced stimulation of liver protein
synthesis in the current study.
In the current study, we found that the physiological elevation of insulin, amino acids, or both
had no effect on protein synthesis in the jejunum. Insulin infusion in healthy adults (42) and in
diabetics (46) reduces protein synthesis in splanchnic tissues, consisting primarily of gut and liver.
However, insulin deprivation modestly reduces, and insulin treatment restores, protein synthesis rates
in the small intestinal mucosa of diabetics (11), suggesting that insulin may be required for the
maintenance of mucosal protein synthesis. Feeding stimulates intestinal protein synthesis in growing
animals (9, 12, 15), but in the current study, amino acids were infused peripherally rather than
enterally. Thus, we postulate that supplying amino acids to the mucosa via the luminal rather than
the arterial route is likely the most important modulator of the feeding-induced stimulation of
intestinal protein synthesis. Indeed, previous studies in neonatal pigs have shown that in the fed
state, dietary amino acids make a greater contribution than arterial amino acids to jejunal protein

R1-E-00517-2001 16
synthesis (52).
Whole-body amino acid and glucose disposal. Previous studies showed that insulin
stimulates the utilization of amino acids for protein deposition in the whole body of the neonatal pig,
and that both the insulin sensitivity and responsiveness of amino acid disposal decrease with
development (61). The current study extends our previous work to show a marked developmental
decline in insulin-stimulated amino acid disposal in the presence of either fasted- or fed-state amino
acid levels. Furthermore, the amount of amino acids required to raise amino acid concentrations to
near the fed state, in the presence of either fasting- or fed-state insulin levels, also decreased with
development. This developmental decrease in amino acid disposal in response to hyperinsulinemia
and/or hyperaminoacidemia paralleled the developmental decline in the ability of these anabolic
agents to stimulate protein synthesis in most tissues. We have recently shown a marked
developmental decline in both insulin receptor abundance and the feeding-induced activation of the
insulin receptor in skeletal muscle of the neonate (53). This developmental change in the activation
of the insulin receptor is propagated down the insulin signaling pathway to many of the eukaryotic
initiation factors that regulate translation (21, 36). Furthermore, the feeding-induced stimulation
of muscle protein synthesis is dependent on activation of a protein kinase, mammalian target of
rapamycin or mTOR (37), which has been shown in vitro to be important in mediating the signal
generated by both amino acids and insulin (31, 50). The lack of developmental decline in insulin-
stimulated glucose disposal in the presence of either euaminoacidemia or hyperaminoacidemia is
surprising, and implies differential developmental regulation of these two insulin-stimulated
metabolic responses.
Perspectives. Feeding stimulates whole body protein synthesis in the newborn infant, but

R1-E-00517-2001 17
the effect is much smaller in the adult (24, 43, 56), implying that different mechanisms regulate
protein synthesis in adults and neonates. Studies using the neonatal pig as an animal model of the
human infant (45) have shown that feeding stimulates protein synthesis in virtually all tissues of the
body of the neonate, a response that decreases with development (12). The results of the current
study strongly support the hypothesis that the feeding-induced stimulation of protein synthesis in
skeletal muscle is mediated by the postprandial rise in both insulin and amino acids. Protein
synthesis in cardiac muscle, skin, and spleen is responsive to insulin but not amino acid stimulation,
whereas liver, kidney, and pancreas are responsive to amino acid but not insulin stimulation.
Because neither insulin nor amino acid infusion altered intestinal protein synthesis, we postulate that
amino acid stimulation by the luminal rather than arterial route may be required for the feeding-
induced stimulation of intestinal protein synthesis. Thus, it appears that the stimulation of protein
synthesis by feeding is mediated by either insulin or amino acids in most tissues of the neonate, but
in skeletal muscle, the feeding-induced stimulation of protein synthesis is regulated by both insulin
and amino acids. The ability of skeletal muscle to respond to the postprandial rise in two anabolic
stimuli likely contributes to the efficient utilization of nutrients for growth in the neonate and the
more rapid gain in protein mass in skeletal muscle than in other tissues of the body in the neonate.

R1-E-00517-2001 18
ACKNOWLEDGMENTS
The authors thank W. Liu, J. Rosenberger, J. Henry, and X. Chang for technical assistance,
J. Cunningham and F. Biggs for care of animals, E. O. Smith for statistical assistance, L. Loddeke
for editorial assistance, A. Gillum for graphics, and J. Croom for secretarial assistance. We
acknowledge Eli Lilly Co. for the generous donation of porcine insulin.
This work is a publication of the USDA/ARS Children’s Nutrition Research Center,
Department of Pediatrics, Baylor College of Medicine and Texas Children’s Hospital, Houston, TX.
This project has been funded in part by National Institute of Arthritis and Musculoskeletal and Skin
Diseases Institute Grant R01 AR44474 and the U.S. Department of Agriculture, Agricultural
Research Service under Cooperative Agreement number 58-6250-6-001. This research was also
supported in part by NIH training grant T32 HD07445. The contents of this publication do not
necessarily reflect the views or policies of the U.S. Department of Agriculture, nor does mention of
trade names, commercial products or organizations imply endorsement by the U.S. Government.
Present address of P. J. Reeds, Ph.D.: University of Illinois, Department of Animal Sciences,
1207 W. Gregory, Room 432, Urbana, IL 61801. Present address of R. Vann, Ph.D.: CMREC,
Mississippi State University, 1320 Seven Springs Road, Raymond, MS 39154.

R1-E-00517-2001 19
REFERENCES
1. Anapol F, Herring, SW. Ontogeny of histochemical fiber types and muscle function in the
masseter muscle of miniature swine. Am J Anthropol 112:595-613,2000.
2. Anthony TG, Anthony JC, Yoshizawa F, Kimball SR, and Jefferson LS. Oral
administration of leucine stimulates ribosomal protein mRNA translation but not global rates
of protein synthesis in the liver of rats. J Nutr 131: 1171-1176, 2001.
3. Ahlman B, Charlton M, Fu A, Berg C, O’Brian P, and Nari KS. Insulin’s effect on
synthesis rates of liver proteins. A swine model comparing various precursors of protein
synthesis. Diabetes 50: 947-954, 2001.
4. Baillie AGS, and Garlick PJ. Attenuated responses of muscle protein synthesis to fasting
and insulin in adult female rats. Am J Physiol Endocrinol Metab 262: E1-E5, 1991.
5. Beckett PR, Hardin DS, Davis TA, Nguyen HV, Wray-Cahen D, and Copeland KC.
Spectrophometric assay for measuring branched chain amino acid concentrations: application
for measuring the sensitivity of protein metabolism to insulin. Anal Biochem 240: 48-53,
1996.
6. Bennet WM, Connacher AA, Scrimgeour CM, and Rennie MJ. The effect of amino acid
infusion on leg protein turnover assessed by L-[15N]phenylalanine and L-[1-13C]leucine
exchange. Eur J Clin Invest 20: 41-50, 1990.
7. Burrin DG, Davis TA, Ebner S, Schoknecht PA, Fiorotto ML, and Reeds PJ. Colostrum
enhances the nutritional stimulation of vital organ protein synthesis in neonatal pigs. J Nutr
127: 1284-1289, 1997.
8. Burrin DG, Davis TA, Ebner S, Schoknecht PA, Fiorotto ML, Reeds PJ, and McAvoy
S. Nutrient-independent and nutrient-dependent factors stimulate protein synthesis in
colostrum-fed newborn pigs. Pediatr Res 37: 593-599, 1995.

R1-E-00517-2001 20
9. Burrin DG, Davis TA, Fiorotto ML, and Reeds PJ. Stage of development and fasting
affect protein synthetic activity in the gastrointestinal tissues of suckling rats. J Nutr 121:
1099-1108, 1991.
10. Burrin DG, Davis TA, Fiorotto ML, and Reeds PJ. Hepatic protein synthesis in suckling
rats: effects of stage of development and fasting. Pediatr Res 31: 247-252, 1992.
11. Charlton M, Ahlman B, and Nair KS. The effect of insulin on human small intestinal
mucosal protein synthesis. Gastroenterology 188: 299-306, 2000.
12. Davis TA, Burrin DG, Fiorotto ML, and Nguyen HV. Protein synthesis in skeletal muscle
and jejunum is more responsive to feeding in 7- than in 26-day-old pigs. Am J Physiol
Endocrinol Metab 270: E802-E809, 1996.
13. Davis TA, Burrin DG, Fiorotto ML, Reeds PJ, and Jahoor F. Roles of insulin and amino
acids in the regulation of protein synthesis in the neonate. J Nutr 128: 347S-350S, 1998.
14. Davis TA, Fiorotto ML, Beckett PR, Burrin DG, Reeds PJ, Wray-Cahen D, Nguyen
HV. Differential effects of insulin on peripheral and visceral tissue protein synthesis in
neonatal pigs. Am J Physiol Endocrinol Metab 280: E770-E779, 2001.
15. Davis TA, Fiorotto ML, Burrin DG, Pond WG, and Nguyen HV. Intrauterine growth
restriction does not alter response of protein synthesis to feeding in newborn pigs. Am J
Physiol Endocrinol Metabol 272: E877-E884, 1997.
16. Davis TA, Fiorotto ML, Nguyen HV, and Burrin DG. Aminoacyl-tRNA and tissue free
amino acid pools are equilibrated after a flooding dose of phenylalanine. Am J Physiol
Endocrinol Metabol 277: E103-E109, 1999.
17. Davis TA, Fiorotto ML, Nguyen HV, Burrin DG, and Reeds PJ. Response of muscle
protein synthesis to fasting in suckling and weaned rats. Am J Physiol Regulatory Integrative
Comp Physiol 261: R1373-1380,1991.

R1-E-00517-2001 21
18. Davis TA, Fiorotto ML, Nguyen HV, and Reeds PJ. Protein turnover in skeletal muscle
of suckling rats. Am J Physiol Regulatory Integrative Comp Physiol 257: R1141-R1146,
1989.
19. Davis TA, Fiorotto ML, Nguyen HV, and Reeds PJ. Enhanced response of muscle protein
synthesis and plasma insulin to food intake in suckled rats. Am J Physiol Regulatory
Integrative Comp Physiol 265: R334-R340, 1993.
20. Davis TA, Fiorotto ML, and Reeds PJ. Amino acid compositions of body and milk protein
change during the suckling period in rats. J Nutr 123: 947-956, 1993.
21. Davis TA, Nguyen HV, Suryawan A, Bush JA, Jefferson LS, and Kimball LS.
Developmental changes in the feeding-induced stimulation of translation initiation in skeletal
muscle and liver of neonatal pigs. Am J Physiol Endocrinol Metab. 279: E1226-1234, 2000.
22. DeFronzo RA, Tobin JD, and Andres R. Glucose clamp technique: a method for
quantifying insulin secretion and resistance. Am J Physiol Endocrinol Metab 237 : E214-
E223, 1979.
23. Denne SC, and Kalhan SC. Leucine metabolism in human newborns. Am J Physiol
Endocrinol Metab 253: E608-E615, 1987.
24. Denne SC, Rossi EM, and Kalhan SC. Leucine kinetics during feeding in normal
newborns. Pediatr Res 30: 23-27, 1991.
25. Fiorotto, ML, Davis TA, and Reeds PJ. Regulation of myofibrillar protein turnover during
maturation in normal and undernourished rat pups. Am J Physiol Regulatory Integrative
Comp Physiol 278: R845-R854, 2000.
26. Fiorotto, ML, Davis TA, Reeds PJ, and Burrin DG. Nonnutritive factors in colostrum
enhance myofibrillar protein synthesis in the newborn pig. Pediatr Res 48: 511-517, 2000.
27. Flaim KE, Peavy DE, Everson WV, and Jefferson LS. The role of amino acids in the

R1-E-00517-2001 22
regulation of protein synthesis in perfused rat liver. J Biol Chem 257: 2932-2938, 1982.
28. Garlick P J, Fern M, and Preedy VR. The effect of insulin infusion and food intake on
muscle protein synthesis in postabsorptive rats. Biochem J 210:669-676, 1983.
29. Garlick PJ, McNurlan MA, and Preedy VR. A rapid and convenient technique for
measuring the rate of protein synthesis in tissues by injection of [3H]phenylalanine. Biochem
J 192: 719-723, 1980.
30. Gelfand RA, and Barrett EJ. Effect of physiologic hyperinsulinemia on skeletal muscle
protein synthesis and breakdown in man. J Clin Invest 80:1-6, 1987.
31. Hara K, Yonezawa K, Weng QP, Kozlowski MT, Belham C, and Avruch J. Amino acid
sufficiency and mTOR regulate p70 S6 kinase and eIF-4E BP1 through a common effector
mechanism. J Biol Chem 273: 1448-124494, 1998.
32. Heslin MJ, Newman E, Wolf RF, Pisters PWT, and Brennan MF. Effect of
hyperinsulinemia on whole body and skeletal muscle leucine carbon kinetics in humans. Am
J Physiol Endocrinol Metab 262: E911-E918, 1992.
33. Hsu C-J, Kimball SR, Antonetti DA, and Jefferson LS. Effects of insulin on total RNA,
poly(A)RNA, and mRNA in primary cultures of rat hepatocytes. Am J Physiol Endocrinol
Metab 263: E1106-E1112, 1992.
34. Jefferson LS, Liao WSL, Peavy DE, Miller TB, Appel MC, Taylor JM. Diabetes-induced
alterations in liver protein synthesis. Changes in the relative abundance of mRNAs for
albumin and other plasma proteins. J Biol Chem 258: 1369-1375, 1983.
35. Kimball SR, and Jefferson LS. Cellular mechanisms involved in the action of insulin on
protein synthesis. Diabetes Metab Rev 4: 773:787, 1988.
36. Kimball SR, Jefferson LS, Nguyen HV, Suryawan A, Bush JA, and Davis TA. Feeding
stimulates protein synthesis in muscle and liver of neonatal pig through an mTOR-dependent

R1-E-00517-2001 23
process. Am J Physiol Endocrinol Metab 279: E1080-E1087, 2000.
37. Kimball SR, Farrell PA, Nguyen HV, Jefferson LS, and Davis TA. Developmental
decline in components of signal transduction pathways regulating muscle protein synthesis
in neonatal pigs. Am J Physiol Endocrinol Metab, In press.
38. Liechty EA, Boyle DW, Moorehead H, Liu YM, and Denne SC. Effect of
hyperinsulinemia on ovine fetal leucine kinetics during prolonged maternal fasting. Am J
Physiol Endocrinol Metab 263: E696-E702, 1992.
39. Louard RJ, Fryburg DA, R. Gelfand A, and Barrett EJ. Insulin sensitivity of protein and
glucose metabolism in human forearm skeletal muscle. J Clin Invest 90: 2348-2354, 1992.
40. McCracken KJ, Eddie SM, and Stevenson WG. Energy and protein nutrition of early-
weaned pigs. Effect of energy intake and energy: protein on growth, efficiency and nitrogen
utilization of pigs between 8-32d. Br J Nutr 43: 289-304, 1980.
41. McNulty PA, Young LH, and Barrett EJ. Response of rat heart and skeletal muscle
protein in vivo to insulin and amino acid infusion. Am J Physiol Endocrinol Metab 264:
E958-E965, 1993.
42. Meek SE, Persson M, Ford GC, and Nair KS. Differential regulation of amino acid
exchange and protein dynamics across splanchnic and skeletal muscle beds by insulin in
healthy human subjects. Diabetes 47: 1824-1835, 1998.
43. Melville S, McNurlan MA, McHardy KC, Broom J, Milne E, Calder AG, and Garlick
PJ. The role of degradation in the acute control of protein balance in adult man: failure of
feeding to stimulate protein synthesis as assessed by L-[1-13C]leucine infusion. Metabolism
38: 248-255, 1989.
44. Mosoni L, Houlier M-L, Mirand PP, Bayle G, and Grizard J. Effect of amino acids alone
or with insulin on muscle and liver protein synthesis in adult and old rats. Am J Physiol

R1-E-00517-2001 24
Endocrinol Metab 624: E613-E620, 1993.
45. Moughan PJ, and Rowan AM. The pig as a model animal for human nutrition research.
Proc Nutr Soc N Z 14: 116-123, 1989.
46. Nair KS, Ford GC, Ekberg K, Fernqvist-Forbes E, and Wahren J. Protein dynamics in
whole body and in splanchnic and leg tissues in type I diabetic patients. J Clin Invest 95:
2926-2937, 1995.
47. Ouali A, Talmant A. Calpains and calpastatin distribution in bovine, porcine and ovine
skeletal muscle. Meat Science 28:331-348, 1990.
48. Pain V, Albertse EC, and Garlick PJ. Protein metabolism in skeletal muscle, diaphragm,
and heart of diabetic rats. Am J Physiol Endocrinol Metab 245: E604-E610, 1983.
49. Preedy VT, Garlick PJ. Response of muscle protein synthesis to parenteral administration
of amino acid mixtures in growing rats. J Parenter Enter Nutr 17: 113-118, 1993.
50. Scott PH, Brunn GJ, Kohn AD, Roth RA, and Lawrence JC Jr. Evidence of insulin-
stimulated phosphorylation and activation of the mammalian target of rapamycin mediated
by a protein kinase B signaling pathway. Proc Natl Acad Sci USA 95: 7772-7777, 1998.
51. Shah OJ, Antonetti DA, Kimball SR, and Jefferson LS. Leucine, glutamine, and tyrosine
reciprocally modulate the translation initiation factors eIF4F and eIF2B in perfused liver. J
Biol Chem 274: 36168-36175, 1999.
52. Stoll B, Burrin DG, Henry JG, Jahoor F, and Reeds PJ. Dietary and systemic
phenylalanine utilization for mucosal and hepatic constitutive protein synthesis in pigs. Am
J Physiol Gastrointest Liver Physiol 276: G49-G57, 1999.
53. Suryawan A, Nguyen HV, Bush JA, and Davis TA. Developmental changes in the
feeding-induced activation of the insulin-signaling pathway in neonatal pigs. Am J Physiol
Endocrinol Metab 281:E908-E915, 2001.

R1-E-00517-2001 25
54. Suzuki A, Cassens RG. A histochemical study of myofiber types in muscle of the growing
pig. J Anim Sci 51:1449-1461, 1980.
55. Tauveron I, Larbaud D, Champredon C, Debras E, Tesseraud S, Bayle G, Bonnet Y,
Thieblot P, and Grizard J. Effect of hyperinsulinemia and hyperaminoacidemia on muscle
and liver protein synthesis in lactating goats. Am J Physiol Endocrinol Metab 267: E877-
E885, 1994.
56. Tessari P, Zanetti M, Barazzoni R, Vettore M, and Michielan F. Mechanism of
postprandial protein accretion in human skeletal muscle. J Clin Invest 98: 1361- 1372, 1996.
57. Thureen PJ, Scheer B, Anderson SM, Tooze JA, Young DA, and Hay WW. Effect of
hyperinsulinemia on amino acid utilization in the ovine fetus. Am J Physiol Endocrinol
Metab 279: E1294-E1304, 2000.
58. Vary TC, Jefferson LS, and Kimball SR. Amino acid-induced stimulation of translation
initiation in rat skeletal muscle. Am J Physiol Endocrinol Metab 277: E1077-E1086, 1999
59. Volpi E, Ferrando AA, Yeckel CW, Tipton KD, and Wolfe RR. Exogenous amino acids
stimulate net muscle protein synthesis in the elderly. J Clin Invest 101: 2000-2007, 1998.
60. Wester TJ, Lobley GE, Birnie LM, and Lomax MA. Insulin stimulates phenylalanine
uptake across the hind limb in fed lambs. J Nutr 130: 608-611, 2000.
61. Wray-Cahen D, Beckett PR, Nguyen HV, and Davis TA. Insulin-stimulated amino acid
utilization during glucose and amino acid clamps decreases with development. Am J Physiol
Endocrinol Metab 273: E305-E314, 1997.
62. Wray-Cahen D, Nguyen HV, Burrin DG, Beckett PR, Fiorotto ML, Reeds PJ, Wester
TJ, and Davis TA. Response of skeletal muscle protein synthesis to insulin in suckling pigs
decreases with development. Am J Physiol Endocrinol Metab 275: E602-E609, 1998.
63. Young VR. The role of skeletal and cardiac muscle in the regulation of protein metabolism.

R1-E-00517-2001 26
In: Mammalian Protein Metabolism, edited by HN Munro. New York:Academic, 1970, vol.
4, p. 585-674.

R1-E-00517-2001 27
Figure Legends
Fig. 1: Plasma concentrations of essential amino acids and nonessential amino acids during
control, hyperinsulinemic-euglycemic-euaminoacidemic (Insulin), euinsulinemic-euglycemic-
hyperaminoacidemic (AA), and hyperinsulinemic-euglycemic-hyperaminoacidemic (Insulin+AA)
clamps in 7- and 26-day-old pigs. Values are means ± SE; n = 7-12 per age and treatment group.
*Amino acid values statistically significantly different from basal values (P < 0.01).
Fig. 2: Whole-body net amino acid and glucose disposal rates during control,
hyperinsulinemic-euglycemic-euaminoacidemic (Insulin), euinsulinemic-euglycemic-
hyperaminoacidemic (AA), and hyperinsulinemic-euglycemic-hyperaminoacidemic (Insulin+AA)
clamps in 7- and 26-day-old pigs. Values are means ± SE; n = 7-12 per age and treatment group.
Amino acid disposal rate was the average rate of infusion of total amino acids (TAA) from an amino
acid mixture necessary to maintain circulating BCAA near either the basal fasting level or twofold
the fasting level from 180 to 240 min of infusion. Amino acids were not infused in control animals.
Amino acid disposal rates were greater in 7- than in 26-day-old pigs (P < 0.01). *Disposal rates
statistically significantly different from control values (P < 0.05)
Fig. 3: Fractional protein synthesis rates in longissimus dorsi, gastrocnemius, and masseter
muscles in 7- and 26-day-old pigs during control, hyperinsulinemic-euglycemic-euaminoacidemic
(Insulin), euinsulinemic-euglycemic-hyperaminoacidemic (AA), and hyperinsulinemic-euglycemic-
hyperaminoacidemic (Insulin+AA) clamps. Values are means ± SE; n = 7-12 per age and treatment
group. Protein synthesis decreased with age in longissimus dorsi, gastrocnemius, and masseter
muscles (P < 0.001). Hyperinsulinemia increased protein synthesis in all muscles (P < 0.005) and
the response decreased with age (P < 0.001). Hyperaminoacidemia increased protein synthesis in
all muscles (P < 0.01) and the response decreased with age (P < 0.05). *Protein synthesis rates
statistically significantly different from control rates (P < 0.05).
Fig. 4: Fractional synthesis rates in total (TP), myofibrillar (MP), and sarcoplasmic (SP)

R1-E-00517-2001 28
proteins in longissimus dorsi muscle of 7-day-old pigs during control, hyperinsulinemic-euglycemic-
euaminoacidemic (Insulin), euinsulinemic-euglycemic-hyperaminoacidemic (AA), and
hyperinsulinemic-euglycemic-hyperaminoacidemic (Insulin+AA) clamps. Values are means ± SE;
n = 7-12 per age and treatment group. Hyperinsulinemia and hyperaminoacidemia increased protein
synthesis in all fractions (P < 0.05). *Protein synthesis rates statistically significantly different from
control rates (P < 0.05).
Fig. 5: Fractional protein synthesis rates in cardiac muscle, skin, and spleen in 7- and 26-day-
old pigs during control, hyperinsulinemic-euglycemic-euaminoacidemic (Insulin), euinsulinemic-
euglycemic-hyperaminoacidemic (AA), and hyperinsulinemic-euglycemic-hyperaminoacidemic
(Insulin+AA) clamps. Values are means ± SE; n = 7-12 per age and treatment group. Protein
synthesis decreased with age in cardiac muscle, skin, and spleen (P < 0.001). Hyperinsulinemia
increased protein synthesis in cardiac muscle (P < 0.001), skin (P < 0.001), and spleen (P < 0.05),
and the responses in cardiac muscle (P < 0.005), skin (P < 0.05), and spleen (P < 0.05) decreased
with age. *Protein synthesis rates statistically significantly different from control rates (P < 0.05).
Fig. 6: Fractional protein synthesis rates in liver, kidney, and pancreas of 7- and 26-day-old
pigs during control, hyperinsulinemic-euglycemic-euaminoacidemic (Insulin), euinsulinemic-
euglycemic-hyperaminoacidemic (AA), and hyperinsulinemic-euglycemic-hyperaminoacidemic
(Insulin+AA) clamps. Values are means ± SE; n = 7-12 per age and treatment group. Protein
synthesis decreased with age in liver (P < 0.001). Hyperaminoacidemia increased protein synthesis
in liver (P < 0.001), kidney (P < 0.005), and pancreas (P < 0.05), and the response in liver decreased
with age (P < 0.05). *Protein synthesis rates statistically significantly different from control rates
(P < 0.05).
Fig. 7: Fractional protein synthesis rates in jejunum of 7- and 26-day-old pigs during control,
hyperinsulinemic-euglycemic-euaminoacidemic (Insulin), euinsulinemic-euglycemic-
hyperaminoacidemic (AA), and hyperinsulinemic-euglycemic-hyperaminoacidemic (Insulin+AA)

R1-E-00517-2001 29
clamps. Values are means ± SE; n = 7-12 per age and treatment group. Protein synthesis decreased
with age in jejunum (P < 0.001).
R1-E-00517-2001 30
Table 1. Plasma insulin and glucose concentrations in response to insulin and/or amino acid infusion
in 7- and 26-day-old pigs
Age Basal Control Insulin AA Insulin + AA
Insulin 7 d 2.6 ± 0.2 2.2 ± 0.2 27.2 ± 2.1* 2.7 ± 0.1 28.3 ± 1.7*26 d 3.2 ± 0.3 2.3 ± 0.2 27.3 ± 1.9* 3.4 ± 0.2 35.0 ± 1.2*
Glucose 7 d 76 ± 2 75 ± 2 78 ± 3 81 ± 3 78 ± 326 d 72 ± 2 71 ± 3 77 ± 3 70 ± 3 72 ± 3
Plasma concentrations of insulin in �U/ml and glucose in mg/dl are means ± SE; n = 7-12
per age and treatment group. * Insulin values in Insulin and Insulin+AA groups, but not Control or
AA groups, were significantly greater than basal, pre-infusion values (P < 0.001). Glucose values
in all treatment groups were not statistically different from basal values (P > 0.05).

R1-E-00517-2001 31
**
**
*
** **
**
**
**
**
*
**
**
**
**
*
**
*
*
*
*
**
**
**
*
**
**
*
*
*
*
*
*
***
**
**
**
**
**
* * *
*
*
* *
**
*
*
*
**
**
***
*
**
*
ALA ASN ASP CIT GLU GLN GLY ORN PRO SER TAU TYR
Nonessential
Baseline Control Insulin AA Insulin + AA
ALA ASN ASP CIT GLU GLN GLY ORN PRO SER TAU TYR0
200
400
600
800
1000
1200
(nm
ol/m
l)
Nonessential
26-day-oldPlasma Amino Acids
ARG HIS ILE LEU LYS MET PHE THR TRP VAL
Essential
7-day-oldPlasma Amino Acids
ARG HIS ILE LEU LYS MET PHE THR TRP VAL0
100
200
300
400
500
600
(nm
ol/m
l)
Essential
Figure 1
R1-E-00517-2001 32
Amino Acid Disposal Rate
0.0
1.0
2.0
3.0
4.0
5.0
6.0
(mm
ol T
AA
/kg/
hr)
*
*
*
* *
*
**
*
*
Glucose Disposal Rate
7-day-old 26-day-old0
5
10
15
20
(mg
gluc
ose/
kg/m
in)
Control Insulin AA Insulin + AA
Figure 2

R1-E-00517-2001 33
7-day-old 26-day-old0
5
10
15
20
25
Ks (%
day)
0
5
10
15
20
25
Ks (%
day)
0
5
10
15
20
25
Ks (%
day)
ControlInsulin AAInsulin + AA
Longissimus Dorsi
Gastrocnemius
Masseter
**
*
* * *
**
*
** *
* *
*
Figure 3

R1-E-00517-2001 34
TP MP SP0
5
10
15
20
25K
s (%da
y)
Control Insulin AA Insulin + AA
* * * * *
*
*
* *
Figure 4

R1-E-00517-2001 35
7-day-old 26-day-old0
10
20
30
40
Ks (%
day)
ControlInsulin AAInsulin + AA
0
10
20
30
40
Ks (%
day)
0
5
10
15
20
25
Ks (%
day)
Heart
Skin
Spleen
*
*
**
**
*
**
Figure 5

R1-E-00517-2001 36
* *
**
**
* *
**
7-day-old 26-day-old0
20
40
60
80
100
Ks (%
day)
0
10
20
30
40
50
Ks (%
day)
0
20
40
60
80
100
Ks (%
day)
ControlInsulin AAInsulin + AA
Liver
Kidney
Pancreas
Figure 6

R1-E-00517-2001 37
Intestine
7-day-old 26-day-old0
10
20
30
40
50K
s (%da
y)
Control Insulin AA Insulin + AA
Figure 7