
Hydrogel membrane electrolyte for electrochemical capacitors

Biomaterials 25 (2004) 2789–2798
ARTICLE IN PRESS
*Correspondin
E-mail addres
0142-9612/$ - see
doi:10.1016/j.bio
Stimulation of in vivo angiogenesis by cytokine-loaded hyaluronicacid hydrogel implants
R.A. Peattiea,*, A.P. Nayateb, M.A. Firpoc, J. Shelbyc, R.J. Fisherd, G.D. Prestwichb
aDepartment of Chemical Engineering, Oregon State University, 103 Gleeson Hall, Corvallis, OR 97331, USAbDepartment of Medicinal Chemistry, The University of Utah, 419 Wakara Way, Suite 205, Salt Lake City, UT 84108, USA
cDepartment of Surgery, The University of Utah, School of Medicine, 30 N., 1930 E., Salt Lake City, UT 84132, USAdDepartment of Chemical Engineering, Massachusetts Institute of Technology, Building 66, Room 446, Cambridge, MA 02139, USA
Received 21 August 2003; accepted 15 September 2003
Abstract
Crosslinked hyaluronic acid (HA) hydrogels were evaluated for their ability to elicit new microvessel growth in vivo when
preloaded with one of two cytokines, vascular endothelial growth factor (VEGF) or basic fibroblast growth factor (bFGF). HA film
samples were surgically implanted in the ear pinnas of mice, and the ears retrieved 7 or 14 days post implantation. Histologic
analysis showed that all groups receiving an implant demonstrated significantly more microvessel density than control ears
undergoing surgery but receiving no implant (po0:01). Moreover, aqueous administration of either growth factor produced
substantially more vessel growth than an HA implant with no cytokine. However, the most striking result obtained was a dramatic
synergistic interaction between HA and VEGF. Presentation of VEGF in crosslinked HA generated vessel density of NI ¼ 6:7 at
day 14, where NI is a neovascularization index defined below, more than twice the effect of the sum of HA alone (NI ¼ 1:8) plusVEGF alone (NI ¼ 1:3). This was twice the vessel density generated by co-addition of HA and bFGF (NI ¼ 3:4; po0:001). Newtherapeutic approaches for numerous pathologies could be notably enhanced by the localized, synergistic angiogenic response
produced by release of VEGF from crosslinked HA films.
r 2003 Elsevier Ltd. All rights reserved.
Keywords: Hyaluronic acid; Hydrogel; Growth factors; Cytokine; VEGF; bFGF; Angiogenesis
1. Introduction
Hyaluronic acid (HA) is widely utilized as a biosyn-thetic biomaterial because of its biocompatibility anddiverse array of physiologic functions. In its naturalsetting, HA is the only non-sulfated glycosaminoglycan(GAG) present in the extracellular matrix (ECM)of vertebrates. Consisting of repeating disaccharideunits (b-1,4-d-glucuronic acid—b-1,3-N-acetyl-d-glucosa-mine) with an overall molecular weight between 100 and5000 kDa, HA is present in all connective tissues as amajor constituent of ECM [1]. HA is also a majorconstituent of the vitreous (0.1–0.4mg/g), synovial jointfluid (3–4mg/ml) and hyaline cartilage, where it reachesapproximately 1mg/g wet weight. Serum HA levelsrange from 10–100 mg/l, but can be elevated during
g author. Tel.: 541-737-9359; fax: 541-737-4600.
s: [email protected] (R.A. Peattie).
front matter r 2003 Elsevier Ltd. All rights reserved.
materials.2003.09.054
disease. Clearance of HA from the systemic circulationresults in a half-life of 2.5–5.5min in plasma.In solution, HA assumes a stiffened helical config-
uration due to hydrogen bonding, and the ensuing coilstructure traps approximately 1000� its weight in water[2]. The highly viscous aqueous solutions thus formedgive HA unique physicochemical and biologic properties[3] that preserve tissue hydration, regulate tissuepermeability through steric exclusion and permit jointlubrication. In the ECM of connective tissues, HA formsa scaffold for binding other large GAGs and proteogly-cans (aggrecans) [4], which are maintained throughspecific HA–protein interactions. Consequently, HAplays important roles in maintaining tissue morphologicorganization, preserving extracellular space and trans-porting ions, solutes and nutrients. Along with ECMproteins, HA binds to specific cell surface receptors suchas CD44 and RHAMM [5]. The resulting activation ofintracellular signaling events leads to cartilage ECMstabilization [6,7], regulates cell adhesion and mobility

ARTICLE IN PRESSR.A. Peattie et al. / Biomaterials 25 (2004) 2789–27982790
[8,9] and promotes cell proliferation and differentiation[10]. Receptors for HA, and the organization of proteinscaffolds and protein–HA complexes, have been recentlyreviewed [10–15].Angiogenesis, the formation of new capillary blood
vessels by outgrowth from existing microvessels, is acentral aspect of normal physiologic processes such astissue and organ development, female reproductivefunctions and wound healing, as well as of pathologictumor development. HA signaling takes place duringmorphogenesis and embryologic development [16],modulation of inflammation [17] and in the stimulationof wound healing [18]. In correspondence with thesefunctions, HA is a strong inducer of angiogenesis,although its biologic activity in tissues has been shownto depend on molecular size [19–21]. Thus highmolecular weight native-HA (n-HA) has been shownto inhibit angiogenesis [22], whereas degradation pro-ducts of low molecular weight stimulate endothelial cellproliferation and migration [23–25]. OligosaccharideHA fragments (o-HA) have been shown to induceangiogenesis in several animal models, including thechick chorioallantois [26], in rat skin [25] and in skingraft preparations [27], as well as within in vitro collagengels [28].The process of angiogenesis is regulated by more than
two dozen known cytokines that appear in complexseries. Of these, the most important and thoroughlyinvestigated are basic fibroblast growth factor (bFGF)and vascular endothelial growth factor (VEGF). Theroles of these growth factors in the process of micro-vessel development have been reviewed by severalauthors in the last 2 years [29–36]. Neovessel formationbegins with growth factor appearance and is followed bythe emergence of matrix metalloproteinase enzymes.Proteinase activity leads to breakdown of the parentcapillary basement membrane and ECM, along withrelease of further ECM-bound growth factors includingVEGF [34]. Proliferating endothelial cells are then ableto migrate into the matrix space and form a capillarysprout. Sprouts grow by continuing endothelial prolif-eration, gradually forming a lumen as they organize intotubules. New microvessels become mature when con-tiguous tubules anastomose with each other and a newbasement membrane is formed [35,37]. Each of thesesteps is regulated by a balance between stimulatory andinhibitory cytokines. In addition, ECM componentsincluding collagen, laminin and other glycoproteins andGAGs have been shown to have substantial roles inmicrovessel formation [38,39].The possibility of a synergistic interaction between
specific growth factors and o-HA in vitro was raised byMontesano et al. [28]. Using a three-dimensionalcollagen gel, the authors showed that a combination ofVEGF and o-HA can induce an in vitro angiogenicresponse greater than the sum of that provoked by each
agent individually. Recently, the development of novel,biocompatible HA hydrogel formulations with mechan-ical properties suitable for tissue implantation [40] hasmade it possible to test the hypothesis that a similarresponse can be obtained in vivo. Previous applicationsof these HA-based biomaterials include scaffolds fortissue engineering, delivery systems for therapeuticpharmaceutics, surface layers to prevent post-surgicaladhesions and dressings for improved wound healing[41–43]. Herein, we describe the use of a modified HAhydrogel for localized delivery of VEGF and bFGF.Such a film can be expected to participate in the tissueresponse itself, not merely serve as a passive deliveryvehicle. We show that a very strong, synergisticangiogenic response can be achieved in vivo if thecytokine is appropriately selected, though not allvasoactive growth factors elicit synergistically increasedmicrovessel growth.
2. Materials and methods
2.1. Materials
Fermentation-derived HA (HA, sodium salt,MW ¼ 1:5� 106 Da) was obtained from Clear SolutionsBiotech, Inc. (Stony Brook, NY). 1-Ethyl-3-[3-(dimethy-lamino)-propyl]carbodiimide (EDCI) and adipic dihy-drazide (ADH) were from Aldrich Chemical Co.(Milwaukee, WI). Chondroitin sulfate C (CS-C), VEGF(human recombinant, 165 amino acid, MW ¼ 46 kDa)and bFGF (human recombinant, 146 amino acid,MW ¼ 18 kDa) were obtained from Sigma (St. Louis,MO). Poly(ethylene glycol)-propiondialdehyde (PEG-diald) (MW ¼ 3400Da) was from Shearwater Polymers,Inc. (Huntsville, AL). Balb/c mice were purchased fromCharles River Laboratories (Wilmington, MA). Isoflur-ane was acquired from Abbott Laboratories (NorthChicago, IL).
2.2. Hydrogel film preparation
HA-ADH and HA hydrogels were synthesized aspreviously described [40,43]. In brief, a 5mg/ml solutionof HA in water was treated with ADH and EDCI, andthe pH was maintained at 4.75 by addition of 1m HCl.The reaction was stopped by raising the solution pH to7.0, and the HA–ADH product was isolated andpurified by extensive dialysis followed by centrifugationand lyophilization. The resulting degree of ADHsubstitution was evaluated by 1H-NMR [40]. A similarprocess was followed using 25mg/ml of CS to make CS–ADH.GAG hydrogels were made by dissolving the purified
HA–ADH or CS–ADH in H2O, at a concentration of25mg/ml. Separately, PEG-diald was also dissolved in

ARTICLE IN PRESS
Fig. 1. Chemical structure of crosslinked HA–ADH.
Fig. 2. Schematic diagram of mouse model, showing implant
placement in the ear pinna.
R.A. Peattie et al. / Biomaterials 25 (2004) 2789–2798 2791
H2O, at 25mg/ml. Volumes of each solution were addedto a small polystyrene dish in a 1:2 ratio of aldehyde tohydrazide functionalities, and the solutions were mixedat 0�C with gentle swirling. A hydrogel formed within3–5min, with PEG-diald as a crosslinking agent for theHA–ADH chains (Fig. 1). However, to insure that thegel was uniform and homogeneous, continuous gentleagitation on an orbital platform was maintained for24 h. Hydrogels thus produced were subsequently keptin open dishes for 24 h at 37�C, which allowed them todehydrate to flexible, dry, durable films 1mm thick.Round disks of 4mm diameter, containing 0.5mg HA–ADH, were cut directly from these films for implanta-tion. All steps were carried out under aseptic conditions.In the hydrated gel state, these HA–ADH hydrogels
have been shown to have an open, highly permeablenetwork structure suitable for sequestering exogenousagents [40]. As a result, when it was desired that agrowth factor be present in the gel, either VEGF orbFGF was incorporated by adding aliquots of a stocksolution into the HA–ADH solution prior to addition ofPEG-diald. Crosslinking then proceeded with no othermodifications. Most experiments were carried out with agrowth factor concentration yielding 25 ng cytokine inthe final 4mm disk, although dose response tests werealso conducted with 0.25, 2.5 and 250 ng. For cases inwhich it was desired to test the effect of the growthfactor alone, without HA, an 0.5ml, aqueous, liquidaliquot containing 25 ng was delivered to the animals.
2.3. Surgical and experimental procedures
The ability of HA hydrogels to stimulate angiogenesiswas evaluated in a mouse model. All procedures andexperiments were carried out with the approval of theUniversity of Utah Institutional Animal Care and UseCommittee. Male Balb/c mice aged 6–8 weeks were
anesthetized with 2.5% isoflurane using an inhalationanesthesia system (VetEquip Inc., Pleasanton, CA).Once a deep general anesthetic plane had been reached,a 4–5mm shallow incision was made through thesuperficial skin of the right ear pinna posterior surface.A blunt probe was inserted into the pinna through thisincision and a 5mm diameter pocket opened immedi-ately subcutaneously (Fig. 2). For those animalsreceiving an implant, a GAG film disk was positionedwithin the pocket and the incision was closed withoutsutures. In addition, there were also control animals inwhich a pocket was created but no implant was placed,as well as treatment cases receiving a liquid aliquot ofgrowth factor rather than a film implant. Micerecovered uneventfully from this brief procedure withinminutes, and the incision normally healed withoutcomplications in approximately 5–7 days.On days 7 or 14 post-surgery, the mice were again
anesthetized by isoflurane and immediately sacrificed bycervical dislocation. Both the implanted and contral-ateral ears were retrieved and fixed in formalin. The earswere then embedded in paraffin, thin-sectioned in planesparallel to the plane of the pinna and stained withhematoxylin and eosin (H&E). Microvessels weredirectly counted in each ear under a conventional lightmicroscope at 400� , after covering identification markson each slide so that, while counting, the observer wasnot aware of the treatment group to which the slide

ARTICLE IN PRESSR.A. Peattie et al. / Biomaterials 25 (2004) 2789–27982792
belonged. At this level of magnification, a very largenumber of fields were potentially available for observa-tion in each ear. Consequently, 10 locations per ear fromwithin the pocket area, containing vessel densityrepresentative of that ear, were selected at random forquantification.
2.4. Data analysis
A total of 6 animals received implants in eachtreatment case, for each time point (n ¼ 6). Microvesseldensity data are accordingly presented as mean7standard deviation. In addition, vessel density in anyimplanted ear could in principle have been influencedboth by the number of vessels present prior to surgeryand by non-specific new vessel growth secondary tosurgical probing. To account for these effects, so thatthe influence of HA and/or growth factor on new vesselformation would be identifiable per se, we defined adimensionless neovascularization index, NI ; as
NI ¼ðtreatment � CLÞ � ðsham � CLÞ
mean CL; ð1Þ
where ðtreatment � CLÞ refers to the vessel count fromthe implanted ear of a particular animal minus that of itscontralateral ear, averaged over all the animals in aparticular treatment group, ðsham � CLÞ represents thesame quantity for a sham surgery control case thatunderwent pocket formation but received no implant,and mean CL is the average count from all contralateralears over all treatment groups. Thus defined, NI
represents the number of additional vessels presentpost-implant in a treatment group, minus the additionalnumber due to the surgical procedure alone, normalizedby the mean contralateral count. (Mathematically, sinceboth (treatment � CL) and (sham � CL) are themselvesrandom variables presumably following Gaussian dis-tributions, NI is a Gaussian random variable as well.Calculation of its mean and standard deviation there-fore requires appropriately linearly combining theproperties of its constituent terms).Statistical significance was determined using 2-way
ANOVA and post-hoc Fisher’s PLSD analysis (Stat-
View 5.0, SAS Institute Inc., Cary, NC), with signifi-cance taken at the level pp0:05:
3. Results
Hydrogel films prepared with ADH derivatives ofHA, in which the hydrazide functionalization permitsgel formation through macromolecular crosslinks, arestable and highly biocompatible [40]. Prior to cross-linking, the degree of ADH substitution on the HA usedin this investigation was determined to be 60%, basedon the number of available glucuronate residues
modified. Gels were formed by reaction with PEG-diald, and rapid gelation occurred as inter- and intra-molecular bis-hydrazone crosslinks formed betweenpendant hydrazide moieties (Fig. 1). Gels having robustbiomechanical properties were obtained for HA–ADHat an aldehyde:ADH ratio of 1:2. These gels can bedehydrated to a film state, which swelled with nodegradation when reversibly rehydrated in aqueoussolution [40]. When growth factors were included duringcrosslinking, the dialdehyde formed robust hydrazonelinks to the HA–ADH or CS–ADH and hydrolyticallylabile Schiff base linkages to the Lys residues of thegrowth factor. These transient linkages, which were notquantified, provided slow release of the growth factorin vivo. In separate studies with the crosslinkedhydrogels, although fluorescent dextran (70 kDa) orproteins larger than BSA (66 kDa) were released fromthe gels at a rate corresponding to that of hydrogelbreakdown, small molecules were released at diffusioncontrolled rates in the absence of a chemical linkage [40].Fig. 2 shows a schematic diagram of the experimental
model used to investigate the capacity of such HA–ADH films to induce angiogenesis. New microvesseldevelopment was assessed for a series of control andexperimental conditions, including (i) surgery with noimplant placement, (ii) an inert implant fabricated fromCS–ADH, (iii) an HA film alone with no growth factor,(iv) VEGF alone, delivered as a liquid aliquot so that itsphysiologic effects would not be altered by interactionwith a hydrogel, (v) aqueous bFGF, then (vi) an HAfilm pre-loaded with VEGF and finally (vii) an HA filmpre-loaded with bFGF. The control experiments (i–v)provided quantitative evaluation of vessel formationsecondary to surgical pocket formation without implantplacement, as well as of tissue response to growth factoror film individually. They thereby permitted directevaluation of the effects attributable to implanted filmspreloaded with cytokines (vi and vii).Representative photographic images of tissue micro-
vascularization at 14 days post-surgery for four of thesecases (Fig. 3) show marked differences that character-ized the differing tissue response to each implant.Contralateral ear sections (Fig. 3a) had very similarappearance for all animals, with no evidence of anymeasurable systemic response to the implant at eithertime point of the experiment for any of the treatmentcases. Chondrocytes were widely distributed in thistissue, as were hair follicles and sebaceous as well asother glands. Few capillaries were evident, but theendothelial borders of those that were present appearedintact. Red cells were confined within defined capillarywalls.In contrast, although by day 14 the implants were
fully absorbed and did not appear in the images, treatedears showed distinct histological differences from thecontralateral controls. Ears implanted with either

ARTICLE IN PRESS
Fig. 3. H&E stained representative images of ear tissue for different implant types, 400� , day 14 post-surgery: (a) contralateral ears; (b) treatment
with inert CS implant; (c) treatment with aqueous VEGF; and (d) treatment with VEGF pre-loaded HA implant. Note the increased microvessel
density in the CS and VEGF-only treated ears, and the greatly increased density along with many extravasated red cells in the HA+VEGF treated
ear. Ca—capillary, Ch—chondrocytes, E—erythrocytes, H—hair follicle, P—polymorphonuclear leukocyte.
R.A. Peattie et al. / Biomaterials 25 (2004) 2789–2798 2793
non-vasoactive CS disks (Fig. 3b) or aqueous VEGF(Fig. 3c) continued to show the chondrocytes, hairfollicles and glands typical of ear pinnas. Inflammatorycells, primarily polymorphonuclear leukocytes recogniz-able by their lobed nuclei, were also identified in thesesections. In addition, there were areas of substantiallyincreased microvessel density beyond that present in thecontralateral ears. Although the walls of these vesselsappeared intact, red cells could also be found that did notappear to be confined within identifiable vessels. Micro-vessel densities were not statistically different in these twocases, and extravasated red cells could be found in each.The greatest density of microvessels, as well as of the
number of areas of high microvessel density, was foundin ears treated with VEGF pre-loaded HA disks. Mostof the tissue in these ears remained undisturbed, withapparently normal distributions of non-inflammatorycells. However, regions of highly perturbed tissue couldalso be found (Fig. 3d). In these regions, large numbersof apparently partially formed new microvessels couldbe identified through associated chains of red cells.These vessels often did not show a complete endothelialborder. Furthermore, in correspondence with the pre-sence of proliferating incomplete neovessels, very highnumbers of extravasated red cells were distributedamong the recognizable vessels. Large numbers ofpolymorphonuclear inflammatory cells were also recog-nizable in these areas.Neovessel density was quantitatively unchanged at
either time in contralateral ears, in all treatment groups,
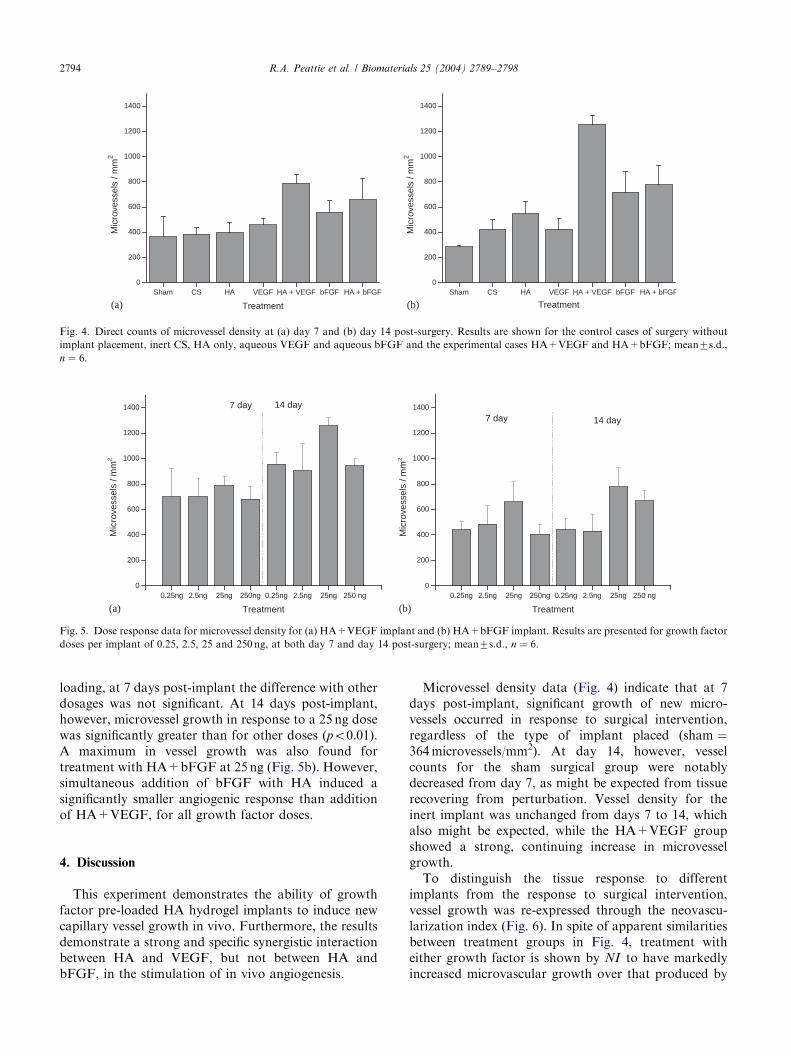
indicating the experiment produced no systemic re-sponse. Quantitative counts of microvessel densities ineach group at 7 and 14 days post-implant are illustratedin Fig. 4. At 7 days post-surgery (Fig. 4a), no significantdifferences in vessel count were observed betweengroups receiving the sham treatment, inert CS, HA orVEGF individually. Although treatment with bFGFalone (mean vessel density 558microvessels/mm2) pro-duced a stronger effect than any of these cases, thegreatest stimulation of vessel growth was shown in thegroup receiving HA+VEGF (789microvessels/mm2;po0:001 except for comparison vs. the group receivingHA+bFGF, for which po0:05).Trends in vessel proliferation at 14 days post-surgery
(Fig. 4b) were generally more clear than at 7 days. Vesseldensities for most groups receiving implants weregreater than those at 7 days, and almost all pairwisecomparisons between treatment groups at 14 days werestatistically significant. However, vessel density in thegroup receiving CS was unchanged from 7 days, whiledensity in the sham group was found to decrease. As atday 7, the most striking vessel growth occurred inresponse to an HA+VEGF implant (1256microvessels/mm2). This was 61% greater vessel density than in thenext largest treatment group, that receiving HA+bFGF(777microvessels/mm2).Co-addition of VEGF with HA resulted in pro-
nounced microvessel growth over all delivered VEGFdosages from 0.25 to 250 ng (Fig. 5a). Although thelargest angiogenic response was produced with 25 ng
ARTICLE IN PRESS
Sham CS HA VEGF HA + VEGF bFGF HA + bFGF0
200
400
600
800
1000
1200
1400M
icro
vess
els
/ mm
2
Treatment
Sham CS HA VEGF HA + VEGF bFGF HA + bFGF0
200
400
600
800
1000
1200
1400
Mic
rove
ssel
s / m
m2
Treatment(a) (b)
Fig. 4. Direct counts of microvessel density at (a) day 7 and (b) day 14 post-surgery. Results are shown for the control cases of surgery without
implant placement, inert CS, HA only, aqueous VEGF and aqueous bFGF and the experimental cases HA+VEGF and HA+bFGF; mean7s.d.,
n ¼ 6:
0.25ng 2.5ng 25ng 250ng 0.25ng 2.5ng 25ng 250 ng0
200
400
600
800
1000
1200
1400 14 day7 day
Mic
rove
ssel
s / m
m2
Treatment
0.25ng 2.5ng 25ng 250ng 0.25ng 2.5ng 25ng 250 ng0
200
400
600
800
1000
1200
1400
14 day7 day
Mic
rove
ssel
s / m
m2
Treatment(a) (b)
Fig. 5. Dose response data for microvessel density for (a) HA+VEGF implant and (b) HA+bFGF implant. Results are presented for growth factor
doses per implant of 0.25, 2.5, 25 and 250 ng, at both day 7 and day 14 post-surgery; mean7s.d., n ¼ 6:
R.A. Peattie et al. / Biomaterials 25 (2004) 2789–27982794
loading, at 7 days post-implant the difference with otherdosages was not significant. At 14 days post-implant,however, microvessel growth in response to a 25 ng dosewas significantly greater than for other doses (po0:01).A maximum in vessel growth was also found fortreatment with HA+bFGF at 25 ng (Fig. 5b). However,simultaneous addition of bFGF with HA induced asignificantly smaller angiogenic response than additionof HA+VEGF, for all growth factor doses.
4. Discussion
This experiment demonstrates the ability of growthfactor pre-loaded HA hydrogel implants to induce newcapillary vessel growth in vivo. Furthermore, the resultsdemonstrate a strong and specific synergistic interactionbetween HA and VEGF, but not between HA andbFGF, in the stimulation of in vivo angiogenesis.
Microvessel density data (Fig. 4) indicate that at 7days post-implant, significant growth of new micro-vessels occurred in response to surgical intervention,regardless of the type of implant placed (sham ¼364microvessels/mm2). At day 14, however, vesselcounts for the sham surgical group were notablydecreased from day 7, as might be expected from tissuerecovering from perturbation. Vessel density for theinert implant was unchanged from days 7 to 14, whichalso might be expected, while the HA+VEGF groupshowed a strong, continuing increase in microvesselgrowth.To distinguish the tissue response to different
implants from the response to surgical intervention,vessel growth was re-expressed through the neovascu-larization index (Fig. 6). In spite of apparent similaritiesbetween treatment groups in Fig. 4, treatment witheither growth factor is shown by NI to have markedlyincreased microvascular growth over that produced by

ARTICLE IN PRESS
Sham CS HA VEGF HA + VEGF bFGF HA + bFGF0
1
2
3
4
5
6
7
8
9
10
NI
Treatment
Sham CS HA VEGF HA + VEGF bFGF HA + bFGF0
1
2
3
4
5
6
7
8
9
10
NI
Treatment(a) (b)
Fig. 6. Neovascularization index at (a) day 7 and (b) day 14 post-surgery. Results are shown for surgery without implant placement, inert CS, HA
only, aqueous VEGF and aqueous bFGF and the experimental cases HA+VEGF and HA+bFGF; mean7s.d., n ¼ 6: NI is defined in the text,
Eq. (1).
R.A. Peattie et al. / Biomaterials 25 (2004) 2789–2798 2795
HA alone. More importantly, a striking finding of thisexperiment is the very strong, synergistic in vivo tissueresponse to co-addition of HA and VEGF, whichproduced NI ¼ 6:7 at day 14 (Fig. 6b). This was nearlytwice the angiogenic effect of any other treatment.Moreover, the response to HA+VEGF exceeded thesum of the tissue response to HA alone (NI ¼ 1:8) plusthe response to VEGF alone (NI ¼ 1:3) by a factor of2.2. In contrast, at both time points, the sum of theangiogenic response to HA alone plus that to bFGFalone (NI ¼ 2:9 at day 14) was greater than the effect ofco-addition of HA+bFGF (NI ¼ 3:4 at day 14). Hence,in contrast to the potent synergistic angiogenic responseelicited by co-addition of HA+VEGF, co-addition ofHA+bFGF produced no such synergy.Montesano and co-authors [28] reported a similarly
strong synergistic interaction between o-HA and VEGFin producing endothelial proliferation into collagen gelsin vitro. Although the authors quantified microvesselgrowth by summation of capillary sprout lengths, adifferent endpoint than total vessel number as countedherein, it is interesting to note that similar ratios werefound in both experiments. Thus the total sprout lengthproduced in vitro by combined loading with o-HA andVEGF exceeded the sum of the lengths produced by o-HA and VEGF individually by approximately a factorof 2.5. That ratio of synergy is highly consistent with theratio of 2.2 obtained in our in vivo experiment, in spiteof the much greater complexity of the tissue environ-ment in vivo than in controlled gel conditions. Takentogether, these findings strongly suggest that angiogenicinteractions between HA and VEGF are intrinsicallysynergistic regardless of any other conditions in whichthey occur, independent of the ECM environment.Both the molecular mechanisms by which HA
hydrogel films induce tissue angiogenesis and themechanisms of interaction between HA and VEGF are
as yet unknown. Of the many known angiogenic factors,VEGF is unique in that it is the only known cytokinewith mitogenic effects primarily confined to endothelialcells. VEGF is produced by a variety of normal andtumor cells. Its expression correlates with periods ofcapillary growth during embryologic development,wound healing and the female reproductive cycle, aswell as with tumor expansion. In addition, suppressionof VEGF expression in adult mice or neutralization ofVEGF receptors suppresses tumor growth [32–37], whileVEGF7 mice are mortal in utero [36]. Consequently,VEGF is thought to be a major promoter of bothphysiologic and pathologic angiogenesis. Furthermore,VEGF has been shown to be an anti-apoptosis survivalfactor for endothelial cells even during periods ofmicrovessel stasis [36]. Although a full molecularexplanation of interactions between VEGF and HAhas not yet been given, it is known that o-HA-inducedcell proliferation in monolayer cultures is not enhancedby co-addition of VEGF [28]. Hence their physiologiceffect is unlikely due solely to a synergistic stimulationof endothelial cell propagation.As was pointed out by Montesano et al. o-HA and
VEGF may activate complementary convergent intra-cellular signaling pathways, with consequent amplifica-tion of capillary growth. Synergy might also result ifeither agent induced up-regulation of receptors for theother. Alternatively, it has been shown that HApromotes cell movement in early granulation tissueduring wound healing [43]. In addition, it is well knownthat VEGF can enhance tissue secretion of a variety ofpro-angiogenic proteases, including uPA, MMP-1 andMMP-2 [44–46]. It has also been shown that inhibitionof VEGF or the VEGF-R2 receptor can suppressexpression of MMP-2 and MMP-3 [47]. These observa-tions together suggest that a combination of VEGF-induced physical disruption of the ECM and

ARTICLE IN PRESSR.A. Peattie et al. / Biomaterials 25 (2004) 2789–27982796
HA-mediated ECM enhancement may condition theECM so as to produce a favorable environment for aprolonged angiogenic response. Nevertheless, the possi-bility of a direct physico-chemical interaction between o-HA and VEGF cannot be dismissed. Once secreted,VEGF in vivo has been shown to largely remain boundto cell surfaces and neighboring ECM [36]. Paracrinesignaling can occur when extracellularly stored VEGF isliberated by proteinase activity. This suggests thatenhanced angiogenic activity produced by the cyto-kine-linked HA hydrogels used in this study may in partbe due to the availability of HA as a sequestration sitemimicking native ECM conditions. In their hydratedstate, the modified-HA films used in this study havepreviously been shown to have an open, networkstructure suitable for storing endogenous molecules aswell as exogenous pharmacologic agents [40]. To theextent that the film provides a repository for retainingVEGF or other cytokines, paracrine hormones andgrowth factors released by proliferating capillaries, thegrowth factor half-life in tissues in vivo would beexpected to increase. Furthermore, interactions betweenVEGF molecules and HA might also facilitate bindingaffinities between VEGF and its receptors throughallosteric modulatory mechanisms. Finally, it shouldbe noted that these potential mechanisms are by nomeans mutually exclusive, and it is not unlikely that thetissue response to co-addition of HA and VEGF mayinvolve more than one of them.One potential drawback of our procedure is that H&E
staining, although demarking red bloods cells clearly,yields relatively poor distinction between endothelialcells and other cells present in the ear tissue. SinceVEGF is known to induce vascular leakage andendothelial fenestration [36], the possibility for extra-vasated red cells in our images leads to the potential foroverestimation of the number of organized capillarytubules present. However, as a systematic error, anysuch overestimation would be expected to affect ourmicrovessel counts for each treatment case. Moreimportantly, as a ratio, NI would not be influenced byoverestimated counts. Our NI measurements were wellcontrolled, self-consistent, and were coherent with thefindings of other authors [28]. Consequently, we see noreason to doubt the veracity of the trends in vesselgrowth suggested by those measurements.An ideal growth factor delivery matrix should have
several important attributes. It should be non-toxic,non-immunogenic, biocompatible, stable and mechani-cally strong enough for feasibility of handling during theimplant procedure. It should provide confined, localizedrelease of the growth factor or factors at a controllablerate. Once released, the delivered agents should main-tain full bioactivity, without secondary systemic effects,and all the growth factor contained within the implantshould be fully delivered with no loss of activity. In
addition, the delivery vehicle should not be costly toprepare. A variety of approaches to optimizing cytokinedelivery have been suggested recently, including me-chanical stimulation of the delivery implant [48], dualgrowth factor combinations [49,50] multi-layer secretorycell encapsulation [51], chemical modification of thecytokine [52] and encapsulation in specifically designedliposome carriers [53]. However, there have been fewreports of GAG-based biomaterials used as cytokinedelivery systems.Compared to those methods, the HA film implants
described herein have a number of significant advan-tages. GAG hydrogel films are stable, strong, highlybiocompatible and non-immunogenic, and biodegradein situ with no systemic sequelae. Moreover, the rate ofdegradation can be controlled by varying the extent ofcrosslinking within the film, a parameter that itself canbe easily altered during the gel fabrication process. Thefilms are simple to fabricate at room temperaturewithout use of toxic or noxious reagents [40]. Theirporous structure when hydrated is ideal for sequesteringtherapeutic agents, and HA gels have been shown to becapable of providing sustained release of those agents[40]. Most importantly, the breakdown products of HAfilms, o-HA fragments, themselves have active, pro-angiogenic properties. Those properties can be takenadvantage of to design an ‘‘intelligent’’ matrix thatparticipates in the tissue response to the implant, ratherthan simply serving as an inert conduit for thesequestered agents. One important application of thoseproperties is the use discussed in this manuscript togenerate a synergistic angiogenic response when used fordelivery of VEGF. This application may ultimately leadto new therapeutic strategies.
5. Conclusions
The angiogenic capacity of hyaluronic acid (HA)hydrogel films loaded with cytokine growth factors hasbeen investigated in vivo in a mouse model. Chemicallymodified HA was crosslinked with PEG-diald after prioraddition of the growth factor, or without addition forcontrol cases, to form reversibly hydratable hydrogelfilms. Round disks cut from these films were thenimplanted in the mouse ear pinna. As evaluated by aneovascularization index, significant differences inmicrovessel density dependant on the implant contentswere found on days 7 and 14 post-implant. All treatmentgroups receiving an implant showed substantially morevessel density than control ears that underwent surgerybut received no implant. Vessel density at day 7 in thegroup receiving an HA implant with no added cytokinewas not statistically different from that of the groupreceiving an inert CS implant, but by day 14 exceededthe CS group by 50% (NI ¼ 1:8 vs. 1.1). Administration

ARTICLE IN PRESSR.A. Peattie et al. / Biomaterials 25 (2004) 2789–2798 2797
of aqueous bFGF produced greater increases in vesseldensity than administration of aqueous VEGF at eithertime point, and produced substantially more vesselgrowth than the pure HA implant. The most prominentresult obtained, however, was a dramatic synergisticinteraction between HA and VEGF. Co-addition of HAand VEGF produced twice the vessel density of the nextgreatest treatment, HA+bFGF, (NI ¼ 6:7 vs. 3.4), andproduced more than twice the density of vessels as thesum of HA alone plus VEGF alone.The ability to stimulate localized microvessel growth
at controlled rates by administration of growth factorsvia GAG carrier films will provide a powerful tool withapplications to a variety of therapeutic interventions.HA films can be fabricated in shapes useful fortreatment of pathologies such as cardiac ischemia,wound healing and surgical bowel adhesions. Manyother processes, for example bone and cartilage regen-eration, may also benefit from this technology. Ther-apeutic approaches to these and many other problemsmay be greatly enhanced by localized, controlled releaseof growth factors.
Acknowledgements
Financial support for this work was provided byawards from the University of Utah Center forBiopolymers at Interfaces and from Connecticut In-novations, Inc. We thank Dr. Yi Luo for many valuablesuggestions, and for preparation of HA–ADH.
References
[1] Knudson CB, Knudson W. Cartilage proteoglycans. Semin Cell
Dev Biol 2001;12:69–78.
[2] Hoekstra D. Hyaluronan-modified surfaces for medical devices.
Med Dev Diagn Ind 1999;48–58.
[3] Laurent TC, Laurent UBG, Fraser JRE. Functions of hyalur-
onan. Ann Rheum Dis 1995;54:429–32.
[4] Farney AC, Cho E, Schweitzer EJ, Dunkin B, Philosophe B,
Colonna J, Jacobs S, Jarrell B, Flowers JL, Bartlett ST.
Simultaneous cadaver pancreas living-donor kidney transplanta-
tion: a new approach for the type I diabetic uremic patient. Ann
Surg 2000;231:696–703.
[5] Aruffo A, Stamenkovic I, Mulnick M, Underhill CB, Steed B.
CD44 is the principal cell surface receptor for hyaluronate. Cell
1990;61:1303–13.
[6] Fraser JRE, Laurent TC, Laurent UBG. Hyaluronan: its nature,
distribution, functions and turnover. J Int Med 1997;242:27–33.
[7] Dowthwaite GP, Edwards JCW, Pitsillides AA. An essential role
for the interaction between hyaluronan and hyaluronan binding
proteins during joint development. J Histochem Cytochem
1998;46:641–51.
[8] Collis L, Hall C, Lange I, Ziebel MR, Prestwich GD, Turley EA.
Rapid hyaluronan uptake is associated with enhanced motility:
implications for an intracellular mode of action. FEBS Lett
1998;440:444–9.
[9] Hardwick C, Hoare K, Owens R, Hohn HP, Hook M, Moore D,
Cripps V, Austen I, Nance DM, Turley EA. Molecular cloning of
a novel hyaluronan receptor that mediates tumor cell motility.
J Cell Biol 1992;117:1343–50.
[10] Entwistle J, Hall CL, Turley EA. Receptors: regulators of
signaling to the cytoskeleton. J Cell Biochem 1996;61:569–77.
[11] Lesley J, Hyman R, English N, Catterall JB, Turner GA. CD44 in
inflammation and metastasis. Glycoconjugate J 1997;14:611–22.
[12] Prevo R, Banerji S, Ferguson DJP, Clasper S, Jackson DG.
Mouse LYVE-1 is an endocytic receptor for hyaluronan in
lymphatic endothelium. J Biol Chem 2001;276:19420–30.
[13] Zhou B, Oka JA, Singh A, Weigel PH. Purification and subunit
characterization of the rat liver endocytic hyaluronan receptor.
J Biol Chem 1999;274:33831–4.
[14] Day AJ, Prestwich GD. Hyaluronan binding proteins: tying up
the giant. J Biol Chem 2001;277:4585–8.
[15] Day AJ, Sheehan J. Hyaluronan: polysaccharide chaos to protein
organization. Curr Opin Struct Biol 2001;11:617–22.
[16] Toole BP. Hyaluronan in morphogenesis. J Int Med 1997;242:
35–40.
[17] Gerdin B, Hallgren R. Dynamic role of hyaluronan (HYA) in
connective tissue activation and inflammation. J Int Med
1997;242:49–55.
[18] Chen WYJ, Abatangelo G. Functions of hyaluronan in wound
repair. Wound Repair Regen 1999;7:79–89.
[19] Forrester JV, Balasz EA. Inhibition of phagocytosis by high
molecular weight hyaluronate. Immunology 1980;40:435–46.
[20] Kujawa MJ, Carrino DA, Caplan AI. Substrate-bonded hyaluro-
nic acid exhibits a size-dependent stimulation of chondrogenic
differentiation of stage 24 limb mesenchymal cells in culture. Dev
Biol 1986;144:519–28.
[21] Goldberg RL, Toole BP. Hyaluronate inhibition of cell prolifera-
tion. Arthritis Rheum 1987;30:769–78.
[22] Rooney P, Kumar S, Ponting J, Wang M. The role of hyaluronan
in tumor neovascularization. Int J Cancer 1995;60:632–6.
[23] West DC, Kumar S. The effects of hyaluronate and it
oligosaccharides on endothelial cell proliferation and monolayer
integrity. Exp Cell Res 1989;183:179–96.
[24] Sattar A, Kumar S, West DC. Does hyaluronan have a role in
endothelial cell proliferation in the synovium? Semin Arthritis
Rheum 1992;22:37–43.
[25] Sattar A, Rooney P, Kumar S, Pye D, West DC, Scott I, Ledger
P. Application of angiogenic oligosaccharides of hyaluronan
increase blood vessel numbers in skin. J Invest Dermatol 1994;
103:573–9.
[26] West DC, Hampson IN, Arnold F, Kumar S. Angiogenesis
induced by degradation products of hyaluronic acid. Science
1985;228:1324–6.
[27] Lees VC, Fan T-PD, West DC. Angiogenesis in a delayed
revascularization model is accelerated by angiogenic oligosac-
charides of hyaluronan. Lab Invest 1995;73:259–66.
[28] Montesano R, Kumar S, Orci L, Pepper MS. Synergistic effect of
hyaluronan oligosaccharides and vascular endothelial growth
factor on angiogenesis in vitro. Lab Invest 1996;75:249–62.
[29] Suhardja A, Hoffman H. Role of growth factors and their
receptors in proliferation of microvascular endothelial cells.
Microsc Res Tech 2003;60:70–5.
[30] Ribatti D, Presta M. The role of fibroblast growth factor-2 in the
vascularization of the chick embryo chorioallantoic membrane.
J Cell Mol Med 2002;6:439–46.
[31] Liekens S. The role of growth factors, angiogenic enzymes and
apoptosis in neovascularization and tumor growth-collected
publications. Verh K Acad Geneeskd Belg 2002;64:197–224.
[32] Ferrara N. Role of vascular endothelial growth factor in
physiologic and pathologic angiogenesis: therapeutic implications.
Semin Oncol 2002;29:10–4.

ARTICLE IN PRESSR.A. Peattie et al. / Biomaterials 25 (2004) 2789–27982798
[33] Jain RK. Tumor angiogenesis and accessibility: role of vascular
endothelial growth factor. Semin Oncol 2002;29:3–9.
[34] Goth MI, Hubina E, Raptis S, Nagy GM, Toth BE. Physiological
and pathological angiogenesis in the endocrine system. Microsc
Res Tech 2003;60:98–106.
[35] Carmeliot P. Angiogenesis in Health and Disease. Nat Med
2003;9:653–60.
[36] Ferrara N, Gerber H-P, LeCouter J. The biology of VEGF and its
receptors. Nat Med 2003;9:669–76.
[37] Paku S, Paweletz N. First steps of tumor-related angiogenesis.
Lab Invest 1991;65:334–46.
[38] Nikosia RF, Bonanno E, Smith M, Yurchenco P. Modulation of
angiogenesis in vitro by laminin–entactin complex. Dev Biol
1994;164:197–206.
[39] Ingber DE, Folkmann J. Mechano-chemical switching between
growth and differentiation during growth factor-stimulated
angiogenesis in vitro: role of the extracellular matrix. J Cell Biol
1989;109:317–30.
[40] Luo Y, Kirker KR, Prestwich GD. Cross-linked hyaluronic acid
hydrogel films: new biomaterials for drug delivery. J Control
Release 2000;69:169–84.
[41] Luo Y, Prestwich GD. Hyaluronic acid-N-hydroxysuccinimide: a
useful intermediate for bioconjugation. Bioconjugugate Chem
2001;12:1085–8.
[42] Luo Y, Ziebell MR, Prestwich GD. A hyaluronic acid-taxol
antitumor bioconjugate targeted to cancer cells. Biomacromole-
cules 2000;1:208–18.
[43] Kirker KR, Luo Y, Nielson JH, Shelby J, Prestwich GD.
Glycosaminoglycan hydrogel films as bio-interactive dressings
for wound healing. Biomaterials 2002;23:3661–71.
[44] Forbes K, Webb MA, Sehgal I. Growth factor regulation of
secreted matrix metalloproteinase and plasminogen activators in
prostate cancer cells, normal prostate fibroblasts and normal
osteoblasts. Prostate Cancer Prostatic Dis 2003;6:148–53.
[45] Kim MH. Flavonoids inhibit VEGF/bFGF-induced angiogenesis
in vitro by inhibiting the matrix-degrading proteases. J Cell
Biochem 2003;89:529–38.
[46] Yabushita H, Shimazu M, Noguchi M, Kishida T, Narumiya H,
Sawaguchi K, Noguchi M. Vascular endothelial growth factor
activating matrix metalloproteinase in ascitic fluid during
peritoneal dissemination of ovarian cancer. Oncol Rep 2003;
10:89–95.
[47] Wagner S, Fueller T, Hummel V, Rieckmann P, Tonn JC.
Influence of VEGF-R2 inhibition on MMP secretion and motility
of microvascular human cerebral endothelial cells (HCEC).
J Neurooncol 2003;62:221–31.
[48] Lee KY, Peters MC, Anderson KW, Mooney DJ. Controlled
growth factor release from synthetic extracellular matrices.
Nature 2000;408:998–1000.
[49] Richardson TP, Peters MC, Ennett AB, Mooney DJ. Polymeric
system for dual growth factor delivery. Nature Biotech 2001;
19:1029–34.
[50] Gillis P, Savia U, Volpert OV, Jimenez B, Waters CM, Panos RJ,
Bouck NP. Keratinocyte growth factor induces angiogenesis and
protects endothelial barrier function. J Cell Sci 1999;112:2049–57.
[51] Rokstad AM, Holtan S, Strand B, Steinkjer B, Ryan I, Kulseng
R, Skjak-Braek G, Espevik T. Microencapsulation of cells
producing therapeutic proteins: optimizing cell growth and
secretion. Cell Transplant 2002;11:313–24.
[52] Luo P, Hayes RJ, Chan C, Stark DM, Hwang MY, Jacinto JM,
Jovvadi P, Chung HS, Kundu A, Ary MI, Dubiyat BI.
Development of a cytokine analog with enhanced stability using
computational ultrahigh throughput screening. Protein Sci
2002;11:1218–26.
[53] Kanaoka F, Takahashi K, Yoshikawa T, Jizomoto H, Nishihara
Y, Hirano K. A novel and simple type of liposome carrier
for recombinant interleukin-2. J Pharm Pharmacol 2001;53:
295–302.
Copyright © 2022 FDOKUMEN




















